Embed Size (px)
Citation preview

Asada et al. 1
The roles of epithelial cell derived type 2 initiating cytokines in experimental 1
allergic conjunctivitis 2
Yosuke Asada1,2, Susumu Nakae2, Waka Ishida3, Kanji Hori1, Jobu Sugita1, Katsuko 3
Sudo4, Ken Fukuda3, Atsuki Fukushima3, Hajime Suto5, Akira Murakami1, Hirohisa 4
Saito6, Nobuyuki Ebihara1, and Akira Matsuda1. 5
1Laboratory of Ocular Atopic Diseases, Department of Ophthalmology, and 6
5Department of Dermatology Juntendo University School of Medicine, Tokyo, Japan. 7
2Frontier Research Initiative, Institute of Medical Science, University of Tokyo, Tokyo, 8
Japan. 3Department of Ophthalmology, Kochi University School of Medicine, Nangoku, 9
Japan. 4Animal Research Center, Tokyo medical University, Tokyo, Japan. 6Department 10
of Allergy and Immunology, National Research Institute for Child Health and 11
Development, Tokyo, Japan. 12
Corresponding author Akira Matsuda, MD, PhD. Laboratory of Ocular Atopic 13
Diseases, Department of Ophthalmology Juntendo University School of Medicine 2-1-1 14
Hongo, Bunkyo-Ku Tokyo 113-8431, Japan. Telephone: +81-3-5802-1228, Fax: 15
+81-3-5689-0394 E-mail: [email protected] 16
Declaration of all sources of funding: This study was supported by Grants-in-Aid from 17
the Japanese Society for the Promotion of Science (No. 24659768 and No. 26462698), 18

Asada et al. 2
and from the Takeda Science Foundation. 19
Word count: 20
21

Asada et al. 3
Abstract 22
Purpose: To clarify the possible involvements of type 2 initiating cytokines, interleukin 23
(IL)-25, IL-33, thymic stromal lymphopoietin (TSLP) for the pathophysiology of 24
allergic conjunctivitis, we evaluated ragweed (RW)-induced experimental allergic 25
conjunctivitis (EAC) models using IL-25 knockout (KO), IL-33 KO, TSLP receptor 26
(TSLPR) KO mice. 27
Methods: IL-25 KO, IL-33 KO, TSLPR KO and BALB/c wild type mice were 28
sensitized twice with RW in alum, and then challenged with RW in eye drops. The 29
clinical scores and eosinophil infiltration were evaluated. Serum IgE and cytokine 30
expression in the conjunctival tissue were quantified and immunohistochemical analysis 31
was carried out. 32
Results: Significant reductions of the clinical scores and the numbers of infiltrating 33
eosinophils were observed in the RW- EAC models using IL-33 KO mice. There were 34
no significant differences in the clinical scores and the numbers of infiltrating 35
eosinophils among IL-25KO, TSLPR KO and wild-type mice. Serum IgE concentration 36
was upregulated after RW challenges, and there were no differences among the mouse 37
genotypes. Il4, il5, il13 and ccl5 mRNA expression was diminished in the conjunctivae 38
of RW-EAC models using IL-33 KO mice compared to those of wild type mice. IL-33 39

Asada et al. 4
expression was upregulated as early as one hour after RW eye drop challenge. The 40
numbers of infiltrated basophils were diminished in the conjunctivae of RW-EAC 41
models using IL-33 KO mice compared to those of wild type mice. 42
Conclusions: Among the type-2 initiating cytokines, IL-33 plays major roles for 43
conjunctival inflammation in RW-EAC model. 44
Key words 45
Type 2 initiating cytokines, IL-25, IL-33, Thymic stromal lymphopoietin, Experimental 46
allergic conjunctivitis. 47
48

Asada et al. 5
INTRODUCTION 49
Type 2 immune responses are inflammatory conditions associated with parasite 50
infections1, or with atopic diseases like asthma, atopic dermatitis and atopic 51
keratoconjunctivitis.2 Type 2 immune responses are characterized by the activation of 52
CD4+ T helper type 2 (Th2) cells along with the production of typical type-2 immunity 53
associated cytokines, [i.e. interleukin-4 (IL-4), IL-5, and IL-13]. Although various 54
external stimuli (including pollens, house dust mites, food allergens and parasites) could 55
induce type 2 responses, these antigens can not directly activate Th2 cells because they 56
are too large to be phagocytosed by antigen-presenting cells.3 The roles of epithelial 57
cell-derived type 2 initiating cytokines [IL-25, IL-33, and thymic stromal lymphopoietin 58
(TSLP) ] were characterized recently as indispensable cytokines for initiating type 2 59
immune responses stimulated by these type 2 immunity-related antigens.3 60
We previously reported the expression of IL-334 and TSLP5 mRNA and protein in the 61
giant papillae tissue obtained from patients of vernal keratoconjunctivitis (VKC) and 62
atopic keratoconjunctivitis (AKC). On the other hand, our study group established IL-33 63
knockout (KO) mouse and reported that IL-33 have essential roles in papain-induced 64
lung inflammation, which considered to be innate immune system dependent type 2 65
inflammation.6 The roles of IL-33 were also reported using ovalbumin (OVA)-induced 66

Asada et al. 6
asthma model7-9 , an established model for acquired immune system dependent type 2 67
inflammation. Ragweed (RW)-induced experimental allergic conjunctivitis (EAC) had 68
been used as a common model for T cell (acquired immunity)-dependent allergic 69
conjunctivitis.10 Matsuba-Kitamura et al. reported that addition of recombinant IL-33 at 70
the time of RW eyedrop challenge developed augmented eosinophil infiltration in the 71
conjunctival tissue in their RW-EAC models.11 72
TSLP is produced by epithelial cells in response to various protein allergens (i.e. OVA) 73
and protease allergens (i.e. pollens and papain).12 TSLP activates dendritic cells (DCs) 74
through TSLP receptor (TSLPR)-IL-7Rα receptor heterodimer complex.13 75
TSLP-activated DCs express OX40L, which initiate T cell mediated type 2 76
inflammation by activating OX40 positive T cells.13 The upregulation of TSLP, TSLPR, 77
and OX40L mRNA expression in RW-EAC models was reported by Zheng et al.14 IL-25 78
was originally discovered in cDNA libraries from highly polarized Th2 cells.15 IL-25 79
also produced by epithelial cell, mast cells, eosinophils, macrophages.3 The importance 80
of IL-25 for type 2 inflammation was demonstrated by the experiments using mouse 81
OVA-induced asthma models, in which attenuated airway inflammation and airway 82
hyper-responsiveness (AHR) were observed by deletion of IL-25 gene.16 There was no 83

Asada et al. 7
report concerning the expression of IL-25 in the conjunctival tissue and its roles for 84
pathophysiology of allergic conjunctivitis. 85
In this study, we investigated the roles of type 2 initiating cytokines in the 86
pathophysiology of allergic conjunctivitis by evaluating RW-EAC models using IL-25 87
KO, IL-33 KO, TSLPR KO mice and congenic wild type mice. 88
MATERIAL and METODS 89
IL-25 KO, IL-33 KO, TSLPR KO mice 90
IL-33 deficient (IL-33 KO) mice6, IL-25 KO mice17, and TSLPR KO mice 18 were 91
generated as previously reported. BALB/c wild type mice purchased from Japan SLC 92
(Shizuoka, Japan) and KO mice were backcrossed with them at least seven generations 93
to establish congenic IL-33 KO, IL-25 KO and TSLPR KO mice. All the animal 94
experiments conformed to the ARVO Statement on the Use of Animals in Ophthalmic 95
and Vision Research. 96
Alam-ragweed immunized experimental allergic conjunctivitis 97
Mouse experimental allergic conjunctivitis (EAC) was prepared as previously 98
described10 with slight modifications. In brief, short ragweed pollen (RW, purchased 99
from Polysciences, Warrington, PA) was emulsified on Imject Alam® (Thermo 100
Scientific, Rockford, IL). At day 0, 50μl of emulsified RW (50ug of RW with 50μl of 101

Asada et al. 8
alum) was injected into the left hind-footpad and the tail base, and blood was collected 102
from tail vein. Two weeks later, second immunization was carried out using the right 103
hind-footpad. From day 26 to 29, the eyes of immunized mice were challenged by RW 104
in phosphate-buffered saline (PBS) (2mg in 10ul per eye) or by PBS alone daily for 4 105
days. 20 minutes after last eye drop challenge, clinical scores for EAC was evaluated by 106
chemosis, redness, lid edema, tearing, discharge and scratching behavior, based on the 107
criteria described by Magone et al. (Supplementary Table 1).19 24 hours after the last 108
eye drop challenge, the eye balls with lids and conjunctival tissue were collected for 109
histological analyses and for quantification of cytokine expression. For IL-33 protein 110
expression analysis, we made ex vivo culture model of resected conjunctivae from the 111
eye challenged by RW eye drop only once at day 26. Blood samples were also collected 112
for the measurement of serum IgE levels at day 30. 113
Measurement of serum IgE levels 114
Total IgE levels in the sera at day 0 and day 30 were quantified using ELISA MAX 115
mouse IgE ELISA kit (Biolegend, San Diego, CA), according to the manufacturer’s 116
protocol. 117
Histological analysis 118
The eye balls were dissected with conjunctival tissue and eye lids, and fixed in 4% 119

Asada et al. 9
paraformaldehyde (PFA) in PBS. Vertical 2 μm-thick paraffin sections were made and 120
stained with Giemsa staining. Infiltrating eosinophils in the lamina propria mucosae of 121
the tarsal and bulbar conjunctivas throughout each section were counted at the central 122
portion of the eye, which included the pupil and optic nerve head as described 123
previously.20 124
Real-time PCR analysis 125
Conjunctival tissue obtained from the mice eyes was immediately submerged in RNA 126
Later solution (Ambion, Austin, TX) to protect RNA. Total RNA was extracted from the 127
tissue using a NucleoSpin® II RNA isolation kit (Macherey-Nagel GmbH, Duren, 128
Germany). cDNAs were prepared using random primers and ReverTra Ace® reverse 129
transcriptase (both from Toyobo, Osaka, Japan) according to the manufacturer’s 130
protocol. Real-time PCR primers specific for mouse il4, il5, il13, il33, ccl5, ccl11, and 131
gapdh mRNA were designed by QuantPrime (http://quantprime.mpimp-golm.mpg.de/), 132
and summarized in the Supplementary Table 2. Real-time PCR analysis was performed 133
with the ABI PRISM 7300 HT Sequence detection system using FAST-SYBER green 134
master mix (Life Technology Japan, Tokyo, Japan). The relative expression of il4, il5, 135
il13, il33, ccl5, and ccl11 was quantified by comparative Ct methods using gapdh 136
mRNA expression in the same cDNA as the internal controls. 137

Asada et al. 10
Immunohistochemistry 138
Immunofluorescent staining was performed to examine the expression of IL-33 in the 139
conjunctival tissue obtained from experimental conjunctivitis. Goat anti-mouse IL-33 140
polyclonal antibody was purchased from R&D sysmtems (Minneapolis, MN), rat 141
anti-mouse F4/80 antibody (clone CI:A3-1) was from BioLegends (San Diego, CA), and 142
sheep anti-mouse mast cell protease (mcp)1 antibody (clone MS-RM8) was from 143
Moredun Scientific (Midlothian, UK). Basophil specific rat anti-mouse mcp 8 antibody 144
was form BioLegends,21 and rat anti-mouse major basic protein (MBP) antibody was 145
provided from Dr. J Lee (Mayo Clinc).22 5 μm frozen sections were made and then 146
immunostained with the anti-IL-33, anti-MBP, anti-F4/80, anti-mcp1 antibodies. 147
Anti-mcp8 immunohistochemicalstaining was carried out using 2 μm paraffin sections. 148
The stained slides were scanned by confocal microscope (FV-1000; Olympus 149
Corporation, Tokyo, Japan). Negative control specimens were immunostained with 150
control goat IgG or rat IgG antibodies (all from Santa Cruz biotechnology, Santa Cruz, 151
CA) instead of primary antibodies. Double-immunostaining was carried out on pairs of 152
the goat anti-IL-33 antibody with the rat anti-MBP antibody, with the rat anti-F4/80 153
antibody or with the sheep anti-mcp1 antibody. Donkey Alexa 488-conjugated anti-rat 154
IgG antibody, donkey Alexa 594-conjugated anti-goat IgG antibody, donkey Alexa 488 155

Asada et al. 11
conjugated anti-sheep IgG antibody (all from Life Technology Japan, Tokyo, Japan) 156
were used as secondary antibodies. 157
Measurement of IL-33 protein concentration in the supernatant of ex vivo 158
conjunctival tissue culture 159
At indicated time period (1hr, 3hr, 6hr, and 12 hr) after the RW eye drop challenge, the 160
mice were sacrificed and conjunctival tissues were sampled. The tissues were cultured 161
in 1.5ml sterile Eppendorf tube (Eppendorf Japan, Tokyo, Japan) for 60 minutes using 162
200 μl of serum free culture medium (OPTI-MEM, Life Technologies Japan, Tokyo 163
Japan) with protease inhibitors (Complete Mini, Roche Diagnostics GmbH, Mannheim, 164
Germany). IL-33 concentration in the supernatant of ex vivo tissue culture was 165
quantified using Mouse IL-33 ELISA Ready-SET-GO (eBioscience, San Diego, CA), 166
according to the manufacturer’s protocol. 167
Statistical analysis 168
Statistical evaluations of cell numbers, cytokine expressions, serum IgE levels, and 169
histological analysis were performed with the two-tail unpaired Mann–Whitney test. P 170
< 0.05 was considered statistically significant. All experiments were repeated at least 171
three times, and representative data were shown. 172
RESULTS 173

Asada et al. 12
Serum IgE concentration in the RW-EAC model 174
To evaluate the effect of RW-sensitization and the roles of type 2 initiating cytokines 175
during sensitization process, we measured the serum IgE level of the day 0 (before 176
sensitization) and day 30 (after eye drop challenge) by ELISA. Significant increase of 177
total serum IgE at day 30 was observed in all type of mice (including wild type and 178
IL-25 KO, IL-33 KO and TSLPR KO). There was no significant difference among the 179
groups (Figure1). 180
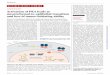
Attenuated clinical symptoms of RW-induced EAC in IL-33 KO mice but not in 181
IL-25 KO or TSLPR KO mice. 182
Mice were immunized with RW in alum at day 0 and day14, and they were challenged 183
from day 26 using RW or PBS eye drops daily for 4 days. 20 min after last challenge, 184
we photographed and scored the severity of EAC by measuring the degree of chemosis, 185
conjunctival redness, lid edema, tearing, discharge and scratching as described in 186
Supplementary Table 1. Clinical score of IL-33KO mice is significantly smaller than 187
that of wild type mice. (Figure 2A and 2B, n=5 per group). There was no significant 188
difference among the clinical scores of IL-25 KO, TSLPR KO and wild type mice 189
(Figure 2B). 190
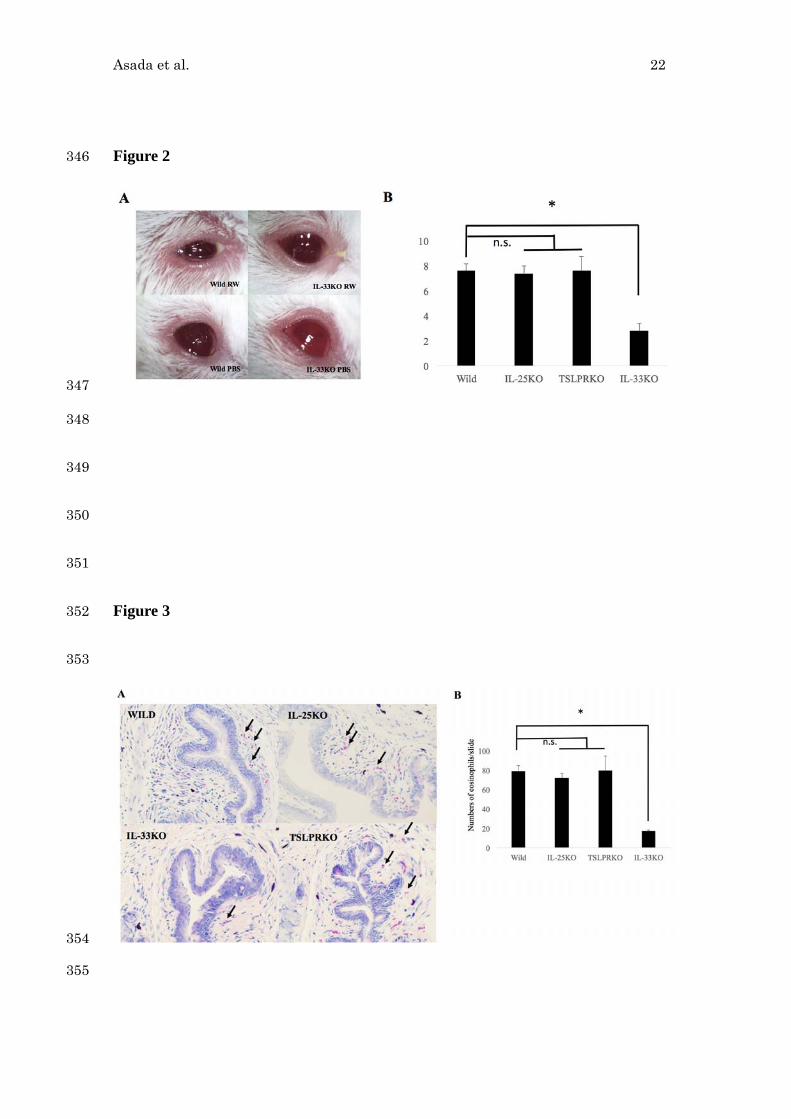
IL-33 deletion diminished the number of infiltrated eosinophils in the conjunctivae 191

Asada et al. 13
of RW-induced EAC. 192
24 hours after the last RW eye drop challenge, the eyes collected and the number of 193
eosinophils was counted using Gimsa stained slides (Figure 3A). The numbers of 194
infiltrating eosinophils in the conjunctivae of IL-33 KO mice were significantly smaller 195
than that of wild type mice. (Figure 3B, n=5 per group). There was no significant 196
difference among the numbers of infiltrating eosinophils of IL-25 KO, TSLPR KO and 197
wild type mice (Figure 3B). 198
Quantification of cytokine expression in the mouse conjunctival tissue obtained 199
from RW-EAC models. 200
The conjunctival tissues were sampled 24 hour after the last RW eye drop challenge, 201
their cytokine expression was quantified. Significant upregulation of il4, il5, il13, and 202
ccl5 mRNA expression was observed in the conjunctival tissue of RW-induced EAC 203
using wild type mice compared to those of PBS-challenged conjunctival tissue of the 204
wild mice (Figure 4, asterisks). No significant upregulation was observed in 205
ccl11-mRNA expression by RW eye drop challenges. Significant attenuation of il4, il5, 206
il13, and ccl5 mRNA expression was observed in IL-33 KO mice compared to those of 207
the wild type mice (Figure 4, double asterisks). Comparable amounts of cytokine 208
expression was observed in the conjunctivae of TSLPR KO mice (Supplementary 209

Asada et al. 14
Figure 1) and of IL-25 KO mice (Supplementary Figure 2) compared to those of the 210
wild type mice, except significantly attenuated il5 mRNA expression in both TSLPR 211
KO mice and IL-25 KO mice (Supplementary Figure 1 and 2, asterisks). In addition, 212
significantly augmented il13 mRNA expression in the conjunctivae of TSLPR KO mice 213
was observed compared to that of wild type mice (Supplementary Figure 1, double 214
asterisks). 215
Time course of il33 mRNA and IL-33 protein expression in RW-EAC models 216
Significant il33 mRNA upregulation was observed in the conjunctival tissue obtained 217
from the wild type mice after 1 hour from the last RW eye drop challenge (Figure 5A). 218
IL-33 protein levels in the ex vivo culture supernatant was significantly upregulated at 219
the conjunctival tissue sample obtained 1 hour after the first RW eye drop challenge 220
(Figure 5B). No IL-33 protein was detected in the conjunctival tissue obtained from RW 221
eye drop challenged IL-33 KO mice (data not shown). 222
Immunohistochemical analysis of EAC 223
Immunofluorescent staining was performed to examine the expression of IL-33, and 224
immunolocalization of eosinophils, macrophages, mast cells and basophils in the 225
conjunctival tissue obtained from EAC. Anti-mcp8 (basophil marker) immunostaining 226
of the RW-EAC models showed basophil infiltration under subepithelial region of 227

Asada et al. 15
RW-challenged eye (Figure 6A and 6B), and the number of infiltrated basophils are 228
significantly higher in RW-EAC models of wild-type mice compared to that of IL-33 229
KO mice (Figure 6C). 230
RW-challenged conjunctivae of wild type mice showed IL-33 protein expression in the 231
conjunctival epithelial cells and in the infiltrating cells of substantia propria of the 232
conjunctival tissue at the vicinity of MBP-positive eosinophils (Figure 7, top row). The 233
PBS-challenged conjunctival tissue of wild type mice showed IL-33 positive 234
immunoreactivity at the epithelilal cell layer, and sparse infiltration of eosinophils in 235
subepithelial region (Figure 7, second row). Conjunctival tissue of RW-EAC model 236
using IL-33KO mice had less MBP-positive eosinophil infiltration compared to those of 237
wild type mice and no IL-33 positive immunostaining was observed (Figure 7, bottom 238
row). Double immunoshistochemical staining using anti-IL-33 antibody and the 239
macrophage marker F4/80 antibody showed that some of the IL-33 positive cells in the 240
substantia propria are also positive for F4/80 antigen (Figure 8 top row, arrows). 241
Similarly, some of the IL-33 positive cells in the substantia propria are also positive for 242
the mast cell marker (mcp1) (Figure 8 bottom row, arrowheads). 243
244
DISCUSSION 245

Asada et al. 16
To explore the roles of type 2 initiating cytokines (IL-25, IL-33, TSLP) in the 246
pathophysiology of allergic conjunctivitis, we made RW-EAC models using IL-25 KO, 247
IL-33 KO and TSLPR KO mice. The measurement of serum total IgE showed clear 248
upregulation of serum IgE in RW-EAC models and no significant difference among the 249
mice groups (Figure 1). The results are consistent with a previous report showing 10 250
folds upregulation of serum IgE in RW-EAC model using BALB/c mice,10 and another 251
report showing the increase of total serum IgE in RW-induced rhinitis models using 252
IL-33 KO mice.23 Similarly, house dust mite (HDM) immunization and subsequent 253
intranasal HDM challenges using TSLPR KO mice or IL-17RB (IL-25 receptor) KO 254
mice showed comparable serum IgE increases to those of wild type mice.24 Taken 255
together, we concluded that the type 2 initiating cytokines do not affect IgE responses 256
on RW-EAC models. 257
Next, we compared clinical scores (Figure 2) and the numbers of infiltrated 258
eosinophils (Figure 3) in the RW-EAC models. The results showed that IL-33 deletion 259
caused attenuated clinical severities and the diminished numbers of eosinophil 260
infiltration in the conjunctivae of RW-EAC models. Attenuation of inflammation of 261
RW-EAC in IL-33 KO mice in both early-phase clinical scores and delayed-phase 262
eosinophilc infiltration was consistent with the previous report showing roles of IL-33 263

Asada et al. 17
during antigen challenge phase of RW-EAC model.11 264
We found no differences among wild type mice, TSLPR KO mice, IL-25 KO mice 265
concerning clinical severities and the numbers of eosinophil infiltration in the 266
conjunctivae of RW-EAC models (Figure 2 and 3). The results of inflammatory 267
cytokine quantification showed attenuated il5 mRNA expression and but no differential 268
expression of il4, ccl5 or ccl11 mRNA in the conjunctival tissue of RW-EAC models 269
using TSLPR-KO mice/IL-25 KO mice compared to wild type mice (Supplementary 270
Figure 1 and 2). Schleimer et al. 25 reported the effects of IL-5, CCL5 and CCL11 for 271
the transendothelial migration of eosinophils. According to their results, IL-5 itself 272
showed minimum effects for eosinophil migration but synergetic effects for eosinophil 273
migration with CCL5 or with CCL11. Absence of differential expression of ccl5 and 274
ccl11 mRNA may account for the lack of difference for eosinophil infiltration among 275
TSLPR KO mice, IL-25 KO mice and wild type mice. 276
To further clarify the roles of IL-33 for the pathophysiology of RW-EAC, 277
chronological changes of IL-33 expression in the conjunctival tissue was examined. 278
Upregulation of il33 mRNA expression, peaked at 1hr after RW eyedrop challenge, was 279
observed (Figure 5A). To quantify IL-33 protein expression at ocular surface in 280
RW-EAC model, we tried to detect IL-33 protein by simply collecting ocular surface 281

Asada et al. 18
exudate using small amount of PBS, however, the IL-33 concentration in the exudates 282
were below the detection level (data not shown). So we measured IL-33 protein 283
concentration of ex vivo culture supernatant of conjunctival tissue obtained after 284
RW-challnege. Significant increase of IL-33 protein in the culture supernatant from 1hr 285
until 12hr after RW eye drop challenge (Figure 5B) suggested continuous IL-33 protein 286
release from IL-33 producing cells. These rapid upregulations of IL-33 mRNA/protein 287
were consistent with the results of RW-induced experimental rhinitis model23 and 288
mixture of allergens (HDM, Aspergillus, Alternaria)-induced lung inflammation 289
models.26 290
We observed significant attenuation of il4 mRNA expression in the conjunctivae of 291
RW-EAC models using IL-33 KO mice compared to those of wild type mice (Figure 4). 292
IL-4 is a key cytokine for type 2 immune responses mediated by adaptive immunity. 293
IL-4 stimulates Th2 cell differentiation from naïve T cell, and induces IL-5 and IL-13 294
expression.27 The effects of IL-33 deletion for il4 mRNA expression were significant 295
even between the eyes with mock eye drop challenges, but it became more apparent 296
when challenged by RW eyedrops (Figure 4). We observed diminished numbers of 297
infiltrating basophils, which is known as the producers of large amount of IL-428, in the 298
conjunctivae of RW-EAC models using IL-33 KO mice compared to those of wild type 299

Asada et al. 19
mice (Figure 6). These results suggested that IL-33 might augment the expression of 300
IL-4 in RW-EAC model by promoting systemic basophil expansion or by promoting 301
basophil infiltration into the EAC tissue. Our hypothesis is further supported by the 302
report showing IL-33 induces murine basophil expansion 29, and by the report showing 303
diminished numbers of basophils in the nasal tissue of RW-induced rhinitis model using 304
IL-33 KO mice.23 305
Differential upregulation of ccl5 (encoding RANTES) mRNA but not of ccl11(encoding 306
eotaxin) mRNA in the RW-EAC models in IL-33 KO mice suggested IL-33 could 307
induce ccl5 expression (Figure 4). These results were consistent with the report of 308
Haenuki et al., showing IL-33 stimulation upregulate CCL5 protein expression but 309
showing marginal effects for CCL11 expression in mast cells and in basophils.23 In 310
RW-EAC model, we observed infiltration of basophils (Figure 6) and mast cells (Figure 311
8) in the conjunctival tissue, therefore IL-33 activated mast cells and basophils may 312
play some roles for eosinophil infiltration in the RW-EAC model through the effect of 313
CCL5, a well-known eosinophil chemoattractant.30 314
The results of immunohistochemical staining revealed that positive IL-33 315
immunostaining not only in the cell nuclei of the conjunctival epithelium as shown in 316
previous reports 11 23 but also in the infiltrating cells located in substantia propria 317

Asada et al. 20
(Figure 7). In Figure 8, we also showed that some of the IL-33 positive cells are double 318
positive with macrophage marker (F4/80) or with mast cell marker (mMCP1). These 319
results are consistent with previous reports showing IL-33 is produced by mast cells31 320
and activated macrophages.32 Taken together, we concluded that not only epithelial cells 321
but also macrophages and mast cells are the source of IL-33 in RW-EAC models. 322
It also should be noted that there are no apparent IL-33 positive staining in the vascular 323
endothelium of conjunctival tissue obtained from RW-EAC models (Figure 7), whereas 324
in human conjunctival tissue obtained from AKC/VKC patients, the vascular 325
endothelium (especially high endothelial venules) was immunopositive for IL-33.4 The 326
limitation of present study is feasibility of RW-induced allergic conjunctivitis as a 327
model for severe human chronic allergic conjunctivitis in which exposure to multiple, 328
and divergent allergens (i.e. HDM, pollens, animal derived antigens) caused severe 329
chronic allergic conjunctivitis, and also the differences of IL-33 expression between 330
human and mice tissue, especially at vascular endothelium. 331
In conclusion, among the epithelial cell derived type 2 initiating cytokines, we showed 332
indispensable roles of IL-33 for the pathophysiology of RW-EAC. Targeting IL-33 333
signaling cascades at ocular surface using the decoy receptor (soluble ST2) for IL-33 334
will be one of the promising therapeutic targets for ocular allergic diseases. 335

Asada et al. 21
Figure 1 336
337
338
339
340
341
342
343
344
345
Asada et al. 22
Figure 2 346
347
348
349
350
351
Figure 3 352
353
354
355

Asada et al. 23
Figure 4 356
Rel
ativ
e IL
-4 m
RN
A e
xpre
ssio
n
0
0.5
1
1.5
2
2.5
3
PBS RW
□Wild ■IL‐33KO
0
0.5
1
1.5
2
2.5
3
PBS RW
Rel
ativ
e C
CL
5 m
RN
A e
xpre
ssio
n
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
PBS RW
Rel
ativ
e C
CL
11 m
RN
A e
xpre
ssio
n
**
**
0
0.5
1
1.5
2
2.5
PBS RW
Rel
ativ
e IL
-13
mR
NA
exp
ress
ion
** (fold)
0
0.5
1
1.5
2
2.5
3
3.5
PBS RW
Rel
ativ
e IL
5 m
RN
A e
xpre
ssio
n
**
* * *
*
357
358
359
360
361
362
363
364
365
366

Asada et al. 24
Figure 5 367

Asada et al. 25
0
1
2
3
4
5
6
PBS challenge 1hr 3hr 6hr 12hr
Rel
ativ
e IL
-33
mR
NA
exp
ress
ion
(fold)
Time after RW challenge
A *
0
100
200
300
400
500
600
700
800
900
PBS challenge 1hr 3hr 6hr 12hr
* B (pg/ml)
Time after RW challenge 368

Asada et al. 26
Figure 6 369
0
2
4
6
8
10
12
Wild 33KO
* C
Wild MCP8
A
IL-33 KO MCP8
B
370
371
Figure 7 372
373
374
375

Asada et al. 27
Figure 8 376
377
378
379
380
381
382
383
384
385
386

Asada et al. 28
Figure Legends 387
Figure 1. Total serum IgE measurement before and after RW-immunizations 388
Total serum IgE concentration was quantified using ELISA at day 0 (before 389
RW-immunization) and on day 30 (after RW eyedrop challenges). The data is a 390
representative data measured in triplicate using five mice per each group, and shown by 391
mean IgE concentration (ng/ml) ± standard deviation (SD). 392
Figure 2. Clinical evaluation of RW-EAC 393
Representative photographs of RW-EAC models, using wild type mice and IL-33 KO 394
mice challenged either by RW-PBS (up row) or by PBS alone (down row) were taken 395
20 minutes after the last eye drop challenge (A). Clinical score of RW-challenged EAC 396
models was shown (B). The data is a representative data of mean ± SD clinical score of 397
5 mice for each group. (*P<0.05, Mann Whitney’s U-test) 398
Figure 3. Eosinophil infiltration in the conjunctivae of RW-EAC 399
The eye of RW-EAC models were sampled 24 hours after last RW challenges and the 400
numbers of infiltrating eosinophils (arrows) in the substantia propria of the conjunctival 401
tissue were counted using Gimsa stained slides (A). The data is a representative data of 402
mean ± SD numbers of the infiltrated eosinophil per slides counting the conjunctivae of 403
5 mice for each group (B). (*P<0.05, Mann Whitney’s U-test) 404

Asada et al. 29
Figure 4. Quantification of cytokine mRNA expression in the conjunctivae of 405
RW-EAC models 406
Expression of inflammatory cytokines/chemokines (il4, il5, il13, ccl5, ccl11) mRNA 407
were quantified by realtime PCR. Relative mRNA expression was shown as fold 408
changes to mRNA expression levels of PBS-challenged conjunctival tissue. The data is 409
normalized by the expression of gapdh mRNA of the same cDNA samples. 410
Significantly elevated il4, il5, il13, ccl5 mRNA expression was observed in the RW 411
challenged conjunctivae of wild type mice compared to PBS challenged conjunctivae 412
(asterisks, *P<0.05). Attenuated il4, il5, il13, ccl5 mRNA expression was observed in 413
the RW-challenged conjunctivae of IL-33 KO mice compared to those of wild type mice 414
(double asterisks, **P<0.05). No differential expression was observed for ccl11 mRNA 415
expression. A representative data (mean fold expression ± SD) measured in triplicate 416
was shown. 417
Figure 5. Time course of IL-33 expression in the conjunctivae of RW-EAC models 418
Expression of il33 mRNA (A) and IL-33 protein (B) at various time points (1, 3, 6, 12 419
hour) after single RW eye drop challenge was quantified by realtime PCR and ELISA 420
analysis, respectively. Relative mRNA expression was shown as fold changes to mRNA 421
expression levels of PBS-challenged conjunctival tissue. The data is normalized by the 422

Asada et al. 30
expression of gapdh mRNA of the same cDNA samples. A representative data (mean 423
fold expression ± SD) measured in triplicate was shown (A). IL-33 concentrations in the 424
culture supernatants of ex vivo cultured conjunctivae tissues were quantified. A 425
representative data (mean fold expression ± SD) using three conjunctivae for each time 426
point measured in duplicate was shown (B). (*P<0.05, Mann Whitney’s U-test) 427
Figure 6. Mcp8 positive basophils in the conjunctivae of RW-EAC models 428
Higher numbers of mcp8 positive basophils (arrows) were found in the subepithelial 429
regions of RW-EAC models in the wild type mice (A) compared to IL-33 KO mice (B). 430
Original magnification x200. Numbers (mean ± SD) of the basophils per slides in the 431
conjunctivae of 8 mice for each group was shown (C). (*P<0.05, Mann Whitney’s 432
U-test) 433
Figure 7. IL-33 expression of in the conjunctivae of RW-EAC models 434
Anti-MBP and anti-IL-33 immunostaining of conjunctivae obtained from RW-EAC 435
models were shown. Massive infiltration of MBP positive eosinophils (green) were 436
observed in the substantia propria of RW-challenged EAC using wild type mice (top 437
left). On the other hand, sparse infiltration of eosinophils was observed in 438
PBS-challenged wild type mice and in RW-challenged IL-33 KO mice (middle and 439
bottom left). IL-33 protein expression (red) was observed in the conjunctival epithelial 440

Asada et al. 31
cells of wild type mice. The infiltrating cells of substantia propria at the vicinity of MBP 441
positive eosinophils (shown in the merged image) were also immunopositive for IL-33 442
(top right). No IL-33 positive cells were observed in the conjunctiva of IL-33 KO mice. 443
Original magnification x200. 444
Figure 8. Colocalization of F4/80 and IL-33, as well as mMCP1 and IL-33 in the 445
substantia propria of RW-EAC model. 446
Some of the IL-33 positive cells in the substantia propria are also immunopositive for 447
F4/80 (arrows) or mMCP1 (arrowheads). Original magnification x200. 448
449
450
451
452
453
454
455
456
457
458

Asada et al. 32
References 459
460
1. Pulendran B, Artis D. New paradigms in type 2 immunity. Science 2012; 461
337:431-5. 462
2. Galli SJ, Tsai M, Piliponsky AM. The development of allergic inflammation. 463
Nature 2008; 454:445-54. 464
3. Oliphant CJ, Barlow JL, McKenzie AN. Insights into the initiation of type 2 465
immune responses. Immunology 2011; 134:378-85. 466
4. Matsuda A, Okayama Y, Terai N, Yokoi N, Ebihara N, Tanioka H, et al. The role 467
of interleukin-33 in chronic allergic conjunctivitis. Invest Ophthalmol Vis Sci 468
2009; 50:4646-52. 469
5. Matsuda A, Ebihara N, Yokoi N, Kawasaki S, Tanioka H, Inatomi T, et al. 470
Functional role of thymic stromal lymphopoietin in chronic allergic 471
keratoconjunctivitis. Invest Ophthalmol Vis Sci 2010; 51:151-5. 472
6. Oboki K, Ohno T, Kajiwara N, Arae K, Morita H, Ishii A, et al. IL-33 is a crucial 473
amplifier of innate rather than acquired immunity. Proc Natl Acad Sci U S A; 474
107:18581-6. 475
7. Stolarski B, Kurowska-Stolarska M, Kewin P, Xu D, Liew FY. IL-33 476
exacerbates eosinophil-mediated airway inflammation. J Immunol 2010; 477
185:3472-80. 478
8. Eiwegger T, Akdis CA. IL-33 links tissue cells, dendritic cells and Th2 cell 479
development in a mouse model of asthma. Eur J Immunol 2011; 41:1535-8. 480
9. Morita H, Arae K, Ohno T, Kajiwara N, Oboki K, Matsuda A, et al. ST2 requires 481
Th2-, but not Th17-, type airway inflammation in epicutaneously antigen- 482
sensitized mice. Allergol Int 2012; 61:265-73. 483
10. Fukushima A, Yamaguchi T, Ishida W, Fukata K, Taniguchi T, Liu FT, et al. 484
Genetic background determines susceptibility to experimental immune-mediated 485
blepharoconjunctivitis: comparison of Balb/c and C57BL/6 mice. Exp Eye Res 486
2006; 82:210-8. 487
11. Matsuba-Kitamura S, Yoshimoto T, Yasuda K, Futatsugi-Yumikura S, Taki Y, 488
Muto T, et al. Contribution of IL-33 to induction and augmentation of 489
experimental allergic conjunctivitis. Int Immunol 2010; 22:479-89. 490
12. Takai T. TSLP expression: cellular sources, triggers, and regulatory mechanisms. 491
Allergol Int 2012; 61:3-17. 492

Asada et al. 33
13. Liu YJ, Soumelis V, Watanabe N, Ito T, Wang YH, Malefyt Rde W, et al. TSLP: 493
an epithelial cell cytokine that regulates T cell differentiation by conditioning 494
dendritic cell maturation. Annu Rev Immunol 2007; 25:193-219. 495
14. Zheng X, Ma P, de Paiva CS, Cunningham MA, Hwang CS, Pflugfelder SC, et 496
al. TSLP and downstream molecules in experimental mouse allergic 497
conjunctivitis. Invest Ophthalmol Vis Sci 2010; 51:3076-82. 498
15. Fort MM, Cheung J, Yen D, Li J, Zurawski SM, Lo S, et al. IL-25 induces IL-4, 499
IL-5, and IL-13 and Th2-associated pathologies in vivo. Immunity 2001; 500
15:985-95. 501
16. Angkasekwinai P, Park H, Wang YH, Wang YH, Chang SH, Corry DB, et al. 502
Interleukin 25 promotes the initiation of proallergic type 2 responses. J Exp Med 503
2007; 204:1509-17. 504
17. Ishii A, Oboki K, Nambu A, Morita H, Ohno T, Kajiwara N, et al. Development 505
of IL-17-mediated delayed-type hypersensitivity is not affected by 506
down-regulation of IL-25 expression. Allergol Int 2010; 59:399-408. 507
18. Carpino N, Thierfelder WE, Chang MS, Saris C, Turner SJ, Ziegler SF, et al. 508
Absence of an essential role for thymic stromal lymphopoietin receptor in 509
murine B-cell development. Mol Cell Biol 2004; 24:2584-92. 510
19. Magone MT, Chan CC, Rizzo LV, Kozhich AT, Whitcup SM. A novel murine 511
model of allergic conjunctivitis. Clin Immunol Immunopathol 1998; 87:75-84. 512
20. Ishida W, Fukuda K, Kajisako M, Sumi T, Matsuda H, Yagita H, et al. B and T 513
lymphocyte attenuator regulates the development of antigen-induced 514
experimental conjunctivitis. Graefes Arch Clin Exp Ophthalmol 2012; 515
250:289-95. 516
21. Ugajin T, Kojima T, Mukai K, Obata K, Kawano Y, Minegishi Y, et al. Basophils 517
preferentially express mouse Mast Cell Protease 11 among the mast cell tryptase 518
family in contrast to mast cells. J Leukoc Biol 2009; 86:1417-25. 519
22. Larson KA, Horton MA, Madden BJ, Gleich GJ, Lee NA, Lee JJ. The 520
identification and cloning of a murine major basic protein gene expressed in 521
eosinophils. J Immunol 1995; 155:3002-12. 522
23. Haenuki Y, Matsushita K, Futatsugi-Yumikura S, Ishii KJ, Kawagoe T, Imoto Y, 523
et al. A critical role of IL-33 in experimental allergic rhinitis. J Allergy Clin 524
Immunol 2012; 130:184-94 e11. 525
24. Chu DK, Llop-Guevara A, Walker TD, Flader K, Goncharova S, Boudreau JE, et 526
al. IL-33, but not thymic stromal lymphopoietin or IL-25, is central to mite and 527
peanut allergic sensitization. J Allergy Clin Immunol 2013; 131:187-200 e1-8. 528

Asada et al. 34
25. Shahabuddin S, Ponath P, Schleimer RP. Migration of eosinophils across 529
endothelial cell monolayers: interactions among IL-5, endothelial-activating 530
cytokines, and C-C chemokines. J Immunol 2000; 164:3847-54. 531
26. Iijima K, Kobayashi T, Hara K, Kephart GM, Ziegler SF, McKenzie AN, et al. 532
IL-33 and thymic stromal lymphopoietin mediate immune pathology in response 533
to chronic airborne allergen exposure. J Immunol 2014; 193:1549-59. 534
27. Paul WE, Zhu J. How are T(H)2-type immune responses initiated and amplified? 535
Nat Rev Immunol 2010; 10:225-35. 536
28. Karasuyama H, Mukai K, Obata K, Tsujimura Y, Wada T. Nonredundant roles of 537
basophils in immunity. Annu Rev Immunol 2011; 29:45-69. 538
29. Schneider E, Petit-Bertron AF, Bricard R, Levasseur M, Ramadan A, Girard JP, 539
et al. IL-33 activates unprimed murine basophils directly in vitro and induces 540
their in vivo expansion indirectly by promoting hematopoietic growth factor 541
production. J Immunol 2009; 183:3591-7. 542
30. Schroder JM, Kameyoshi Y, Christophers E. RANTES, a novel 543
eosinophil-chemotactic cytokine. Ann N Y Acad Sci 1994; 725:91-103. 544
31. Hsu CL, Neilsen CV, Bryce PJ. IL-33 is produced by mast cells and regulates 545
IgE-dependent inflammation. PLoS One 2010; 5:e11944. 546
32. Talabot-Ayer D, Calo N, Vigne S, Lamacchia C, Gabay C, Palmer G. The mouse 547
interleukin (Il)33 gene is expressed in a cell type- and stimulus-dependent 548
manner from two alternative promoters. J Leukoc Biol 2012; 91:119-25. 549
550
551
552
553
554
555
556

Asada et al. 35
Supplementary Figure 1 557
0
0.5
1
1.5
2
2.5
PBS RW
Rel
ativ
e C
CL
11 m
RN
A e
xpre
ssio
n
Rel
ativ
e C
CL
5 m
RN
A e
xpre
ssio
n R
elat
ive
IL-4
mR
NA
exp
ress
ion
0
0.5
1
1.5
2
2.5
3
3.5
PBS RW
0
0.5
1
1.5
2
2.5
3
3.5
4
PBS RW
□Wild ■TSLPRKO
Rel
ativ
e IL
-13
mR
NA
exp
ress
ion
0
1
2
3
4
5
6
PBS RW
**
**
0
0.5
1
1.5
2
2.5
3
3.5
PBS RW
Rel
ativ
e IL
5 m
RN
A e
xpre
ssio
n
*
558
Supplementary Figure 2 559
□Wild ■IL‐25KO
Rel
ativ
e C
CL
11 m
RN
A e
xpre
ssio
n
Rel
ativ
e C
CL
5 m
RN
A e
xpre
ssio
n R
elat
ive
IL-4
mR
NA
exp
ress
ion
0
0.5
1
1.5
2
2.5
3
3.5
PBS RW
Rel
ativ
e IL
-13
mR
NA
exp
ress
ion
0
0.5
1
1.5
2
2.5
3
3.5
4
PBS RW
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
PBS RW 0
0.5
1
1.5
2
2.5
PBS RW
0
0.5
1
1.5
2
2.5
3
3.5
PBS RW
Rel
ativ
e IL
5 m
RN
A e
xpre
ssio
n
*
560

Asada et al. 36
Legends for Supplementary Figure 1 561
Expression of inflammatory cytokines/chemokines (il4, il5, il13, ccl5, ccl11) mRNA 562
were quantified by realtime PCR analysis. Relative mRNA expression was shown as 563
fold changes to mRNA expression levels of PBS-challenged conjunctival tissue. The 564
data is normalized by the expression of gapdh mRNA of the same cDNA samples. 565
Attenuated il5 mRNA expression (asterisk, *P<0.05) and augmented il13 mRNA 566
expression (double asterisks, **P<0.05) was observed in the RW challenged 567
conjunctivae of TSLPR KO mice compared to those of wild type mice. A representative 568
data (mean fold expression ± SD) measured in triplicate was shown. 569
Legends for Supplementary Figure 2 570
Expression of inflammatory cytokines/chemokines (il4, il5, il13, ccl5, ccl11) mRNA 571
were quantified by realtime PCR analysis. Relative mRNA expression was shown as 572
fold changes to mRNA expression levels of PBS-challenged conjunctival tissue. The 573
data is normalized by the expression of gapdh mRNA of the same cDNA samples. 574
Attenuated il5 mRNA expression (asterisk, *P<0.05) was observed in the RW 575
challenged conjunctivae of IL-25 KO mice compared to those of wild type mice. A 576
representative data (mean fold expression ± SD) measured in triplicate was shown. 577
578