Embed Size (px)
Citation preview

SAC
Sa
bTb
w
ArlaptdCdectaAnidtdAahArinAAsI
Kst
1
E*EAnsalPpsTet
Neuroscience 134 (2005) 155–163
0d
YNAPTIC AND SUBCELLULAR LOCALIZATION OF A-KINASENCHORING PROTEIN 150 IN RAT HIPPOCAMPAL CA1 PYRAMIDAL
ELLS: CO-LOCALIZATION WITH EXCITATORY SYNAPTIC MARKERSAtpdDbswak(tspar1ets
hct(2p(2tAtLAfo
pdosMcsNMGe
. M. LILLY,a F. J. ALVAREZb1 AND E. I. TIETZa1*
Department of Pharmacology and the Cellular and Molecular Neuro-iology Program, 3035 Arlington Avenue, Medical University of Ohio,oledo, OH 43614, USA
Department of Anatomy and Physiology, 3640 Colonel Glenn High-ay, Wright State University, Dayton, OH 45435, USA
bstract—Excitatory and inhibitory ionotropic receptors areegulated by protein kinases and phosphatases, which areocalized to specific subcellular locations by one of severalnchoring proteins. One of these is the A-kinase anchoringrotein (AKAP150), which confers spatial specificity to pro-ein kinase A and protein phosphatase 2B in the rat brain. Theistribution of AKAP150 was examined at rat hippocampalA1 pyramidal cell asymmetric and symmetric post-synapticensities and with respect to the distribution of markers ofxcitatory (vesicular glutamate transporter 1, glutamate re-eptor subunit 1) and inhibitory receptors (vesicular GABAransporter, GABA receptor type A �2/3 subunits, gephyrin)nd the Golgi marker, trans-Golgi network glycoprotein 38.KAP150 was close to asymmetric synapses, consistent withumerous molecular and biochemical studies suggesting its
nteraction with components of the excitatory postsynapticensity. In contrast, we did not find AKAP150-immunoreac-ivity associated with inhibitory synapses in rat CA1 neurons,espite reports demonstrating an in vitro interaction betweenKAP150 and GABA receptor type A receptor � subunits,nd the reported co-localization of these proteins in ratippocampal cultures. There was some overlap betweenKAP150 and GABA receptor type A receptor �2/3-immuno-
eactivity intracellularly in perinuclear clusters. These find-ngs support previous work indicating the integration of ki-ase and phosphatase activity at excitatory synapses byKAP150, but do not support a role for selective targeting ofKAP150 and its accompanying proteins to inhibitoryynapses. © 2005 Published by Elsevier Ltd on behalf ofBRO.
ey words: protein kinase A, calcineurin, post-synaptic den-ity, GABAA receptors, receptor phosphorylation, receptorargeting.
An equal contribution was made from the laboratories of F.J.A. and.I.T.
Corresponding author. Tel: �1-419-383-4170, fax: �1-419-383-2871.-mail address: [email protected] (E. Tietz).bbreviations: AKAP, A-kinase anchoring protein; AKAP150-IR, A-ki-ase anchoring protein 150 immunoreactivity; EM, electron micro-copic; GABAA, GABA receptor type A; GABARAP, GABAA receptorssociated protein; GluR1-4, glutamate receptor subunits 1-4; LM,
ight microscopic; LTP, long term potentiation; PKA, protein kinase A;KC, protein kinase C; PP2B, protein phosphatase type B; PSD,ost-synaptic density; RACK1, receptor for activated C-kinase-1; SO,tratum oriens; SP, stratum pyramidale; SR, stratum radiatum;GN38, trans-Golgi network glycoprotein 38; TMB, Tris–maleate buff-
Gr; VGAT, vesicular GABA transporter; VGLUT1, vesicular glutamateransporter 1.
306-4522/05$30.00�0.00 © 2005 Published by Elsevier Ltd on behalf of IBRO.oi:10.1016/j.neuroscience.2005.03.039
155
-kinase anchoring proteins (AKAPs) are a group of func-ionally related proteins that bind the regulatory subunits ofrotein kinase A (PKA), and target the holoenzyme toiscrete subcellular locations (Colledge and Scott, 1999;iviani and Scott, 2001). The various AKAP isoforms cane discerned by their specific molecular interactions andubcellular distribution (Colledge and Scott, 1999; Ed-ards and Scott, 2000). AKAP150 (human AKAP79) inter-cts with the regulatory (RII�) subunit of PKA, proteininase C (PKC), and protein phosphatase type B (PP2B)calcineurin). Moreover, this AKAP isoform is believed toarget its associated proteins to neuronal membranes andynapses, through domains capable of interacting withost-synaptic scaffold proteins including PSD-95, SAP-97,nd F-actin, as well as the intracellular loops of GABAeceptor type A (GABAA) receptor � subunits (Carr et al.,992; Colledge et al., 2000; Gomez et al., 2002; Brandont al., 2003). Thus AKAP150 may confer spatial specificityo serine/threonine kinases and phosphatases at synapticites in situ.
Studies in recombinant and native neuronal systemsave indicated that the cytoplasmic tails of glutamate re-eptor subunits (GluR)1, GluR2, and GluR4 AMPA recep-or subunits are substrates for PKA, PKC, and CaMKIIRoche et al., 1996; Barria et al., 1997; McDonald et al.,001). PKA modulates AMPA receptor currents via phos-horylation of GluR1-containing receptors at Ser845Greengard et al., 1991; Roche et al., 1996; Banke et al.,000). In the hippocampus, PKA-mediated phosphoryla-ion of Ser845 is permissive for synaptic incorporation ofMPA receptors during long term potentiation (LTP) induc-
ion and required for LTP maintenance (Nayak et al., 1998;ee et al., 2000; Esteban et al., 2003). In addition, it is theKAP150-anchored PKA and PP2B that are responsible
or the phosphorylation and dephosphorylation of Ser845n GluR1 (Tavalin et al., 2002).
Similarly, GABAA receptors are regulated by proteinhosphorylation (Swope et al., 1999; Olsen and Mac-onald, 2002; Brandon et al., 2002). The cytoplasmic loopsf GABAA receptor �1–3 subunits contain consensus PKAubstrate sequences (Moss et al., 1992; McDonald andoss, 1994), and increases or decreases in GABAA re-
eptor function have been reported depending on the �ubunit residue phosphorylated (McDonald et al., 1998;usser et al., 1999; Poisbeau et al., 1999; Hinkle andacdonald, 2003). The effects of phosphorylation onABAA receptor function may be mediated by PKA teth-red to the receptor by AKAP150. Indeed, AKAP150 and
ABAA receptor � subunits co-immunoprecipitate from
wc
capcAcoiheanfyrTabctGg
T
EaNt(ebhtmsRp
I
AGgUUcfi
(ehaa(td(
m(cSRic(
hfAf(tG(aoa((maiHo0cpodntaAt�tt
(msawtmfwa0tTifma1rTcW3El
S. M. Lilly et al. / Neuroscience 134 (2005) 155–163156
hole rat brain lysates, and co-localize in rat hippocampalulture (Brandon et al., 2003).
Modulation of receptor trafficking and function via re-eptor phosphorylation could occur locally at synapses ort a distance from synaptic sites, likely directed by therecise subcellular localization of kinase/phosphatase an-horing complexes. Unfortunately the exact location ofKAP150 in neurons is uncertain. In one electron micros-opy study (Sik et al., 2000) AKAP 79, the human orthologf rodent AKAP150, was found postsynaptically in proxim-
ty to excitatory (asymmetrical) synapses in the humanippocampus. However, a light microscopy study (Glantzt al., 1992) failed to reveal AKAP150 immunoreactivityssociated with excitatory synapses in rat hippocampaleurons. Moreover, despite the wealth of biochemical in-
ormation relating AKAP150 to GABAA receptor phosphor-lation, neither of these studies found AKAP150 immuno-eactivity at, or in close proximity to, inhibitory synapses.herefore in this study, the distribution of AKAP150 wasssessed in rat CA1 neuron synapses using both preem-edding immunohistochemistry and dual immunofluores-ence with markers of excitatory (vesicular glutamateransporter 1 (VGLUT1), GluR1) and inhibitory (vesicularABA transporter (VGAT), GABAA receptor �2/3 subunits,ephyrin) synapses.
EXPERIMENTAL PROCEDURES
issue preparation
ight adult male Sprague–Dawley rats (250–500 g, Harlan, Indi-napolis, IN, USA), were perfused under anesthesia (120 mg/kga pentobarbital, i.p.) via the left ventricle with 0.9% NaCl con-
aining 4% sucrose followed by 4% paraformaldehyde in 0.1 M PBpH 7.4) with or without 0.25% glutaraldehyde for light (LM) andlectron microscopic (EM) studies, respectively. For LM studies,locks of tissue containing the hippocampus were post-fixed (1–6at 4 °C), equilibrated in 15% sucrose in PB, then frozen-sec-
ioned in the coronal or sagittal plane at 40 �m on a slidingicrotome. Tissue intended for EM studies was blocked and
ectioned at 50 �m on a vibratome® (Pelco101, Ted Pella Inc.,edding, CA, USA). All sections were collected in PBS (0.01 M,H 7.4) and stored at 4 °C until further processing.
mmunohistochemistry
Antibodies. The following antibodies were used: goat anti-KAP150 (N-19, Santa Cruz, Santa Cruz, CA, USA), rabbit anti-luR1 (R. Wenthold NIDCD, Bethesda, MD, USA), mouse anti-ephyrin (mAb 7a, Boehringer Mannheim, Indianapolis, IN,SA), guinea-pig anti-VGLUT1 (Chemicon, Temecula, CA,SA), mouse anti-GABAA receptor �2/3 (mAb341, bd-17, Chemi-on), rabbit anti-VGAT (Chemicon), and mouse-anti-TGN-38 (Af-nity Bioreagents, Golden, CO, USA).
LM immunohistochemistry. Free-floating sagittal sections40 �m) were slide-mounted (Colorfrost®/Plus slides, Fisher Sci-ntific, Pittsburgh, PA, USA), blocked for 30 min in 10% normalorse serum diluted in PBS-T (0.01 M PBS�0.1% Triton X-100),nd incubated overnight at 4 °C in anti-AKAP150 (1:200), rabbitnti-GluR1 (1:1000, 1 �g/ml) or mouse anti-GABAA receptor �2/31:100, 10 �g/ml). After rinsing (3�5 min, 1�10 min PBS-T)issues were exposed to HRP-conjugated biotin-sp-IgG raised inonkey, against goat (AKAP150), rabbit (GluR1), or mouse
GABAA receptor �2/3). Sections were rinsed, developed 5–10 ein with a nickel-enhanced DAB (diaminobenzidine) reactionVector Laboratories, Burlingame, CA, USA), dehydrated andleared with xylene, then coverslipped with Permount™ (Fisher).lide-mounted sections were visualized on a lightbox (Imagingesearch Inc., St. Catherine, Ontario, Canada) under constant
llumination and images were acquired with a high resolution CCDamera (Sierra Scientific, Sunnyvale, CA, USA) using NIH imagev. 1.59) software.
Double immunofluorescence. After 30 min in 10% normalorse serum diluted in PBS-T (0.01 M PBS�0.1% Triton X-100),ree-floating sections were incubated overnight at 4 °C in anti-KAP150 (1:100–1:200) alone or in combination with one of the
ollowing: rabbit anti-GluR1 (1:1000), mouse anti-gephyrin1:100), guinea-pig VGLUT1 (1:4000), mouse anti-GABAA recep-or �2/3 (1:100), rabbit anti-VGAT (1:200), or mouse anti-trans-olgi network glycoprotein 38 (TGN38) (1:1000). After rinsing
3�5 min, 1�10 min PBS-T) tissues were exposed to Alexanti-goat-568 (1:200, Molecular Probes, Eugene, OR, USA) aloner in combination with the appropriate secondary antibody: Alexanti-mouse-488 (1:200, Molecular Probes), Alexa anti-rabbit-4881:200, Molecular Probes), or anti-guinea-pig-FITC antibodies1:50, Jackson Laboratories). Sections were then rinsed andounted on glass slides coated with 0.5% gelatin/0.05% chromelum and coverslipped with Vectashield (Vector). Immunoreactiv-
ties were analyzed with an Olympus FX confocal microscope.igh magnification was achieved using a Plan APO Olympus 60�bjective lens (1.4 numerical aperture, theoretical resolution.17 �m). Cross-talk between the emission of different fluoro-hromes at 488 nm with confocal microscopy was minimized asreviously described (Geiman et al., 2002). Controls included themission of the primary or secondary antibody in both single- andouble-labeling experiments. These controls verified lack of sig-ificant auto-fluorescence, lack of secondary antibodies’ interac-ions with naïve tissue, and the lack of secondary antibody inter-ctions with primary antibodies of different species. In addition,KAP150 antiserum specificity was tested by pre-absorption with
he 19-amino acid immunizing peptide (20 �g antiserum and 20g peptide for 60 min at room temperature), before being applied
o tissue sections. No staining was observed following preabsorp-ion of the primary AKAP150 antiserum.
Preembedding immunohistochemistry. Vibratome sections50 �m) were exposed to 1% sodium borohydride in PBS for 30in, rinsed 6�5 min in PBS, and incubated in 10% normal chicken
erum (Vector) for 3 h. Sections were then incubated in goatnti-AKAP150 (1:50) overnight with agitation at 4 °C. Tissuesere rinsed (3�5 min; 1�10 min) and incubated for 3 h at room
emperature in anti-goat-biotin (Vector), rinsed (3�5 min; 1�10in) and exposed to avidin–biotin-peroxidase complex (Vector)
or 2 h. After rinsing, the immunoreactive sites were visualizedith DAB (0.005% a DAB reaction with 0.01% H2O2), rinsedgain, fixed for 60 min in 2.5% glutaraldehyde, and washed in.1 M Tris–maleate buffer (TMB, pH 7.4) overnight. Sections werehen silver-intensified with a silver nitrate solution (6 min), rinsed inMB (3�5 min), treated with 0.05% gold chloride (5 min), rinsed,
ncubated with 2.5% sodium thiosulfate (2 min), rinsed in TMB,ollowed by PBS, then fixed with 0.5% osmium tetroxide for 20in. To prepare for embedding, sections were dehydrated throughn ascending series of ethanols (50%, 2�70%, 2�95%, 3�100%,0 min each), propylene oxide (2�10 min), 50% epon/aralditeesin in propylene oxide (overnight) and then 100% resin (6 h).issues were flat-embedded in between glass coverslips wereoated with formen-trenmittel (Electron Microscopy Sciences, Ft.ashington, PA, USA) and the resin was polymerized at 55 °C fordays. Areas of CA1 were excised from cured resin, mounted inM capsules for recutting, ultrathin sections obtained and col-
ected on nickel grids. The sections were analyzed in a Philips 201
lectron microscope. Digital images were obtained using a Bio-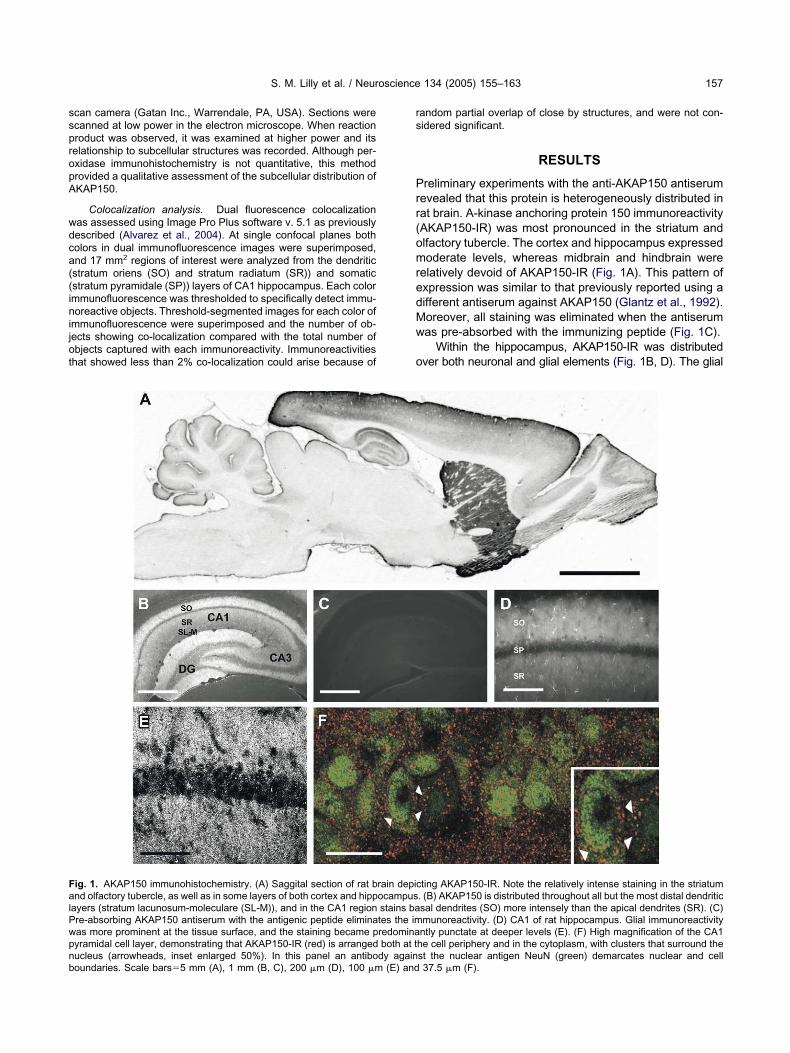
sspropA
wdca((inijot
rs
Prr(omredMw
o
FalPwpnb
S. M. Lilly et al. / Neuroscience 134 (2005) 155–163 157
can camera (Gatan Inc., Warrendale, PA, USA). Sections werecanned at low power in the electron microscope. When reactionroduct was observed, it was examined at higher power and itselationship to subcellular structures was recorded. Although per-xidase immunohistochemistry is not quantitative, this methodrovided a qualitative assessment of the subcellular distribution ofKAP150.
Colocalization analysis. Dual fluorescence colocalizationas assessed using Image Pro Plus software v. 5.1 as previouslyescribed (Alvarez et al., 2004). At single confocal planes botholors in dual immunofluorescence images were superimposed,nd 17 mm2 regions of interest were analyzed from the dendriticstratum oriens (SO) and stratum radiatum (SR)) and somaticstratum pyramidale (SP)) layers of CA1 hippocampus. Each colormmunofluorescence was thresholded to specifically detect immu-oreactive objects. Threshold-segmented images for each color of
mmunofluorescence were superimposed and the number of ob-ects showing co-localization compared with the total number ofbjects captured with each immunoreactivity. Immunoreactivitieshat showed less than 2% co-localization could arise because of
ig. 1. AKAP150 immunohistochemistry. (A) Saggital section of rat bnd olfactory tubercle, as well as in some layers of both cortex and hipp
ayers (stratum lacunosum-moleculare (SL-M)), and in the CA1 regionre-absorbing AKAP150 antiserum with the antigenic peptide eliminaas more prominent at the tissue surface, and the staining became pyramidal cell layer, demonstrating that AKAP150-IR (red) is arranged
ucleus (arrowheads, inset enlarged 50%). In this panel an antibody againoundaries. Scale bars�5 mm (A), 1 mm (B, C), 200 �m (D), 100 �m (E) andandom partial overlap of close by structures, and were not con-idered significant.
RESULTS
reliminary experiments with the anti-AKAP150 antiserumevealed that this protein is heterogeneously distributed inat brain. A-kinase anchoring protein 150 immunoreactivityAKAP150-IR) was most pronounced in the striatum andlfactory tubercle. The cortex and hippocampus expressedoderate levels, whereas midbrain and hindbrain were
elatively devoid of AKAP150-IR (Fig. 1A). This pattern ofxpression was similar to that previously reported using aifferent antiserum against AKAP150 (Glantz et al., 1992).oreover, all staining was eliminated when the antiserumas pre-absorbed with the immunizing peptide (Fig. 1C).
Within the hippocampus, AKAP150-IR was distributedver both neuronal and glial elements (Fig. 1B, D). The glial
ting AKAP150-IR. Note the relatively intense staining in the striatum. (B) AKAP150 is distributed throughout all but the most distal dendriticsal dendrites (SO) more intensely than the apical dendrites (SR). (C)munoreactivity. (D) CA1 of rat hippocampus. Glial immunoreactivityntly punctate at deeper levels (E). (F) High magnification of the CA1he cell periphery and in the cytoplasm, with clusters that surround the
rain depicocampusstains bates the imredominaboth at t
st the nuclear antigen NeuN (green) demarcates nuclear and cell37.5 �m (F).

saousbtmwdic
apaDmimFd2ws
tdAo
miicaswpPpHVAwA(apw
Fpp erved ina ts (arrow
S. M. Lilly et al. / Neuroscience 134 (2005) 155–163158
taining was most obvious at the surface of the tissue section,nd when shorter post-fixation was utilized. The significancef this staining is presently unclear. AKAP150-IR was notniform in all regions. For example, CA1 and dentate gyrushowed stronger immunoreactivity than CA3. In addition,asal dendritic regions (SO) displayed greater immunoreac-ivity than did apical dendritic regions (SR; Fig. 1B). At higheragnification, AKAP150-IR was punctate (Fig. 1E), andhen superimposed on NeuN-immunolabeled CA1 pyrami-al cell somata revealed that most AKAP150-IR was located
n the cytoplasm, while a smaller proportion of puncta werelose to the cell surface (Fig. 1E, F).
The subcellular distribution of AKAP150 was further ex-mined with electron microscopy and silver-intensified ABCeroxidase preembedding immunolabeling. AKAP150-IRppeared as clusters of silver particles over a weak diffuseAB precipitate that frequently was not well resolved inetal-contrasted sections. Most AKAP150-IR was found
n dendritic spines, usually in spine cytoplasm or theembrane region outside the PSD (post-synaptic density;ig. 2A–C). Some AKAP150-IR was found in other den-ritic regions and also on glial profiles (arrowheads, Fig.D). Although multiple CA1 layers were analyzed, thereas no indication of AKAP150-IR in close association with
ig. 2. Electron micrographs of AKAP150-IR in rat hippocampal CA1ost-synapses. AKAP150-IR was concentrated in the neck of dendost-synaptic densities (arrows in A–C). No immunoreactivity was obsctivity was also noted in glial elements, associated with glial filamen
ymmetric, presumed inhibitory synapses. Control sec- 3
ions, including those from the thalamus (a region thatoes not express AKAP150) and those not exposed toKAP antibodies showed no reaction product in neuronalr glial elements.
Although the ultrastructural studies indicated enrich-ent of AKAP150 in association with excitatory, but not
nhibitory, synapses in CA1 of rat hippocampus, it wasmportant to reevaluate these findings with light micros-opy, where less stringent fixation and membrane perme-bilization could be used, allowing for increased detectionensitivity. First, immunofluorescent labeling for AKAP150as compared with markers of presynaptic (VGLUT1) orostsynaptic elements (GluR1) of excitatory synapses.reliminary observations confirmed the robust hippocam-al expression of VGLUT1 reported elsewhere (Fig. 3A;erzog et al., 2001). AKAP150-IR did not overlap withGLUT1 staining in any hippocampal layer suggesting thatKAP150 is not localized presynaptically. However, thereere regions where VGLUT1-IR was apposed toKAP150-IR in the cellular and dendritic layers of CA1
arrowheads, Fig. 3B, C). GluR1 immunoreactivity waslso strong in the rat hippocampus, and also reflected theattern previously reported (Petralia and Wenthold, 1992)ith a relatively dense distribution in the CA1 region (Fig.
Most immunoreactive profiles were found in proximity of asymmetrics (reaction product denoted by arrowheads in A–C), proximate tothe vicinity of symmetric, presumed inhibitory, synapses. Immunore-
head in D). Scale bars�0.5 �m.
neurons.ritic spine
D). Although there was occasional overlap of AKAP150

aapA
pco(pisCcicr(m4aGlAaG
Apaw
iAawtf
swGAin5
TApsspiphfat
FS(t 0 (arrowhb
S. M. Lilly et al. / Neuroscience 134 (2005) 155–163 159
nd GluR1, less than 1% of AKAP clusters were associ-ted with GluR1 immunoreactivity (Fig. 3D–F). This iserhaps a reflection of the subsynaptic distribution ofKAP150 noted with electron microscopy.
To assess whether AKAP150 was present at rat hip-ocampal CA1 inhibitory synapses, dual immunofluores-ence studies were carried out with AKAP150 and markersf inhibitory terminals (VGAT) and postsynaptic elementsgephyrin and GABAA receptor �2/3 subunit). VGAT isresent in the synaptic vesicles of inhibitory terminals, and
n the rat hippocampus clearly demarcates the cellularomatic layer of CA1 (Fig. 4A). VGAT immunoreactivity inA1 somatic (Fig. 4B) or dendritic (Fig. 4C) layers did noto-localize with AKAP150-IR in these layers. Interneuronsn CA1 dendritic layers often exhibited perinuclear AKAPlusters usually more prominent than those evident in py-amidal neurons. Gephyrin, a marker of inhibitory PSDsSassoè-Pognetto and Fritschy, 2000), likewise exhibitedore pronounced staining in the somatic layer of CA1 (Fig.D). As the gephyrin antibody is more sensitive to fixation,shorter post-fixation (1 h) was used to assess overlap.lial staining for AKAP was appreciably greater in the
ighter-fixed tissue. There was no significant enrichment ofKAP-immunoreactivity (i.e. �1% colocalization) associ-ted with gephyrin clusters in either the cellular (Fig. 4E–), or dendritic layers of CA1.
In hippocampal cell cultures co-localization between ofKAP150 and �2/3 immunoreactivities have been re-orted (Brandon et al., 2003). We utilized a monoclonalntibody to GABA receptor �2/3 subunits in conjunction
ig. 3. Immunohistochemistry of AKAP150 and excitatory synaptic maP. Note that VGLUT1 (green) and AKAP150 (red) do not overlap, bu
SR, panel C). (D) GluR1-IR in the CA1 region. (E) High magnification imhe pyramidal cell cytoplasm, and partially overlaps with AKAP15ars�100 �m (A, D); 25 �m (B, C, E, F).
A
ith the antiserum to AKAP150 to explore this possible a
nteraction in intact hippocampus. Again, less than 1% ofKAP150 clusters were associated with �2/3 immunore-ctivity (Fig. 4J). Interestingly, the limited overlap detectedas largely cytoplasmic and distributed in clusters around
he nuclear periphery (Fig. 4J). Whether this overlap isunctionally significant is presently unclear.
AKAP150-IR was clearly associated with intracellulartructures surrounding the nucleus, where it co-localizedith TGN38, a transport protein specifically located in theolgi apparatus (Fig. 5A–C). In the CA1 region, 10% ofKAP150 clusters contained also TGN-38 immunoreactiv-
ty. This co-localization was most apparent in the promi-ent Golgi apparatus displayed by hilar interneurons (Fig.D–F).
DISCUSSION
his study extends previous studies that describeKAP150 immunoreactivity in multiple layers of the hip-ocampal CA1 region and provides the first direct ultra-tructural evidence that AKAP150 immunoreactivity is as-ociated with excitatory synaptic profiles in rat hippocam-al CA1 pyramidal cells, in situ. Using both EM and LM
mmunochemical techniques, we found no evidence for theresence of AKAP150 at inhibitory synapses in the ratippocampus, suggesting that there is no anatomical basisor a stable interaction between AKAP150-anchored PKAnd GABAA receptor � subunits at the synapse. The iden-ification of AKAP150 in clusters around the nucleus and in
) VGLUT1-IR in the CA1 region. (B) High magnification image of CA1rspersed (see arrowheads) in both this layer, and in dendritic layersLUR1-IR in CA1 SP. Note the GluR1-IR (green) is present throughouteads, lower right) (F), enlarged 200% in G (arrowheads). Scale
rkers. (At are inteage of G
ssociation with a marker of the Golgi apparatus offers the

po
mwughctpKfisr
ee
gt1mpaSpcsi
FoIrcmt s in interb
S. M. Lilly et al. / Neuroscience 134 (2005) 155–163160
ossibility that the AKAP150 plays a role in the traffickingf membrane-targeted proteins.
AKAP150 is present throughout rat hippocampal for-ation, but restricted to CA1 in human hippocampus. Not-ithstanding these differences, here we report that theltrastructural distribution of AKAP150 in the rat CA1 re-ion is similar to the distribution of its ortholog in CA1 ofuman hippocampus (Sik et al., 2000). Our findings areonsistent with multiple biochemical and molecular studieshat support the association of AKAP150 and excitatoryost-synapses (Bregman et al., 1989; Coghlan et al., 1995;lauck et al., 1996; Colledge et al., 2000). Moreover, thendings provide a direct structural correlate for functionaltudies that implicate the PKA-AKAP150-PP2B complex in
ig. 4. Immunohistochemistry of AKAP150 with inhibitory synaptic maf CA1 SP. Although the VGAT (green) and AKAP150 (red) clustersmage from SR depicting a VGAT immunoreactive interneuron, with Aegion. When higher magnification images of gephyrin (E) and AKAP1o-localize. (H) GABAA receptor �2/3-IR in CA1 of hippocampus.agnification image depicting �2/3-IR (green) and AKAP150-IR (red) in
he overlap that is present between �2/3 and AKAP150 is more obviouars�100 �m (A, D, H); 25 �m (B, C, E–G, I).
egulating short- and long-term changes in hippocampal t
xcitatory receptor function (Rosenmund et al., 1994; Abelt al., 1997; Tavalin et al., 2002).
Asymmetric synapses contain NMDA- and AMPA-typelutamate receptors embedded in a prominent membranehickening composed of multiple proteins, the PSD (Gray,959; Kennedy, 1997; Walikonis et al., 2000). Scaffoldingolecules, like those in the AKAP family, are proposed tolay a role in recruiting signaling molecules to the postsyn-ptic membrane (Colledge and Scott, 1999; Edwards andcott, 2000; Diviani and Scott, 2001). However, in theresent immunofluorescence studies, most AKAP150lusters were not associated with GluR1 clusters. Ultra-tructural analysis revealed that AKAP150 immunoreactiv-
ty was clearly associated with dendritic spines, subjacent
VGAT-IR in the CA1 region. (B) High magnification photomicrographpersed, a common overlapping pattern is not easily discernable. (C)rranged around the nucleus (arrowhead). (D) Gephyrin-IR in the CA1P of CA1 are overlaid (G), puncta representing these proteins do notpresence of numerous well-stained interneurons in SP. (I) High
midal cell layer of CA1. Panel I is enlarged 200% in J. Although limited,neurons, and present within cytoplasmic clusters (arrowheads). Scale
rkers. (A)are intersKAP-IR a50 (F) in SNote thethe pyra
o the post-synapse, consistent with the lack of co-local-

iittlgsAntbmdmnsAptmArt
lSt1At
sdSarA(
lAteipta
G1H(GAwrshsP
Fo(T gi apparac
S. M. Lilly et al. / Neuroscience 134 (2005) 155–163 161
zation between AKAP150 and GluR1. The immunoperox-dase label offers the advantage of greater sensitivity, buthe reaction product tends to diffuse, and therefore lackshe spatial resolution of immunogold labeling. However,ittle or no immunoreactivity was detected in PSDs sug-esting that diffusion of the reaction product from thesetructures is unlikely to have caused errant localization ofKAP150-immunoreactivity in the cytoplasm. Unfortu-ately we were unable to obtain adequate labeling usinghis antibody with post-embedding techniques. It is possi-le that AKAP150-immunoreactivity could have beenasked by reduced antibody accessibility or steric hin-rance with the macromolecular PSD complex. Using im-unogold, Sik et al. (2000) also reported AKAP79 immu-oreactivity near, but not within PSDs. Considering theimilarity between the ultrastructural distribution ofKAP79/150 immunoreactivity in the human and rat hip-ocampal CA1 regions reported by Sik et al. (2000), and in
his study using two different immunocytochemical EMethods and different antibodies, we conclude thatKAP150 may be preferentially located in cytoplasmic
egions in proximity to excitatory synapses, but not withinhe PSD.
Molecular studies have indicated that AKAP150 isinked to the GluR1 subunit by the post-synaptic proteinAP97 (Colledge et al., 2000) and maintains AMPA recep-
or function by locally recruiting PKA (Rosenmund et al.,994; Tavalin et al., 2002). In contrast, yotiao, an alternateKAP isoform, directly interacts with the NR1 subunit of
ig. 5. (A) AKAP150-IR in the CA1 pyramidal cell layer of the hippocaf pyramidal cells (arrowheads) and interneurons (arrows). (C) Whenarrowheads) and interneurons (arrows), although those on interneuronGN38-IR at the same plane as in (D), depicting the extensive Gololocalize in these interneurons (arrowheads). Scale bars�25 �m.
he NMDA receptor and active protein phosphatase 1, f
uppressing NMDA receptor function by promoting NR1ephosphorylation (Westphal et al., 1999; Edwards andcott, 2000). Although PKA bound to AKAPs may be in-ctive (Faux et al., 1999; Edwards and Scott, 2000), dis-uption of the AKAP150-PKA interaction suppressesMPA receptor function and phosphorylation on Ser845
Rosenmund et al., 1994; Tavalin et al., 2002).The subsynaptic localization of AKAP150 may allow for
ocal sequestration of PKA, which upon dissociation fromKAP150 is catalytically active and required to maintain
he phosphorylation of synaptic GluR1 subunits. Whenxcitatory synaptic strength is increased following LTP
nduction, AKAP150 is upregulated (Genin et al., 2003),erhaps recruiting more PKA. This might increase excita-ory synaptic function and receptor incorporation (Lee etl., 2000; Esteban et al., 2003).
There is ample evidence for PKA-mediated regulation ofABAA receptors (McDonald et al., 1998; Poisbeau et al.,999; Nusser et al., 1999; Olsen and Macdonald, 2002;inkle and Macdonald, 2003). Moreover, Brandon et al.
2003) reported an association between AKAP150 andABAA receptor � subunits, and co-localization ofKAP150 and �2/3 in rat hippocampal culture. However,e did not find AKAP150 at inhibitory synapses in CA1 of
at hippocampus. This finding is consistent with the ab-ence of AKAP79 at inhibitory synapses in the humanippocampus (Sik et al., 2000), and fails to provide atructural basis for local regulation by AKAP150-anchoredKA of GABA receptors at the synapse. Similarly, receptor
) TGN38-IR at the same plane as in (A), depicting the Golgi apparatus, there are multiple areas of colocalization over both pyramidal cellsre apparent. (D) AKAP150-IR in the hilus of the rat dentate gyrus. (E)tus in hilar interneurons. (F) When overlaid AKAP150 and TGN38
mpus. (Boverlaids are mo
or activated C-kinase-1 (RACK1) and PKC co-immuno-

pat(hwaaa(psktzptkb
wsoNt1rtfeaccrrftettcatae
TthtafiArrp
AtM9
A
A
B
B
B
B
B
C
C
C
C
D
E
E
F
G
G
G
S. M. Lilly et al. / Neuroscience 134 (2005) 155–163162
recipitate with cortical GABAA receptor � subunits, inter-ctions that are necessary for the PKC-mediated attenua-ion of whole-cell GABA currents in cultured neuronsBrandon et al., 2002). However, neither PKC nor RACK1ave been identified immunohistochemically in associationith inhibitory synapses in the CNS. Unlike excitatory syn-pses, inhibitory profiles do not exhibit a prominent PSD,re not associated with spines, and in the hippocampusre concentrated at the cell soma and proximal processesMegias et al., 2001). In the absence of an anchoringrotein, one possibility is that the receptors at inhibitoryynapse PSDs are regulated by cytoplasmic changes ininase and phosphatase activity, rather than being con-rolled by compartmentalized aggregates of regulatory en-ymes, including PKA and PKC in the PSDs. Alternatively,ost-translational modifications of synaptic GABAA recep-
ors may occur at the synapse through recruitment ofinases and phosphatases by scaffolding molecules yet toe identified.
The perinuclear staining of AKAP150 and its overlapith the Golgi-apparatus marker TGN38 is intriguing. Aimilar pattern of Golgi-staining was observed in a varietyf cell lines with an alternate AKAP isoform, AKAP350.otably, AKAP350 has multiple splice variants, including
he NMDA receptor binding AKAP, yotiao (Schmidt et al.,999). It is interesting to speculate that the Golgi- andeceptor-associated AKAP variants regulate excitatoryone by affecting receptor trafficking and synaptic receptorunction, following transcription of a single gene. The pres-nce of AKAP150 immunoreactivity in perinuclear clustersnd in association with TGN38 may indicate a Golgi-asso-iated function for AKAP150. Like AKAP150, GABAA re-eptor associated protein (GABARAP) binds to GABAA
eceptor subunits in vitro and co-localizes with GABAA
eceptors in neuronal culture. Additionally, GABARAP isound in the Golgi apparatus and now thought related tohe intracellular trafficking of GABAA receptors (Kneusselt al., 2000; Kittler et al., 2001). An alternate possibility is
hat AKAP150 is clustered at the Golgi-apparatus in transito the sub-synapse, where it tethers local signaling mole-ules. Albeit limited, the co-localization between AKAP150nd the GABAA receptor �2/3 subunit in perinuclear clus-
ers offers the possibility that the previously reported inter-ction between AKAP150 and GABAA receptors (Brandont al., 2002) in part occurs in the Golgi apparatus.
CONCLUSIONS
he present study indicates that AKAP150 immunoreac-ivity is present in proximity of excitatory synapses in ratippocampal CA1 neurons. Together with previous func-ional studies pertaining to the effect of AKAP150 and itsssociated kinases on excitatory receptor function, thesendings confirm the existence of an endogenous pool ofKAP150 in the proximity of synaptic excitatory ionotropic
eceptors. Conversely, we did not find AKAP150 immuno-eactivity in association with inhibitory synapses in rat CA1
yramidal cells.cknowledgments—This work was supported by NIDA pre-doc-oral fellowship F30-DA041412 (S.M.L), NIDA RO1 DA04075 andCO Drug Abuse Research Program (E.I.T), and NSF grant984441 (F.J.A.).
REFERENCES
bel T, Nguyen PV, Barad M, Deuel TA, Kandel ER, BourtchouladzeR (1997) Genetic demonstration of a role for PKA in the late phaseof LTP and in hippocampus-based long-term memory. Cell88:615–626.
lvarez FJ, Villalba RM, Zerda R, Schneider SP (2004) Vesicularglutamate transporters in the spinal cord, with special reference tosensory primary afferent synapses. J Comp Neurol 472:257–280.
anke TG, Bowie D, Lee HK, Huganir RL, Schousboe A, Traynelis SF(2000) Control of GluR1 AMPA receptor function by cAMP-depen-dent protein kinase. J Neurosci 20:89–102.
arria A, Derkach V, Soderling T (1997) Identification of the Ca2�/calmodulin-dependent protein kinase II regulatory phosphorylationsite in the alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate-type glutamate receptor. J Biol Chem 272:32727–32730.
randon NJ, Jovanovic JN, Moss S (2002) Multiple roles of proteinkinases in the modulation of gamma-aminobutyric acid(A) receptorfunction and cell surface expression. Pharmacol Ther 94:113–122.
randon NJ, Jovanovic JN, Colledge M, Kittler JT, Brandon JM, ScottJD, Moss SJ (2003) A-kinase anchoring protein 79/150 facilitatesthe phosphorylation of GABA(A) receptors by cAMP-dependentprotein kinase via selective interaction with receptor beta subunits.Mol Cell Neurosci 22:87–97.
regman DB, Bhattacharyya N, Rubin CS (1989) High affinity bindingprotein for the regulatory subunit of cAMP-dependent protein ki-nase II-B. J Biol Chem 264:4648–4656.
arr DW, Stofko-Hahn RE, Fraser IDC, Cone RD, Scott JD (1992)Localization of the cAMP-dependent protein kinase to postsynapticdensities by A-kinase anchoring proteins: Characterization ofAKAP79. J Biol Chem 267:16816–16823.
oghlan VM, Perino BA, Howard M, Langeberg LK, Hicks JB, GallatinWM, Scott JD (1995) Association of protein kinase A and proteinphosphatase 2B with a common anchoring protein. Science267:108–111.
olledge M, Scott JD (1999) AKAPs: from structure to function. TrendsCell Biol 9:216–221.
olledge M, Dean RA, Scott GK, Langeberg LK, Huganir RL, Scott JD(2000) Targeting of PKA to glutamate receptors through aMAGUK-AKAP complex. Neuron 27:107–119.
iviani D, Scott JD (2001) AKAP signaling complexes at the cytoskel-eton. J Cell Sci 113:1431–1437.
dwards AS, Scott JD (2000) A-kinase anchoring proteins: proteinkinase A and beyond. Curr Opin Cell Biol 12:217–221.
steban JA, Shi SH, Wilson C, Nuriya M, Huganir RL, Malinow R(2003) PKA phosphorylation of AMPA receptor subunits controlssynaptic trafficking underlying plasticity. Nat Neurosci 6:136–143.
aux MC, Rollins EN, Edwards AS, Langeberg LK, Newton AC, ScottJF (1999) Mechanism of A-kinase-anchoring protein 79 (AKAP79)and protein kinase C interaction. Biochem J 343:443–452.
eiman EJ, Zheng W, Fritschy J-M, Alvarez FJ (2002) Glycine andGABAA receptor subunits on Renshaw cells: Relationship withpresynaptic neurotransmitters and postsynaptic gephyrin clusters.J Comp Neurol 444:275–289.
enin A, French P, Doyere V, Davis S, Errington ML, Maroun M, SteanT, Truchet B, Webber M, Wills T, Richter-Levin G, Hunt SP, MalletJ, Laroche S, Bliss TVP, Oconnor V (2003) LTP but not seizure isassociated with up-regulation of AKAP-150. Eur J Neurosci17:331–340.
lantz SB, Amat JA, Rubin CS (1992) cAMP signaling in neurons:patterns of neuronal expression and intracellular localization for a
novel protein, AKAP150, that anchors the regulatory subunit of
G
G
G
H
H
K
K
K
K
L
M
M
M
M
M
N
N
O
P
P
R
R
S
S
S
S
T
W
W
S. M. Lilly et al. / Neuroscience 134 (2005) 155–163 163
cAMP-dependent protein kinase II beta. Mol Biol Cell 3:1215–1228.
omez LL, Alam S, Smith KE, Horne E, Dell’Acqua ML (2002) Reg-ulation of A-kinase anchoring protein 79/150-cAMP-dependentprotein kinase postsynaptic targeting by NMDA receptor activationof calcineurin and remodeling of dendritic actin. J Neurosci 22:7027–7044.
ray EG (1959) Axo-somatic and axo-dendritic synapses of the cere-bral cortex: an electron microscopic study. J Anat 93:420–433.
reengard P, Jen J, Nairn AC, Stevens CF (1991) Enhancement ofglutamate responses by cAMP-dependent protein kinase in hip-pocampal neurons. Science 253:1135–1138.
erzog E, Bellenchi GC, Grac C, Bernard V, Ravassard P, Bedet C,Gasnier B, Giros B, El Mestikawy S (2001) The existence of asecond vesicular glutamate transporter species specifies subpopu-lations of glutamatergic neurons. J Neurosci 21:RC181.
inkle DJ, Macdonald RL (2003) � Subunit phosphorylation selec-tively increases fast desensitization and prolongs deactivationof �1�1�2L and �1�3�2L GABAA receptor currents. J Neurosci23:11698 –11710.
ennedy MB (1997) The postsynaptic density at glutamatergic syn-apses. Trends Neurosci 20:264–268.
ittler JT, Rostaing P, Schiavo G, Fritschy J-M, Olsen R, Triller A,Moss SJ (2001) The subcellular distribution of GABARAP and itsability to interact with NSF suggest a role for this protein in theintracellular transport of GABAA receptors. Mol Cell Neuro 18:13–25.
lauck TM, Faux MC, Labudda K, Langeberg LK, Jaken S, Scott JD(1996) Coordination of three signaling enzymes by AKAP79, amammalian scaffold protein. Science 271:1589–1592.
neussel M, Haverkamp S, Fuhrmann JC, Want H, Wassle H, OlsenRW, Betz H (2000) The �-aminobutyric acid type A receptor(GABAAR)-associated protein GABARAP interactions with gephy-rin but is not involved in receptor anchoring at the synapse. ProcNatl Acad Sci U S A 97:8594–8599.
ee HK, Barbarosie M, Kamayama K, Bear MF, Huganir RL (2000)Regulation of distinct AMPA receptor phosphorylation sites duringbi-directional synaptic plasticity. Nature 405:955–959.
cDonald BJ, Moss SJ (1994) Differential phosphorylation of intracel-lular domains of GABAA receptor subunits by calcium/calmodulintype 2-dependent protein kinase and cGMP-dependent kinase.J Biol Chem 269:18111–18117.
cDonald BJ, Amato A, Connolly CN, Benke D, Moss SJ, Smart TG(1998) Adjacent phosphorylation sites on GABAA receptor � sub-units determine regulation by cAMP-dependent protein kinase. NatNeurosci 1:23–28.
cDonald BJ, Chung HJ, Huganir RL (2001) Identification of proteinkinase C phosphorylation sites within the AMPA receptor GluR2subunit. Neuropharmacology 41:672–679.
egias M, Emri Z, Freund TF, Gulyas AI (2001) Total number anddistribution of inhibitory and excitatory synapses on hippocampal
CA1 pyramidal cells. Neuroscience 102:527–540.oss SJ, Doherty CA, Huganir RL (1992) Identification of the cAMP-dependent protein kinase and protein kinase C phosphorylationsites within the major intracellular domains of the �1, �2s and �2Lsubunits of the GABAA receptor. J Biol Chem 267:14470–14476.
ayak A, Zastrow DJ, Lickteig R, Zahniser NR, Browning MD (1998)Maintenance of late-phase LTP is accompanied by PKA-depen-dent increase in AMPA receptor synthesis. Nature 394:680–683.
usser Z, Sieghart W, Mody I (1999) Differential regulation of synapticGABAA receptors by cAMP-dependent protein kinase in mousecerebellar and olfactory bulb neurons. J Physiol 521:421–435.
lsen RW, Macdonald RL (2002) GABAA receptor complex: Structureand function. In: Glutamate and GABA receptors: structure, func-tion and pharmacology (Egebjerg J, Schousboe A, Krogsgaard-Larsen P, eds), pp 203–235. London: Taylor & Francis.
etralia RS, Wenthold RJ (1992) Light and electron immunocytochem-ical localization of AMPA-selective glutamate receptors in the ratbrain. J Comp Neurol 318:329–354.
oisbeau P, Cheney MC, Browning MD, Mody I (1999) Modulation ofsynaptic GABAA receptor function by PKA and PKC in adult hip-pocampal neurons. J Neurosci 19:674–683.
oche KW, O’Brien RJ, Mammen AL, Bernhardt J, Huganir RL (1996)Characterization of multiple phosphorylation sites on the AMPAreceptor GLUR1 subunit. Neuron 16:1179–1188.
osenmund C, Carr DW, Bergeson SE, Nilaver G, Scott JD, West-brook GL (1994) Anchoring of protein kinase A is required formodulation of AMPA/kainite receptors on hippocampal neurons.Nature 368:853–856.
assoè-Pognetto M, Fritschy JM (2000) Mini-review: gephyrin, a majorpostsynaptic protein of GABAergic synapses. Eur J Neurosci12:2205–2210.
chmidt PH, Dransfield DT, Claudio JO, Hawley RG, Trotter KW,Milgram SL, Goldenring JR (1999) AKAP350, a multiply splicedprotein kinase A-anchoring protein associated with centrosomes.J Biol Chem 274:3055–3066.
ik A, Gulacsi A, Lai Y, Doyle WK, Pacia S, Mody I, Freund TF (2000)Localization of the A kinase anchoring protein AKAP79 in thehuman hippocampus. Eur J Neurosci 12:1155–1164.
wope SL, Moss SJ, Raymond LA, Huganir RL (1999) Regulation ofligand-gated ion channels by protein phosphorylation. Adv SecondMessenger Phosphoprotein Res 33:49–78.
avalin SJ, Colledge M, Hell JW, Langeberg LK, Huganir RL, Scott JD(2002) Regulation of GluR1 by the A-kinase anchoring protein 79(AKAP79) signaling complex shares properties with long-term de-pression. J Neurosci 22:3044–3051.
estphal RS, Tavalin SJ, Lin JW, Alto NM, Fraser ID, Langeberg LK,Sheng M, Scott JD (1999) Regulation of NMDA receptors by anassociated phosphatase-kinase signaling complex. Science 285:9–96.
alikonis RS, Jensen ON, Mann M, Provance DW Jr, Mercer JA,Kennedy MB (2000) Identification of proteins in the postsynaptic
density fraction by mass spectrometry. J Neurosci 20:4069–4080.(Accepted 25 March 2005)





























