Embed Size (px)
Citation preview

B R A I N R E S E A R C H 1 2 4 2 ( 2 0 0 8 ) 3 7 – 4 4
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
Temporo-nasal asymmetry in multisensory integrationmediated by the Superior Colliculus
Caterina Bertinia,b, Fabrizio Leoa,b, Elisabetta Làdavasa,b,⁎aDipartimento di Psicologia, Università di Bologna, Bologna, ItalybCentro Studi e Ricerche in Neuroscienze Cognitive, Polo Didattico e Scientifico, Cesena, Italy
A R T I C L E I N F O
⁎ Corresponding author. Dipartimento di PsicE-mail address: [email protected]: A, Auditory; V, Visual; AV,
Model Inequality
0006-8993/$ – see front matter © 2008 Elsevidoi:10.1016/j.brainres.2008.03.087
A B S T R A C T
Article history:Accepted 19 March 2008Available online 12 April 2008
Temporo-nasal asymmetry in visual responses has been observed in many behaviouralstudies. These observations have typically been attributed to the anatomical asymmetry offibres projecting to the Superior Colliculus (SC), even though this attribution is debated. Thepresent study investigates temporo-nasal asymmetry in multisensory integration, and, byexploiting the absence of S-cone input to the SC, measures a behavioural responsedependent strictly on the activity of the SC itself. We used a redundant signal paradigm forsimple reaction times, with visual stimuli (red or purple) presented in either the temporal orthe nasal hemifield. Participants responded more quickly to concurrent audio-visual (AV)stimuli than to either an auditory or a visual stimulus alone, an established phenomenonknown as the Redundant Target Effect (RTE). The nature of this effect was dependent on thecolour of the visual stimuli, suggesting its modulation by collicular circuits. When spatially-coincident audio-visual stimuli were visible to the SC (i.e. red stimuli), the RTE depended ona neural coactivation mechanism, suggesting an integration of multisensory information.When using stimuli invisible to the SC (i.e. purple stimuli), the RTE depended only on asimple statistical facilitation effect, in which the two sensory stimuli were processed byindependent channels. Finally, we demonstrate that the multisensory integration effectwas stronger for stimuli presented to the temporal hemifield than to the nasal hemifield.Taken together, these findings suggested thatmultisensory stimulation can be differentiallyeffective depending on specific stimulus parameters.
© 2008 Elsevier B.V. All rights reserved.
Keywords:Temporo-nasal asymmetryMultisensory integrationSuperior ColliculusVisual orienting response
1. Introduction
The existence of a biased representation favouring thetemporal hemifield in the Superior Colliculus (SC) has beeninvestigated in several behavioural studies and its functionalrelevance in humans has been shown in a wide range of visualorienting behaviours. For instance, adult subjects show a biasto saccade towards temporal visual stimuli under conditions of
ologia, Viale Berti Pichat,it (E. Làdavas).Audio-visual; SP, Same P
er B.V. All rights reserved
bilateral, simultaneous stimulation (Posner and Cohen, 1980).Additionally, a temporo-nasal asymmetry has been found incovert orienting behaviour (Rafal et al., 1991), in unconsciouspriming effects (Ansorge, 2003) and in more subtle measuressuch as saccadic latencies (Kristjansson et al., 2004). A pro-nounced bias in orienting saccades towards stimuli in thetemporal hemifield versus the nasal hemifield has also beenshown in newborns (Lewis and Maurer, 1992). Moreover,
5, 40127 Bologna, Italy. Fax: +39 051 209 1844.
osition; DP, Different Position; T, Temporal; N, Nasal; RMI, Race
.

1 Note: To qualify the nature of the RTE (Redundant TargetEffect: RTs to bi-sensory stimuli are faster, on average, thanresponses to unisensory ones; Todd, 1912), Miller (Miller, 1982)devised a mathematical method (race inequality test) that, usingthe cumulative frequency distribution (CFD) of RTs in the singleversus double-target condition, sets an upper limit for the CDFwhen redundant target are presented (Miller, 1982, 1986): p(tb t0 |AV)≤p(tb t0 |A)+p(tb t0 |V). If this limit is violated, then a statisticalfacilitation is no longer tenable, and the RTE can be attributed to aneural coactivation mechanism.
38 B R A I N R E S E A R C H 1 2 4 2 ( 2 0 0 8 ) 3 7 – 4 4
hemianopic patients demonstrate a temporal hemifieldadvantage in both the oculomotor distractor effect (Rafalet al., 1990) and in a forced choice localization paradigm(Dodds et al., 2002). Such advantages in the representation ofthe temporal hemifield have been attributed to an asymmetryin projections from the hemiretinae to the Superior Colliculus;this anatomical argument is based on the observation that thebehavioural asymmetries are present only in subjects whoseretinotectal pathways are fully functional, but whose genicu-lostriate pathways are either not yet mature, as in infants(Johnson, 1990), or are disrupted, as in hemianopic patients.
Neuroanatomical studies on cats support this anatomicalaccount; it has been shown that the nasal hemiretina (pro-cessing the temporal hemifield) has a strongerdirect input to theSuperior Colliculus than the temporal hemiretina (processingthe nasal hemifield) (Sherman, 1974). However, inmonkeys, thepresence of this asymmetry exclusively in retinotectal projec-tions has been questioned (Hubel et al., 1975): in a study on theprimate visual system, this temporo-nasal asymmetry has beenobserved for both the projections from the retina to the SuperiorColliculus and fromthe retina to thedLGN (Williams et al., 1995).
Recently, an fMRI study (Sylvester et al., 2007) testing thisasymmetry directly in the human brain revealed a strongeractivation of the SC in response to stimuli presented in thetemporal hemifield than in the nasal hemifield, whereas nodifferential activity was observed in the lateral geniculatenucleus (LGN) or in the retinotopic cortical areas V1 and V3.Although these results do not distinguish between structuralasymmetries (i.e.moreneurons responding to temporal stimuli)and functional ones (i.e. larger gainassociatedwith responses totemporal stimuli), they constitute a direct physiological demon-stration of the neural correlates of characteristic behaviouraltemporo-nasal asymmetries.
Theaimof thepresent study is to investigate theproperties ofthe collicular temporo-nasal asymmetry by using a newbehavioural method that directly tests a Superior Colliculusdependent response, namely multisensory integration. The roleof SC in mediating multisensory integration has been exten-sively investigated in animals using neurophysiological record-ings (Stein and Meredith, 1993). These studies showed thatmultisensory neurons in the deep layers of the SC typically havespatially-coincident receptive fields, and that they exhibitmulti-sensory response enhancement (i.e. the multisensory responseexceeds the most effective unisensory component response) tomultimodal stimulation delivered in close spatial and temporalproximity (Stein and Meredith, 1993; Wallace et al., 1992).
To provide evidence of temporo-nasal asymmetry in multi-sensory integration, we exploited the fact that neurophysiolo-gical studies have consistently reported the lack of retinal inputto the SC from short-wave sensitive S-cones (de Monasterio,1978; Marrocco and Li, 1977; Schiller and Malpeli, 1977; Sumneret al., 2002).
The argument that the SC receives no S-cone input isbolstered by evidence that S-cones signals reach the koniocel-lular layers of the lateral geniculate nucleus, from where theyproject to layers 2 and 3 of the striate cortex (Dacey and Lee,1994;White et al., 1998).Moreover, S-cones stimuli are invisibleto the magnocellular pathway, which rules out the possibilityof feedback projections running from the visual cortex to theSC.
A recent study by Leo et al. (2007) took advantage of theSuperior Colliculus' insensitivity to S-cone activity in order todemonstrate the pivotal role of the SC in mediating multi-sensory integration when a spatial orienting response isrequired. In a redundant signal paradigm for simple reactiontime, the researchers presented peripheral monochromatic red(i.e. long-wavelength stimulus, visible to the SC) or purple (i.e.short-wavelength S-cone dependent stimulus, invisible to theSC) visual stimuli with concurrent acoustic stimuli in either thesame or different locations within the visual field. Resultsprovided evidence that RTs to spatially-coincident audio-visual(AV) stimuli were significantly faster than RTs to spatially-disparate AV stimuli when red stimuli were used, but not whenpurple stimuli were presented. Moreover, a second analysisbased on Miller's race inequality model (RMI; Miller, 1982)showed that the RTs to red (but not purple) spatially-coincidentAV stimuli could be ascribed to a neural coactivation mechan-ism, suggesting in this way amultisensory integration effect. Inother words, the effect could not be explained exclusively as astatistical facilitation1.
Given that multisensory spatial integration requires theactivation of the retinotectal pathway, different behaviouraleffects should be expected when presenting stimuli in thetemporal hemifield than in the nasal, a result of the temporo-nasal asymmetry present in the SC. To test this hypothesis,subjects were presented with peripheral red (i.e. visible to theSC) or purple (i.e. invisible to the SC) visual stimuli, eitherwithor without a spatial disparity to concurrent auditory stimuli,in a redundant signal paradigm for simple reaction time.Subjects performed the task monocularly, which created asimple way to isolate the temporal and nasal hemifields:when patching a single eye, the contralateral hemifieldbecomes temporal and the ipsilateral field nasal. As predictedby Leo et al.'s study, an effect of spatial congruency (RTsshorter to spatially-coincident AV stimuli than to spatially-disparate AV stimuli) was expected only when red stimuliwere presented; moreover, multisensory spatial integration(i.e. violation of the race model) should occur only with AVspatially-coincident stimuli visible to the SC. In accordancewith temporo-nasal asymmetry, the magnitude of the viola-tion of the race model was expected to be stronger for stimulipresented in the temporal rather than in the nasal hemifield.Conversely, no effect of spatial congruency was expectedwhen purple stimuli (i.e. invisible to the SC) were presented.
2. Results
Participants responded to 1% of catch trials. RTs faster than140 ms and slower than 650 ms were considered anticipations

Table 1 – Redundancy gain (RG)
Red Purple
Stimulus M SD RG Stimulus M SD RG
T V 335 36 T V 343 29N V 335 31 N V 346 30T A 304 76N A 308 79T SP-AV 259 36 62 T SP-AV 265 39 60N SP-AV 260 37 59 N SP-AV 267 40 58T DP-AV 267 39 52 T DP-AV 269 41 55N DP-AV 267 40 55 N DP-AV 269 40 58
Mean RTs (M) and standard deviations (SD) in milliseconds foreach stimulation condition, separately for the red visual stimulusand purple visual stimulus. RG is the difference betweenmean RTsin unimodal conditions and mean RTs in bimodal conditions.Auditory stimuli are labelled as temporal (T) and nasal (N): whensubjects perform the task with the right eye patched, temporalauditory (TA) refers to an auditory stimulus presented in the lefthemifield,whereas nasal auditory (NA) refers to an auditory stimuluspresented in the right hemifield. On the contrary, when subjectsperform the task with the left eye patched, temporal auditory (TA)refers to an auditory stimulus presented in the right hemifield,whereasnasal auditory (NA) refers toanauditory stimuli presented inthe left hemifield. Note that the same RT data from unisensoryauditory (T A and N A) conditions were used to calculate RG forred and purple visual stimuli. Abbreviations: T V=temporal visual,N V=nasal visual, T A=temporal auditory, N A=nasal auditory,T SP-AV=temporal spatially-coincident bimodal condition,N SP-AV=nasal spatially-coincident bimodal condition, T DP-AV=temporalspatially-disparate bimodal condition, N DP-AV=nasal spatially-disparate bimodal condition.
Fig. 1 – Mean reaction times. Mean RTs (S.E.M. indicated) foreach experimental condition (V=visual alone; A=auditoryalone; SP-AV=same-position audio-visual condition;DP-AV=different-position audio-visual condition). Grey barsrepresent the conditions in which the visual stimulus wasred; white bar represents the auditory alone condition; blackbars represent the conditions in which the visual stimuluswas purple.
39B R A I N R E S E A R C H 1 2 4 2 ( 2 0 0 8 ) 3 7 – 4 4
and delayed responses respectively, and were discarded (b1%of trials). Given that no differences between eyes were foundin a preliminary analysis, reaction time data were analysedwith a three-way ANOVA with Colour (red versus purplestimuli), Hemifield (temporal versus nasal) and Condition(unimodal visual, unimodal auditory, bimodal SP-AV, bimodalDP-AV) as within-subject factors.
Pairwise comparisons were conducted using the Newman–Keuls test. Overall, we found that responses to long-wave-length red stimuli (292 ms) were faster than responses toshort-wavelength purple stimuli (296.4 ms), evidenced by thesignificant main effect of Colour [F (1,21)=16.7, pb .0006].
Wealso foundasignificantmaineffect ofCondition [F (3,63)=41.6, pb .000001]. Responses to unimodal visual stimuli (340ms)were significantly slower than responses to AV stimuli both inSP-AV condition (263 ms; pb .0002) and in the DP-AV condition(268 ms; pb .0002). Likewise, RTs in the unimodal auditorycondition (306ms)were significantlyslower thanRTs inboth theSP-AV condition (pb .0002) and DP-AV condition (pb .0002).Finally, the redundancy gain (i.e., the RT difference betweensingleanddoublestimulation)was similar inbothhemifields forred and purple stimuli (see Table 1).
Moreover, we observed a significant interaction betweenColour and Condition [F (3, 63)=7.8, pb .0002]. That is, when thevisual stimulus was red, RTs in the SP-AV condition werefaster than RTs in the DP-AV condition (260 and 267 ms,respectively, pb .0002), whereas, when the visual stimulus waspurple, there was no significant difference between SP-AV(266 ms) and DP-AV (269 ms) conditions. In addition,
responses were significantly faster in the red SP-AV conditionthan in the purple SP-AV condition (pb .0003) (see Fig. 1).
RT data were then analysed with Miller's race inequalitytest to determine whether the observed redundancy gainscould be explained in terms of either a race or a neuralcoactivation model (Miller, 1982; Ulrich et al., 2007). Asignificant violation of race inequality was found only forred stimuli, in the spatially-coincident AV condition, in boththe temporal and nasal hemifields (see Figs. 2 and 3). The greyrectangles in Fig. 3 indicate the percentiles in which theviolation was significantly different from zero, as assessed bya t-test. Therefore, the RTE with red SP-AV stimuli violated therace inequality, and can therefore best be explained by neuralcoactivation. On the other hand, with the red DP-AV and withthe purple (either SP-AV or DP-AV) stimuli, there was no raceinequality violation, and therefore the RTE in these cases isbest explained by statistical facilitation.
Given that a violation of the race inequality occurred withred SP-AV stimuli both in the temporal and nasal hemifields,we evaluated whether the magnitude of violation wassignificantly greater in the temporal compared to the nasalcondition. To this aim, we computed the differences betweenthe observed cumulative distribution functions (CDFs) and theCDFs predicted by the RMI for both the conditions (T SP-AVand N SP-AV), for all percentiles included in the violationareas, and then carried out a nonparametric inferentialanalysis (Wilcoxon matched pairs test). The result showedthat the magnitude of the violation was reliably greater in thetemporal SP-AV (median=14.63) than in the nasal SP-AVcondition (median=10.83; z=2.80, pb .006) (see Fig. 4).
Moreover, we computed the means of the differences be-tween the observed and the predicted CDFs for each condition

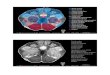
Fig. 2 – Cumulative reaction times. Observed cumulative reaction time (RT) distribution functions (CDFs) in the fourspatially-coincident bimodal conditions and race model predictions from the respective RT CDFs to unimodal target stimuli.The two panels in the first column (A and C) show the conditions with red audio-visual stimuli presented in either the temporal(A) or nasal (C) visual field. The two panels in the second column (B and D) show the conditions with audio-visual purple visualstimuli in either the temporal (B) or nasal (D) visual field. Both the red spatially-coincident CDF presented in the temporal(T SP-AV) and nasal (N SP-AV) hemifield significantly outgrow the summed unimodal distributions, thus violating the racemodel. Asterisks are plotted for the percentiles where this violation is significant.
40 B R A I N R E S E A R C H 1 2 4 2 ( 2 0 0 8 ) 3 7 – 4 4
(T SP-AV and N SP-AV; 13 and 9.83, respectively). The ratiobetween the mean in the temporal condition and the mean inthe nasal condition (1.32) was compared to the neurophysio-logic naso-temporal ratio (1.57) reported by Williams et al.(1995). The result of a Fisher test (exact, 2-tail) demonstratedthat the two ratios did not differ significantly (p=.403).
3. Discussion
Neurophysiological evidence in animals and humans hasconsistently highlighted the critical involvement of the Super-ior Colliculus in visual orienting behaviour. Previous findingsin monkeys have demonstrated that preparation of saccadesand covert shifts of attention share a commonneural network,in which the SC may play a pivotal role by determiningorienting behaviour towards peripheral visual locations (Kus-tov and Robinson, 1996; Ignashchenkova et al., 2004).
A recent behavioural study of humans (Leo et al., 2007)provides further evidence of this collicular involvement invisual orienting responses. This study exploited the lack of S-cones inputs to the SC (Marrocco and Li, 1977; Schiller andMalpeli, 1977; deMonasterio, 1978; Sumner et al., 2002) and therelevance of this neural structure in the integration of differentsensory inputs (Stein and Meredith, 1993). The results of theLeo et al.'s study showed that, when subjects were presentedwithperipheral visual stimuli,multisensory integration effectsoccurred only with stimuli visible to the SC; this importantresult suggests that the activity of the SC is absolutelynecessary when a spatial orienting response is required.
The present study investigates temporo-nasal asymmetryin the SC, taking advantage of the behavioural techniqueproposed in Leo et al.'s study, which constitutes a direct wayto study SC-mediated responses. To this end, a redundanttarget paradigm for simple reaction times was used, and AVstimuli were presented with either spatial coincidence or at aspatial disparity. The visual stimuli themselves could be either

Fig. 3 – Violation of the race inequality. Violation of the raceinequality test for the four red (panel A) and purple (panelB) bimodal AV conditions of the experiment. The greyrectangles in the red condition mark the areas in which theviolation is significantly different from zero (T SP-AV; NSP-AV), as assessed by one-sample t-test.
Fig. 4 – Hemifield violation of the race inequality.Comparison of the magnitude of violation of red temporalsame-position audio-visual condition (T SP-AV) and rednasal same-position audio-visual condition (N SP-AV), for allpercentiles included in the violation areas. Filled linerepresents the median of T SP-AV distribution. Dashed linerepresents the median of N SP-AV distribution.
41B R A I N R E S E A R C H 1 2 4 2 ( 2 0 0 8 ) 3 7 – 4 4
long-wavelength red stimuli (i.e. visible to the SC) or short-wavelength purple stimuli (i.e. invisible to the SC).
First, our results revealed an effect of spatial congruency onRTs depending on the colour of the visual stimulus: RTs tospatially-coincident AV stimuli were found to be significantlyfaster than RTs to spatially-disparate AV stimuli only with redvisual stimuli (i.e. visible to the SC), but not when purple visualstimuli (i.e. invisible to the SC) were presented.
In order to disambiguate the mechanism underlying thereduction of RTs to double stimuli, data were analysedaccordingly to Miller's race inequality test (Miller, 1982,1986). Results showed a significant race model violation forspatially-coincident AV stimuli only when a red visual sti-mulus was used; the effect was present in both temporal andnasal hemifields. This significant violation, although it doesnot rule out the possibility of a probabilistic race betweensignals transmitted along different channels (statistical facil-itation; Colonius and Diederich, 2006), suggests that the
reduction of RTs to double stimuli may be due to a neuralcoactivation mechanism resulting from the convergence ofthe incoming signals onto one or more brain centres (multi-sensory integration).
Taken together, the present findings are consistent withthe results presented in Leo et al.'s study and reveal thepresence of a multisensory integration effect in a taskrequiring a visual orientation response, only when the visualinformation reaches the SC and when the different sensorymodalities originate from the same position in the space.
To characterize the temporo-nasal asymmetry in multi-sensory integration, a further analysis on the magnitude ofviolation of the race model was carried out with the resultsobtained in the temporal and nasal conditions. This analysisdemonstrated a significantly greater violation of the raceinequality in the temporal hemifield than in thenasal hemifieldwhen a red visual stimulus was presented with a concurrentacoustic stimulus in the same spatial position. Moreover, theratio between the magnitude of the violation in both theconditions seems to reflect the ratio of the fibres projecting tothe SC observed at neural level: an electrophysiological studyexamining nasal and temporal retinal ganglion cells, retro-gradely labelled in the SC of four macaquemonkeys, revealed amean naso-temporal ratio in the collicular projection of 1.57(Williams et al., 1995). The present study describes an asym-metry inmultisensory effects that appears to be linearly relatedto the asymmetry of the afferent fibres to the SC.
These findings corroborate the notion that an anatomicalbias favours the temporal hemifield in the SC. They areconsistent with the results of a recent fMRI study revealinggreater SC responses to visual stimuli in the temporalhemifield than in the nasal hemifield; no such bias wasapparent in geniculostriate pathway structures such as LGNand cortical areas V1 and V3 (Sylvester et al., 2007).
The notion that the stimulation of the temporal hemifieldleads to a greater multisensory response enhancement,

42 B R A I N R E S E A R C H 1 2 4 2 ( 2 0 0 8 ) 3 7 – 4 4
comparing to the stimulation of the nasal hemifield, and therelevance of SC in visual orienting behaviour, suggest anadaptive function of this midbrain structure. Both the abilityto integrate different sensory modalities into a unified multi-sensory map of the external environment, and the bias inorienting towards stimuli in the periphery of the visual field,might have evolved because of significant survival advantagesin rapidly orienting towards salient peripheral stimuli.
The results of the present study have several operationalimplications. Recent clinical studies on hemianopic patients(Bolognini et al., 2005) have demonstrated that multisensorystimulation can systematically activate the SC and, therefore,can constitute a therapeutic tool in the recovery from spatialrepresentational disorders. From this perspective, the ob-served temporo-nasal asymmetry lends support to the sug-gestion that stimulating the temporal hemifield shouldimprove the therapeutic efficacy of rehabilitative treatmentsbased on multisensory integration.
To sumup, thepresent studyprovides evidenceof a temporo-nasal asymmetry in a multisensory integration effect mediatedby the SC. By taking advantage of the absence of retinal input tothe SC from short-wave sensitive S-cones, we have shown thepresence of a multisensory spatial integration effect that occursonly with visual stimuli visible to the SC. In addition, we havedemonstrated that this effect is stronger for stimuli presented tothe temporal hemifield than to the nasal hemifield.
4. Experimental procedures
4.1. Participants
Eleven healthy subjects (age range: 21–31 years; three malesand eight females) took part in the experiment. All had normalhearing and normal or corrected-to-normal vision and were
Fig. 5 – Schematic diagram of the experimental setup. Panel A rexperiment. Panel B shows schematic, top-down views of a subjehere is a long-wavelength stimulus (red). In the experiment, loudbelow the guide boxes. Diagrams not to scale.
naive as to the purpose of the experiment. Participants re-ceived course credit for their participation and gave informedconsent prior to beginning. The experimental procedures wereapproved by the Ethical Committee of the Department ofPsychology, University of Bologna. The experiment was car-ried out according to the principles laid out in the 1964Declaration of Helsinki.
4.2. Stimuli and procedures
Subjects sat in a completely dark room in front of a SamsungSyncMaster 153B 17 inch PCmonitor at a distance of 57 cm. Allstimuli were generated by a PC with MATLAB 6.1 and PSY-CHTOOLBOX 2.50 (Brainard, 1997; Pelli, 1997). Subjects wererequired to hold constant fixation on a central white dotagainst a black background (0.05 cd/m2). The experimenter,standing behind themonitor and facing the subject, initializedeach trial onlywhen subject's eyewere at fixation. In each trial(see Fig. 5A for the sequence of events in a trial) the fixationpoint lasted 500ms and then, after an interval whose durationwas randomised in a 100–1100 ms time window, two 3°×3°square “guide boxes” were displayed 6° to the right and to theleft of the fixation point with an exposure duration rangingbetween 750 and 1000ms. Each boxwasmade up of nine smallsquares changing luminance every 67 ms to a value drawnrandomly from the range 4.05–6.75 cd/m2. This luminancenoise ensured that the colour changes described below couldbe detected only by a chromatic channel (Birch et al., 1992; Leoet al., 2007; Mollon, 1982; Savazzi and Marzi, 2004; Sumneret al., 2002).
The visual stimuli were 1°×1° squares randomly presented atthe centre of one of the two guide boxes, in the last 100 ms ofguide boxes' presentation. The visual stimuli could be eithershort-wavelength (purple; colorimetric values: x=0.1826,y=0.0869) or long-wavelength (red; colorimetric values: x=
epresents a schematic diagram of a typical trial in thect, below the experimental displays. The visual target shownspeakers (not reported in the figure) were placed immediately

43B R A I N R E S E A R C H 1 2 4 2 ( 2 0 0 8 ) 3 7 – 4 4
0.6194, y=0.3455). For all stimuli, exposure duration was 100 msand the luminance was 5.4 cd/m2. Chromaticities and lumi-nanceswere calculated from the spectra of each stimulus, whichwas measured directly with a Minolta chromameter CL-200.
The auditory stimuli comprised a 100 ms broadband burstof pure tones (2000 Hz), emitted by one of two piezoelectricloudspeakers (0.4 W, 8 W) located in front of the PC monitor,~4 cm below the visual stimuli. The intensity level of theauditory stimuli was 66.3 dB SPL.
Subjects completed two different sessions varying bymonocular condition (i.e. one with a patch covering the lefteye and one with a patch covering the right eye). Whenpatching the right eye, the left hemifield becomes temporaland the right nasal, whereas the reverse holdswith the left eyepatched instead (see Fig. 5B). Sessions' order was counter-balanced across subjects.
There were 14 randomly intermingled stimulus conditions:
1) unimodal auditory condition: the auditory stimulus waspresented alone in the right or left hemifield;2) unimodal visual condition: the visual stimulus (purpleor red) was presented alone in the temporal (T V) or nasal(N V) hemifield;3) same-position audio-visual condition: the visual (purpleor red presented in the temporal or nasal hemifield, T SP-AV and N SP-AV, respectively) and the auditory stimuluswere presented simultaneously, in close spatial proximity;4) different-position audio-visual condition: the visual(purple or red, presented in the temporal or nasal hemi-field, T DP-AV and N DP-AV, respectively) and the auditorystimulus were presented simultaneously and in differentpositions (i.e., opposite hemifields).
Subjects were instructed to maintain fixation on the centralwhite dot, and to respond by pressing the space bar of thekeyboardwith the index finger of the dominant hand as quicklyas possible in response to any unimodal (visual or auditory) orbimodal stimulus.
A 5-min practice, run for each subject, preceded the ex-periment. Each session was subdivided into four blocks oftrials with an overall number of 747 presentations for eachparticipant. There were 40 trials for each type of stimuluscondition, and 187 catch trials in which only the guide boxeswere presented without the target stimuli. Catch trials wereintroduced to discourage the participant from responding tothe guide box rather than to the target stimulus.
4.3. Statistical analysis
For each subject, the average RTs were calculated and theirdistributions were recorded for each stimulus condition. Totest for the presence of an RTE, planned comparisons betweeneach of the unimodal stimulus conditions and the bimodalstimulus conditions were performed.
To determine if a statistical facilitation explanation for theRTE was tenable, or if a coactivation explanation of the RTEwas necessary, Miller's test of the race model was implemen-ted by using RMITest software (Miller, 1982; Ulrich et al., 2007).The program computes the estimated cumulative distributionfunctions (CDFs) of RT for each condition and the estimated
race model inequality bound. It also computes a t-test at eachpercentile to see whether the race model violation isstatistically significant. The difference between the cumula-tive probability for the double stimuli and the sum of thecumulative probability for the single stimuli indicates themagnitude of violation.
Acknowledgments
This work was supported by grants from MURST to EL. Wethank all the participants for their cooperation and MarioMazzocchi for his helpful insights in statistical analysis.
R E F E R E N C E S
Ansorge, U., 2003. Asymmetric influences of temporally vs. nasallypresented masked visual information: Evidence for collicularcontributions to nonconscious priming effects. Brain Cogn. 51,317–325.
Birch, J., Barbur, J.L., Harlow, A.J., 1992. New method based onrandom luminancemasking formeasuring isochromatic zonesusing high resolution colour displays. Ophthalmic Physiol. Opt.12, 133–136.
Bolognini, N., Rasi, F., Coccia, M., Làdavas, E., 2005. Visual searchimprovement in hemianopic patients after audio-visualstimulation. Brain 128, 2830–2842.
Brainard, D.H., 1997. The Psychophysics toolbox. Spat. Vis. 10,433–436.
Colonius, H., Diederich, A., 2006. The race model inequality:interpreting a geometric measure of the amount of violation.Psychol. Rev. 113, 148–154.
Dacey, D.M., Lee, B.B., 1994. The “blue on" opponent pathway inprimate retina originates from a distinct bistratified ganglioncell type. Nature 367, 731–735.
deMonasterio, F.M., 1978. Properties of ganglion cells with atypicalreceptive-field organisation in the retina of macaques.J. Neurophysiol. 41, 1435–1449.
Dodds, C., Machado, L., Rafal, R., Ro, T., 2002. A temporal/nasalasymmetry for blindsight in a localisation task: evidence forextrageniculate mediation. Neuroreport 13, 655–658.
Hubel, D.H., LeVay, S., Wiesel, T.N., 1975. Mode of termination ofretinotectal fibers in macaque monkey: an autoradiographicstudy. Brain Res. 96, 25–40.
Ignashchenkova, A., Dicke, P.W., Haarmeier, T., Thier, P., 2004.Neuron specific contribution of the superior colliculus to overtand covert shifts of attention. Nature Neurosci. 7, 56–64.
Johnson, M.H., 1990. Cortical maturation and the developmentof visual attention in early infancy. J. Cogn. Neurosci. 2, 81–95.
Kristjansson, A., Vandenbroucke, M.W., Driver, J., 2004. When prosbecome cons for anti- versus prosaccades: factors withopposite or common effects on different saccade types. Exp.Brain Res. 155, 231–244.
Kustov, A.A., Robinson, D.L., 1996. Shared neural control ofattentional shifts and eye movements. Nature 384, 74–77.
Leo, F., Bertini, C., Di Pellegrino, G., Làdavas, E., 2007. Multisensoryintegration for orienting responses in humans requires theactivation of the superior colliculus. Exp. Brain Res. 186, 66–77.
Lewis, T.L., Maurer, D., 1992. The development of the temporal andnasal visual fields during infancy. Vis. Res. 32, 903–911.
Marrocco, R.T., Li, R.H., 1977. Monkey superior colliculus:properties of single cells and their afferent inputs.J. Neurophysiol. 40, 844–860.
Miller, J., 1982. Divided attention: evidence for coactivation withredundant signals. Cogn. Psychol. 14, 247–279.

44 B R A I N R E S E A R C H 1 2 4 2 ( 2 0 0 8 ) 3 7 – 4 4
Miller, J., 1986. Time course of coactivation in bimodal dividedattention. Percept. Psychophys. 40, 331–343.
Mollon, J.D., 1982. Color vision. Ann. Rev. Psychol. 33, 41–85.Pelli, D.G., 1997. The Video Toolbox software for visual
psychophysics: transforming numbers into movies. Spat. Vis.10, 437–442.
Posner, M.I., Cohen, Y., 1980. Attention and the control of eyemovements. In: Stelmach, G.E., Requin, J. (Eds.), Tutorials inMotor Behavior. North Holland, Amsterdam, pp. 243–258.
Rafal, R., Smith, J., Krantz, J., Cohen, A., Brennan, C., 1990.Extrageniculate vision in hemianopic humans: saccadeinhibition by signals in the blind field. Science 250,118–121.
Rafal, R., Henik, A., Smith, J., 1991. Extrageniculate contributions toreflex visual orientating in normal humans: a temporalhemifield advantage. J. Cogn. Neurosci. 3, 322–328.
Savazzi, S., Marzi, C.A., 2004. The superior colliculus subservesinterhemispheric neural summation in both normals andpatients with a total section or agenesis of the corpus callosum.Neuropsychologia 42, 1608–1618.
Schiller, P.H., Malpeli, J.G., 1977. Properties and tectal projectionsof monkey retinal ganglion cells. J. Neurophysiol. 40, 428–445.
Sherman, S.M., 1974. Visual fields of cats with cortical and tectallesions. Science 185, 355–357.
Stein, B.E., Meredith, M.A., 1993. Merging of the Senses. MIT Press,Cambridge, MA.
Sumner, P., Adamjee, T., Mollon, J.D., 2002. Signals invisible to thecollicular and magnocellular pathways can capture visualattention. Curr. Biol. 12, 1312–1316.
Sylvester, R., Josephs, O., Driver, J., Rees, G., 2007. Visual FMRIresponses in human superior colliculus show a temporal-nasalasymmetry that is absent in lateral geniculate and visualcortex. J. Neurophysiol. 97, 1495–1502.
Todd, J.W., 1912. Reaction to multiple stimuli. Science Press,Oxford.
Ulrich, R., Miller, J., Schroter, H., 2007. Testing the race modelinequality: an algorithm and computer programs. Behav. Res.Meth. 39, 291–302.
Wallace, M.T., Meredith, M.A., Stein, B.E., 1992. Integration ofmultiple sensory modalities in cat cortex. Exp. Brain Res. 91,484–488.
White, A.J.R., Wilder, H.D., Goodchild, A.K., Sefton, A.J., Martin, R.,1998. Segregation of receptive field properties in the lateralgeniculate nucleus of a New-World monkey, the marmosetCallithrix jacchus. J. Neurophysiol. 80, 2063–2076.
Williams, C., Azzopardi, P., Cowey, A., 1995. Nasal and temporalretinal ganglion cells projecting to the midbrain: implicationsfor “blindsight". Neuroscience 65, 577–586.