Embed Size (px)
Citation preview

1
Running head: Arsenic speciation in phloem and xylem exudates 1
Corresponding author: Fang-Jie Zhao 2
Address: Rothamsted Research, Harpenden, Hertfordshire AL5 2JQ, UK 3
Phone: 44 1582 763133 4
Email: [email protected] 5
6
Research category: Environmental Stress and Adaptation 7
8
Plant Physiology Preview. Published on September 24, 2010, as DOI:10.1104/pp.110.163261
Copyright 2010 by the American Society of Plant Biologists
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

2
Arsenic speciation in phloem and xylem exudates of Ricinus communis 9
10
Wen-Ling Ye, B. Alan Wood, Jacqueline L. Stroud, P. John Andralojc, Andrea Raab, 11
Steve P. McGrath, Jörg Feldmann, Fang-Jie Zhao* 12
13
1. School of Earth and Space Sciences, University of Science and Technology of China, Hefei, 14
Anhui 230026, China (W.-L.Y.) 15
2. Rothamsted Research, Harpenden, Hertfordshire AL5 2JQ, United Kingdom (W.-L.Y., J. 16
L.S., P.J.A., S.P.M., F.-J.Z.) 17
3. College of Physical Science, Trace Element Speciation Laboratory (TESLA) Chemistry, 18
University of Aberdeen, Meston Building, Aberdeen AB24 3UE, United Kingdom (B. A.W., 19
A.R., J.F.) 20
21
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

3
Footnotes: 22
Rothamsted research is an institute of the Biotechnology and Biological Sciences Research 23
Council of the United Kingdom. 24
* Corresponding author: Fang-Jie Zhao (Email: [email protected]) 25
26
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

4
Abstract 27
How arsenic (As) is transported in phloem remains unknown. To help answer this question, 28
we quantified the chemical species of As in phloem and xylem exudates of Ricinus communis 29
exposed to arsenate [As(V)], arsenite [As(III)], monomethylarsonic acid [MMA(V)] or 30
dimethylarsinic acid [DMA(V)]. In the As(V)- and As(III)-exposed plants, As(V) was the 31
main species in xylem exudate (55 - 83%) whereas As(III) predominated in phloem exudate 32
(70 – 94%). The ratio of As concentrations in phloem to xylem exudate varied from 0.7 to 3.9. 33
Analyses of phloem exudate using high-resolution ICP-MS and accurate mass ES-MS coupled 34
to HPLC identified high concentrations of reduced and oxidized glutathione and some 35
oxidized phytochelatin (PC2), but no As(III)-thiol complexes. It is thought that As(III)-thiol 36
complexes would not be stable in the alkaline conditions of phloem sap. Small concentrations 37
of oxidized glutathione and oxidized PC2 were found in xylem exudate, where there was also 38
no evidence of As(III)-thiol complexes. MMA(V) was partially reduced to MMA(III) in roots, 39
but only MMA(V) was found in xylem and phloem exudate. Despite the smallest uptake 40
among the four As species supplied to plants, DMA(V) was most efficiently transported in 41
both xylem and phloem, and its phloem concentration was 3.2 times that in xylem. Our results 42
show that free inorganic As, mainly As(III), was transported in the phloem of R. communis 43
exposed to either As(V) or As(III), and that methylated As species were more mobile than 44
inorganic As in the phloem. 45
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

5
INTRODUCTION 46
47
Arsenic (As) is an environmental and food chain contaminant that has attracted much 48
attention in recent years. Soil contamination with As may lead to phytotoxicity and reduced 49
crop yield (Panaullah et al., 2009). Food crops are also an important source of inorganic As, a 50
class-one carcinogen, in humans dietary intake, and there is a need to decrease the exposure to 51
this toxin (European Food Safety Authority, 2009). Paddy rice (Oryza sativa) is particularly 52
efficient in As accumulation, which poses a potential risk to the population based on a rice 53
diet (Meharg et al., 2009; Zhao et al., 2010a). Other terrestrial food crops generally do not 54
accumulate as much As as paddy rice; however, where soils are contaminated, relatively high 55
concentrations of As in wheat (Triticum aestivum) grain have been reported (Williams et al., 56
2007; Zhao et al., 2010b). On the other hand, some fern species in the Pteridaceae family are 57
able to tolerate and hyperaccumulate As in the above-ground part to >1000 mg kg-1 dry weight 58
(e.g. Ma et al., 2001; Zhao et al., 2002); these plants offer the possibility for remediation of 59
As-contaminated soil or water (Salido et al., 2003; Huang et al., 2004). A better understanding 60
of As uptake and long-distance transport, metabolism and detoxification is needed for 61
developing strategies for mitigating As contamination, through either decreased As 62
accumulation in food crops or enhanced As accumulation for phytoremediation. 63
The pathways of As uptake by plant roots differ between different As species; arsenate 64
[As(V)] enters plant cells via phosphate transporters, whereas arsenite [As(III)] is taken up via 65
some aquaporins (reviewed by Zhao et al., 2009). In rice, a silicic acid efflux protein also 66
mediates As(III) efflux towards stele for xylem loading (Ma et al., 2008). Methylated As 67
species, such as monomethylarsonic acid (MMA(V)) and dimethylarsinic acid (DMA(V)), 68
which may be present in the environment as products of microbial or algal methylation of 69
inorganic As or from past uses of methylated As pesticides, are taken up by rice roots partly 70
through the aquaporin NIP2;1 (also named Lsi1) (Li et al., 2009). Once inside plant cells, 71
As(V) is reduced to As(III), possibly catalyzed by arsenate reductase(s) such as the plant 72
homologues of the yeast ACR2 (Bleeker et al., 2006; Dhankher et al., 2006; Ellis et al., 2006; 73
Duan et al., 2007). As(III) has a high affinity to thiol (-SH) groups and is detoxified by 74
complexation with thiol-rich phytochelatins (PCs) (Pickering et al., 2000; Schmöger et al., 75
2000; Raab et al., 2005; Bluemlein et al., 2009; Liu et al., 2010). As(III)-PC complexation in 76
roots was found to result in reduced mobility for efflux and for long-distance transport, 77
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

6
possibly because the complexes are stored in the vacuoles (Liu et al., 2010). Excess As(III) 78
causes cellular toxicity by binding to the vicinal thiol groups of enzymes, such as the plastidial 79
lipoamide dehydrogenase, which has been shown to be a sensitive target of As toxicity (Chen 80
et al., 2010). The As hyperaccumulating Pteris species differ from non-hyperaccumulating 81
plants because of enhanced arsenate uptake (Wang et al., 2002; Poynton et al., 2004), little 82
As(III)-thiol complexation (Zhao et al., 2003; Raab et al., 2004) and efficient xylem loading of 83
As(III) (Su et al., 2008). Recently, an As(III) efflux transporter, PvACR3, has been found to 84
play an important role in As(III) detoxification by transporting As(III) into vacuoles in Pteris 85
vittata (Indriolo et al., 2010). 86
With the exception of As hyperaccumulators, most plant species have a limited root-87
to-shoot translocation of As (Zhao et al., 2009). The chemical species of As in xylem exudate 88
have been determined in a number of plant species. As(III) was found to be the predominant 89
species (80 – 100%) in the xylem sap of rice, tomato (Lycopersicon esculentum), cucumber 90
(Cucumis sativus) and P. vittata even when these plants were fed As(V) (Mihucz et al., 2005; 91
Xu et al., 2007; Ma et al., 2008; Su et al., 2010), suggesting that As(V) is reduced in roots 92
before being loaded into the xylem. In other plant species, such as Brassica juncea 93
(Pickering et al., 2000), wheat (Triticum aestivum) and barley (Hordeum vulgare) (Su et al., 94
2010), As(V) accounted for larger proportions (40 – 50%) of the total As in the xylem sap. 95
Studies using HPLC-ICP-MS coupled with electrospray mass spectrometry (ES-MS) showed 96
no evidence of As(III)-thiol complexation in the xylem sap of sunflower (Helianthus annuus) 97
(Raab et al., 2005). When rice plants were exposed to MMA(V) or DMA(V), both As species 98
were found in the xylem sap (Li et al., 2009). Generally, methylated As species are taken up 99
by roots at slower rates than inorganic As, but they are more mobile during the xylem 100
transport from roots to shoots (Marin et al., 1992; Raab et al., 2007; Li et al., 2009). 101
It has been shown that phloem transport contributes substantially to As accumulation 102
in rice grain (Carey et al., 2010). However, little is known about how As is transported in 103
phloem (Zhao et al., 2009). There are no reports on the chemical species of As in phloem 104
exudate. The speciation of As in phloem is important because it dictates how As is loaded in 105
the source tissues and unloaded in the sink tissues, such as grain. Questions with regard to the 106
oxidation state, methylation and complexation of As in phloem sap remain to be answered. 107
Unlike xylem sap, phloem sap is much more difficult to obtain in sufficient quantities for 108
analysis. In the present study, we investigated As speciation in phloem and xylem exudates of 109
Ricinus communis (castor bean), which is widely used as a model plant to investigate phloem 110
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

7
transport of solutes (e.g. Hall et al., 1971; Hall and Baker, 1972; Allen and Smith, 1986; 111
Bromilow et al., 1987). 112
113
RESULTS 114
115
Sugar composition in phloem and xylem exudates of R. communis 116
The purity of phloem and xylem exudates was checked by analyzing the sugar 117
composition. Phloem exudate contained high concentrations of sucrose with the values similar 118
to those reported before (Hall and Baker, 1972; Smith and Milburn, 1980), whilst reducing 119
sugars were absent (fructose) or present at only trace level (glucose) (Table 1). Xylem exudate 120
contained very small concentrations of sucrose, fructose and glucose. These compositions are 121
characteristic of phloem and xylem saps, suggesting little cross contamination. 122
123
Arsenic speciation in R. communis exposed to As(V) or As(III) 124
In the first experiment, 18-d old plants were exposed to 10 μM As(III) or As(V) for 24 125
– 48 h. Total As concentration in phloem exudate increased slightly from 24 to 48 h after As 126
exposure, from approximately 0.8 to 0.9 – 1.2 μM (Fig. 1a). The concentrations were not 127
significantly different between the As(V) and As(III) treatments at either sampling times. The 128
majority of As in phloem exudates was As(III), which accounted for 67 – 79% and 86 – 90% 129
of the total As at 24 and 48 h, respectively, and the remainder was As(V). Again, there was no 130
significant difference in this percentage between the As(V) and As(III) treatments. Xylem 131
exudate was collected after 48 h exposure. Total As concentration in the xylem exudate was 132
about half of that in the phloem exudate (Fig. 1b), with As(III) accounting for 44% in both 133
treatments and the rest being As(V). 134
Analysis of the plant samples collected after 48 h exposure showed much higher As 135
concentrations in roots than in leaves (Fig. 1c, d). 79% of the As in roots was As(III), 136
compared with 96% in leaves, and there was no significant difference in this percentage 137
between the As(III) and As(V) treatments. The sum of all As species detected in roots was 138
smaller in the As(V) than in the As(III) treatment (p = 0.056), but the opposite was observed 139
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

8
for the concentrations in leaves (p < 0.01). Total As concentrations were also determined by 140
acid digestion followed by ICP-MS determination. The sum of As species determined by 141
extraction with a phosphate buffer solution and HPLC-ICP-MS analysis represented 81% and 142
43% of the total As in roots and shoots, respectively. 143
A considerable proportion (43% at 24 h) of As(III) in the nutrient solution was 144
oxidized to As(V), whereas 9% of the As in the As(V) treatment was reduced to As(III) (data 145
not shown). It has been shown that plant roots extrude As(III) to the external medium 146
following As(V) uptake and reduction (Xu et al. 2007). Conversely, As(III) in the medium 147
may be oxidized to As(V) because the nutrient solution was continuously aerated. The partial 148
inter-conversion of As species in the medium may partly explain the lack of significant 149
differences between the As(V) and As(III) treatments. 150
151
Arsenic speciation in R. communis exposed to increasing concentration of As(V) 152
Total As concentration in phloem exudate collected 24 h after As(V) exposure 153
increased with the increasing As(V) concentration in the nutrient solution (p < 0.001; Fig. 2a). 154
The proportion of As(III) varied little (84 – 90%) in the 2 – 50 μM As(V) treatment, but 155
decreased to 71% in the 100 μM As(V) treatment. Xylem exudate could only be collected 156
from the low As(V) treatments (2 – 10 μM) and from one of the four replicates of the 50 μM 157
As(V)-treated plants; As toxicity at high As(V) concentrations might have affected water 158
uptake and/or xylem exudation. As(V) was the main As species in the xylem exudate, 159
accounting for 67% and 79% of the total As in the 2 and 10 μM treatments, respectively, with 160
the rest being As(III) (Fig. 2b). The ratio of phloem to xylem As concentrations was 3.9 and 161
0.7 in the 2 and 10 μM treatments, respectively. No As species were detected in either xylem 162
or phloem exudate from the control plants in the 0 μM As(V) treatment. 163
Arsenic concentrations in roots and leaves increased with the external As(V) 164
concentrations (Fig. 2c, d). Most of the As in leaves (96%) was As(III) and the proportion was 165
not significantly affected by the concentration of As(V) in the medium. In contrast, the 166
proportion of As(III) in roots decreased progressively with the concentration of As(V) 167
exposure, from 81% at 2 μM to 36% at 100 μM As(V). In fact, whilst the concentration of 168
As(V) in roots increased with the As(V) exposure concentration, that of As(III) did not 169
increase beyond the 10 μM As(V) treatment. In this experiment, the extraction and As 170
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

9
speciation method recovered on average 84% and 59% of the total As in roots and leaves, 171
respectively. 172
173
Phosphate supply affects As concentration but not speciation in phloem and xylem 174
exudates of R. communis 175
In this experiment, R. communis plants were exposed to 10 μM As(V) with either 10 or 176
200 μM phosphate. Phloem exudate was dominated by As(III), which accounted for 89 - 94% 177
of the total As at 24 h and 48 h, and the level of phosphate supply had no significant effect on 178
this percentage (p > 0.80) (Fig. 3a). Increasing phosphate from 10 to 200 μM did significantly 179
decrease the concentration of As(III) in phloem exudate, by 50% (p < 0.001) and 26% (p < 180
0.05) at 24 and 48 h, respectively. In contrast, xylem exudate was dominated by As(V), with 181
As(III) accounting for only 17 – 23% of the total As (Fig. 3b). The concentrations of As(V) 182
and As(III) in xylem exudate were decreased by 40% (p < 0.05) and 60% (p < 0.01), 183
respectively, by increasing phosphate concentration in the medium. The percentage of As(III) 184
was not significantly affected. At 48 h, total As concentrations in phloem exudate were 15 – 185
60% higher than those in xylem exudate. 186
187
No evidence of As(III)-thiol complexation in the phloem and xylem exudates of R. 188
communis 189
Phloem and xylem exudates collected after 48 h exposure to 10 μM As(V) were 190
analyzed for As-thiol complexes and free thiol compounds using reverse phase HPLC-high 191
resolution ICP-MS / high resolution ES-MS. The use of high resolution ICP-MS allows 192
quantification of S (m/z 32), which is not possible with normal ICP-MS due to the polyatomic 193
interference of O2 (m/z 32), in addition to simultaneous quantification of As (m/z 75). In 194
addition, the use of high resolution ES-MS allows molecular identification of the compounds. 195
In both phloem and xylem exudate samples, As was eluted in the void volume corresponding 196
to inorganic As; also eluted was sulfate. No As-thiol complexes were detected in either 197
exudate (Fig. 4a, b). High concentrations of reduced and oxidized glutathione (GSH and 198
GSSG) were found in the phloem exudate, as well as some oxidized phytochelatin (PC2) 199
(Table 2). GSSG and oxidized PC2 were detected in the xylem exudate, but at much lower 200
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

10
concentrations than those in the phloem exudate. No GSH was present in the xylem exudate. 201
Inorganic S (sulfate) was abundant in both exudates. 202
203
Methylated As has a greater mobility in phloem and xylem than inorganic As 204
In this experiment, As speciation and the relative mobility were compared among 205
plants exposed to 10 μM As(V), MMA(V) or DMA(V). Only inorganic As species were found 206
in the phloem exudate from the As(V)-treated plants, with As(III) being the predominant 207
species (88% and 92% at 24 h and 48 h, respectively) (Fig. 5a). In the MMA(V)-treated plants, 208
MMA(V) was the only species detected in the phloem exudate, whilst only DMA(V) was 209
found in the DMA(V)-treated plants. Arsenic concentrations in phloem exudate were similar 210
in both 24 h and 48 h samples. Among the three treatments, As concentration in phloem 211
exudate was the highest in the DMA(V) treatment (p < 0.001; Fig. 5a). 212
In xylem exudate, total As concentration followed the order of treatment of MMA(V) 213
< As(V) < DMA(V) (Fig. 5b). The xylem exudate from the As(V)-treated plants contained 214
38% and 62% As(III) and As(V), respectively. The xylem exudate from the MMA(V)-treated 215
plants contained 67% MMA(V), 15% As(III) and 18% As(V). However, the MMA(V) stock 216
solution used in the experiment contained a small amount (~3%) of As(V) as an impurity, 217
which likely explains the occurrence of As(V) and As(III) in the xylem exudate from the 218
MMA(V)-treated plants. Only DMA(V) was found in the xylem exudate of the DMA(V)-219
treated plants. The ratios of As concentration in phloem exudate to that in xylem exudate were 220
2.3, 7.3 and 3.2 in the As(V), MMA(V) and DMA(V) treatments, respectively. 221
The concentrations and species of As in roots and leaves differed between the three 222
treatments (Fig. 5c, d). Similar to the experiments described above, As(III) was the dominant 223
species in roots (87%) and leaves (91%) of the As(V)-treated plants, with As(V) being the 224
minor As species. Arsenic speciation in the roots of the MMA(V)-treated plants was more 225
complex, showing the presence of MMA(V), MMA(III), As(III) and As(V) with percentages 226
of 60, 14, 23 and 3%, respectively (Supplemental Fig. 1). In the leaves of this treatment, As 227
species comprised 89% and 11% MMA(V) and As(III), respectively. In the plants exposed to 228
DMA(V), only DMA(V) was found in both roots and leaves. The MMA(V)-treated plants had 229
smaller (by about 75%) concentrations of total As (sum of all As species) in roots and leaves 230
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

11
than the As(V)-treated plants. In the DMA(V)-treated plants, total As concentrations in roots 231
and leaves were 2% and 61%, respectively, of those in the As(V)-treated plants. 232
In this experiment, the sum of As species accounted for 40 – 60% of the total As 233
concentration by acid digestion, with the exception of a complete extraction (105%) in the 234
DMA(V)-treated shoots. Based on the total As concentrations by acid digestion, uptake of As 235
during the 48 h exposure were 358, 80 and 8.5 nmol g-1 root fresh weight in the As(V), 236
MMA(V) and DMA(V) treatments, respectively. 237
238
DISCUSSION 239
240
The present study was motivated by the gap of knowledge of As speciation in phloem 241
exudate, which has important implications for how As is redistributed around the plant, 242
especially towards the seeds (Zhao et al., 2009). R. communis plants were exposed to four 243
different As species, and As speciation in phloem exudate was compared to that in xylem 244
exudate and roots and leaves. 245
In the plants exposed to either As(V) or As(III), As(III) was found to be the 246
predominant As species in phloem exudate and As(V) the minor species. The proportion of 247
As(III) varied somewhat among different experiments and was also influenced by the 248
concentrations of As(V) in the external solution, but generally fell within the range of 80 – 249
90% (Figs 1, 2, 3, 5). The proportion dropped to around 70% when plants were exposed to a 250
high concentration (100 μM) of As(V). Furthermore, inorganic As in the phloem exudate was 251
not complexed with thiol compounds, even though the concentrations of GSH, GSSG and PC2 252
far exceeded that of inorganic As (Fig. 4 and Table 2). The lack of thiolated arsenic is not an 253
analytical artefact, because the extraction and the chromatographic separation has been 254
validated by using XANES/EXAFS of plant tissues directly (Bluemlein et al., 2008). This lack 255
of thiol complexation may be explained by the high pH of phloem sap in R. communis (7.4 – 256
8.2) (Hall and Baker, 1972; Allen and Smith, 1986). It is known that As(III) forms stable 257
complexes with thiol compounds under acidic conditions but not in neutral-alkaline 258
environments (Schmöger et al., 2000). A number of As(III)-thiol complexes have been 259
identified in roots and shoots of different plant species (Raab et al., 2004; Raab et al., 2005; 260
Liu et al., 2010), whilst X-ray absorption studies indicate that most of As(III) in non-261
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

12
hyperaccumulating plant species is bonded with S (Pickering et al., 2000; Dhankher et al., 262
2002). As(III)-thiol complexes are likely to be stored in vacuoles where the acidic pH favors 263
the stability of the complexes. Increasing As(III)-thiol complexation was found to decrease 264
xylem transport of As(III) (Liu et al., 2010), and may also affect As(III) transport in phloem in 265
the same way. Given that uncomplexed As(III) or at least only weakly complexed As(III), 266
which is not detectable by chromatographic methods (Feldmann et al., 2009), and, to a smaller 267
extent, As(V), are the only species of As in phloem exudate, transporters for these species are 268
likely to be involved in the loading and unloading during phloem transport, e.g. some of the 269
aquaporins, Lsi2-like or ACR3-like proteins for As(III) and phosphate transporters for As(V). 270
Similar to our study, thiol compounds (mainly glutathione and PC2) were found to be 271
abundant in the phloem sap from Brassica napus exposed to Cd (Mendoza-Cózatl et al., 2008). 272
Experiments with grafting and shoot-specific expression of the genes involved in glutathione 273
synthesis have shown that glutathione and PCs are transported via phloem from shoots to 274
roots of Arabidopsis thaliana (Chen et al., 2006; Li et al., 2006). Unlike As, it is possible that 275
Cd-PC complexes are transported in phloem (Mendoza-Cózatl et al., 2008), because these 276
complexes are stable in the slightly alkaline conditions of phloem sap, but not in acidic 277
environments (Johanning and Strasdeit, 1998). 278
Different from phloem exudate, more As(V) than As(III) was found in xylem exudate 279
of R. communis, with the former typically accounting for 60 - 80% of the total As (Figs. 1, 2, 3 280
and 5). There was also no evidence of As(III)-thiol complexes (Fig. 4), even though xylem sap 281
pH (~5.0) in R. communis (Allen and Smith, 1986) is favorable to the stability of As(III)-thiol 282
complexes. Small concentrations of GSSG and oxidized PC2 were found in xylem sap of R. 283
communis (Table 2), which was consistent with a previous study on sunflower (Raab et al., 284
2005). The study of Raab et al. (2005) also found no evidence of As(III)-thiol complexation in 285
the xylem sap of sunflower. The presence of only low concentrations of non-reactive 286
(oxidized) thiols in xylem sap probably explains the lack of As(III)-thiol complexation. 287
Similarly, Mendoza-Cózatl et al. (2008) found small concentrations of glutathione and traces 288
of PC2 in the xylem sap of Brassica napus exposed to Cd, although the analytical method used 289
did not separate oxidized from reduced thiols. 290
Previous studies have shown that As(III) dominates in the xylem sap collected from a 291
number of plant species exposed to As(V), including tomato, rice, cucumber and the As 292
hyperaccumulator Pteris vittata (Mihucz et al., 2005; Xu et al., 2007; Su et al., 2008; Su et al., 293
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

13
2010). In contrast, As(V) was found to be an important As species in the xylem sap collected 294
from Brassica juncea, Holcus lanatus and wheat, accounting for between 30 – 60% of the 295
total As (Pickering et al., 2000; Su et al., 2010). The relative percentages of As(III) and As(V) 296
in xylem sap may depend on a number of factors, such as the capacity of arsenate reduction in 297
roots, the abundance and the relative affinities of As(III) or As(V) transporters involved in 298
xylem loading, the degree of As(III)-thiol complexation in roots and the presence of 299
competing ions. There is evidence that more As(V) was loaded into xylem when the arsenate 300
reduction capacity in roots was exceeded in the treatments with high As(V) concentrations 301
(Fig. 2); this also had the effect of increasing As(V) percentage in the phloem exudate. 302
Complexation of As(III) with thiol compounds in roots may decrease As(III) loading into the 303
xylem (Liu et al. 2010), resulting in a larger As(V)% in the xylem exudate than that in the root 304
tissues. This explanation may also apply to the higher As(V)% in phloem sap than in leaves. 305
Phosphate is known to inhibit As(V) uptake (Ullrich-Eberius et al., 1989; Abedin et al., 2002; 306
Wang et al., 2002), and may also inhibit As(V) loading into xylem or phloem. However, this 307
latter effect was not observed in our experiment (Fig. 3). Increasing phosphate supply 308
decreased As concentrations in both xylem and phloem without significantly affecting the 309
percentages of the two As species, suggesting that phosphate inhibits the influx of arsenate 310
into roots rather than its loading into xylem. Comparing xylem and phloem exudates, the 311
concentrations of As in the latter were, in most cases, higher than those in the former. The fact 312
that both leaves and phloem exudate contained much higher proportions of As(III) than xylem 313
exudate indicates that As(V) is reduced to As(III) in leaves as well as in roots. 314
Previous studies have shown that methylated As species are generally taken up more 315
slowly than inorganic As, but translocated more efficiently from roots to shoots (Marin et al., 316
1992; Abedin et al., 2002; Raab et al., 2007; Li et al., 2009). Similar results were obtained 317
with R. communis, with As uptake following the order of As(V) > MMA(V) > DMA(V). In R. 318
communis roots, MMA(V) was partly reduced to MMA(III), although only MMA(V) was 319
loaded into the xylem and phloem (Fig. 5, Supplemental Fig. 1). According to the Challenger 320
pathway of As methylation established in microorganisms (Bentley and Chasteen, 2002), 321
reduction of MMA(V) to MMA(III) is a prerequisite step before further addition of the methyl 322
group. However, no DMA(V) was produced in the MMA(V)-treated plants, indicating no 323
further methylation of MMA(III) in R. communis within the 48 h exposure period. Despite a 324
very limited uptake of DMA(V), its concentration was the highest in both xylem and phloem 325
exudates among the three As treatments, with the phloem concentration being about 3 times 326
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

14
the xylem concentration (Fig. 5). MMA(V) concentrations in phloem exudate were also 327
comparable to or larger than inorganic As despite smaller uptake of the former species. These 328
results indicate that methylated As, particularly the dimethyl species, are more mobile than 329
inorganic As in the phloem. Consistent with this finding, Carey et al. (2010) showed that 330
DMA(V) was transported to rice grain much more efficiently than inorganic As. Uptake of 331
undissociated MMA(V) and DMA(V) by rice roots is partly mediated by the aquaporin 332
NIP2,1 (Li et al., 2009), but membrane transporters responsible for loading and unloading of 333
methylated As species in phloem remain unknown. It has been suggested that a lack of thiol 334
complexation of MMA(V) and DMA(V) may explain their high mobility in xylem transport 335
(Raab et al., 2007); this explanation may also apply to the high mobility in phloem observed in 336
the present study. Increased lipophilicity with methylation may be another reason, although 337
this is apparently associated with decreased uptake by roots. 338
The identification of the forms of As in phloem exudate should help future research to 339
elucidate the mechanisms of As loading and unloading in the phloem. Future work should also 340
investigate if the forms of As transported in phloem are similar in food crops such as rice. 341
342
MATERIALS AND METHODS 343
344
Plant culture 345
Seeds of R .communis var. Gibsonii were surface sterilized by immersing them in 0.5% 346
NaOCl for 15 min. After rinsing, seeds were soaked in deionized water overnight and then 347
germinated in moist vermiculite. After germination, seedlings were grown in hydroponic 348
culture for 18 d with a modified ½ strength Hoagland nutrient solution (1 seedling per 1-L 349
pot). The composition of the nutrient solution were 3 mM KNO3, 2 mM Ca(NO3)2, 0.5 mM 350
NH4H2PO4, 1 mM MgSO4, 2 mM MES (2-morpholinoethanesulfonic acid; pH adjusted to 6.0 351
with KOH), 100 µM Fe(III)-EDTA, 0.55 µM MnCl2, 46 µM H3BO3,9 µM MnCl2,0.75 µM 352
ZnSO4 and 0.35 µM CuSO4. The nutrient solution was aerated continuously and renewed once 353
every 3 d. The growth conditions were 16 h photoperiod with a light intensity of 350 µmol m-354
2s-1, day : night temperatures 28:25ºC and 70% relative humidity. After 18-d growth, the fresh 355
weight of shoots and roots were on average 30 and 18 g per plant. 356
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

15
357
Arsenic exposure and collection of phloem and xylem exudates 358
A series of experiments were conducted with 18-d old R. communis plants, varying in 359
the species and concentrations of As added to the nutrient solution. The first experiment 360
compared As exposure in the forms of As(V) (Na2HAsO4) or As(III) (NaAsO2) at 10 μM. The 361
second experiment investigated the dose response to increasing As(V) concentrations (0, 2, 10, 362
50 and 100 μM). In the third experiment, plants were exposed to 10 μM As(V) with either 10 363
or 200 μM phosphate. In the fourth experiment, xylem and phloem exudates were collected 364
from plants exposed to 10 μM As(V) for the analysis of thiol compounds and As(III)-thiol 365
complexes. The fifth experiment compared the As mobility and speciation in xylem and 366
phloem exudates after R. communis plants were exposed to 10 μM As(V), MMA(V) or 367
DMA(V). Each treatment was replicated in 3 – 4 pots. During As exposure, the phosphate 368
concentration in the nutrient solution was reduced to 10 μM to allow more uptake of As(V) 369
(except the high P treatment in the third experiment). Phloem exudate was collected at 24 and 370
48 h after As exposure, and xylem exudate at 48 h exposure, except in the second experiment 371
when both exudates were collected at 24 h. To collect phloem exudate, a shallow, V-shaped 372
incision was made into the bark of the stem at about 2 cm above the stem/root junction with a 373
razor blade (Hall et al., 1971). Phloem exudate was collected in a glass capillary tube (50 or 374
100 µl) for 1 h. After collection of phloem exudate, the bark (containing the phloem tissue) 375
was removed from a 30-40 mm section of stem near the base by ring-girdling (Allen and 376
Smith, 1986). One h later, the stem was cut at the top of the ring-girdled section and xylem 377
exudate collected by pipette for 1 h after decapitation. The collected sap samples were stored 378
on ice, and diluted with a phosphate buffer solution (PBS) containing 2 mM NaH2PO4 and 0.2 379
mM Na2-EDTA (pH 6.0) before As speciation analysis. For analysis of thiols and As(III)-thiol 380
complexes, phloem and xylem saps were diluted with 1% formic acid in 1:5 ratio. After 381
collection of phloem and xylem exudates, plant roots were rinsed briefly in an ice-cold 382
desorption solution containing 1 mM K2HPO4, 0.5 mM Ca(NO3)2 and 5 mM MES (pH 6.0), 383
and immersed in 1 L of the same solution for 10 min to remove apoplastic As (Xu et al., 2007). 384
Shoots were rinsed in deionized water. Root and leaf samples were blotted dry, frozen in 385
liquid N2 and were ground to fine powder with a mortar and pestle. Aliquots of nutrient 386
solutions were taken at 24 and 48 h to monitor the changes in As species. 387
388
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

16
Analysis of sugar composition in xylem and phloem exudates 389
The predominant soluble sugars in both xylem and phloem exudates were identified 390
and quantified by high-pH anion exchange HPLC using a Dionex CarboPac PA1 guard (4 x 391
50 mm) and analytical (4 x 250 mm) column connected to a Dionex DX500 (Sunnyvale, CA, 392
USA), with refrigerated autosampler. Each separation was achieved within 20 min by isocratic 393
elution using 40 mM sodium hydroxide at a flow rate of 1 mL min-1. Peak detection was by 394
pulsed amperometry using a gold working electrode and an Ag/AgCl reference electrode 395
together with the Manufacturer’s electrode potential settings, optimized for carbohydrate 396
detection. The calibration standards were linearly related to detector response over the range 397
employed (0, 1.25, 2.5 and 5 nmols of glucose, fructose and sucrose) and the exudates were 398
diluted as necessary to give component signals which fell within the standard range (xylem 399
20-fold; phloem 2000-fold). An injection volume of 25 µL was used throughout. 400
401
Analysis of As species using anion exchange HPLC-ICP-MS 402
Aliquots (0.2 - 0.5 g) of the ground materials were extracted with 20 ml PBS for 1 h 403
under sonication. The extracts were filtered through 0.45 µm filters. Arsenic speciation in 404
phloem and xylem saps, nutrient solutions and plant extracts were determined using HPLC-405
ICP-MS (Agilent LC1100 series and Agilent ICP-MS 7500ce, Agilent Technologies, Santa 406
Clara, CA, US). Arsenic species [As(III), As(V), DMA(V), MMA(V), and MMA(III)] were 407
separated by an anion-exchange column (Hamilton PRP X-100, Reno, NV, USA) as described 408
previously (Li et al., 2009). The mobile phase was either 6.6 mM NH4H2PO4 and 6.6 mM 409
NH4NO3 (pH 6.3) with a flow rate of 0.6 ml min-1 (method 1), or 2 mM NH4H2PO4 and 2 mM 410
NH4NO3 (pH 6.3) with a flow rate of 0.8 ml min-1 (method 2). Method 1 was used to separate 411
As(III), As(V), MMA(V) and DMA(V), and method 2 As(III), MMA(V), MMA(III) and 412
DMA(V) (Li et al., 2009). The outlet of the separation column was connected to a concentric 413
nebulizer and a water-jacketed cyclonic spray chamber of the ICP-MS. Germanium (Ge) was 414
used as the internal standard which was mixed continuously with the post-column solution 415
through a peristaltic pump. Signals at mass-to-charge ratio (m/z) 75 (As), 72 (Ge), and 35 416
(chlorine [Cl]) were collected with a dwell time of 0.3 s for As and Ge and 0.1 s for Cl. 417
Possible polyatomic interference of 40Ar35Cl on m/z 75 was removed by the Agilent octopole 418
reaction system using helium as the reaction gas. The As signal was normalized by the Ge 419
signal to correct any signal drift during the analysis. Arsenic peaks were identified by 420
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

17
comparisons with the retention times of standard compounds. Analysis of As species was 421
carried out immediately following sample collection or extraction. For each batch of samples, 422
the analysis was completed within 12 h; repeated analyses showed no changes in As 423
speciation during this period of time. To test the stability of As species in xylem exudate 424
during collection and subsequent analysis, As(III) or As(V) was spiked to the exudate 425
collected from plants untreated with As. No evidence of As(III) oxidation or As(V) reduction 426
was found. 427
428
Analysis of thiols and As(III)-thiol complexes using reverse phase HPLC-high resolution 429
ICP-MS – high resolution ES-MS 430
Analysis of thiols and As(III)-thiol complexes in phloem and xylem saps was carried 431
out using HPLC coupled with high-resolution ICP-MS (Element 2, Thermo Fisher Scientific, 432
Bremen, Germany) and high resolution ES-MS (LTQ Orbitrap Discovery, Thermo Fisher 433
Scientific, Bremen, Germany). Separation was performed on a C18 reverse-phase column, 434
using a water-methanol gradient described previously (Bluemlein et al., 2008; Bluemlein et al., 435
2009; Liu et al., 2010). For the ICP-MS analysis, As and sulfur (S) were measured on m/z 75 436
and 32, respectively. Rhodium (Rh) was added post-column as internal standard and its 437
signal was measured on m/z 103. The ICP-MS was used in medium resolution. The molecular 438
masses of the eluted compounds were determined with high resolution on the ES-MS during a 439
chromatographic run, where glutathione from a xylem sap sample was used as an internal 440
standard for mass determination and correction. A retention time dependent element-specific 441
response using dimethylarsinic acid and methionine was used for quantification of any As and 442
S containing molecules as described in Bluemlein et al. (2008). 443
444
Analysis of total As in plant tissues 445
Ground plant samples were digested in 5 ml high purity HNO3/HClO4 (87/13, v/v). 446
Total As concentrations in the samples were determined by ICP-MS (Agilent 7500ce) 447
operating in the helium gas mode to remove possible interference of 40Ar35Cl on m/z 75. 448
449
Data analysis 450
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

18
Significance of treatment effects was tested by analysis of variance (ANOVA). Where 451
necessary, data were logarithmically transformed prior to ANOVA. Treatment means were 452
compared using the least significant difference (LSD). 453
454
Supplemental Materials: 455
Figure 1. HPLC-ICP-MS chromatograms of As species in roots, leaves, xylem and phloem 456
exudates of Ricinus communis exposed to 10 μM arsenate, DMA or MMA. 457
458
ACKNOWLEDGEMENT 459
We thank Dr. Richard Bromilow for advice on phloem sap collection. 460
461
Literature cited 462
463
Abedin MJ, Feldmann J, Meharg AA (2002) Uptake kinetics of arsenic species in rice 464
plants. Plant Physiol 128: 1120-1128 465
Allen S, Smith JAC (1986) Ammonium nutrition in Ricinus communis - its effect on plant 466
growth and the chemical composition of the whole plant, xylem and phloem saps. J 467
Exp Bot 37: 1599-1610 468
Bentley R, Chasteen TG (2002) Microbial methylation of metalloids: Arsenic, antimony, and 469
bismuth. Microbiology and Molecular Biology Reviews 66: 250-271 470
Bleeker PM, Hakvoort HWJ, Bliek M, Souer E, Schat H (2006) Enhanced arsenate 471
reduction by a CDC25-like tyrosine phosphatase explains increased phytochelatin 472
accumulation in arsenate-tolerant Holcus lanatus. Plant J 45: 917-929 473
Bluemlein K, Raab A, Feldmann J (2009) Stability of arsenic peptides in plant extracts: off-474
line versus on-line parallel elemental and molecular mass spectrometric detection for 475
liquid chromatographic separation. Anal Bioanal Chem 393: 357-366 476
Bluemlein K, Raab A, Meharg AA, Charnock JM, Feldmann J (2008) Can we trust mass 477
spectrometry for determination of arsenic peptides in plants: comparison of LC-ICP-478
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

19
MS and LC-ES-MS/ICP-MS with XANES/EXAFS in analysis of Thunbergia alata. 479
Anal Bioanal Chem 390: 1739-1751 480
Bromilow RH, Rigitano RLO, Briggs GG, Chamberlain K (1987) Phloem translocation of 481
non-ionised chemicals in Ricinus communis Pesticide Science 19: 85-99 482
Carey AM, Scheckel KG, Lombi E, Newville M, Choi Y, Norton GJ, Charnock JM, 483
Feldmann J, Price AH, Meharg AA (2010) Grain unloading of arsenic species in rice. 484
Plant Physiol 152: 309-319 485
Chen A, Komives EA, Schroeder JI (2006) An improved grafting technique for mature 486
Arabidopsis plants demonstrates long-distance shoot-to-root transport of 487
phytochelatins in Arabidopsis. Plant Physiol 141: 108-120 488
Chen W, Chi Y, Taylor NL, Lambers H, Finnegan PM (2010) Disruption of ptLPD1 or 489
ptLPD2, genes that encode isoforms of the plastidial lipoamide dehydrogenase, confers 490
arsenate hypersensitivity in Arabidopsis thaliana. Plant Physiol 153: 1385–1397 491
Dhankher OP, Li YJ, Rosen BP, Shi J, Salt D, Senecoff JF, Sashti NA, Meagher RB 492
(2002) Engineering tolerance and hyperaccumulation of arsenic in plants by combining 493
arsenate reductase and gamma-glutamylcysteine synthetase expression. Nature 494
Biotechnology 20: 1140-1145 495
Dhankher OP, Rosen BP, McKinney EC, Meagher RB (2006) Hyperaccumulation of 496
arsenic in the shoots of Arabidopsis silenced for arsenate reductase (ACR2). Proc Nat 497
Acad Sci USA 103: 5413-5418 498
Duan GL, Zhou Y, Tong YP, Mukhopadhyay R, Rosen BP, Zhu YG (2007) A CDC25 499
homologue from rice functions as an arsenate reductase New Phytol 174: 311-321 500
Ellis DR, Gumaelius L, Indriolo E, Pickering IJ, Banks JA, Salt DE (2006) A novel 501
arsenate reductase from the arsenic hyperaccumulating fern Pteris vittata. Plant 502
Physiol 141: 1544-1554 503
European Food Safety Authority (2009) Scientific opinion on arsenic in food. EFSA Journal 504
7: 1351 505
Feldmann J, Salaun P, Lombi E (2009) Elemental speciation analysis methods in 506
environmental chemistry - moving towards methodological integration. Environ Chem 507
6: 275-289 508
Hall SM, Baker DA (1972) Chemical composition of Ricinus phloem exudate. Planta 106: 509
131-& 510
Hall SM, Baker DA, Milburn JA (1971) Phloem transport of 14C-labelled assimilates in 511
Ricinus. Planta 100: 200-& 512
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

20
Huang JWW, Poynton CY, Kochian LV, Elless MP (2004) Phytofiltration of arsenic from 513
drinking water using arsenic-hyperaccumulating ferns. Environ Sci Technol 38: 3412-514
3417 515
Indriolo E, Na GN, Ellis D, Salt DE, Banks JA (2010) A vacuolar arsenite transporter 516
necessary for arsenic tolerance in the arsenic hyperaccumulating fern Pteris vittata is 517
missing in flowering plants. Plant Cell: in press 518
Johanning J, Strasdeit H (1998) A coordination-chemical basis for the biological function of 519
the phytochelatins. Angewandte Chemie-International Edition 37: 2464-2466 520
Li RY, Ago Y, Liu WJ, Mitani N, Feldmann J, McGrath SP, Ma JF, Zhao FJ (2009) The 521
rice aquaporin Lsi1 mediates uptake of methylated arsenic species. Plant Physiol 150: 522
2071-2080 523
Li YJ, Dankher OP, Carreira L, Smith AP, Meagher RB (2006) The shoot-specific 524
expression of gamma-glutamylcysteine synthetase directs the long-distance transport 525
of thiol-peptides to roots conferring tolerance to mercury and arsenic. Plant Physiol 526
141: 288-298 527
Liu WJ, Wood BA, Raab A, McGrath SP, Zhao FJ, Feldmann J (2010) Complexation of 528
Arsenite with Phytochelatins Reduces Arsenite Efflux and Translocation from Roots to 529
Shoots in Arabidopsis. Plant Physiol 152: 2211-2221 530
Ma JF, Yamaji N, Mitani N, Xu XY, Su YH, McGrath SP, Zhao FJ (2008) Transporters 531
of arsenite in rice and their role in arsenic accumulation in rice grain. Proc Nat Acad 532
Sci USA 105: 9931–9935 533
Ma LQ, Komar KM, Tu C, Zhang WH, Cai Y, Kennelley ED (2001) A fern that 534
hyperaccumulates arsenic. Nature 409: 579-579 535
Marin AR, Masscheleyn PH, Patrick WH (1992) The influence of chemical form and 536
concentration of arsenic on rice growth and tissue arsenic concentration. Plant Soil 139: 537
175-183 538
Meharg AA, Williams PN, Adomako E, Lawgali YY, Deacon C, Villada A, Cambell RCJ, 539
Sun G, Zhu YG, Feldmann J, Raab A, Zhao FJ, Islam R, Hossain S, Yanai J 540
(2009) Geographical variation in total and inorganic arsenic content of polished (white) 541
rice. Environ Sci Technol 43: 1612-1617 542
Mendoza-Cózatl DG, Butko E, Springer F, Torpey JW, Komives EA, Kehr J, Schroeder 543
JI (2008) Identification of high levels of phytochelatins, glutathione and cadmium in 544
the phloem sap of Brassica napus. A role for thiol-peptides in the long-distance 545
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

21
transport of cadmium and the effect of cadmium on iron translocation. Plant J 54: 249-546
259 547
Mihucz VG, Tatar E, Virag I, Cseh E, Fodor F, Zaray G (2005) Arsenic speciation in 548
xylem sap of cucumber (Cucumis sativus L.). Anal Bioanal Chem 383: 461-466 549
Panaullah GM, Alam T, Hossain MB, Loeppert RH, Lauren JG, Meisner CA, Ahmed 550
ZU, Duxbury JM (2009) Arsenic toxicity to rice (Oryza sativa L.) in Bangladesh. 551
Plant Soil 317: 31–39 552
Pickering IJ, Prince RC, George MJ, Smith RD, George GN, Salt DE (2000) Reduction 553
and coordination of arsenic in Indian mustard. Plant Physiol 122: 1171-1177 554
Poynton CY, Huang JWW, Blaylock MJ, Kochian LV, Elless MP (2004) Mechanisms of 555
arsenic hyperaccumulation in Pteris species: root As influx and translocation. Planta 556
219: 1080-1088 557
Raab A, Feldmann J, Meharg AA (2004) The nature of arsenic-phytochelatin complexes in 558
Holcus lanatus and Pteris cretica. Plant Physiol 134: 1113-1122 559
Raab A, Schat H, Meharg AA, Feldmann J (2005) Uptake, translocation and transformation 560
of arsenate and arsenite in sunflower (Helianthus annuus): formation of arsenic-561
phytochelatin complexes during exposure to high arsenic concentrations. New Phytol 562
168: 551-558 563
Raab A, Williams PN, Meharg A, Feldmann J (2007) Uptake and translocation of inorganic 564
and methylated arsenic species by plants. Environ Chem 4: 197-203 565
Salido AL, Hasty KL, Lim JM, Butcher DJ (2003) Phytoremediation of arsenic and lead in 566
contaminated soil using Chinese Brake Ferns (Pteris vittata) and Indian mustard 567
(Brassica juncea). International Journal of Phytoremediation 5: 89-103 568
Schmöger MEV, Oven M, Grill E (2000) Detoxification of arsenic by phytochelatins in 569
plants. Plant Physiol 122: 793-801 570
Smith JAC, Milburn JA (1980) Osmoregulation and the control of phloem sap composition 571
in Ricinus communis L. Planta 148: 28-34 572
Su YH, McGrath SP, Zhao FJ (2010) Rice is more efficient in arsenite uptake and 573
translocation than wheat and barley. Plant Soil 328: 27-34 574
Su YH, McGrath SP, Zhu YG, Zhao FJ (2008) Highly efficient xylem transport of arsenite 575
in the arsenic hyperaccumulator Pteris vittata. New Phytol 180: 434-441 576
Ullrich-Eberius CI, Sanz A, Novacky AJ (1989) Evaluation of arsenate- and vanadate-577
associated changes of electrical membrane potential and phosphate transport in Lemna 578
gibba-G1. J Exp Bot 40: 119-128 579
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

22
Wang JR, Zhao FJ, Meharg AA, Raab A, Feldmann J, McGrath SP (2002) Mechanisms 580
of arsenic hyperaccumulation in Pteris vittata. Uptake kinetics, interactions with 581
phosphate, and arsenic speciation. Plant Physiol 130: 1552-1561 582
Williams PN, Villada A, Deacon C, Raab A, Figuerola J, Green AJ, Feldmann J, Meharg 583
AA (2007) Greatly enhanced arsenic shoot assimilation in rice leads to elevated grain 584
levels compared to wheat and barley. Environ Sci Technol 41: 6854-6859 585
Xu XY, McGrath SP, Zhao FJ (2007) Rapid reduction of arsenate in the medium mediated 586
by plant roots. New Phytol 176: 590-599 587
Zhao FJ, Dunham SJ, McGrath SP (2002) Arsenic hyperaccumulation by different fern 588
species. New Phytol 156: 27-31 589
Zhao FJ, Ma JF, Meharg AA, McGrath SP (2009) Arsenic uptake and metabolism in plants. 590
New Phytol 181: 777-794. 591
Zhao FJ, McGrath SP, Meharg AA (2010a) Arsenic as a food-chain contaminant: 592
mechanisms of plant uptake and metabolism and mitigation strategies. Ann Rev Plant 593
Biol 61: 535–559 594
Zhao FJ, Stroud JL, Eagling T, Dunham SJ, McGrath SP, Shewry PR (2010b) 595
Accumulation, distribution and speciation of arsenic in wheat grain. Environ Sci 596
Technol 44: 5464–5468 597
Zhao FJ, Wang JR, Barker JHA, Schat H, Bleeker PM, McGrath SP (2003) The role of 598
phytochelatins in arsenic tolerance in the hyperaccumulator Pteris vittata. New Phytol 599
159: 403-410 600
601
602
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

23
Table 1. The composition of sugars in phloem and xylem exudates of Ricinus communis 603
(mean ± SE, n = 4). 604
Exudate Sucrose (mM)
Glucose (mM)
Fructose (mM)
Phloem 238 ± 15.8 0.12 ± 0.12 0 ± 0
Xylem 1.20 ± 0.32 0.29 ± 0.08 0.36 ± 0.11 605
606
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

24
Table 2. The concentrations of sulfate, reduced glutathione (GSH) and oxidized glutathione 607
(GSSG) and oxidized PC2 in phloem and xylem exudates of Ricinus communis exposed to 10 608
μM arsenate for 48 h (mean ± SE, n = 2). 609
Exudate
Sulfur species (nmol g-1)
Sulfate GSH GSSG Oxidized PC2
Phloem 141 ± 56 64 ± 33 22 ± 1.4 1.2 ± 0.7
Xylem 131 ± 8 0 ± 0 5.8 ± 0.8 0.2 ± 0.01 610
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

25
Figure captions: 611
Fig. 1. Arsenic speciation in phloem exudate (a), xylem exudate (b), roots (c) and leaves of 612
Ricinus communis exposed to 10 µM arsenate or arsenite for 24 or 48 h. Error bars are one SE 613
(n = 3 - 4). 614
Fig. 2. Arsenic speciation in phloem exudate (a), xylem exudate (b), roots (c) and leaves of 615
Ricinus communis exposed to 2 - 100 µM arsenate for 24 h. Error bars are one SE (n = 3 - 4, 616
except the 50 µM treatment in which only one replicate produced xylem exudate; the 100 µM 617
treatment produced no xylem exudate). 618
Fig. 3. Arsenic speciation in phloem exudate (a) and xylem exudate (b) of Ricinus communis 619
exposed to 10 µM arsenate with either 10 µM (low P) or 200 µM phosphate (high P) for 24 or 620
48 h. Error bars are one SE (n = 3 - 4). 621
Fig. 4. Overlaid RP-HPLC chromatograms for the phloem (a) and xylem (b) exudates detected 622
online simultaneously by using ICP-MS (m/z 32 and 75) and ES-MS (m/z 308, 538 and 613, 623
[M+H]+). The sulfur trace (m/z 32) shows the relative quantity of the inorganic sulfur 624
compounds eluting near the void at 2 min compared to GSSG and oxidized PC2. 625
Fig. 5. Arsenic speciation in phloem exudate (a), xylem exudate (b), roots (c) and leaves of 626
Ricinus communis exposed to 10 µM arsenate, MMA(V) or DMA(V) for 24 or 48 h. Error 627
bars are one SE (n = 3 – 4). 628
www.plantphysiol.orgon May 13, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.

Figure 1. Arsenic speciation in phloem exudate (a), xylem exudate (b), roots (c) and leaves of Ricinus communis
exposed to 10 µM arsenate or arsenite for 24 or 48 h. Error bars are one SE (n = 3 - 4).
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
As(III), 24h As(V), 24h As(III), 48h As(V), 48h
Con
cent
ratio
n (μ
M)
PhloemAs(V)
As(III)
0
0.1
0.2
0.3
0.4
0.5
0.6
As(III), 48h As(V), 48h
Con
cent
ratio
n (μ
M)
XylemAs(V)
As(III)
0
50
100
150
200
250
300
As(III) As(V)
Con
cent
ratio
n (n
mol
/g F
W)
Roots
As(V)
As(III)
0
2
4
6
8
10
As(III) As(V)
Con
cent
ratio
n (n
mol
/g F
W)
Leaves
As(V)
As(III)
(c) (d)
(b)(a)
w
ww
.plantphysiol.orgon M
ay 13, 2018 - Published by
Dow
nloaded from
Copyright ©
2010 Am
erican Society of P
lant Biologists. A
ll rights reserved.
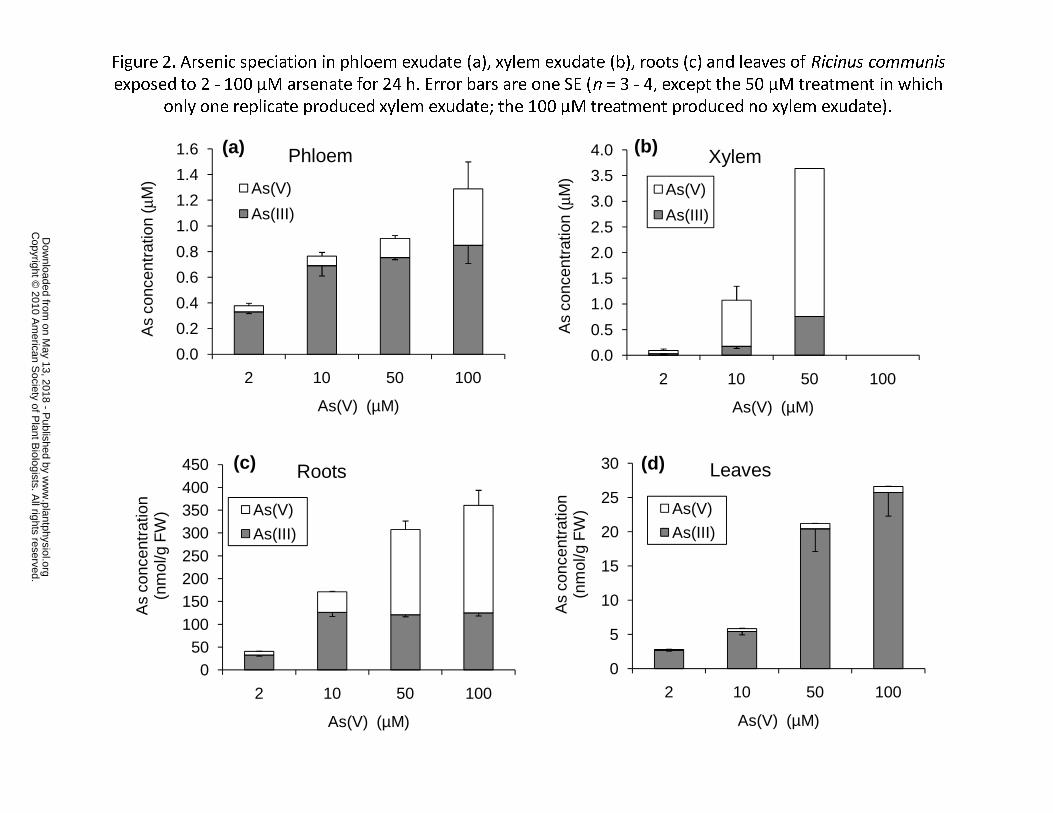
Figure 2. Arsenic speciation in phloem exudate (a), xylem exudate (b), roots (c) and leaves of Ricinus communis
exposed to 2 - 100 µM arsenate for 24 h. Error bars are one SE (n = 3 - 4, except the 50 µM treatment in which
only one replicate produced xylem exudate; the 100 µM treatment produced no xylem exudate).
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
2 10 50 100
As
conc
entr
atio
n (μ
M)
As(V) (µM)
Phloem
As(V)
As(III)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
2 10 50 100
As
conc
entr
atio
n (μ
M)
As(V) (µM)
Xylem
As(V)
As(III)
0
5
10
15
20
25
30
2 10 50 100
As
conc
entr
atio
n (n
mol
/g F
W)
As(V) (µM)
Leaves
As(V)
As(III)
050
100150200250300350400450
2 10 50 100
As
conc
entr
atio
n (n
mol
/g F
W)
As(V) (µM)
Roots
As(V)
As(III)
(a) (b)
(c) (d)
w
ww
.plantphysiol.orgon M
ay 13, 2018 - Published by
Dow
nloaded from
Copyright ©
2010 Am
erican Society of P
lant Biologists. A
ll rights reserved.

Figure. 3. Arsenic speciation in phloem exudate (a) and xylem exudate (b) of Ricinus communis exposed to 10 µM
arsenate with either 10 µM (low P) or 200 µM phosphate (high P) for 24 or 48 h. Error bars are one SE (n = 3 - 4).
0.0
0.2
0.4
0.6
0.8
1.0
Low P, 24h High P, 24 h Low P, 48h High P, 48 h
Con
cent
ratio
n (μ
M)
Phloem As(V)
As(III)
0.0
0.2
0.4
0.6
0.8
1.0
Low P, 48h High P, 48 h
Con
cent
ratio
n (μ
M)
Xylem
As(V)
As(III)
(a) (b)
w
ww
.plantphysiol.orgon M
ay 13, 2018 - Published by
Dow
nloaded from
Copyright ©
2010 Am
erican Society of P
lant Biologists. A
ll rights reserved.

Figure 4. Overlaid RP-HPLC
chromatograms for the phloem (a) and
xylem (b) exudates detected online
simultaneously by using ICP-MS (m/z 32
and 75) and ES-MS (m/z 308, 538 and 613,
[M+H]+). The sulfur trace (m/z 32) shows
the relative quantity of the inorganic
sulfur compounds eluting near the void at
2 min compared to GSSG and oxidized PC2.
Retention time, min
0 10 20 30 40
Intensity, arb
itrary
m/z 75 (As)
m/z 32 (S)
m/z 613 (GSSG)
m/z 538 (ox PC2)
b)
Retention time, min
0 10 20 30 40
Intensity, a
rbitra
ry
m/z 75 (As)
m/z 32 (S)
m/z 308 (GSH)
m/z 613 (GSSG)
m/z 538 (ox PC2)a)
w
ww
.plantphysiol.orgon M
ay 13, 2018 - Published by
Dow
nloaded from
Copyright ©
2010 Am
erican Society of P
lant Biologists. A
ll rights reserved.

0
1
2
3
4
5
As(V) MMA DMA
Con
cent
ratio
n (n
mol
/g F
W) Shoots DMA(V)
MMA(V)
As(V)
As(III)
0
20
40
60
80
100
120
140
As(V) MMA DMA
Con
cent
ratio
n (n
mol
/g F
W) Roots DMA(V)
MMA(III)
MMA(V)
As(V)
As(III)
0.0
0.2
0.4
0.6
0.8
1.0
Con
cent
ratio
n (μ
M) Xylem DMA(V)
MMA(V)
As(V)
As(III)
0
1
2
3
Con
cent
ratio
n (μ
M) Phloem DMA(V)
MMA(V)
As(V)
As(III)
Figure 5. Arsenic speciation in phloem exudate (a), xylem exudate (b), roots (c) and leaves of Ricinus communis exposed
to 10 µM arsenate, MMA(V) or DMA(V) for 24 or 48 h. Error bars are one SE (n = 3 – 4).
(b)
(c) (d)
(a)
w
ww
.plantphysiol.orgon M
ay 13, 2018 - Published by
Dow
nloaded from
Copyright ©
2010 Am
erican Society of P
lant Biologists. A
ll rights reserved.





























