Embed Size (px)
Citation preview

Site-directed Mutagenesis of Cys-15 and Cys-20 of PulmonarySurfactant Protein DEXPRESSION OF A TRIMERIC PROTEIN WITH ALTERED ANTI-VIRAL PROPERTIES*
(Received for publication, December 4, 1995, and in revised form, February 27, 1996)
Patricia Brown-Augsburger‡, Kevan Hartshorn¶, Donald Chang‡, Kevin Rust‡,Catherine Fliszar§, Howard G. Welgus§, and Edmond C. Crouch‡i
From the ‡Department of Pathology and §Division of Dermatology, Washington University School of Medicine,St. Louis, Missouri 63110 and the ¶Departments of Medicine and Pathology, Boston UniversitySchool of Medicine, Boston, Massachusetts 02118
Surfactant protein D (SP-D) molecules are preferen-tially assembled as dodecamers consisting of trimericsubunits associated at their amino termini. The NH2-terminal sequence of each monomer contains two con-served cysteine residues, which participate in inter-chain disulfide bonds. In order to study the roles ofthese residues in SP-D assembly and function, we em-ployed site-directedmutagenesis to substitute serine forcysteine 15 and 20 in recombinant rat SP-D (RrSP-D),and have expressed the mutant (RrSP-Dser15/20) in Chi-nese hamster ovary (CHO-K1) cells. The mutant, whichwas efficiently secreted, bound to maltosyl-agarose, butunlike RrSP-D, was assembled exclusively as trimers.The constituent monomers showed a decreased mobilityon SDS-polyacrylamide gel electrophoresis resultingfrom an increase in the size and sialylation of the N-linked oligosaccharide at Asn-70. Although RrSP-Dser15/20 contained a pepsin-resistant triple helical do-main, it showed a decreased Tm, and acquiredsusceptibility to proteolytic degradation. Like RrSP-D,RrSP-Dser15/20 bound to the hemagglutinin of influenzaA. However, it showed no viral aggregation and did notenhance the binding of influenza A to neutrophils(PMN), augment PMN respiratory burst, or protectPMNs from deactivation. These studies indicate thatamino-terminal disulfides are required to stabilize do-decamers, and support our hypothesis that the oligo-merization of trimeric subunits contributes to the anti-microbial properties of SP-D.
Surfactant protein D (SP-D)1 is a collagenous C-type lectin(collectin) that is secreted into the distal airways and alveoli ofthe lung (1, 2). Recent studies have demonstrated binding ofSP-D to a variety of bacterial, viral, and fungal pathogens (3–5)and effects of SP-D on leukocyte function in vitro (4, 6, 7). For
example, SP-D has been shown to enhance the binding ofinfluenza A to neutrophils, to inhibit the viral hemagglutininand inhibit infectivity in vitro, to potentiate the neutrophilrespiratory burst in response to bound virus, and to protectneutrophils from viral deactivation (4).Cloning and sequence analysis of SP-D has shown that the
43-kDa monomer has four major structural domains (8–11).The short, non-collagenous amino-terminal domain containstwo absolutely conserved cysteine residues (Cys-15 and Cys-20)involved in interchain disulfide bonding. The uninterruptedcollagenous domain contains 59 Gly-X-Y repeats (in rat andhuman), and is connected to the carboxyl-terminal carbohy-drate recognition domain (CRD) by a short linking sequence(L). A single N-linked glycosylation site at Asn-70 is foundwithin the collagenous domain, and studies of SP-D from nat-ural and recombinant sources have shown that this site isutilized and contains a sialylated sugar moiety (12, 13).Other collagenous C-type lectins include: pulmonary surfac-
tant protein A (SP-A), serum mannose-binding protein (MBP),bovine serum conglutinin, and bovine serum lectin 43 (CL-43)(14, 15). SP-D is predominantly assembled as a dodecamerconsisting of four homotrimeric subunits associated at theiramino termini (12, 16). Analysis of proteolytic fragments andlimited reduction and alkylation of SP-D dodecamers suggeststhat the amino-terminal cysteine residues form intra- and in-tertrimeric disulfide bonds, which stabilize the fully assembleddodecamer (12).Trimeric CRDs (but not monomers) show high affinity bind-
ing to various saccharide ligands in vitro (17). In addition,ultrastructural studies suggest that the rigid collagenous armsrestrict the spatial distribution of the lectin domains and allowfor bridging interactions between ligands separated by up to100 nm (8, 12). Accordingly, we hypothesized that amino-ter-minal cross-linking of trimeric subunits is critical for SP-D-mediated microbial agglutination. We further hypothesizedthat trimeric subunits of SP-D (i.e. single arms) will bind tomicroorganisms but show decreased agglutinating activity. Al-though SP-D “arms” can be generated by reduction and alkyl-ation, the intrachain bonds required for lectin activity are alsodisrupted (12). We therefore sought to test these hypotheses bymutating the amino-terminal cysteine residues in rat SP-D andutilizing a CHO K1 expression system. Previous work in ourlaboratory has shown that wild type recombinant rat SP-D(RrSP-D) produced by this system is chemically, morphologi-cally, and functionally indistinguishable from natural rat SP-D(13).Using this approach, we have produced a full-length mutant
SP-D molecule, RrSP-Dser15/20, that is secreted as a trimer.These studies indicate that amino-terminal disulfide bonds are
* This work was supported by National Institutes of Health GrantsHL29594, HL44015, and AR35805. The costs of publication of thisarticle were defrayed in part by the payment of page charges. Thisarticle must therefore be hereby marked “advertisement” in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
i To whom correspondence should be addressed: Dept. of Pathology,Jewish Hospital at Washington University Medical Center, 216 S.Kingshighway, St. Louis, MO 63110. Tel.: 314-454-8462; Fax: 314-454-5505.
1 The abbreviations used are: SP-D, surfactant protein D; EndoF,endoglycosidase F; CRD, carbohydrate recognition domain; MBP, man-nose-binding protein; PCR, polymerase chain reaction; CHO, Chinesehamster ovary; DMEM, Dulbecco’s modified Eagle’s medium; PAGE,polyacrylamide gel electrophoresis; PMSF, phenylmethylsulfonyl fluo-ride; MMP, matrilysin; TBS, Tris-buffered saline; IAV, influenza Avirus; FMLP, formylmethionylleucylphenylalanine.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 271, No. 23, Issue of June 7, pp. 13724–13730, 1996© 1996 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
13724
by guest on April 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

required to form or stabilize dodecamers, and provide supportfor our hypothesis that the oligomerization of trimeric subunitsto form dodecamers is required for some of the previouslyobserved interactions of SP-D with influenza A. Our resultsalso show that the cysteine substitutions decrease the thermalstability of the collagen helix and increase the protein’s suscep-tibility to proteolytic degradation.
MATERIALS AND METHODS
Site-directed Mutagenesis of rSP-D—Site-directed mutagenesis wasperformed on a full-length rat SP-D cDNA clone provided by Drs. J. H.Fisher and D. R. Voelker, Denver, CO (10), using the PCR overlapextension method (18). Primers for substituting serine for Cys-15 andCys-20 of the mature protein were: CAATAACCAACACGAGCACCC-TAGTCTTGAGTAGTCCAACAG (forward) and CTGTTGGACTACTC-AAGACTAGGGTGCTCGTGTTGGTTATTG (reverse). Sp6 and T7 wereused as outside primers. PCR reactions were performed using 400 ng ofScaI linearized rSP-DpGEM-3Z template. The reaction buffer con-tained 2 mM MgCl2, 2 mM dithiothreitol, and 5 units of Taq polymerase(Promega). Separate reactions containing the forward and T7 primersor reverse and Sp6 primers were performed for 15 cycles with a 45 °Cannealing temperature. The resulting products were gel-purified andcombined in a second PCR reaction for 10 cycles at 52 °C annealing,followed by 20 cycles at 42 °C annealing using Sp6 and T7 as primers.The ;1.3-kilobase pair DNA product was gel-purified, digested withEcoRI, and subcloned into pGEM-3Z. Clones were sequenced up to theApaI site within rSP-D to verify the presence of the desired mutationsand the absence of any additional point mutations. An EcoRI/ApaIfragment containing the mutated region was then subcloned into thecorresponding site in wild type rSP-DpGEM-3Z. Subcloning of the mu-tated region was performed in order to avoid introduction of extraneousTaq generated mutations.Expression of Rat SP-D cDNA Mutants in CHO K1—Mutated rSP-D
cDNAs were excised from pGEM-3Z with EcoRI and subcloned into thecorresponding site within the multiple cloning site of pEE14 (13, 19,20). Orientation of the subclones was determined by restriction map-ping and DNA sequencing. Transfection of pEE14 constructs into CHOK1 cells and selection of stably expressing cell lines was performed asdescribed previously (13).Metabolic Labeling and Purification of Recombinant SP-D—Conflu-
ent cultures were washed in serum-free DMEM and then incubated for16 h in DMEM containing 1% (v/v) dialyzed fetal calf serum, 50 mg/mlascorbic acid and 5 mCi/ml L-[14C]proline (283 mCi/mmol; DuPontNEN). Secreted recombinant protein was isolated from the culturemedium by sequential maltosyl-agarose and gel filtration chromatog-raphy as described previously (13). The elution of radiolabeled proteinwas monitored by liquid scintillation counting and/or SDS-PAGE andautoradiography.Immunoprecipitation—Cells were plated in six-well tissue culture
plates and grown to confluence. Metabolic labeling was performed withL-[14C]proline as described above or with Tran35S-label (ICN, 1255Ci/mmol). For labeling with Tran35S-label, cells were washed and pre-incubated for 30 min with DMEM deficient in cysteine and methionine,DMEM(2Cys/Met), supplemented with 50 mg/ml ascorbate. Cells werethen incubated in 1 ml of DMEM(2Cys/Met) plus 10 mCi/ml Tran35S-label for up to 8 h. Conditioned medium was harvested and centrifugedat 1000 3 g to pellet any cells or debris, and the supernatant was usedin subsequent steps. For analysis of intracellular protein, cell layerswere washed twice with cold DMEM and lysed with 1 ml of immuno-precipitation buffer (50 mM Tris, pH 7.5, 150 mM NaCl, 0.5% (w/v)deoxycholate, 1% (v/v) Triton X-100, 0.1% (w/v) SDS, 2.5 mM EDTA, 0.1mM PMSF) supplemented with 50 mM iodoacetamide to trap any freesulfhydryls. Cell lysates were incubated on ice in the dark for 30 minand centrifuged at 12,000 3 g to pellet cell debris. Immunoprecipita-tions were performed by mixing 200–800 ml of cell lysate or conditionedmedium with 500 ml of immunoprecipitation buffer and 5 ml of rabbitanti-rat SP-D antibody for 2 h at room temperature or overnight at 4 °C.Samples were then mixed with 300 ml of a 10% suspension of IgGsorb(The Enzyme Center, Malden, MA) for 1 h at room temperature. Aftercentrifugation at 12,000 3 g for 30 s, the supernatant was discardedand the pellets were washed three times with immunoprecipitationbuffer. Immunoprecipitates were resolved by 5/10% or 5/12% SDS-PAGE and visualized by fluorography.Degradative Analysis—Immune complexes were washed with dis-
tilled water and then resuspended in 50 ml of endoglycosidase F buffer(100 mM sodium phosphate, pH 6.1, 50 mM EDTA, 1% (v/v) b-mercap-toethanol, 0.1% (w/v) SDS, 1%(v/v) Nonidet P-40, 2 mM PMSF) or 50 ml
of neuraminidase buffer (100 mM sodium acetate, pH 5.5, 0.1%(v/v)Nonidet P-40, 0.2 mM PMSF, and 2 mM EDTA). Samples were boiledand then incubated overnight at 37 °C with 0.025 units of neuramini-dase (Genzyme) or 0.050 units of endoglycosidase F (Sigma). Collagen-ase and pepsin digestions were performed as described (21, 22).Thermal Stability Analysis—Thermal denaturation temperatures
were determined essentially as described (23). L-[3H]Proline-labeledwild type RrSP-D and L-[14C]proline-labeled RrSP-Dcys3ser15/20 werediluted in 100 mM Tris, 400 mM NaCl (pH 7.4), and 45 ml was added to200 ml microcentrifuge tubes. Two tubes of each sample were kept atroom temperature. The remaining tubes were placed in a Perkin-ElmerCetus thermal cycler at 30 °C and programmed to increase 2 °C every10 min between 30° and 44 °C. At the end of each 10-min plateau, onetube of each sample was removed, quenched at 20 °C for 30 s, anddigested with 5 ml of 1 mg/ml L-1-tosylamido-2-phenylethyl chloro-methyl ketone-treated trypsin (Sigma, 12,000 units/mg) for 2 min at20 °C. Digestion was stopped by addition of reducing SDS-PAGE sam-ple buffer containing PMSF, followed by boiling.Protease Susceptibility—The susceptibility of wild type SP-D and
RrSP-Dser15/20 to proteolytic degradation was further tested using avariety of highly purified proteinases including human leukocyte elas-tase (Elastin Products, Owensville, MO), human matrilysin (MMP-7),human 72-kDa (MMP-2) and 92-kDa (MMP-9) gelatinases, human in-terstitial collagenase (MMP-1), rat interstitial collagenase (MMP-13),stromelysin (MMP-3), and human and mouse metalloelastases (MMP-12). Human matrilysin, 92-kDa gelatinase, interstitial collagenase,metalloelastase, and mouse metalloelastase were expressed and puri-fied as we have reported previously (24–27). Human 72-kDa gelatinasefree of TIMP-2 was kindly provided by William Stetler-Stevenson (Na-tional Cancer Institute), rat interstitial collagenase was supplied byJohn J. Jeffrey (Albany Medical College), and human stromelysin cat-alytic domain was obtained from Qi-Zhuang Ye (Parke-Davis, AnnArbor, MI).To test susceptibility to proteolytic degradation, wild type SP-D and
the Cys mutant (1.5 mg each) were incubated with 1 3 1026 M of eachenzyme in a total mixture volume of 50 ml (buffer 5 0.05 M Tris, pH 7.5,0.01 M CaCl2, 0.15 M NaCl, 0.02% Brij) at 37 °C overnight. The reactionswere terminated by the addition of 1 mM o-phenanthroline for themetalloproteinases and 1 mM PMSF for leukocyte elastase. The mix-tures were then boiled in reducing sample buffer. Proteins were re-solved by SDS-PAGE and visualized by silver staining.Preparation of Virus—Virus stocks were grown in the chorioallantoic
fluid of 10-day-old embryonated hen’s eggs, purified on a discontinuoussucrose density gradient, stored, and titered as described previously(28). The A/Texas 77/H3N2 (Texas 77) and A/PR/8/34/H1N1 (PR8)strains of IAV were gracious gifts of Dr. Jon Abramson (Bowman-GraySchool of Medicine, Winston-Salem, NC). The A/Mem71H-BelN (H3N1)and its glycosylation mutant derivative strain Mem71H-BelN/BS werekindly provided by Dr. E. Margot Anders (University of Melbourne,Melbourne, Australia) (29). Incubation of SP-Ds with IAV stocks wascarried out at 37 °C in TBS with 2 mM Ca21 except in indicated exper-iments in which maltose was added, or in which the buffer contained 10mM EDTA and no added Ca21 (28).Assessment of Viral Aggregation—Aggregation was assessed by
measuring changes in light transmission through suspensions of IAVafter addition of various concentrations of SP-D (28). Addition of SP-Dalone in the presence of 2 mM Ca21 did not cause any decrease in lighttransmission.Measurement of Neutrophil Activation and Protection—Neutrophils
(PMN) from healthy volunteer donors were isolated to .95% purityusing dextran sedimentation, followed by a Ficoll-Hypaque gradientcentrifugation for removal of mononuclear cells, and hypotonic lysis toremove contaminating erythrocytes (28). H2O2 production was meas-ured by the oxidation of scopolectin, and O2
. assessed by monitoring thesuperoxide dismutase inhibitable reduction of cytochrome c (30). PMNprotection assays measured the superoxide response to FMLP (5 3 1027
M) after exposure to IAV preincubated in the absence or presence ofSP-D (4, 31, 32).
RESULTS
Construction and Expression of RrSP-D Site-directed Mu-tants—Sequence analysis of the constructs showed that we hadgenerated the expected Cys-15/Cys-20 double substitution mu-tant, designated RrSP-Dser15/20. RrSP-Dser15/20 was effi-ciently secreted into the culture medium, and bound quantita-tively to maltosyl-agarose (Figs. 1 and 2). The recovery wascomparable to that obtained for wild type SP-D under compa-
Surfactant Protein D 13725
by guest on April 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from
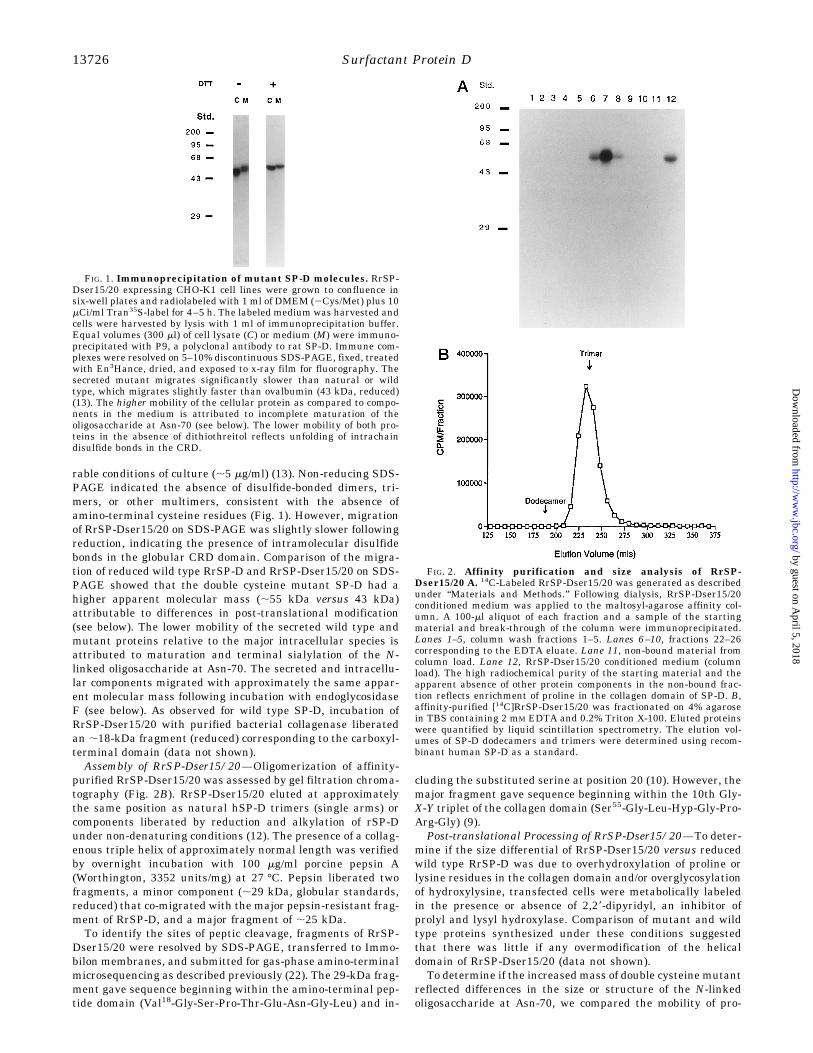
rable conditions of culture (;5 mg/ml) (13). Non-reducing SDS-PAGE indicated the absence of disulfide-bonded dimers, tri-mers, or other multimers, consistent with the absence ofamino-terminal cysteine residues (Fig. 1). However, migrationof RrSP-Dser15/20 on SDS-PAGE was slightly slower followingreduction, indicating the presence of intramolecular disulfidebonds in the globular CRD domain. Comparison of the migra-tion of reduced wild type RrSP-D and RrSP-Dser15/20 on SDS-PAGE showed that the double cysteine mutant SP-D had ahigher apparent molecular mass (;55 kDa versus 43 kDa)attributable to differences in post-translational modification(see below). The lower mobility of the secreted wild type andmutant proteins relative to the major intracellular species isattributed to maturation and terminal sialylation of the N-linked oligosaccharide at Asn-70. The secreted and intracellu-lar components migrated with approximately the same appar-ent molecular mass following incubation with endoglycosidaseF (see below). As observed for wild type SP-D, incubation ofRrSP-Dser15/20 with purified bacterial collagenase liberatedan ;18-kDa fragment (reduced) corresponding to the carboxyl-terminal domain (data not shown).Assembly of RrSP-Dser15/20—Oligomerization of affinity-
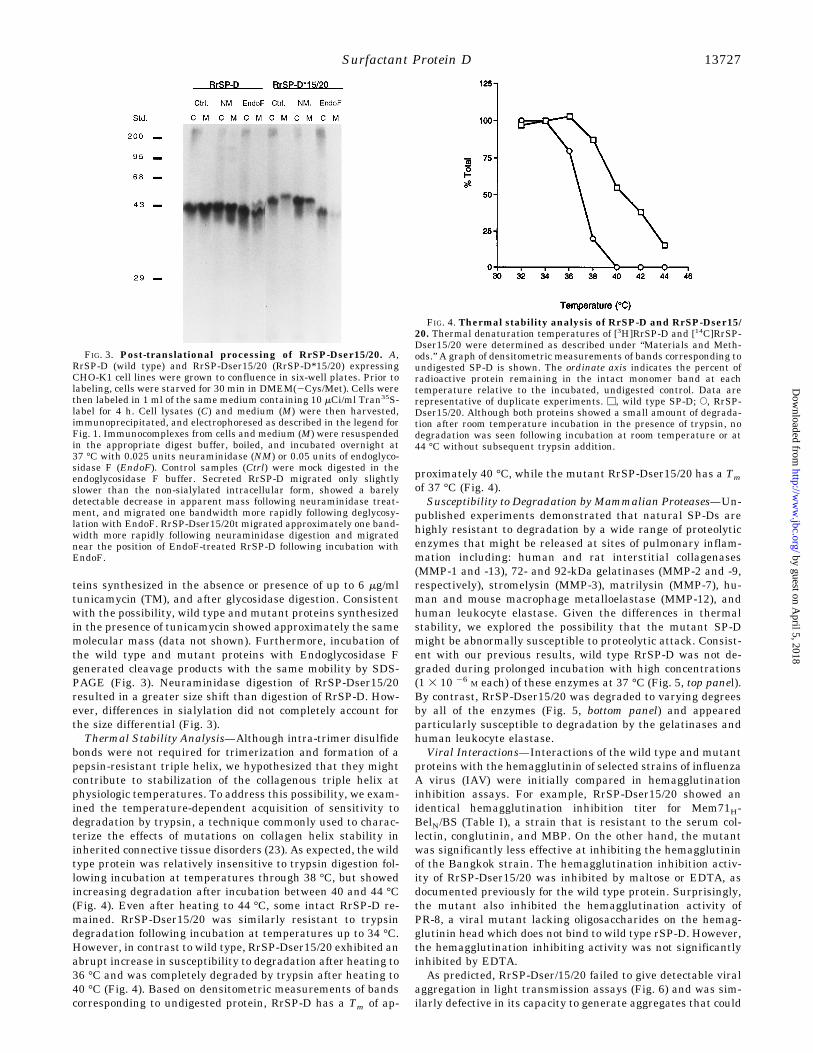
purified RrSP-Dser15/20 was assessed by gel filtration chroma-tography (Fig. 2B). RrSP-Dser15/20 eluted at approximatelythe same position as natural hSP-D trimers (single arms) orcomponents liberated by reduction and alkylation of rSP-Dunder non-denaturing conditions (12). The presence of a collag-enous triple helix of approximately normal length was verifiedby overnight incubation with 100 mg/ml porcine pepsin A(Worthington, 3352 units/mg) at 27 °C. Pepsin liberated twofragments, a minor component (;29 kDa, globular standards,reduced) that co-migrated with the major pepsin-resistant frag-ment of RrSP-D, and a major fragment of ;25 kDa.To identify the sites of peptic cleavage, fragments of RrSP-
Dser15/20 were resolved by SDS-PAGE, transferred to Immo-bilon membranes, and submitted for gas-phase amino-terminalmicrosequencing as described previously (22). The 29-kDa frag-ment gave sequence beginning within the amino-terminal pep-tide domain (Val18-Gly-Ser-Pro-Thr-Glu-Asn-Gly-Leu) and in-
cluding the substituted serine at position 20 (10). However, themajor fragment gave sequence beginning within the 10th Gly-X-Y triplet of the collagen domain (Ser55-Gly-Leu-Hyp-Gly-Pro-Arg-Gly) (9).Post-translational Processing of RrSP-Dser15/20—To deter-
mine if the size differential of RrSP-Dser15/20 versus reducedwild type RrSP-D was due to overhydroxylation of proline orlysine residues in the collagen domain and/or overglycosylationof hydroxylysine, transfected cells were metabolically labeledin the presence or absence of 2,29-dipyridyl, an inhibitor ofprolyl and lysyl hydroxylase. Comparison of mutant and wildtype proteins synthesized under these conditions suggestedthat there was little if any overmodification of the helicaldomain of RrSP-Dser15/20 (data not shown).To determine if the increased mass of double cysteine mutant
reflected differences in the size or structure of the N-linkedoligosaccharide at Asn-70, we compared the mobility of pro-
FIG. 1. Immunoprecipitation of mutant SP-D molecules. RrSP-Dser15/20 expressing CHO-K1 cell lines were grown to confluence insix-well plates and radiolabeled with 1 ml of DMEM (2Cys/Met) plus 10mCi/ml Tran35S-label for 4–5 h. The labeled medium was harvested andcells were harvested by lysis with 1 ml of immunoprecipitation buffer.Equal volumes (300 ml) of cell lysate (C) or medium (M) were immuno-precipitated with P9, a polyclonal antibody to rat SP-D. Immune com-plexes were resolved on 5–10% discontinuous SDS-PAGE, fixed, treatedwith En3Hance, dried, and exposed to x-ray film for fluorography. Thesecreted mutant migrates significantly slower than natural or wildtype, which migrates slightly faster than ovalbumin (43 kDa, reduced)(13). The higher mobility of the cellular protein as compared to compo-nents in the medium is attributed to incomplete maturation of theoligosaccharide at Asn-70 (see below). The lower mobility of both pro-teins in the absence of dithiothreitol reflects unfolding of intrachaindisulfide bonds in the CRD.
FIG. 2. Affinity purification and size analysis of RrSP-Dser15/20 A. 14C-Labeled RrSP-Dser15/20 was generated as describedunder “Materials and Methods.” Following dialysis, RrSP-Dser15/20conditioned medium was applied to the maltosyl-agarose affinity col-umn. A 100-ml aliquot of each fraction and a sample of the startingmaterial and break-through of the column were immunoprecipitated.Lanes 1–5, column wash fractions 1–5. Lanes 6–10, fractions 22–26corresponding to the EDTA eluate. Lane 11, non-bound material fromcolumn load. Lane 12, RrSP-Dser15/20 conditioned medium (columnload). The high radiochemical purity of the starting material and theapparent absence of other protein components in the non-bound frac-tion reflects enrichment of proline in the collagen domain of SP-D. B,affinity-purified [14C]RrSP-Dser15/20 was fractionated on 4% agarosein TBS containing 2 mM EDTA and 0.2% Triton X-100. Eluted proteinswere quantified by liquid scintillation spectrometry. The elution vol-umes of SP-D dodecamers and trimers were determined using recom-binant human SP-D as a standard.
Surfactant Protein D13726
by guest on April 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from
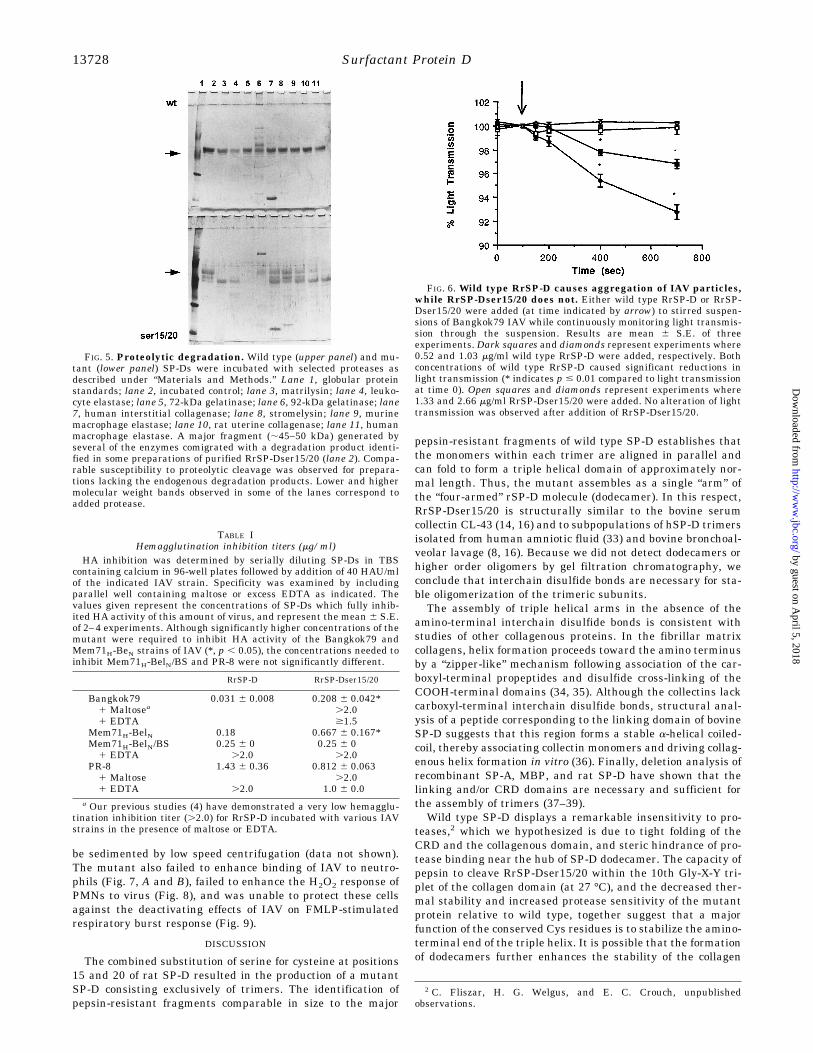
teins synthesized in the absence or presence of up to 6 mg/mltunicamycin (TM), and after glycosidase digestion. Consistentwith the possibility, wild type and mutant proteins synthesizedin the presence of tunicamycin showed approximately the samemolecular mass (data not shown). Furthermore, incubation ofthe wild type and mutant proteins with Endoglycosidase Fgenerated cleavage products with the same mobility by SDS-PAGE (Fig. 3). Neuraminidase digestion of RrSP-Dser15/20resulted in a greater size shift than digestion of RrSP-D. How-ever, differences in sialylation did not completely account forthe size differential (Fig. 3).Thermal Stability Analysis—Although intra-trimer disulfide
bonds were not required for trimerization and formation of apepsin-resistant triple helix, we hypothesized that they mightcontribute to stabilization of the collagenous triple helix atphysiologic temperatures. To address this possibility, we exam-ined the temperature-dependent acquisition of sensitivity todegradation by trypsin, a technique commonly used to charac-terize the effects of mutations on collagen helix stability ininherited connective tissue disorders (23). As expected, the wildtype protein was relatively insensitive to trypsin digestion fol-lowing incubation at temperatures through 38 °C, but showedincreasing degradation after incubation between 40 and 44 °C(Fig. 4). Even after heating to 44 °C, some intact RrSP-D re-mained. RrSP-Dser15/20 was similarly resistant to trypsindegradation following incubation at temperatures up to 34 °C.However, in contrast to wild type, RrSP-Dser15/20 exhibited anabrupt increase in susceptibility to degradation after heating to36 °C and was completely degraded by trypsin after heating to40 °C (Fig. 4). Based on densitometric measurements of bandscorresponding to undigested protein, RrSP-D has a Tm of ap-
proximately 40 °C, while the mutant RrSP-Dser15/20 has a Tmof 37 °C (Fig. 4).Susceptibility to Degradation by Mammalian Proteases—Un-
published experiments demonstrated that natural SP-Ds arehighly resistant to degradation by a wide range of proteolyticenzymes that might be released at sites of pulmonary inflam-mation including: human and rat interstitial collagenases(MMP-1 and -13), 72- and 92-kDa gelatinases (MMP-2 and -9,respectively), stromelysin (MMP-3), matrilysin (MMP-7), hu-man and mouse macrophage metalloelastase (MMP-12), andhuman leukocyte elastase. Given the differences in thermalstability, we explored the possibility that the mutant SP-Dmight be abnormally susceptible to proteolytic attack. Consist-ent with our previous results, wild type RrSP-D was not de-graded during prolonged incubation with high concentrations(1 3 10 26 M each) of these enzymes at 37 °C (Fig. 5, top panel).By contrast, RrSP-Dser15/20 was degraded to varying degreesby all of the enzymes (Fig. 5, bottom panel) and appearedparticularly susceptible to degradation by the gelatinases andhuman leukocyte elastase.Viral Interactions—Interactions of the wild type and mutant
proteins with the hemagglutinin of selected strains of influenzaA virus (IAV) were initially compared in hemagglutinationinhibition assays. For example, RrSP-Dser15/20 showed anidentical hemagglutination inhibition titer for Mem71H-BelN/BS (Table I), a strain that is resistant to the serum col-lectin, conglutinin, and MBP. On the other hand, the mutantwas significantly less effective at inhibiting the hemagglutininof the Bangkok strain. The hemagglutination inhibition activ-ity of RrSP-Dser15/20 was inhibited by maltose or EDTA, asdocumented previously for the wild type protein. Surprisingly,the mutant also inhibited the hemagglutination activity ofPR-8, a viral mutant lacking oligosaccharides on the hemag-glutinin head which does not bind to wild type rSP-D. However,the hemagglutination inhibiting activity was not significantlyinhibited by EDTA.As predicted, RrSP-Dser/15/20 failed to give detectable viral
aggregation in light transmission assays (Fig. 6) and was sim-ilarly defective in its capacity to generate aggregates that could
FIG. 3. Post-translational processing of RrSP-Dser15/20. A,RrSP-D (wild type) and RrSP-Dser15/20 (RrSP-D*15/20) expressingCHO-K1 cell lines were grown to confluence in six-well plates. Prior tolabeling, cells were starved for 30 min in DMEM(2Cys/Met). Cells werethen labeled in 1 ml of the same medium containing 10 mCi/ml Tran35S-label for 4 h. Cell lysates (C) and medium (M) were then harvested,immunoprecipitated, and electrophoresed as described in the legend forFig. 1. Immunocomplexes from cells and medium (M) were resuspendedin the appropriate digest buffer, boiled, and incubated overnight at37 °C with 0.025 units neuraminidase (NM) or 0.05 units of endoglyco-sidase F (EndoF). Control samples (Ctrl) were mock digested in theendoglycosidase F buffer. Secreted RrSP-D migrated only slightlyslower than the non-sialylated intracellular form, showed a barelydetectable decrease in apparent mass following neuraminidase treat-ment, and migrated one bandwidth more rapidly following deglycosy-lation with EndoF. RrSP-Dser15/20t migrated approximately one band-width more rapidly following neuraminidase digestion and migratednear the position of EndoF-treated RrSP-D following incubation withEndoF.
FIG. 4. Thermal stability analysis of RrSP-D and RrSP-Dser15/20. Thermal denaturation temperatures of [3H]RrSP-D and [14C]RrSP-Dser15/20 were determined as described under “Materials and Meth-ods.” A graph of densitometric measurements of bands corresponding toundigested SP-D is shown. The ordinate axis indicates the percent ofradioactive protein remaining in the intact monomer band at eachtemperature relative to the incubated, undigested control. Data arerepresentative of duplicate experiments. M, wild type SP-D; E, RrSP-Dser15/20. Although both proteins showed a small amount of degrada-tion after room temperature incubation in the presence of trypsin, nodegradation was seen following incubation at room temperature or at44 °C without subsequent trypsin addition.
Surfactant Protein D 13727
by guest on April 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from
be sedimented by low speed centrifugation (data not shown).The mutant also failed to enhance binding of IAV to neutro-phils (Fig. 7, A and B), failed to enhance the H2O2 response ofPMNs to virus (Fig. 8), and was unable to protect these cellsagainst the deactivating effects of IAV on FMLP-stimulatedrespiratory burst response (Fig. 9).
DISCUSSION
The combined substitution of serine for cysteine at positions15 and 20 of rat SP-D resulted in the production of a mutantSP-D consisting exclusively of trimers. The identification ofpepsin-resistant fragments comparable in size to the major
pepsin-resistant fragments of wild type SP-D establishes thatthe monomers within each trimer are aligned in parallel andcan fold to form a triple helical domain of approximately nor-mal length. Thus, the mutant assembles as a single “arm” ofthe “four-armed” rSP-D molecule (dodecamer). In this respect,RrSP-Dser15/20 is structurally similar to the bovine serumcollectin CL-43 (14, 16) and to subpopulations of hSP-D trimersisolated from human amniotic fluid (33) and bovine bronchoal-veolar lavage (8, 16). Because we did not detect dodecamers orhigher order oligomers by gel filtration chromatography, weconclude that interchain disulfide bonds are necessary for sta-ble oligomerization of the trimeric subunits.The assembly of triple helical arms in the absence of the
amino-terminal interchain disulfide bonds is consistent withstudies of other collagenous proteins. In the fibrillar matrixcollagens, helix formation proceeds toward the amino terminusby a “zipper-like” mechanism following association of the car-boxyl-terminal propeptides and disulfide cross-linking of theCOOH-terminal domains (34, 35). Although the collectins lackcarboxyl-terminal interchain disulfide bonds, structural anal-ysis of a peptide corresponding to the linking domain of bovineSP-D suggests that this region forms a stable a-helical coiled-coil, thereby associating collectin monomers and driving collag-enous helix formation in vitro (36). Finally, deletion analysis ofrecombinant SP-A, MBP, and rat SP-D have shown that thelinking and/or CRD domains are necessary and sufficient forthe assembly of trimers (37–39).Wild type SP-D displays a remarkable insensitivity to pro-
teases,2 which we hypothesized is due to tight folding of theCRD and the collagenous domain, and steric hindrance of pro-tease binding near the hub of SP-D dodecamer. The capacity ofpepsin to cleave RrSP-Dser15/20 within the 10th Gly-X-Y tri-plet of the collagen domain (at 27 °C), and the decreased ther-mal stability and increased protease sensitivity of the mutantprotein relative to wild type, together suggest that a majorfunction of the conserved Cys residues is to stabilize the amino-terminal end of the triple helix. It is possible that the formationof dodecamers further enhances the stability of the collagen
2 C. Fliszar, H. G. Welgus, and E. C. Crouch, unpublishedobservations.
FIG. 5. Proteolytic degradation. Wild type (upper panel) and mu-tant (lower panel) SP-Ds were incubated with selected proteases asdescribed under “Materials and Methods.” Lane 1, globular proteinstandards; lane 2, incubated control; lane 3, matrilysin; lane 4, leuko-cyte elastase; lane 5, 72-kDa gelatinase; lane 6, 92-kDa gelatinase; lane7, human interstitial collagenase; lane 8, stromelysin; lane 9, murinemacrophage elastase; lane 10, rat uterine collagenase; lane 11, humanmacrophage elastase. A major fragment (;45–50 kDa) generated byseveral of the enzymes comigrated with a degradation product identi-fied in some preparations of purified RrSP-Dser15/20 (lane 2). Compa-rable susceptibility to proteolytic cleavage was observed for prepara-tions lacking the endogenous degradation products. Lower and highermolecular weight bands observed in some of the lanes correspond toadded protease.
TABLE IHemagglutination inhibition titers (mg/ml)
HA inhibition was determined by serially diluting SP-Ds in TBScontaining calcium in 96-well plates followed by addition of 40 HAU/mlof the indicated IAV strain. Specificity was examined by includingparallel well containing maltose or excess EDTA as indicated. Thevalues given represent the concentrations of SP-Ds which fully inhib-ited HA activity of this amount of virus, and represent the mean 6 S.E.of 2–4 experiments. Although significantly higher concentrations of themutant were required to inhibit HA activity of the Bangkok79 andMem71H-BeN strains of IAV (*, p , 0.05), the concentrations needed toinhibit Mem71H-BelN/BS and PR-8 were not significantly different.
RrSP-D RrSP-Dser15/20
Bangkok79 0.031 6 0.008 0.208 6 0.042*1 Maltosea .2.01 EDTA $1.5
Mem71H-BelN 0.18 0.667 6 0.167*Mem71H-BelN/BS 0.25 6 0 0.25 6 0
1 EDTA .2.0 .2.0PR-8 1.43 6 0.36 0.812 6 0.063
1 Maltose .2.01 EDTA .2.0 1.0 6 0.0
a Our previous studies (4) have demonstrated a very low hemagglu-tination inhibition titer (.2.0) for RrSP-D incubated with various IAVstrains in the presence of maltose or EDTA.
FIG. 6. Wild type RrSP-D causes aggregation of IAV particles,while RrSP-Dser15/20 does not. Either wild type RrSP-D or RrSP-Dser15/20 were added (at time indicated by arrow) to stirred suspen-sions of Bangkok79 IAV while continuously monitoring light transmis-sion through the suspension. Results are mean 6 S.E. of threeexperiments. Dark squares and diamonds represent experiments where0.52 and 1.03 mg/ml wild type RrSP-D were added, respectively. Bothconcentrations of wild type RrSP-D caused significant reductions inlight transmission (* indicates p # 0.01 compared to light transmissionat time 0). Open squares and diamonds represent experiments where1.33 and 2.66 mg/ml RrSP-Dser15/20 were added. No alteration of lighttransmission was observed after addition of RrSP-Dser15/20.
Surfactant Protein D13728
by guest on April 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from
domain or limits accessibility of proteases to cleavage sitesnear the amino terminus in RrSP-D. Although the measuredTm for the wild type protein is consistent with the formation ofa stable triple helix in vivo, the melting temperature of themutant (Tm 37 °C) suggests that its helical domain is incom-pletely folded in the cell. To our knowledge, these studies
provide the first demonstration of stabilization of a contiguouscollagenous helix by interchain disulfide bonds. We speculatethat the amino-terminal and intrahelical cysteine residuesfound in other collectins play a similar role.To our surprise, the mutant showed a slower mobility on
SDS-PAGE than wild type RrSP-D or the predominant forms ofnatural SP-D. Although some degree of overhydroxylation ofthe collagen domain cannot be excluded, our data indicate thatlower mobility of mutant monomers primarily results from anincrease in the apparent size and sialylation of the N-linkedsugar at Asn-70. We speculate that the mutant offers a lessrestricted access of Golgi enzymes to the glycosylation site,which is close to the hub of cross-linked dodecamers (12). In
FIG. 9. Ability of wild type RrSP-D or RrSP-Dser15/20 to pro-tect neutrophils against the deactivating effects of IAV onFMLP-stimulated respiratory burst responses. Neutrophils weretreated with control buffer, Bangkok79 IAV, or Bangkok79 IV that hadbeen preincubated with the indicated concentrations of wild typeRrSP-D or RrSP-Dser15/20. FMLP (5 3 1027 M) was then added to allsamples, and the resulting superoxide (O2
.) response measured over 10min. The amount of O2
. produced by neutrophils treated with IAVsamples was divided by that produced by neutrophils treated withcontrol buffer to obtain the percent of control O2
. response. Resultsshown are mean 6 S.E. of four experiments. Neutrophils treated withunopsonized IV had significantly depressed O2
. responses to FMLP asreported previously (4). Preincubation with wild type RrSP-D signifi-cantly protected neutrophils against IAV-induced depression of O2
. re-sponses to FMLP (as indicated by *), while preincubation with RrSP-Dser15/20 (even at higher concentrations) did not.
FIG. 7. Effect of wild type RrSP-D and RrSP-Dser15/20 on IAVbinding to neutrophils. A, neutrophils were allowed to adhere toplastic coverslips followed by incubation of the cells with either FITC-labeled Bangkok79 IAV alone (top panels), or the same amount of IAV,which had been preincubated with either 1.6 mg/ml RrSP-D (middlepanels) or RrSP-Dser15/20 (bottom panels). Phase contrast images areshown on the left and fluorescence images of the same fields on the right(magnification, 3100). In the absence of SP-D, faint fluorescence is seendistributed over the surface indicating bound virus; negative controlswere devoid of surface immunostaining. In the presence of RrSP-D,coarse aggregates of viral particles (e.g. white arrows) were observed inassociation with the cell surface. The identification of viral aggregatesin association with the cell surface was confirmed by electron micros-copy (40). In the presence of the mutant, there was a marked decreasein the binding of virus and no coarse viral aggregates were observed. B,FITC-labeled Bangkok79 IAV was preincubated with the indicatedconcentrations of either wild type RrSP-D (dark squares) or RrSP-Dser15/20 (open squares), followed by assessment of binding of thesesamples to human neutrophils by flow cytometry. Results shown aremean 6 S.E. of four experiments. Viral binding was significantly en-hanced by wild type RrSP-D (* indicates p # 0.05 compared to bindingof untreated IAV). In contrast, RrSP-Dser15/20 did not cause any en-hancement of viral binding. In fact, significantly reduced IAV bindingwas seen using samples preincubated with the highest concentration ofRrSP-Dser15/20 (indicated by **).
FIG. 8. Effect of wild type RrSP-D and RrSP-Dser15/20 on neu-trophil H2O2 responses to IAV. Samples of Bangkok79 IAV werepreincubated with the indicated concentrations of wild type RrSP-D(dark squares) or RrSP-Dser15/20 (open squares) as described in Fig. 7.Neutrophils were then added to these samples (final concentration 4 3106 cells/2 ml) and H2O2 production measured using the scopolectinassay as described under “Materials and Methods.” Results shown aremean 6 S.E. of three experiments. Preincubation with wild typeRrSP-D caused significant enhancement of neutrophil H2O2 production(* indicates p # 0.05 compared to response elicited by unopsonizedIAV), while preincubation with RrSP-Dser15/20 did not.
Surfactant Protein D 13729
by guest on April 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

this regard, other studies indicate that wild type dodecamersassemble in the rough endoplasmic reticulum, and that conver-sion of the N-linked sugar from a high mannose to maturesialylated form occurs shortly prior to secretion.3
The failure of RrSP-Dser15/20 to aggregate IAV providesstrong support for our hypothesis that the multimers of trimersare necessary for SP-D to agglutinate viral particles and othermicroorganisms. While RrSP-Dser15/20 bound to IAV virionsand showed significant hemagglutination inhibition activity,there was no detectable agglutinating activity. The protein wasalso deficient in other activities believed to be dependent on thestate of viral aggregation including: IAV binding to neutro-phils, augmentation of the neutrophil respiratory burst re-sponse to bound virus, and protection of neutrophils from viraldeactivation (4).Although distant conformational effects transmitted from
the mutated amino terminus through the helical arms to theCRD cannot be entirely excluded, the available data stronglysuggest that the trimeric CRD of SP-D is functionally univalentwith regard to IAV binding, i.e. that each trimeric CRD inter-acts with one or more hemagglutinin molecules on a single viralparticle. In this regard, crystallographic studies of trimericcarboxyl-terminal fragments of MBP suggest that the threesugar binding sites are located near the tip of the carboxyl-terminal globule (i.e. oriented at ;180° relative to the axis ofthe coiled-coil) and form a relatively planar binding surfacewhich could favor cooperative interactions with up to threeindependent glycoconjugates displayed on a microbial surface(17).We speculate that the enhanced anti-IAV activities of SP-D
(and conglutinin) relative to SP-A, MBP, and monoclonal IgGantibodies to the hemagglutinin (40), primarily result fromdifferences in the valency and spatial distribution of the he-magglutinin binding domains (i.e. CRD or Fab). The long armlength of SP-D may favor bridging interactions between viralparticles, leading to the formation of large viral aggregates,thereby modifying the interactions of IAV with the leukocytesurface. The failure of the mutant SP-D to promote viral ag-gregation suggests that single-arm forms of natural SP-D aresimilarly “deficient” in aggregation-dependent anti-viralactivity.An unexpected finding relates to the capacity of RrSP-
Dser15/20 to bind to the PR-8 strain of IAV in a calcium-independent manner (Table I). Previous studies have shownnegligible binding of rSP-D, RrSP-D, and hSP-D to PR-8, astrain that lacks N-linked oligosaccharides on the head of thehemagglutinin. We speculate that RrSP-Dser15/20 binding tothis strain reflects increased accessibility of the sialylated oli-gosaccharide at Asn-70 to sialic acid binding sites on the he-magglutinin or neuraminidase.In conclusion, we have expressed a mutated SP-D molecule
that is secreted as a trimer corresponding to a single “arm” ofthe SP-D dodecamer. Characterization of this mutant hasshown that the amino-terminal disulfide bonds of SP-D play animportant role in stabilizing the collagen triple helix and pro-tecting the molecule from degradation by several proteasesthat may be released at sites of pulmonary inflammation.These studies have also confirmed that important anti-viral
properties of SP-D are entirely dependent on the oligomeriza-tion of trimeric subunits.
Acknowledgments— We thank Dr. Robert Mecham, Department ofCell Biology, Washington University, for performing the microsequenc-ing. We also thank Janet North for excellent secretarial assistance.
REFERENCES
1. Crouch, E., Parghi, D., Kuan, S. F., and Persson, A. (1992) Am. J. Physiol. 263,L60–L66
2. Voorhout, W. F., Veenendaal, T., Kuroki, Y., Ogasawara, Y., Van Golde, L. M.,and Geuze, H. J. (1992) J. Histochem. Cytochem. 40, 1589–1597
3. Kuan, S. F., Rust, K., and Crouch, E. (1992) J. Clin. Invest. 90, 97–1064. Hartshorn, K. L., Crouch, E. C., White, M. R., Eggleton, P., Tauber, A. I.,
Chang, D., and Sastry, K. (1994) J. Clin. Invest. 94, 311–3195. O’Riordan, D. M., Standing, J. E., Kwon, K., Crouch, E. C., and Limper, A. H.
(1995) J. Clin. Invest. 95, 2699–27106. Crouch, E. C., Persson, A., Griffin, G. L., Chang, D., and Senior, R. M. (1995)
Am. J. Respir. Cell Mol. Biol. 12, 410–4157. Miyamura, K., Leigh, L. E., Lu, J., Hopkin, J., Lopez Bernal, A., and Reid, K.
B. (1994) Biochem. J. 300, 237–2428. Lu, J., Wiedemann, H., Holmskov, U., Thiel, S., Timpl, R., and Reid, K. B.
(1993) Eur. J. Biochem. 215, 793–7999. Crouch, E., Rust, K., Veile, R., Donis-Keller, H., and Grosso, L. (1993) J. Biol.
Chem. 268, 2976–298310. Shimizu, H., Fisher, J. H., Papst, P., Benson, B., Lau, K., Mason, R. J., and
Voelker, D. R. (1992) J. Biol. Chem. 267, 1853–185711. Rust, K., Grosso, L., Zhang, V., Chang, D., Persson, A., Longmore, W., Cai, G.
Z., and Crouch, E. (1991) Arch. Biochem. Biophys. 290, 116–12612. Crouch, E., Persson, A., Chang, D., and Heuser, J. (1994) J. Biol. Chem. 269,
17311–1731913. Crouch, E., Chang, D., Rust, K., Persson, A., and Heuser, J. (1994) J. Biol.
Chem. 269, 15808–1581314. Lim, B. L., Willis, A. C., Reid, K. B., Lu, J., Laursen, S. B., Jensenius, J. C., and
Holmskov, U. (1994) J. Biol. Chem. 269, 11820–1182415. Reid, K. B. (1993) Biochem. Soc. Trans. 21, 464–46816. Holmskov, U., Laursen, S. B., Malhotra, R., Wiedemann, H., Timpl, R., Stuart,
G. R., Tornoe, I., Madsen, P. S., Reid, K. B., and Jensenius, J. C. (1995)Biochem. J. 305, 889–896
17. Weis, W. I., and Drickamer, K. (1994) Structure (Lond.) 2, 1227–124018. Horton, R. M., Ho, S. N., Pullen, J. K., Hunt, H. D., Cai, Z., and Pease, L. R.
(1993) Methods Enzymol. 217, 270–27919. Ausubel, R. M., Brent, R., Kingston, R. E., Moore, S. S., Seidman, J. G., Smith,
J. A., and Struhl, K. (eds) (1992) Current Protocols in Molecular Biology, pp.14.1–16.14.13, John Wiley & Sons, Inc., New York
20. Cockett, M. I., Bebbington, C. R., and Yarranton, G. T. (1990) Bio/Technology8, 662–668
21. Persson, A., Rust, K., Chang, D., Moxley, M., Longmore, W., and Crouch, E.(1988) Biochemistry 27, 8576–8584
22. Persson, A., Chang, D., Rust, K., Moxley, M., Longmore, W., and Crouch, E.(1989) Biochemistry 28, 6361–6367
23. Wenstrup, R. J., Shrago-Howe, A. W., Lever, L. W., Phillips, C. L., Byers, P. H.,and Cohn, D. H. (1991) J. Biol. Chem. 266, 2590–2594
24. Saarinen, J., Kalkkinen, N., Welgus, H. G., and Kavanen, P. T. (1994) J. Biol.Chem. 269, 18134–18140
25. Shapiro, S. D., Kobayashi, D. K., and Ley, T. J. (1993) J. Biol. Chem. 268,23824–23829
26. Shapiro, S. D., Fliszar, T., Broeckelman, R. P., Mecham, R. P., Senior, R. M.,and Welgus, H. G. (1995) J. Biol. Chem. 270, 6351–6356
27. Sires, U. I., Murphy, G., Welgus, H. G., and Senior, R. M. (1994) Biochem.Biophys. Res. Commun. 204, 613–620
28. Hartshorn, K. L., Collamer, M., Auerbach, M., Myers, J. B., Pavlotsky, N., andTauber, A. I. (1988) J. Immunology 141, 1295–1301
29. Anders, E. M., Hartley, C. A., and Jackson, D. C. (1990) Proc. Natl. Acad. Sci.U. S. A. 87, 4485–4489
30. Hartshorn, K. L., Collamer, M., White, M. R., Schwartz, J. H., and Tauber, A.I. (1990) Blood 75, 218–226
31. Hartshorn, K. L., Sastry, K., Brown, D., White, M. R., Okarma, T. B., Lee, Y.M., and Tauber, A. I. (1993) J. Immunol. 151, 6265–6273
32. Hartshorn, K. L., Sastry, K., White, M. R., Anders, E. M., Super, M.,Ezekowitz, R. A., and Tauber, A. I. (1993) J. Clin. Invest. 91, 1414–1420
33. Lu, J., Willis, A. C., and Reid, K. B. (1992) Biochem. J. 284, 795–80234. Olsen, B. R. (1991) in Cell Biology of Extracellular Matrix (Hay, E. D., ed) pp.
177–220, Plenum Publishing Corp., New York35. Engel, J., and Prockop, D. J. (1991) Annu. Rev. Biophys. Biophys. Chem. 20,
137–15236. Hoppe, H. J., Barlow, P. N., and Reid, K. B. (1994) FEBS Lett. 344, 191–19537. Spissinger, T., Schafer, K. P., and Voss, T. (1991) Eur. J. Biochem. 199, 65–7138. Weis, W. I., Kahn, R., Fourme, R., Drickamer, K., and Hendrickson, W. A.
(1991) Science 254, 1608–161539. Ogasawara, Y., and Voelker, D. R. (1995) J. Biol. Chem. 270, 19052–1905840. Hartshorn, K., Reid, K., White, M., Jensenius, J., Tauber, A., and Crouch, E.
(1996) Blood, 87, 3450–3461
3 P. Brown-Augsburger, D. Chang, K. Rust, and E. C. Crouch, sub-mitted for publication.
Surfactant Protein D13730
by guest on April 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Fliszar, Howard G. Welgus and Edmond C. CrouchPatricia Brown-Augsburger, Kevan Hartshorn, Donald Chang, Kevin Rust, Catherine
PROPERTIESEXPRESSION OF A TRIMERIC PROTEIN WITH ALTERED ANTI-VIRAL
Site-directed Mutagenesis of Cys-15 and Cys-20 of Pulmonary Surfactant Protein D:
doi: 10.1074/jbc.271.23.137241996, 271:13724-13730.J. Biol. Chem.
http://www.jbc.org/content/271/23/13724Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/271/23/13724.full.html#ref-list-1
This article cites 38 references, 20 of which can be accessed free at
by guest on April 5, 2018
http://ww
w.jbc.org/
Dow
nloaded from






![Modèles structuraux et fonctionnels du site actif des … · 2014. 10. 4. · Ni Cys-S S S Cys-S Fe CN CN CO Cys Cys Site actif des H2ases [NiFe] Dérivés du [Ni(xbsms)]: 1. Synthèse](https://img.dokumen.tips/doc/110x75/5fc526ed9695db7c55538df1/modles-structuraux-et-fonctionnels-du-site-actif-des-2014-10-4-ni-cys-s-s.jpg)






















