Embed Size (px)
Citation preview

SOME EFFECTS OF BRAIN STIMULATIONON TIMING BEHA VIOR
J. V. BRADY and D. G. CONRADWALTER REED ARMY INSTITUTE OF RESEARCH
When reinforcement of a free operant is contingent upon a specified temporal delay be-tween responses, an organism develops a pattern of "spaced responding" or "timing be-havior" in accordance with the requirements of such a reward schedule (Anger, 1956;Conrad, Sidman, & Herrnstein, 1958; Sidman, 1956; Skinner, 1938; Wilson & Keller, 1953).The differential reinforcement of low rates (DRL) in this way generates a timing processwhich makes it possible for the organism to discriminate the duration of the intervalelapsing since the last response. Since there is no exteroceptive discriminative stimulus inthis situation, the event or events upon which reinforcement is contingent must be pro-vided by the organism's own behavior. The specific aspects of the organism's interactionwith its internal or external environment which mediate this timing process, however, arefar from clear.
Recent reports of the rewarding properties of electrical brain stimulation (Olds & Milner,1954; Brady, 1956; Brady, 1958) suggest an interesting approach to the exploration of cen-tral-nervous-system participation in such timing behavior. It has been demonstrated thatstable lever pressing on several different schedules of reinforcement can be maintained inlaboratory animals for an intracranial electrical stimulus in selected areas of the brainwithout any other reward (Sidman, Brady, Boren, Conrad, & Schulman, 1955). The purposeof the present study is to investigate the interaction between such rewarding brain stimu-lation and the timing process generated by the DRL schedule. Specifically, this report de-scribes a preliminary comparative experimental analysis of DRL performance under con-ditions of intracranial electrical self-stimulation reward and food reinforcement in therhesus monkey.
METHOD
General ProcedureThree mature, male, rhesus monkeys served as subjects. All animals were trained in lever
pressing for both brain-shock reinforcement and food reward. Timing behavior was estab-lished on a DRL schedule requiring that lever responses be spaced at least 20 seconds apartto produce reinforcement. With stereotaxically implanted chronic electrodes in selectedareas of the brain, DRL performance was compared under various conditions of electricalself-stimulation and/or food reinforcement. Upon completion of the behavioral observ-ations, all animals were sacrificed and the electrode placements verified histologically.
ApparatusThe monkeys were housed, trained, and tested in a specially constructed primate chair
previously described in some detail (Mason, 1958). This restraining device was equippedwith an automatic pellet feeder (Davis) and mouth hopper (Foringer), a visual stimuluspanel with five different-colored lights (Foringer), and a modified telegraph-key lever(Foringer). An electrical junction box and multiconductor cable attached to the chair pro-vided for interconnections between the experimental space and the control, programming,and recording equipment in an adjoining room. Responses and reinforcements were re-corded on electrical impulse counters, and a cumulative-response recorder (Gerbrands) pro-
93

J. V. BRADY and D. G. CONRAD
vided continuous performance measures. A system of timers and counters automaticallytabulated inter-response times in class intervals of 2 seconds and primed the reinforce-ment circuit at appropriate times. The output of a white-noise generator (Grason-Stadler)through a speaker in the experimental room masked sounds from the programming equip-ment which might have provided unwanted exteroceptive discriminative stimuli for themonkey.The electrical brain stimulus used to reinforce lever pressing in the self-stimulation
portions of these experiments was provided by a Tektronix stimulating unit modified togive a biphasic pulse similar to that described by Lilly, Hughes, Alvord, and Galkin (1955).The parameters of this intracranial electrical stimulus remained constant for each animalthroughout the experiment. Each brain shock consisted of a 100-cycle-per-second,0.2-millisecond pulse duration, 20-milliampere electrical stimulus applied for a train dura-tion of 0.5 second. Tissue resistance values in the monkeys ranged between 1500 and 2000ohms.
Operative ProcedureThe electrode construction and implanation procedure involved the use of a stereotaxi-
cally placed multilead electrode array and common-socket pedestal rigidly fixed to the skullso that stimulating leads could be firmly attached according to a procedure recently de-scribed by Sheatz (1959). All surgery was performed aseptically under pentobarbital anes-thesia, and the animals were permitted a postoperative recovery period of I to 2 weeks be-fore behavioral procedures were introduced.
Behavioral ProcedureAll animals were initially deprived of food for 24 hours and given a single 3-hour,
"'magazine-training" session during which sugar pellets (48 milligrams each) were auto-matically delivered into the food hopper on the chair on the average of once every 60 sec-onds. Then, they were trained in lever pressing on a continuous-reinforcement schedule (200sugar pellets) followed by' a series of 2-hour daily sessions on the DRL schedule. Reinforce-ment of the lever response with a sugar pellet occurred only if an interval of 20 secondshad elapsed since the last lever press. This differential reinforcement of low rates was con-tinued with sugar pellets alone as reward until the average response rate per session andinter-response time distributions (IRT's) for each animal showed no consistent change fromday to day.
Brain-stimulation ProcedureFollowing stabilization of the DRL performance, the brain stimulation was substituted
for sugar pellets as reinforcement during a 2-hour experimental session. Responses spaced20 seconds or more apart were reinforced during this session with a 20-milliampere elec-trical stimulus for 0.5 second at the electrode locations indicated in Fig. 1. All animalswere then continued on an alternating-day procedure which provided a 2-hour DRL sessionfor sugar pellets on one day followed the next day by a 2-hour DRL session for brain stimu-lation. This procedure was continued for 10 consecutive days, 5 days for sugar pellets and 5days for brain stimulation, with comparison of the DRL performance under the two con-ditions. Then, with all three monkeys, several experimental sessions explored the effects ofconcurrent food and brain-shock reward upon the timing schedule. During these sessions,responses spaced 20 seconds or more apart were reinforced with both a sugar pellet and theelectrical brain stimulus at the same time. Finally, one of the monkeys, M-12, was exposed
94

TIMING BEHA VIOR
to a similar series of experimental sessions involving various combinations of food andbrain stimulation alternatively and food and brain stimulation concurrently using the sec-
ond electrode placement indicated in Fig. 1.
Histological ProcedureHistological verification of the electrode placements was obtained for all three monkeys
upon completion of the experiment. The animals were perfused with physiological salinefollowed by 10% formalin in which the brains were then fixed for I to 4 weeks before em-
bedding in paraffin. Serial sections were cut at 15,u and stained with thionin. The elec-trode placements were reconstructed after microscopic examination of all brains andphotomicrographs of selected sections for each animal are shown in Fig. 1. Section M-197is cut approximately through frontal level A-17 (Horsley-Clark coordinates), and shows theelectrode track coursing through the caudate nucleus, internal capsule, and globus pallidus,with the stimulating tip in the area of the medial forebrain bundle. Section M-2 is cut
approximately through frontal level A-16 (Horsley-Clark coordinates) and shows the elec-trode track leaving the caudate nucleus and internal capsule to terminate with the stimu-lating tip in the medial aspect of the globus pallidus. Section M-12 is cut approximatelythrough frontal level A-11 (Horsley-Clark coordinates) and shows the two electrode place-ments used in this experiment. The rostral-medial aspects of the section on the left sideshow one electrode coursing through the cingulate gyrus, corpus callosum, and anteriorventral nucleus of the thalamus to terminate with the stimulating tip in the anteriormedial nucleus of the thalamus. The more lateral aspects of the section on the right sideshow the other electrode leaving the internal capsule and terminating with the electrode tipin the most medial portion of the globus pallidus.
RESULTS, DISCUSSION, AND CONCLUSIONS
Following introduction of the 20-second DRL requirement for sugar-pellet reward, thelever-pressing rates for all three monkeys showed a gradual and orderly decline similar tothat described by Wilson and Keller (1953) for the rat. Within 10 to 14 daily 2-hour ses-
sions for sugar pellets alone, the animals developed a clear and relatively stable time dis-crimination as reflected in the distribution of inter-response times (IRT's) recorded foreach monkey. With the exception of the previously reported (Sidman, 1956) high proportionof responses in the zero-to-2-second interval, the distribution of lever presses displayed thetypically gradual rise to a peak around the 20-second interval, and then a gradual declinein the frequency of longer inter-response times. The characteristics of this base-line per-
formance were common to all three monkeys prior to introduction of the brain stimulation.The cumulative curves in Fig. 2 illustrate typical DRL performances for Monkey M-12
under conditions of sugar-pellet reinforcement (upper section) and brain-stimulation re-ward (lower section) through the electrode placement in the globus pallidus (lower right
arrow, Section M-12, Fig. 1). The stable time discrimination generated by the "spaced-responding" contingency is clearly reflected in the low steady rate and high frequency ofreinforcement (vertical pips indicate reinforcements) when sugar pellets alone reward thelever response. In contrast, the response rate for the brain-stimulus reward on the sameDRL schedule shows a marked increase over the optimal "spaced-responding" performancewith a consequent low frequency of reinforcement.
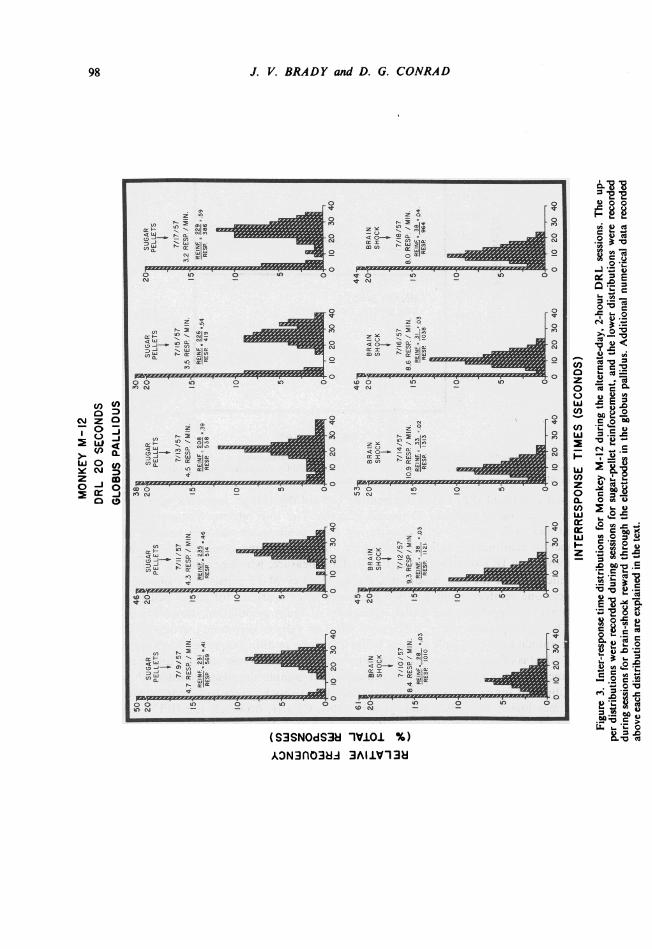
Figure 3 shows the IRT's recorded for Monkey M-12 during the alternate-day 2-hourDRL session5 for sugar pellets and brain stimulation in the globus pallidus. Thetypica!
95

J. V. BRADY and D. G. CONRAD
"timing" performance during the sugar-pellet sessions shows a peak in the response dis-tribution at or slightly above the 20-second interval required for reinforcement. The al-ternate-day IRT's recorded during DRL performance for brain shock, however, show a
M-2 M-197
M-12
Figure 1. Low-power photomicrographs of brain sections for Monkeys M-2, M-197, and M-12. The locationof the stimulating electrode tips are indicated by the arrows on each section.
96

TIMING BEHA VIOR
D R L-20 SECONDSMONKEY M-12
500 SUGAR PELLETS400
300
c> 200wc' 100z00- 0enC')w
z 500 BRAIN SHOCK> 400 (GLOBUS PALLIDUS)- X
300200--100-
0 ~ ~ ~ 330 60
TIME-MINUTESFigure 2. Sample cumulative-response curves showing the differential effect of reinforcement with sugar
pellets (upper section) and with brain stimulation in the globus pallidus (lower section) upon DRL performancefor Monkey M-12. Pips on the record indicate reinforcements.
marked and consistent shift of the distribution toward the shorter intervals with the peakfrequency occurring around the 10-second interval. The low relative frequency of inter-response times of 20 seconds or longer during these brain-shock sessions reduces sharplythe number of reinforcements as indicated by the "reinforcement-per-response" ratio re-corded above each IRT distribution in Fig. 3. This measure, which provides a rough indexof "efficiency" under the two conditions, shows Monkey M-12 consistently obtaining ap-proximately one reinforcement per two responses when working for sugar pellets (ratiosaround 0.50). The performance under conditions of brain stimulation, however, is char-acterized by a sharp drop in the "efficiency" ratio to values consistently around 0.03 andshows the animal obtaining only slightly better than one reinforcement per forty responses.A related effect is of course reflected in the markedly different response rates under the tworeinforcement conditions. The rates in responses per minute are also recorded above theIRT distributions for each session and show the "brain-shock" rates to be consistently twoto three times higher than the "sugar-pellet" rates. Clearly, however, the IRT's recordedduring the brain-stimulation sessions continue to show a temporal discrimination as evi-denced by the relatively sharp peaks around the 10-second interval. The animal continues tospace responses but makes a rather consistent error in the duration of the delay intervalrequired for brain-shock reinforcement.
Figure 4 shows a similar effect for Monkey M-2 with an electrode placement in a relatedportion of the globus pallidus (Section M-2, Fig. 1). The IRT's recorded during DRL ses-sions for sugar pellets show a peak frequency slightly below the required 20-second delayinterval. The gradual decline in the relative frequency of longer inter-response times, how-
97
J. V. BRADY and D. G. CONRAD
a ..r'n
,r ,,, r, z|U)~CL
lb-i
Zi s7
-J )
W N _ _
t
Z Q
zO_ X
"
;i sn 11 X S
_.90w_
O I0In
0
2
00FJ 0_0a) N
F ) F)I- t40~~~~~~~~~~~0
0
. ~~~~~~~~~~r°-3 IAJt 3
U) ff *
0-'- _) M
0 0
0
00 CM 0 )
F)n w -at -o Loi 2
In, 0
2R.0N
0zQ
c -i r-'
tooe o e >0
0
z I0
NN
0
Z 1z' c
co F0)a:
2-~~~~~'0
0
Ni 0
0
st C-i _
0 4ION w- co
0On F-Or O
'_ o
_.CD_VW ~~~~~
(S3SNOdS38 10VJOi %)AON3nO3HA 3A1.V-13i
98
Cf) U)c
0
>- O 0.
Y C\ -.3z mo J o
0c-
' -o
400
coo.00
'-: D4
*0 ' 0
4)-
o E
wCO > C
w 0
I . _;', 7
0 0ci) > 1l_4
mo oZ 2
.o0 °'Q
'0- 2.
10
fi o2 0.0X
e 0, _
I I

TIMING BEHAVIOR
00. -s
go
0 0N -
OW n
N _N
':w - .:(A C. a£ b
goa
00xi
0A 2 1-
z _re Wma1
p1ZZ °
4 a
0 _I. e
u *-.
a~1
ON - -
z*5O h-K I
0 ......._U
0 I0 0
Loe
N -P.- ,I32Z
mnI- -j
a w
0T*
-00i 0
- o I,
Z _
#A - a Ir
2
N b
-U * 0
0t 0 O O
0 0
- 0
e w b- ~~~~~~~~~'o
0
V.z
;!Iw 2ON" - -0
-u 0
N
00 O~~:
_
0_ o C~~~~~~~(S3SNOdS3H I1V.O %)ADN3nO384 3A1I.V 138
99
/) U)Cy 0 D
I Z a
Y 0
C
o .
0
2 0-w =
Cl_
o. C
W QC
ooQU)
U c
Qz;0^;
Cl) )
z 0Dz
E M-
CT
r .2oNbo -
-In W _
--------------
or . s___ w v w TI "IM

J. V. BRADY and D. G. CONRAD
ever, can be seen to extend well into the 30- and even 40-second delay interval compart-ments. In contrast, the DRL performance elicited with brain-shock reinforcement on al-ternate days is characterized by a shift in the IRT's to a peak frequency around the10-second delay interval. The complete absence of inter-response times longer than26 seconds under the brain-stimulation condition provides another striking similarity tothe brain-shock performance of Monkey M-12 shown in Fig. 3. The relationships betweenthe lever-pressing response rates and between the "reinforcement-per-response" ratiosunder the two different reinforcement conditions for Monkey M-2 are also reflected in thenumerical recordings above the IRT's for each experimental session in Fig. 4. In general,these data show the same effects on response rate and reinforcement frequency describedabove for Monkey M-12.A dramatically different picture emerged with Monkey M-197 under the same general ex-
perimental conditions, but with the self-stimulation electrode placement in the area of themedial forebrain bundle (Section M-197, Fig. 1). Figure 5 shows the IRT's for MonkeyM-197 throughout the 10-day alternating sugar pellet and brain-shock reinforcement periodand indicates that this animal maintained a remarkably stable and accurate time discrimina-tion under both conditions. For all 10 experimental sessions, the IRT's show a sharp peak atthe 20-second delay interval required for reinforcement, and neither the lever-pressing re-sponse rates nor the "reinforcement-per-response" ratios appear to differ significantly underthe two conditions. Clearly, an effect on timing behavior does not appear to be a necessaryconsequence either of electrical stimulation of brain tissue per se or of the reinforcingproperties of such brain stimulation. Indeed, these findings suggest that the particular tissueregions involved in the electrical stimulation play a critical role in the determination ofthe timing performance.
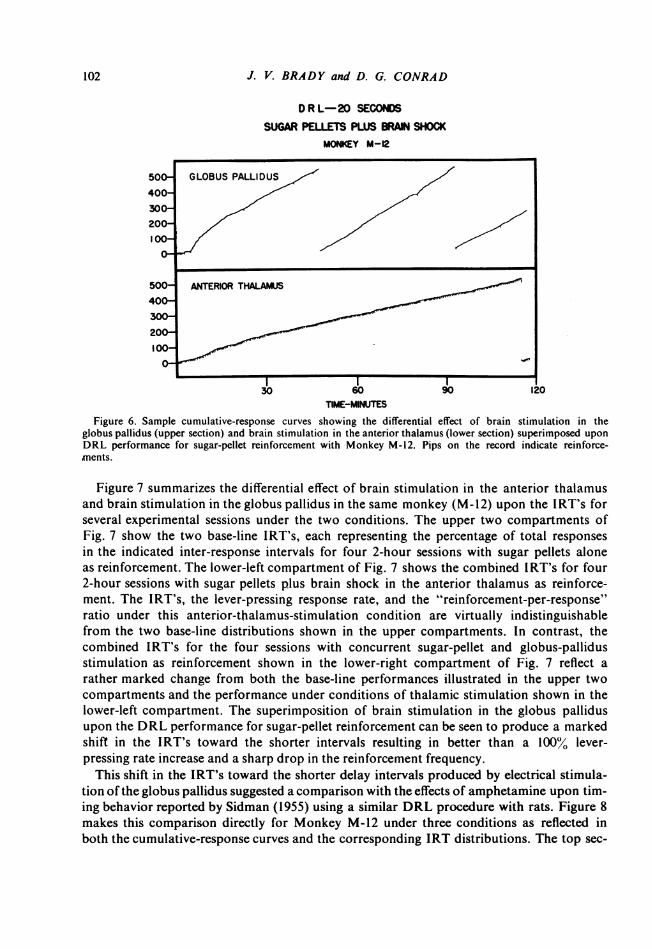
Additional data relevant to the neural "localization" of timing processes were obtainedfrom further experiments with Monkey M-12 involving the second electrode placement inthe anterior thalamus (upper left arrow, Section M-12, Fig. 1). Although electrical stimu-lation in this region did reinforce lever pressing, the effect on DRL performance was quitedifferent from that observed with the same animal working for brain shock in the globuspallidus (lower right arrow, Section M-12, Fig. 1). Performance on DRL 20 for the anteriorthalamus brain-shock reward did not produce the clear shift in IRT's to the 10-second peakseen in Fig. 3 with stimulation of the globus pallidus as reinforcement. The data obtainedwith Monkey M-12 under conditions of concurrent brain shock and sugar-pellet reinforce-ment shows further that this differential effect on the timing process is specifically relatedto the neural tissue regions affected by the electrical stimulus per se. Figure 6 shows theeffect on the lever-pressing rate of superimposing electrical stimulation of the globuspallidus and the anterior thalamus upon the sugar-pellet reinforcement during the DRL per-formance. When both sugar pellets and brain shock in the globus pallidus are delivered con-currently to reward the 20-second spaced responding, a marked elevation in the lever-press-ing rate and a relatively low frequency of reinforcements (pips on the cumulative curve) arethe result (upper portion of Fig. 6). In contrast, however, the DRL performance underconditions of concurrent sugar-pellet and anterior-thalamus-stimulation reinforcement(lower portion of Fig. 6) is quite similar to the performance recorded when sugar pelletsalone reinforce spaced responding in this same animal (upper portion of Fig. 2). Superim-posing electrical stimulation of the anterior thalamus upon sugar-pellet reward in thismonkey neither elevates the lever-pressing rate nor reduces the reinforcement frequency.
100

101TIMING BEHA VIOR
MONKEY M-197(MEDIAL FOREBRAIN BUNDLE)
DRL 20 SECONDS
35S30
25
20
I0-
lS0
25
20
Is
I0
i
F
SUGARPELLETS
10/23/574 6 RESP / MIN.REINF. 243E
. * 44RESP 549
O24
0 10
43
40
3S
30
25
F
20
.1
1015
00
43
40
35
SUGARfELLETS
10/25/5746 RESP/MIN.REINF. 282RESP 557
30
25
20.
IS5
IO.
S'
0-
BRAI N BRAINi-SHOCK 33 SHOCK
fi30-A10/24/57 3 10/26/573.7 RESP / MIN. 4.4 RESP / MIN.REINF. 212 REINF 193RESP 446 *RESP 525 -'25J
20
10
5 0
20 30 40. 0 10 20 30 40
F
0
SUGARPELLETS
410/27/57
4.9 RESP /MINREINF . 323.55RESP 590
5250
45
40
35
30
25S
20
IS.
15-10-
0-
BRAINSHOCK
30 10/218/574.4 RESP. / MIN.REINF 205RESP. 523 39
25
20
10 1aoi i ;
3230
25
2
1111
10
0. a02 304
SUGARPELLETS
4:1/29/57
54 RESP/MIN.REINF, 319,RESP 648
BRAINSH CK
1
10/30/574.3 RESP MIN.REINFP 203
REPi?'3
INTERRESPONSE TIMES (SECONDS)
Figure 5. Inter-response time distributions for Monkey M-197 during alternate-day, 2-hour DRL sessions.The upper distributions were recorded during sessions for sugar-pellet reinforcement, and the lower distribu-tions were recorded during sessions for brain-shock reward through the electrodes in the area of the medianforebrain bundle. Additional numerical data recorded above each distribution are explained in the text.
SUGARPELLETS
10/21/5748 RESP / MIN.REINF 219
-._ * 37RESP R81
U35
3
25
20
Is
10
5
o
ImanZ W,,
W t)D Za C)
IL1 Li>1C(
"-_-
It OU BRAIN
SHOCK
30 10/22/57
4. RESP. / MIN,REINF. 206RESp. - 42
25
20
1510
5 f
O 10 20 30 40
5
-
!O
1-
I
2
J. V. BRADY and D. G. CONRAD
D R L-20 SECONDS
SUGAR PELLETS PLUS BRAIN SHOCKMONEY M-12
50 GLOBUS PALLIDUS
100-500 ANTERIOR THALAMUS
30 60 90 120TIME-MINUTES
Figure 6. Sample cumulative-response curves showing the differential effect of brain stimulation in theglobus pallidus (upper section) and brain stimulation in the anterior thalamus (lower section) superimposed uponDRL performance for sugar-pellet reinforcement with Monkey M-12. Pips on the record indicate reinforce-ments.
Figure 7 summarizes the differential effect of brain stimulation in the anterior thalamusand brain stimulation in the globus pallidus in the same monkey (M-12) upon the IRT's forseveral experimental sessions under the two conditions. The upper two compartments ofFig. 7 show the two base-line IRT's, each representing the percentage of total responsesin the indicated inter-response intervals for four 2-hour sessions with sugar pellets aloneas reinforcement. The lower-left compartment of Fig. 7 shows the combined IRT's for four2-hour sessions with sugar pellets plus brain shock in the anterior thalamus as reinforce-ment. The IRT's, the lever-pressing response rate, and the "reinforcement-per-response"ratio under this anterior-thalamus-stimulation condition are virtually indistinguishablefrom the two base-line distributions shown in the upper compartments. In contrast, thecombined IRT's for the four sessions with concurrent sugar-pellet and globus-pallidusstimulation as reinforcement shown in the lower-right compartment of Fig. 7 reflect arather marked change from both the base-line performances illustrated in the upper twocompartments and the performance under conditions of thalamic stimulation shown in thelower-left compartment. The superimposition of brain stimulation in the globus pallidusupon the DRL performance for sugar-pellet reinforcement can be seen to produce a markedshift in the IRT's toward the shorter intervals resulting in better than a 100% lever-pressing rate increase and a sharp drop in the reinforcement frequency.This shift in the IRT's toward the shorter delay intervals produced by electrical stimula-
tion of the globus pallidus suggested a comparison with the effects of amphetamine upon tim-ing behavior reported by Sidman (1955) using a similar DRL procedure with rats. Figure 8makes this comparison directly for Monkey M-12 under three conditions as reflected inboth the cumulative-response curves and the corresponding IRT distributions. The top sec-
102

TIMING BEHA VIOR 103
MONKEY M- 12DRL 20 SECONDS
INTERRESPONSE TIMES ( SECONDS)
Figure 7. Inter-response time distributions for Monkey M-12 under conditions of sugar-pellet reinforce-ment alone (upper distributions), sugar pellets plus brain shock in the anterior thalamus as reinforcement(lower-left distribution), and sugar pellets plus brain shock in the globus pallidus as reinforcement (lower-right distribution). Each distribution represents the combined percentage of total responses in the indicatedinter-response intervals for four separate sessions. Additional numerical data recorded above each distributionare explained in the text.
tion of Fig. 8 shows a typically stable DRL performance for sugar pellets alone as rein-forcement: the lever-pressing rate is appropriately low, the reinforcement frequency is rel-atively high, and the IRT's show a peak around the 20-second delay interval. In contrast,the DRL performance for sugar pellets plus electrical stimulation of the globus pallidus as
reinforcement shown in the middle section of Fig. 8 reflects the now familiar lever-pressing-rate increase, drop in reinforcement frequency, and shift in IRT's to the shorter delay in-tervals. Similarly, the lowest section of Fig. 8 shows the DRL performance for this same
44
I*. .-0.J
ILz
20 SUAR |20 SUGARPELAES A PELLET AUG
2~~~ ~ ~~~~~ ~j 24
6~~~~~~~~~~~~~~~~~~~~1
15 3.9 RESP/MWN. 15 5.1 RESP/MIN.
RENF. 136 RN 902.45-
N~~~~~~~ E WP iW3" 3
0 ~~~~~~~~~~~~~~~0
O 10 20 30 40 0 10 20 30 40
44 1SUGAR PELLETS 42 SUGAR PELLETS
20hAAMUS
AG20 RANO GLOBUS AUGBRAlSIOCK ITHAL 3 BRAINgSHOCK PALLIOUS5 5
7 17
15 4.6 REIP/MIN. 15 12.7 IESP/MIN.NEIW 671 RE 4
NE~~~~~~~~~~~iSP 6114
o o
O 10 t0 30 40 O 10 D w 000 10 203040 ~~~~0 10 20 30 40
L
r 57

J. V. BRADY and D. G. CONRAD
DRL-20 SECONDSMONKEY M-12
C,)w)z0Q-C/)w
LU
w-J
TIME-MINUTESFigure 8. Sample cumulative-response curves and inter-response time distributions recorded under conditions
of sugar-pellet reinforcement alone (top section), sugar pellets plus brain shock in the globus pallidus as rein-forcement (middle section), and sugar-pellet reinforcement plus amphetamine (bottom section). Pips on therecord indicate reinforcements. Additional numerical data recorded above each distribution are explained in thetext.
monkey recorded during a 2-hour experimental session for sugar pellets alone following in-traperitoneal administration of 2 milligrams per kilogram of amphetamine sulfate. A com-parison between the two lower sections of Fig. 8 reveals the striking similarity in lever-pressing rates, reinforcement frequencies, and IRT distributions recorded under the twoconditions of "sugar pellets plus brain shock" and "sugar pellets plus amphetamine."These two operations, electrical stimulation of the globus pallidus and administration ofamphetamine, clearly produce similar effects upon timing behavior under the conditions ofthis experiment. However, only further experimental analysis can determine whether or notthese effects are produced in the same way by these two independent operations.One further comparison between the effects of brain stimulation and of food reinforce-
ment was provided by an experiment with Monkey M-197 performing on a variable-interval
104

TIMING BEHA VIOR
schedule. Figure 9 shows the cumulative curves recorded for this animal during the two1-hour experimental sessions on the same VI schedule (average 60 seconds). The top sectionof Fig. 9 shows the VI performance with sugar pellets alone as reinforcement, and the bot-tom section shows the VI performance for brain-shock reinforcement alone in the area of
M-197
500 60 SEC. VI-SUGAR PELLETS
400
300
200LiJU) 100z0 3Uf)
Vlprfoman f r Mo60 SEC. VI-ICS-MFB
500
0
0 15 30 45 60
TIME IN MINUTESFigure 9. Sample cumulative-response curves showing the differential effect of reinforcement with sugar pel-
lets (upper section) and with brain stimulation in the area of the median forebrain bundle (lower section) uponVI performance for Monkey M-197. Pips on the record indicate reinforcements.
the medial forebrain bundle. The sharp rate increase illustrated in the lower section ofFig. 9 is similar to that observed with Monkeys M-2 and M-12, and indicates that the ef-fects of brain stimulation upon the response rate are not specific to the DRL schedule. It isalso clear, however, that these effects of brain stimulation upon the response rate do notnecessarily reflect the same process under the two different reinforcement-schedule condi-tions. Monkey M-197 showed little or no change in the base-line DRL performance (Fig. 5)with the same electrical stimulation reinforcement in the area of the median forebrainbundle which produced the rate increase during the VI performance shown in Fig. 9. Un-fortunately, Monkeys M-2 and M-12 were not available for similar experiments on the ef-fects of brain stimulation upon VI performance. Comparison of the effects upon both VIand DRL performance with an electrical brain stimulus producing temporal effects on theDRL might well provide valuable additional data on the neural-behavioral relationshipsunder experimental analysis.
105

J. V. BRADY and D. G. CONRAD
REFERENCES
Anger, D. The dependence of interresponse times upon the relative reinforcement of different interresponsetimes. J. exp. Psychol., 1956, 52, 145-161.
Brady, J. V. Emotional behavior and the nervous system. Trans. N. Y. A cad. Sci., 1956, 18, 601-612.Brady, J. V. The paleocortex and behavioral motivation. In H. F. Harlow and C. N. Woolsey (Eds.) Biological and
Biochemical Bases of Behavior. Madison: Univer. Wisconsin Press, 1958.Conrad, D. G., Sidman, M., and Herrnstein, R. J. The effects of deprivation upon temporally spaced respond-
ing. J. exper. anal. Behav., 1958, 1, 59-65.Lilly, J. C., Hughes, J. R., Alvord, E. C., and Galkin, T. W. Brief.non-injurious electric waveform for stimula-
tion of the brain. Science, 1955, 121, 468.Mason, J. W. Restraining chair for the experimental study of primates, J. applied Physiol., 1958, 12, 130-133.Olds, J., and Milner, P. Positive reinforcement produced by electrical stimulation of septal area and other
regions of the rat brain. J. comp. physiol. Psychol., 1954, 47, 419-427.Sheatz, G. C. Multilead electrode holders in chronic preparations. In D. E. Sheer (Ed.) Electrical stimulation of
the brain: Subcortical integrative systems. Houston: Univer. Texas Press, 1959.Sidman, M. Technique for the assessment of drug effects on timing behavior. Science, 1955, 122, 925.Sidman, M., Brady, J. V., Boren, J. J., Conrad, D. G., and Schulman, A. Reward schedules and behavior main-
tained by intracranial self-stimulation. Science, 1955, 22, 830-831.Skinner, B. F. The behavior of organisms. New York: Appleton Century Co., 1938.Wilson, M. P., and Keller, F. S. On the selective reinforcement of spaced responses. J. comp. physiol. Psychol.,
1953, 46, 190-193.
Received November 19, 1959
106





























