Embed Size (px)
Citation preview

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 1/121
PRINCÍPIOS DE NEFROLOGIA EDISTÚRBIOS HIDROELETROLÍTICOS

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 2/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 3/121
PRINCÍPIOS DENEFROLOGIA
E
DISTÚRBIOS HIDROELETROLÍTICOS
Miguel Carlos RiellaProfessor Titular de Clínica Médica da Pontifícia Universidade Católica do Paraná.
Professor Titular de Clínica Médica e Diagnóstico da Faculdade Evangélica de Medicina do Paraná.Diretor do Serviço de Nefrologia do Hospital Universitário Evangélico de Curitiba.Ex-Research Fellow em Nefrologia na University of Washington (Seattle, U.S.A.).
Doutor em Medicina pela UNIFESP
QUARTA EDIÇÃO

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 4/121
NOTA DA EDITORA: A área da saúde é um campo em constante mudança. As normasde segurança padronizadas precisam ser obedecidas; contudo, à medida que as novaspesquisas ampliam nossos conhecimentos, tornam-se necessárias e adequadas modificaçõesterapêuticas e medicamentosas. O autor desta obra verificou cuidadosamente os nomesgenéricos e comerciais dos medicamentos mencionados, bem como conferiu os dadosreferentes à posologia, de modo que as informações fossem acuradas e de acordo com ospadrões aceitos por ocasião da publicação. Todavia, os leitores devem prestar atenção àsinformações fornecidas pelos fabricantes, a fim de se certificarem de que as dosespreconizadas ou as contra-indicações não sofreram modificações. Isso é importante,sobretudo em relação a substâncias novas ou prescritas com pouca freqüência. O autor e aeditora não podem ser responsabilizados pelo uso impróprio ou pela aplicação incorreta doproduto apresentado nesta obra.
No interesse de difusão da cultura e do conhecimento, o autor e os editores envidaram omáximo esforço para localizar os detentores dos direitos autorais de qualquer material
utilizado, dispondo-se a possíveis acertos posteriores caso, inadvertidamente, a identificaçãode algum deles tenha sido omitida.
Capa: Leonardo Vidal Riella
Direitos exclusivos para a língua portuguesaCopyright © 2003 byEDITORA GUANABARA KOOGAN S.A.
Travessa do Ouvidor, 11Rio de Janeiro, RJ — CEP 20040-040Tel.: 21–2221-9621Fax: 21–2221-3202www.editoraguanabara.com.br
Reservados todos os direitos. É proibida a duplicaçãoou reprodução deste volume, no todo ou em parte,sob quaisquer formas ou por quaisquer meios(eletrônico, mecânico, gravação, fotocópia,distribuição na Web, ou outros),sem permissão expressa da Editora.
Editoração Eletrônica:

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 5/121
COLABORADORES
ABRAHÃO SALOMÃO FILHOProfessor Adjunto do Departamento de Clínica Médica da FM/ UFMG. Coordenador do Grupo de Transplantes Renais do HU/ UFMG
ALEXANDER J. ROUCH Associate Professor of Physiology in the Department of Physiologyat Oklahoma State University Center for Health Sciences
ALUIZIO BARBOSA DE CARVALHOProfessor Afiliado Doutor da Disciplina de Nefrologia da EscolaPaulista de Medicina — UNIFESP. Responsável pelo Setor de Do-enças Ósseas do Hospital do Rim da Fundação Oswaldo Ramos —
Hospital do Rim e Hipertensão
AMÉRICO LOURENÇO CUVELLO NETODoutor em Nefrologia pela Faculdade de Medicina da Universidadede São Paulo (FMUSP). Médico Assistente do Grupo de Insuficiên-cia Renal Aguda, Hospital das Clínicas — FMUSP
ANTONIO BARBIERIProfessor Adjunto do Departamento de Diagnóstico por Imagem daUniversidade Federal de São Paulo. Coordenadoria de Radioisóto-
pos da Escola Paulista de Medicina
ANTONIO CARLOS SEGUROProfessor Livre-Docente de Nefrologia do Hospital das Clínicas daFaculdade de Medicina da USP
ANTONIO JOSÉ BARROS MAGALDIProfessor Assistente Doutor e Professor Colaborador da USP-Labo-ratório e Pesquisa Básica LIM 12 — Nefrologia — Hospital dasClínicas da FMUSP
ANTONIO MARMO LUCONProfessor Assistente da Disciplina de Urologia. Departamen-to Cirúrgico da Faculdade de Medicina da Universidade de SãoPaulo
CARLOS JADER FELDMAN
Médico-Chefe do Serviço de Radiologia do Instituto de Cardiologiado Rio Grande do Sul/FUC
CARLOS PEREZ GOMES Mestre em Nefrologia. Nefrologista do Hospital da Polícia Civil JCM. Médico Nefrologista da Pró-Renal
CESAR COSTAProfessor Titular do Departamento de Medicina Interna, Faculdadede Medicina da Universidade Federal do Rio Grande do Sul. Médi-co Assistente do Serviço de Nefrologia do Hospital de Clínicas dePorto Alegre, RS
CHRISTOPHE LEGENDREProfessor Adjunto da Universidade de Paris V, Université RenéDescartes, Faculté de Médécine Necker-Enfants Malades, Serviço de
Transplante Renal do Hospital Necker, Paris, França
CIBELE ISAAC SAAD RODRÍGUEZProfessora Titular do Departamento de Medicina, Disciplina deNefrologia, da Faculdade de Ciências Médicas de Sorocaba — PUC/ SP. Mestrado e Doutorado em Nefrologia na UNIFESP-EMP
CLÁUDIA MARIA DE BARROS HELOUProfessora Assistente Doutora, Disciplina de Nefrologia, do Hospitaldas Clínicas da Faculdade de Medicina da Universidade de São Paulo
CRISTINA MARTINS Mestre em Nutrição Clínica pela New York University, U.S.A.Doutoranda em Ciências Médicas, Disciplina de Nefrologia, UFRGS.Nutricionista-Chefe, Clínica de Doenças Renais — Curitiba, PR.Professora Assistente — Curso de Nutrição PUC/PR. Diretora Ge-ral da Nutroclínica
DALTRO ZUNINO Mestre em Pediatria. Especialista em Nefrologia Pediátrica. Respon-sável pelo Setor de Nefropediatria dos Serviços de Pediatria eNefrologia do Hospital Universitário Evangélico de Curitiba, PR
DÉCIO MION JR.Professor Livre-Docente. Chefe da Unidade de Hipertensão do Hos-
pital das Clínicas, Disciplina de Nefrologia, da Faculdade de Medi-cina da Universidade de São Paulo
E. BARSANULFO PEREIRAProfessor Titular de Clínica Médica, Disciplinas de Nefrologiae Farmacologia Clínica, Universidade Federal de Mato Grosso do Sul
EDISON MATOS NÓVAKNeurologista. Professor Adjunto de Neurologia — UFPR. Mestreem Medicina Interna. Membro Titular da Academia Brasileira deNeurologia. Membro Titular da Sociedade Brasileira de Investiga-ção Neurológica

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 6/121
vi Colaboradores
EDIVALDO CELSO VIDALProfessor Doutor da Faculdade de Medicina da Universidade Fede-ral de Uberlândia, MG
EDNA REGINA SILVA PEREIRAProfessora Adjunta. Chefe do Serviço de Nefrologia do Hospital dasClínicas, Departamento de Clínica Médica da Universidade Federalde Goiás
ELIAS ASSAD WARRAKProfessor Auxiliar da Disciplina de Nefrologia da UniversidadeFederal Fluminense
EMIL SABBAGAProfessor Associado Livre-Docente de Nefrologia da Faculdade de
Medicina da Universidade de São Paulo
EMMANUEL DE ALMEIDA BURDMANNProfessor Livre-Docente da Faculdade de Medicina da Universi-dade de São Paulo. Professor Adjunto da Disciplina de Nefrologiada Faculdade de Medicina de São José do Rio Preto, SP
ERIC ROGER WROCLAWSKIProfessor Adjunto e Regente da Disciplina de Urologia da Facul-dade de Medicina do ABC. Responsável pelo Serviço de Urologiado Hospital Universitário da Fundação do ABC. Urologista do Hos-
pital Israelita Albert Einstein. Chefe do Setor Cirúrgico da Uni-dade de Transplante Renal do Instituto Dante Pazzanese de Car-diologia de São Paulo. Presidente da Sociedade Brasileira deUrologia
EUTHYMIA BRANDÃO A. PRADODocente do Curso de Pós-Graduação em Nefrologia da Faculdade de
Medicina da Universidade de São Paulo
FERNANDO ANTONIO DE ALMEIDA
Professor Titular do Departamento de Medicina, Disciplina deNefrologia, Faculdade de Ciências Médicas e Biológicas de Sorocaba
— PUC/SP. Doutor em Nefrologia — Escola Paulista de Medicina.Post-Doctoral Fellow Cornell University Medical College, NewYork, U.S.A.
FERNANDO MEYER Mestre em Clínica Cirúrgica pela UFPR. Professor Assistente doCurso de Medicina da PUC/PR
FLÁVIO M. BARBOSA Médico Residente do Serviço de Radiologia do Instituto de Cardio-logia do Rio Grande do Sul/FUC
GILBERTO ALONSOProfessor Adjunto do Departamento Diagnóstico por Imagem da
Escola Paulista de Medicina — Universidade Federal de São Paulo
GILVAN NEIVA FONSECADoutor em Urologia pela UNIFESP — Escola Paulista de Medicina.Professor Adjunto de Urologia do Serviço de Urologia da Faculdadede Medicina da Universidade Federal de Goiás
HENRI KREISProfessor Adjunto da Universidade de Paris V, Université RenéDescartes, Faculté de Médécine Necker-Enfants Malades, Serviço deTransplante Renal do Hospital Necker, Paris, França
HENRY DE HOLANDA CAMPOSProfessor Adjunto do Departamento de Medicina Clínica da Uni-versidade Federal do Ceará. Professor Associado da Universidade deParis V, Université René Descartes, Faculté de Médécine Necker-Enfants Malades, Paris, França
HEONIR ROCHAProfessor Titular do Departamento de Medicina — Faculdade de
Medicina da Universidade Federal da Bahia
IRENE L. NORONHADoutora em Imunologia de Transplantes pela Universidade de
Heidelberg, Alemanha. Professora Livre-Docente Doutora da Dis-ciplina de Nefrologia do Hospital das Clínicas da Faculdade de
Medicina da USP. Staff da Clínica de Nefrologia do Hospital daBeneficência Portuguesa de São Paulo. Coordenadora do Progra-ma Clínico de Transplante de Pâncreas-Rim do Hospital da Be-neficência Portuguesa de São Paulo e Hospital Israelita AlbertEinstein
IRINA ANTUNESDoutora em Nefrologia pela Universidade de São Paulo. MédicaColaboradora do Serviço de Nefrologia do Hospital das Clínicas daFMUSP
ISTÊNIO FERNANDES PASCOALDoutor em Medicina pela Universidade de São Paulo. Pós-Douto-rado pela Universidade de Chicago. Nefrologista Clínico em Brasí-lia, DF
JOÃO PROCÓPIO FORTES JÚNIOR
Mestre em Radiologia pela Universidade Federal de São Paulo. Físi-co Especialista em Medicina Nuclear
JOCEMIR RONALDO LUGONProfessor Titular de Nefrologia da Universidade Federal Fluminense.Rio de Janeiro, RJ
JORGE KALILProfessor Titular de Imunologia Clínica e Alergia, Diretor do Labo-ratório de Imunologia do Instituto do Coração, Faculdade de Medi-cina da Universidade de São Paulo
JORGE LUIZ GROSSProfessor Titular do Departamento de Medicina Interna da Facul-dade de Medicina da Universidade Federal do Rio Grande do Sul.Chefe do Serviço de Endocrinologia do Hospital de Clínicas de Por-
to Alegre, RS
JORGE PAULO STROGOFF DE MATOS Mestre em Nefrologia pela Universidade do Estado do Rio de Janeiro. Médico Nefrologista da Clínica de Doenças Renais, Rio de Janeiro

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 7/121
Colaboradores vii
JOSÉ HERMÓGENES ROCCO SUASSUNA Mestre em Medicina — Nefrologia. Doutor em Ciências — Microbiologia e Imunologia. Médico da Kidney Assistance/CDR
Serviços Hospitalares. Professor Adjunto, Disciplina de Nefrologia,da Faculdade de Ciências Médicas da Universidade do Estado do Riode Janeiro (UERJ)
JOSÉ LUIZ MONTEIROProfessor Assistente Doutor da Disciplina de Nefrologia da Facul-dade de Medicina da Universidade de São Paulo
JOSÉ MAURO VIEIRA JÚNIOR Assistente Doutor do LIM 16 da Faculdade de Medicina da Univer-sidade de São Paulo
JOSÉ NERY PRAXEDESProfessor Doutor da Disciplina de Nefrologia da Faculdade de Me-dicina da Universidade de São Paulo
LEONARDO VIDAL RIELLA
Doutorando em Medicina pela Universidade Federal do Paraná
LINEU CÉSAR WERNECKProfessor Titular de Neurologia — UFPR. Doutor em Neurologia.Chefe do Serviço de Neurologia e Doenças Neuromusculares do Hos-
pital de Clínicas — UFPR. Membro Titular da Academia Brasileirade Neurologia. Membro Titular da Sociedade Brasileira de Investi-
gação Neurológica. Membro da American Academy of Neurology. Membro da American Neurological Association
LUCIA H. KUDOProfessora Aposentada pela Universidade de São Paulo e Atual Re-search Scientist no OSU Center for Health Sciences, Tulsa, Oklaho-ma, U.S.A.
LÚCIO SILVA
Médico Nefrologista do Instituto Mineiro de Nefrologia. Coordena-dor da Clínica de Nefrologia e Hemodiálise do Hospital Belo Hori-zonte. Preceptor da Residência Médica do Hospital Belo Horizonte
LUÍS HENRIQUE CANANIPesquisador Associado do Serviço de Endocrinologia do Hospital deClínicas de Porto Alegre. Pós-Doutorado na Unidade de Genética eEpidemiologia da Clínica Joslin, Boston, U.S.A.
LUIS YUProfessor Livre-Docente da Disciplina de Nefrologia da Faculdadede Medicina da Universidade de São Paulo (FMUSP). Médico Res-
ponsável pelo Grupo de Insuficiência Renal Aguda, Hospital dasClínicas — FMUSP
LUIZ ANTONIO RIBEIRO DE MOURA
Professor Adjunto Doutor, Departamento de Patologia, Faculdadede Medicina da Universidade Federal de São Paulo
LUIZ APARECIDO BORTOLOTTODoutor em Cardiologia. Médico Assistente da Unidade de Hiperten-são do Instituto do Coração
LUIZ ESTEVAM IANHEZProfessor Livre-Docente de Nefrologia da Faculdade de Medicina daUSP — São Paulo. Chefe do Setor Clínico da Unidade de Transplante
Renal da Divisão de Clínica Urológica — Hospital das Clínicas daFMUSP. Médico Nefrologista do Hospital Alemão Oswaldo Cruz
LUIZ SERGIO SANTOSProfessor Adjunto da Disciplina de Urologia da PUC/PR
MARCELLO FABIANO DE FRANCOProfessor Titular de Patologia da Escola Paulista de Medicina —Universidade Federal de São Paulo
MARCELO MAZZA DO NASCIMENTO Mestre em Medicina Interna pela Universidade Federal do Paraná.Doutor em Medicina (2003), Universidade Federal do Rio Grande doSul. Chefe do Serviço de Hemodiálise do Hospital Universitário Evan-
gélico de Curitiba da Faculdade Evangélica de Medicina do Paraná
MÁRCIA CRISTINA DE ALMEIDA
Mestre em Nefrologia pela Universidade Federal de São Paulo. Es- pecialista em Medicina Nuclear
MARCOS ALEXANDRE VIEIRA Médico da Fundação Pró-Rim de Santa Catarina. Nefrologista For-mado pelo Hospital Universitário Evangélico de Curitiba
MARIA APARECIDA PACHALLYNefrologista do Hospital Universitário de Curitiba. Especialista emTerapia Intensiva
MARIA DE FÁTIMA SANTOS BANDEIRANefrologista da Clínica de Doenças Renais, RJ
MARIA FERNANDA CORDEIRO DE CARVALHOProfessora Doutora em Nefrologia da Faculdade de Medicina de
Botucatu — UNESP
MARIANGELA M. COSNER Médica Radiologista do Serviço de Radiologia do Instituto de Car-diologia do Rio Grande do Sul/FUC
MÁRIO ABBUD FILHOProfessor Adjunto, Departamento de Medicina, Faculdade de Me-dicina de São José do Rio Preto (FAMERP). Diretor do Laboratóriode Imunologia de Transplantes, Instituto de Urologia e Nefrologiade S. J. do Rio Preto, SP. Diretor do Centro Interdepartamental deTransplantes de Órgãos da FAMERP/FUNFARME
MARISTELA CARVALHO DA COSTADoutora em Nefrologia pela Faculdade de Medicina da Universi-dade de São Paulo (FMUSP). Médica Responsável pela UTI
Nefrologia do Hospital das Clínicas — FMUSP
MAURI FÉLIX DE SOUZA Mestre em Nefrologia pela Faculdade de Medicina da UFRGS. Pro- fessor Assistente de Nefrologia da Faculdade de Medicina da UFG.Pós-Graduando da Faculdade de Medicina — USP a nível de doutorado

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 8/121
viii Colaboradores
MAURÍCIO DE CARVALHOProfessor Adjunto, Disciplina de Nefrologia, da Pontifícia Univer-sidade Católica do Paraná. Preceptor da Residência em Medicina
Interna da Universidade Federal do Paraná. Ex-Research Fellow emNefrologia, Universidade de Chicago, U.S.A.
MIGUEL CARLOS RIELLAProfessor Titular de Clínica Médica da Pontifícia UniversidadeCatólica do Paraná. Professor Titular de Clínica Médica e Diag-nóstico da Faculdade Evangélica de Medicina do Paraná. Dire-tor do Serviço de Nefrologia do Hospital Univers itário Evangéli-co de Curitiba. Ex-Research Fellow em Nefrologia na Universityof Washington (Seattle, U.S.A.). Doutor em Medicina pelaUNIFESP
MIGUEL CENDOROGLO NETOProfessor Adjunto da Disciplina de Nefrologia da Escola Paulistade Medicina — Universidade Federal de São Paulo
MIGUEL LUIS GRACIANO
Pós-Graduando em Nefrologia (Doutorado), USP, São Paulo, SP. Mestre em Nefrologia, UFRJ, Rio de Janeiro, RJ. Nefrologista, Hos- pital Universitário Antônio Pedro, UFF, Niterói, RJ. Nefrologista, Hospital Geral de Bonsucesso, Rio de Janeiro, RJ
MIRELA JOBIM DE AZEVEDOProfessora Adjunta do Departamento de Medicina Interna da Fa-culdade de Medicina da Universidade Federal do Rio Grande do Sul.Livre-Docente pela Disciplina de Endocrinologia da Escola Paulis-ta de Medicina — Universidade Federal de São Paulo
MIRIAN APARECIDA BOIMProfessora Afiliada da Disciplina de Nefrologia da Escola Paulistade Medicina — Universidade Federal de São Paulo
NESTOR SCHOR
Pró-Reitor de Pós-Graduação e Pesquisa. Professor Titular da Dis-ciplina de Nefrologia da Escola Paulista de Medicina — Universi-dade Federal de São Paulo
NICOLAS PANAJOTOPOULOSPesquisador do Laboratório de Imunologia do Instituto do Coração,Faculdade de Medicina da Universidade de São Paulo
NOEMIA PERLI GOLDRAICHDoutora em Nefrologia pela UNIFESP. Professora Adjunta do De-
partamento de Pediatria e Puericultura da Faculdade de Medicinada Universidade Federal do Rio Grande do Sul. Chefe da Unidadede Nefrologia Pediátrica do Hospital de Clínicas de Porto Alegre, RS
OSCAR FERNANDO PAVÃO DOS SANTOSProfessor Associado da Disciplina de Nefrologia da Escola Paulista
de Medicina — Universidade Federal de São Paulo
PAULO HENRIQUE FRAXINONefrologista da Santa Casa de Irati. Diretor Médico da Clínica Re-nal Irati. Ex-Residente do Serviço de Nefrologia do Hospital Uni-versitário Evangélico de Curitiba, PR
PAULO SCHIAVOM DUARTEEspecialista em Medicina Nuclear
REINALDO MARTINELLIProfessor Titular, Departamento de Medicina, Faculdade de Medi-cina da Universidade Federal da Bahia
RICARDO AUGUSTO FARIA Mestre em Nefrologia. Médico Nefrologista do Ministério da Saú-de. Médico da Kidney Assistance/CDR Serviços Hospitalares
ROBERTO C. MANFROProfessor Adjunto do Departamento de Medicina Interna, Facul-dade de Medicina, Universidade Federal do Rio Grande do Sul. Mé-dico Assistente do Serviço de Nefrologia do Hospital de Clínicas dePorto Alegre, RS
ROBERTO FLÁVIO DA SILVA PECOITS FILHOProfessor Adjunto de Clínica Médica da Pontifícia Universidade Cató-lica do Paraná. Doutor em Nefrologia pela Universidade de São Paulo.Pesquisador Associado ao Karolinska Institutet, Estocolmo, Suécia
ROBERTO ZATZProfessor Associado de Disciplina de Nefrologia, Departamento deClínica Médica, Faculdade de Medicina da USP
RODRIGO VIEIRA OZELAMEEspecialista em Radiologia Pediátrica
RONALDO ROBERTO BÉRGAMOProfessor Titular da Disciplina de Nefrologia da Faculdade de Me-dicina do ABC
RUI TOLEDO BARROSProfessor Assistente da Disciplina de Nefrologia da Faculdade de Me-dicina da Universidade de São Paulo. Coordenador do Grupo de
Nefrologia Clínica do Serviço de Nefrologia do Hospital das Clíni-cas da FMUSP
SANDRA BALEEIRO ABRAHÃODoutora em Nefrologia pela Faculdade de Medicina da Universidadede São Paulo. Médica do Serviço-Disciplina de Nefrologia do HCFMUSP
SANDRA PINHO SILVEIROProfessora Adjunta do Departamento de Medicina Interna da Fa-culdade de Medicina da Universidade Federal do Rio Grande do Sul
SEBASTIÃO O. L. DE CARVALHORadiologista do Hospital de Clínicas da Universidade Federal do Paraná
SERGIO ANTONIO DRAIBEProfessor Associado da Disciplina de Nefrologia da Escola Paulista
de Medicina — Universidade Federal de São Paulo
SÉRGIO AUGUSTO DE MUNHOZ PITAKI Médico Especialista em Radiologia e Diagnóstico por Imagem — Colé- gio Brasileiro de Radiologia (CBR). Membro da Sociedade Norte-Ameri-cana de Radiologia (RSNA). Diretor da Clínica Alphasonic-Curitiba, PR

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 9/121
Colaboradores ix
TAÍS TINNUCCIDoutora em Nefrologia pela Faculdade de Medicina da Universi-dade de São Paulo. Docente pela Escola de Educação Física e Espor-
tes da Universidade de São PauloTELMA SAKUNO
Especialista em Radiologia Pediátrica. Radiologista do Hospital In- fantil Joana de Gusmão e do Hospital Universitário da Faculdade de Medicina da Universidade Federal de Santa Catarina
VANDA JORGETTIDoutora em Nefrologia pela USP. Médica Assistente do Serviço de Nefro-logia do Hospital das Clínicas da Universidade de São Paulo. Responsável
pelo Laboratório de Osteodistrofia Renal e Doenças Ósseas Metabólicas
VANILDO JOSÉ OZELAMEDoutor em Radiologia. Professor Titular de Radiologia da Uni-versidade Federal de Santa Catarina. Especialista em Radiologia Pe-
diátricaVIKTÓRIA WORONIK
Professora Assistente da Disciplina de Nefrologia da Faculdade de Medicina da Universidade de São Paulo. Responsável pelo Ambu-latório de Glomerulopatias do Serviço de Nefrologia do Hospital dasClínicas da FMUSP
VITOR AUGUSTO SOARES ( IN MEMORIAM )Professor Livre-Docente em Nefrologia e Professor Emérito da Fa-culdade de Medicina de Botucatu — UNESP.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 10/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 11/121
A meus pais,
Edith e Carlos, uma homenagem de gratidão.
A minha esposa
Marila e a meus filhos Leonardo e Cristian, pelo incentivo, compreensão e apoio.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 12/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 13/121
APRESENTAÇÃO DA QUARTA EDIÇÃO
No ano de 1980, o Prof. Miguel Carlos Riella lançou, com osucesso esperado, o seu livro de Nefrologia, o qual, de modoprogressivo, foi se tornando o livro de informação nefrológica queconsidero o mais completo entre os textos usados nas escolasmédicas de nosso País, além de ser, acredito, o mais informativoem língua portuguesa.
Engano pensar que a reedição de um livro como este, que tenhoa satisfação especial de apresentar, seja um trabalho relativamentesimples de realizar, pelo fato de grande parte já ter sido feita.
A nova edição de um livro deste porte implica a necessidadede novos capítulos, cuja escolha precisa ser criteriosa, adicionandoinformações necessárias e complementares. No caso atual, aos 51capítulos já existentes, foram adicionados mais três, cobrindo im-portantes áreas. Além de investigação através de imagem do apa-relho urinário, foi incluída a imagem renal por ressonância mag-nética, sendo discutidas suas indicações, vantagens e limites.
Pela importância do tema, foi acrescentado um capítulo sobrea hipertensão arterial na mulher, assim como um capítulo espe-cífico e destacado sobre diálise peritoneal, além de métodoshemodialíticos contínuos para tratamento da insuficiência renalaguda.
Tudo isso vem acrescentar conhecimentos práticos e objetivosna conduta diagnóstica e terapêutica no cuidado com o nefropata.
Há necessidade de aprimorar as informações sobre patogeniae fisiopatologia das nefropatias, sobretudo pelo volume de no-
vas informações de ordem imunológica, assim como pelas novi-
dades trazidas por avanços da biologia molecular. Ao lado dis-so, a necessidade cada vez maior de termos evidências essenci-ais que fundamentem o entendimento do processo patológico. Anefrologia é uma rica área nesse sentido.
Um livro moderno deve facilitar o leitor, acompanhando osavanços na área. Principalmente deve ajudar o professor no pre-paro de suas aulas e conferências, bem como na revisão de tópi-cos que se atualizam continuamente. Nesse particular, a apresen-tação moderna da formatação dos capítulos e um site na Internet
no final de cada um deles, além de todo o conteúdo do livro apre-sentado em um CD, vão ajudar o manuseio prático e a realizaçãode revisões e atualizações mais freqüentes e mais fáceis de seremfeitas e aproveitadas.
O acompanhamento de um livro como este, através de edi-ções sucessivas e aprofundadas e cuidadosamente aprimoradas,serve também para definir e apresentar o vigor científico dessaárea motivadora e estimulante sobre que se detém o trabalhoapresentado.
De parabéns o Prof. Riella e seus colaboradores; seu livro deNefrologia melhora a cada nova edição, demonstrando a serie-dade com que realiza o seu trabalho. A Nefrologia brasileira seenriquece com mais esta contribuição.
HEONIR ROCHAFaculdade de Medicina
Universidade Federal da Bahia

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 14/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 15/121
PREFÁCIO DA QUARTA EDIÇÃO
Agora já se passaram 23 anos desde que escrevemos o prefá-cio da primeira edição, publicada em setembro de 1980. Princí-
pios de Nefrologia e Distúrbios Hidroeletrolíticos é o livro deNefrologia adotado pela maioria das escolas de medicina do Bra-sil. Em função disso, a sua editoração tornou-se cada vez maiscriteriosa. Algumas inovações foram introduzidas nesta edição:esta é a primeira vez que o texto é impresso em duas cores; pon-tos-chave destacam tópicos importantes a serem lembrados peloleitor, e, para complementar a informação, são fornecidos diver-
sos endereços eletrônicos que poderão auxiliar sobremaneira oleitor. E, como se isso não bastasse, o livro é apresentado tambémsob a forma de CD, que permite uma pesquisa rápida e uma vi-são abrangente das figuras e fotos coloridas. Na era da medicinabaseada em evidências, procurou-se referenciar extensamente o tex-
to. Capítulos foram atualizados e outros desdobrados, como mé-todos de depuração extra-renal: hemodiálise, diálise peritoneale métodos contínuos são agora apresentados separadamente.Além disso, expandiu-se a abordagem aos métodos de imagemdo aparelho urinário e introduziu-se um novo capítulo: hiperten-são na mulher.
Esperamos novamente que o livro satisfaça aos nossos estu-dantes de medicina e jovens nefrologistas. Reiteramos o com-promisso de mantê-lo atualizado. Agradecemos mais uma vez
aos nossos colaboradores, dignos representantes da Nefrologia brasileira.
MIGUEL CARLOS RIELLACuritiba, março de 2003

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 16/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 17/121
APRESENTAÇÃO DA TERCEIRA EDIÇÃO
Dr. Miguel Carlos Riella tomou a decisão muito acertada delançar, no mercado de livros científicos, a terceira edição de Prin-cípios de Nefrologia e Distúrbios Hidroeletrolíticos.
O prefácio da primeira edição (1980) foi escrito pelo Dr.Belding H. Scribner, com quem Riella fez seu fellowship. Scribner,a meu ver, deveria já ter recebido o prêmio Nobel de Medicinapelos seus trabalhos e pesquisas no tratamento do renal crônico.
A segunda edição (1988) foi prefaciada pelo Prof. OswaldoLuiz Ramos, Professor de Nefrologia da Escola Paulista de Me-dicina, local onde Riella defendeu seu doutorado.
Esta edição, muito mais ampla que as anteriores, com 51 capí-tulos, é prefaciada por mim, cujo único mérito reside no fato deser um amigo e admirador do Dr. Riella de longa data.
É preciso ressaltar a importância de um livro deste padrão den-tro do contexto do ensino de nefrologia no Brasil. Os graduandosde medicina, os residentes e pós-graduandos de Nefrologia encon-traram nele um instrumento muito adequado para seu aprendiza-
do. A meu ver, é o único livro nacional que cumpre esta finalida-de. É extremamente abrangente, analisando não só as alteraçõeseletrolíticas do paciente nefropata, como a fisiopatologia, a clínicanefrológica e a terapêutica.
A medicina, da qual a Nefrologia é um capítulo substancial, é umaciência profundamente dinâmica, o que determinou esta nova edi-ção de 1996 e que permite prever que novas edições se sucederão.
Professor Miguel Riella é um nefrologista extremamente mo-tivado em ensino, pioneiro no Brasil em algumas técnicasnefrológicas, e lança a terceira edição do seu livro quando ocupamuito meritoriamente o cargo de Presidente da Sociedade Brasi-leira de Nefrologia.
Parabéns, meu amigo Riella, por ter contribuído mais uma vezpara o progresso da nossa especialidade.
EMIL SABBAGAFaculdade de Medicina da Universidade de São Paulo

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 18/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 19/121
PREFÁCIO DA TERCEIRA EDIÇÃO
A primeira edição de Princípios de Nefrologia e Distúrbios Hi-droeletrolíticos foi apresentada em 1980 com 30 capítulos e ten-do como alvo principal os nossos estudantes de medicina. A se-gunda edição é de 1988 com 35 capítulos, consolidando-se comolivro-texto nas escolas de medicina e bastante útil também paraos jovens médicos residentes e nefrologistas. Esta terceira edi-ção, de 1996, é ampliada para 51 capítulos, procurando tradu-zir o avanço da Nefrologia, tornando-se mais atualizada para o
jovem nefrologista, sem contudo perder as características dos
temas básicos para o estudante de medicina. A seção de distúr- bios hidroeletrolíticos é mantida e de certa forma tem umaconotação histórica, já que é responsável pela minha escolha daNefrologia como especialização clínica. Durante todo o cursode medicina preparava-me para ser um cirurgião. A noção deque seria importante para o cirurgião o domínio da reposiçãohidroeletrolítica, do plano parenteral, levou-me mais de per-to a conhecer e encantar-me com a Nefrologia. O interesse de
Belding H. Scribner, meu mentor na Universidade de Washing-ton em 1973, consolidou a minha formação nesta área, que hojefaz parte integrante de Princípios de Nefrologia e Distúrbios Hi-droeletrolíticos .
Se me coube o mérito pela iniciativa da primeira edição, hojeem dia o partilho com a Nefrologia brasileira. Tenho ao longo dosanos abdicado de escrever certos capítulos para que novos cola-
boradores participem, estimulando-os a escrever, expor suas idéi-as e trabalho e, de certa forma, procurando traduzir a pujança, o
crescimento e a valorização da nossa especialidade. O lançamentodesta terceira edição, estando eu Presidente da Sociedade Brasi-leira de Nefrologia, muito me honra e de certa forma coroa a mi-nha trajetória na Nefrologia brasileira. Aos amigos e colaborado-res, o meu sincero agradecimento.
MIGUEL CARLOS RIELLACuritiba, setembro de 1996

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 20/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 21/121
APRESENTAÇÃO DA SEGUNDA EDIÇÃO
Temos enorme prazer em apresentar a 2.ª edição de Princípios deNefrologia e Distúrbios Hidroeletrolíticos, de autoria do Professor Mi-guel Carlos Riella. A 1.ª edição cumpriu integralmente sua finalida-de, cobrindo, de maneira direta e precisa, os principais conhecimen-tos indispensáveis à formação de um nefrologista. O livro se mostrouextremamente útil a estudantes, residentes e pós-graduandos daárea. Passados sete anos, sentiu o autor que, para manter o mesmopadrão de qualidade, era necessário haver uma atualização. Assim,
surge esta nova edição com pelo menos quatro novos capítulos e umareal atualização dos restantes, conservando, entretanto, as caracte-rísticas de precisão e concisão que nortearam a edição antecedente.
Parabéns ao autor e à Nefrologia brasileira por mais esta demons-tração de pujança e competência.
OSWALDO LUIZ RAMOSEscola Paulista de Medicina

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 22/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 23/121
PREFÁCIO DA SEGUNDA EDIÇÃO
Após sete anos do lançamento da primeira edição deste li-vro, sinto-me gratificado pela sua aceitação entre os estudan-tes e jovens médicos das escolas de Medicina deste País. Pas-sou-me, no entanto, despercebida, na época, a responsabilida-de que assumia perante a comunidade científica de manteresta obra atualizada e, portanto, útil a gerações vindouras.Isto significa, na prática, que mal sai uma nova edição come-çamos a nos preocupar com a seguinte. É um trabalho e umaresponsabilidade contínua. Mais uma vez coube-me o privilé-
gio e a honra de coordenar esta edição. Dela participam gru-pos nefrológicos os mais representativos do País, refletindo,de certa forma, o aprimoramento e progresso da Nefrologia
brasileira.
Nesta 2.ª edição, além de atualização de cada capítulo, introdu-zimos outros novos: Fisiopatologia do Edema; Modernos MétodosDiagnósticos: Radioisótopos, Ultra-sonografia e Tomografia Computa-dorizada; Métodos de Depuração Extra-renal; Síndrome Nefrítica e En-
fermidades Túbulo-intersticiais. Acredito que esta renovação apri-mora e enriquece a nova edição.
O sentimento de que este texto poderá contribuir para a for-mação de nossos médicos e jovens nefrologistas deixa-me orgu-lhoso e gratificado, sentimento este que partilho com todos aque-
les que aqui colaboram.
MIGUEL CARLOS RIELLACuritiba, 12 de setembro de 1987

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 24/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 25/121
APRESENTAÇÃO DA PRIMEIRA EDIÇÃO
Este único volume contém uma quantidade quase inacreditávelde informações úteis e práticas. Abrangendo ambas as áreas de
balanço hidroeletrolítico e nutrição parenteral, o Dr. Riella e seuscolaboradores produziram um livro que deve provar ser muito útil.Além disto, os vários capítulos de Nefrologia abordam este cam-po de maneira a satisfazer as necessidades de qualquer clínico geralou internista, assim como de nefrologistas e urologistas.
Através de uma cuidadosa seleção de tópicos, o Dr. Riella eseus co-autores criaram uma fonte única de informações. Agra-
da-me o fato de que alguns conhecimentos que o Dr. Riella apren-deu em Seattle, durante o seu Research Fellowship, fazem partedeste novo e importante texto.
BELDING H. SCRIBNER, M. D.Professor de Medicina Interna. Chefe, Divisão de Nefrologia,
University of Washington, Seattle, USA.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 26/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 27/121
PREFÁCIO DA PRIMEIRA EDIÇÃO
A idéia inicial de organizar este texto de Nefrologia surgiudurante a minha permanência na Universidade de Washingtonem Seattle. Impressionou-me sobremaneira a preocupação doseducadores em orientar os estudantes de Medicina e proporcio-nar-lhes textos atualizados da matéria a ser ministrada. O ensinoera encarado com seriedade e responsabilidade. As anotações emclasse, que tanto distraem o estudante, tornavam-se desneces-sárias. Desapareciam as aulas magistrais e surgia um proveitosodiálogo professor-alunos.
Esta experiência inicial foi marcante e constituiu-se, sem dú-vida, numa fonte geradora de energia e estímulos para a organi-zação deste livro, cuja finalidade primordial é a de proporcionaraos nossos estudantes de Medicina um texto de Nefrologia sim-ples, prático e atualizado. É óbvio que, dada a profundidade comque alguns temas foram abordados, poderá ser útil a residentes,clínicos gerais e nefrologistas. Além dos temas básicos deNefrologia, abordamos neste volume a Terapia Parenteral, inici-ando com os distúrbios hidroeletrolíticos, que sem dúvida enri-quecem o texto e suprem um vazio há muito sentido pelos estu-dantes nos textos nefrológicos, e finalizando com noções de Nu-trição Parenteral, que, embora atraia os mais diversos especialis-tas (clínicos, cirurgiões, anestesiologistas etc.), pode ser de extre-ma utilidade para o nefrologista, freqüentemente envolvido nocuidado do paciente gravemente enfermo.
Apenas aqueles que se defrontaram com a árdua tarefa de or-
ganizar e editar um livro sabem do esforço e dificuldades envol-vidos. A tentativa de agilizar a publicação e, por conseguinte,evitar a disseminação de informação ultrapassada traz seus ris-
cos. A participação de vários co-autores compromete a homoge-neidade do texto, por mais bem familiarizados que estejam como espírito da obra. Estes fatos, aliados a nossa inexperiência nocampo editorial, trarão à tona, sem dúvida, erros e falhas. Noentanto, conscientes das deficiências, antecipamos o nosso espí-rito receptivo a críticas e sugestões, procurando aprimorar estetrabalho inicial. Colaboram, neste livro, nefrologistas e pesqui-sadores de diversas escolas médicas do Brasil, cada um trazendoo melhor da sua experiência pessoal. Sem eles, esta obra não se-
ria uma realidade. Além do mais, muitos foram aqueles que meapoiaram e incentivaram na organização deste texto. Inicialmenteo meu agradecimento e admiração aos mestres que serviram deestímulo: Belding H. Scribner, Gary E. Striker, Robert Hickman,Leonard J. Quadracci e Ralph Cutler, da Universidade de Wa-shington, Seattle. A minha gratidão aos meus colegas de traba-lho, João Loewen e Luiz Sallim Emed, que arcaram com grandeparte das minhas obrigações no Serviço de Nefrologia, proporci-onando-me o tempo e a tranqüilidade que um trabalho destanatureza requer. Agradeço ainda aos jovens médicos Marco PoloRauth e Angel A. J. Mate que, com seus trabalhos artísticos, ilus-tram alguns capítulos deste livro; à minha secretária Cecília HalluPalma, pela sua paciência e incansável dedicação no preparo dosmanuscritos; ao Dr. Antonino S. Rocha, pela análise e comentá-rio de alguns capítulos de fisiologia renal. E, por fim, mas nãomenos importante, o agradecimento a minha esposa Marila, pela
compreensão e apoio.MIGUEL CARLOS RIELLA

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 28/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 29/121
26 Nefropatia do Refluxo, 50727 Doenças Vasculares dos Rins, 51928 Nefropatia e Gestação, 53729 Hipertensão na Mulher, 54630 Tubulopatias Hereditárias, 55731 Doenças Císticas Renais, 58032 Nefropatia Diabética, 59733 Nefrolitíase, 609
34 Uropatia Obstrutiva, 62035 Tumores Renais, 631
IV. FISIOPATOLOGIA DAS NEFROPATIAS
36 Insuficiência Renal Crônica (IRC), 64937 Insuficiência Renal Crônica: Fisiopatologia da Uremia, 66138 Conseqüências Hematológicas da Uremia, 69139 O Sistema Nervoso na Insuficiência Renal, 70540 Fisiopatologia, Clínica e Tratamento da
Osteodistrofia Renal, 71741 Hipertensão Arterial Primária, 73042 Hipertensão Renovascular, 75743 Hipertensão Arterial e Doença Renal Parenquimatosa, 769
V. MANEJO CLÍNICO DO PACIENTECOM INSUFICIÊNCIA RENAL
44 Diuréticos. Mecanismo de Ação e Uso Clínico, 77545 Drogas Anti-hipertensivas, 78746 Uso de Medicamentos em Insuficiência Renal, 79947 Manejo e Terapia Nutricional do Urêmico, 84348 Nutrição Parenteral Intradialítica, 86149 Hemodiálise, 86950 Métodos Hemodialíticos Contínuos para Tratamento da
Insuficiência Renal Aguda, 90851 Diálise Peritoneal, 91952 Imunologia do Transplante Renal, 929
53 Manejo Clínico do Transplante Renal, 94454 Infecções em Transplante Renal, 974
Índice Alfabético, 988
CONTEÚDO
I. ESTRUTURA E FUNÇÃO RENAL
1 Anatomia Renal, 12 Circulação Renal, 203 Filtração Glomerular, 304 Função Tubular, 375 Mecanismos de Acidificação Urinária, 496 Mecanismo de Concentração e de Diluição Urinária, 58
7 Peptídeos Vasoativos e o Rim, 69
II. DISTÚRBIOS HIDROELETROLÍTICOS
8 Compartimentos Líquidos do Organismo, 909 Metabolismo da Água, 100
10 Metabolismo do Sódio e Fisiopatologia do Edema, 13211 Metabolismo Ácido-Básico, 16212 Metabolismo do Potássio, 18913 Metabolismo do Cálcio, Fósforo e Magnésio, 21314 Metabolismo do Ácido Úrico, 23815 Terapia Parenteral. Reposição Hidroeletrolítica, 254
III. PATOGENIA DAS NEFROPATIAS
16 Avaliação Clínica e Laboratorial da Função Renal, 26717 Investigação por Imagem do Aparelho Urinário no
Adulto, 294I. Introdução aos Métodos de Diagnóstico por Imagem
da Doença Renal, 294II. Radiologia do Aparelho Urinário no Adulto, 295
18 Ultra-sonografia Renal e Imagem Renal por RessonânciaMagnética, 315I. Ultra-sonografia Renal, 315
II. Imagem Renal por Ressonância Magnética, 32619 Investigação por Imagem do Trato Urinário na Criança, 33620 Radioisótopos em Nefrourologia, 372
21 Insuficiência Renal Aguda, 38822 Glomerulonefrites Primárias, 40223 Glomerulopatias Secundárias, 42424 Nefropatia Tóxica e Tubulointersticial, 45025 Infecção do Trato Urinário, 490

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 30/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 31/121
PRINCÍPIOS DE NEFROLOGIA EDISTÚRBIOS HIDROELETROLÍTICOS

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 32/121

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 33/121
Capítulo
1Anatomia Renal
Leonardo Vidal Riella, Luiz Antonio Ribeiro de Moura e Miguel Carlos Riella
MACROSCOPIA
VASCULARIZAÇÃO
CIRCULAÇÃO LINFÁTICA
INERVAÇÃO
EMBRIOLOGIA
Anomalias do desenvolvimento
O NÉFRON
Glomérulo
Células endoteliais
Células mesangiais
Células epiteliais viscerais
Células epiteliais parietais
Aparelho justaglomerular
Células peripolares
Túbulo proximal
Alça de Henle
Túbulo distal
Ducto coletor
INTERSTÍCIO RENAL
REFERÊNCIAS BIBLIOGRÁFICAS
ENDEREÇOS RELEVANTES NA INTERNET
MACROSCOPIAOs rins, em número de dois, são órgãos que lembram a
forma de um grão de feijão, de coloração marrom-averme-lhada, situados no espaço retroperitoneal, um de cada ladoda coluna vertebral, de tal forma que seu eixo longitudi-nal corre paralelamente ao músculo psoas maior.
Na posição ortostática, sua margem superior encontra-se ao nível da primeira vértebra lombar e a inferior, daquarta vértebra lombar. Em decúbito dorsal, as margenssuperior e inferior dos rins elevam-se ao nível do bordosuperior da 12-ª vértebra torácica e da terceira vértebra lom- bar, respectivamente.1 Com a respiração os rins podem des-locar-se cerca de 1,9 cm, chegando a 4,1 cm na inspiração
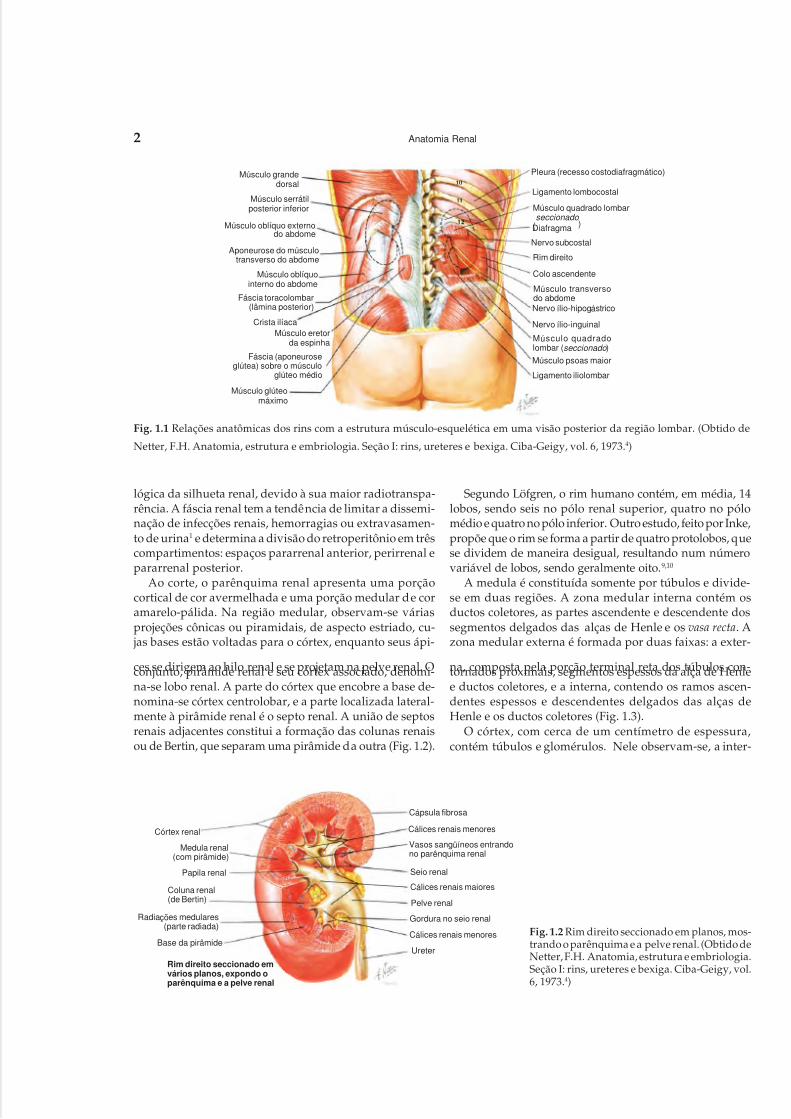
profunda. Normalmente, o rim direito é um centímetro me-nor e encontra-se ligeiramente mais caudal em relação aoesquerdo (Fig. 1.1).
O rim de um indivíduo adulto mede de 11 a 13 cm decomprimento, 5 a 7,5 cm de largura e 2,5 a 3 cm de espessu-ra, pesando entre 125 e 170 gramas, no homem, e 115 e 155gramas, na mulher. Com o envelhecimento, há uma dimi-nuição do peso renal.6 Em recém-nascidos este peso varia
de 13 a 44 gramas.7
A variação do tamanho e do peso dosrins na população demonstrou estar mais relacionada coma superfície corporal do indivíduo, não sendo influenciadapor sexo, idade ou raça, quando se leva em consideração otipo de constituição corporal. Outros estudos demonstraramtambém que o nível de hidratação do organismo e a pres-são arterial provocam variações no tamanho do rim.8
Na parte medial côncava de cada rim, localiza-se o hilorenal, local onde se encontram a artéria e a veia renal, vasoslinfáticos, plexos nervosos e o ureter, que se expande den-tro do seio renal, formando a pelve. O rim é envolvido emtoda sua superfície por membrana fibroelástica muito finae brilhante, denominada cápsula renal. Esta adere à pelve eaos vasos sanguíneos na região do hilo. No rim sadio, con-segue-se destacar facilmente a cápsula renal do restante do
órgão, sendo que o mesmo não acontece no rim doente.Ao redor dos rins, no espaço retroperitoneal, tem-se umacondensação de tecido conjuntivo, que representa a fásciade Gerota ou fáscia renal. Ela divide-se em fáscias renaisanterior e posterior, envolvendo um tecido adiposo, deno-minado gordura perirrenal, que contorna o rim e a glân-dula adrenal de cada lado, constituindo o espaço perirre-nal. Essa gordura é a responsável pela visualização radio-
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 34/121
2 Anatomia Renal
lógica da silhueta renal, devido à sua maior radiotranspa-rência. A fáscia renal tem a tendência de limitar a dissemi-nação de infecções renais, hemorragias ou extravasamen-to de urina1 e determina a divisão do retroperitônio em trêscompartimentos: espaços pararrenal anterior, perirrenal epararrenal posterior.
Ao corte, o parênquima renal apresenta uma porçãocortical de cor avermelhada e uma porção medular de coramarelo-pálida. Na região medular, observam-se váriasprojeções cônicas ou piramidais, de aspecto estriado, cu- jas bases estão voltadas para o córtex, enquanto seus ápi-
ces se dirigem ao hilo renal e se projetam na pelve renal. Oconjunto, pirâmide renal e seu córtex associado, denomi-na-se lobo renal. A parte do córtex que encobre a base de-nomina-se córtex centrolobar, e a parte localizada lateral-mente à pirâmide renal é o septo renal. A união de septosrenais adjacentes constitui a formação das colunas renaisou de Bertin, que separam uma pirâmide da outra (Fig. 1.2).
Segundo Löfgren, o rim humano contém, em média, 14lobos, sendo seis no pólo renal superior, quatro no pólomédio e quatro no pólo inferior. Outro estudo, feito por Inke,propõe que o rim se forma a partir de quatro protolobos, quese dividem de maneira desigual, resultando num númerovariável de lobos, sendo geralmente oito.9,10
A medula é constituída somente por túbulos e divide-se em duas regiões. A zona medular interna contém osductos coletores, as partes ascendente e descendente dossegmentos delgados das alças de Henle e os vasa recta. Azona medular externa é formada por duas faixas: a exter-
na, composta pela porção terminal reta dos túbulos con-tornados proximais, segmentos espessos da alça de Henlee ductos coletores, e a interna, contendo os ramos ascen-dentes espessos e descendentes delgados das alças deHenle e os ductos coletores (Fig. 1.3).
O córtex, com cerca de um centímetro de espessura,contém túbulos e glomérulos. Nele observam-se, a inter-
Músculo grandedorsal
Músculo serrátilposterior inferior
Músculo oblíquo externodo abdome
Aponeurose do músculotransverso do abdome
Músculo oblíquointerno do abdome
Fáscia toracolombar(lâmina posterior)
Crista ilíacaMúsculo eretor
da espinha
Fáscia (aponeuroseglútea) sobre o músculo
glúteo médio
Músculo glúteomáximo
Pleura (recesso costodiafragmático)
Ligamento lombocostal
Músculo quadrado lombar
(seccionado
)Diafragma
Nervo subcostal
Rim direito
Colo ascendente
Músculo transversodo abdomeNervo ílio-hipogástrico
Nervo ílio-inguinal
Músculo quadradolombar (seccionado )
Músculo psoas maior
Ligamento iliolombar
Fig. 1.1 Relações anatômicas dos rins com a estrutura músculo-esquelética em uma visão posterior da região lombar. (Obtido de
Netter, F.H. Anatomia, estrutura e embriologia. Seção I: rins, ureteres e bexiga. Ciba-Geigy, vol. 6, 1973.4)
Córtex renal
Medula renal(com pirâmide)
Papila renal
Coluna renal(de Bertin)
Radiações medulares(parte radiada)
Base da pirâmide
Rim direito seccionado emvários planos, expondo oparênquima e a pelve renal
Cápsula fibrosa
Cálices renais menores
Vasos sangüíneos entrandono parênquima renal
Seio renal
Cálices renais maiores
Pelve renal
Gordura no seio renal
Cálices renais menores
Ureter
Fig. 1.2 Rim direito seccionado em planos, mos-trando o parênquima e a pelve renal. (Obtido deNetter, F.H. Anatomia, estrutura e embriologia.Seção I: rins, ureteres e bexiga. Ciba-Geigy, vol.6, 1973.4)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 35/121
capítulo 1 3
valos regulares, estriações denominadas raios medulares.Estes raios originam-se das bases das pirâmides e contêmtúbulos coletores, ramos ascendentes da alça de Henle e asporções retas terminais dos túbulos contornados proxi-mais, cuja disposição em paralelo é responsável pelo as-pecto estriado das pirâmides (Fig. 1.2).
Cada raio medular ocupa o centro de um lóbulo renal, uma
pequena e cilíndrica área de córtex, delimitada por artériasinterlobulares. O termo lóbulo renal, apesar de descrito, nãoé muito empregado, uma vez que não se consegue definiruma importância anatomofuncional para o mesmo.
Alguns dos túbulos se unem para formar ductos coleto-res. Os ductos coletores maiores, ou ductos de Bellini, abrem-se no ápice da pirâmide, na papila renal, região que contéma área crivosa com cerca de 10 a 25 perfurações. A urina, quedaí drena, cai num receptáculo chamado cálice menor.
Até a 28-ª semana de gestação existem 14 cálices, de talmaneira que cada cálice se associa apenas a uma papila.Após este período, dá-se início a um processo de fusãolobar, que pode prolongar-se até após o nascimento, e quedetermina a diminuição do número de cálices e de papilasrenais. O grau de fusão calicial é maior que o de fusão pa-
pilar, o que determina o aparecimento de cálices compos-tos, ou seja, cálices que recebem mais que uma papila.Aparecem também papilas compostas, que drenam maisde um lobo. Este processo mostra-se mais evidente nospólos superior e inferior do rim, sendo que na região cen-tral predominam os cálices e papilas simples.9
As papilas simples possuem extremidades convexas,enquanto as compostas, dependendo do número de fusões,
apresentam formato circular, rígido, achatado, ou até mes-mo côncavo, predispondo ao surgimento do fenômeno dorefluxo intra-renal, relacionado na etiologia da pielonefri-
te crônica e da nefropatia do refluxo. Seqüelas de pielone-frite são mais observadas nos pólos renais, locais de maiorocorrência de papilas compostas.11
A porção do cálice menor que se projeta para cima, aoredor da papila, é chamada de fórnix e é importante por-que os primeiros sinais de infecção ou obstrução ocorrema este nível (v. Fig. 1.2).
Os cálices menores unem-se para formar os cálices mai-ores, que são em número de dois a quatro. Comumente,apenas três cálices são vistos no urograma excretor (v. Cap.17). Os cálices maiores, por sua vez, unem-se para formarum funil curvo, chamado pelve renal, que se curva no sen-tido medial e caudal, para tornar-se o ureter a um pontodenominado junção ureteropélvica.
Pontos-chave:
• Órgão retroperitoneal localizado entre asvértebras L1 e L4, apresentaaproximadamente 12 cm de comprimento.Seu peso médio é de 150 g. A diminuição dotamanho renal está principalmenteassociada à nefropatia crônica
• Macroscopicamente, pode ser dividido emcórtex e medula. O córtex se constitui deglomérulos, túbulos contorcidos proximaise distais; já a medula contém as alças deHenle e os túbulos coletores, os quais se
abrem nas papilas dos cálices menores• A gordura perirrenal, localizada entre o rim
e a fáscia renal, é a responsável pelavisualização radiológica da silhueta renal
• Cálculos renais obstruem os ureteresprincipalmente em três regiões: junçãoureteropiélica, porção anterior à bifurcaçãoda artéria ilíaca comum e junçãoureterovesical
• Na cirurgia de histerectomia, especialatenção deve ser tomada na hora de ligar aartéria uterina, devido à sua relação íntimacom o ureter, o qual passa posteriormente
O ureter é um tubo muscular que se estende da pelverenal à bexiga urinária. Localiza-se no compartimento re-troperitoneal e descende anteriormente ao músculo psoas.Em seu trajeto apresenta algumas relações importantescom outras estruturas: é cruzado anteriormente pelos va-sos gonadais; passa anteriormente à bifurcação da artériailíaca comum na entrada da pelve, e se situa posteriormente
Fig. 1.3 Relações entre os vários segmentos do néfron e o córtex emedula renal. (Obtido de Netter, F.H. Anatomia, estrutura e em-
briologia. Seção I: rins, ureteres e bexiga. Ciba-Geigy, vol. 6, 1973.4)
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 36/121
4 Anatomia Renal
ao ducto deferente no homem e posteriormente à artériarenal na mulher. Esta última relação é especialmente im-portante nas cirurgias de histerectomia, em que o ureter
pode ser inadvertidamente ligado ou clampeado junto coma artéria uterina. O ureter apresenta três segmentos ondea impactação de um cálculo é mais freqüente: na junçãoureteropiélica, na porção ureteral anterior à bifurcação dasartérias ilíacas comuns e na junção ureterovesical.
VASCULARIZAÇÃO
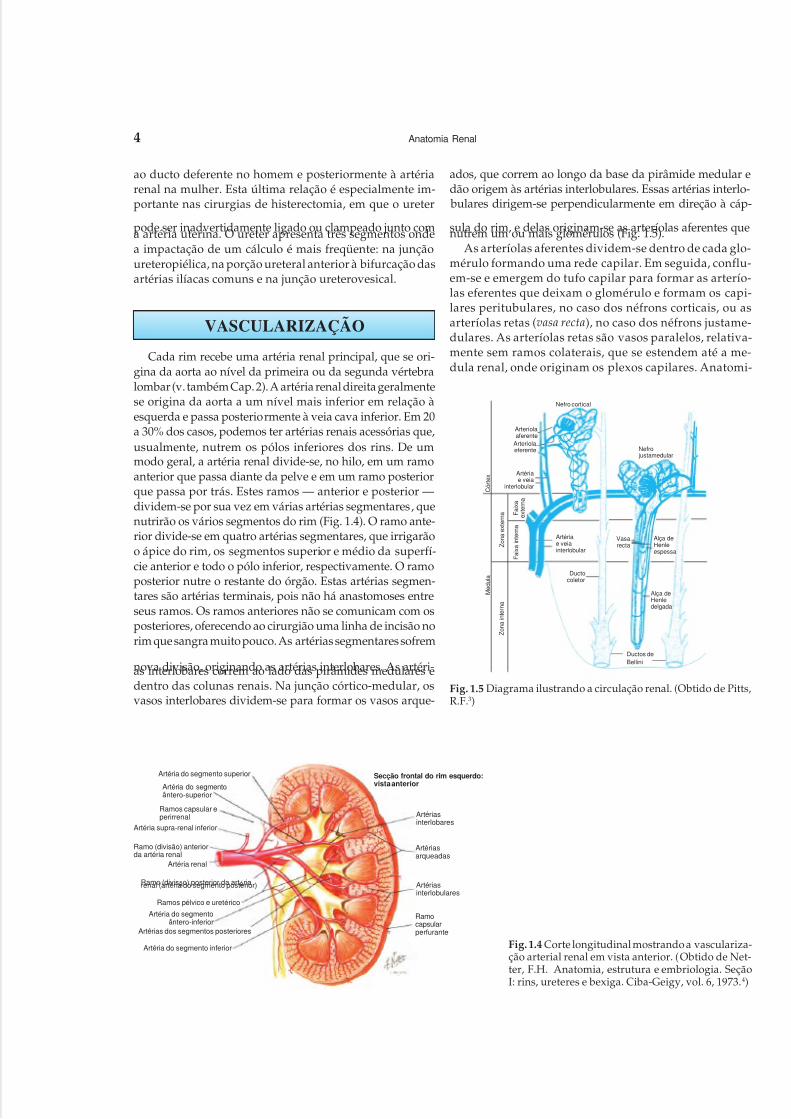
Cada rim recebe uma artéria renal principal, que se ori-gina da aorta ao nível da primeira ou da segunda vértebralombar (v. também Cap. 2). A artéria renal direita geralmentese origina da aorta a um nível mais inferior em relação àesquerda e passa posteriormente à veia cava inferior. Em 20a 30% dos casos, podemos ter artérias renais acessórias que,usualmente, nutrem os pólos inferiores dos rins. De ummodo geral, a artéria renal divide-se, no hilo, em um ramoanterior que passa diante da pelve e em um ramo posteriorque passa por trás. Estes ramos — anterior e posterior —dividem-se por sua vez em várias artérias segmentares, quenutrirão os vários segmentos do rim (Fig. 1.4). O ramo ante-rior divide-se em quatro artérias segmentares, que irrigarãoo ápice do rim, os segmentos superior e médio da superfí-cie anterior e todo o pólo inferior, respectivamente. O ramoposterior nutre o restante do órgão. Estas artérias segmen-tares são artérias terminais, pois não há anastomoses entreseus ramos. Os ramos anteriores não se comunicam com osposteriores, oferecendo ao cirurgião uma linha de incisão norim que sangra muito pouco. As artérias segmentares sofrem
nova divisão, originando as artérias interlobares. As artéri-as interlobares correm ao lado das pirâmides medulares edentro das colunas renais. Na junção córtico-medular, osvasos interlobares dividem-se para formar os vasos arque-
ados, que correm ao longo da base da pirâmide medular edão origem às artérias interlobulares. Essas artérias interlo- bulares dirigem-se perpendicularmente em direção à cáp-
sula do rim, e delas originam-se as arteríolas aferentes quenutrem um ou mais glomérulos (Fig. 1.5).As arteríolas aferentes dividem-se dentro de cada glo-
mérulo formando uma rede capilar. Em seguida, conflu-em-se e emergem do tufo capilar para formar as arterío-las eferentes que deixam o glomérulo e formam os capi-lares peritubulares, no caso dos néfrons corticais, ou asarteríolas retas (vasa recta), no caso dos néfrons justame-dulares. As arteríolas retas são vasos paralelos, relativa-mente sem ramos colaterais, que se estendem até a me-dula renal, onde originam os plexos capilares. Anatomi-
Fig. 1.4 Corte longitudinal mostrando a vasculariza-ção arterial renal em vista anterior. (Obtido de Net-ter, F.H. Anatomia, estrutura e embriologia. SeçãoI: rins, ureteres e bexiga. Ciba-Geigy, vol. 6, 1973.4)
Nefro cortical
Arteríolaaferente
Artériae veia
interlobularC
ó rtex
Nefro justamedular
Zonaexterna
Zonainterna
Medula
Faixa
externa
Faixainterna
Artériae veiainterlobular
Ductocoletor
Vasarecta
Alça deHenleespessa
Alça deHenledelgada
Ductos de
Bellini
Fig. 1.5 Diagrama ilustrando a circulação renal. (Obtido de Pitts,R.F.3)
Arteríolaeferente
Artéria do segmento superior
Artéria do segmentoântero-superior
Ramos capsular eperirrenal
Artéria supra-renal inferior
Ramo (divisão) anteriorda artéria renal
Artéria renal
Ramo (divisão) posterior da artériarenal (artéria do segmento posterior)
Ramos pélvico e uretérico
Artéria do segmentoântero-inferior
Artérias dos segmentos posteriores
Artéria do segmento inferior
Secção frontal do rim esquerdo:vista anterior
Artériasinterlobares
Artériasarqueadas
Artériasinterlobulares
Ramocapsularperfurante

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 37/121
capítulo 1 5
camente, a circulação venosa costuma seguir paralela-mente o trajeto do sistema arterial. As veias são forma-das perto da superfície do rim por confluência dos capi-
lares do córtex. Elas drenam nas veias interlobulares etornam-se veias arqueadas na junção do córtex com amedula (v. Fig. 1.5). As vênulas retas na medula tambémdrenam nas veias arqueadas, que então formam as veiasinterlobares. Estas veias interlobares drenam em veiassegmentares, as quais, eventualmente, formam as veiasrenais. A veia renal esquerda recebe a veia adrenal esquer-da e a veia gonadal esquerda e passa inferiormente à ar-téria mesentérica superior antes de entrar na veia cavainferior. As veias adrenal e gonadal direita entram dire-tamente na veia cava inferior. A veia renal direita é me-nor e situa-se dorsalmente ao duodeno.
Devido à migração dos rins durante seu desenvolvimen-to, a vascularização renal também apresenta um processode transformação no qual as artérias mais inferiores regri-
dem e novas artérias mais superiores surgem, acompa-nhando o trajeto renal. Quando as artérias inferiores nãoregridem, os rins podem apresentar artérias acessórias, asquais devem ser identificadas pela arteriografia antes deum procedimento cirúrgico renal.
CIRCULAÇÃO LINFÁTICA
Aparentemente, nos animais estudados, os linfáticosintra-renais distribuem-se, primariamente, ao longo dasartérias interlobulares e arqueadas, não penetrando noparênquima propriamente. Os linfáticos corticais origi-nam-se ao nível do tecido conjuntivo que envolve as arté-
rias interlobulares, drenam nos linfáticos arqueados na junção córtico-medular e atingem os linfáticos do hilo atra-vés dos linfáticos interlobares. Há também uma rede lin-fática no interior e sob a cápsula renal, comunicando-secom os linfáticos intra-renais.5
INERVAÇÃO
Origina-se basicamente do plexo celíaco. Há, no entan-to, contribuições do plexo hipogástrico superficial e denervos intermesentéricos, esplâncnicos superiores e torá-cicos. A distribuição das fibras nervosas segue os vasosarteriais através do córtex e medula externa. Parece nãohaver inervação nos túbulos renais (com exceção do apa-
relho justaglomerular) e tampouco terminações nervosasnos glomérulos. No entanto, uma extensa inervação dosvasos arteriolares eferentes foi descrita. Há evidência dapresença, no parênquima renal, de fibras nervosas colinér-gicas e adrenérgicas. As fibras para a sensibilidade dolo-rosa, principalmente a partir da pelve renal e da parte su-perior do ureter, penetram na medula espinhal através dosnervos esplâncnicos.12
Pontos-chave:
• A circulação renal apresenta uma
característica única: duas redes capilares seencontram em série em um mesmo órgão —rede capilar e peritubular
• A artéria renal se divide em ramo anterior eramo posterior. Algumas vezes é possívelencontrar artérias acessórias renais, as quaisapresentam importância cirúrgica, porexemplo na nefrectomia
• A inervação simpática renal atuaprincipalmente nas arteríolas aferentes eeferentes e no aparelho justaglomerular.Nesta estimula a secreção de renina enaquela atua na musculatura lisa
• A inervação aferente da dor tambémapresenta papel importante, pois podeajudar a localizar a altura de um cálculo emmigração. O rim distendido estimula asterminações nervosas da cápsula renal eprovoca dor em região lombar agravada àpunho-percussão. Já a dilatação ureteral porcálculo causa dor que segue o trajeto doureter à medida que o cálculo desce, comirradiação para a genitália quando localizadoprincipalmente no segmento inferior ureteral
EMBRIOLOGIAO desenvolvimento do sistema urinário está intimamente
relacionado com o do sistema genital, sendo estes os últimossistemas a se desenvolverem durante a embriogênese. Ambostêm origem mesodérmica, e seus canais excretores penetraminicialmente numa cavidade comum denominada cloaca.Durante o desenvolvimento embrionário, identificam-se trêssistemas de excreção: pronefro, mesonefro, que são transitó-rios, e metanefro, ou rim permanente. Estes sistemas originam-se do mesoderma intermediário ou cordão nefrogênico.
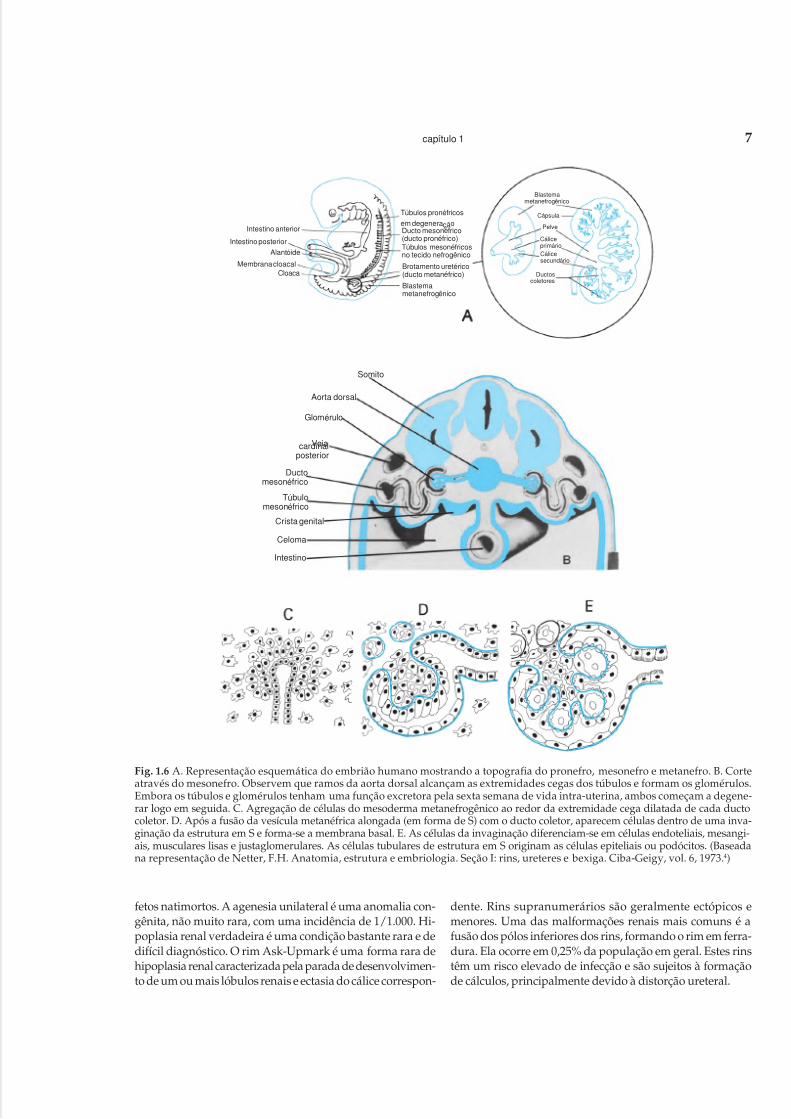
PronefroA sua formação se inicia por volta da terceira semana de
vida. Cada pronefro é composto de aproximadamente setetúbulos. A porção cefálica degenera-se e forma nefrostomasque se abrem na cavidade celômica (Fig. 1.6 A). A porçãocaudal funde-se com a do lado oposto, formando o ductopronéfrico (mesonefro), que se abre na cloaca.
Na maioria dos vertebrados adultos, o pronefro é vesti-gial ou nem sempre existe, embora na lampreia, o mais in-ferior dos vertebrados, funcione como um rim permanente.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 38/121
6 Anatomia Renal
MesonefroDesenvolve-se a partir da quarta semana, numa posição
caudal à do pronefro. Cada túbulo mesonéfrico possui uma
estrutura glomerular proximal, um segmento tubular pro-ximal e um distal, que se abre no ducto mesonéfrico (Fig.1.6 B). Nos peixes superiores e nos anfíbios, o mesonefro éo órgão excretor final. Nos répteis, aves e mamíferos, omesonefro também degenera, formando o metanefro emposição mais caudal.
No homem, os túbulos e ductos mesonéfricos originamvários componentes do sistema reprodutor masculino: epi-dídimo, ducto deferente e vesículas seminais. Na mulher osmesonefros degeneram e os ductos de Müller, que aparecemna oitava semana, originarão o útero, a vagina e as trompas.
MetanefroRepresenta o desenvolvimento final do rim do mamí-
fero. Sua formação resulta da interação entre o broto ure-teral, que surge a partir do ducto metanéfrico por volta daquarta e da quinta semana, com o blastema metanéfrico,derivado da parte caudal do mesoderma intermediário(Fig. 1.6 A). Estudos atribuem ao broto ureteral um papelimportante como indutor da gênese renal, uma vez que naausência ou no distúrbio de sua interação com a massametanéfrica o metanefro não se forma, constituindo oscasos de agenesia renal.
Após se dilatar e se subdividir em cálices primários esecundários, seguindo um padrão muito bem estudado porOsathanondh e Potter, o broto ureteral formará o sistemacoletor do rim: pelve, cálices e os ductos coletores; enquan-to o blastema formará o sistema excretor: corpúsculo re-nal, túbulos proximais e distais e alça de Henle.13,14
O blastema metanefrogênico origina-se de pequenos focosde mesênquima condensado, localizados ao lado do brotoureteral. As células do mesoderma metanefrogênico, estimu-ladas pelo epitélio da extremidade cega dilatada de cadaducto coletor, agregam-se ao redor desta (Fig. 1.6 C), sofremdiversas mitoses e estágios de diferenciação, formando inici-almente uma vesícula que se alonga e se une à luz do ductocoletor. Esta vesícula alongada tem a forma de um S; umadepressão na extremidade do S é o local do futuro gloméru-lo. Nesta depressão aparecem células mesenquimais e, a se-guir, forma-se uma membrana basal ao redor da estrutura emS (Fig. 1.6 D). Algumas células tubulares da estrutura em Sformarão as células epiteliais ou podócitos, e as células me-senquimais darão origem às células endoteliais e mesangiais.Outras células mesenquimais se diferenciam em células mus-
culares lisas das arteríolas aferentes e eferentes (Fig. 1.6 E).Portanto, esta vesícula alongada em S origina, na suaporção inferior, o corpúsculo renal, e na outra porção ori-gina os túbulos proximal e distal da alça de Henle.
Com o crescimento do sistema coletor e a indução do blastema metanéfrico simultaneamente, tem-se um padrãode crescimento centrífugo ao longo do córtex renal, de talmaneira que os primeiros néfrons passam a ocupar uma
posição justamedular, enquanto os últimos encontram-semais externamente no córtex.
Embora os néfrons do metanefro comecem a funcionar em
torno da 11-ª e 12-ª semanas de vida, a maturação renal conti-nua após o nascimento. Alguns autores sugerem que o cor-púsculo renal pode aumentar por 20 anos e os túbulos con-tornados proximal e distal chegam a atingir um comprimen-to 10 vezes maior, desde o nascimento até a vida adulta.
Naturalmente, o que foi descrito é uma explicação sim-plificada do desenvolvimento do rim, e não envolve as vá-rias teorias e fatores existentes para explicar este complexoprocesso. Não foram enfatizados aqui os mecanismos celu-lares e moleculares da formação renal. Segundo Clapp eAbrahamson, estes parecem estar relacionados também comas desordens genéticas primárias do rim, com o tipo de res-posta e com a recuperação renal frente às doenças, o quetorna o seu entendimento de grande importância.15
Anomalias do Desenvolvimento
Podemos observar ausência congênita ou agenesia de umou ambos os rins, assim como hipoplasia renal. Agenesia bi-lateral, quando presente, é freqüentemente observada em
Pontos-chave:
• O sistema urinário tem origem namesoderma intermediária. Durante aembriogênese, ocorre regressão de algumasestruturas, sendo que são os metanefros osresponsáveis pela formação do rim adulto
•
A agenesia renal bilateral não é compatívelcom a vida e deve ser suspeitada napresença de oligoidrâmnio por volta da 14-ªsemana de gestação
• A doença policística da infância é outragrave enfermidade que leva a insuficiênciarenal e morte, caso um transplante renal nãoseja realizado. É uma doença autossômicarecessiva, diferente da forma do adulto, aqual é autossômica dominante
• A migração deficiente do rim pode levar alocalização pélvica renal, a qual apresentaprincipal importância no diagnósticodiferencial de massa pélvica
• O rim em ferradura é uma anormalidaderelativamente comum causada pela fusãodos pólos inferiores dos rins. O rim se situaem região lombar baixa devido àincapacidade de migração superiormente àraiz da artéria mesentérica inferior
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 39/121
capítulo 1 7
fetos natimortos. A agenesia unilateral é uma anomalia con-gênita, não muito rara, com uma incidência de 1/1.000. Hi-poplasia renal verdadeira é uma condição bastante rara e dedifícil diagnóstico. O rim Ask-Upmark é uma forma rara dehipoplasia renal caracterizada pela parada de desenvolvimen-to de um ou mais lóbulos renais e ectasia do cálice correspon-
dente. Rins supranumerários são geralmente ectópicos emenores. Uma das malformações renais mais comuns é afusão dos pólos inferiores dos rins, formando o rim em ferra-dura. Ela ocorre em 0,25% da população em geral. Estes rinstêm um risco elevado de infecção e são sujeitos à formaçãode cálculos, principalmente devido à distorção ureteral.
Intestino anterior
Intestino posterior
Alantóide
Membrana cloacal
Cloaca
Túbulos pronéfricos
em degeneraçãoDucto mesonéfrico(ducto pronéfrico)Túbulos mesonéfricosno tecido nefrogênico
Blastemametanefrogênico
Blastemametanefrogênico
Cápsula
Pelve
Cáliceprimário
Cálicesecundário
Ductoscoletores
Brotamento uretérico(ducto metanéfrico)
Somito
Aorta dorsal
Glomérulo
Veiacardinalposterior
Ductomesonéfrico
Túbulomesonéfrico
Crista genital
Celoma
Intestino
Fig. 1.6 A. Representação esquemática do embrião humano mostrando a topografia do pronefro, mesonefro e metanefro. B. Corteatravés do mesonefro. Observem que ramos da aorta dorsal alcançam as extremidades cegas dos túbulos e formam os glomérulos.Embora os túbulos e glomérulos tenham uma função excretora pela sexta semana de vida intra-uterina, ambos começam a degene-rar logo em seguida. C. Agregação de células do mesoderma metanefrogênico ao redor da extremidade cega dilatada de cada ductocoletor. D. Após a fusão da vesícula metanéfrica alongada (em forma de S) com o ducto coletor, aparecem células dentro de uma inva-ginação da estrutura em S e forma-se a membrana basal. E. As células da invaginação diferenciam-se em células endoteliais, mesangi-ais, musculares lisas e justaglomerulares. As células tubulares de estrutura em S originam as células epiteliais ou podócitos. (Baseadana representação de Netter, F.H. Anatomia, estrutura e embriologia. Seção I: rins, ureteres e bexiga. Ciba-Geigy, vol. 6, 1973.4)
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 40/121
8 Anatomia Renal
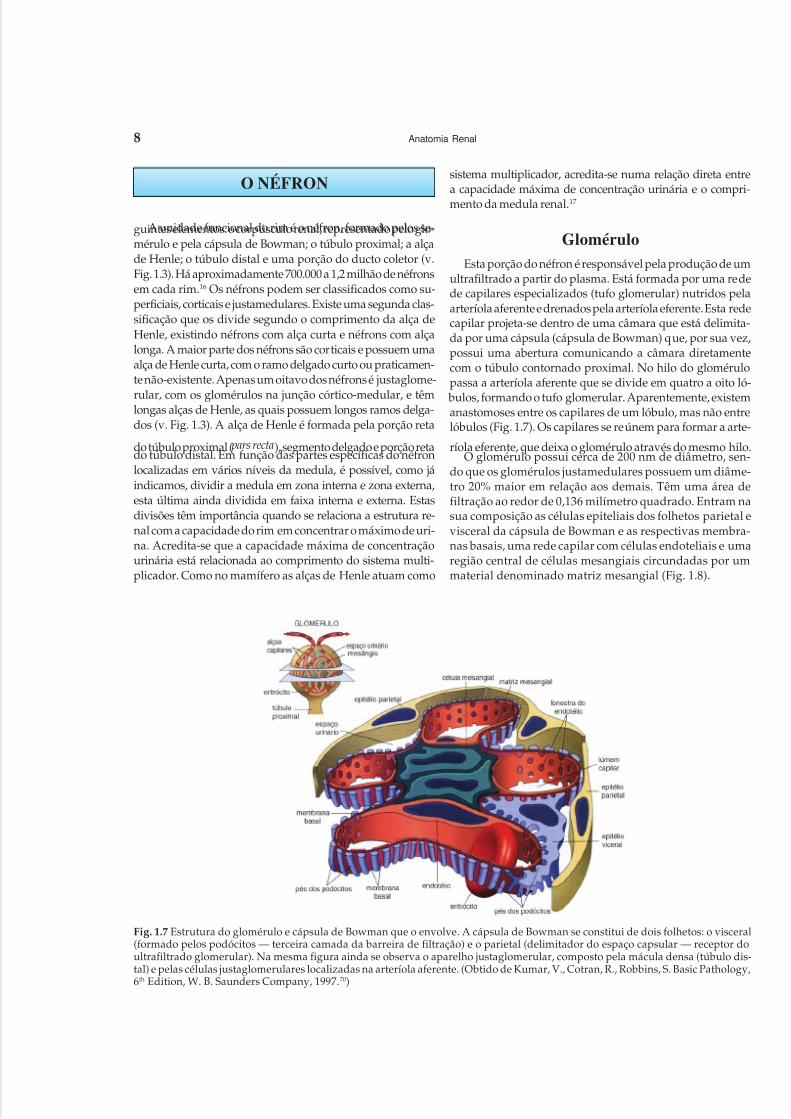
O NÉFRON
A unidade funcional do rim é o néfron, formado pelos se-guintes elementos: o corpúsculo renal, representado pelo glo-mérulo e pela cápsula de Bowman; o túbulo proximal; a alçade Henle; o túbulo distal e uma porção do ducto coletor (v.Fig. 1.3). Há aproximadamente 700.000 a 1,2 milhão de néfronsem cada rim.16 Os néfrons podem ser classificados como su-perficiais, corticais e justamedulares. Existe uma segunda clas-sificação que os divide segundo o comprimento da alça deHenle, existindo néfrons com alça curta e néfrons com alçalonga. A maior parte dos néfrons são corticais e possuem umaalça de Henle curta, com o ramo delgado curto ou praticamen-te não-existente. Apenas um oitavo dos néfrons é justaglome-rular, com os glomérulos na junção córtico-medular, e têmlongas alças de Henle, as quais possuem longos ramos delga-dos (v. Fig. 1.3). A alça de Henle é formada pela porção reta
do túbulo proximal ( pars recta), segmento delgado e porção retado túbulo distal. Em função das partes específicas do néfronlocalizadas em vários níveis da medula, é possível, como jáindicamos, dividir a medula em zona interna e zona externa,esta última ainda dividida em faixa interna e externa. Estasdivisões têm importância quando se relaciona a estrutura re-nal com a capacidade do rim em concentrar o máximo de uri-na. Acredita-se que a capacidade máxima de concentraçãourinária está relacionada ao comprimento do sistema multi-plicador. Como no mamífero as alças de Henle atuam como
sistema multiplicador, acredita-se numa relação direta entrea capacidade máxima de concentração urinária e o compri-mento da medula renal.17
GloméruloEsta porção do néfron é responsável pela produção de um
ultrafiltrado a partir do plasma. Está formada por uma redede capilares especializados (tufo glomerular) nutridos pelaarteríola aferente e drenados pela arteríola eferente. Esta redecapilar projeta-se dentro de uma câmara que está delimita-da por uma cápsula (cápsula de Bowman) que, por sua vez,possui uma abertura comunicando a câmara diretamentecom o túbulo contornado proximal. No hilo do glomérulopassa a arteríola aferente que se divide em quatro a oito ló- bulos, formando o tufo glomerular. Aparentemente, existemanastomoses entre os capilares de um lóbulo, mas não entrelóbulos (Fig. 1.7). Os capilares se reúnem para formar a arte-
ríola eferente, que deixa o glomérulo através do mesmo hilo.O glomérulo possui cerca de 200 nm de diâmetro, sen-
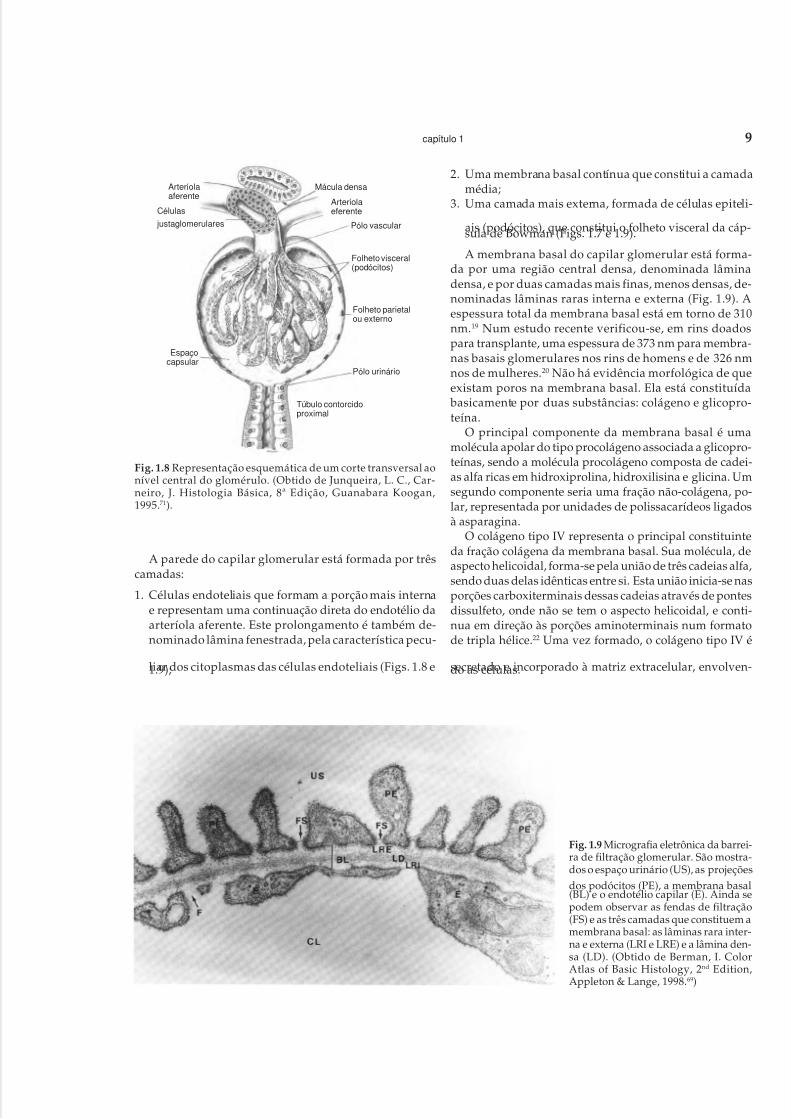
do que os glomérulos justamedulares possuem um diâme-tro 20% maior em relação aos demais. Têm uma área defiltração ao redor de 0,136 milímetro quadrado. Entram nasua composição as células epiteliais dos folhetos parietal evisceral da cápsula de Bowman e as respectivas membra-nas basais, uma rede capilar com células endoteliais e umaregião central de células mesangiais circundadas por ummaterial denominado matriz mesangial (Fig. 1.8).
Fig. 1.7 Estrutura do glomérulo e cápsula de Bowman que o envolve. A cápsula de Bowman se constitui de dois folhetos: o visceral(formado pelos podócitos — terceira camada da barreira de filtração) e o parietal (delimitador do espaço capsular — receptor doultrafiltrado glomerular). Na mesma figura ainda se observa o aparelho justaglomerular, composto pela mácula densa (túbulo dis-tal) e pelas células justaglomerulares localizadas na arteríola aferente. (Obtido de Kumar, V., Cotran, R., Robbins, S. Basic Pathology,6th Edition, W. B. Saunders Company, 1997.70)
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 41/121
capítulo 1 9
A parede do capilar glomerular está formada por trêscamadas:
1. Células endoteliais que formam a porção mais internae representam uma continuação direta do endotélio daarteríola aferente. Este prolongamento é também de-nominado lâmina fenestrada, pela característica pecu-
liar dos citoplasmas das células endoteliais (Figs. 1.8 e1.9);
2. Uma membrana basal contínua que constitui a camadamédia;
3. Uma camada mais externa, formada de células epiteli-
ais (podócitos), que constitui o folheto visceral da cáp-sula de Bowman (Figs. 1.7 e 1.9).
A membrana basal do capilar glomerular está forma-da por uma região central densa, denominada lâminadensa, e por duas camadas mais finas, menos densas, de-nominadas lâminas raras interna e externa (Fig. 1.9). Aespessura total da membrana basal está em torno de 310nm.19 Num estudo recente verificou-se, em rins doadospara transplante, uma espessura de 373 nm para membra-nas basais glomerulares nos rins de homens e de 326 nmnos de mulheres.20 Não há evidência morfológica de queexistam poros na membrana basal. Ela está constituída basicamente por duas substâncias: colágeno e glicopro-teína.
O principal componente da membrana basal é umamolécula apolar do tipo procolágeno associada a glicopro-teínas, sendo a molécula procolágeno composta de cadei-as alfa ricas em hidroxiprolina, hidroxilisina e glicina. Umsegundo componente seria uma fração não-colágena, po-lar, representada por unidades de polissacarídeos ligadosà asparagina.
O colágeno tipo IV representa o principal constituinteda fração colágena da membrana basal. Sua molécula, deaspecto helicoidal, forma-se pela união de três cadeias alfa,sendo duas delas idênticas entre si. Esta união inicia-se nasporções carboxiterminais dessas cadeias através de pontesdissulfeto, onde não se tem o aspecto helicoidal, e conti-nua em direção às porções aminoterminais num formatode tripla hélice.22 Uma vez formado, o colágeno tipo IV é
secretado e incorporado à matriz extracelular, envolven-do as células.
Arteríolaaferente
Células
justaglomerulares
Mácula densa
Arteríolaeferente
Pólo vascular
Folheto visceral(podócitos)
Folheto parietalou externo
Pólo urinário
Túbulo contorcidoproximal
Espaçocapsular
Fig. 1.8 Representação esquemática de um corte transversal aonível central do glomérulo. (Obtido de Junqueira, L. C., Car-neiro, J. Histologia Básica, 8ª Edição, Guanabara Koogan,1995.71).
Fig. 1.9 Micrografia eletrônica da barrei-ra de filtração glomerular. São mostra-dos o espaço urinário (US), as projeçõesdos podócitos (PE), a membrana basal(BL) e o endotélio capilar (E). Ainda sepodem observar as fendas de filtração(FS) e as três camadas que constituem amembrana basal: as lâminas rara inter-na e externa (LRI e LRE) e a lâmina den-sa (LD). (Obtido de Berman, I. ColorAtlas of Basic Histology, 2nd Edition,Appleton & Lange, 1998.69)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 42/121
10 Anatomia Renal
Já foram identificados tipos diferentes de cadeias alfaformadoras de colágeno tipo IV. A cadeia alfa-1, codifica-da pelo gene COL4A1, e a cadeia alfa-2,23 codificada pelo
gene COL4A2, ambos situados no cromossomo 13, apare-cem no mesângio, na membrana basal glomerular (suben-dotelial), na cápsula de Bowman, em toda membrana ba-sal tubular e vasos. A cadeia alfa-3,24 codificada pelo geneCOL4A3, a cadeia alfa-4,25 codificada pelo gene COL4A4,localizado no cromossomo 2, e a cadeia alfa-5, codificadapelo gene COL4A5, situado no braço longo do cromosso-mo X,26 aparecem na membrana basal glomerular (lâminadensa), na cápsula de Bowman e na membrana basal dotúbulo distal.
Alterações nessas cadeias podem levar ao surgimentode alterações estruturais com conseqüências mórbidas,como a síndrome de Alport, onde foi detectada ausênciadas cadeias alfa-3 e alfa-4 na membrana basal glomerular,27
em função de uma mutação do gene da cadeia alfa-5.28 Esta
mutação impede a formação do colágeno tipo IV, uma vezque as cadeias alfa-3 e alfa-4 necessitam da cadeia alfa-5para formar a tripla hélice. Como conseqüência, observam-se graus variados de malformação estrutural da membra-na basal, com repercussões na filtração e seletividade damesma ao longo do tempo.
Ao contrário dos outros tipos de colágeno, o colágenotipo IV apresenta nas suas cadeias numerosas seqüênciasGly-X-Y, onde X e Y representam outros tipos de aminoá-cidos, aumentando a flexibilidade da molécula.29 Alémdisso, o colágeno tipo IV não perde sua porção carboxiter-minal após ser secretado pela célula, o que possibilita trêstipos diferentes de interações entre as moléculas: porçãocarboxiterminal de uma molécula com porção carboxiter-minal de outra (head-to-head); porção carboxiterminal deuma com porção lateral da tripla hélice de outra;30 e, final-mente, porção aminoterminal de uma com porção amino-terminal de outras três moléculas (tail-to-tail). Com isso,temos a formação de uma rede poligonal, não-fibrilar e fle-xível que servirá de arcabouço para o depósito de glicopro-teínas e para a fixação das células.31
Colágeno tipo V,32 laminina, fibronectina33 e entactina/nidógeno34 também foram identificados na membrana basal.
Dados recentes indicam que a membrana basal do glo-mérulo possui locais fixos de cargas negativas capazes deinfluenciar a filtração de macromoléculas.35 Ela seria a prin-cipal responsável pela seletividade da filtração glomeru-lar, permitindo ou não a passagem de moléculas, de acor-do com a carga elétrica e com o tamanho destas. Num ex-
perimento, empregando-se o processo de digestão enzimá-tica, retiraram-se os glicosaminoglicanos ricos em heparansulfato, presentes no lado aniônico da membrana basal, enotou-se um aumento da permeabilidade à ferritina36 e àalbumina sérica em bovinos.37
Os efeitos de danos glomerulares, alterando a seletivida-de e a permeabilidade da membrana basal, foram estuda-dos utilizando-se o modelo experimental de nefrite causa-
da por soro nefrotóxico.38 Evidenciou-se que nessa situaçãoexperimental há perda ou diminuição do conteúdo polianiô-nico da membrana basal, explicando um aumento na filtra-
ção de poliânions circulantes, incluindo a albumina. Outrosexperimentos evidenciaram, também, que a perda de cargasnegativas pode influenciar na localização e na magnitude dadeposição de imunocomplexos, bem como na deposição deagregados circulantes não-imunes no mesângio e na paredeglomerular.39 Esses agregados levam a um estímulo contínuoà produção de matriz mesangial, que, quando se estende pormuito tempo, pode levar à esclerose nodular.
CÉLULAS ENDOTELIAISRevestem o lúmen dos capilares glomerulares. O núcleo
e a maior parte do citoplasma estão no lado mesangial docapilar, sendo que uma estreita faixa do citoplasma esten-de-se ao longo da parede capilar (Fig. 1.13). Esta faixa decitoplasma é contínua, mas apresenta várias fenestras ou
poros, cujo diâmetro aproximado é de 70 a 100 nm (Fig. 1.9).Membranas delgadas, ou diafragmas, foram observadasentre poros (Fig. 1.12). Alguns acreditam que estes diafrag-mas são altamente permeáveis e não constituem barreiraà passagem de moléculas maiores.
Estas células possuem uma superfície carregada nega-tivamente devido à presença de glicoproteínas polianiôni-cas, como a podocalixina.40 Na sua membrana são apresen-tados antígenos como os de grupo sanguíneo ABO e HLAde tipos I e II.
CÉLULAS MESANGIAISMuitos acreditam serem de origem mesenquimal, pois
apresentam certas propriedades características das células
do músculo liso. As células têm forma irregular, com vári-os processos citoplasmáticos estendendo-se do corpo dacélula. Na região paramesangial e ao longo dos processoscitoplasmáticos mesangiais justamedulares, foi evidenci-ada uma extensa rede de microfilamentos compostos pelomenos em parte por actina, alfa-actina e miosina.41 Suamembrana plasmática apresenta receptores de B1-integri-na para fibronectina e, talvez, também para laminina.42
O material que as circunda, aparentemente sintetizadopelas próprias células, chama-se matriz mesangial. Nela seencontram glicosaminoglicanos sulfatados, laminina e fi- bronectina.43 É similar na aparência mas não idêntica àmembrana basal do glomérulo.
Ao conjunto célula mesangial e matriz dá-se o nome demesângio. Este está separado da luz capilar pelo endotélio.
A função da célula mesangial não está bem definida,mas, além da função de suporte estrutural, ela provavel-mente participa de mecanismos de fagocitose e da modu-lação da filtração glomerular, regulando o fluxo sanguíneonos capilares glomerulares através de suas propriedadesmusculares de contração e relaxamento. A célula mesan-gial também produz muitos agentes vasoativos, sintetizae degrada várias substâncias do tufo glomerular.44

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 43/121
capítulo 1 11
Segundo Schlondorff, substâncias como vasopressina, an-giotensina II, fator de ativação plaquetária, tromboxane, leu-cotrienos e fator de crescimento derivado de plaqueta atuam
na indução da contração da célula mesangial.44 A produçãolocal de prostaglandina E2, pela própria célula mesangial, fariao papel contrário dos vasoconstritores anteriormente citados.
Acredita-se, no entanto, que esse mecanismo de contra-ção seria mais para prevenir a distensão da parede capilare para elevar a pressão hidrostática intracapilar45 e não tan-to para ser o controle da filtração glomerular.
Há evidências de que células mesangiais tenham proprie-dades de endocitose de imunocomplexos, fagocitose, de pro-duzir e de ser alvo de substâncias reguladoras de crescimentocelular, além de atuarem na modulação de dano celular glo-merular.44 A produção de prostaglandinas influencia a proli-feração celular local, a produção de citocinas, a produção e adestruição de matriz mesangial e de membrana basal. A inte-ração entre células mesangiais, prostaglandinas e citocinas
deve fornecer pistas importantes para a compreensão da le-são glomerular presente nos processos patológicos.
Além do mais, é provável que a célula mesangial possatransformar-se em célula endotelial quando houver neces-sidade da expansão da rede capilar.
CÉLULAS EPITELIAIS VISCERAISConhecidas também como podócitos, são as maiores
células do glomérulo. Possuem lisossomos proeminentes,um aparelho de Golgi bem desenvolvido e muitos filamen-tos de actina. Do corpo da célula, estendem-se trabéculasalongadas, das quais se originam processos denominadospedicelos ou pés dos podócitos, que ficam em contato coma lâmina rara externa da membrana basal do glomérulo
(Figs. 1.7 e 1.10). A distância entre os pés dos podócitos
varia de 25 a 60 nm, ao nível da membrana basal. Este es-paço é também referido como fenda de filtração ou, impro-priamente, poro (Fig. 1.12). Aqui também há uma membra-
na delgada ou diafragma entre os pés dos podócitos. Nelaencontrou-se, por estudos imuno-histoquímicos,46 a prote-ína ZO-1, específica dos complexos unitivos intercelulares(tight junctions). Uma densidade central com um diâmetrode 11 nm é observada neste diafragma. Esta densidaderepresenta um filamento central contínuo conectado àmembrana plasmática do pedicelo adjacente por pontesespaçadas regularmente com 7 nm de diâmetro e 14 nm decomprimento, dando uma configuração semelhante a umzíper.47 Discute-se se esta estrutura também entra na de-terminação da seletividade da barreira de filtração.
Na superfície das células epiteliais viscerais notou-se oreceptor C3b em glomérulos humanos,48 bem como o antí-geno de Heymann, gp 330.49 A superfície negativa que co- bre os pedicelos é rica em ácido siálico. Encontrou-se tam- bém podoxilina na superfície urinária, mas não na super-fície basal, dos podócitos.50
Em várias nefropatias associadas com proteinúria, os pésdos podócitos são substituídos por uma faixa contínua decitoplasma adjacente à lâmina rara externa. Este aspecto temsido denominado fusão dos pés dos podócitos. É uma ex-pressão errônea porque não se sabe se realmente há umafusão, e tudo indica que alguns pés na verdade se retraem eos que permanecem expandem-se. A fusão dos pés dospodócitos resulta, pelo menos em parte, da perda de forçaseletrostáticas repulsoras normais entre os processos adjacen-tes, devido à neutralização (ou perda) de sua cobertura ani-ônica. Em estudos experimentais, com a perfusão de rins derato com neuroaminidase, que remove ácido siálico, obser-
vou-se que tanto as células viscerais quanto as epiteliais
Fig. 1.10 Imagem de microscopia eletrônica das células viscerais da cápsula de Bowman (podócitos). São visualizados os corposcelulares dos podócitos (CB) e as projeções citoplasmáticas primárias (PB) e secundárias (SB). (Obtido de Berman, I. Color Atlas ofBasic Histology, 2nd Edition, Appleton & Lange, 1998.69)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 44/121
12 Anatomia Renal
descolam-se da membrana basal glomerular.51 Portanto,sugere-se que os campos de carga negativa da membranadestas células sejam muito importantes na manutenção da
estrutura e da função da barreira de filtração.Acredita-se que a célula epitelial visceral seja capaz defazer endocitose, capturando proteínas e outros componen-tes do ultrafiltrado, e que ela também seja responsável, pelomenos em parte, pela síntese e manutenção da membrana basal do glomérulo,52 embora ainda se conheça pouco so- bre a dinâmica desse processo.
Pontos-chave:
• O néfron é a unidade funcional do rim e éconstituído pelo corpúsculo renal(glomérulo cápsula de Bowman), túbulocontorcido proximal, alça de Henle, túbulo
contorcido distal e ducto coletor• A barreira de filtração glomerular é
constituída por três camadas:-Endotélio fenestrado do capilar glomerular-Membrana basal-Células epiteliais especializadas (podócitos),as quais circunscrevem os capilares com suasprojeções citoplasmáticas, formandoinúmeras fendas de filtração
• Esta complexa barreira permite a passagemseletiva de água e pequenos solutos.Moléculas de carga negativa apresentam umamenor taxa de filtração em relação a cátions
devido à negatividade da barreira glomerular• Alterações estruturais na barreira podemlevar a uma série de doenças renais, dentreelas as glomerulonefrites primárias
• A fusão dos pés dos podócitos está presentena nefrose lipoídica e na glomeruloesclerosefocal e segmentar, levando a um quadro desíndrome nefrótica com proteinúria maciça
• As glomerulonefrites rapidamenteprogressivas apresentam à microscopiaóptica uma proliferação anormal das célulasepiteliais parietais associada à infiltração demonócitos e macrófagos, formando ascrescentes glomerulares
• A nefropatia por IgA é uma doençaglomerular extremamente comumcaracterizada por hematúria recorrente,freqüentemente seguindo um quadroinfeccioso. As imunoglobulinas A sãodepositadas no mesângio glomerular
CÉLULAS EPITELIAIS PARIETAISSão células escamosas que revestem a parede externa da
cápsula de Bowman (v. Fig. 1.7). Possuem esparsas orga-
nelas, pequenas mitocôndrias e numerosas vesículas, de 40a 90. Apresentam microvilosidades de até 600 nm de com-primento na superfície livre e, freqüentemente, em cadacélula encontra-se um longo cílio. Estas células são respon-sáveis pela manutenção da integridade da cápsula. Emalgumas nefropatias, como na glomerulonefrite rapida-mente progressiva, estas células parietais podem vir a pro-liferar, vindo a constituir um dos elementos das semiluasou crescentes. O estímulo para esta proliferação parece sera presença de fibrina ou material proteináceo e hemáciasno espaço urinário.
Aparelho Justaglomerular
Está situado no hilo do glomérulo e é formado pelosseguintes elementos:
1. porção terminal da arteríola aferente;2. mácula densa;3. uma região mesangial extraglomerular;4. a arteríola eferente.
A região mesangial extraglomerular está localizada en-tre a mácula densa e as células mesangiais do tufo glome-rular (Figs. 1.7 e 1.11). Nesta região, encontram-se dois ti-pos de células: agranulares e granulares.
As células agranulares ocupam o centro dessa região esão as mais abundantes. As células granulares ou mioepi-teliais (pois parecem representar células especializadas domúsculo liso) estão localizadas principalmente no interior
das paredes das arteríolas glomerulares aferentes e eferen-tes. Os grânulos representam o hormônio renina ou o seuprecursor. Durante o desenvolvimento renal a expressãoda renina aparece ao longo de todas as arteríolas do glo-mérulo em formação. Especula-se que a alta expressão derenina esteja relacionada à proliferação vascular.53
Fig. 1.11 Diagrama do aparelho justaglomerular.
Células agranulares
Célulasgranulares
Mácula densa A r t é r i a
a f e
r e n t
eA r t e
r í o l a e f e r e
n t e

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 45/121
capítulo 1 13
A mácula densa deriva de células epiteliais da bordasuperior da fissura vascular, que se estabelecem no seg-mento ascendente espesso da alça de Henle, parte do tú-
bulo distal. O túbulo distal está em extenso contato com aarteríola eferente e com a região mesangial extraglomerulare possui um contato menos extenso com a arteríola aferen-te. O corte transversal do túbulo distal, a este nível, mos-tra que as células adjacentes do hilo são distintas das de-mais: são colunares, com um núcleo apical (v. Fig. 1.11). Amicroscopia eletrônica mostra interdigitações entre a baseda célula e as células mesangiais extraglomerulares. Oaparelho justaglomerular é a estrutura mais importante dosistema renina-angiotensina. Ele parece participar do me-canismo de feedback entre o túbulo distal e as arteríolas afe-rentes e eferentes, atuando ativamente na regulação daexcreção de sódio pelo organismo (v. Cap. 10). Há duasteorias para explicar o mecanismo de liberação de reninapelo aparelho justaglomerular: a da mácula densa e a do
receptor de volume.A primeira infere que a concentração de sódio na má-
cula densa controla a liberação de renina;54 a segunda, quealterações no volume da arteríola aferente seriam respon-sáveis pelo fato.55
Posteriormente, as duas teorias foram integradas naexplicação deste mecanismo, que se baseia no seguinteprincípio: quanto maior for o contato entre o túbulo e oscomponentes vasculares do aparelho justaglomerular,menor quantidade de renina é secretada; quanto menor foreste contato, maior será a secreção da substância. Assim,pela teoria da mácula densa, quanto menos sódio atingiro túbulo distal, menor o diâmetro do túbulo e, portanto,menor o contato com os componentes vasculares, haven-do, então, um aumento da secreção de renina. O inversoocorre quando muito sódio chega ao túbulo distal. Pelaoutra teoria, um aumento do volume arteriolar aumenta ocontato dos componentes vasculares com o túbulo distal,e logo menos renina é liberada. Havendo um volume arte-riolar reduzido, ocorrerá o contrário.56
Atualmente, sabe-se que o sistema simpático também écapaz de estimular a secreção de renina.
Células Peripolares
Acredita-se que sejam um componente adicional doaparelho justaglomerular. Encontram-se interpostas entrecélulas epiteliais parietais e viscerais na origem do tufoglomerular da cápsula de Bowman, estando comumente
separadas da arteríola aferente pela membrana basal dacápsula. Têm seu lado oposto voltado para o espaço uri-nário ou espaço de Bowman.
Estas células possuem grânulos eletrondensos que seacredita serem do tipo secretório. Evidenciaram-seexocitoses desse material granular em rins de ovelhasdepletadas de sódio. Acredita-se que as células peripolaresestejam envolvidas no controle da função do aparelho
justaglomerular e especula-se que a liberação de seus fa-tores no espaço de Bowman afete o transporte de elemen-tos distalmente do corpúsculo renal.
Túbulo Proximal
Com cerca de 14 nm de comprimento, inicia-se no pólourinário do glomérulo, forma vários contornos próximos aoglomérulo de origem e depois desce, sob a forma de segmen-to reto, em direção à medula. O segmento inicial é geralmen-te denominado pars convoluta e o mais distal, pars recta, sen-do que estes últimos constituem parte dos raios medulares.As células da pars convoluta são colunares e possuem um bordo em escova, devido às projeções da membrana plas-mática, denominadas microvilos (Figs. 1.12 e 1.13).
Há numerosas mitocôndrias alongadas, estendendo-se da base ao ápice da célula, possuindo ramificações e anastomo-ses entre elas.61 Essas células também possuem numerososprocessos interdigitais laterais de outras células, o que au-menta o espaço intercelular. A microscopia eletrônica reve-la numerosas mitocôndrias de forma alongada, situadasdentro desses compartimentos formados pelos processosinterdigitais entre células adjacentes. Como resultado des-sa extensa interdigitação lateral entre células adjacentes,forma-se um complexo compartimento extracelular, deno-minado espaço intercelular lateral. Este espaço intercelular
Túbulo contorcido
proximal
Túbulo contorcido distale parte espessa da alçade Henle
Parte delgada daalça de Henle
Tubo coletor
Fig. 1.12 Representação esquemática da ultra-estrutura celulardos vários segmentos do néfron. Apesar da semelhança das cé-lulas da parte espessa da alça de Henle e as do túbulo distal, suasfunções são diferentes. (Obtido de Junqueira, L.C. , Carneiro, J.Histologia Básica, 8ª Edição, Guanabara Koogan, 1995.71)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 46/121
14 Anatomia Renal
está separado do lúmen tubular por uma estrutura especi-alizada, localizada na parte superior do espaço e denomi-nada tight junction ou zonula occludens (Fig. 1.13). A impor-tância deste espaço intercelular está na sua participação ati-va na reabsorção de água e de solutos no túbulo proximal,assunto particularmente abordado no Cap. 10. Além disso,a pars convoluta reabsorve várias substâncias protéicas (p. ex.,albumina) e não-protéicas (p. ex., carboidratos).
O epitélio da pars recta é geralmente cubóide. A super-fície apical da célula é convexa e recoberta de microvilos.É uma célula mais simples, com menos vesículas, vacúo-los, mitocôndrias e interdigitações entre as células. Estaredução de complexidade morfológica sugere que esta re-gião está menos envolvida no transporte ativo de sódio eágua quando comparada com a pars convoluta. Essa impres-são é corroborada por estudos experimentais.
O túbulo proximal promove uma reabsorção, quase isos-mótica, de 2/3 do ultrafiltrado, acoplada a transporte ati-vo de sódio. Qualquer doença que afete essa região causaum desequilíbrio hidroeletrolítico mais importante. Ascélulas do túbulo contornado proximal possuem um sis-
tema vacúolo-lisossomal muito bem desenvolvido. Assim,uma importante função da pars convoluta e, em menor grau,da pars recta é a reabsorção e a degradação de várias ma-cromoléculas, inclusive a albumina e proteínas de baixopeso molecular do filtrado glomerular. As proteínas sãoreabsorvidas, levadas ao lisossomo e degradadas. A reab-sorção dá-se juntamente com o transporte ativo de sódio,constituindo um transporte ativo secundário. É um proces-
so seletivo determinado pela carga elétrica e pela distribui-ção desta carga na molécula, além do tamanho e configu-ração moleculares da proteína.
Há evidências também de endocitose mediada por re-ceptor nessas células.
O túbulo proximal é importante na formação de amô-nia e na secreção de íons de hidrogênio.
Foi bem estabelecido que bases fracas, como cloreto deamônio e cloroquina, acumulam-se nos compartimentosacídicos intracelulares, incluindo endossomos e lisosso-mos. Este mecanismo talvez explique o acúmulo de dro-gas catiônicas anfifílicas, como a cloroquina, os antidepres-sivos tricíclicos e os antibióticos aminoglicosídeos. Metaispesados também se acumulam nos lisossomos, provavel-mente porque estão ligados a proteínas.
Por muitos anos sabe-se que a pars recta do túbulo pro-ximal está envolvida na secreção de ácidos e bases orgâni-cas. Assim, essa porção é freqüentemente lesada por com-postos nefrotóxicos, incluindo várias drogas e metais pe-sados, secretados por essa via de transporte.
Alça de HenleA transição entre o túbulo contornado proximal e o seg-
mento delgado da alça de Henle é abrupta e marca a divi-são entre a faixa externa e a faixa interna da zona externa damedula. As células do segmento delgado ascendente têmaspecto morfológico distinto das células do segmento del-gado descendente (Fig. 1.12). Estas últimas são mais com-
Fig. 1.13 Microscopia eletrônica do túbulo proximal mostrando os inúmeros microvilos que constituem o característico bordo emescova da microscopia óptica. Ainda se observam mitocôndrias (M) — responsáveis pela energia para o transporte ativo; lisossomos(L) e vesículas pinocíticas (V), e os complexos juncionais próximos à superfície luminal (J). (Obtido de Burkitt, H.G., YOUNG, B.,HELATH, J.W. Weather’s Functional Histology, 3rd Edition, Churchill Livingston, 1993.68)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 47/121
capítulo 1 15
plexas, irregulares na configuração e apresentam extensasinterdigitações entre si. Este segmento delgado da alça deHenle possui grande importância no mecanismo de concen-
tração da urina, participando do mecanismo de contracor-rente e gerando um interstício medular hipertônico (v. Cap.6). O segmento ascendente é relativamente impermeável àágua, mas bastante permeável a sódio e cloro, enquanto, nosegmento descendente, a água passa passivamente para ointerstício hipertônico e sódio e cloro praticamente não pas-sam. Não há evidências de que nesses segmentos delgadoshaja um transporte ativo de sódio e cloro. Estudos recentesindicam que a concentração de urina na medula interna éum processo puramente passivo, embora o debate aindapersista. Verificou-se que a saída de sódio e cloro do segmen-to ascendente é maior que a entrada de uréia, o que ajudana formação do gradiente osmótico da medula interna.
Túbulo DistalConstitui-se através do segmento ascendente espesso da
alça de Henle ( pars recta), da mácula densa e do túbulocontornado distal ( pars convoluta).
A pars recta atravessa a medula externa e sobe no raiomedular do córtex até ficar em contato com o seu próprioglomérulo. Esta porção tubular contígua ao glomérulo for-ma a mácula densa. As células neste segmento aumentamde altura, tornando-se cubóides na parte média do segmen-to (Fig. 1.11). A transição entre o segmento ascendentedelgado e o segmento espesso marca a divisão entre zonaexterna e zona interna da medula.
A pars recta possui um alto metabolismo, sendo especi-almente sensível à isquemia.62 Nos processos laterais de
suas células e próximas à membrana basal aparecem mui-tas mitocôndrias alongadas, contendo vários tipos de fila-mentos e inclusões cristalinas. A principal função da parsrecta encontra-se no transporte de cloreto de sódio (trans-porte ativo de sódio, ATPase sódio/potássio, e passivo decloro) para o interstício, função muito importante para omecanismo contracorrente58 (v. Cap. 10). A pars recta temsua atividade influenciada por hormônios, como parator-mônio (PTH), vasopressina, calcitonina e glucagon, atra-vés da ativação do sistema adenilato ciclase. O PTH esti-mula a reabsorção de cálcio e magnésio no segmento as-cendente, parte cortical.
A pars convoluta estende-se da mácula densa ao início doducto coletor. As células desse segmento são muito seme-lhantes às da pars recta.
A relação entre a estrutura e a função neste segmentodo néfron é um pouco complicada, pela diferente termino-logia usada por anatomistas e fisiologistas. Para os fisiolo-gistas dedicados à micropunção, o túbulo distal é defini-do como aquela região do néfron que se inicia após a má-cula densa e se estende até a junção com outro túbulo dis-tal (TD). Mas em muitas ocasiões o segmento cortical doramo ascendente da alça de Henle se estende além da
mácula densa e há também evidência anatômica para apresença de uma região de conexão ou transição entre a pars convoluta do túbulo distal e o ducto coletor. Desta
maneira, o túbulo distal pode ser formado por quatro ti-pos diferentes de epitélio.Em geral, a porção inicial do túbulo distal correspon-
de ao túbulo contornado distal ou pars convoluta do ana-tomista. Este possui a maior atividade sódio/potássioATPase, comparando-se com os demais segmentos. Pos-sui também muitas mitocôndrias e está associada à reab-sorção de cálcio e magnésio, apresentando, em estudoshistoquímicos, uma elevada reatividade imunológicapara uma proteína carreadora de cálcio, vitamina D-de-pendente. A porção mais distal do túbulo distal está re-presentada pelo túbulo conector e a primeira porção doducto coletor, habitualmente referido como túbulo cole-tor inicial (v. Cap. 4).
O túbulo conector é uma região de transição e parece estarenvolvido com a secreção de potássio, pelo menos em parte,regulada por mineralocorticóides, e na secreção de íons H.
Ducto Coletor
Deriva-se do broto ureteral. De acordo com a localiza-ção no rim, costuma-se dividir o ducto coletor (DC) em trêssegmentos: segmento coletor cortical, segmento medularinterno e externo. O segmento coletor cortical está forma-do no começo pelo túbulo coletor inicial e, depois, conti-nua com uma porção arqueada e medular. O segmentomedular interno termina na papila.5
Fig. 1.14 Microscopia óptica do ducto coletor (CD), segmentodelgado da alça de Henle (TL) e algumas arteríolas retas (V).(Obtido de Berman, I. Color Atlas of Basic Histology, 2nd Edition,Appleton & Lange, 1998.69)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 48/121
16 Anatomia Renal
A célula mais abundante no ducto coletor é uma célulaclara, contendo um núcleo central cercado por um citoplas-ma claro e um pequeno número de mitocôndrias (Fig. 1.14).
Um outro tipo de célula encontrado é uma célula escu-ra ou intercalada: citoplasma escuro com numerosas mi-tocôndrias. Estudos imuno-histoquímicos demonstraramaltos níveis de atividade da anidrase carbônica nessas cé-lulas, sugerindo que elas estejam envolvidas no processode acidificação da urina.
As funções do ducto coletor são muitas, embora às ve-zes seja difícil separá-las das funções do túbulo contorna-do distal. Juntos, ducto coletor e túbulo contornado distalformam o néfron distal, onde vários processos fisiológicosocorrem: reabsorção de bicarbonato, secreção de hidrogê-nio, reabsorção e secreção de potássio, secreção de amô-nia, reabsorção de água, etc. Evidência experimental do-cumenta nitidamente que todo ducto coletor reabsorveágua sob a influência de vasopressina (Fig. 1.15).
Na presença de vasopressina, sendo a água reabsorvi-da do interior do ducto coletor, há uma maior concentra-ção de uréia no interior do ducto coletor, cujos segmentoscortical e medular são impermeáveis à uréia. Os segmen-tos medular interno e papilar são permeáveis à uréia, faci-litando a passagem desta para o interstício medular, fatomuito importante no mecanismo de concentração de uri-na (v. Cap. 9). Além disso, há evidência de que o DC par-ticipa da reabsorção de cloreto de sódio, secreção ou reab-sorção de potássio, secreção de íons hidrogênio e do pro-cesso de acidificação urinária, como já foi citado.
INTERSTÍCIO RENAL
O interstício renal engloba tudo o que se encontra noespaço extravascular e intertubular do rim, estando limi-tado pelas membranas basais dos vasos e túbulos. Segun-do Lemley e Kriz,63 o interstício não se constitui de um sim-ples espaço com elementos celulares e uma matriz extra-celular que envolve as “estruturas funcionais dos rins”,néfrons e túbulos. Há evidências de que ele não só fornecesuporte estrutural, mas também funciona como mediador,ou mais exatamente como modulador de quase todas astrocas que ocorrem ao longo dos vasos capilares e túbulosdo parênquima renal. Considera-se provável sua influên-cia na filtração glomerular, através de seus efeitos no feed-back túbulo-glomerular. Ele também tem muita importân-cia no crescimento e na diferenciação das células do parên-quima renal, bem como na determinação da distribuição
da microvasculatura peritubular e na circulação linfática.Além disso, produz fatores autacóides e hormônios de açãolocal, como a adenosina e a prostaglandina, e sistêmica,como a eritropoetina. Alterações no interstício renal con-tribuem para as manifestações clínicas da doença renal.
O interstício renal divide-se nos compartimentos corti-cal e medular, que por sua vez têm suas subdivisões. Nocórtex têm-se as partes peritubular, periarterial e especial,formada pelo mesângio glomerular e extraglomerular. Namedula observam-se as faixas externa e interna da medu-lar externa e a medular interna. Na região periarterial dointerstício cortical encontram-se os vasos linfáticos renais,particularmente abundantes ao redor das artérias arquea-das e corticais radiais ou interlobulares. Eles possuem umendotélio perfurado e sem membrana basal. Não existemvasos linfáticos na medula renal.
O volume do interstício em relação ao parênquima vaiaumentando em direção à papila renal, a partir do córtex.Assim, temos um volume relativo intersticial de 30 a 40%na medula interna de rins de animais de laboratório, en-quanto a parte intersticial cortical tem apenas 7 a 9%. Emrins de adultos jovens normais o volume relativo do inters-tício varia de 5 a 10% no córtex e aumenta com a idade.64
No córtex, identificam-se basicamente dois tipos de cé-lulas intersticiais. O tipo mais freqüente assemelha-se a fi- broblastos, e o outro lembra células mononucleares (macró-fagos). A produção de adenosina por células semelhantes afibroblastos da parte cortical inibe a liberação de renina ediminui a reabsorção de sódio, tendo-se revelado parte do
mecanismo de proteção renal frente a situações de hipóxia.Durante a hipóxia, há evidências de aumento de adenosinae de eritropoetina. Sugere-se que a síntese desta última éestimulada pela adenosina, representando a resposta celu-lar a um sinal de diminuição do O2 disponível.
Na medula, especialmente na medula interna, as célu-las intersticiais são numerosas e vários tipos foram identi-ficados. Através de microscopia eletrônica identificaram-
Fig. 1.15 Representação esquemática do nefro procurando sali-entar as diferenças morfológicas e funcionais da porção inicial edistal do túbulo distal. Observem que o túbulo contornado dis-tal é impermeável à água, como o ramo ascendente da alça deHenle. A porção distal do TD (túbulo coletor) responde ao HAD,como todo o ducto coletor.
Isotônico
Hipertônico
Túbulo contornado distalTúbulo coletor
Epitélio permeável à água
Epitélio impermeável à água
Epitélio permeável à águasomente em presença de HAD

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 49/121
capítulo 1 17
se, inclusive, partículas de gordura em determinadas cé-lulas, muito abundantes nessa região. Por meio de reaçõeshistoquímicas, revelou-se que estas partículas são compos-
tas de ácidos graxos saturados e insaturados. Esses ácidossão precursores de prostaglandinas, formando assim aevidência de que essas células intersticiais medulares es-tejam envolvidas na síntese de prostaglandinas renais, sen-do a medula o principal sítio de produção. Evidenciou-setambém que elas participam da síntese de glicosaminogli-canos presentes na matriz do interstício e que têm umafunção endócrina anti-hipertensiva.65
As células mononucleares têm a capacidade de fagoci-tose e estão freqüentemente associadas às células dendrí-ticas intersticiais, que não se diferenciam claramente dascélulas semelhantes a fibroblastos e funcionam como ex-celentes apresentadoras de antígenos, como se observouem trabalhos experimentais.66 Em humanos, as célulashomólogas a essas células dendríticas intersticiais encon-
tram-se mais no parênquima, como células endoteliais, eexpressam o antígeno comum leucocitário CD45.67
REFERÊNCIAS BIBLIOGRÁFICAS
1. CHAPMAN, W.H.et al. The Urinary System. An Integrated Approach.W.B. Saunders Co., 1973.
2. HEPTINSTALL, R.H. Pathology of the Kidney, 2nd edition. Little, Bro-wn and Co., cap. 1, 1974.
3. PITTS, R.F. Physiology of the Kidney and Body Fluids. Year Book Me-dical Publishers, 1972.
4. NETTER, F.H. Anatomia, estrutura e embriologia. Seção I: rins, ure-teres e bexiga. Ciba-Geigy e Guanabara Koogan, vol. 6, 1973.
5. TISHER, C.C. & MADSEN, K.M. Anatomy of the kidney. In Brenner,B.M. & Rector Jr, F.C. The Kidney. W.B. Saunders Co., p. 3, 1986.
6. WALD, H. The weight of normal adult human kidneys and itsvariability. Arch. Pathol. Lab. Med., 23:493-500, 1937.7. EMERY, J.L.; MITHAL, A. The weight of kidneys in late intra-uterine
life and childhood. J. Clin. Pathol., 13:490-3, 1960.8. KASISKE, B.L.; UMEN, A.J. The influence of age, sex, race and body
habitus on kidney weight in humans. Arch. Pathol. Lab. Med., 110 :55-60, 1986.
9. SYKES, D. The morphology of renal lobulations and calyces, andtheir relationship to partial nephrectomy. Br. J. Surg., 51:294-304,1964.
10. INKE, G. The Protolobar Structure of the Human Kidney: Its Biologic andClinical Significance. New York, Alan R. Liss, 1988.
11. HODSON, C.J. Reflux nephropathy: A personal historical review. Am. J. Roentgenol., 137:451-62, 1981.
12. GARDNER, GRAY, O’RAHILLY. Anatomia do Corpo Humano — Es-tudo Regional, 4-ª edição. Ed. Guanabara Koogan, Caps. 37 e 38, 1988.
13. OSATHANONDH, V.; POTTER, E.L. Development of humankidney as shown by microdissection: II. Renal pelvis, calyces, and
papillae. Arch. Pathol., 76:277-89, 1963.14. OSATHANONDH, V.; POTTER, E.L. Development of human kid-ney as shown by microdissection: III. Formation and interrelati-onships of collecting tubules and nephrons. Arch. Pathol ., 76:290-302,1963.
15. CLAPP, W.L.; ABRAHAMSON, D. Development and gross ana-tomy of the kidney. In Tisher, C.C. & Brenner, B.M. Renal Pathology,2nd edition. Philadelphia, J.B. Lippincott Company, pp. 3-59, 1994.
16. DUNNIL, M.S.; HALLEY, W. Some observations on the quantitati-ve anatomy of the kidney. J. Pathol., 110:113-21, 1973.
Pontos-chave:
• O aparelho justaglomerular éprincipalmente formado pelas célulasgranulares da arteríola aferente (secretorasda renina) e pela mácula densa(diferenciação celular do túbulo distal). Estaestrutura é a principal responsável pelocontrole do sistema renina-angiotensina-aldosterona (SRAA), o qual tem comofunção a regulação do metabolismo desódio
• A estenose de artéria renal diminui o fluxoglomerular, atuando diretamente noaparelho justaglomerular. Ocorre, então,uma estimulação do SRAA, o qual leva a
um quadro de hipertensão arterial sistêmicade causa renovascular
• O túbulo proximal é responsável pelareabsorção da maioria dos pequenossolutos filtrados, e dentre eles temos osíons sódio, cloreto, potássio, cálcio e
bicarbonato, assim como moléculas deaminoácidos e glucose. A água épermeável neste segmento, sendoreabsorvida passivamente. Uma disfunçãohereditária ou adquirida no túbuloproximal leva à síndrome de Fanconi
• A alça de Henle possui grande importância
na concentração da urina, participando nacriação do mecanismo de contracorrenteatravés da criação de um interstíciomedular hipertônico
• Os túbulos distais, junto com os ductoscoletores, formam os néfrons distais.Nestes segmentos agem a aldosterona(reabsorção de sódio e secreção depotássio), o hormônio antidiurético(reabsorção de água) e o fator natriuréticoatrial (inibe reabsorção de sódio). Alémdisto, o ducto coletor tem papel importantena secreção de ácido através do amônio eno mecanismo de contracorrente com auréia
• A nefrite intersticial é um quadro deinflamação aguda do interstício renalprovocada principalmente por drogas,como derivados da penicilina eantiinflamatórios não-esteroidais

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 50/121
18 Anatomia Renal
17. BANKIR, B.; de ROUFFIGNAC, C. Urinary concentrating ability:Insights from comparative anatomy. Am. J. Physiol., 249 (RegulatoryInteg. Comp. Physiol., 18):R643-66, 1985.
18. TISHER, C.C.; BRENNER, B.M. Structure and function of the
glomerulus. In Renal Pathology, 2nd edition. Philadelphia, J.B. Lippin-cott Company, pp. 143-61, 1994.
19. OSTERBY, R. Morphometric studies of the peripheral glomeru-lar basement membrane in early juvenile diabetes: Developmentof initial basement membrane thickening. Diabetologia, 8:84-92,1972.
20. STEFFES, M.W.; BARBOSA, J.; BASGEN, J.M.; SUTHERLAND,D.E.R.; NAJARIAN, J.S.; MAUER, S.M. Quantitative glomerularmorphology of the normal human kidney. Lab. Invest., 49:82-6,1983.
21. KASHTAN, C.E.; MICHAEL, A.F.; SIBLEY, R.K.; VERNIER, R.L.Hereditary nephritis — Alport syndrome and thin glomerular basement disease. In Tisher, C.C. & Brenner, B.M. Renal Patholo- gy, 2nd edition. Philadelphia, J.B. Lippincott Company, p. 1250,1994.
22. WEBER, S.; ENGEL, J.; WIEDEMANN, H.; GLANVILLE, R.W.; TIMPL,R. Subunit structure and assembly of the globular domain of basementmembrane collagen type IV. Eur. J. Biochem., 139:401-10, 1984.
23. BOYD, C.D.; TOTH-FEJEL, S.; GADI, I.K.; LITT, M.; CONDON,M.R.; KOLBE, M.; HAGEN, I.K.; KURKINEN, M.; MACKENZIE, J.W.; MAGENIS, E. The genes coding for human pro alpha 1(IV)and pro alpha 2(IV) collagen are both located at the end of thelong arm of the chromossome 13. Am. J. Hum. Genet., 42:309-14,1988.
24. MORRISON, K.E.; GERMINO, G.G.; REEDERS, S.T. Use of the po-lymerase chain reaction to clone and sequence a cDNA encoding the bovine alpha-3 chain of the type IV collagen. J. Biol. Chem., 266 :34-9,1991.
25. MARIYAMA, M.; KALLURI, R.; HUDSON, B.J.; REEDERS, S.T. Thealpha-4(IV) chain of basement membrane collagen: isolation ofcDNAs encoding bovine alpha-4(IV) and comparison with othertype of collagens. J. Biol. Chem., 67:1253-8, 1991.
26. HOSTIKKA, S.L.; EDDY, R.L.; BYERS, M.G.; HOYHTYA, M.;SHOWS, T.B.; TRYGGVASON, K. Identification of a distinct typeIV collagen alpha chain with restricted kidney distribution andassignment of its gene to the locus of X chromossome-linked Alport
syndrome. Proc. Natl. Acad. Sci. USA, 87:1606-10, 1990.27. KLEPPEL, M.M.; KASHTAN, C.E.; BUTKOWSKI, R.J.; FISH, A.J.;MICHAEL, A.F. Alport familial nephritis — absence of 28 kilodaltonnon-collagenous monomers of type IV collagen in glomerular basement membrane. J. Clin. Invest., 80:263-6, 1987.
28. ANTIGNAC, C.; DECHENES, G.; GROS, F.; KNEBELMANN, B.;TRYGGVASON, K.; GUBLER, M.C. Mutations in the COL4A5 genein Alport syndrome. J. Am. Soc. Nephrol., 2:249, 1991.
29. BRAZEL, D.; OBERBAUMER, I.; DIERINGER, H.; BABEL, W.;GLANVILLE, R.W.; DEUTZMANN, R.; KUHN, K. Completion ofthe amino acid sequence of the alfa-1 chain of human basementmembrane collagen (type IV) reveals 21 non-triplet interruptionslocated within the collagenous domain. Eur. J. Biochem., 168:529-36,1987.
30. TSILIBARY, E.; CHARONIS, A. The role of the main noncol-lagenous domain (NC1) in type IV collagen assembly. J. Cell. Biol.,103:2467-73, 1986.
31. AUMAILLEY, M.; TIMPL, R. Attachment of cells to basement mem- brane collagen type IV. J. Cell Biol., 103:1569-75, 1986.
32. MARTINEZ-HERNANDEZ, A.S.; GAY, S.; MILLER, E.J. Ultrastruc-tural localization of type V collagen in rat kidney. J. Cell Biol., 92 :343-9, 1982.
33. MADRI, J.A.; ROLL, F.J.; FURTHMAYR, H.; FOIDART, J.M. Ultras-tructural localization of fibronectin and laminin in basement mem- branes of murine kidney. J. Cell Biol., 86:682-7, 1980.
34. KATZ, A.; FISH, A.J.; KLEPPEL, M.M.; HAGEN, S.G.; MICHAEL,A.F.; BUTKOWSKI, R.J. Renal entactin (nidogen): isolation, charac-terization and tissue distribution. Kidney Int., 40:643-52, 1991.
35. FARQUHAR, M.G. The glomerular basement membrane — Aselective macromolecular filter. In Hay, E.D. Cell Biology of Extracellular Matrix. New York, Plenum Press, p. 335-78, 1981.
36. KANVAR, Y.S.; LINKER, A.; FARQUHAR, M.G. Increased perme-
ability of the glomerular basement membrane to ferritin after remo-val of glycosaminoglycans (heparan sulfate) by enzyme digestion. J. Cell Biol., 86:688-93, 1980.
37. ROSENZWEIG, L.J.; KANVAR, Y.S. Removal of sulfated(heparan sulfate) or nonsulfated (hyaluronic acid) glycosamino-glycans results in increased permeability of the glomerular basement membrane to 123I-bovine serum albumin. Lab. Invest.,47:177-84, 1982.
38. BOHRER, M.P.; BAYLIS, C.; HUMES, H.D.; GLASSOCK, R.J.; RO-BERTSON, C.R.; BRENNER, B.M. Permselectivity of the glomeru-lar capillary wall: Facilitated filtration of circulating polycations. J.Clin. Invest., 61:72-8, 1978.
39. COUSER, W.G.; HOYER, J.R.; STILMANT, M.M.; JERMANOVICH,N.B.; BELOCK, S. Effect of aminonucleoside nephrosis on immunecomplex localization in autologous immune complex nephritis in therat. J. Clin. Invest., 61:561-72, 1978.
40. HORVAT, R.; HOVOKA, A.; DEKAN, G.; POCZEWSKI, H.;KERJASCHKI, D. Endothelial cell membranes contain podocalyxin
— the major sialoprotein of visceral glomerular epithelial cells. J. CellBiol., 102:484-91, 1986.41. DRENCKHAHN, D.; SCHNITTLER, H.; NOBILING, R.; KRIZ, W.
Ultrastructural organization of contractile proteins in rat glomeru-lar mesangial cells. Am. J. Pathol., 137:1343-52, 1990.
42. GEHLSEN, K.R.; DILLNER, L.; ENGVALL, E.; RUOSLAHTI, E. Thehuman laminin receptor is a member of the integrin family of celladhesion receptors. Science, 241:1228-9, 1988.
43. LAITINEN, L.; VARTIO, T.; VIRTANEN, I. Cellular fibronectins aredifferentially expressed in human fetal and adult kidney. Lab. Invest.,64:492-8, 1991.
44. SCHLONDORFF, D. The glomerular mesangial cell — An expan-ding role for a specialized pericyte. FASEB J. 1:272-81, 1987.
45. KRIZ, W.; ELGER, M.; LEMLEY, K.; SAKAI, T. Structure of the glo-merular mesangium: A biomechanical interpretation. Kidney Int.,38(Suppl 30):S2-9, 1990.
46. SCHNABEL, E.; ANDERSON, J.M.; FARQUHAR, M.G. The tight junction protein ZO-1 is concentrated along the slit diaphragms ofthe glomerular epithelium. J. Cell Biol., 111:1255-63, 1990.
47. SCHNEEBERGER, E.E.; LEVEY, R.H.; McCLUSKEY, R.I.;KARNOVSKY, M.J. The isoporous substructure of the human glo-merular slit diaphragm. Kidney Int., 8:48-52, 1975.
48. KAZATCHKINE, M.D.; FEARON, D.T.; APPAY, M.D.; MANDET,C.; BARIETY, J. Immunohistochemical study of the human glome-rular C3b receptor in normal kidney and in 75 cases of renal disea-ses. J. Clin. Invest., 69:900-12, 1982.
49. KERJASCHKI, D.; FARQUHAR, M.G. Immunocytochemical loca-lization of the Heymann antigen (gp 330) in glomerular epithelialcells of normal Lewis rats. J. Exp. Med., 157:667-86, 1983.
50. KERJASCHKI, D.; SHARKEY, D.J.; FARQUHAR, M.G. Identifica-tion and characterization podocalyxin — the major sialoprotein ofrenal glomerular epithelial cell. J. Cell Biol., 98:1591-6, 1984.
51. KANVAR, Y.S.; FARQUHAR, M.G. Detachment of the endotheliumand epithelium from the glomerular basement membrane produced by kidney perfusion with neuraminidase. Lab. Invest., 42:375-84,1980.
52. KURTZ, S.M.; FELDMAN, J.D. Experimental studies on the
formation of the glomerular basement membrane. J. Ultrastr. Res.,6:19-27, 1962.
53. GOMEZ, R.A.; PUPILLI, C.; EVERETT, A.D. Molecular aspects ofrenin during kidney ontogeny. Pediatr. Nephrol., 5:80-7, 1991.
54. THURAU et al. Composition of tubular fluid in the macula densasegment as a factor regulating the function of the juxtaglomerularapparatus. Circ. Res., 20:suppl. 2:79, 1967.
55. TOBIAN et al. Correlation between granulation of juxtaglomerularcells and extractable renin in rats with experimental hypertension.Proc. Soc. Exp. Biol. Med., 100:94, 1959.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 51/121
capítulo 1 19
56. BARAJAS, L. Renin secretion: an anatomical basis for tubular con-trol. Science, 172:485, 1971.
57. RHODIN, J.A.G. Structure of the kidney. InDisease of the Kidney, 2nd
edition. Little, Brown and Co., 1971.
58. ROCHA, A.S. and KOKKO, J.P. Sodium chloride and water trans-port in the medullary thick ascending limb of Henle: evidence foractive chloride transport. J. Clin. Invest., 52:612, 1973.
59. WOODHALL, P.B. and TISHER, C.C. Response of the distal tubuleand cortical collecting duct to vasopressin in the rat. J. Clin. Invest.,47:203, 1968.
60. MADSEN, K.M., BRENNER, B.M. Structure and function of the renaltubule and interstitium. In Tisher, C.C. & Brenner, B.M. Renal Patholo- gy, 2nd edition. Philadelphia, J.B. Lippincott Company, pp. 661-98, 1994.
61. BERGERON, M.; GUERETTE, D.; FORGET, J.; THIÉRY, G. Three-dimentional characteristics of the mitochondria of the rat nephron.Kidney Int., 17:175-85, 1980.
62. ALLEN, F.; TISHER, C.C. Morphology of the ascending thick limbof Henle. Kidney Int., 9:8-22, 1976.
63. LEMLEY, K.V.; KRIZ, W. Anatomy of the renal interstitium. KidneyInt., 39:370-81, 1991.
64. KAPPEL, B.; OLSEN, S. Cortical interstitial tissue and sclerosed glo-meruli in the human kidney related to age and sex: A quantitative
study. Virchows Arch.(A), 387:271-7, 1980.65. MUIRHEAD, E.E.; GERMAIN, G.S.; ARMSTRONG, F.B.; BROOKS,B.; LEACH, B.E.; BYERS, L.W.; PITCOCK, J.A.; BROWN, P. Endo-crine-type antihypertensive function of renomedullary interstitialcells. Kidney Int., 8:S271-82, 1975.
66. GURNER, A.C.; SMITH, J.; CATTEL, V. The origin of Ia antigen-expressing cells in the rat kidney. Am. J. Pathol., 127:169-75, 1984.
67. ALEXPOULOS, E.; SERON, D.; HARTLEY, R.B.; CAMERON, J.S.Lupus nephritis: correlation of interstitial cells with glomerular func-tion. Kidney Int., 37:100-9, 1990.
68. BURKITT, H.G.; YUONG, B.; HELATH, J.W. Wheather’s Functio-nal Histology, 3rd Edition, Churchill Livingston, 1993.
69. BERMAN, I. Color Atlas of Basic Histology, 2nd Edition, Appleton &Lange, 1998.
70. KUMAR, V.; COTRAN, R.; ROBBINS, S.Basic Pathology, 6th Edition,W.B. Saunders Company, 1997.
71. JUNQUEIRA, L.C.; CARNEIRO, J. Histologia Básica , 8-ª Edição, Gua-nabara Koogan, 1995.
ENDEREÇOS RELEVANTES NA INTERNET
http://www.nlm.nih.gov/medlineplus/aboutmedli-neplus.html — site do MEDLINEplus, uma mina de ourode informações na área da saúde da maior biblioteca mé-dica do mundo, a National Library of Medicine.http://www.e-kidneys.net/ — site do ALtruis Biomedi-cal Network, uma colecão de websites com foco na pesqui-sa médica e cuidado da saúde.http://www.mhhe.com/biosci/ap/saladin2e/student/adam/aoa.mhtml — acesso a Anatomia e Fisiologia deKenneth Saladin.http://www.gradesummit.com/studentinfo-q1.htm —um instrumento de auto-avaliacão e preparação para exa-mes. Há uma seção de anatomia e fisiologia.http://www.abdn.ac.uk/~phy050/kidney.htm — é umprograma tutorial desenvolvido com Asymetrix Toolbooke serve como uma introdução e revisão tutorial para estu-dantes de medicina do primeiro ano. Introduz o estudan-te na anatomia e histologia do rim e complementa as au-las.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 52/121
Capítulo
2Circulação Renal
José Luiz Monteiro e Claudia Maria de Barros Helou
INTRODUÇÃO
ANATOMIA VASCULAR RENAL
MEDIDAS DO FLUXO SANGUÍNEO RENALFluxo sanguíneo renal total
Métodos de medida do fluxo sanguíneo renal
Método das microesferas radioativas
Fluxômetro eletromagnético
Técnicas de processamento de imagem
DISTRIBUIÇÃO INTRA-RENAL DO FLUXO SANGUÍNEO
Fluxo sanguíneo cortical
Técnica dos gases inertes
Método das microesferas marcadas com isótopos
radioativos
Fluxo sanguíneo medularDoppler
REGULAÇÃO DA CIRCULAÇÃO RENAL
Inervação renal
Auto-regulação do fluxo sanguíneo renal
Teoria miogênica
Teoria do feedback túbulo-glomerular
Regulação parácrina da microcirculação renal
Mecanismos de ativação em resposta a estímulo
BIBLIOGRAFIA SELECIONADA
pulações de néfrons superficiais e profundos (justamedu-lares), tanto quanto ao fluxo sanguíneo como à taxa de fil-tração glomerular.
Com os avanços recentes através de técnicas de micro-dissecção, conseguiu-se isolar heterogêneas arteríolas afe-rentes e eferentes, como será descrito adiante.
As células endoteliais eram consideradas no passadocomo simples membranas semipermeáveis, que impediama passagem principalmente de proteínas. Atualmente, atu-am como verdadeiros órgãos, dotados de propriedadesmetabólicas autócrinas e parácrinas, isto é, com síntese defatores vasomotores agindo nas próprias células ou nos
tecidos adjacentes. Para que ocorra um verdadeiro equilí- brio na regulação da circulação renal, são evidenciadosvasodilatadores representados pelo óxido nítrico, pelasprostaglandinas e pelo fator hiperpolarizante derivado doendotélio (EDHF) e os vasoconstritores, endotelina e trom- boxane. Estes agonistas, uma vez liberados pelo endotélio,exercem a sua função na musculatura lisa das arteríolasrenais.
INTRODUÇÃO
Os rins humanos pesam cerca de 300 g, representandoaproximadamente 0,5% do peso corpóreo. Apesar deste baixo peso, recebem de 20 a 25% do débito cardíaco, cor-respondendo a 400 ml de fluxo por 100 g de tecido renalpor minuto, 5 a 50 vezes maior que o de outros órgãos tam- bém importantes, coração, cérebro e fígado. Devido à sua baixa resistência vascular, associada à grande capacidadefiltrante, possui, portanto, a maior taxa de perfusão entretodos os tecidos dos mamíferos.
A circulação renal apresenta certas características inte-ressantes: ocorre uma baixa diferença arteriovenosa de oxi-gênio, indicando que o alto fluxo sanguíneo é muito mai-or que sua necessidade metabólica. Por outro lado, em es-tado de choque circulatório sistêmico, uma freqüente com-plicação é a ocorrência de insuficiência renal aguda, mui-to mais comum que lesões no coração, cérebro e fígado.
Outra peculiaridade do rim refere-se às diferentes po-

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 53/121
capítulo 2 21
queadas. A partir destas, formam-se as artérias radiais corti-cais, que se dirigem perpendicularmente ao córtex super-ficial, dividindo-o em lóbulos, e por isso eram antigamen-
te denominadas de artérias interlobulares.Das artérias radiais corticais originam-se as arteríolas afe-
rentes, cuja porção distal penetra na cápsula de Bowmanramificando-se em múltiplos capilares que convergem eformam as arteríolas eferentes (Fig. 2.1). Estas arteríolas sãoimportantes na regulação da resistência vascular glomeru-lar por apresentarem estruturas esfinctéricas modulando,então, a hemodinâmica renal e a filtração glomerular.
Essa rede capilar formada no interior da cápsula deBowman tem o endotélio envolvido por prolongamentosdas células epiteliais da cápsula, os podócitos. Além dascélulas endoteliais e epiteliais, há um terceiro tipo de célu-las que são responsáveis pela matriz e denominadas decélulas mesangiais. As células mesangiais, além da impor-tante participação no arcabouço glomerular, também de-
sempenham papel na regulação da filtração glomerulardevido à presença de elementos contráteis que induzemvariações das áreas filtrantes. A esse conjunto de estrutu-ras vasculares, epiteliais e mesangiais é dado o nome de glomérulo.
Os diâmetros glomerulares são heterogêneos ao longodo córtex renal, sendo maiores os justamedulares em rela-ção aos superficiais, correspondendo também a uma mai-or filtração por cada unidade funcional renal, o néfron.
As arteríolas aferentes, que são os vasos pré-glomerula-res, caracterizam-se por apresentarem parede espessa eregular devido à distribuição homogênea das fibras circu-lares de músculo liso independentemente de sua localiza-
Pontos-chave:
• Os rins são órgãos de baixa resistência
vascular cujo fluxo sanguíneo corrigido porgrama de tecido é o maior do organismo.
• A circulação renal não é homogênea.• As células endoteliais sintetizam e/ou
liberam agonistas que modulam atonicidade da musculatura lisa dasarteríolas renais.
ANATOMIA VASCULAR RENAL
As artérias renais usualmente são únicas, dividindo-se junto ao hilo em um ramo anterior e outro posterior. O ramoanterior se divide em quatro artérias segmentares respon-
sáveis pela irrigação de todo o pólo inferior, do ápice e dossegmentos superior e médio da face anterior renal. Os seg-mentos restantes são irrigados pelo ramo posterior. Nãoexistem anastomoses entre estes ramos iniciais da artériarenal, subentendendo-se daí que a obstrução de qualquerdeles levará à isquemia de todo o tecido para o qual o flu-xo sanguíneo se distribui.
Essas artérias segmentares dividem-se em várias outrasque se dirigem até a junção córtico-medular, delimitandoespaços denominados de lobos, e por isso elas recebem onome de artérias interlobares.
Na região córtico-medular, as artérias interlobares assu-mem forma encurvada, originando-se então as artérias ar-
Fig. 2.1 Fotomicrografia dos vasos do córtex renal humano, fixados com silicone. As f lechas indicam as artérias radiais corticais (in-terlobulares), perpendiculares à superfície renal, e os glomérulos são visíveis como pequenos objetos arredondados. (Obtido deBrenner, B.M. et al. The renal circulations. In: Brenner, B.M. and Rector, F.C., Jr. (eds). The Kidney, 4th ed. W.B. Saunders Company,Philadelphia, 1991, p. 165.)
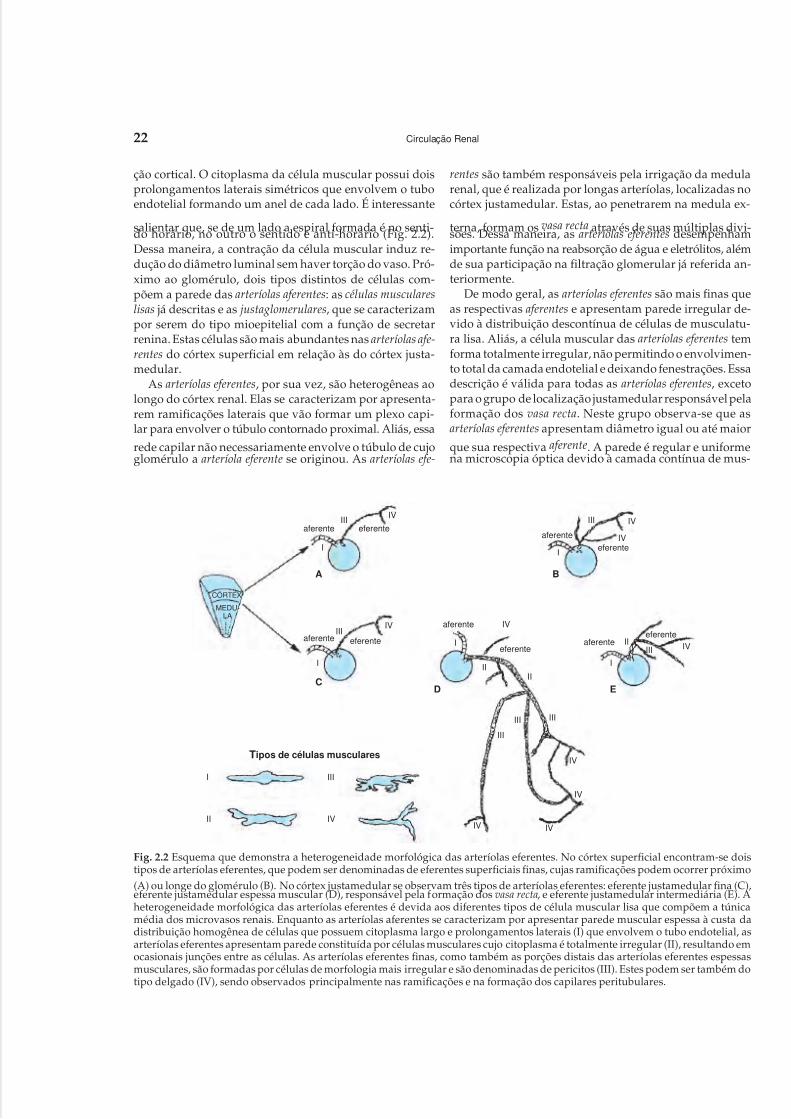
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 54/121
22 Circulação Renal
ção cortical. O citoplasma da célula muscular possui doisprolongamentos laterais simétricos que envolvem o tuboendotelial formando um anel de cada lado. É interessante
salientar que, se de um lado a espiral formada é no senti-do horário, no outro o sentido é anti-horário (Fig. 2.2).Dessa maneira, a contração da célula muscular induz re-dução do diâmetro luminal sem haver torção do vaso. Pró-ximo ao glomérulo, dois tipos distintos de células com-põem a parede das arteríolas aferentes: as células musculareslisas já descritas e as justaglomerulares, que se caracterizampor serem do tipo mioepitelial com a função de secretarrenina. Estas células são mais abundantes nas arteríolas afe-rentes do córtex superficial em relação às do córtex justa-medular.
As arteríolas eferentes, por sua vez, são heterogêneas aolongo do córtex renal. Elas se caracterizam por apresenta-rem ramificações laterais que vão formar um plexo capi-lar para envolver o túbulo contornado proximal. Aliás, essa
rede capilar não necessariamente envolve o túbulo de cujoglomérulo a arteríola eferente se originou. As arteríolas efe-
rentes são também responsáveis pela irrigação da medularenal, que é realizada por longas arteríolas, localizadas nocórtex justamedular. Estas, ao penetrarem na medula ex-
terna, formam os vasa recta através de suas múltiplas divi-sões. Dessa maneira, as arteríolas eferentes desempenhamimportante função na reabsorção de água e eletrólitos, alémde sua participação na filtração glomerular já referida an-teriormente.
De modo geral, as arteríolas eferentes são mais finas queas respectivas aferentes e apresentam parede irregular de-vido à distribuição descontínua de células de musculatu-ra lisa. Aliás, a célula muscular das arteríolas eferentes temforma totalmente irregular, não permitindo o envolvimen-to total da camada endotelial e deixando fenestrações. Essadescrição é válida para todas as arteríolas eferentes, excetopara o grupo de localização justamedular responsável pelaformação dos vasa recta. Neste grupo observa-se que asarteríolas eferentes apresentam diâmetro igual ou até maior
que sua respectiva aferente. A parede é regular e uniformena microscopia óptica devido à camada contínua de mus-
Fig. 2.2 Esquema que demonstra a heterogeneidade morfológica das arteríolas eferentes. No córtex superficial encontram-se doistipos de arteríolas eferentes, que podem ser denominadas de eferentes superficiais finas, cujas ramificações podem ocorrer próximo(A) ou longe do glomérulo (B). No córtex justamedular se observam três tipos de arteríolas eferentes: eferente justamedular fina (C),eferente justamedular espessa muscular (D), responsável pela formação dos vasa recta, e eferente justamedular intermediária (E). Aheterogeneidade morfológica das arteríolas eferentes é devida aos diferentes tipos de célula muscular lisa que compõem a túnicamédia dos microvasos renais. Enquanto as arteríolas aferentes se caracterizam por apresentar parede muscular espessa à custa dadistribuição homogênea de células que possuem citoplasma largo e prolongamentos laterais (I) que envolvem o tubo endotelial, asarteríolas eferentes apresentam parede constituída por células musculares cujo citoplasma é totalmente irregular (II), resultando emocasionais junções entre as células. As arteríolas eferentes finas, como também as porções distais das arteríolas eferentes espessasmusculares, são formadas por células de morfologia mais irregular e são denominadas de pericitos (III). Estes podem ser também dotipo delgado (IV), sendo observados principalmente nas ramificações e na formação dos capilares peritubulares.
IIIIV
I
IVIII
I
I
IV
IIII
III III
III
III
III
IV
IV
IV
I
IV
IV
IVIV
III
IV
I
II
eferente
eferente
eferente
eferente
eferente
aferente
aferente
aferente
aferente
IIaferente
Tipos de células musculares
A B
CD E
CÓRTEX
MEDU-LA
I

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 55/121
capítulo 2 23
culatura lisa e é somente pela presença de ramificações quese permite distingui-las das aferentes. Entretanto, na mi-croscopia eletrônica também se observam fenestrações na
parede muscular, uma vez que a irregularidade dos pro-longamentos laterais dessas células não permite a forma-ção de um anel contínuo muscular sobre o tubo endotelial(Fig. 2.2).
No córtex superficial as arteríolas eferentes são sempre fi-nas (16 a 18 µm de diâmetro no rim do rato) e de parede ir-regular. Entretanto, elas podem mostrar padrão heterogê-neo quanto ao local da ramificação. Algumas se ramificam bem próximas ao glomérulo e em outras as ramificações sóocorrerão a partir de 100 a 200 m. Ao local onde ocorremas ramificações é dado o nome de “welling point”, ou “vasoestrelado”, como preferiam os micropuncionadores.
No córtex intermediário, as arteríolas eferentes também
são finas e de parede irregular, mas elas são extremamen-te curtas devido às múltiplas ramificações para formar oplexo capilar que envolve o túbulo contornado proximal.
Esta rede vascular é tão complexa que impede distinguiro caminho individual de um capilar.No córtex profundo ou justamedular, também se obser-
vam arteríolas eferentes finas com parede irregular e rami-ficações laterais situadas longe do glomérulo e portanto
Pontos-chave:
• As artérias renais são únicas e se dividemsucessivamente até a formação doglomérulo (a. renal a. segmentar a.interlobar a. arqueada a. radial cortical arteríola aferente)
• Devido à ausência de anastomoses entre asmúltiplas divisões da artéria renal, aobstrução de uma dessas divisões ocasionaisquemia parcial do órgão
• As arteríolas aferentes apresentam o mesmopadrão morfológico por todo o córtex renal
• As arteríolas eferentes apresentamheterogeneidade morfológica ecaracterizam-se pela presença deramificações laterais que formam os plexoscapilares que envolvem os túbulos. Nocórtex justamedular, as a. eferentes espessasmusculares penetram na medula e formamos vasa recta através de múltiplas divisõeslongitudinais
• As arteríolas eferentes participam docontrole da filtração glomerular, dairrigação medular e da reabsorção de água eeletrólitos através da formação dos plexos
capilares e dos vasa recta• O sangue retorna à circulação através dos
vasa recta ascendentes, de anastomosesvenosas entre os capilares peritubulares e asveias na região cortical que drena para v.interlobulares v. interlobares v. renal v. cava inferior
Fig. 2.3 Representação esquemática da organização microvascu-lar e tubular do rim de cão. Notam-se três tipos de néfrons comseus glomérulos situados no córtex externo, médio e interno. Àdireita, há a divisão do córtex (C), medula externa (OM) e medu-la interna (IM). À esquerda, notam-se o glomérulo (G), arteríolasaferentes (AA) e os vasos eferentes (EV), dos quais formam-se oscapilares peritubulares. Na superfície renal, os túbulos contorna-dos proximais (PCT) são vistos ao lado de densa rede capilarperitubular (retângulo 1). No córtex médio e interno, os túbulosproximais localizam-se ao lado de artérias interlobulares e redecapilar adjacentes (retângulos 2 e 4). Vêem-se também ductoscoletores (CD), entre vasos interlobulares e alças de Henle. Oscapilares peritubulares desta região, derivados de arteríolas efe-rentes do córtex médio, orientam-se paralelamente às estruturastubulares do raio medular (retângulo 3). No córtex interno ou
justamedular, os glomérulos têm arteríolas eferentes que se diri-gem para baixo, dividindo-se para formar uma extensa rede vas-cular da medula externa. Em direção à medular interna, há a for-mação dos vasa recta, ao lado de alças de Henle e ductos coleto-res. (Obtido de Beeuwkes, R. e Bonventre, J.V. Tubular organi-zation and vascular tubular relations in the dog kidney. Am. J. Phy-siol., 229:695, 1975.)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 56/121
24 Circulação Renal
com aspecto morfológico semelhante ao daquelas do cór-tex superficial. Como já foi referido em parágrafos anteri-ores, nesta região se localizam as arteríolas eferentes espes-
sas musculares (diâmetro de 23,0 1,5 m em ratos) quese dirigem à medula para formar os vasa recta. Além disso,nesta região também se reconhece um outro tipo de arterí-ola eferente de diâmetro (19,3 0,5m) e morfologia inter-mediários entre as eferentes finas e as espessas musculares.
Do plexo capilar oriundo dos vasa recta descendentesformam-se a circulação venosa e os vasa recta ascendentes.Esses capilares, além de suprirem as necessidades meta- bólicas locais, são também responsáveis pela captação eremoção de água extraída dos ductos coletores durante oprocesso de formação da urina. Para manter a tonicidadedo interstício, o fluxo sanguíneo medular desempenhaimportante função na formação de gradiente de solutos. Arepresentação esquemática desta microcirculação é mos-trada na Fig. 2.3.
Anastomoses venosas entre capilares peritubulares eveias são encontradas na região cortical. A circulação ve-nosa inicia-se então através das veias corticais superficiaisque formam as veias interlobulares. Estas, na região córtico-medular, originam as veias arqueadas, saindo destas as vei-as interlobares, que vão formar finalmente a veia renal prin-cipal, saindo do hilo renal em direção à veia cava inferior.
MEDIDAS DO FLUXOSANGUÍNEO RENAL
Fluxo Sanguíneo Renal Total
Como foi dito anteriormente, o fluxo sanguíneo renalcorresponde a 1/4 do débito cardíaco, ou seja, em torno de1.200 ml/min no homem adulto. É um pouco maior no sexomasculino, e em crianças de até um ano de idade corres-ponde à metade dos valores do adulto, índice que alcançaao redor dos três anos de vida. A partir da terceira décadacomeça a decrescer, chegando à metade dos valores nor-mais aos 80 anos. Baseado no peso renal, o fluxo sanguí-neo renal total é aproximadamente de 4 ml/min/g de te-cido. O fluxo cortical é cerca de duas vezes e meia maiorem relação ao medular.
Métodos de Medida do Fluxo
Sanguíneo RenalO método mais utilizado em clínica baseia-se na aplica-
ção do princípio de Fick. Se uma substância não é sinteti-zada nem metabolizada dentro do rim, a sua passagempelo órgão e posterior aparecimento na urina poderá sercalculada através da equação:
Ux Vu (Ax Vx) FPR (1)
onde Ux concentração do indicador x na urina; Vu fluxo urinário; Ax e Vx concentração do indicador naartéria e veia renal, respectivamente; e FPR fluxo plas-
mático renal. Portanto:
FPRUx Vu
Ax 2 Vx
(2)
Esta remoção da substância do sangue arterial renal édesignada de extração renal do referido indicador. O maisutilizado é o ácido paraminoipúrico (PAH), o qual é ativa-mente secretado pelos tú bulos proximais. Sua extração écerca de 0,7 a 0,9 em humanos. Com infusão constanteexógena do PAH, em clínica assume-se este valor igual a 1e, nestas condições, a equação (2) representaria o clearancede PAH. É importante salientar que este método somenteé válido quando a substância é administrada continuamen-te, mantido um bom fluxo urinário, e a mesma não seja
sintetizada nem metabolizada pelos rins.O fluxo sanguíneo renal (FSR) pode ser calculado atra-vés da correção pelo hematócrito (Ht):
FSR FPR/1 Ht
MÉTODO DAS MICROESFERAS RADIOATIVASÉ utilizado especificamente para condições experimen-
tais.Microesferas são partículas plásticas de dimensões uni-
formes de 15 5 m de diâmetro, com propriedades quí-micas inertes e densidade específica muito próxima à dosangue. Possuem a vantagem de poderem ser marcadascom isótopos radioativos e ser extraídas pelo leito capilarde um órgão, distribuindo-se de acordo com o fluxo san-
guíneo do mesmo. No rim, são captadas pelas arteríolas oucapilares glomerulares, sem alterar a hemodinâmica local.
Quando injetadas no ventrículo esquerdo ou aorta, dis-tribuem-se homogeneamente por toda a circulação. Aquantidade de microesferas que atinge o rim, ou seja, amedida da radioatividade renal total (Qt), é proporcionalao fluxo sanguíneo renal total (FSR), assim como a radioa-tividade por minuto de amostra de sangue coletada poraspiração na artéria femoral durante a administração dasmicroesferas (qt) é proporcional ao fluxo sanguíneo (ml/min) na artéria femoral (Ff) coletado por bomba de aspira-ção contínua. Assim:
FSR Qt Ff/qt
FLUXÔMETRO ELETROMAGNÉTICOPara medidas diretas do fluxo sanguíneo renal total, estemétodo oferece a vantagem de monitorização contínua dataxa de perfusão do rim. Baseia-se na implantação de ele-trodos circulares ao redor da artéria renal, sendo captadasondas magnéticas oriundas do volume líquido em movi-mento nas mesmas, registrando-se os valores em velocida-de do fluxo sanguíneo renal. Pode ser utilizado em condi-

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 57/121
capítulo 2 25
ções experimentais e mesmo no homem, quando em cirur-gias com acesso às artérias renais.
TÉCNICAS DE PROCESSAMENTO DE IMAGEMBaseiam-se no método de processamento eletrônico da
imagem microscópica capilar, com a determinação da ve-locidade do eritrócito e o tempo necessário para percorreruma distância conhecida. Medidas simultâneas do diâme-tro do vaso e o hematócrito determinam o fluxo sanguíneoneste vaso. Através de filmagem de vídeo, as imagenspodem ser transferidas para um sistema com avaliação emmaior grau do fluxo arterial e, por conseguinte, o fluxosanguíneo renal. A região medular é mais bem examina-da por este método. A principal crítica a esta técnica refe-
re-se à distribuição preferencialmente axial das hemácias,ocorrendo, portanto, superestimação do fluxo sanguíneocapilar.
Pontos-chave:
• Diversos métodos vêm sendo utilizadospara medida do fluxo sanguíneo renal totalem pesquisa clínica e/ou experimental
• O método do clearance de PAH permiteestimar o fluxo sanguíneo renal total emhumanos
DISTRIBUIÇÃO INTRA-RENALDO FLUXO SANGUÍNEO
Fluxo Sanguíneo CorticalA distribuição intra-renal do fluxo sanguíneo e a fun-
ção renal parecem não se correlacionar com os métodos deestudo até o momento realizados. Exemplo neste sentidosão os resultados, principalmente após trauma e hemorra-gia, com as diferentes taxas de perfusão nas regiões inter-nas do rim. Como foi dito anteriormente, existem hetero-gêneas populações de néfrons, não só quanto à sua anato-mia microvascular, como também quanto à função eà taxade perfusão de cada uma.
TÉCNICA DOS GASES INERTESBaseia-se na administração endovenosa ou diretamen-
te em artéria renal de um marcador (criptônio ou xenônio)com captação externa em região lombar com detectorcintilográfico da passagem pelo rim deste marcador. Ten-ta correlacionar os vários componentes de uma curvamultiexponencial, obtida com o detector, com as diversasregiões córtico-medulares, baseado em comparações auto-radiográficas. São descritos quatro componentes: cortical,medular externo, medular interno e perirrenal e gordura
hilar. Devido a várias dificuldades técnicas relacionadas aeste método, como distribuição do gás no tecido renal e ofato de a medida do fluxo ser dada por volume, pratica-
mente não está mais sendo utilizado.
MÉTODO DAS MICROESFERAS MARCADASCOM ISÓTOPOS RADIOATIVOS
É o método mais utilizado experimentalmente. Asmicroesferas marcadas com isótopos são injetadas direta-mente no ventrículo esquerdo ou aorta, distribuindo-separa todos os órgãos de forma homogênea, sendo propor-cional ao fluxo sanguíneo de cada um. No rim ficam im-pactadas nas arteríolas ou capilares glomerulares.
A medida do fluxo sanguíneo para diferentes regiões dorim pode ser determinada através de cortes paralelos, nosentido horizontal da superfície para o córtex mais inter-no. Com a retirada de um fragmento do córtex, seccionan-
do-se três fatias paralelas, da superfície externa para a in-terna, de igual espessura, designamos, respectivamente,zona 1 o córtex externo, zona 2 o córtex médio e zona 3 ocórtex interno. Determinando-se a radioatividade e o pesode cada zona, a porcentagem de fluxo sanguíneo de cadazona (Pz) é calculada pela fórmula:
Pz qz/qt
onde qz é a radioatividade (contagens) por minuto porgrama de uma determinada zona do córtex renal e qt é aradioatividade das três zonas (qz1 qz2 qz3).
As críticas a este método relacionam-se, na distribuiçãoaxial das microesferas, à semelhança das hemácias. Destamaneira, em vasos menores, tipo artéria interlobular, suaconcentração mediana poderia estar superestimando o flu-
xo sanguíneo cortical superficial, local de maior popula-ção destas artérias e das arteríolas aferentes.
Fluxo Sanguíneo Medular
A circulação medular provém das arteríolas pós-glome-rulares dos néfrons justamedulares.
O fluxo medular, mesmo sendo menor que o cortical,assemelha-se ao de outros órgãos. Característica importan-te é a baixa pressão parcial de oxigênio nesta região, emtorno de 10 a 20 mmHg, ao contrário da cortical, cerca de50 mmHg. Esta hipóxia medular é considerada como ten-do papel fisiológico para que ocorra uma eficiente concen-tração urinária. Se o fluxo sanguíneo for excessivo, rompe
o gradiente osmolar do interstício, alterando o mecanismode contracorrente multiplicador. Se for baixo, ocorre lesãoisquêmica das células tubulares. Pré-requisito crítico paraque a urina se concentre devidamente é uma exata perfu-são de oxigênio e a demanda, através de um preciso equi-lí brio entre a regulação do fluxo sanguíneo medular e otrabalho tubular.
As técnicas de medida do fluxo sanguíneo medular ne-

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 58/121
26 Circulação Renal
cessitam de estudos em conjunto, ou seja, avaliação do flu-xo dos néfrons justamedulares combinados com estudosanatômicos detalhados da região medular. Os mais utili-
zados são os realizados através de indicadores não-difusí-veis, tais como albumina marcada com 131I, eritrócitos mar-cados com 32P e rubídio radioativo.
DOPPLERO princípio do Doppler é utilizado na transmissão do som
do sangue fluindo através dos vasos numa freqüência queé captada por um transdutor. As imagens detectadasquantificam o fluxo sanguíneo correspondente ao vaso es-pecífico, indicando também a direção do respectivo fluxo.
A maior importância deste método aplica-se nos estu-
dos de anastomoses de vasos em transplante renal, tantoartérias como veias, identificando-se possíveis estenoses ouoclusões.
Pontos-chave:
• A distribuição do fluxo sanguíneo éheterogênea no rim, sendo que 80% destefluxo destina-se à região cortical
• A medula renal apresenta baixa pressãoparcial de oxigênio
REGULAÇÃO DACIRCULAÇÃO RENAL
Os vasos renais possuem musculatura lisa em várias
camadas, porém a partir das arteríolas aferentes elas serestringem a uma única camada.
A vasoconstrição ou dilatação arteriolar manifesta-sedependendo de fatores físicos intra-renais, humorais eneurogênicos agindo na arteríola aferente e/ou eferente.Este aumento ou diminuição da resistência vascular alte-ra tanto a filtração glomerular como o fluxo sanguíneo re-nal, desde que a pressão de perfusão não se altere.
A Fig. 2.4 ilustra as várias mudanças que ocorrem nofluxo sanguíneo e na filtração glomerular quando a resis-tência é alterada.
Inervação Renal
No rim são encontradas terminações nervosas simpáti-cas ao longo das arteríolas aferentes e eferentes até o com-plexo justaglomerular. Através de microscopia eletrônica,revelou-se a presença de vesículas granulares em nervosrenais, que são típicas de fibras adrenérgicas, e vesicularesagranulares, provavelmente de natureza colinérgica. Entreos tú bulos renais, existem também ocasionalmente nervos,que podem influenciar os processos de reabsorção tubular.
À estimulação do nervo renal, ocorre imediatamentecontração da musculatura lisa dos vasos, com conseqüen-te queda do fluxo sanguíneo. Este efeito pode ser minimi-
zado com pequenas doses de noradrenalina e totalmenteabolido com agentes bloqueadores alfa-adrenérgicos. Sãoencontrados também receptores beta-adrenérgicos e dopa-minérgicos.
O efeito da estimulação do nervo renal sobre a micro-circulação renal mostra um aumento na resistência arteri-olar aferente e eferente, com grande redução do fluxo san-guíneo glomerular, com semelhantes alterações no coefi-ciente de ultrafiltração do capilar glomerular e vasos peri-tubulares.
Auto-Regulação do FluxoSanguíneo Renal
A maioria dos órgãos são capazes de manter o seu flu-xo sanguíneo quando ocorrem alterações da pressão deperfusão.
O fenômeno da auto-regulação no rim é demonstradocom variações da pressão arterial entre 80 e 180 mmHg. Umaumento da pressão de perfusão é acompanhado por umequivalente aumento da resistência vascular, tornando-seinalterado o fluxo sanguíneo renal total.
A auto-regulação persiste mesmo após denervação re-nal, em rim isolado e perfundido in vitro com plasma e apósretirada da medula adrenal (que previne a produção decatecolaminas). Portanto, auto-regulação é um fenômenointrínseco que ocorre dentro do rim e só não está presente
Fig. 2.4 Efeito das alterações da resistência das arteríolas aferen-te e eferente sobre o fluxo sanguíneo renal (FSR) e filtração glo-merular (TFG), mantendo-se constante a pressão de perfusão.
RESISTÊNCIA NAS ARTERÍOLAS FSR TFG
Af Ef
CONTROLE
REDUÇÃO NAAFERENTE
AUMENTO NAAFERENTE
REDUÇÃO NAEFERENTE
AUMENTO NAEFERENTE

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 59/121
capítulo 2 27
quando existem grandes alterações da pressão de perfu-são arterial.
É importante salientar que a auto-regulação também se
aplica à taxa de filtração glomerular, de tal maneira que nocaso de alterações mais profundas da pressão de perfusão,por exemplo quando da administração do vasodilatadorpapaverina, ocorre abolição do efeito da auto-regulaçãotanto do fluxo como da filtração.
As teorias envolvidas, muito provavelmente em com- binação, no processo da auto-regulação são: miogênica e feedback tú bulo-glomerular.
TEORIA MIOGÊNICASegundo esta teoria, a musculatura lisa arterial contrai-
se e relaxa-se em resposta a um aumento ou redução datensão na parede vascular, respectivamente. Perante umaelevação abrupta da pressão de perfusão, há um aumento
do raio do vaso. Entretanto, quase imediatamente, a mus-culatura lisa se contrai, permitindo que o fluxo sanguíneose mantenha constante. O contrário existe quando há umaqueda da pressão de perfusão. Crê-se atualmente que omediador deste processo de relaxamento e constrição vas-cular seja a entrada de cálcio nas células musculares lisas
dos vasos. Nas situações de aumento de pressão intravas-cular, o estímulo mecânico exercido na parede do vasodeflagra a despolarização da membrana da célula muscu-lar lisa. Os canais de cálcio operados por voltagem (VOCC)se abrem, permitindo a entrada de cálcio do extracelularpara o intracelular. A elevação deste íon no citosol deflagraa fosforilação das pontes de miosina, resultando na con-tração da célula muscular.
O mecanismo miogênico baseia-se na lei de Laplace,
pela equação:
T R/(Pi Pe)
onde T é a tensão na parede do vaso, R é o seu raio inter-no, Pi é a pressão hidrostática intravascular e Pe é a pres-são hidrostática extravascular. O gradiente de pressãotransmural (PiPe) reduzindo-se, diminuiria a tensão naparede e a resistência vascular. Quando a pressão de per-fusão renal cai, reduz-se também a pressão transmural e atensão na parede do vaso, e a resistência na arteríola afe-rente diminui igualmente para manter relativamente cons-tante o fluxo sanguíneo renal.
Convém também lembrar que a regulação miogênica sóocorre nos vasos pré-glomerulares, ou seja, ao nível daartéria interlobular e principalmente da arteríola aferente.
Teoria do Feedback Túbulo-Glomerular
O mecanismo tú bulo-glomerular na auto-regulação dofluxo sanguíneo renal envolve também a taxa de filtraçãoglomerular. Sugere-se que, quando ocorre uma elevaçãoda pressão arterial, há um aumento do fluxo sanguíneo
renal e da pressão hidráulica do capilar glomerular. Estasalterações causam um aumento na taxa de filtração glome-rular, elevando-se o fluxo de fluido ao tú bulo distal. O
aumento de oferta de fluido a este segmento sensibilizariaa mácula densa, que ativaria mecanismos efetores, aumen-tando a resistência pré-glomerular, reduzindo o fluxo san-guíneo renal, a pressão glomerular e, por conseguinte, ataxa de filtração glomerular.
O principal soluto envolvido nesta resposta da máculadensa alterando o tônus da musculatura lisa das arterío-las aferentes talvez seja o cloreto de sódio. Algumas evi-dências experimentais foram demonstradas, principalmen-te após perfusão intratubular em velocidades crescentes deNaCl. Entretanto, o verdadeiro mecanismo efetor não estáesclarecido, podendo ser através da ativação do sistemarenina-angiotensina, da adenosina, do ATP ou de algumprostanóide não-ciclooxigenase.
Convém salientar que tanto a resposta miogênica como
o feedback tú bulo-glomerular são indispensáveis para queocorra a auto-regulação renal. Estes dois mecanismos nãosão apenas aditivos, pois existe uma interação complexapara que a auto-regulação ocorra em sua eficiência máxima.
Regulação Parácrina daMicrocirculação Renal
Além da ação dos hormônios circulantes, a microcircu-lação renal pode ser controlada néfron a néfron através deagonistas liberados pelo endotélio, pelo epitélio ou pelointerstício. Esta regulação local recebe o nome de regula-ção parácrina. Entre os vários sistemas que exercem estafunção, podemos citar: 1) sistema renina-angiotensina in-
tra-renal, 2) mediadores purinérgicos, 3) metabólitos doácido araquidônico, 4) agonistas liberados ou sintetizadospelo endotélio (endotelina, óxido nítrico, fator hiperpola-rizante derivado do endotélio).
Atualmente, reconhecem-se dois sistemas renina-angi-otensina, que podem ser denominados como circulante eintra-renal. No primeiro, a síntese de angiotensina II é re-alizada pela ação integrada do rim, fígado e endotélio pul-monar. No segundo, a angiotensina II é formada localmen-te no rim. Em ambas as situações a angiotensina II exerceação de constricção dos vasos pré- e pós-glomerulares. Háindícios de que esta ação seja preferencial nas arteríolasaferentes em relação às eferentes no córtex superficial. Poroutro lado, estudos vêm demonstrando ação semelhante
entre as arteríolas aferentes e eferentes no córtex justame-dular. Assim, a regulação da microcirculação renal feitapela angiotensina II não é homogênea, como se pensoudurante tantos anos. E o conhecimento da regulação pará-crina permite compreender melhor a heterogeneidade queexiste na microcirculação renal.
Os compostos purinérgicos, em especial o ATP, vêmsendo citados como importantes reguladores parácrinos.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 60/121
28 Circulação Renal
A ação do ATP ou da adenosina seria através da ativaçãodos receptores P2 presentes apenas nas arteríolas aferen-tes induzindo a vasoconstrição. A hipótese aventada é de
que grandes quantidades de ATP seriam liberadas pelamácula densa em resposta a um aumento do aporte deNaCl aos segmentos distais do néfron. Assim, o ATP seriao mediador parácrino do feedback tú bulo-glomerular. Umdos argumentos para esta hipótese é devido ao fato de as
células da mácula densa serem ricas em mitocôndrias e dea atividade da Na-K-ATPase ser baixa em relação à quan-tidade de ATP que é gerada nestas células.
Importante ação parácrina é exercida pelos metabólitosdo ácido araquidônico que são também conhecidos comoeicosanóides. Estes metabólitos apresentam importanteação reguladora principalmente ao nível da arteríola afe-rente mediando tanto a vasoconstrição quanto a vasodila-tação. Atualmente são reconhecidas três vias enzimáticas:a da ciclooxigenase (COX), a da lipooxigenase e a do cito-cromo P-450 (CYP450). Os eicosanóides podem ser origi-nários das células endoteliais, epiteliais ou intersticiais.Entre os mediadores da vasoconstrição podemos citar otromboxane, os leucotrienes e os ácidos hidroxieicosatetra-enóicos (HETEs). Na vasodilatação geralmente são descri-tas as ações das prostaglandinas PGE
2e PGI
2, como tam-
bém as dos ácidos epoxieicotrienóicos (11,12-EET).E por fim, outro importante sistema na regulação pará-
crina é representado pelos agonistas e/ou metabólitos que
são gerados ou liberados pelo endotélio. Na vasoconstri-ção, as endotelinas exercem importante ação tanto nas ar-teríolas aferentes quanto nas eferentes. Na vasodilatação,o óxido nítrico, a bradicinina e o fator hiperpolarizantederivado do endotélio (EDHF) são os mediadores. A ação
do óxido nítrico é evidente nas arteríolas aferentes masdiscutível nas eferentes. Provavelmente, o não-reconheci-mento de que grupo morfológico a arteríola eferente estu-dada pertença seja responsável pelos resultados contradi-tórios. Assim, o óxido nítrico exerce possivelmente ação
vasodilatadora nas arteríolas eferentes que formam os vasarecta e não tem ação nos outros grupos morfológicos. Maisuma vez, o conhecimento da regulação parácrina permitecompreender melhor a heterogeneidade que existe na mi-crocirculação renal.
Mecanismos de Ativação emResposta a Estímulo
Além da já mencionada heterogeneidade morfológicaexistente entre as arteríolas glomerulares, a microcircula-ção renal é dotada de distintos mecanismos de ativação emresposta a estímulo mecânico ou induzido por agonistas.
Assim, no córtex superficial, canais de cálcio operadospor voltagem (VOCC) participam dos mecanismos para aresposta vascular apenas nas arteríolas aferentes. Então, oaumento do cálcio citosólico ([Ca2]i) ocorre principalmen-
te pela abertura dos VOCC presentes na membrana damusculatura lisa. Entretanto, outros mecanismos como aliberação do cálcio estocado nas organelas também contri-
buem para elevação da [Ca2
]i. As arteríolas eferentes docórtex superficial não possuem VOCC e a sinalização viacálcio é feita preferencialmente por outros mecanismos deentrada deste cátion do extracelular para o intracelular,como também pela liberação deste íon estocado nas orga-nelas citoplasmáticas. No córtex justamedular, os canais decálcio operados por voltagem estão presentes tanto nasarteríolas aferentes como também nas arteríolas eferentesespessas musculares que são responsáveis pela formaçãodos vasa recta. Então, a regulação da microcirculação renaldeve ser heterogênea, ou seja, os fatores que influenciamo córtex superficial não necessariamente influenciam a re-gião medular e vice-versa.
Pontos-chave:• A circulação renal é regulada pelas
terminações simpáticas presentes nasarteríolas glomerulares e pela ação deagonistas circulantes ou sintetizadoslocalmente pelo endotélio, pelo epitélio oupelo interstício (regulação parácrina)
• O rim possui um sistema de auto-regulaçãode fluxo sanguíneo a fim de que estepermaneça constante independente davariação da pressão arterial. A auto-regulação renal ocorre quando a pressãoarterial está entre 80 e 180 mmHg
• A resposta miogênica exercida pelos vasospré-glomerulares e o feedback tú bulo-glomerular são os fatores determinantespara que ocorra a auto-regulação do fluxosanguíneo renal
• A microcirculação renal pode ser reguladalocalmente, néfron a néfron, através deagonistas parácrinos: a) sistema renina-angiotensina, b) mediadores purinérgicos,c) metabólitos do ácido araquidônico, d)agonistas liberados ou sintetizados peloendotélio (endotelina, óxido nítrico, fatorhiperpolarizante derivado do endotélio)
BIBLIOGRAFIA SELECIONADA
ARENDSHORST, W.J. and NAVAR, L.G. Renal Circulation and Glomeru-lar Hemodynamics. In: Schrier, R.W. and Gottschalck C.W. (eds). Disea-ses of the Kidney, 6th ed. Little, Brown and Company, pp. 59-106, 1996.
BREZIS, M. and ROSEN, S. Hypoxia of the renal medulla — Its implica-tions for disease. The New England Journal of Medicine, 332:647-655, 1995.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 61/121
capítulo 2 29
DWORKIN, L.D., SUN A.M. and BRENNER, B.M. The renal circulations.In: Brenner, B.M. and Rector, F.C., Jr. (eds). The Kidney, 6 th ed. W.B.Saunders Company, Philadelphia, pp. 277-318, 2000.
EVAN, A.P. and DAIL, W.G., Jr. Efferent arterioles in the cortex of the
rat kidney. Anat. Rec., 187:135-145, 1977.GATTONE II, V.H.; LUFT, F.C. and EVAN, A.P. Renal afferent and
efferent arterioles of the rabbit. American Journal of Physiology, 247:F219-F228, 1984.
NAVAR, LG. Integrating multiple paracrine regulators of renal micro-vascular dynamics. American Journal of Physiology, 274:F433-F444, 1998.
VALTIN, H. and SCHAFER, J.A. Renal hemodynamics and oxygen con-sumption. In: Valtin, H. and Schaffer, J.A. (eds). Renal Function, 3rd ed.
Little, Brown and Company, pp. 95-114, 1995.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 62/121
Capítulo
3Filtração Glomerular
Antonio Carlos Seguro e Luis Yu
DETERMINANTES DA FILTRAÇÃO GLOMERULAR
FILTRAÇÃO GLOMERULAR POR NEFRO
REGULAÇÃO HORMONAL DA FILTRAÇÃO GLOMERULARPERMEABILIDADE SELETIVA GLOMERULAR
Os rins recebem normalmente 20% do débito cardíaco,o que representa um fluxo sanguíneo de 1.000 a 1.200 ml/min para um homem de 70-75 kg. Este alto fluxo é aindamais significativo se considerado pelo peso dos rins, cercade 300 gramas. Assim, o fluxo sanguíneo por grama de rimé de cerca de 4 ml/min, um fluxo 5 a 50 vezes maior que
em outros órgãos. Este sangue que atinge o rim passa ini-cialmente pelos glomérulos, onde cerca de 20% do plasmaé filtrado, totalizando uma taxa de filtração glomerular de120 ml/min ou 170 litros/dia. Os estudos de micropunçãomostraram que o líquido filtrado tem composição iônica ede substâncias cristalóides (glicose, aminoácidos etc.) idên-tica ao plasma, porém sem a presença de elementos figu-rados do sangue (hemácias, leucócitos, plaquetas) e comquantidades mínimas de proteínas e macromoléculas,constituindo-se, portanto, em um ultrafiltrado do plasma.
DETERMINANTES DAFILTRAÇÃO GLOMERULAR
A passagem de água e moléculas através do capilar glo-merular é governada pelas mesmas forças que atuam emqualquer outro capilar do organismo.
Tomando-se um determinado ponto do capilar glome-rular, o ritmo de ultrafiltração (Jv) neste local é dado pelaequação:
Jv K (P )
HIPERFILTRAÇÃO GLOMERULAR
MEDIDA DA FILTRACÃO GLOMERULAR
BIBLIOGRAFIA SELECIONADAENDEREÇO RELEVANTE NA INTERNET
onde K é o coeficiente de permeabilidade hidráulica docapilar glomerular; P é a diferença entre a pressão hidros-tática do capilar glomerular (Pcg) e a pressão hidrostáticado fluido da cápsula de Bowman, que é igual à pressãointratubular (PT); é a diferença entre a pressão oncóti-ca do capilar glomerular (cg), que é uma força que se opõe
à ultrafiltração, e a pressão oncótica do fluido da cápsulade Bowman, esta última igual a zero, uma vez que este flui-do é um ultrafiltrado, portanto, isento de proteínas. Assim,a equação pode ser estendida para:
Jv K (Pcg PT cg),
onde Pcg PT cg é igual à pressão de ultrafiltração(Puf).
Com a descoberta de uma raça mutante de ratos Wistar(ratos Wistar de Munique), que apresentam glomérulos nasuperfície renal, portanto, acessíveis à micropunção, foipossível fazer medidas diretas da pressão capilar glome-rular e estimar todos os determinantes da ultrafiltração.
Desta forma, a pressão capilar glomerular, em condi-ções de hidropenia, tem um valor de 45 mmHg e se man-
tém praticamente constante ao longo do capilar glomeru-lar. A pressão intratubular é em torno de 10 mmHg. Apressão oncótica no início do capilar glomerular é de 20mmHg, sendo igual à pressão oncótica da artéria renal. Àmedida que vai havendo saída de água ao longo do capi-lar glomerular, aumenta a concentração de proteína intra-capilar, traduzindo-se por uma pressão oncótica mais ele-vada (Fig. 3.1). A determinação direta da pressão oncóti-

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 63/121
capítulo 3 31
ca do capilar glomerular ao nível da arteríola eferente,através de ultramicrométodo, revela uma pressão em tor-no de 35 mmHg.
A pressão de ultrafiltração pode, então, ser calculada emdois pontos:
Puf no início do capilar glomerular 45 mmHg
10 mmHg 20 mmHg 15 mmHg.
Puf no fim do capilar glomerular
45 mmHg
10mmHg 35 mmHg 0 mmHg.
A esta condição observada em ratos e macacos, em quea pressão de ultrafiltração chega a zero no fim do capilarglomerular, chama-se de equilíbrio de pressão de filtração.
A pressão de filtração, nesta condição de equilíbrio, nãopode ser calculada, pois poderia ser 0 em qualquer ponto
intermediário do capilar glomerular. A Fig. 3.2 mostra duasdas infinitas possibilidades de valores da Puf na condiçãode equilíbrio.
Pontos-chave:
• A pressão capilar glomerular é uma forçaque favorece a filtração glomerular
• A pressão intratubular e a pressão oncóticado capilar glomerular são forças que seopõem à filtração
• A filtração glomerular depende dapermeabilidade do capilar glomerular
FILTRAÇÃO GLOMERULAR
POR NEFROConsiderando-se a filtração glomerular de um único
glomérulo (RFGn), pode-se escrever:
RFGn Kf Puf
onde Kf, o coeficiente de permeabilidade glomerular, é igualao produto de k e S, sendo k o coeficiente de permeabilida-de hidráulica do capilar glomerular, anteriormente descri-to, e S é a área, ou superfície filtrante de todo o glomérulo.
Vários estudos mostraram que a filtração glomerularpor nefro nos ratos Wistar é altamente dependente do flu-xo plasmático glomerular, isto é, o aumento do fluxo plas-mático glomerular leva ao aumento da filtração glomeru-lar por aumento da pressão de ultrafiltração, deslocando
o ponto de equilíbrio para mais próximo do fim do capilarglomerular, como, por exemplo, na Fig. 3.2, levando dacondição A para a condição B.
Através de infusões endovenosas isoncóticas de plasmaem ratos, pode-se aumentar o fluxo plasmático glomeru-lar a níveis três vezes maiores que o normal, até um pontoem que a pressão oncótica não se iguala à pressão hidros-
Fig. 3.1 Determinantes da pressão de ultrafiltração. Representa-
ção esquemática de um capilar glomerular. Pcg é a pressão hi-drostática do capilar glomerular, constante ao longo de toda suaextensão. Pt é a pressão intratubular e cg é a pressão oncóticadas proteínas do capilar glomerular, que aumenta progressiva-mente ao longo do capilar, à medida que a água vai sendo filtra-da, concentrando-se as proteínas.
Fig. 3.2 Equilíbrio da pressão de filtração. Em abscissa está representada a distância do capilar glomerular. Zero corresponde aoinício do capilar, e 1, ao fim. Em ordenadas, os valores de pressão em mmHg. A diferença de pressão hidrostática (∆p) é praticamen-te constante ao longo do capilar. A diferença de pressão oncótica (∆π) aumenta progressivamente. A pressão de ultrafiltração (Puf)é representada pela área entre as duas curvas. Os gráficos A e B representam duas das infinitas possibilidades de valores de Puf emcondição de equilíbrio de filtração. Em ambas (A e B), se iguala a P antes do fim do capilar glomerular.
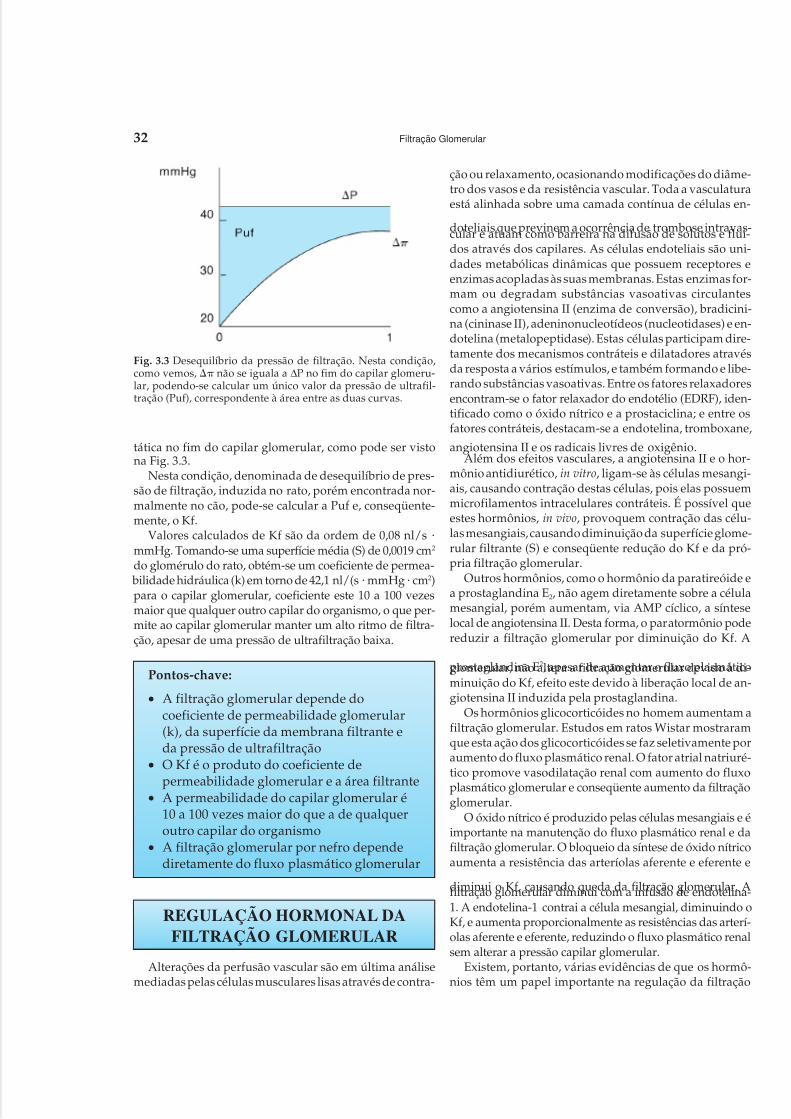
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 64/121
32 Filtração Glomerular
tática no fim do capilar glomerular, como pode ser vistona Fig. 3.3.
Nesta condição, denominada de desequilíbrio de pres-são de filtração, induzida no rato, porém encontrada nor-malmente no cão, pode-se calcular a Puf e, conseqüente-mente, o Kf.
Valores calculados de Kf são da ordem de 0,08 nl/s
mmHg. Tomando-se uma superfície média (S) de 0,0019 cm2
do glomérulo do rato, obtém-se um coeficiente de permea- bilidade hidráulica (k) em torno de 42,1 nl/(s mmHg cm2)para o capilar glomerular, coeficiente este 10 a 100 vezesmaior que qualquer outro capilar do organismo, o que per-mite ao capilar glomerular manter um alto ritmo de filtra-ção, apesar de uma pressão de ultrafiltração baixa.
Pontos-chave:
• A filtração glomerular depende docoeficiente de permeabilidade glomerular(k), da superfície da membrana filtrante eda pressão de ultrafiltração
• O Kf é o produto do coeficiente depermeabilidade glomerular e a área filtrante
• A permeabilidade do capilar glomerular é10 a 100 vezes maior do que a de qualqueroutro capilar do organismo
• A filtração glomerular por nefro dependediretamente do fluxo plasmático glomerular
REGULAÇÃO HORMONAL DAFILTRAÇÃO GLOMERULAR
Alterações da perfusão vascular são em última análisemediadas pelas células musculares lisas através de contra-
ção ou relaxamento, ocasionando modificações do diâme-tro dos vasos e da resistência vascular. Toda a vasculaturaestá alinhada sobre uma camada contínua de células en-
doteliais que previnem a ocorrência de trombose intravas-cular e atuam como barreira na difusão de solutos e flui-dos através dos capilares. As células endoteliais são uni-dades metabólicas dinâmicas que possuem receptores eenzimas acopladas às suas membranas. Estas enzimas for-mam ou degradam substâncias vasoativas circulantescomo a angiotensina II (enzima de conversão), bradicini-na (cininase II), adeninonucleotídeos (nucleotidases) e en-dotelina (metalopeptidase). Estas células participam dire-tamente dos mecanismos contráteis e dilatadores atravésda resposta a vários estímulos, e também formando e libe-rando substâncias vasoativas. Entre os fatores relaxadoresencontram-se o fator relaxador do endotélio (EDRF), iden-tificado como o óxido nítrico e a prostaciclina; e entre osfatores contráteis, destacam-se a endotelina, tromboxane,
angiotensina II e os radicais livres de oxigênio.Além dos efeitos vasculares, a angiotensina II e o hor-
mônio antidiurético, in vitro, ligam-se às células mesangi-ais, causando contração destas células, pois elas possuemmicrofilamentos intracelulares contráteis. É possível queestes hormônios, in vivo, provoquem contração das célu-las mesangiais, causando diminuição da superfície glome-rular filtrante (S) e conseqüente redução do Kf e da pró-pria filtração glomerular.
Outros hormônios, como o hormônio da paratireóide ea prostaglandina E2, não agem diretamente sobre a célulamesangial, porém aumentam, via AMP cíclico, a sínteselocal de angiotensina II. Desta forma, o paratormônio podereduzir a filtração glomerular por diminuição do Kf. A
prostaglandina E2, apesar de aumentar o fluxo plasmáticoglomerular, não altera a filtração glomerular devido à di-minuição do Kf, efeito este devido à liberação local de an-giotensina II induzida pela prostaglandina.
Os hormônios glicocorticóides no homem aumentam afiltração glomerular. Estudos em ratos Wistar mostraramque esta ação dos glicocorticóides se faz seletivamente poraumento do fluxo plasmático renal. O fator atrial natriuré-tico promove vasodilatação renal com aumento do fluxoplasmático glomerular e conseqüente aumento da filtraçãoglomerular.
O óxido nítrico é produzido pelas células mesangiais e éimportante na manutenção do fluxo plasmático renal e dafiltração glomerular. O bloqueio da síntese de óxido nítricoaumenta a resistência das arteríolas aferente e eferente e
diminui o Kf, causando queda da filtração glomerular. Afiltração glomerular diminui com a infusão de endotelina-1. A endotelina-1 contrai a célula mesangial, diminuindo oKf, e aumenta proporcionalmente as resistências das arterí-olas aferente e eferente, reduzindo o fluxo plasmático renalsem alterar a pressão capilar glomerular.
Existem, portanto, várias evidências de que os hormô-nios têm um papel importante na regulação da filtração
Fig. 3.3 Desequilíbrio da pressão de filtração. Nesta condição,como vemos, não se iguala a ∆P no fim do capilar glomeru-lar, podendo-se calcular um único valor da pressão de ultrafil-tração (Puf), correspondente à área entre as duas curvas.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 65/121
capítulo 3 33
glomerular e podem também estar envolvidos nas altera-ções da filtração glomerular, observados em condiçõespatológicas ou induzidas por drogas.
O uso crônico da gentamicina induz queda da filtraçãoglomerular. Estudos com ratos Wistar mostraram que estaqueda ocorre principalmente devido à redução do Kf, efeitoeste que pode ser atenuado por ingestão de dieta rica emsal, ou pela administração crônica de captopril, situaçõesestas que diminuem a geração de angiotensina II, sugerin-do um papel deste hormônio na insuficiência renal agudanefrotóxica causada por aminoglicosídeos. A ciclosporinadiminui a filtração glomerular por nefro devido ao aumen-to das resistências das arteríolas aferente e eferente comdiminuição do fluxo plasmático glomerular e do Kf.
Em modelos experimentais de obstrução renal parcial,demonstrou-se que a filtração glomerular por nefro pou-co se altera, embora ocorra queda do Kf, e esta é contraba-lançada por aumento do gradiente de pressão hidrostáti-
ca (∆P). Entretanto, se a síntese de prostaglandina for ini- bida pela indometacina, os valores da filtração glomeru-lar por nefro no rim parcialmente obstruído caem intensa-mente, sugerindo que durante a obstrução ureteral parci-al o efeito vasodilatador da prostaglandina antagoniza oefeito vasoconstritor simultâneo, provavelmente da angi-otensina II.
Experimentalmente, tem sido demonstrado que nas le-sões glomerulares primárias há mediação da angiotensi-na II. O aminonucleosídeo puromicina, quando adminis-trado em ratos, causa proteinúria, acompanhada por que-da da filtração glomerular devido principalmente à dimi-nuição do Kf, que pode ser parcialmente revertida pelainfusão de um antagonista da angiotensina II (saralasina).
Em resumo, a filtração glomerular é regulada por umasérie de substâncias vasoativas sistêmicas ou localmentesintetizadas pelas células glomerulares, incluindo-se ascélulas endoteliais e musculares lisas. A célula mesangialpode ser o alvo destas substâncias devido à sua capacida-
de de contração, com conseqüente redução da área filtran-te (S) e do Kf. Estes mecanismos reguladores podem estarafetados e contribuir para a queda da filtração glomerular
observada em doenças renais.
PERMEABILIDADE SELETIVAGLOMERULAR
Os capilares glomerulares permitem a passagem livrede pequenas moléculas como a água, uréia, sódio, clore-tos e glicose; mas não permitem a passagem de moléculasmaiores como eritrócitos ou proteínas plasmáticas. O ca-pilar glomerular comporta-se como uma membrana filtran-te contendo canais aquosos localizados entre as células e amembrana basal do capilar glomerular. Além destes com-ponentes, as células epiteliais com seus podócitos também
fazem parte desta barreira filtrante. Estima-se que o diâ-metro desses canais varie entre 75 e 100 Å devido à per-meabilidade seletiva que eles apresentam.
Vários estudos foram feitos, tanto no homem como emanimais, para se estudar a permeabilidade seletiva do ca-pilar glomerular. A maioria destes estudos foram feitosutilizando-se macromoléculas, como o dextran, uma subs-tância homogênea quanto à estrutura química e formamolecular, porém encontrado em tamanhos diferentes, osquais podem ser utilizados para o estudo da permeabili-dade glomerular.
O dextran, uma vez filtrado, não é reabsorvido nem se-cretado pelos túbulos renais. Pode-se comparar o clearan-ce do dextran com o clearance de inulina, molécula peque-na que é filtrada pelo rim, cuja concentração no fluido dacápsula de Bowman é a mesma do plasma, e também nãoé reabsorvida nem secretada pelos túbulos. Desta forma, arazão entre o clearance do dextran e o clearance de inulina éuma medida indireta da permeabilidade seletiva. Esta ra-zão pode variar de 0 (zero), quando determinada molécu-la de dextran não é filtrada pelo rim, até 1 (um), quando amolécula atravessa livremente o filtro glomerular, como ainulina.
A Fig. 3.4 mostra a variação do clearance fracional dedextran em função do raio da molécula.
Verifica-se que não ocorre qualquer restrição à passa-gem de dextran com raio molecular até 20 Å (clearance fra-cional igual a 1). A partir deste valor, à medida que se au-menta o raio molecular, a molécula vai sendo menos fil-
trada pelo rim até se tornar impermeável (raio de 42 Å).Estes dados não explicam por que uma molécula comoa albumina, de raio molecular de aproximadamente 36 Å,não é filtrada pelo rim, visto que uma molécula de dextrande mesmo raio ainda atravessa o filtro glomerular.
Outros estudos mostraram que a permeabilidade glo-merular não depende só do tamanho da molécula, mastambém da forma, flexibilidade, e especialmente da carga
Pontos-chave:
• A angiotensina II e o hormônioantidiurético promovem contração dascélulas mesangiais e redução do Kf
• A endotelina-1 e o bloqueio do óxido nítricodiminuem o Kf
• O fator atrial natriurético aumenta o fluxoplasmático glomerular
• Os glicocorticóides aumentam o fluxoplasmático glomerular
• A gentamicina diminui o Kf• A ciclosporina diminui o fluxo plasmático
glomerular e o Kf
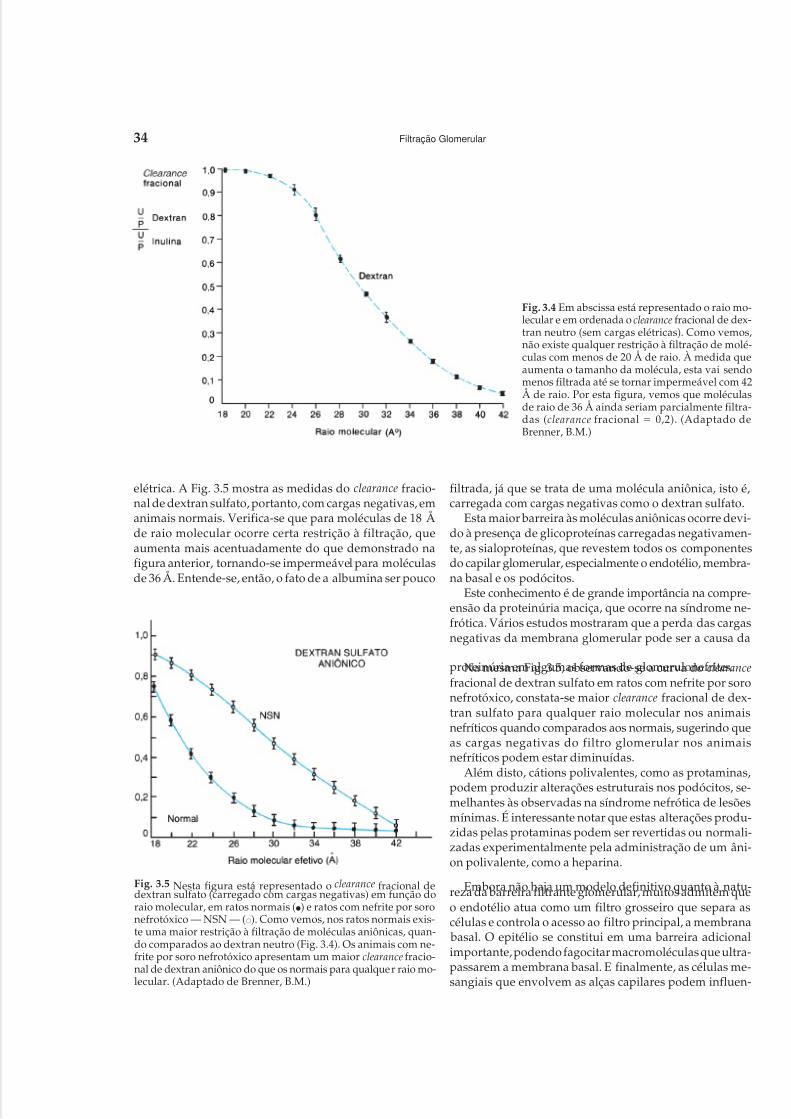
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 66/121
34 Filtração Glomerular
elétrica. A Fig. 3.5 mostra as medidas do clearance fracio-nal de dextran sulfato, portanto, com cargas negativas, emanimais normais. Verifica-se que para moléculas de 18 Åde raio molecular ocorre certa restrição à filtração, queaumenta mais acentuadamente do que demonstrado nafigura anterior, tornando-se impermeável para moléculasde 36 Å. Entende-se, então, o fato de a albumina ser pouco
filtrada, já que se trata de uma molécula aniônica, isto é,carregada com cargas negativas como o dextran sulfato.
Esta maior barreira às moléculas aniônicas ocorre devi-do à presença de glicoproteínas carregadas negativamen-te, as sialoproteínas, que revestem todos os componentesdo capilar glomerular, especialmente o endotélio, membra-na basal e os podócitos.
Este conhecimento é de grande importância na compre-ensão da proteinúria maciça, que ocorre na síndrome ne-frótica. Vários estudos mostraram que a perda das cargasnegativas da membrana glomerular pode ser a causa da
proteinúria em algumas formas de glomerulonefrites.Na mesma Fig. 3.5, observando-se a curva do clearancefracional de dextran sulfato em ratos com nefrite por soronefrotóxico, constata-se maior clearance fracional de dex-tran sulfato para qualquer raio molecular nos animaisnefríticos quando comparados aos normais, sugerindo queas cargas negativas do filtro glomerular nos animaisnefríticos podem estar diminuídas.
Além disto, cátions polivalentes, como as protaminas,podem produzir alterações estruturais nos podócitos, se-melhantes às observadas na síndrome nefrótica de lesõesmínimas. É interessante notar que estas alterações produ-zidas pelas protaminas podem ser revertidas ou normali-zadas experimentalmente pela administração de um âni-on polivalente, como a heparina.
Embora não haja um modelo definitivo quanto à natu-reza da barreira filtrante glomerular, muitos admitem queo endotélio atua como um filtro grosseiro que separa ascélulas e controla o acesso ao filtro principal, a membrana basal. O epitélio se constitui em uma barreira adicionalimportante, podendo fagocitar macromoléculas que ultra-passarem a membrana basal. E finalmente, as células me-sangiais que envolvem as alças capilares podem influen-
Fig. 3.5 Nesta figura está representado o clearance fracional dedextran sulfato (carregado com cargas negativas) em função doraio molecular, em ratos normais () e ratos com nefrite por soronefrotóxico — NSN — (). Como vemos, nos ratos normais exis-te uma maior restrição à filtração de moléculas aniônicas, quan-do comparados ao dextran neutro (Fig. 3.4). Os animais com ne-frite por soro nefrotóxico apresentam um maior clearance fracio-nal de dextran aniônico do que os normais para qualquer raio mo-lecular. (Adaptado de Brenner, B.M.)
Fig. 3.4 Em abscissa está representado o raio mo-lecular e em ordenada o clearance fracional de dex-tran neutro (sem cargas elétricas). Como vemos,não existe qualquer restrição à filtração de molé-culas com menos de 20 Å de raio. À medida queaumenta o tamanho da molécula, esta vai sendomenos filtrada até se tornar impermeável com 42Å de raio. Por esta figura, vemos que moléculasde raio de 36 Å ainda seriam parcialmente filtra-das (clearance fracional 0,2). (Adaptado deBrenner, B.M.)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 67/121
capítulo 3 35
ciar o fluxo plasmático e conseqüentemente a filtração glo-merular devido às suas propriedades contráteis.
Pontos-chave:
• A permeabilidade seletiva da barreiraglomerular depende do tamanho, da formae especialmente da carga da molécula
• A albumina tem raio molecular de 32 Å e émuito pouco filtrada por se tratar demolécula aniônica
• Nas glomerulonefrites a perda das cargasnegativas da membrana glomerularaumenta a filtração de proteínas
HIPERFILTRAÇÃOGLOMERULAR
A redução da massa renal, cirúrgica ou por lesão doparênquima renal, induz aumento da filtração glomerulardos nefros remanescentes, principalmente devido ao au-mento do fluxo plasmático glomerular e do gradiente depressão hidrostática (∆P). O aumento da filtração glome-rular por nefro é tanto maior, quanto maior a redução damassa renal.
A hiperfiltração glomerular é também observada emcrianças e adultos jovens com diabetes mellitus e parece con-tribuir com o início e a manutenção da glomerulopatia fre-qüentemente encontrada na doença. Estudos em ratos com
diabetes induzido pela administração de estreptozocinmostraram que estes animais apresentam aumento da fil-tração glomerular devido ao aumento do fluxo plasmáti-co e da pressão capilar glomerular.
Outro fator que pode levar ao aumento da filtração glo-merular é a ingestão protéica. Ratos mantidos em dieta com35% de proteínas apresentam filtração glomerular 70%maior que animais mantidos com apenas 6% de proteínasna dieta. Este efeito parece ser devido à vasodilatação re-nal induzida pelas proteínas ou aminoácidos. Há evidên-cias recentes sugerindo que este efeito seja mediado vialiberação de óxido nítrico.
Vários estudos sugerem que a hiperfiltração leva, aolongo do tempo, à lesão glomerular com aumento da per-meabilidade glomerular às macromoléculas aniônicas, re-
sultando no aparecimento de proteinúria. Este aumento deproteínas no mesângio serve como estímulo para a proli-feração das células mesangiais e maior produção de ma-triz mesangial, causando a glomeruloesclerose. A esclero-se glomerular reduz ainda mais o número de nefros funci-onantes, com conseqüente maior redução de massa renal,conduzindo a uma progressão inexorável para a insufici-ência renal crônica terminal.
Tem sido demonstrado que a redução da ingesta pro-téica retarda a deterioração da função renal nestas condi-ções, assim como a hiperfiltração do diabetes pode ser
normalizada com um tratamento adequado com insulina.
Pontos-chave:
• Na redução de massa renal, no diabetesmellitus e no aumento da ingestão protéicaocorre hiperfiltração glomerular
• O aumento do fluxo plasmático glomerulare da pressão capilar glomerular são osresponsáveis pelo aumento da filtraçãoglomerular por nefro
MEDIDA DA FILTRAÇÃOGLOMERULAR
A quantidade de plasma filtrado por minuto pode serdeterminada pela depuração plasmática de alguma subs-tância livre no plasma, que não esteja ligada às proteínasplasmáticas, com diâmetro menor que 75 Å, sem cargaselétricas e que passe prontamente pela membrana capilarglomerular. Além disso, não deve ser reabsorvida, secre-tada ou metabolizada pelos túbulos renais. Uma destassubstâncias é a inulina, que possui um diâmetro aproxima-do de 30 Å. Assim, a filtração glomerular pode ser avalia-da pela medida da depuração ou clearance da inulina. Estamedida é feita após infusão endovenosa contínua de inu-lina, envolvendo as seguintes etapas, conforme o exemplo
abaixo em seres humanos:1) Medida do fluxo urinário (V) em ml/min: 1,0 ml/min2) Medida da concentração urinária de inulina (Uin): 60 mg/
ml3) Cálculo da quantidade de inulina excretada por mi-
nuto:
Uin V 60 mg/ml 1,0 ml/min 60 mg/min
Uma vez que toda a inulina alcançou os rins por filtra-ção e não foi secretada, reabsorvida ou metabolizada pe-los túbulos renais e a concentração plasmática de inulina(Pin) medida foi de 0,5 mg/ml, pode-se afirmar que 120ml de plasma foram filtrados por minuto para haver umaexcreção urinária (Uin V) de 60 mg/min, ou seja:
60 mg/min 5 mg/ml 60 mg/min 1 ml/0,5 mg 120 ml/min
Desta forma, em 1 minuto, 120 ml de plasma e os solu-tos foram separados por ultrafiltração do sangue e dasproteínas plasmáticas. Esta medida da filtração glomeru-lar é o clearance de inulina, cuja fórmula é esta:
Cin Uin V/Pin

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 68/121
36 Filtração Glomerular
O resultado é expresso em ml/min/1,73 m2 de superfí-cie corpórea, significando o volume de plasma no qual todaa inulina é retirada em 1 minuto.
O clearance de inulina é muito utilizado para estudosexperimentais e clínicos, porém é pouco utilizado na prá-tica médica diária devido à necessidade de infusão plas-mática contínua da inulina. Por esta razão, geralmente uti-liza-se o clearance de creatinina, que é uma substância en-dógena e não necessita de infusão venosa, para avaliaçãorotineira da filtração glomerular.
A creatinina não é um marcador ideal da filtração glo-merular, pois existe uma pequena secreção tubular destasubstância. Como outras substâncias endógenas do plas-ma interferem com a dosagem sérica de creatinina supe-restimando sua concentração plasmática, estes dois efeitoscontrários acabam se compensando, o que faz com que oclearance de creatinina seja uma medida bastante razoávelda filtração glomerular na clínica, exceto em pacientes com
filtração glomerular muito baixa, situação na qual a secre-ção tubular de creatinina aumenta muito.
Mais recentemente um outro composto endógeno, acistatina C, tem-se mostrado promissor como marcadorda filtração glomerular. A cistatina C é produzida portodas as células nucleadas e seu ritmo de produção é cons-tante. A cistatina C é livremente filtrada pelo gloméruloe primariamente catabolizada pelos túbulos, de tal formaque como molécula intacta não é reabsorvida nem secre-
tada pelos túbulos. Os níveis plasmáticos da cistatina C já aumentam quando a filtração glomerular cai para 88ml/min/1,73 m2, sugerindo que a medida da cistatina C
sérica pode ser importante na clínica para se detectar ainsuficiência renal inicial que acontece em uma série dedoenças renais para as quais um tratamento precoce écrítico.
BIBLIOGRAFIA SELECIONADA
BOIM, M.A.; TEIXEIRA, V.P.C.; SCHOR, N. Rim e compostos vasoati-vos. In: Zatz, R. Fisiopatologia Renal. Atheneu 2000, p. 21-39.
BRENNER, B.M. The Kidney., 6th ed. W.B. Saunders Company, 2000.COLL, E.; BOTEY, A.; ALVAREZ, L. et al. Serum cystatin C — a new
marker for noninvasive estimation of glomerular filtration rate andas a marker for early renal impairment. Am. J. Kidney Dis., 36:29-34,2000.
STANTON, B.A.; KOEPPEN, B.M. Elements of renal function. In: Berne,
R.M.; Levy, M.N. Physiology, 4th
ed.
Mosby, 1998, p. 677-698.VALTIN, H. and SCHAFER, J.A. Renal Function, 3rd ed. Little, Brown andCompany, 1995, p. 41.
ZATZ, R. Distúrbios da filtração glomerular. In: Zatz, R. FisiopatologiaRenal. Atheneu, 2000, p. 1-20.
ZATZ, R. Proteinúria. In: Zatz, R. Fisiopatologia Renal. Atheneu, 2000, p.41-55.
ENDEREÇO RELEVANTE NA INTERNET
www.renalnet.org

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 69/121
Capítulo
4Função Tubular
Antonio Carlos Seguro, Lúcia H. Kudo e Claudia M. de B. Helou
INTRODUÇÃO
TRANSPORTE ATRAVÉS DA MEMBRANA EPITELIAL
PROCESSOS REGULADORES DE TRANSPORTETRANSPORTE AO LONGO DO NÉFRON
Túbulo proximal
Alça de Henle
características de transporte de suas duas membranas:apical e basolateral (Fig. 4.2).
A membrana apical ou luminal, que está em contatodireto com o fluido tubular, apresenta diferentes canais
iônicos, carregadores, trocadores e co-transportadores, deacordo com as necessidades de transporte do segmento,além de bombas de transporte ativo, como a H-ATPase.
A membrana basolateral é a que está em contato com oespaço intercelular e o capilar peritubular. Além de canaise outros tipos de transportes facilitados, a membrana ba-solateral apresenta uma densidade variável de bombas,que utilizam a energia liberada pela hidrólise do ATP paratransportar ativamente o Na para fora e o K para o inte-rior da célula (Fig. 4.3). Essas bombas são na verdade en-zimas transportadoras e são denominadas de Na,K- ATPases. Em condições normais as Na,K-ATPases distri- buem-se apenas na face basolateral das células tubularesrenais. Como esta enzima necessita de ATP, a sua distri- buição nos segmentos do néfron é diretamente proporcio-
nal aos segmentos que possuem maior quantidade de mi-tocôndrias. Portanto, o túbulo contornado proximal e aporção espessa ascendente da alça de Henle são os segmen-tos do néfron que apresentam maior distribuição quanti-tativa da Na,K-ATPase.
A maior parte do transporte de solutos e de água noepitélio renal é realizada pela via transcelular, ou seja, atra-vés da célula. Mas o fluido e os solutos podem atingir o
Túbulo contornado distal
Túbulo de conexão
Ducto coletorBIBLIOGRAFIA SELECIONADA
ENDEREÇOS RELEVANTES NA INTERNET
INTRODUÇÃO
O néfron é a unidade funcional do rim e é constituídopelo glomérulo e 14 segmentos tubulares. O trabalho de
milhões de néfrons resulta na formação da urina. Cerca de25% do plasma que atinge o rim são ultrafiltrados pelos glo-mérulos, levando à formação de 100 a 120 ml/min deultrafiltrado em média no homem. Entretanto, apenas 1,2%desse volume é eliminado, e o restante reabsorvido da luztubular para o espaço peritubular (Fig. 4.1).
Ao lado deste intenso processo de reabsorção temosoutro, não menos importante, o de secreção tubular. Este secaracteriza pelo transporte de substâncias do espaço peri-tubular (vasos e interstício) para a luz tubular. Este pro-cesso permite a excreção pela urina de substâncias que nãopassaram pela barreira dos capilares glomerulares, comomacromoléculas ou partículas ligadas a proteínas.
Portanto, a formação da urina resulta de três processos:
1. Filtração glomerular2. Reabsorção tubular3. Secreção tubular
O túbulo renal é formado por uma parede de epitéliosimples, ou seja, uma única camada de células que repou-sa sobre a membrana basal birrefringente. As células epi-teliais renais são ditas polarizadas devido às diferentes
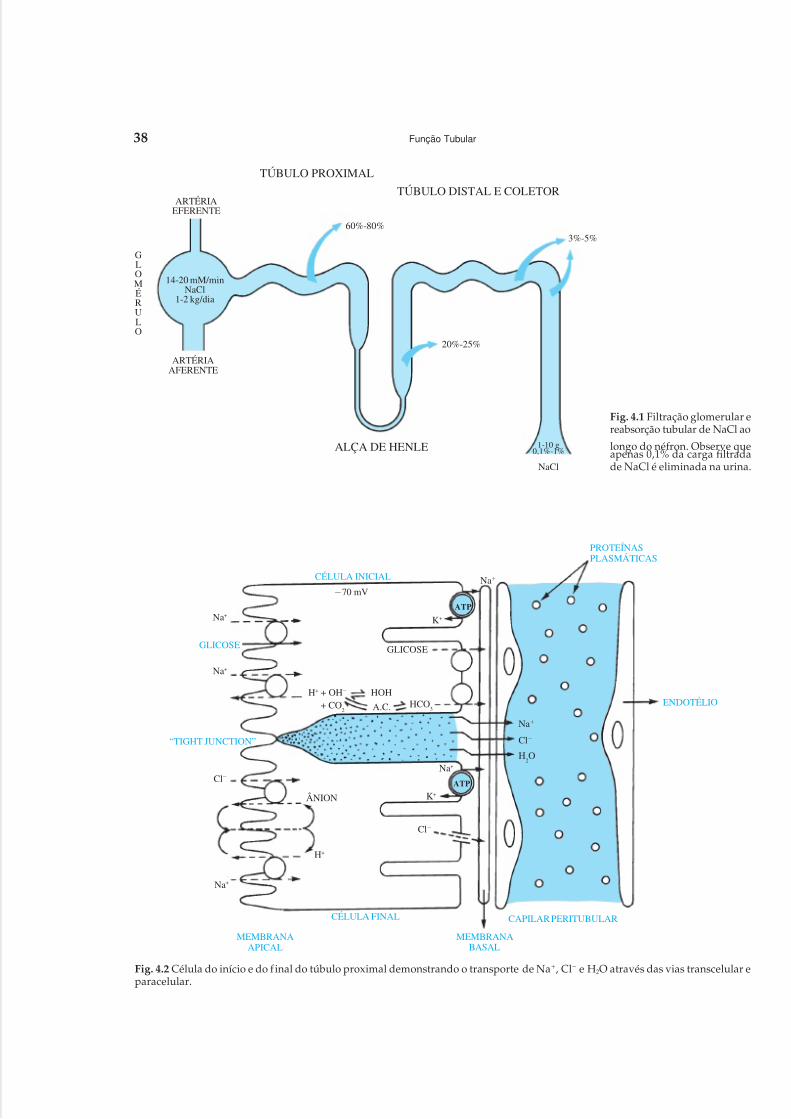
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 70/121
38 Função Tubular
GLOMÉRULO
ARTÉRIAEFERENTE
14-20 mM/minNaCl
1-2 kg/dia
ARTÉRIAAFERENTE
TÚBULO DISTAL E COLETOR
60%-80%
TÚBULO PROXIMAL
3%-5%
20%-25%
ALÇA DE HENLE 1-10 g0,1%-1%
NaCl
Fig. 4.1 Filtração glomerular ereabsorção tubular de NaCl ao
longo do néfron. Observe queapenas 0,1% da carga filtradade NaCl é eliminada na urina.
Fig. 4.2 Célula do início e do f inal do túbulo proximal demonstrando o transporte de Na, Cl e H2O através das vias transcelular eparacelular.
PROTEÍNASPLASMÁTICAS
Na
Na
Cl
H2O
CAPILAR PERITUBULAR
K+
ATP
GLICOSE
CÉLULA INICIAL
70 mV
GLICOSE
H+ + OH
+ CO2
HOH
A.C. HCO3
“TIGHT JUNCTION”
Cl
ATP
K+
Cl
CÉLULA FINAL
MEMBRANAAPICAL
MEMBRANABASAL
Na+
Na+
Na+
H+
ÂNION
ENDOTÉLIO
Na+

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 71/121
capítulo 4 39
capilar pela via paracelular, que é através das junções estrei-tas (tight junctions) e do espaço intercelular, portanto, omovimento é realizado pela face lateral das células. As junções estreitas variam de morfologia e de componentesdependendo do segmento, e por isso são denominadasatualmente de complexos juncionais. É através da alta ou da
baixa condutância dos complexos juncionais que se deter-mina a resistência ao movimento molecular pela via para-celular em muitas células. Pode-se citar como exemplo o
túbulo contornado proximal, que é considerado como seg-mento do néfron cujo epitélio é de vazamento devido à altacondutância do complexo juncional (Fig. 4.2). O contrárioé observado no ducto coletor medular interno, onde ascélulas epiteliais são fortemente aderidas devido à presençade complexos juncionais de baixa condutância, além dedesmossomos.
TRANSPORTE ATRAVÉS DAMEMBRANA EPITELIAL
O transporte de uma substância através de uma mem- brana epitelial pode ser feito por:
1. Mecanismo passivo2. Mecanismo ativo
Nos processos de transporte passivo, o movimentotransepitelial (reabsorção ou secreção) se faz sem gasto deenergia, obedecendo às forças físicas como gradiente quí-mico (reabsorção de uréia), pressão hidrostática (filtraçãoglomerular), gradiente elétrico (reabsorção de cloretos notúbulo proximal) ou pela diferença de potencial eletroquí-mico ocorrido pelo transporte de algum íon, ou então pelaforça física resultante do movimento do arrasto do solvente(solvent drag). O transporte passivo pode ser então por sim- ples difusão ou por difusão facilitada através de poros, carre- gadores ou canais existentes na membrana.
O processo de difusão simples através do epitélio ocor-re com muitas substâncias ao longo do néfron, caracteri-zando-se pela migração transmembrana de uma substân-cia apenas sob a ação do gradiente químico, elétrico ouentão de pH. Neste caso a quantidade transportada depen-derá apenas do gradiente existente e da maior ou menorpermeabilidade da membrana em relação à substância a sertransportada.
Com relação ao solvente como a água, que também éreabsorvida em muitos segmentos do néfron, a difusãopassiva se dá no túbulo renal por osmose, isto é, a água semovimenta do meio menos concentrado (com menor os-molalidade) para o mais concentrado (com maior osmola-lidade). O coeficiente de reflexão do soluto, que pode va-riar de zero a um, é que determina o movimento da água
através da membrana. Quanto maior o coeficiente de re-flexão, maior a capacidade do soluto de produzir um mo-vimento de água através da membrana. Isto é, o soluto quepossui alto coeficiente de reflexão exerce maior pressãoosmótica para um mesmo gradiente de concentração. Aosmose determina a reabsorção de 99% da água filtradapelo glomérulo, e é este tipo de transporte que permite aformação de urina concentrada (alta osmolalidade).
Fig. 4.3 Estrutura da Na-K-ATPase. ( A) A bomba pode ser umheterodímero , . A subunidade contém os sítios de ligaçãopara Na (1), para ATP (4), para fosforilação (5), para K (2) e paraouabaína (3). (B) O painel inferior mostra a subunidade atra-vessando a membrana sete a oito vezes. A subunidade , que églicosilada em sua porção extracelular, atravessa somente umavez a membrana. A função da subunidade não é conhecida, masela é indispensável para o completo funcionamento da Na-K-ATPase.
A
B
2K+
Ouabaína
Citoplasma
3 Na+
Mg ATP Mg ADP + Pi
subunidade subunidade
32
1
4 5
C
C
2 3
N N1
4 5
C
?
?
c
a
b
Pontos-chave:
• A formação da urina se deve à filtraçãoglomerular e ao trabalho do epitélio tubularem processos de reabsorção e secreção
• O transporte tubular se faz pelas vias
transcelular e paracelular através doscomplexos juncionais• O gradiente eletroquímico gerado pela
Na,K-ATPase inserida na membrana basolateral é o responsável por diversostransportes que ocorrem na membranaluminal

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 72/121
40 Função Tubular
O gradiente gerado por pH também pode induzir difu-são passiva de uma substância pela membrana epitelial.Provavelmente devido à natureza hidrofóbica da membra-
na celular, formas não-ionizadas de ácidos e bases fracaspenetram mais rapidamente do que formas ionizadas.Considerando que em muitos segmentos do néfron o pHdo fluido tubular difere do existente no espaço peritubu-lar, a geração de um gradiente de pH favorece a difusãode ácidos e bases fracas pelo epitélio. Se o pH do fluidotubular for mais ácido, como ocorre normalmente, o gra-diente resultante favorecerá a reabsorção de ácidos fracosdo lúmen para o espaço peritubular. Mesmo que a concen-tração do ácido fraco seja idêntica nos dois lados do epité-lio, o baixo pH luminal favorecerá a não-dissociação doácido e portanto a sua difusão do espaço luminal para operitubular. Entretanto, se o pH luminal for mais elevadoque o do espaço peritubular, a dissociação do ácido seráfavorecida, resultando em menor reabsorção, por ser esta
forma menos permeável (Fig. 4.4).O inverso ocorre com bases fracas. A acidificação do flui-
do tubular aumenta a dissociação de bases fracas, dificul-tando então a sua difusão do lúmen para o espaço peritu- bular (Fig. 4.4).
Em resumo, a evidência de transporte passivo origina-se de duas observações básicas: 1a) desaparecimento dotransporte quando se abole ou anula o gradiente elétricoe/ou químico; 2a) quando o uso de inibidores metabólicosnão altera o transporte da substância em estudo.
No caso de transporte ativo, a reabsorção ou a secreçãode uma determinada substância se faz contra gradienteelétrico, químico ou ambos, e por conseguinte é feita à custade energia. No transporte ativo temos uma dependência
imediata do metabolismo celular, e a inibição deste deter-mina a parada do transporte.
Os transportadores que utilizam diretamente a energialiberada pela hidrólise do ATP são considerados como ele-mentos de transporte ativo primário e são chamados de bom-
bas. Na verdade, as bombas são enzimas que possuem umsítio de ligação para o ATP e por isso são também conheci-das como ATPases. A fosforilação destas enzimas permiteque íons sejam transportados contra gradientes químicose/ou elétricos (Fig. 4.3). Um bom exemplo é a Ca-ATPa-se, que ativamente transporta o Ca do intracelular, cujaconcentração é de 100 a 150 nM, para o interstício, onde aconcentração deste íon é aproximadamente 6.000 a 10.000vezes maior (1 mM).
A energia liberada por uma ATPase para o transportede um íon pode induzir um gradiente eletroquímico quefacilita o movimento desse íon a favor do gradiente gera-do. A este transporte iônico pode-se acoplar um outro so-luto que poderá ser na mesma direção, co-transporte, ou emsentido oposto, antiporte. Por isso, este transporte acopla-
do é tido como transporte secundariamente ativo (Fig. 4.2).Como exemplo de co-transporte secundariamente ati-
vo podemos citar o de Na-glicose que existe na face lu-minal das células do túbulo proximal. As Na,K-ATPasespresentes na face basolateral dessas células geram um gra-diente eletroquímico que facilita a entrada de Na pela faceluminal (Fig. 4.2). Esta entrada pode ser através de umaproteína transportadora que possui sítios específicos paraNa e para glicose (Fig. 4.5). Primeiro, o Na se liga ao seurespectivo sítio e produz uma alteração na conformaçãoprotéica do carregador, expondo o sítio para a ligação daglicose. Essa segunda ligação (glicose e receptor) provocauma nova alteração na estrutura da proteína, permitindoque tanto o Na quanto a glicose atravessem a membrana.
Portanto, Na
e glicose passam pela membrana lipoprotéi-ca utilizando a energia liberada pela Na,K-ATPase. Aflorizina pode inibir este co-transporte, competindo coma glicose pelo mesmo sítio de ligação no carregador. A li-gação da florizina ao sítio não promove a segunda altera-ção na proteína carregadora, impedindo então o co-trans-porte Na-glicose (Fig. 4.5).
Em muitos segmentos do néfron a secreção de H ocor-re através do transportador Na-H . Este sistema trocador deíons é também secundariamente ativo, pois a secreção deH para a luz tubular é feita acoplada a um movimentocontrário de Na. O Na movimenta-se da luz para o in-tracelular a favor de gradiente eletroquímico gerado pelaatividade da Na,K-ATPase (Fig. 4.2).
Convém também citar um tipo especial de transporte
ativo, que é a endocitose. Macromoléculas são reabsorvidasatravés do seu envolvimento pela membrana apical, resul-tando em invaginações e formação de vacúolos. Quandoo conteúdo dos vacúolos é de substâncias sólidas, esseprocesso recebe o nome de fagocitose, e quando o vacúoloé formado por fluido, a denominação é de pinocitose. Nocitoplasma, o material fagocitado pode sofrer ações de di-gestão. A extrusão do conteúdo vacuolar para o extracelu-
A
HA HA H A
B OH
H
A
pH 5,5pH 7,4PERITUBULAR
LUZ TUBULAR
BOH BOH
pH 8,5
B
OH
pH 7,4PERITUBULARLUZ TUBULAR
B
Fig. 4.4 Difusão transtubular à custa de um gradiente de pH.Esquema A: reabsorção de um ácido fraco (HA) e ausência dereabsorção de base fraca (BOH) em virtude de o pH do fluidotubular ser inferior ao peritubular. Esquema B: reabsorção de uma
base fraca (BOH) e não-reabsorção de ácido fraco decorrente deum pH urinário alcalino.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 73/121
capítulo 4 41
lar recebe o nome de exocitose e consiste na fusão da mem- brana vacuolar à membrana basolateral da célula e conse-qüente extrusão do conteúdo do vacúolo para o espaçoextracelular.
Nos túbulos renais o transporte de macromoléculas érepresentado principalmente pela reabsorção de proteínasfiltradas pelo glomérulo, que ocorre logo no primeiro seg-
mento do néfron, túbulo contornado proximal.
Pontos-chave:
• Transporte passivo: difusão, difusãofacilitada, “solvent-drag”
• O transporte ativo é realizado por ATPases,enzimas que hidrolisam o ATP
• O gradiente eletroquímico gerado pelasATPases pode permitir o transportesecundário de outros íons
PROCESSOS REGULADORES DETRANSPORTE
Didaticamente podemos dividir os processos regulado-res de transporte em: fatores cinéticos, endocitoses-exocitoses e segundos mensageiros.
Os fatores cinéticos modulam a velocidade de transportealterando a concentração de solutos. O transporte de uma
substância pode ser saturável ou insaturável, independentede ele ser ativo ou passivo.
Um transporte é classificado como saturável quando aquantidade da substância transportada na unidade de tem-po aumenta até um certo limite, acima do qual o aumentoda substância a ser transportada não mais incrementa otransporte, pois alcançou o transporte máximo, Tm. Portan-
to, quando se atinge o Tm de uma substância, nem a adi-ção de energia, no caso de transporte ativo, nem o aumen-to do gradiente químico e/ou elétrico, no caso de transpor-te passivo, aumenta o transporte.
A existência de um transporte máximo saturável podeser decorrente de vários mecanismos:
1. Existência de um carregador auxiliando no transporte.Então, o Tm da substância a ser transportada é deter-minado pela quantidade de carregadores existentes, ou,então, se o sítio de ligação a uma determinada substân-cia apresenta afinidade a uma outra, resultando em umprocesso de competição. A galactose por exemplo com-pete com a glicose pelos mesmos receptores da proteí-na carregadora presente no túbulo contornado proxi-mal.
2. Limite de energia para transporte ativo. Por exemplo, oTm de glicose pode ser diminuído pela presença detransporte de fosfato que compete pela energia libera-da pela Na,K-ATPase.
3. Limite do gradiente eletroquímico gerado pelo transpor-te ativo. Assim, uma substância ou íon sendo transpor-tado da luz tubular para o espaço peritubular por um
Na
Na
Na
FLORIZINA
TIRO-SINA
LISINA
GLICOSE
Na
GLICOSE OUFLORIZINA
FLORIZINA
GLICOSE
Na
Fig. 4.5 Representação esquemática do co-transporte Na-glicose. Os sítios de ligação de sódio e glicose na proteína transportadoralocalizam-se no lado externo da membrana celular. A ligação do sódio causa alteração estrutural na enzima transportadora, resul-tando na exposição do sítio de ligação à glicose. A interação glicose e receptor induz uma segunda alteração estrutural que permitea passagem do Na e da glicose para o interior da célula. A florizina pode competir com a glicose pelo receptor. Entretanto, a ligaçãoflorizina-receptor não induz alteração estrutural, impedindo então que tanto florizina quanto Na sejam transportados para o intra-celular.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 74/121
42 Função Tubular
mecanismo ativo diminuiria progressivamente sua con-centração luminal, aumentando-a no espaço peritubu-lar se esses fluidos não fossem removidos. Este aumen-
to de concentração no espaço peritubular e o gradienteelétrico criado pelo transporte favorecem a volta desteíon ou da substância para a luz tubular, anulando o tra- balho ativo efetuado.
O processo de endocitose-exocitose é considerado comoregulador de transporte, pois em condições de repouso ostransportadores podem estar seqüestrados em vesículaslogo abaixo da membrana apical. É necessário um estímu-lo apropriado para que ocorra a inserção dessas proteínasformando evaginações na face luminal da membrana. Oaumento de inserções dessas proteínas favorece o transpor-te da substância em questão. Como exemplos podemoscitar a secreção de H e o fluxo de água induzido pela va-sopressina. No caso da secreção de H, a acidificação da
célula é o estímulo para a inserção na borda luminal dasvesículas que contêm as H-ATPases. No caso do transpor-te de água estimulado pela vasopressina, os canais de água(aquaporinas) são ancorados à membrana através da ge-ração de AMP cíclico e portanto com a utilização de umsegundo mensageiro. A exocitose por sua vez requer a açãointegrada do citoesqueleto celular. Assim, um estímulo in-duz o aumento de circulação de vesículas ativando tantoa endocitose quanto a exocitose.
A regulação de transporte através da ação de segundosmensageiros vem sendo amplamente estudada, principal-mente nos últimos anos. Entre eles podemos citar a gera-ção do AMP e GMP cíclicos e a variação da concentraçãodo Ca livre intracelular ([Cai]), que podem modulardiretamente as proteínas transportadoras ou afetar a aber-
tura de um canal iônico.Na regulação de transporte existe ainda o fenômeno deadaptação ao longo do tempo. O melhor exemplo é o da esti-mulação da reabsorção de Na no ducto coletor induzidopela aldosterona. Este hormônio estimula a produção deproteínas que ativam os canais de Na+ existentes na mem- brana luminal, como também aumenta a síntese de Na,K-ATPase. Este mineralocorticóide também favorece a inser-
ção e a ativação desta bomba na membrana basolateral. Acapacidade metabólica da célula também é influenciadapela mediação da aldosterona a nível de mitocôndria e
portanto pela produção de ATP (Fig. 4.3). Assim, a aldos-terona é um agonista que participa da adaptação da célulado ducto coletor para aumentar o transporte de Na nestesegmento do néfron.
TRANSPORTE AO LONGO DONÉFRON
Túbulo Proximal
O túbulo proximal, segmento que segue imediatamen-te o glomérulo, é responsável pela reabsorção da maiorparte das substâncias que são filtradas pelo glomérulo. Por
isso, este segmento do néfron desempenha importantepapel no controle da eliminação de diversas substâncias.Assim, pequenas alterações na intensidade de reabsorçãoao nível do túbulo proximal podem causar variações sig-nificantes na excreção urinária de uma dada substância.
O túbulo proximal é constituído por três segmentos. Osdois primeiros, que são denominados de S1 e S2, correspon-dem à parte convoluta do túbulo e a eles se segue umaporção retificada, S3, conhecida também como pars recta. Amaior parte de água, sódio e cloro filtrados pelo gloméru-lo (60% a 70% da carga filtrada) é reabsorvida pelo túbuloproximal (Fig. 4.1).
A análise da composição química do fluido obtido dotúbulo proximal mostra que a concentração de Na perma-nece idêntica à do plasma (140 mEq/L), assim como a
osmolaridade. Estes dados indicam, então, que a reabsor-ção do Na nesta região do néfron é acompanhada pelamesma proporção de água, portanto, uma reabsorção isotô-nica.
Como já foi referido em parágrafos anteriores, a entra-da do Na pela membrana apical das células do túbuloproximal ocorre através de mecanismos passivos a favorde um gradiente eletroquímico gerado pelas Na,K-ATPases presentes na membrana basolateral. Na verdade,esses mecanismos são secundariamente ativos, pois utili-zam a energia liberada pela quebra do ATP. A entrada desódio na célula se faz através de dois mecanismos:
1. co-transporte que pode ser com a glicose, com o fosfatoinorgânico, com os aminoácidos, com os sulfatos ou
então com os outros ácidos orgânicos (Fig. 4.2). Este sis-tema ocorre principalmente nos segmentos S1 e S2 e éatravés de um processo de difusão facilitada que essassubstâncias saem passivamente da célula pela membra-na basolateral;
2. trocador Na-H. Através da quebra da molécula daágua o íon H+ é liberado e secretado para a luz tubularatravés de uma troca com o Na. A hidroxila, por sua
Pontos-chave:
• Certos transportadores como o da glicosesão saturáveis. Portanto, atingem um
transporte máximo (Tm)• O processo de endocitose permite estocardentro das células ATPases e outrasproteínas, como por exemplo asaquaporinas. O inverso, a exocitose, permitea inserção dessas proteínas na membranacelular em condições de estímulo

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 75/121
capítulo 4 43
vez, em presença da anidrase carbônica, reage com oCO2 formando o HCO3
que sai da célula pela membra-na basolateral por um co-transporte ligado ao Na na
proporção de 1 cátion para 3 ânions (Fig. 4.2).No início do túbulo proximal, o gradiente elétrico entre
a luz tubular e o espaço peritubular é da ordem de 2 a4 mV, lúmen negativo (Fig. 4.6). Estes dados sugerem quea reabsorção de Na se faz contra gradiente elétrico. Omovimento de cargas positivas devido à ação das Na,K-ATPases existentes na face basolateral das células seriaresponsável por essa diferença de potencial transtubular.Entretanto, nos segmentos finais do túbulo proximal ondepraticamente toda a glicose, o fosfato e os aminoácidosforam reabsorvidos, a diferença de potencial transtubularpassa a ser de 1 a 2 mV, lúmen positivo (Fig. 4.6). Istoé explicado pela difusão de íons cloro, cuja concentraçãoaumenta progressivamente ao longo do túbulo proximal.
No início do túbulo proximal, a reabsorção de sódio é pre-ferencialmente acompanhada pela reabsorção do bicarbo-nato. Dessa maneira, a concentração de cloro na luz tubu-lar aumenta progressivamente ao longo deste túbulo, atin-gindo a concentração de 135 mEq/L no segmento S3, valoreste superior à do plasma e à do espaço peritubular, que éde 105 a 110 mEq/L, como está ilustrado na Fig. 4.6.
A reabsorção de cloro se faz tanto pela via paracelularquanto pela transcelular. Neste último caso, o cloro entrapela membrana apical através de um trocador de Cl aco-plado a outro ânion, e através de gradiente eletroquímicofavorável, o cloro se difunde pela membrana basolateralda célula. Em conseqüência à difusão passiva dos íons Cl,o gradiente elétrico é gerado com lúmen positivo, favore-cendo portanto a reabsorção passiva de cátions como Na,
K e Ca neste segmento do néfron.Outro importante íon reabsorvido pelo túbulo proximalé o potássio. Este íon utiliza principalmente a via parace-
lular e mecanismos passivos. O fato de a água ser ampla-mente reabsorvida ao longo do néfron induz um aumentona concentração de potássio na luz tubular, criando-se en-
tão um gradiente químico que facilita a sua reabsorção.Além desse mecanismo, também se tem sugerido a possi- bilidade de o K ser reabsorvido neste segmento por umtransporte ativo. Experimentos inibindo a reabsorção deNa com acetazolamida (inibidor da anidrase carbônica)mostraram que a concentração de potássio no fluido tubu-lar diminui, atingindo valores inferiores aos observados noespaço peritubular e plasma, indicando que a reabsorçãode potássio no túbulo contornado proximal envolve tam- bém um mecanismo ativo de transporte.
O transporte de água através do túbulo proximal se faztanto pela via transcelular quanto paracelular devido aogradiente de pressão osmótica existente entre o fluido tu- bular e o espaço peritubular. Apesar do baixo gradienteosmótico, de 2 a 5 mOsm/kg H2O, ele é suficiente parainduzir a reabsorção da água, uma vez que as membranasapical, basolateral e complexo juncional das células do tú- bulo proximal são muito permeáveis a este solvente. Porisso, como já foi referido anteriormente, este epitélio é con-siderado como de vazamento.
Nas porções iniciais do túbulo proximal essa ligeira hi-pertonicidade do fluido peritubular em relação ao lúmené induzida pela reabsorção de Na acoplada ao HCO3
ouao co-transporte com outros solutos como a glicose. Nametade final deste túbulo, embora a concentração luminalde Cl (135 mEq/L) seja maior que a do espaço peritu- bular, a reabsorção da água também é feita por osmose,uma vez que o sódio, o bicarbonato e os outros solutos queforam reabsorvidos na porção inicial geram um gradiente
osmótico maior que o Cl
.A intensa reabsorção de Na e água ao longo do túbulo
contornado proximal forma o gradiente químico que fa-
Fig. 4.6 Transporte de água e solutos ao longo do túbulo contornado proximal.
TÚBULO CONTORNADO PROXIMALA. EFERENTE
GLOMÉRULO
Cl 110
Na+
A. AFERENTE
INICIAL
4 mV
HCO
3
FOSFATO
GLICOSE
AMINOÁCIDOS
FINAL
+ 1,0 mV
CAPILARPERITUBULAR
Cl 135
Cl
Na+
H2O
Ca2+

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 76/121
44 Função Tubular
vorece a reabsorção passiva de outras substâncias perme-áveis a este epitélio, como a uréia, o ácido úrico e os íonsK e Cl. Desta maneira, a diminuição na reabsorção pro-
ximal de Na acarreta também a diminuição da reabsor-ção desses outros solutos. O transporte de Ca e Mg émodulado por fatores hormonais, mas existem evidênciasde que também está relacionado com o transporte ativo deNa. O fosfato também é intensamente reabsorvido, prin-cipalmente nas porções iniciais do túbulo contornado pro-ximal. Este transporte diminui com a redução na quanti-dade de Na reabsorvida e com o aumento da concentra-ção de paratormônio através do estímulo da adenilciclase.
Ainda em relação ao transporte de Na no túbulo pro-ximal, é importante descrever a teoria do balanço gloméru-lo-tubular. Verifica-se que frente a variações fisiológicas dafiltração glomerular ocorrem alterações paralelas da reab-sorção de Na no túbulo proximal, de modo que perma-nece constante a quantidade do íon reabsorvido em rela-
ção à sua carga filtrada, ou seja, a fração de reabsorção de Na
mantém-se inalterada. O balanço glomérulo-tubular é de-corrente pelo menos em grande parte das variações daconcentração de proteínas nos capilares, pressão oncótica,que ocorre durante as alterações da filtração glomerular,como mostra a Fig. 4.7. Quanto à finalidade da existênciado balanço glomérulo-tubular, acredita-se que esse proces-so, juntamente com o feedback túbulo-glomerular, que serádescrito adiante, constituem os dois mecanismos pelosquais o rim impede a perda de sódio durante variações fi-siológicas da carga filtrada de sódio devido a alterações dafiltração glomerular.
Quanto às proteínas, que eventualmente escapam noprocesso de ultrafiltração glomerular, são reabsorvidas atra-vés de mecanismo de endocitose já descrito anteriormente.
A pars recta ou segmento S3 do túbulo proximal se iniciano córtex renal a partir da última alça da parte convoluta ese dirige em linha reta para a medula terminando ao nívelde medula externa. Na microscopia óptica as células des-se segmento são semelhantes às da parte convoluta. Entre-tanto, os estudos de microscopia eletrônica revelam que a pars recta é constituída por células epiteliais retangularescom grande quantidade de mitocôndrias junto à membra-na peritubular, mas com menor número de invaginaçõesna membrana basolateral.
Quanto à fisiologia da pars recta, a reabsorção de sódiotambém se faz à custa da geração de gradiente eletroquími-co induzido pelas Na-K-ATPases presentes na membra-na basolateral. O gradiente elétrico e químico criado pelotransporte de Na é que determina a reabsorção passiva de
Cl, cuja concentração é elevada neste segmento. A reabsor-ção de Na também é do tipo isotônica, pois a mesma quan-tidade de água acompanha este cátion (Fig. 4.8).
Apesar de a pars recta dos néfrons superficiais possuirum comprimento de 5 mm, a quantidade reabsorvida deNaCl e água é apenas em torno de 5 a 10% da carga fil-trada, e portanto significativamente menor do que nasporções convolutas.
Entretanto, analisando a capacidade de secreção de áci-dos orgânicos, verifica-se que a pars recta tem maior capa-cidade em secretar ácido úrico, para-amino-hipurato eoutros ácidos que os segmentos S1 e S2. O transporte des-
Fig. 4.7 Mecanismos que impediriam a perda de NaCl: balançoglomérulo-tubular e feedback túbulo-glomerular.
BALANÇO
GLOMÉRULO-TUBULAR
70%
14 mEq/min
Na+
A. A F E R
E N T E
MÁ C U L
A
D E N SA
10%
N a + 0
, 9 m E q /
m i n
Cl 15%
3-5%
“FEEDBACK”TÚBULO-GLOMERULAR
2,8mEq/min
Fig. 4.8 Processos de reabsorção e secreção na pars recta do túbu-lo proximal.
G L O M É R U L O
T.C. Proximal
Na+ClH
2O
Ác. Orgânicos
K+
UréiaNa+ClH
2O
Ác. Orgânicos
PARS
RECTA
A. A F E R
E N T E

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 77/121
capítulo 4 45
ses ácidos orgânicos é mediado por carregadores e portantopor mecanismo saturável. Do ponto de vista clínico e far-macológico, a alta capacidade do segmento S3 em secretar
ácidos orgânicos constitui uma via importante de excreçãode muitos medicamentos como a aspirina, antibióticos ediuréticos.
Uma outra função muito importante atribuída ao seg-mento S3 é a sua capacidade de secretar K e uréia. Portan-to, a pars recta participa dos mecanismos de concentraçãourinária como elemento integrante no sistema de contra-corrente.
Pontos-chave:
• O túbulo proximal é responsável pelareabsorção isotônica de 60 a 70% da cargafiltrada de NaCl e água
• O sódio é reabsorvido na membranaluminal através de diferentes mecanismos:trocador Na-H, co-transporte com glicose,fosfato e aminoácido
• O bicarbonato é preferencialmentereabsorvido nos segmentos S1 e S2
• Na pars recta (segmento S3) ocorrereabsorção preferencial de Cl e secreção deácidos orgânicos
Alça de Henle
A alça de Henle é dividida em porção fina descendente, porção fina ascendente, porção espessa ascendente medular e porção espessa ascendente cortical.
A porção fina descendente é altamente permeável àágua e pouco permeável a solutos. Aproximadamente 20%da água filtrada é reabsorvida neste segmento. A diferen-ça de potencial transtubular é próxima a zero com lúmennegativo (2 a 4 mV).
O segmento que se segue à porção fina descendente daalça de Henle é a curvatura. Esta porção do néfron é mui-to utilizada pelos micropuncionadores para o estudo dafunção dos néfrons justamedulares.
A porção fina ascendente da alça de Henle apresentacomo característica ser impermeável à água mas permeá-vel a Cl e a Na, que são reabsorvidos por um processopassivo na sua maior parte.
A porção espessa ascendente da alça de Henle que tam- bém é impermeável à água é responsável pela reabsorçãode 25% da carga filtrada de sódio. A Na, K-ATPase pre-sente na membrana basolateral gera um gradiente eletro-químico que favorece a entrada do Na pela membranaapical através de um co-transporte Na-K-2Cl (Fig. 4.9).
Existem indícios de que o co-transporte Na-K-2Cl
obedece a uma seqüência de ligações iônicas que se suce-
dem resultando em alterações na estrutura do co-transpor-tador para poder permitir as uniões seguintes. Primeiro éo Na que se liga, seguindo-se um íon Cl e em terceirolugar o K, e só então é que se liga o segundo Cl. A furo-semida e a bumetanida podem inibir este sistema de co-transporte ao se ligarem no lugar do segundo Cl na últi-
ma etapa.Uma vez no intracelular, o Na é ativamente transporta-do para o interstício através da ação da Na-K-ATPase namembrana basal, mas o K e o Cl são transportados passi-
Fig. 4.9 Célula da porção espessa ascendente da alça de Henlemostrando o co-transporte Na-K-2Cl e o contratransporteNa-Hna membrana luminal. Os íons Na são ativamente trans-portados através da membrana basal pela Na-K-ATPase e osíons K e Cl saem passivamente da célula através de canais.Outro dado importante a assinalar é o potencial positivo do flui-do tubular em relação ao peritubular.
PORÇÃO ESPESSA ASCENDENTE DA ALÇA DE HENLE
LÚMEN
+3 a +10 mV
CÉLULA
70 mV
PERITUBULAR
0 mV
1 Na+
2 Cl
1 K+
Na+
Na+
K++ CO
2
H+ + OH HOHA.C.
HCO
3
K+
K+
Cl
Na+ e
OUTROSCÁTIONS
ATP
Pontos-chave:
• A porção fina descendente é permeável àágua e muito pouco a solutos
• 25% da carga filtrada de NaCl éreabsorvida nas porções ascendentes da alçade Henle
• Presença do co-transportador Na-K-2Cl,sensível ao furosemide, na membranaluminal da porção espessa ascendente. Esteco-transportador é elemento muitoimportante nos mecanismos deconcentração e diluição urinárias
• Ca2 e Mg2 são reabsorvidos pela viaparacelular

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 78/121
46 Função Tubular
vamente. O K retorna ao lúmen através de um canal espe-cífico (pertencente à família ROMK) na membrana apical eo Cl sai da célula pela membrana basal através de um ca-
nal específico a este ânion. A saída de carga positiva para olúmen e de uma carga negativa para o interstício gera umpotencial positivo luminal de cerca de7 mV. Esta diferen-ça de potencial permite que o Na+ e outros cátions como opróprio K, Ca e Mg sejam reabsorvidos passivamentepelos espaços intercelulares laterais, como ilustra a Fig. 4.9.
No segmento cortical da porção espessa ascendente daalça de Henle é descrita também a secreção de H atravésdo trocador Na-H presente na membrana luminal; cer-ca de 10% da carga filtrada de bicarbonato são reabsorvi-dos neste segmento.
Túbulo Contornado Distal
O túbulo contornado distal é a continuidade do segmen-to cortical da porção espessa ascendente da alça de Henlese estendendo da mácula densa até a região de transiçãocom o ducto coletor. Este segmento do néfron também éimpermeável à água e apresenta características especiaisquanto ao transporte de sódio e cloro. Através de um co-transporte com o Cl, o Na é transportado pela membra-na luminal de maneira passiva. Este co-transporte pode serinibido por tiazídicos e é secundariamente ativo à ação daNa-K-ATPase que transporta ativamente o Na+ pelamembrana basal, mas o Cl sai do interior da célula atra-vés de mecanismo passivo por canal específico (Fig. 4.10).
A teoria do feedback túbulo-glomerular relaciona a quan-tidade de Na que chega aos segmentos distais do néfron
e a regulação da filtração glomerular. De acordo com estateoria, a quantidade de Na ao atingir o início do túbulodistal sensibiliza a mácula densa, que por sua vez ativa
mecanismos efetores que irão modular a resistência dosvasos pré-glomerulares. Portanto, se grande quantidade deNa atinge o início do túbulo distal, a renina é liberada,induzindo vasoconstrição da arteríola aferente com con-seqüente redução do fluxo sanguíneo renal, pressão glo-merular e filtração glomerular (Fig. 4.7).
Ponto-chave:
• Presença do co-transportador Na-Cl
sensível a tiazídicos na membrana luminal
Túbulo de Conexão
A região de transição entre o túbulo contornado distale o ducto coletor pode ser abrupta ou gradual, e como nestelocal vários túbulos distais se reúnem para formar o ductocoletor, esse segmento do néfron recebe então o nome detúbulo de conexão. Ele é constituído por dois tipos de célu-las: as do túbulo de conexão e as intercaladas. As célulasdo túbulo de conexão apresentam características morfoló-gicas compatíveis com a transição entre as células do dis-tal e as principais do ducto coletor. A principal funçãodessas células está relacionada à secreção de potássio, queé em parte regulada por mineralocorticóides. As célulasintercaladas, por sua vez, desempenham importante papelnos mecanismos de secreção de H, que será descrito emdetalhes no parágrafo referente ao túbulo coletor.
Neste segmento do néfron, o sódio pode ser reabsorvi-
do através de um co-transporte acoplado ao Cl semelhan-te ao acima descrito nas células do túbulo contornado dis-tal. Aliás, estudos em coelhos onde o túbulo de conexão é bem evidente sugerem que o co-transporte Na-Cl sensí-vel a tiazídico ocorre apenas neste segmento. O Na podetambém ser transportado para o intracelular através decanais sensíveis ao amiloride e do trocador Na-H aco-plado a um trocador Cl-HCO3
. A ação ativa da Na,K-ATPase presente na membrana basal também é respon-sável pela saída do Na da célula (Fig. 4.11).
É importante ressaltar que o túbulo de conexão e o cole-tor cortical são os únicos segmentos do néfron onde o bicar-
Fig. 4.10 Célula do túbulo distal inicial mostrando o transportede Na acoplado a Cl sensível ao tiazídico na membrana lumi-nal secundariamente ativo à ação de Na-K-ATPase da mem-
brana basal.
TÚBULO DISTAL INICIAL
LÚMENNEGATIVO
CÉLULA70 mV
PERITUBULAR0 mV
Na+
ATP
ClINIBIÇÃOPELOTIAZÍDICO
K+
Na+
Cl
Pontos-chave:
• Constituído por células de transição entre asdo túbulo distal e do ducto coletor
• Presença do trocador Cl-HCO3 na
membrana luminal pode permitir a secreçãode bicarbonato em certas condições dealcalemia

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 79/121
capítulo 4 47
bonato pode ser secretado para a luz tubular utilizando paratal o trocador Cl-HCO3
presente na membrana apical.Também é descrita a presença de uma ATPase na face
luminal das células intercaladas neste segmento, denomi-nada H,K-ATPase. Esta enzima seria estimulada peladepleção de K.
Ducto Coletor
O ducto coletor é dividido em cortical, medular exter-no e medular interno, apresentando dois tipos de células:as principais e as intercaladas.
As células principais caracterizam-se na microscopia ele-trônica por apresentarem um cílio central. O sódio é reabsor-vido nestas células por mecanismo passivo através de canaisna membrana luminal sensíveis ao amiloride ou trianterene,denominados ENaC. É também através da Na,K-ATPaseque o transporte ativo de Na gera potencial negativo nolúmen na ordem de30 mV no coletor cortical (Fig. 4.12).
A concentração de K no intracelular das células princi-pais do ducto coletor é elevada devido à alta atividade dasNa,K-ATPases presentes na membrana basal. É através decanais específicos (ROMK) tanto na membrana apical quan-to na basolateral que o Ké transportado passivamente para
fora da célula (Fig. 4.12). A secreção de K
está diretamenterelacionada à diferença de potencial gerada pela quantida-de de Na reabsorvida.
Tanto a reabsorção de sódio quanto a secreção de potás-sio nas células principais do ducto coletor são moduladaspela aldosterona. Este mineralocorticóide entra no citoplas-ma da célula induzindo através de RNAm a síntese de pro-teínas pelo núcleo, que aumentam o número de canais de
sódio da membrana apical, a densidade de Na,K-ATPa-ses da membrana basal e por fim estimulam a produção deATP pelas mitocôndrias, resultando em aumento na ativi-
dade das Na
,K
-ATPases. A espironolactona interfere como sítio citoplasmático da aldosterona impedindo a produ-ção do RNAm, e dessa maneira a reabsorção de Na e a se-creção de K ficam prejudicadas.
As células intercaladas caracterizam-se por serem célu-las escuras devido à presença de grânulos em seu citoplas-ma. São descritos atualmente dois tipos de células interca-ladas: as e as . Elas estão relacionadas ao transporte ati-vo de H através de H-ATPases. Estas se localizam namembrana luminal nas células do tipo e na membrana basal nas do tipo . No caso das células intercaladas do tipoα a secreção luminal de H está acoplada ao sistema troca-dor Cl-HCO3
na membrana basal. O inverso é observadonas células intercaladas do tipo , onde o H é transporta-do pela H-ATPase agora localizada na membrana basal da
célula e o sistema trocador Cl-HCO3 tem localização na
membrana apical. Acredita-se que as condições ácido-bási-cas determinam a quantidade de células ou. Na acidosepredominam as células do tipo e na alcalose, as do tipo .
Como já foi referido no túbulo de conexão, uma ATPa-se relacionada à reabsorção de K está presente nas célu-las intercaladas do ducto coletor. A H,K-ATPase é maisabundante nos segmentos corticais e diminui à medida quese aproxima da papila. Ainda é controverso na literaturase alterações ácido-básicas modulam a atividade dessaenzima. Há indícios de que no ducto coletor medular in-terno outros cátions possam ocupar o lugar do H naH,K-ATPase, e por isso ela tem sido denominada comoX,K-ATPase, como o amônio (NH4
).
O ducto coletor medular interno é dividido em três seg-mentos devido à sua heterogeneidade morfológica e fun-cional: IMCD1, IMCD2 e IMCD3. As células que compõemo IMCD1 são muito semelhantes às do ducto coletor me-dular externo, estando presentes as células do tipo princi-pal e cerca de 10% de intercaladas. Entretanto, as porçõesIMCD2 e IMCD3 parecem representar um segmento distin-to. Estudos recentes têm considerado que o ducto coletormedular interno apresenta dois segmentos funcionalmentedistintos: a porção inicial que corresponde ao IMCD1 e aporção distal com os segmentos IMCD2 e IMCD3.
Este último segmento do néfron tem importante papelna regulação final da composição da urina pelo ajuste dareabsorção de sódio, potássio, uréia e água. Convém sali-entar que o ducto coletor medular interno é o único seg-
mento do néfron que possui sítio de ação aos peptídiosatriais natriuréticos, e também existem evidências da pre-sença do co-transporte Na-K-2Cl.
O transporte de água no túbulo distal final e ducto co-letor varia com a concentração plasmática do hormônio an-tidiurético, HAD, que altera a permeabilidade destes seg-mentos à água. A ação do hormônio antidiurético tornatambém o epitélio do coletor medular permeável à uréia.
Fig. 4.11 Célula do túbulo de conexão mostrando na membranaluminal os canais de Na sensíveis ao amiloride, o co-transporteNa-Cl e os co-transportes Na-He Cl-HCO3
secundariamen-te ativos à Na-K-ATPase da membrana basal.
TÚBULO DE CONEXÃO
LÚMEN5 mV
CÉLULA85 mV
Na+
Na+INIBIÇÃOPELO
AMILORIDE Na+
Cl
K+
ATP
KCl KCl
Cl Na+
PERITUBULAR0 mV
H+
HCO
3

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 80/121
48 Função Tubular
Esta permeabilidade aumenta em direção à papila, promo-vendo a hipertonicidade do interstício. Esta propriedade,que é importante na determinação da osmolaridade daurina, será discutida com mais detalhes em outro capítuloreferente a mecanismos de concentração urinária.
Antes de finalizar este capítulo sobre a função tubular,é importante salientar dois aspectos da função renal:
1.º) A descrição da função tubular foi feita considerandoo rim como constituído por uma população homogênea denéfrons; entretanto, sabemos que existem diferenças mor-fológicas e funcionais entre os néfrons justamedulares (cór-tex profundo) e os néfrons superficiais. Entre as diferençasdeve-se destacar a maior capacidade dos néfrons justame-
dulares de variar a excreção de NaCl frente a variaçõesdo volume extracelular. Em condições de depleção intensado volume extracelular, observa-se uma maior reabsorçãode NaCl pelos néfrons justamedulares e, em condições deexpansão, também são os néfrons profundos os que apre-sentam a maior capacidade de excreção de NaCl.
2.º) A função renal foi apresentada como simplesmenteum processo de filtração glomerular, reabsorção e secreção
Fig. 4.12 Representação dos três tipos de células do ducto coletor: intercaladaα, intercalada e principal. Observa-se que na inter-caladaα a inserção da H-ATPase é na membrana luminal, e do contratransporte Cl-HCO3
, na membrana basolateral, o que favo-rece a secreção de ácidos. O inverso é observado nas células intercaladas . Na face luminal das células principais o Na é reabsor-vido através de canais sensíveis ao amiloride. O K é secretado passivamente para a luz tubular. Ambos os transportes são decor-rentes do gradiente eletroquímico gerado pela Na-K-ATPase na membrana basal.
CÉLULA
Cl
PERITUBULAR
ATPCl
CÉLULA CÉLULA PRINCIPAL
ATP
PERITUBULAR
Cl Cl
H+
Cl
Cl
H+
K+
Na+
K+ K+
Na+
K+
ATP
PERITUBULAR
LUMINAL LUMINAL
Cl
HCO
3
Cl
HCO
3
tubular que permite a manutenção do balanço de sódio,potássio, hidrogênio e água; entretanto, o rim tem outrasimportantes funções do ponto de vista metabólico, como,por exemplo: no metabolismo de hidratos de carbono pelasíntese de glicose que ocorre no córtex renal e pela inativa-ção de insulina e glucagon, diminuindo a meia-vida desseshormônios; no metabolismo ósseo pela regulação da excre-ção de cálcio, fósforo, ativação de vitamina D e inativaçãode paratormônio. Não devemos esquecer também o impor-tante papel do rim na regulação da pressão arterial.
BIBLIOGRAFIA SELECIONADAKRIZ, W. and BANKIR, L. A standard nomenclature for structures of the
kidney. American Journal of Physiology, 254(23):F1-F8, 1988.MOE, O.W.; BERRY, C.A. and RECTOR JR, F.C. Renal transport of
glucose, amino acids, sodium, chloride, and water. In: Brenner, B.M.and Rector, F.C., Jr. (eds.) The Kidney, 6th ed. W.B. Saunders Company,Philadelphia, pp. 375-415, 2000.
SEGURO, A.C.; MAGALDI, A.J.B. ; HELOU, C.M.B.; MALNIC, G. e ZATZ,R. Processamento de água e eletrólitos pelos túbulos renais. In Zatz, R.(ed.) Fisiopatologia Renal, 1.ª edição, Atheneu, pp. 71-96, 2000.
STOKES, J.B. Principles of epithelial transport. In: Narins, R.G. (ed.) Maxwel & Kleeman’s Clinical Disorders of Fluid and Electrolyte Metabo-lism, 5th ed. McGraw-Hill, Inc. New York, pp. 21-44, 1994.
VALTIN, H. and SCHAFER, J.A. Tubular reabsorption.In: Valtin, H. andSchafer, J.A. (eds.) Renal Function, 3rd ed. Little, Brown and Company,pp. 62-82, 1995.
VALTIN, H. and SCHAFER, J.A. Tubular secretion. In: Valtin, H. and
Schafer, J.A. (eds.) Renal Function, 3rd
ed. Little, Brown and Company,pp. 84-93, 1995.
ENDEREÇOS RELEVANTES NA INTERNET
http://www.hdcn.comhttp://www.nephron.comhttp://www.renalnet.org
Pontos-chave:
• As células principais são responsáveis pelareabsorção de sódio e secreção de potássio,
sendo estes processos modulados pelaaldosterona• As células intercaladas são células escuras
responsáveis pela acidificação urinária• O ducto coletor medular interno é a porção
final do néfron, onde ocorrem os ajustesfinais para a formação da urina
LUMINAL

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 81/121
Capítulo
5Mecanismos de Acidificação Urinária
Alexander J. Rouch
INTRODUÇÃO
SECREÇÃO DE H E REABSORÇÃO DE HCO3 NO
TÚBULO PROXIMALSECREÇÃO DE H E REABSORÇÃO DE HCO3
NO
TÚBULO DISTAL
FORMAÇÃO E EXCREÇÃO DE ÁCIDO TITULÁVEL
SÍNTESE E EXCREÇÃO RENAL DE AMÔNIA
REGULAÇÃO DA ACIDIFICAÇÃO URINÁRIA
FUTUROS ESTUDOSRESUMO
REFERÊNCIAS BIBLIOGRÁFICAS
ENDEREÇOS RELEVANTES NA INTERNET
INTRODUÇÃO
O rim, para cumprir o seu papel na manutenção do ba-lanço ácido-básico, deve excretar ácido não-volátil numaproporção igual à sua produção metabólica, que sob ascondições de dieta normal é cerca de 1,0 mEq/kg por dia.Portanto, a excreção de ácido (EA) para um indivíduo quepesa 70 kg deve ser de aproximadamente 70 mEq/dia. Aexcreção renal de ácido (EA) pode ser determinada atra-vés da seguinte equação:
EA UNH4V UTAV UHCO3
V
Esta equação mostra que a excreção deácido, EA, é igualà taxa de excreção de amônia (U V)NH4
mais a taxa de ex-creção de ácido titulável (UTAV) menos a taxa de excreção
de bicarbonato (UHCO3V).Para excretar esse ácido, o rim deve acidificar a urina.
Mecanismos de acidificação urinária envolvem secreção deH ao longo do néfron, particularmente no tú bulo contor-nado proximal e no ducto coletor. É importante observarque a taxa de secreção de H foi “projetada” para manter
normal a concentração plasmática de [HCO3]. Isto requer
que o rim reabsorva todo o HCO3 filtrado e gere novo
HCO3 para substituir aquele que foi usado para tampo-
nar o ácido não-volátil.
Pontos-chave:
• O principal papel do rim é no balançoácido-básico de todo o organismo
• O rim acidifica a urina a fim de excretarácido não-volátil e manter normal aconcentração plasmática de bicarbonato[HCO3
]
Investigações sobre os mecanismos de acidificação uri-nária têm enfocado o transporte celular da secreção renal
de H
e a reabsorção de HCO3
. Neste capítulo, discutire-mos primeiro estes mecanismos de transporte celular notú bulo proximal e no tú bulo distal. A importância da ani-drase carbônica (AC) será enfatizada. Os fatores determi-nantes da excreção de ácido titulável serão então apresen-tados seguidos por uma discussão sobre a síntese e a ex-creção renal de amônia e, finalmente, a regulação da aci-dificação urinária.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 82/121
50 Mecanismos de Acidificação Urinária
Pontos-chave:
• EA (UNH4V) (UTAV) – (UHCO3
V)• EA produção de ácido não-volátil• Mecanismos de ação celular• Tú bulo proximal• Tú bulo distal• Excreção de ácido titulável• Síntese e excreção de amônia• Regulação de acidificação urinária
SECREÇÃO DE H EREABSORÇÃO DE HCO3
NOTÚBULO PROXIMAL
O tú bulo proximal reabsorve cerca de 80% do HCO3
filtrado.Pesquisadores acreditam que a reabsorção de HCO3
pode ocorrer através de dois mecanismos: 1) secreção depróton pela célula para o lúmen; ou 2) transporte direto doHCO3
filtrado através da membrana apical. Estudos uti-lizando diferentes tipos de eletrodos sensíveis ao pH e me-dindo o pH intracelular na presença e na ausência de ini- bição da anidrase carbônica (AC) demonstraram que a se-creção de H é o mecanismo responsável pela acidificaçãodo fluido luminal e pela reabsorção de HCO3
no tú buloproximal, e não o transporte direto de HCO3
. O leitor podeobter mais detalhes desta técnica nos trabalhos indicadosnas referências 1 a 3.
A Fig. 5.1 ilustra os mecanismos fundamentais da secre-ção de H e da reabsorção de HCO3
no tú bulo proximal.A secreção de H ocorre primariamente através da troca
Na/H na membrana apical, embora haja também a par-ticipação da H-ATPase. O primeiro processo está ligadoà entrada passiva de Na para o interior da célula e o últi-
mo é um mecanismo de transporte primariamente ativo.O H secretado combina-se com o HCO3
filtrado paraformar H2CO3, que é convertido em CO2 e H2O no lúmen.O CO2 difunde para o interior da célula, onde a reaçãoquímica é revertida e então formando íons H para poste-rior secreção de íons HCO3
, que deixa a célula pelo lado basolateral e entra para o fluido peritubular. A anidrasecarbônica (AC) estimula a reação química tanto no lúmenquanto no citoplasma celular e mantém alta a taxa de se-creção de H (ver adiante). O transporte de HCO3
parafora da célula pela membrana basolateral ocorre passiva-mente através de dois principais processos: cotransporteNa/3HCO3
e troca HCO3/Cl. Algumas discussões
sobre esses transportadores estão em ordem.
Pontos-chave:
• O tú bulo proximal reabsorve cerca de 80%do HCO3
filtrado• Secreção de H é o mecanismo responsável
pela acidificação do fluido luminal e pelareabsorção de HCO3
no tú bulo proximal enão o transporte direto de HCO3
• A secreção de H ocorre primariamenteatravés da troca Na/H na membranaapical, embora haja também a participaçãoda H-ATPase
Troca Na /H. Vários estudos têm sido realizados para
examinar as características funcionais e moleculares doantiporte Na/H na membrana apical do tú bulo proximal(ver Cap. 4). A troca Na/H foi primeiro demonstrada nasvesículas de membrana da borda em escova do córtex re-nal de ratos.4 Outros trabalhos demonstraram que oantiporte apresenta uma saturação cinética de Michaelis-Menten e que pode ser inibida pelo amiloride. 5,6 Estudosutilizando medidas de pH intracelular no tú bulo proximale técnicas de substituição de Na confirmaram a existên-
cia de um transportador Na/H na membrana apical dotú bulo proximal.7-9 Além disso, este transportador é dife-rente do chamado “housekeeping” trocador Na/H, queajuda a controlar o pH e o volume celular.10
Foram identificados cinco isoformas de trocador Na
/H
,(NHE 1-5) através de estudos em ratos e coelhos, e NHE-1 aNHE-4 foram identificados no rim.11,12 Pesquisas empregan-do técnicas de imuno-histoquímica, expressão molecular deRNAm e métodos de “knockout” de gens confirmam que aisoforma NHE-3 é a proteína da membrana apical do tú bulo
proximal responsável pela secreção de H.13,14 Esta isoformaé sensível ao amiloride e apresenta um peso molecular apa-
Fig. 5.1 Mecanismos de secreção e reabsorção de HCO3 no tú-
bulo proximal.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 83/121
capítulo 5 51
rente de 92.997, e assim como as outras NHEs ela possui sí-tios de fosforilação para proteína-quinase.
A maior parte da secreção de H no tú bulo proximal
ocorre via trocador NHE-3 dependente de Na. O restan-te, via H-ATPase, independente de Na. Isso tem sido de-monstrado nas vesículas de membrana da borda em esco-va e tú bulo proximal de ratos.15,16 Existem evidências quedemonstram que as células do tú bulo proximal podemdefender o pH intracelular em presença de uma carga deácido na ausência de Na extracelular, e este mecanismo éeletrogênico, criando uma voltagem luminal positiva sobcondições apropriadas.17,18
Quando a célula secreta Hpara o interior do lúmen, uma base equivalente é formada e esta deve ser transportada.Existem dois mecanismos aparentes para a saída da baseatravés da membrana basolateral: transporte eletrogênicoNa/3HCO3
e troca Cl/HCO3. Pesquisas mostram que
a maior parte do transporte de HCO3 ocorre via transpor-
tador Na/3HCO3. Técnicas eletrofisiológicas bem como
termodinâmicas foram usadas para determinar a estequio-metria de três HCO3
para um Nano efluxo de HCO3atra-
vés da membrana basolateral.19,20 Além disso, tem sido mos-trado que a absorção de HCO3
no tú bulo proximal é muitomais dependente de Nado que de Cl.21 Estudos empregan-do técnicas de cloning sobre esse transporte certamente for-necerão mais detalhes para melhor entendimento da regu-lação da absorção de HCO3
.22
Pontos-chave:
• Mecanismos de secreção de H e absorçãode HCO3
no tú bulo proximal
• A maior parte da secreção de H
no tú buloproximal ocorre via trocador NHE-3dependente de Na
• Secreção de H através da membrana apical– NHE-3 e H-ATPase• A absorção de HCO3
no tú bulo proximal émuito mais dependente de Na do que deCl
• Transporte de HCO3 através da membrana
basolateral– Na/3HCO3
Cl/HCO3
Porção espessa ascendente. Na alça de Henle, particu-
larmente na porção espessa ascendente, a secreção de H
e a reabsorção de HCO3 ocorrem através de mecanismos
semelhantes que ocorrem no tú bulo proximal. 23,24 A secre-ção de H é sensível ao amiloride e mediada pelo antiporteNHE-3, e a absorção de HCO3
ocorre via cotransporteNa/3HCO3
. A alça de Henle reabsorve cerca de 10 a 15%do HCO3
filtrado. Portanto, apenas 5 a 10% do HCO3 fil-
trado é liberado para o tú bulo distal e ducto coletor.
SECREÇÃO DE H E REABSORÇÃODE HCO3
NO TÚBULO DISTAL
O néfron distal reabsorve o restante do HCO3 filtrado
através de mecanismos que secretam H. A secreção de H
no néfron distal também resulta em titulação de ânionstampões, primariamente fosfato, e em amônia impulsio-nando dois processos necessários para a formação de novo bicarbonato (discutidos adiante). A pequena quantidadedo HCO3
filtrado é liberada para o ducto coletor e reab-sorvida da mesma forma básica que foi descrita no tú buloproximal. Isto é, o íon H secretado combina-se com o íonHCO3
e forma CO2 e H2O no lúmen. O CO2 então se di-funde para o interior da célula e é convertido em HCO3
,que deixa a célula através da membrana basolateral.
O néfron distal apresenta uma considerável heterogenei-dade em relação à morfologia e à função celular. Segmen-
tos distais que precedem o ducto coletor parecem acidificaro lúmen via membrana apical pela troca Na/H mediadapela NHE-2.25,26 O tú bulo coletor cortical (TCC) e o ductocoletor da medula externa (DCME) acidificam o lúmen poruma via eletrogênica, mecanismo independente de Naquetem sido atribuído ao H-ATPase na membrana apical dascélulas intercaladas.27,28 Técnicas de imuno-histoquímica têmsido empregadas para identificar H-ATPase na membra-na apical e na basolateral do tú bulo coletor cortical e na mem- brana apical do ducto coletor da medula externa.15 Estes re-sultados são consistentes com o modelo atual de transportefuncional das células intercaladas do tú bulo distal. Estas cé-lulas podem ser divididas em tipo alfa e tipo beta (ICe IC).O mecanismo básico destas células é ilustrado na Fig 5.2. AIC secreta H e absorve HCO3
, enquanto a IC secreta
HCO3 e absorve H. Pesquisas têm mostrado que no TCC
a absorção de HCO3 predomina durante as condições de
acidose, enquanto a secreção de HCO3 predomina duran-
te as condições de alcalose.29
Pontos-chave:
• A alça de Henle reabsorve cerca de 10 a 15%do HCO3
filtrado• O néfron distal reabsorve o restante do
HCO3 filtrado através de mecanismos que
secretam H
• A secreção de H no néfron distal tambémresulta em titulação de ânions tampões,
primariamente fosfato, e em amônia,impulsionando dois processos necessáriospara a formação de novo bicarbonato
• No TCC, a absorção de HCO3 predomina
durante as condições de acidose, enquanto asecreção de HCO3
predomina durante ascondições de alcalose

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 84/121
52 Mecanismos de Acidificação Urinária
Evidências também indicam que H/K-ATPase noducto coletor funciona para promover a absorção de K esecreção de H.30 A depleção de K estimula a H/K-
ATPase no ducto coletor.31,32 A significância deste transpor-tador sobre regulação global da acidificação urinária nãoestá ainda esclarecida, e é a H-ATPase o método predo-minante para secreção de H no tú bulo distal.
Assim como no tú bulo proximal, a secreção de um pró-ton através da membrana apical do néfron distal obriga asaída de uma base equivalente através da membrana ba-solateral. Evidências indicam que no tú bulo distal o troca-dor Cl/HCO3
na membrana basolateral é o mediadorneste mecanismo.33 O íon Cl que entrou na célula via estetrocador deixa a célula através da membrana basolateralvia canal condutor de Cl. 34
Pontos-chave:
Mecanismos de secreção de H1 e absorção deHCO3
no túbulo distal• IC• Membrana apical — H-ATPase, H/K-
ATPase• Membrana basolateral — troca H/HCO3
• IC• Membrana apical — trocador Cl/HCO3
• Membrana basolateral — H-ATPase
Pontos-chave:
Papel funcional da anidrase carbônica• Esta enzima catalisa a reação entre CO2 e
OH para formar HCO3
• CO2 OH HCO3
• Acelera a secreção de H e a absorção deHCO3
FORMAÇÃO E EXCREÇÃO DEÁCIDO TITULÁVEL
Como já foi mencionado anteriormente, o organismoproduz uma quantidade de ácido fixo ou não-volátil atra-vés do metabolismo de uma dieta normal. Estes ácidosprimariamente incluem ácido sulfúrico produzido a par-tir do metabolismo de aminoácidos contendo enxofre, áci-do fosfórico produzido pelo metabolismo de fosfolípidese ácido clorídrico produzido pelo metabolismo de amino-ácidos catiônicos. A produção de ácido metabólico é par-
Importância da anidrase carbônica. Esta enzima já foimencionada anteriormente, sendo importante reconhecera sua função na acidificação urinária. Ela catalisa a reação
entre CO2 e OH para formar HCO3.35 Sem a anidrase car- bônica (AC), as reações químicas no lúmen e na célulaocorreriam num ritmo muito lento e a secreção de H e areabsorção de HCO3
seriam significantemente reduzidas.Ânions monovalentes e sulfonamidas inibem a AC.36 É bem conhecido que a acetazolamida, inibidor da AC, re-duz a secreção de H e a reabsorção de HCO3
e pode cau-sar uma acidose metabólica.
No organismo existem quatro isoformas de AC (I-IV) esua distribuição ao longo do néfron tem sido muito estu-dada.37,38 A isoforma tipo II é encontrada no citoplasma dascélulas renais, enquanto a do tipo IV é encontrada na mem- brana plasmática. Nas células do tú bulo contornado pro-ximal, a AC é encontrada tanto no citoplasma quanto na
membrana apical e na basolateral. A atividade enzimáticaé aparentemente reduzida nos segmentos terminais dotú bulo proximal. A porção espessa ascendente da maioriadas espécies estudadas mostrou uma atividade da AC nocitoplasma e nas membranas apical e basolateral, emborao segmento do coelho seja uma exceção. As células interca-ladas no ducto coletor contêm altos níveis de AC, que estácorrelacionada com o seu papel na acidificação urinária.
Fig. 5.2 Transporte de H e HCO3pelas células intercaladas (Cl
e Cl).

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 85/121
capítulo 5 53
cialmente balanceada pela produção de HCO3 através do
metabolismo de aminoácidos aniônicos e ácidos orgânicos.Entretanto, o efeito global do metabolismo é a produção
resultante de ácido que entra na corrente sanguínea e étamponado pelo HCO3
extracelular. Estes ácidos são fil-trados no glomérulo e titulados pelo mecanismo de secre-ção de H, descrito anteriormente.
A Fig. 5.3 ilustra um exemplo de titulação de fosfatoem que a quantidade de ácido titulável na urina pode serdeterminada medindo a quantidade de base forte neces-sária para trazer de volta o pH urinário para o mesmo pHplasmático, que é em torno de 7,4. Quantitativamente, éo ácido titulável mais importante. O H secretado com- bina-se com o fosfato monoidrog ênico para formar o fos-fato diidrogênico que é excretado pela urina final. O im-portante evento neste processo é que um novo íon HCO3
(isto é, aquele que não foi filtrado) é formado na célula eabsorvido para repor o HCO3
usado no tamponamento
extracelular.Três fatores determinam a eficácia da excreção de áci-
do titulável: 1) a quantidade total de ácido fraco disponí-vel para a titulação; 2) o pK do ácido fraco; e 3) o limite decapacidade do rim acidificar a urina (isto é, o menor pHurinário). A reação de equilí brio de fosfato é um bom exem-
plo:
HPO42 H ↔ H2PO4
pK 6,8
H2PO4 H ↔ H3PO4 pK 2,1
O menor pH urinário que o rim é capaz de produzir écerca de 4,5 ([H] 3,2 105 M). A este nível, H-ATPa-se não é capaz de bombear íons H contra um gradientede concentração (entre a célula e o lúmen) tão grande (mai-or que 1:1.000). A um pH de 4,5, virtualmente todo o fos-fato está sob a forma de fosfato diidrogênico (H2PO4).H3PO4 não pode ser formado em grande quantidade naurina porque o baixo pK desta reação está fora da ampli-tude de variação do pH urinário. Deve ser observado que
o menor pH urinário é obtido no ducto coletor, enquantoo pH no final do tú bulo proximal é cerca de 6,5.
Se mais fosfato puder ser liberado para o ducto coletor,
então mais H2PO4 poderá ser formado no lúmen e maisHCO3
novo poderá ser liberado para a corrente sanguí-nea. Outros ácidos que são titulados são: citrato acetato ecreatinina, mas eles contribuem muito pouco para a quan-tidade total de ácido titulável comparado ao fosfato. Sim-plesmente não há ácido titulável suficiente para o rim re-por completamente o HCO3
usado no tamponamento deácidos não-voláteis. Felizmente, o rim utiliza um outromecanismo para gerar novo HCO3
, que é a s íntese e aexcreção de amônia.
Pontos-chave:
Ácido titulável (AT)•
A quantidade de ácido titulável na urinapode ser determinada medindo aquantidade de base forte necessária paratrazer de volta o pH urinário para o mesmopH plasmático, que é em torno de 7,4
• Forma novo HCO3
• Fosfato — o AT mais abundante
Importantes fatores na excreção de AT• Quantidade total de AT• pK• pH urinário
SÍNTESE E EXCREÇÃO RENALDE AMÔNIA
Amônia é uma base fraca existente em equilí brio comoamônia iônica, NH4
, e não-iônica, NH3:
NH3 H ↔ NH4 pK 9,2
Tanto a amônia quanto o fosfato são tampões urinári-
os. Por definição, a amônia não é um ácido titulável. Como já foi mencionado anteriormente, o ácido titulável é deter-minado pela quantidade de base forte necessária para tra-zer de volta o pH urinário para 7,4. Como o pK da reaçãoda amônia é 9,2 num pH de 7,4, a maior parte da amôniapermanece ainda como NH4
e não se dissocia até o pH al-
cançar 8,0 a 8,5. Sob condições normais, a maioria do H
na urina final é excretada como NH4 e a excreção renal de
NH4 aumenta significantemente durante condições de
acidose. Portanto, quantitativamente, embora o fosfato sejao ácido titulável mais importante, a amônia é o mais im-portante tampão urinário.
O mecanismo básico da síntese renal de amônia é ilus-trado na Fig. 5.4. NH4
é produzido primariamente no tú-Fig. 5.3 Mecanismo de titulação do fosfato.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 86/121
54 Mecanismos de Acidificação Urinária
bulo proximal a partir do metabolismo de aminoácido,glutamina, via enzima glutaminase I. A amônia pode tam- bém ser sintetizada em outros segmentos do néfron, massua síntese é mais abundante e fisiologicamente mais rele-vante no tú bulo contornado proximal.39,40
A síntese envolve a entrada de glutamina nas células dotú bulo proximal e uma série de passos bioquímicos paraformar dois íons NH4
e dois íons HCO3. O NH4
é secre-tado no lúmen pela substituição de H do trocador Na/H na membrana apical. O HCO3
é transportado para acorrente sanguínea pelo mesmo mecanismo ilustrado naFig. 5.1.
A manipulação renal da amônia secretada é ilustrada na
Fig. 5.5. Este mecanismo é relativamente complexo e não écompletamente conhecido. Uma parte da NH4
secretadano tú bulo proximal é reabsorvida na porção espessa ascen-dente pela substituição de íon K do cotransportador Na/K/2Cl na membrana apical. Uma parte da NH4
reab-sorvida entra na porção fina descendente da alça de Hen-le e portanto é reciclada. Este processo produz um gradi-ente córtico-medular para NH4
semelhante ao gradiente
Fig. 5.4 Mecanismo da síntese renal de amônia.
Fig. 5.5 Manipulação renal do NH4
excretado.
para NaCl estabelecido pelo mecanismo de contracorren-te multiplicador.
O epitélio renal é relativamente impermeável ao íon
NH4
mas é permeável ao NH3, não-iônico, que se difun-de passivamente para o lúmen do ducto coletor onde ocor-re a secreção de H pelas células intercaladas , formandoentão NH4
. A NH4 é assim captada no lúmen e excreta-
da. Como o pK da amônia é 9,2, sua maior parte está sob aforma de NH4
na urina ácida. Este mecanismo é referidocomo “difusão-captação” da amônia. Observe que, diferen-te do fosfato que é derivado somente da dieta, NH4
é sin-tetizada pelo rim e portanto está sujeita a regulação fisio-lógica (ver adiante).
Pontos-chave:
• Significância da síntese de amônia — geranovo HCO
3
• Tanto a amônia quanto o fosfato sãotampões urinários
• Quantitativamente, embora o fosfato seja oácido titulável mais importante, a amônia éo mais importante tampão urinário
• Síntese da amônia– Metabolismo de glutamina no tú bulo
proximal
Manipulação renal de amônia• Secreção de NH4
no tú bulo proximal• Absorção de NH4
na porção espessaascendente
•
Difusão de NH4
para o tú bulo distal• “Difusão-captação” via secreção de H no
tú bulo distal
REGULAÇÃO DA ACIDIFICAÇÃOURINÁRIA
Existem vários fatores que influenciam os mecanismosde acidificação já descritos. Alguns desses fatores regulama quantidade e o ritmo de acidificação, enquanto outrossimplesmente afetam esses mecanismos.
Um fator importante que afeta a reabsorção de HCO3
é a sua carga filtrada. O balanço glomérulo-tubular para
HCO3
ocorre da mesma forma que ocorre para o Na
,onde a mudança na carga filtrada de HCO3
resulta numamudança apropriada na reabsorção de HCO3
no tú buloproximal. Um aumento na carga filtrada resulta num au-mento na reabsorção, prevenindo assim a bicarbonatúria.O mecanismo para esta resposta adaptativa é ainda des-conhecido. Aumentando a taxa de filtração glomerular(RFG), aumenta o fluxo luminal no tú bulo proximal, e isso

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 87/121
capítulo 5 55
de alguma forma estimula a troca Na/H e/ou transpor-te Na/3HCO3
.41
Como a troca Na/H é responsável pela maior parte
da secreção de H
no tú bulo proximal, fatores que regu-lam a reabsorção de Na afetarão também a secreção de H
neste segmento do néfron. Obviamente, estes fatores tam- bém afetarão a reabsorção de HCO3
no tú bulo proximal.Talvez o fator mais importante seja o volume extracelular(VEC). Quando o VEC se expande, a reabsorção de Na notú bulo proximal diminui e portanto a reabsorção de HCO3
também diminui. Quando o VEC se contrai, aumenta areabsorção de Na e de HCO3
no tú bulo proximal.O status ácido-base influencia a secreção de H. Tanto a
acidose metabólica quanto a respiratória estimulam a se-creção de Hno tú bulo proximal, porção espessa ascenden-te e ducto coletor. A acidose aumenta a secreção porquequando o fluido intracelular acidifica, o gradiente célula-lúmen torna-se favorável para a secreção de H através de
ambas as vias: troca Na/H e H-ATPase. Como era es-perado, a alcalose diminui a secreção de H. Existem evi-dências interessantes indicando que mudanças no pH pe-ritubular resultam na inserção de novos transportadoresde H na membrana apical do tú bulo proximal e ductocoletor.42
Acidose, tanto a crônica quanto a aguda, também esti-mula a síntese renal de amônia, provavelmente ativandoenzimas envolvidas no metabolismo da glutamina. Dimi-nuição do pH intracelular aparentemente estimula a pro-dução enzimática da amônia e conseqüentemente a pro-dução de novos HCO3
. A concentração plasmática de K
também influencia a síntese de amônia. O mecanismo paraeste efeito não está totalmente esclarecido, entretanto, acre-
dita-se que está relacionado às mudanças na concentraçãointracelular de H. Hipercalemia diminui a concentraçãointracelular de H, assim inibindo a síntese de amônia,enquanto a hipocalemia apresenta efeitos opostos. Doucetmostrou que a deficiência de K estimula a secreção de H
via o aumento da expressão aparente de H/K-ATPasena membrana apical do ducto coletor.31
Efeitos hormonais sobre a acidificação urinária. Vári-os hormônios afetam a secreção de H no néfron. Existemnovamente questões como: se estes efeitos hormonais sãodestinados a regular a acidificação urinária ou se eles sãosimplesmente destinados a alterar a secreção de H. Doishormônios que têm sido muito estudados são a aldostero-na e o hormônio da paratireóide, o paratormônio (PTH).
Aldosterona estimula a secreção de H no ducto cole-
tor através de diferentes mecanismos. Um é pela estimu-lação da reabsorção de Na nas células principais. Como já foi dito anteriormente, o ducto coletor apresenta célulasprincipais e células intercaladas. As células principais reab-sorvem o Na e secretam o K, e a aldosterona estimulaambos os processos. A estimulação da reabsorção de Na
no ducto coletor pela aldosterona produz uma voltagemnegativa no lúmen que aumenta o gradiente eletroquími-
co para secreção de H. Isto representa um mecanismoindireto da aldosterona na secreção de H no ducto cole-tor. Há também evidências que mostram que a aldostero-
na estimula diretamente a secreção de H
e a troca Cl
/HCO3
no ducto coletor.43 Um outro mecanismo indiretopelo qual a aldosterona afeta a acidificação urinária estárelacionado com a síntese de amônia. Aldosterona estimulaa secreção de K nas células principais e isto pode levar àhipocalemia, que aumenta a produção de amônia, como jáfoi explicado. Com maior produção de amônia no tú buloproximal, maior reabsorção de HCO3
pode ocorrer noducto coletor.
Assim como a aldosterona, o PTH afeta a secreção deH de várias formas. Evidências mostram que o PTH esti-mula a adenil-ciclase no tú bulo proximal, e o aumento deconcentração celular de AMPc inibe a troca Na/H. OPTH pode estimular a fosfolipase C levando a um aumen-to de cálcio intracelular e ativando a proteinoquinase C.
Estas vias celulares podem estar ligadas à inibição da aci-dificação. PTH também inibe a reabsorção de HCO3
naporção espessa ascendente. Evidências também indicamque o PTH aumenta a secreção de H no ducto coletor,aumentando a excreção deânions não-reabsorvíveis e agin-do como um tampão urinário, tal como o fosfato. Isso podeser um mecanismo compensatório para a inibição de secre-ção de H induzido pelo PTH no tú bulo proximal. As in-fluências do PTH sobre a acidificação urinária continuam
Pontos-chave:
Regulação da acidificação urinária•
Carga filtrada de HCO3
– Aumento da carga filtrada aumenta aabsorção e vice-versa
• Status ácido-base– Acidose aumenta a secreção de H e a
alcalose diminui a secreção de H
• Status volume extracelular (VEC)– Expansão de VEC inibe a secreção de H
e a contração do VEC estimula a secreçãode H
• Amoniagênese– Acidose aumenta a síntese e a alcalose
diminui a síntese– Hipocalemia aumenta a síntese, e a
hipercalemia diminui a síntese• Hormônios– Aldosterona aumenta a secreção de H
direta e indiretamente– PTH inibe a secreção de H no tú bulo
proximal

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 88/121
56 Mecanismos de Acidificação Urinária
controversas, e mais estudos são necessários para melhorentendimento.
Glicocorticóides aumentam a troca Na/H no tú bulo
proximal, aumentando a quantidade de proteína NHE-3 ede RNAm.44 Outros hormônios que também aumentam asecreção de H e HCO3
são: adenosina, catecolaminas,endotelinas e angiotensina II. Provavelmente existem ou-tros envolvidos.
FUTUROS ESTUDOS
A evidência com relação ao efeito de glicocorticóides eda proteína NHE-3 enfatiza a necessidade de futuras pes-quisas nesta área. O transporte de proteínas responsáveispela acidificação urinária ao nível molecular está agoraesclarecido, e evidências sobre a regulação destas proteí-
nas a este nível certamente irão fornecer respostas às ques-tões de como a acidificação urinária é regulada e irão dis-tinguir os reguladores e os efetores.
RESUMO
O complexo mecanismo de acidificação urinária é desti-nado a manter o balanço ácido-básico que requer um pHplasmático estável (entre 7,35 e 7,40 ) e a concentração plas-mática de HCO3
(entre 23 e 25 mM). O rim deve reabsor-ver todo o HCO3
e fabricar novo HCO3 para substituir
aquele que foi usado para tamponar o ácido não-volátil. Orim reabsorve HCO3
através da secreção de H que ocorreatravés de três mecanismos primários: troca Na/H, H-ATPase e troca K/H. O tú bulo proximal e a porção espessaascendente secretam Hprimariamente pela troca Na/H
via proteína NHE-3, e esses segmentos do néfron reabsor-vem a maior parte da carga filtrada de HCO3
. O tú bulodistal e o ducto coletor secretam Hprimariamente pela H-ATPase nas células intercaladas alfa (CI). Novo HCO3
égerado pela excreção deácido titulável e síntese de amônia.Quantitativamente, o ácido titulável mais importante é ofosfato. Amônia, o tampão mais importante, é sintetizadaatravés do metabolismo de glutamina no tú bulo proximal.
A carga filtrada de HCO3 afeta a sua reabsorção no
tú bulo proximal assim como o status de VEC, alterando areabsorção de Na. Acidose estimula e alcalose inibe a se-creção de H. Acidose e hipocalemia estimulam a síntese
de amônia, enquanto alcalose e hipercalemia apresentamefeitos inibitórios. Aldosterona estimula a secreção de H
no ducto coletor através de um mecanismo indireto asso-ciado ao aumento de reabsorção de Na e um mecanismo
direto associado à H-ATPase, e por um outro mecanismoindireto associado à hipocalemia e à síntese de amônia.PTH inibe a troca Na/H no tú bulo proximal estimulan-do a adenil-ciclase.
Para obter uma discussão mais completa e detalhada,aconselhamos o leitor a consultar a seguinte referência:Alpern, R.J. Renal acidification mechanisms. Brenner, B.M.
(Ed.) The Kidney, 6th ed. Philadelphia: Saunders, 2000,Capítulo 11.
REFERÊNCIAS BIBLIOGRÁFICAS
1. DUBOSE, T.D.J.; PUCACCO, L.R. and CARTER, N.W. Determina-tion of disequilibrium pH in the rat kidney in vivo: evidence forhydrogen secretion. Am. J. Physiol ., 240 (Renal Fluid Electrolyte Phy-siol., 9):F138-F146, 1981.
2. KURTZ, I.; STAR, R.; BALABAN, R.S.; GARVIN, J.L. and KNEPPER,M.A. Spontaneous luminal disequilibrium pH in S3 proximal tubu-les. J. Clin. Invest., 78:989-996, 1986.
3. VIEIRA, G.L. and MALNIC, B. Hydrogen ion secretion by rat renalcortical tubules as studied by an antimony microelectrode. Am. J.Physiol., 214: 710-718, 1968.
4. MURER, H.; HOPFER, U. and KINNE, R. Sodium/proton antiportin brush-border membrane vesicles isolated from rat small intestineand kidney. Biochem. J ., 154: 597-604, 1976.
5. KINSELLA, J.L. and ARONSON, P.S. Amiloride inhibition of the Na-H exchanger in renal microvillus membrane vesicles. Am. J. Physi-ol., 241:F374-F379, 1981.
6. WARNOCK, D.G.; REENSTRA, W.W. and YEE, V.J. Na/H antipor-ter of brush border vesicles: Studies with acridine orange uptake. Am. J. Physiol., 242:F733-F739, 1982.
7. ALPERN, R.J. and CHAMBERS, M. Cell pH in the rat proximal con-voluted tubule. J. Clin. Invest., 78:502-510, 1986.
8. BORON, W.F. and BOULPAEP, E.L. Intracellular pH regulation inthe renal proximal tubule of the salamander. J. Gen. Physiol., 81:53-94, 1983.
9. SASAKI, S.; SHIIGAI, T. and TAKEUCHI, J. Intracellular pH in theisolated perfused rabbit proximal straight tubule. Am. J. Physiol.,249(Renal Fluid Electrolyte Physiol., 18):F417-F423, 1985.
10. HAGGERTY, J.G.; AGARWAL, N. and REILLY, R.F. Pharmacolo-gically different Na/H antiporters on the apical and basolateral
surfaces of cultured porcine kidney cells (LLC-PK1). Proc. Natl. Acad.Sci., 85:6797-6801, 1988.
11. ORLOWSKI, J.; KANDASAMY, R.A. and SHULL, G.E. Molecularcloning of putative members of Na/H exchanger gene family. J. Biol.Chem., 267:9331-9339, 1992.
12. TSE, C.M.; BRANT, S.R.; WALKER, M.S.; POUYSSEGUR, J. andDONOWITZ, M. Cloning and sequencing of a rabbit cDNA enco-ding an intestinal and kidney-specific Na/H exchanger isoform(NHE-3). J. Biol. Chem., 267:9340-9346, 1992.
13. ORLOWSKI, J. Heterologous expression and functional propertiesof amiloride high affinity (NHE-1) and low affinity (NHE-3) isoformsof the rat Na/H exchanger. J. Biol. Chem., 268:16369-16377, 1993.
14. TSE, C.M.; LEVINE, S.A.; YUN, C.H.C.; BRANT, S.R.; POUSYSSE-GUR, J.; MONTROSE, M.H. and DONOWITZ, M. Functional cha-racteristics of a cloned epithelial Na/H exchanger (NHE3): resis-tance to amiloride and inhibition by protein kinase C. Proc. Natl. Acad. Sci., 90:9110-9114, 1993.
15. BROWN, D.; HIRSCH, S. and GLUCK, S. Localization of a proton-
pumping ATPase in rat kidney. J. Clin. Invest., 82:2114-2126, 1988.16. KINNE-SAFFRAN, E.; BEAUWENS, R. and KINNE, R. An ATP-driven proton pump in brush-border membranes from rat renalcortex. J. Membrane Biol., 64:67-76, 1982.
17. KURTZ, I. Apical Na/H antiporter and glycolysis-dependent H-ATPase regulate intracellular pH in the rabbit S3 proximal tubule. J.Clin. Invest., 80:928-935, 1987.
18. YOSHITOMI, K.; BURCKHARDT, B.C. and FROMTER, E. Rheoge-nic sodium-bicarbonate cotransport in the peritubular cell membra-ne of rat renal proximal tubule. Pflugers Arch., 405:360-366, 1985.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 89/121
capítulo 5 57
19. ALPERN, R.J. Cell mechanisms of proximal tubule acidification.Physiol. Rev., 70:79-114, 1990.
20. PREISIG, P.A. and ALPERN, R.J. Basolateral membrane H/OH/HCO3 transport in the proximal tubule. Am. J. Physiol . 256:F751-F765,
1989.21. SASAKI, S. and BERRY, C.A. Mechanism of bicarbonate exit across
basolateral membrane of the rabbit proximal convoluted tubule. Am. J. Physiol., 246:F889-F896, 1984.
22. ROMERO, M.F.; HEDIGER, M.A.; BOULPAEP, E.L. and BORON,W.F. Expression cloning and characterization of a renal electrogenicNa/HCO3
cotransporter. Nature., 387:409-413, 1997.23. GOOD, D.W. Ammonia and bicarbonate transport by thick
ascending limb of rat kidney. Am. J. Physiol., 248:F821-F829, 1984.24. KRAPF, R. Basolateral membrane H/OH/HCO3 transport in the rat
cortical thick ascending limb. J. Clin. Invest., 82:234-241, 1988.25. CHAMBREY, R.; WARNOCK, D.G.; PODEVIN, R.A. BRUNEVAL,
P.; MANDET, C.; BELAIR, M.F.; BARIETY, J. and PAILLARD, M.Immunolocalization of the Na/H exchanger isoform NHE2 in ratkidney. Am. J. Physiol., 275:F379-F386, 1998.
26. WANG, T.; MALNIC, G.; GIEBISCH, G. and CHAN, Y.L. Renal bicarbonate reabsorption in the rat. IV. Bicarbonate transport me-chanisms in the early and late distal tubule. J. Clin. Invest., 91:2776-
2784, 1993.27. KOEPPEN, B.M. and HELMAN, S.I. Acidification of luminal fluid by the rabbit cortical collecting tubule perfused in vitro. Am. J. Phy-siol. 242(Renal Fluid Electrolyte Physiol., 11):F521-F531, 1982.
28. LOMBARD, W.E.; KOKKO, J.P. and JACOBSON, H.R. Bicarbonatetransport in cortical and outer medullary collecting tubules. Am. J.Physiol., 244(Renal Fluid Electrolyte Physiol., 13):F289-F296, 1983.
29. MCKINNEY, T.D. and BURG, M.B. Bicarbonate transport by rabbitcortical collecting tubules. J. Clin. Invest., 60:766-768, 1977.
30. NAKAMURA, S.; WANG, Z.; GALLA, J.H. and SOLEIMANI, M. K
depletion increases HCO3 reabsorption in OMCD by activation of
colonic H-K-ATPase. Am. J. Physiol., 274:F687-F692, 1998.31. DOUCET, A. and MARSY, S. Characterization of K-ATPase activi-
ty in distal nephron: Stimulation by potassium depletion. Am. J. Phy-siol., 253:F418-F423, 1987.
32. WINGO, C.S. Active proton secretion and potassium absorption inthe rabbit outer medullary collecting duct. J. Clin. Invest ., 84:361-365,1989.
33. HAYS, S.R. and ALPERN, R.J. Apical and basolateral membrane H
extrusion mechanisms in inner stripe of rabbit outer medullary col-lecting duct. Am. J. Physiol., 259(Renal Fluid Electrolyte Physiol.,28):F628-F635, 1990.
34. KOEPPEN, B.M. Conductive properties of the rabbit outer medul-lary collecting duct: Inner stripe. Am. J. Physiol ., 248:F500-F506, 1985.
35. SILVERMAN, D.N. and VINCENT, S.H. Proton transfer in thecatalytic mechanism of carbonic anhydrase. CRC Crit. Rev. Biochem.,14:207-255, 1983.
36. MAREN, T.H. Carbonic anhydrase: Chemistry, physiology, andinhibition. Physiol. Rev., 47:595-781, 1967.
37. DOBYAN, D.C. and BULGER, R.E. Renal carbonic anhydrase. Am. J. Physiol., 243:F311-F324, 1982.
38. PREISIG, P.A.; TOTO, R.D. and ALPERN, R.J. Carbonic anhydraseinhibitors. Renal Physiol., 10:136-159, 1987.
39. CURTHOYS, N.P. and LOWRY, O.H. The distribution of glutami-nase isoenzymes in the various structures of the nephron in normal,acidotic, and alkalotic rat kidney. J. Biol. Chem.; 248:162-168, 1973.
40. GOOD, D.W. and BURG, M.B. Ammonia production by individualsegments of the rat nephron. J. Clin. Invest., 73:602-610, 1984.
41. ALPERN, R.J.; COGAN, M.G. and RECTOR, F.C.J. Flow dependenceof proximal tubular bicarbonate absorption. Am. J. Physiol ., 245:F478-
F484, 1983.42. SCHWARTZ, G.J. and AL-AWQATI, Q. Carbon dioxide causesexocytosis of vesicles containing H pumps in isolated perfused pro-ximal and collecting tubules. J. Clin. Invest., 75:1638-1644, 1985.
43. HAYS, S.R. Mineralocorticoid modulation of apical and basolateralmembrane H/OH/HCO3 transport processes in the rabbit innerstripe of outer medullary collecting duct. J. Clin. Invest., 90:180-187,1992.
44. BAUM, M.; MOE, O.W.; GENTRY, D.L. and ALPERN, R.J. Effect ofglucocorticoids on renal cortical NHE-3 mRNA. Am. J. Physiol.,267:F437-F442, 1994.
ENDEREÇOS RELEVANTES NA INTERNET
http://www.qldanaesthesia.com/AcidBaseBook/AB8-5.htmhttp:// www.ha.org .hk/tmh/medicine/rta.htmlhttp://www.niddk.nih.gov/health/kidney/pubs/rta/rta.htm

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 90/121
Capítulo
6Mecanismo de Concentração e de Diluição Urinária
Antonio José Barros Magaldi
INTRODUÇÃO
FORMAÇÃO DA MEDULA HIPERTÔNICA
AÇÃO DO HORMÔNIO ANTIDIURÉTICOBALANÇO HÍDRICO
INTRODUÇÃO
O estudo do mecanismo de concentração e de diluiçãourinária constitui um dos capítulos mais fascinantes da fi-siologia renal. Os recentes avanços na metodologia de pes-quisa e as admiráveis descobertas acerca da secreção e do
mecanismo de ação do hormônio antidiurético mostramcomo o rim, com um mínimo gasto de energia, conseguevariar a osmolaridade da urina e a excreção de água deacordo com as necessidades do organismo.
A eliminação de urina concentrada resulta da reabsor-ção de água no ducto coletor. Para que esta reabsorçãoaconteça, dois fatos devem ocorrer: 1.º) formação de me-dula hipertônica em relação ao fluido do ducto coletor e2.º) permeabilidade do ducto coletor à água aumentadapelo hormônio antidiurético (HAD). Portanto, a análise domecanismo de concentração e diluição urinária resume-seno estudo do processo pelo qual o rim acumula solutos nointerstício medular durante os estados hidropênicos e omodo de ação do hormônio antidiurético.
Pontos-chave:
Condições para a reabsorção de água no
ducto coletor medular
• Formação de uma medula hipertônica• Ação do hormônio antidiurético
SECREÇÃO DE HAD
REGULAÇÃO DA INGESTA — MECANISMO DA SEDE
BIBLIOGRAFIA SELECIONADAENDEREÇO RELEVANTE NA INTERNET
FORMAÇÃO DA MEDULAHIPERTÔNICA
O estudo da medula renal de animais em estado de res-trição aquosa mostra que a hipertonicidade aí existentedecorre, fundamentalmente, da acumulação de uréia esolutos, principalmente NaCl. O mecanismo pelo qual es-tes solutos se depositam no interstício medular foi genial-mente idealizado em 1942 por Werner e Kuhn, com a hi-pótese da existência de um sistema de contracorrentemultiplicador nos ramos em “U” da alça de Henle. Estesistema produziria um aumento progressivo da osmolari-dade da medula renal do córtex em direção à papila, compouco gasto de energia. Esse modelo foi baseado no siste-ma multiplicador de calor, utilizado na indústria, ondeuma fonte constante de calor aquece o fluido em um pon-to na alça de um tubo em forma de “U”, promovendo umaumento progressivo da temperatura desse fluido, semgrande consumo de energia. Este tubo dobrado e justapostoum ao lado do outro faz com que exista um fluxo do mes-
mo fluido em sentidos opostos proporcionando troca decalor contínua a partir do ponto que recebe o calor, forman-do um gradiente de temperatura (Fig. 6.1). Um sistemasemelhante existe nos membros inferiores das aves pernal-tas que ficam com os pés mergulhados em águas de baixatemperatura, onde a artéria descendente fica justaposta àsveias ascendentes, ajudando a aumentar gradualmente atemperatura do sangue que se dirige dos pés ao coração.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 91/121
capítulo 6 59
No rim, este tubo dobrado corresponde às alças descenden-tes e ascendentes de Henle.
O sistema de contracorrente multiplicador inicialmen-te idealizado e aplicado à medula renal é apresentado naFig. 6.2. A energia inicial que movimentaria este sistemaseria dada pelo transporte ativo de NaCl da luz tubularpara o interstício medular na porção ascendente da alça de
Henle. Este transporte de NaCl é que, aumentando a os-molaridade do interstício, promoveria a reabsorção deágua no ramo descendente da alça, com conseqüente au-mento progressivo da osmolaridade do seu fluido tubularem direção à papila. Este efeito inicial seria multiplicado eo gradiente osmótico então criado determinaria maior re-absorção de água no ducto coletor.
Alguns estudos experimentais que se seguiram à pro-posta do sistema de contracorrente multiplicador na me-dula renal foram compatíveis com a sua existência. Assim,observou-se que o fluido no início do túbulo distal é hipo-tônico (100 mOsm/kg H2O) em relação ao filtrado glome-rular (289 mOsm/kg H2O) e que está de acordo com a exis-tência de uma reabsorção ativa de NaCl, na ausência detransporte de água no ramo ascendente da alça de Henle.
Observou-se, também, que o aumento da osmolaridade damedula externa em direção à papila é diretamente propor-cional ao comprimento da alça de Henle do animal emestudo. São semelhantes as osmolaridades dos fluidos co-lhidos dos vasa recta e da porção fina descendente da alçade Henle.
A maioria das proposições para explicar o mecanismode contracorrente foram elucidadas através de estudos efe-
tuados com a técnica de microperfusão em porções isola-das do néfron de coelhos, que permitiram a análise diretadas características de permeabilidade e de transporte nos
segmentos medulares do néfron. Os estudos funcionais daporção espessa da alça de Henle, tanto da região medularcomo da região cortical (segmento diluidor), mostraram sereles impermeáveis à água, mesmo na presença de hormô-nio antidiurético.
Neste segmento, na membrana luminal ocorre uma re-absorção de Na acoplado a Cl e K em um co-transporteNa:K:2Cl, secundariamente ativo ao transporte de Na ati-vo pela Na-K-ATPase na membrana basolateral. Este trans-porte ativo propicia um gradiente eletroquímico favorávelà entrada de Na na célula. A passagem de Na da luz tubu-lar para o interstício, retirando Na do fluido filtrado e adi-cionando-o ao interstício, constitui o chamado efeito uni-Fig. 6.1 Princípio da contracorrente. Tanto no sistema reto A quan-
to no sistema dobrado B a fonte de calor produz a mesma quan-tidade de calorias; no entanto, no sistema B ocorre a formação de
um gradiente de temperatura, em decorrência da conformaçãode dois tubos justapostos com fluxos inversos. Este modelo ex-plica a formação de gradiente de osmolaridade que ocorre namedula renal com a conformação idêntica à existente na alça deHenle. Adaptado de Berliner, R.W., Lewinsky, N.G., Davidson,D.G., Eden, M. Am. J. Med., 24:730-744, 1958.
Fontede
Calor
Fluido30°
40°
40°
A40° Fonte de Calor
30°
Fluido30°
30°
60°
90°
40°
70°
100°
B
Fig. 6.2 Mecanismo de concentração urinária. 1) Transporte ativode cloreto de sódio no ramo ascendente espesso da alça de Henle— efeito inicial — aumentando a quantidade de sódio no interstício.2) Reabsorção de água no ducto coletor cortical, medular externoe interno, na presença de HAD ocasionado pelo gradiente osmóti-co resultante do transporte ativo de NaCl na porção espessa ascen-dente. 3) Reabsorção de água na alça descendente de Henle frenteao gradiente osmótico entre a luz tubular e a medula. 4) Adição deuréia à medula interna na presença de HAD. 5) Efluxo de NaCl daporção fina ascendente, na ausência de transporte de água, aumen-tando a osmolaridade medular e multiplicando o efeito inicial. 6)Aumento da reabsorção de água no ducto coletor medular inter-no, decorrente do aumento da tonicidade intersticial dado pelo sis-tema multiplicador. 7) Reabsorção de solutos e de água pelos vasarecta, fazendo a recirculação de uréia e mantendo a medula hiper-tônica.
7
MEDULAEXTERNA
VASARECTA
7
3
CÓRTEX
MEDULAINTERNA
NaCl
H2O
Uréia
NaCl
7
Uréia
UréiaH2ONaCl
H2O
H2O
H2O
H2O
NaCl
NaCl
NaCl
Uréia
NaCl
H2O
3
Uréia
NaCl
H2O
Uréia
NaCl
NaCl
NaCl
NaCl
NaCl
Uréia
NaCl
H2O
Uréia
6
4
5
Uréia
NaCl
UréiaHAD
H2O
4
H2O
2H2O
6
5 2
UréiaH2O
6NaCl
HAD
HAD Uréia
HAD
H2O
H2O
HAD
2
1
NaCl
NaCl
HADUréia
H2O
H2O
HAD
1 2
NaCl
Uréia

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 92/121
60 Mecanismo de Concentração e de Diluição Urinária
tário do mecanismo de contracorrente multiplicador. Esteco-transportador Na:K:2Cl já foi clonado e seqüenciadopela técnica de biologia molecular. A porção espessa ascen-
dente da alça de Henle é uma região importante para omecanismo de concentração urinária e é o local de ação doschamados diuréticos de alça, como a furosemida e a bu-metanida, que se ligando ao sítio do íon Cl promovem ainibição do co-transportador Na:K:2Cl.
Os ramos finos da alça de Henle, tanto ascendentescomo descendentes, são formados por um epitélio simplesescamoso, que repousa sobre uma membrana basal, e ocitoplasma de suas células é escasso em mitocôndrias. Estepadrão morfológico é de um epitélio favorável ao equilí- brio osmótico entre o lúmen e o interstício, e não a umtransporte ativo com gasto de energia.
O estudo funcional da porção fina descendente da alçade Henle mostra que este ramo é altamente permeável àágua e pouco permeável ao sódio e a outros solutos, suge-
rindo que o equilíbrio osmótico com o interstício medularocorra à custa da reabsorção de água, com conseqüenteaumento da concentração de cloreto de sódio, uréia e ou-tros solutos no fluido tubular, em direção à papila renal,como está apresentado na Fig. 6.2.
A porção fina ascendente da alça de Henle apresentacaracterísticas opostas às descritas acima para o ramo des-cendente. Observa-se que a porção ascendente é imperme-ável à água e é altamente permeável a Na e Cl, sendo queo movimento transtubular de cloretos deve ocorrer por ummecanismo passivo facilitado.
Nestas condições, a mudança de características de per-meabilidade à água e solutos nos ramos finos descenden-tes e ascendentes permite que o acúmulo de NaCl que
ocorre na porção descendente da alça de Henle por reab-sorção de água se desfaça, pelo menos em parte, na por-ção fina ascendente, como ilustra a Fig. 6.2. No entanto,neste segmento ascendente o equilíbrio osmótico com ointerstício medular dá-se à custa do efluxo de NaCl rápi-do e influxo de uréia mais lento, o que resulta na forma-ção de um fluido tubular com menor concentração deNaCl que o interstício. Este fluido, agora atingindo a por-ção espessa ascendente onde ocorre grande reabsorção deNaCl ativamente, ficará cada vez mais hipotônico, sendoque a sua osmolaridade pode atingir valores inferiores a100 mOsm/kg H2O no início do túbulo distal. Assim, poreste fato, este segmento é chamado de segmento diluidor.Vê-se, portanto, que apenas as características opostas depermeabilidade dos ramos finos, descendentes e ascen-
dentes, proporcionam um meio genial de adicionar solu-to (NaCl) ao nível da região medular interna e de formarum fluido hipotônico à custa, unicamente, da reabsorçãoativa de NaCl da região medular externa, como estáesquematizado na Fig. 6.2.
Esses dados sobre as características de transporte deNa, Cl, H2O e uréia nas várias porções da alça de Henlesão capazes de explicar, pelo menos qualitativamente, o
acúmulo de NaCl e uréia no interstício papilar. Entretan-to, como descrevemos no início, a hipertonicidade medu-lar se faz à custa de NaCl e uréia e, por conseguinte, preci-
samos explicar como se forma o gradiente túbulo-intersti-cial desse soluto (uréia). Como veremos a seguir, ele é tam- bém o resultado de diferenças nas características de per-meabilidade dos vários segmentos medulares e principal-mente das várias porções do túbulo coletor.
O papel importante da uréia no mecanismo de concen-tração urinária já era conhecido de longa data pelas obser-vações de que animais submetidos a dieta pobre em pro-teínas tinham menor capacidade de formar urina hipertô-nica. Contudo, foi só recentemente que as investigaçõesacerca do transporte de solutos nos vários segmentos donéfron trouxeram a explicação para esse fato.
O mecanismo de conservação de uréia no rim é dado porvias de recirculação indicadas na Fig. 6.3. A uréia filtradapelo glomérulo e não reabsorvida pelo túbulo contornado
proximal junta-se à secretada pela pars recta antes de atin-gir a porção fina descendente da alça de Henle. Nesse seg-mento, o equilíbrio osmótico com o interstício se faz prin-cipalmente à custa da saída de água e aumento da concen-tração de solutos do fluido tubular. No rato a permeabili-dade à uréia aí existente permite que, em parte, ocorra in-fluxo desse soluto, elevando ainda mais a concentraçãoluminal. A seguir, no ramo fino ascendente, relativamen-te permeável à uréia, impermeável à água e altamente per-meável a Na e Cl, o equilíbrio osmótico com o interstí-cio se faz à custa de saída rápida de NaCl e entrada lentade uréia. Vemos, portanto, que na porção fina ascendenteda alça de Henle ocorre adição de uréia ao fluido tubular.Por outro lado, no ramo espesso ascendente, túbulo distal
e túbulo coletor cortical não temos nenhum movimento
Fig. 6.3 Recirculação da uréia. Mecanismo pelo qual alta concen-tração de uréia é mantida na medula (1) à custa da sua difusãoda luz do ducto coletor medular interno para a papila e reabsor-ção ao nível da porção fina ascendente da alça de Henle (etapas 1a 7) e (2) à custa da sua retirada do interstício pelos vasa recta,sendo novamente filtrada e lançada na luz tubular. Adaptado deValtin, H. e Schafer, J.A. Renal Function, Little, Brown and Com-pany, 1995.
zonaexterna
Rotas de Recirculaçãoda Uréia
Néfrons Corticais eJustamedulares
MedulaExterna
zonainterna
MedulaInterna
Córtex

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 93/121
capítulo 6 61
transtubular de uréia. No túbulo coletor distal, a reabsor-ção de água em presença de hormônio antidiurético deter-mina elevação na concentração luminal de uréia até atin-
gir o coletor papilar. Nesta porção final do coletor existepermeabilidade transtubular à uréia que permite que estesoluto mais concentrado na luz tubular eflua para o inters-tício papilar. Esta uréia adicionada ao interstício medularpromoverá maior reabsorção de água no ramo fino descen-dente da alça de Henle, acionando, ainda mais, o mecanis-mo de contracorrente multiplicador passivo. O equilíbrioosmótico medular é conseguido pela circulação sanguínealenta e pela entrada de uréia, novamente, para a alça finaascendente, conservando-a dentro do néfron.
Segundo alguns autores, o epitélio que separa a papilarenal da pelve é constituído por células relativamente per-meáveis à uréia, que permitiriam que uma parte desse so-luto eliminado pela urina se retrodifunda para a papilarenal, constituindo outro mecanismo para conservação de
soluto dentro da medula renal.No processo de formação da medula hipertônica os vasa
recta possuem um papel importante, pois deve existir umatroca intensa entre o interstício medular e a luz dos vasosque penetram neste interstício para que se mantenha o gra-diente de concentração medular (Fig. 6.4). Cerca de 5% dofluxo renal plasmático são dirigidos para os vasos da me-dula externa e interna, e como o fluxo plasmático renal é alto,o fluxo plasmático nos vasa recta descendente e ascendenteé cerca de 10 vezes mais intenso que o fluxo do fluido tubu-
lar no começo do ducto coletor medular externo, isto é, en-tra 10 vezes mais plasma que fluido tubular numa mesmaregião da medula. A alta permeabilidade à água e a solutos
de suas paredes, associada à sua disposição em forma dehairpin, como a alça de Henle, permite que seja possível aremoção de água e solutos do interstício medular tambématravés de um mecanismo de troca em contracorrente semalterar a formação do gradiente de concentração medular eauxiliando diretamente o mecanismo de contracorrentemultiplicador que ocorre na luz tubular. Trabalhos publi-cados recentemente evidenciaram nestes vasos a existênciade receptores do tipo V1 e V2 da vasopressina, mostrandoque este hormônio também pode regular o fluxo medular— a estimulação do receptor V1 diminui o fluxo medular,enquanto a estimulação do receptor V2 aumentaria este flu-xo — e evidenciaram também canais de água do tipoaquaporin 1 nos vasa recta descendentes.
Vemos, portanto, que esquematicamente a formação deuma medula hipertônica consta de duas partes: A) umarelacionada com as diferenças de permeabilidade a NaCle água nos ramos finos da alça de Henle, que leva à adiçãode NaCl ao interstício papilar a partir da reabsorção deNaCl na porção espessa ascendente, e B) outra que deter-mina a adição de uréia ao interstício papilar, resultante dasdiferenças de permeabilidade à uréia entre o túbulo cole-tor cortical e o ducto coletor papilar. Esta uréia adiciona-da à papila constitui uma segunda força que promove areabsorção de água no ramo fino descendente da alça deHenle, acelerando o mecanismo de contracorrentemultiplicador passivo aí localizado. Estas idéias aqui apre-sentadas com base nos estudos experimentais constituemo modelo de contracorrente multiplicador atualmente acei-
to e que é esquematizado na Fig. 6.2.
Pontos-chave:
• Heterogeneidade tubular• Efeito unitário na porção espessa da alça de
Henle• Sistema de contracorrente multiplicador• Recirculação da uréia
AÇÃO DO HORMÔNIOANTIDIURÉTICO
O conhecimento dos eventos celulares envolvidos na açãodo hormônio antidiurético (HAD) expandiu-se considera-velmente nos últimos anos. O HAD é um hormônio capazde induzir alterações estruturais na parede luminal das cé-lulas principais, determinando um aumento da permeabi-lidade à água e à uréia. O HAD, que evoca a resposta celu-lar, é o “primeiro mensageiro”, e o seu efeito intracelular é
Fig. 6.4 Contracorrente nos vasa recta. Os números referem-se àsosmolaridades (mOsm/kg) no sangue e no fluido intersticial.Note-se que estes vasos propiciam a retirada da medula de partedos solutos (principalmente NaCl e uréia) e da água, ajudandona formação e na manutenção da medula hipertônica. Adaptadode Berliner, R.W., Lewinsky, N.G., Davidson, D.G. and Eden,M.A. Am. J. Med., 24:730-744, 1958.
Troca em contracorrente nos vasa recta
400
600
450
750
1.200
1.050
900
1.025
875
725
575
425
350
291
1.200
1.200
1.050
900
750
600
450
400
325
1.200
1.075
925
775
625
475Medula Externa
Córtex
Medula Interna
Papila
Difusão passiva de solutos
Difusão passiva de água

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 94/121
62 Mecanismo de Concentração e de Diluição Urinária
mediado por um “segundo mensageiro”, que é produzidocomo resultado da interação do hormônio com o seu recep-tor específico. Os dois mais importantes sistemas de “segun-
dos mensageiros” conhecidos são os sistemas da adenosinamonofosfato cíclico (AMPc) e o do Ca. O HAD exerce seuefeito hormonal estimulando dois tipos de receptores, cha-mados de V1 e V2, sendo que respectivamente utilizam oCa e o AMPc como “segundos mensageiros”. Estes recep-tores estão localizados na membrana basolateral da célulaprincipal e quando estimulados determinam alterações bio-químicas intracelulares que, por sua vez, acarretam modifi-cações na membrana luminal modulando ou regulando apermeabilidade à água, como mostra a Fig. 6.5.
O receptor V2 do HAD é uma estrutura inserida namembrana e que contém sete domínios intramembranosos,quatro extracelulares e quatro intracelulares formandoquatro alças intracelulares. O receptor, uma vez estimula-do pela inserção do HAD no seu locus específico, promo-
ve o estímulo do complexo proteína G, que contém trêsunidades: unidades , e , formando um complexoheterotrimérico. Existe uma família de proteínas G, e aproteína G acoplada ao receptor V2 é do grupo s. Este com-plexo de proteína-Gs, através da unidade , é capaz de seligar na guanidina trifosfato (GTP) formando a G s-GTP,que, por sua vez, auxiliada pelas unidades , vão estimu-lar uma enzima chamada de adenilciclase (AC). A AC é
uma estrutura complexa que também está inserida namembrana celular e contém doze domínios intramembra-nosos divididos em dois grupos de seis domínios, mais oito
domínios extracelulares e oito domínios intracelulares. AAC que atua na cascata do HAD é a de número IV e per-tence a uma família de nove componentes. A ação da AC écatalisar a passagem da adenosina trifosfato (ATP) paraadenosina monofosfato cíclico (AMPc, 3',5',AMPc), já re-ferida acima como sendo o segundo mensageiro do HAD.A quantidade de AMPc intracelular é regulada pela fosfo-diesterase, que é uma enzima que o transforma em umaforma inativa, o 3' AMPc. Prosseguindo na ativação dacascata do HAD, o AMPc vai estimular a proteinoquinaseA (PKA), que é uma proteína multimérica que contém nasua forma inativa duas subunidades catalíticas e duas su- bunidades reguladoras. A unidade reguladora é compos-ta por quatro tipos de proteína ( I e II e I e II), enquantoa unidade catalítica é composta por três tipos (, e ).
Quando o AMPc se liga nas unidades reguladoras, estasse dissociam das unidades catalíticas, resultando na ativi-dade quinásica das subunidades catalíticas. A PKA vaientão fosforilar canais de água que se encontram inseridosna superfície de microvesículas livres do citoplasma. Pro-teínas dos microtúbulos (dineínas e dinactinas) e dos mi-crofilamentos, que são sistemas motores citosólicos, e re-ceptores localizados na superfície destas vesículas (VAMP-
Fig. 6.5 Esquema mostrando uma célula principal do ducto coletor e a translocação das vesículas contendo AQP. A cascata do HADpromove a formação de PKA que vai fosforilar a AQP contida nas vesículas. Proteínas motoras dos microtúbulos (dineínas e dinactinas)e receptores localizados nas vesículas (Vamp-2, sintaxina-4 e NSF) participam da fixação das vesículas na membrana luminal. APKA, acredita-se, também agiria no núcleo celular fosforilando fatores nucleares (CREB-P e AP-1) e aumentando a transcrição gê-nica de AQP, resultando na sua síntese e na sua liberação para o citosol, e entrando no processo de traficking e docking. Adaptado deNielsen, S., Kwon, T.-H., Christensen, B.M., Promeneur D., Frøkiær, J., Marples, D. JASN , 10:647-663,1999.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 95/121
capítulo 6 63
2, sintaxina-4, NSF) participam do processo de trafficking edocking, isto é, de translocação destas vesículas em direçãoà membrana celular e que termina com a sua inserção na
membrana apical da célula, expondo finalmente os canaisde água na superfície luminal e aumentando a permeabi-lidade à água (Fig. 6.5).
No modelo da Fig. 6.6 a AC estaria ligada a dois recep-tores de naturezas opostas: um deles a estimularia (Rs),enquanto o outro a inibiria (Ri), através das unidades re-guladoras da proteína G, respectivamente Gs e Gi. Estesreceptores ativariam (Gs) ou inibiriam (Gi) a adenilcicla-se, quando o receptor estimulador ou inibidor fossem ocu-pados, respectivamente. O receptor do HAD é o receptorestimulador (Rs), enquanto o receptor ocupado pelos agen-tes -2 adrenérgicos seria inibidor (Ri), uma vez que estesagentes inibem o transporte de água. Este processo, noentanto, pode ser modulado intracelularmente, como já foidito acima, pela atividade da AMPc fosfodiesterase (que
transformaria o AMPc na sua forma inativa, a 5’adenosi-na monofosfato, 5’AMPc), bem como por autacóides comoas prostaglandinas e por outras substâncias como o Ca
e a proteinoquinase C (PKC). No Quadro 6.1 podemos vervárias substâncias que estão envolvidas na geração doAMPc e na sua modulação.
Recentemente foi descrita a presença de receptores dotipo V1 nas células principais dos túbulos distais (Fig. 6.5).Este receptor, quando ocupado pelo HAD, desencadeariauma reação em cascata da seguinte forma: ativação de umafosfolipase C (PLC) de membrana que clivaria o fosfatidil-inositol-bifosfato (PIP2) em dois segundos mensageiros —o diacilglicerol (DAG) e o inositol-trifosfato (ITP). O DAG, junto com os Ca, ativaria uma proteinoquinase C (PKC),e o ITP estimularia a liberação de cálcio das organelas parao citosol. O aumento do cálcio intracelular mais a PKC re-gulariam a atividade da adenilciclase, exercendo sobre eleum efeito inibitório.
O HAD também estimula uma fosfolipase A de mem- brana que, agindo sobre o ácido araquidônico (AA), trans-forma-o em prostaglandina (PGE2), que por sua vez temum efeito inibitório sobre a adenilciclase, constituindodesta forma um sistema de feedback negativo modulando
a ação do próprio HAD.Com a técnica recente da biologia molecular, foi de-monstrado que existem vários tipos de canais de água noreino animal. Muitos são proteínas de baixo peso molecu-lar (25.000 a 30.000 daltons) que pertencem a famílias decanais de água chamadas MIP 26 ( Membrane Integral Pro-tein com PM 26.000 daltons). São encontrados em grandevariedade em tecidos transportadores de fluidos, como o
Fig. 6.6 Representação esquemática dos efeitos da vasopressinanas células do DCMI. Abreviações: V1 e V2 — subtipo de recep-tores; Rs e Ri — receptores para agentes estimuladores e inibido-res, agindo sobre a adenilciclase; Gs e Gi — unidades guanina re-guladoras, estimuladora e inibidora; AC — unidade catalítica ade-nilciclase; AMPc — adenosina monofosfato cíclica; ATP — ade-nosina trifosfato; PGE2; PL — pool de fosfolípides; AA — ácidoaraquidônico; PLA — fosfolipase A; PLC — fosfolipase C; PIP 2
— fosfatidilinositol-4-5-bifosfato; ITP — inositol-trifosfato; Cacálcio citosólico livre. Adaptado de Kinter, L.B., Huffman, W.F.,Stassen, F.L. Am. J. Physiol., 254:F165-177, 1988.
2-agonista
O2
Ri
Gi
PLCPLA
vasopressina
PGE2
EXTRA-CELULAR
MEMBRANACELULAR
CITOSOL
MEMBRANABASOLATERAL
Gs
Rs
V2 PGE2
V1
AC PGE2
PGE2 AA PL
PIP2ITP
EFEITO V1
EFEITO V2
(SUPERFÍCIE APICAL)
PROTEINOQUINASE
ATP AMPc
Ca++
Quadro 6.1 Eventos que envolvem a geraçãode AMPc
A nível de receptorOcupação do receptor V2
Agonistas — DDAVP (desmopressina)Antagonistas — d (CH2) 5 Tyr (Et) VAVP etc.*[Ca] sérico
Ocupação do receptor V1 — ativação da via dofosfoinositolAVPAgentes 1-adrenérgicosSomatostatinaAcetilcolinaCarbacol
Modulação do complexo adenilciclase A nível da proteína reguladora
GTPToxina da cólera (Ns)Toxina pertussis (Ni)
Prostaglandina (Ns)Bradicinina (Ni)Agentes 2-adrenérgicos
Ao nível da unidade catalítica (adenilciclase)Forskolin (Ni)Calmodulina (?)
Ao nível ainda não determinadoFator atrial natriurético
*Atualmente são conhecidos inúmeros agonistas e antagonistas do hor-mônio antidiurético. Adaptado de Abramov. M. et al. Kidney Int., 32 Su-ppl 21:S56-S66, 1987.
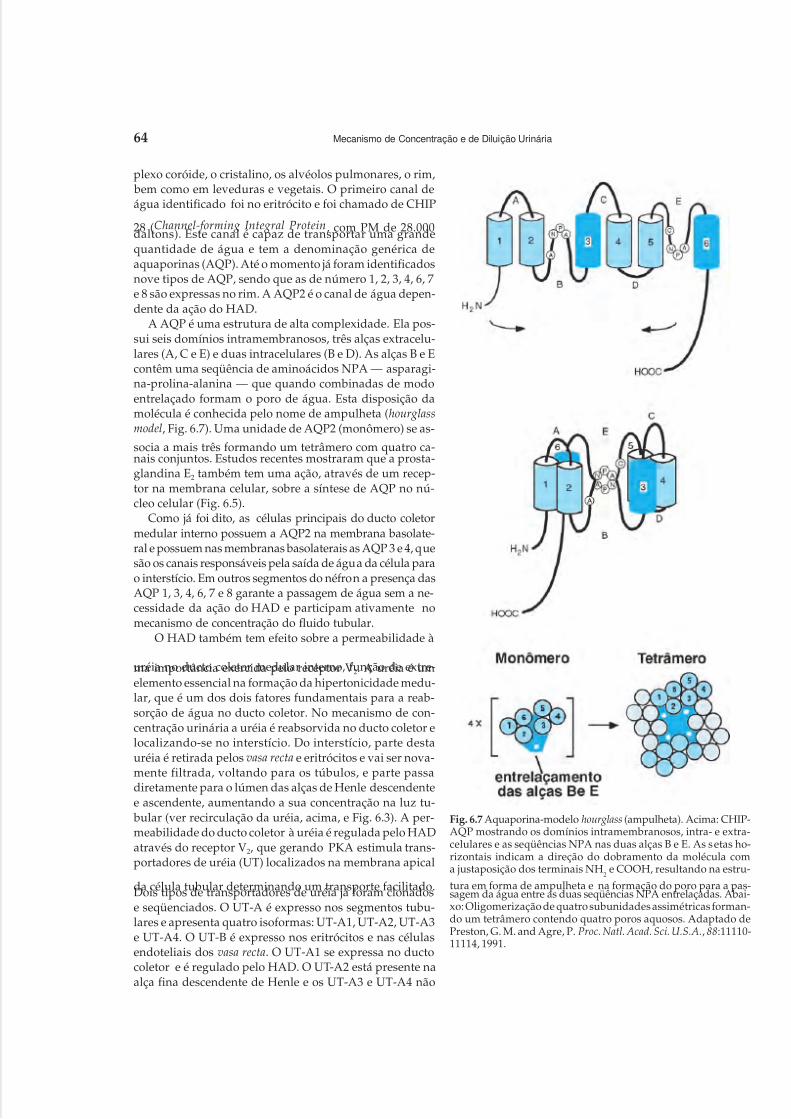
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 96/121
64 Mecanismo de Concentração e de Diluição Urinária
plexo coróide, o cristalino, os alvéolos pulmonares, o rim, bem como em leveduras e vegetais. O primeiro canal deágua identificado foi no eritrócito e foi chamado de CHIP
28 (Channel-forming Integral Protein, com PM de 28.000daltons). Este canal é capaz de transportar uma grandequantidade de água e tem a denominação genérica deaquaporinas (AQP). Até o momento já foram identificadosnove tipos de AQP, sendo que as de número 1, 2, 3, 4, 6, 7e 8 são expressas no rim. A AQP2 é o canal de água depen-dente da ação do HAD.
A AQP é uma estrutura de alta complexidade. Ela pos-sui seis domínios intramembranosos, três alças extracelu-lares (A, C e E) e duas intracelulares (B e D). As alças B e Econtêm uma seqüência de aminoácidos NPA — asparagi-na-prolina-alanina — que quando combinadas de modoentrelaçado formam o poro de água. Esta disposição damolécula é conhecida pelo nome de ampulheta (hourglassmodel, Fig. 6.7). Uma unidade de AQP2 (monômero) se as-
socia a mais três formando um tetrâmero com quatro ca-nais conjuntos. Estudos recentes mostraram que a prosta-glandina E2 também tem uma ação, através de um recep-tor na membrana celular, sobre a síntese de AQP no nú-cleo celular (Fig. 6.5).
Como já foi dito, as células principais do ducto coletormedular interno possuem a AQP2 na membrana basolate-ral e possuem nas membranas basolaterais as AQP 3 e 4, quesão os canais responsáveis pela saída de água da célula parao interstício. Em outros segmentos do néfron a presença dasAQP 1, 3, 4, 6, 7 e 8 garante a passagem de água sem a ne-cessidade da ação do HAD e participam ativamente nomecanismo de concentração do fluido tubular.
O HAD também tem efeito sobre a permeabilidade à
uréia no ducto coletor medular interno, função de extre-ma importância exercida pelo receptor V2. A uréia é umelemento essencial na formação da hipertonicidade medu-lar, que é um dos dois fatores fundamentais para a reab-sorção de água no ducto coletor. No mecanismo de con-centração urinária a uréia é reabsorvida no ducto coletor elocalizando-se no interstício. Do interstício, parte destauréia é retirada pelos vasa recta e eritrócitos e vai ser nova-mente filtrada, voltando para os túbulos, e parte passadiretamente para o lúmen das alças de Henle descendentee ascendente, aumentando a sua concentração na luz tu- bular (ver recirculação da uréia, acima, e Fig. 6.3). A per-meabilidade do ducto coletor à uréia é regulada pelo HADatravés do receptor V2, que gerando PKA estimula trans-portadores de uréia (UT) localizados na membrana apical
da célula tubular determinando um transporte facilitado.Dois tipos de transportadores de uréia já foram clonadose seqüenciados. O UT-A é expresso nos segmentos tubu-lares e apresenta quatro isoformas: UT-A1, UT-A2, UT-A3e UT-A4. O UT-B é expresso nos eritrócitos e nas célulasendoteliais dos vasa recta. O UT-A1 se expressa no ductocoletor e é regulado pelo HAD. O UT-A2 está presente naalça fina descendente de Henle e os UT-A3 e UT-A4 não
Fig. 6.7 Aquaporina-modelo hourglass (ampulheta). Acima: CHIP-AQP mostrando os domínios intramembranosos, intra- e extra-celulares e as seqüências NPA nas duas alças B e E. As setas ho-rizontais indicam a direção do dobramento da molécula coma justaposição dos terminais NH
2e COOH, resultando na estru-
tura em forma de ampulheta e na formação do poro para a pas-sagem da água entre as duas seqüências NPA entrelaçadas. Abai-xo: Oligomerização de quatro subunidades assimétricas forman-do um tetrâmero contendo quatro poros aquosos. Adaptado dePreston, G. M. and Agre, P. Proc. Natl. Acad. Sci. U.S.A., 88:11110-11114, 1991.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 97/121
capítulo 6 65
têm ainda bem definidos os papéis que efetuam, apesar deserem expressos no ducto coletor.
A uréia é o produto final do metabolismo das proteínas
e o seu excesso deve ser eliminado pelo rim. Este processode secreção se dá principalmente no terço final do DCMI enão é dependente da ação do HAD, envolvendo um me-canismo de transporte secundariamente ativo acoplado aosódio, um contratransporte na membrana apical das célu-las destes segmentos.
Pontos-chave:
• Receptor V2-membrana basolateral• Geração de AMPc-segundo mensageiro• Inserção da aquaporina 2 na membrana
luminal
BALANÇO HÍDRICO
O balanço de água do organismo é dado pela quantida-de de água que é ingerida, comparada com a quantidadede água que é excretada. Sob condições basais, as perdashídricas e a ingesta aquosa variam em torno de 2 a 2,5 li-tros. Assim, o balanço aquoso pode ser mantido por lon-gos períodos sem a intervenção de mecanismos regulado-res específicos. No entanto, esta condição ideal pode serrompida pela atividade física, por alterações climáticas, porvariação de dieta ou outras alterações ambientais. Sempreque tais desvios ocorrem, um poderoso mecanismo home-ostático entra em ação, aumentando ou diminuindo a in-
gesta ou a excreção de água e solutos. Esta homeostase sefaz pela regulação da secreção de HAD (eliminação) e pelaregulação da sede (ingestão).
Pontos-chave:
• Água ingerida• Perdas hídricas
SECREÇÃO DE HAD
O principal meio pelo qual o organismo elimina a águasem movimento resultante de solutos ocorre no rim, pela
ação do hormônio antidiurético nos túbulos renais, como já foi mencionado anteriormente.Quimicamente o hormônio antidiurético, na maioria dos
mamíferos, é a arginina vasopressina. Nos suínos é cons-tituído pela lisina vasopressina. Ambos são octapeptídiosde aproximadamente 1.100 daltons. Nos animais vertebra-dos mais inferiores, o hormônio antidiurético é a argininavasotocina. Até o momento já foram identificados sete octa-
peptídios na neuro-hipófise de vertebrados e mais de 200análogos já foram sintetizados. O grande progresso obti-do na química desse hormônio trouxe a descoberta de com-postos sintéticos de variável potência, tempo de ação pro-longada, fácil absorção etc., o que é de extrema importân-cia no tratamento substitutivo nos casos de portadores dediabetes insipidus.
O hormônio antidiurético, ou arginina vasopressina nosmamíferos, é secretado pelos corpos celulares dos neurô-nios existentes nos núcleos supra-ópticos e paraventricu-lares do hipotálamo em forma de grânulos. Há uma estreitacorrelação entre o número desses grânulos nas células ner-vosas secretoras e o estado de hidratação do animal. O hor-mônio antidiurético está como que “empacotado” nessesgrânulos que percorrem o axoplasma dos nervos em dire-ção à glândula pituitária posterior (Fig. 6.9). Dentro des-ses grânulos o hormônio antidiurético está ligado a umaproteína específica chamada neurofisina A ou neurofisinaII, formando um complexo. Ambos, tanto o hormôniocomo a neurofisina, podem originar-se de um mesmo pre-cursor biológico. As células secretoras da oxitocina naneuro-hipófise também têm grânulos nos quais a oxitoci-
na está ligada a uma outra proteína carregadora, a neuro-fisina B ou neurofisina I. As neurofisinas são cadeias depolipeptídios contendo 90 a 100 aminoácidos de aproxima-damente 10.000 daltons. Estudos com a técnica de freeze- fracture e estudos eletromicroscópicos mostram que a se-creção na neuro-hipófise ocorre por exocitose. Tem sidosugerido que na neuro-hipófise existem dois pools de hor-mônio antidiurético: um pronto para ser liberado e outro
Fig. 6.8 Modelo esquemático representando o CHIP-aquaporin
inserido na membrana de bicamada lipídica compreendendo umcomplexo homotetramérico com uma subunidade de polilacto-saminoglicano e possíveis canais de água individuais dentro decada subunidade. Adaptado de Agre, P. et al. Am. J. Physiol.,265:F463-476, 1993.
H2O
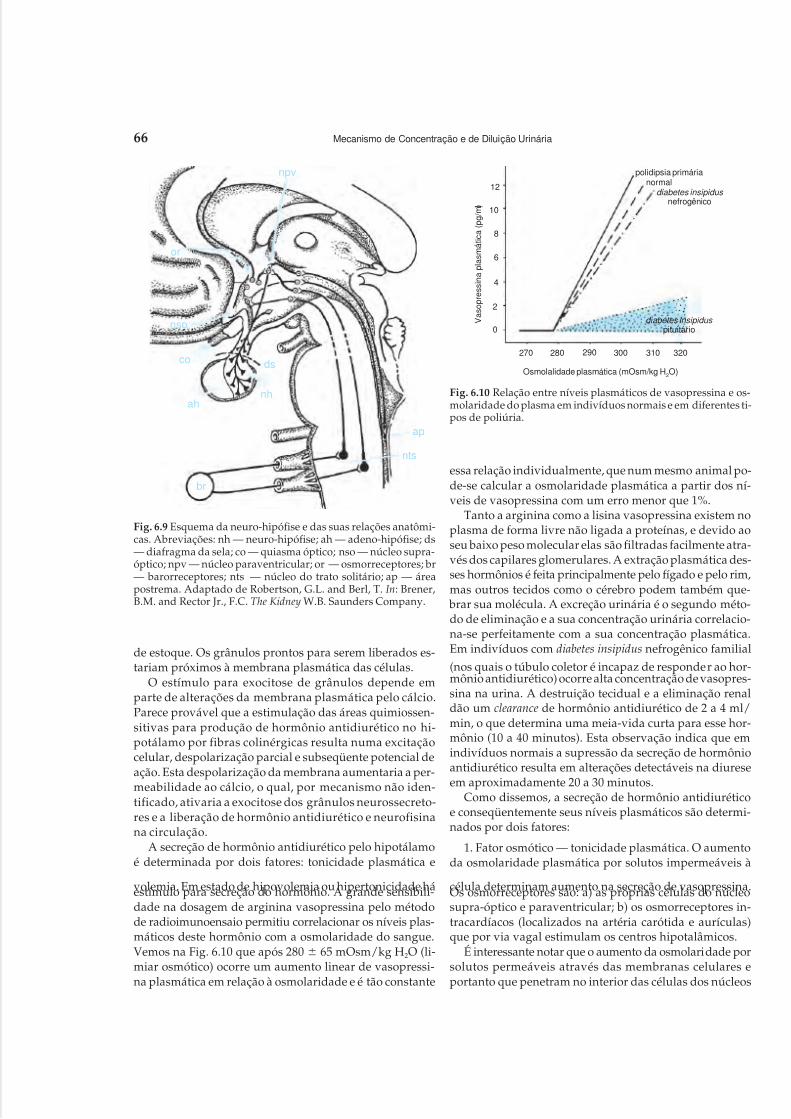
5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 98/121
66 Mecanismo de Concentração e de Diluição Urinária
de estoque. Os grânulos prontos para serem liberados es-tariam próximos à membrana plasmática das células.
O estímulo para exocitose de grânulos depende emparte de alterações da membrana plasmática pelo cálcio.Parece provável que a estimulação das áreas quimiossen-sitivas para produção de hormônio antidiurético no hi-potálamo por fibras colinérgicas resulta numa excitaçãocelular, despolarização parcial e subseqüente potencial deação. Esta despolarização da membrana aumentaria a per-meabilidade ao cálcio, o qual, por mecanismo não iden-tificado, ativaria a exocitose dos grânulos neurossecreto-res e a liberação de hormônio antidiurético e neurofisinana circulação.
A secreção de hormônio antidiurético pelo hipotálamoé determinada por dois fatores: tonicidade plasmática e
volemia. Em estado de hipovolemia ou hipertonicidade háestímulo para secreção do hormônio. A grande sensibili-dade na dosagem de arginina vasopressina pelo métodode radioimunoensaio permitiu correlacionar os níveis plas-máticos deste hormônio com a osmolaridade do sangue.Vemos na Fig. 6.10 que após 280 65 mOsm/kg H2O (li-miar osmótico) ocorre um aumento linear de vasopressi-na plasmática em relação à osmolaridade e é tão constante
essa relação individualmente, que num mesmo animal po-de-se calcular a osmolaridade plasmática a partir dos ní-veis de vasopressina com um erro menor que 1%.
Tanto a arginina como a lisina vasopressina existem noplasma de forma livre não ligada a proteínas, e devido aoseu baixo peso molecular elas são filtradas facilmente atra-vés dos capilares glomerulares. A extração plasmática des-ses hormônios é feita principalmente pelo fígado e pelo rim,mas outros tecidos como o cérebro podem também que- brar sua molécula. A excreção urinária é o segundo méto-do de eliminação e a sua concentração urinária correlacio-na-se perfeitamente com a sua concentração plasmática.Em indivíduos com diabetes insipidus nefrogênico familial
(nos quais o túbulo coletor é incapaz de responder ao hor-mônio antidiurético) ocorre alta concentração de vasopres-sina na urina. A destruição tecidual e a eliminação renaldão um clearance de hormônio antidiurético de 2 a 4 ml/min, o que determina uma meia-vida curta para esse hor-mônio (10 a 40 minutos). Esta observação indica que emindivíduos normais a supressão da secreção de hormônioantidiurético resulta em alterações detectáveis na diureseem aproximadamente 20 a 30 minutos.
Como dissemos, a secreção de hormônio antidiuréticoe conseqüentemente seus níveis plasmáticos são determi-nados por dois fatores:
1. Fator osmótico — tonicidade plasmática. O aumentoda osmolaridade plasmática por solutos impermeáveis à
célula determinam aumento na secreção de vasopressina.Os osmorreceptores são: a) as próprias células do núcleosupra-óptico e paraventricular; b) os osmorreceptores in-tracardíacos (localizados na artéria carótida e aurículas)que por via vagal estimulam os centros hipotalâmicos.
É interessante notar que o aumento da osmolaridade porsolutos permeáveis através das membranas celulares eportanto que penetram no interior das células dos núcleos
Fig. 6.9 Esquema da neuro-hipófise e das suas relações anatômi-cas. Abreviações: nh — neuro-hipófise; ah — adeno-hipófise; ds— diafragma da sela; co — quiasma óptico; nso — núcleo supra-óptico; npv — núcleo paraventricular; or — osmorreceptores; br— barorreceptores; nts — núcleo do trato solitário; ap — áreapostrema. Adaptado de Robertson, G.L. and Berl, T. In: Brener,B.M. and Rector Jr., F.C. The Kidney W.B. Saunders Company.
npv
or
nso
co
ah
br
ds
nh
nts
ap
Fig. 6.10 Relação entre níveis plasmáticos de vasopressina e os-molaridade do plasma em indivíduos normais e em diferentes ti-pos de poliúria.
0
2
4
6
8
10
12
Vasopressinaplasm
á tica(pg/ml)
270 280 290 300 310 320
Osmolalidade plasmática (mOsm/kg H2O)
polidipsia primárianormal
diabetes insipidus nefrogênico
diabetes insipidus pituitário

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 99/121
capítulo 6 67
hipotalâmicos não determinam aumento da secreção dehormônio antidiurético. Assim, por exemplo, tanto a infu-são de uréia como o aumento da glicemia no diabetes melli-
tus (em ausência de hipovolemia e desidratação), apesarde aumentarem a osmolaridade do plasma, não acarretamaumento da secreção de hormônio antidiurético. Ao con-trário, a hiperglicemia (na ausência de hipovolemia) deter-mina uma diminuição na liberação de vasopressina, o queindica ser a poliúria do diabetes mellitus o resultado de, pelomenos, dois fatores: diurese osmótica inibição do hor-mônio antidiurético, causando uma menor reabsorção deágua no túbulo e ducto coletor.
2. Fatores não-osmóticos — volemia. Em condições dedepleção de volume, o fator volemia é mais importantecomo estímulo que a osmolaridade plasmática. Assim, emcondições de hipovolemia, mesmo com hipotonicidade doplasma, observa-se um aumento na secreção de hormônioantidiurético. Ao contrário, em condições de hipertonici-
dade (osmolaridade plasmática acima de 280 mOsm/kgH2O) o fator tonicidade predomina, observando-se au-mento da secreção do hormônio mesmo em condições deexpansão do volume extracelular. Os receptores de volu-me para secreção de hormônio antidiurético podem ser di-vididos em: de baixa pressão (localizados no setor veno-so — aurícula direita) e de alta pressão (localizados no se-tor arterial — aurícula esquerda, carótida etc.). A via afe-rente desses receptores é o vago e glossofaríngeo. O siste-ma de baixa pressão é mais sensível que o de alta pressão, bastando ocorrer uma depleção de volume de 10%, mes-mo sem alterações da pressão arterial, para que se obser-ve um aumento de 6 vezes na secreção de hormônio anti-diurético através de estímulos recebidos no sistema de baixa pressão.
Além desses fatores volêmicos e osmóticos, outros,como a ação de drogas vasoativas, levam a alterações nasecreção desse hormônio. É comum a observação de anti-diurese durante a infusão de isoproterenol em animais emdiurese aquosa. Também a infusão de noradrenalina, empequenas doses, pode determinar aumento da diurese poraumento da pressão arterial ou menor reabsorção de águano túbulo coletor.
Inúmeros trabalhos têm chamado a atenção para a par-ticipação do sistema renina-angiotensina na regulação daexcreção urinária de urina. Estudos têm demonstrado quea administração sistêmica ou intracerebral (liquórica) deangiotensina II determina aumento na secreção de hor-mônio antidiurético. Por outro lado, a administração de
HAD exógeno a ratos Brattleboro, que apresentam diabe-tes insipidus hipotalâmico hereditário, produz diminuiçãoda secreção renal e níveis plasmáticos de renina. Foi ve-rificado, também, que o sistema renina-angiotensina podedesempenhar um papel importante no centro reguladorda sede.
Recentemente foi descrito que um heptapeptídio forma-do diretamente da angiotensina I sem a participação da
enzima conversora e denominado de angiotensina 1-7 pos-sui efeitos semelhantes ao do HAD, isto é, é capaz de au-mentar a permeabilidade à água no ducto coletor medu-
lar interno.É importante salientar que em certas condições patoló-gicas observa-se quadro de oligúria (antidiurese) em pre-sença de hipotonicidade plasmática, mesmo com volemianormal ou aumentada, como nos casos de:
1. Síndrome de secreção inapropriada de hormônio anti-diurético que ocorre associada a tumores, patologiaspulmonares, lesões cranianas etc.
2. Decorrente da administração de drogas que estimulama produção de hormônio antidiurético, como morfina, barbitúricos e clofibrato, ou mesmo de drogas que au-mentam a atividade do hormônio, como clorpropami-da e carbamazepina (Tegretol).
3. Endocrinopatias, como o mixedema, no qual os fatores
responsáveis pela incapacidade de eliminar urina dilu-ída permanecem ainda por serem esclarecidos, e nadoença de Addison, onde se observa também uma in-suficiente excreção de água conseqüente à falta de gli-cocorticóides.
A combinação da excreção de urina hipertônica associ-ada à hipotonicidade do sangue ocorre em condições dehipovolemia, depleção ou má distribuição de volume.Observamos, também, em condições de redução da pres-são na aurícula esquerda durante a comissurotomia mitral,ventilação pulmonar prolongada, insuficiência cardíacagrave e cirrose hepática avançada.
Pontos-chave:
• Fator osmótico• Fator volêmico• Outros fatores — secreção inapropriada de
HAD
REGULAÇÃO DA INGESTA –MECANISMO DA SEDE
A sede é a maior defesa do organismo contra a deple-ção dos fluidos corporais e é definida como a sensação quecompele o indivíduo a beber água. É estimulada pelasmesmas variáveis que estimulam o HAD, ou seja, o aumen-
to da osmolaridade plasmática ou a diminuição da vole-mia, mas a hipertonicidade do plasma parece ser maispotente que a hipovolemia. No homem, um aumento deapenas 2% a 3% acima do nível basal produz um desejointenso de ingestão de água. O nível efetivo de osmolari-dade plasmática que provoca um desejo urgente conscientede ingestão de água é chamado de limiar da sede e é ligei-ramente diferente de indivíduo para indivíduo e varia em

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 100/121
68 Mecanismo de Concentração e de Diluição Urinária
torno de 295 mOsm/kg. O limiar para o estímulo da sedeestá ligeiramente abaixo do limiar para o estímulo de libe-ração do HAD. As vias neuronais que medeiam a dipso-
gênese osmótica não estão ainda bem definidas, mas pa-rece que envolvem osmorreceptores localizados na áreaventromedial do hipotálamo próximo àquelas que regulama secreção de HAD e devem ter uma representação no cór-tex cerebral, a fim de permitir que o indivíduo tenha cons-ciência da necessidade de ingerir líquidos. Além dos fato-res já descritos, o sistema renina-angiotensina e mesmo opróprio HAD exercem uma mediação parcial sobre a dip-sogênese.
Pontos-chave:
• Fator osmótico• Fator volêmico
BIBLIOGRAFIA SELECIONADA
AGRE, P. et al. Aquaporin CHIP: the archetypal molecular water channel. Am. J. Physiol., 265:F463, 1993.
BRENNER, B.M. and RECTOR Jr., F.C. (eds) The Kidney, 6th edition. W.B. Saunders Company, Phil., 2000.
KINTER, L.B. et al. Antagonist of the antidiuretic activity of vasopressin. Am. J. Physiol., 254:F165, 1988.
KOVÁCS, L. and LICHARDUS, B. Vasopressin — Disturbed Secretion andits Effects, 1.ª ed., Kluwer Academic Publishers, 1989.
NIELSEN, S. et al. Aquaporins in the kidney: from molecules to medici-ne. Physiol. Ver., 82:205, 2002.
NIELSEN, S. et al. Physiology and pathophysiology of renal aquaporins. JASN , 10:647, 1999.
SANDS, J.M. Regulation of urea transporters. JASN , 10:635, 1999.SANDS, J.M. et al. Urea transporters in the kidney and erythrocytes. Am.
J. Physiol., 273:F321, 1997.
ENDEREÇO RELEVANTE NA INTERNET
http://www.ndif.org

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 101/121
Capítulo
7Peptídeos Vasoativos e o Rim
Irene L. Noronha e Miguel Luis Graciano
INTRODUÇÃO
SISTEMA RENINA-ANGIOTENSINA (SRA)
Componentes do SRA
Angiotensinogênio
Renina
Biossíntese da Renina
ESTRUTURA DO APARELHO JUSTAGLOMERULAR
CONTROLE DA SECREÇÃO DE RENINA
QUANTIFICAÇÃO DA ATIVIDADE DA RENINA
ENZIMA CONVERSORA DE ANGIOTENSINA (ECA)
Propriedades bioquímicas
Distribuição tecidual
ANGIOTENSINA II
Ações na vasculatura
Ações renais
Ações na supra-renal
Ações no sistema nervoso central
Ações da Ang II no crescimento celular
Receptores para Ang II e mecanismo de ação celular
ANGIOTENSINASES
SISTEMA RENINA-ANGIOTENSINA EM SITUAÇÕES
FISIOPATOLÓGICAS
Hipertensão arterial
Nefropatia diabética
Nefropatias crônicas
PEPTÍDEOS DERIVADOS DAS ANGIOTENSINAS
Ang-(I-7)
Ang III
Ang IV
SISTEMA RENAL CALICREÍNA-CININA
Componentes do sistema renal calicreína-cinina
Cininogênios
Calicreínas
Cininas
Cininases
Receptores para cininas
Localização renal dos componentes do sistema
calicreína-cinina
Ações do sistema calicreína-cinina no rim
Efeitos na hemodinâmica renal
Efeitos na excreção de sódio e água
Inter-relações entre sistema calicreína-cinina e outros
sistemas
Sistema renina-angiotensina
Eicosanóides
Peptídeo natriurético atrial (ANP)
Óxido nítrico
SISTEMA CALICREÍNA-CININA EM SITUAÇÕES
FISIOPATOLÓGICAS
Cirrose hepática
Hipertensão arterial
Fibrose miointimal
Infarto do miocárdio
PEPTÍDEOS NATRIURÉTICOS
Peptídeo natriurético atrial (ANP)
Síntese e estrutura
Regulação da secreção do ANP
Receptores para ANP
Ações do ANP
Efeitos na hemodinâmica renal e efeitos na indução de
natriurese
Efeitos no sistema renina-angiotensina-aldosterona,
vasopressina e endotelina
Efeitos na vasculatura
ANP em situações fisiopatológicas
Insuficiência cardíaca congestiva
Taquicardia supraventricular
Hipertensão arterial
Cirrose hepática
Doença renal

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 102/121
70 Peptídeos Vasoativos e o Rim
PEPTÍDEO NATRIURÉTICO CEREBRAL (BNP)
Peptídeo natriurético do tipo C (CNP)
URODILATINA
ENDOTELINA
Biossíntese e estrutura
Receptores para endotelina e mecanismo de ação celular
Ações da endotelina no rim
Endotelina em situações fisiopatológicas
Insuficiência renal aguda
Hipertensão arterial
Ciclosporina A
Doenças glomerulares
Fibrose renal e insuficiência renal crônica
ADRENOMEDULINA
REFERÊNCIAS BIBLIOGRÁFICAS
ENDEREÇOS RELEVANTES NA INTERNET
INTRODUÇÃO
Existe no rim um importante balanço entre agentes va-soconstritores e vasodilatadores, responsável pela regula-ção da filtração glomerular. Dentre os agentes vasocons-tritores devem ser ressaltados: peptídeos como a angioten-sina (Ang II) e endotelina, além de outras substâncias comoadenosina, PAF (fator ativador de plaquetas) e metabóli-tos do ácido araquidônico (PGF2a, tromboxane A2 e leu-cotrienos). Os agentes vasodilatadores incluem: peptíde-os como bradicinina e peptídeos natriuréticos, além deoutras substâncias como óxido nítrico, PGE2, PGI2 e do-pamina. No presente capítulo, serão abordados alguns dosmais relevantes peptídeos vasoativos que desempenhampapel fundamental na regulação da hemodinâmica renal.
Pontos-chave:
Principais peptídeos vasoconstritores
• Angiotensina II• Endotelina
Principais peptídeos vasodilatadores
• Bradicinina• Peptídeo atrial natriurético
SISTEMA RENINA-ANGIOTENSINA (SRA)
O sistema renina-angiotensina (SRA) tem como impor-tante função a regulação da pressão arterial e do volumeintravascular. Este controle é possível graças à ação da an-giotensina II (Ang II), que promove uma potente vasocons-trição na musculatura lisa dos vasos (e assim aumenta aresistência vascular periférica), além de diminuir a excreçãorenal de sódio (mediado pela aldosterona). Deste modo,
mantém uma adequada perfusão capilar principalmentequando ocorrem alterações do volume de água corporal.
A atividade do SRA é regulada pela renina, que é pro-duzida e armazenada no aparelho justaglomerular renale liberada em resposta a uma série de sinais. A renina éuma enzima proteolítica que quebra o angiotensinogênioexistente no plasma, formando o decapeptídeo angioten-sina I (Fig. 7.1). Este peptídeo, biologicamente inativo, éconvertido pela enzima conversora formando o hormônioativo Ang II. A Ang II é clivada por aminopeptidases for-mando a angiotensina III, que é subseqüentemente que- brada em fragmentos inativos.1,2,3 Atualmente tem sidocada vez mais conhecido o papel fisiológico de metabóli-
Fig. 7.1 Componentes do sistema renina-angiotensina-aldosterona.
reabsorção deNa e H2O
ANGIOTENSINA II
ANGIOTENSINOGÊNIO
ANGIOTENSINA I
renina
pró-renina
pressão de perfusão
VASOCONSTRIÇÃO
enzima conversora
síntese deALDOSTERONA

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 103/121
capítulo 7 71
tos das angiotensinas, como por exemplo a ação vasodi-latadora da Ang-(1-7).4
Componentes do SRAANGIOTENSINOGÊNIO
O angiotensinogênio plasmático é uma glicoproteína depeso molecular que varia de 52 a 60 kDa, produzido pri-mariamente no fígado. No entanto, RNAm para angioten-sinogênio também foi encontrado no cérebro, rim, supra-renal, coração, pulmão, vasos e trato gastrintestinal,5 de-monstrando que o angiotensinogênio pode ser produzidoem diversos tecidos extra-hepáticos. O sítio de produçãodo angiotensinogênio pode ter relevância fisiopatológica,uma vez que pode contribuir para a formação local de AngII, que por sua vez pode ser importante na regulação localde diversas funções nos tecidos.
RENINA
Biossíntese da ReninaRenina foi o nome dado em 1898 por Tigerstedt e
Bergamann6 à substância com capacidade pressora extraí-da de rins de coelhos. A renina é uma protease que quebraespecificamente a ligação LEU-VAL da região aminotermi-nal do angiotensinogênio, formando a Ang I.
O gene da renina humana (localizado no cromossoma1) codifica uma proteína precursora de 45 kDa chamadade PRÉ-PRÓ-renina, que rapidamente é quebrada forman-do a PRÓ-renina. Tanto a PRÉ-PRÓ-renina como a PRÓ-reni-na são completamente inativas. A PRÓ-renina é clivadaformando a enzima ativa de 40 kDa denominada renina.
Apesar de o rim ser a principal fonte da produção derenina, existem diversos órgãos que têm a capacidade desintetizar renina: fígado, cérebro, próstata, testículo, baço,timo e pulmão.5
ESTRUTURA DO APARELHOJUSTAGLOMERULAR
O aparelho justaglomerular, situado no hilo glomerular,é formado pelas células justaglomerulares, pela mácula den-sa e por um tecido interposto entre eles, o mesângio extra-glomerular, que é um prolongamento do mesângio glome-rular. As células justaglomerulares são células modificadas
da musculatura lisa encontradas na parede da arteríola afe-rente. A renina é produzida e armazenada nas células jus-taglomerulares, que aparecem à microscopia eletrônicacomo grânulos eletrodensos, sendo secretada por exocito-se. Utilizando-se imuno-histoquímica, foi possível demons-trar que os grânulos intracelulares contêm renina e Ang II.7,8,9
Estudos utilizando-se hibridização in situ demonstraramclaramente uma grande concentração de grânulos negros,
que correspondem ao mRNA para renina, localizados nohilo renal (Fig. 7.2). Tal localização corresponde à topogra-fia das arteríolas aferentes, o que é confirmado pelo exame
da imuno-histoquímica (Fig. 7.3). A mácula densa é a parteda alça ascendente espessa de Henle que se encontra próxi-ma à região da arteríola aferente.
Pontos-chave:
Principais componentes do sistema renina-
angiotensina
• Renina• Angiotensinogênio• Enzima conversora da angiotensina II• Angiotensina II• Aldosterona• Receptores AT1 e AT2 da angiotensina II
CONTROLE DA SECREÇÃO DERENINA
Inúmeros fatores estão envolvidos no controle da secre-ção da renina (Quadro 7.1). Os estímulos primários para aliberação de renina renal dependem, basicamente, da re-dução da pressão de perfusão renal e da restrição da in-gesta de sódio ou perda de sódio. O grau de estimulaçãodo eixo renina-angiotensina depende do grau de depleçãode volume.1,3
A diminuição da pressão de perfusão renal pode serdecorrente de hemorragia aguda, estenose crônica de ar-téria renal ou depleção do fluido de volume extracelular(resultado da restrição de sódio ou da administração de
Fig. 7.2 Expressão de mRNA para renina utilizando-se hibridi-zação in situ em rim de rato submetido a restrição de sódio nadieta associado ao uso de diurético de alça (furosemida). Note agrande concentração de grânulos negros, que correspondem aomRNA para renina, localizados no hilo renal.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 104/121
72 Peptídeos Vasoativos e o Rim
diuréticos). Além disso, situações clínicas que cursam com baixa perfusão renal, como é o caso da insuficiência cardía-ca congestiva descompensada e cirrose hepática, estão fre-qüentemente associadas ao aumento da liberação renalde renina. Ao contrário, a expansão de volume e a dieta ricaem sódio levam à supressão da liberação de renina.6,10
Assim, a liberação de renina responde inversamente aalterações da perfusão renal. Esta resposta parece ser me-diada por mecanismo barorreceptor renal localizado nascélulas justaglomerulares da arteríola aferente, sensíveis apequenas alterações de pressão transmural e de estiramen-to da parede da arteríola. O aumento da pressão de perfu-são estira a parede da arteríola aferente induzindo dimi-nuição da secreção de renina, enquanto a redução da pres-
são de perfusão renal aumenta a secreção de renina.Existe também uma relação inversa entre a ingestão desódio e a atividade da renina. Devido à disposição anatô-mica especial da alça de Henle (no local da mácula densa)com as células justaglomerulares produtoras de renina (naarteríola aferente), a concentração de cloreto de sódio dofluido tubular é detectada pela mácula densa, regulando asecreção de renina. Dieta rica em sódio e expansão do vo-lume estão associadas com baixos níveis plasmáticos derenina, enquanto dieta pobre em sal e depleção de volumesão acompanhadas por baixos níveis de sódio e cloro nofluido tubular distal, que estimula renina.6,10
Foi objeto de grande discussão se era o conteúdo desódio ou de cloro do fluido tubular o responsável pela sen-sibilização da mácula densa. A favor da possibilidade do
cloro como principal modulador da secreção de renina, osestudos de Hackenthall e cols demonstraram que a infu-são de sais de sódio (outros que não o cloreto de sódio) nãoconsegue diminuir a liberação de renina, enquanto a infu-são de variadas concentrações de cloro produz alteraçõesda secreção de renina.10 No entanto, é importante observarque este padrão de resposta é idêntico ao que se observana alça ascendente espessa de Henle quando se estuda o
comportamento do transportador sensível à furosemida(Na-K-2 Cl). Na verdade as células da mácula densa sãocélulas da alça de Henle e o fato de ser o canal Na-K-2 Cl omediador do sinal para a secreção de renina explica por queo uso de furosemida aumenta em grande monta a secre-ção deste hormônio. Isto ocorre porque quanto menor otransporte de NaCl na mácula densa, maior é a liberaçãode renina pelas células justaglomerulares.11
O estímulo adrenérgico desempenha um papel relevan-te na regulação da secreção de renina. As células justaglo-merulares são inervadas por fibras simpáticas e apresentamreceptores -adrenérgicos. Diversos estudos demonstraramque a estimulação elétrica dos nervos renais12 ou infusões
de concentrações farmacológicas de epinefrina e norepine-frina13 estimulam diretamente a secreção de renina, sem al-terar o fluxo sanguíneo renal nem a excreção de sódio. Adopamina também influencia a secreção de renina, uma vezque existe inervação dopaminérgica específica no aparelho justaglomerular. Infusão intra-renal de dopamina produzum aumento da secreção de renina que é dose-dependentee bloqueada por drogas bloqueadoras dopaminérgicas.
Fig. 7.3 Expressão de renina em arteríola aferente. Imuno-histo-química. Note a arteríola marcada em vermelho. (Ver o Cd-ROM.)
Quadro 7.1 Condições que alteram níveis de renina
Estimula Secreção Inibe Secreção
de RENINA de RENINA
pressão de perfusão renal pressão deperfusão renal
restrição na ingesta de Na dieta rica em Naperda de Na (uso de diuréticos) expansão de volumedepleção de volumehipovolemia funcional(ICC, cirrose hepática comascite, síndrome nefrótica)
estímulo adrenérgico (epinefrina, deficiência denorepinefrina, dopamina) catecolamina
inibição de AII AII
Íons Íons Ca intracelular Ca intracelular
(quelação de Ca comEDTA, bloqueador decanais de Ca)
Mg K
K
prostaglandinas inibição de(PGE2 e PGI2) prostaglandinas
adenosina ADH
Fatores de crescimento Fatores deTNF, IL-1 crescimentoIGF EGFTGF-
tumor secretor de renina

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 105/121
capítulo 7 73
A secreção de renina pode ser influenciada de maneirasignificativa por substâncias circulantes e por substânciasproduzidas localmente. Neste contexto, destaca-se a parti-
cipação da Ang II, que inibe diretamente a liberação de re-nina.10 Assim, a inibição de Ang II por qualquer modalida-de (por exemplo, inibidores da enzima de conversão) é umpotente estímulo para a liberação de renina. Prostaglandi-nas, tais como PGE2 ou PGI2, estimulam a secreção de reni-na,14 e a inibição da síntese de prostaglandinas bloqueia aliberação de renina. As cininas são vasodilatadores que tam- bém estimulam a liberação de renina. O hormônio antidiuré-tico (ADH) inibe a liberação de renina estimulada, porémnão está claro se esta inibição é devida a uma ação direta nascélulas justaglomerulares ou à expansão do volume plasmá-tico. A ação do peptídeo atrial natriurético (ANP) na secre-ção de renina é controversa. Na maioria dos estudos o ANPdiminui a atividade da renina plasmática.
O óxido nítrico pode tanto inibir quanto estimular a
secreção de renina.15 Como a mácula densa tem grandequantidade de sintetase do óxido nítrico do tipo b (bNOS),que é uma das enzimas que sintetizam óxido nítrico, é bastante provável que o óxido nítrico participe do sinalpara produção de renina gerado na mácula densa.
Alterações nas concentrações extracelulares de diversosíons podem também alterar a liberação de renina. O cálciotem um papel central no controle da secreção de renina: di-minuição do cálcio citosólico estimula a secreção de renina,enquanto aumento do cálcio intracelular está associado comdiminuição da liberação de renina.10 Tanto a quelação docálcio com EDTA quanto o uso de bloqueadores de canaisde cálcio estimulam a secreção de renina. O aumento da con-centração de magnésio estimula a secreção de renina prova-
velmente por hiperpolarização da membrana celular, queinibe o influxo de cálcio. Existe uma correlação entre potás-sio e liberação de renina. Aumento de potássio despolarizaa membrana celular, aumenta a permeabilidade da célula aocálcio e assim permite um aumento do influxo de cálcio.
A adenosina parece ser um sinal adicional inibindo aliberação de renina. A adenosina exógena, in vivo, leva auma vasoconstrição renal passageira, com redução da taxade filtração glomerular e inibição da secreção de renina.Estudos com bloqueadores do receptor da adenosina mos-traram que a adenosina é um mediador parcial da libera-ção de renina dependente da mácula densa.16 Assim, seusefeitos na secreção de renina podem também ser secundá-rios às alterações na hemodinâmica renal.
Mais recentemente, tem sido analisado o papel dos fa-
tores de crescimento na hemodinâmica renal e sistêmica.O fator de necrose tumoral (TNF) e a interleucina-1 (IL-1)são potentes indutores da secreção de renina mas inibema secreção de aldosterona. Estas citocinas foram implica-das na síndrome do hipoaldosteronismo hiper-reninêmicoobservado em pacientes graves. Concentrações fisiológicasde insulina e fator de crescimento semelhante à insulina(IGF) também estimulam a renina. Os fatores transforma-
dores do crescimento 1 e 2 (TGF-1 e TGF-2) estimu-lam a renina e aparentemente seus efeitos são mediadosvia prostaglandinas. A privação de água aumenta a expres-
são de TGF- com aumento da atividade da renina plas-mática. Ao contrário, o fator de crescimento epidérmico(EGF) (que apresenta muitas propriedades em comum comAng II) é mais potente inibidor de renina que de Ang II.
Pontos-chave:
Principais estímulos para secreção de renina
• Diminuição da volemia• Hipoperfusão renal
Principais estímulos para diminuição da
secreção de renina
• Aumento da volemia•
Aumento da perfusão dos rins
QUANTIFICAÇÃO DAATIVIDADE DA RENINA
A renina plasmática circulante é constituída por suaforma precursora inativa (PRÓ-renina) e sua forma ativa(renina). A atividade da renina plasmática é determinadaatravés da medida da taxa de geração de Ang I a partir doangiotensinogênio plasmático endógeno. A quantidade deAng I produzida durante um determinado período é me-dida por radioimunoensaio e expressa em unidade denanogramas por ml/min.
A coleta de sangue de veia renal para dosagem de reni-na é realizada para auxiliar o diagnóstico de estenose deartéria renal unilateral (hipertensão arterial renovascular).Nos pacientes com suspeita de estenose de artéria renal aadministração de inibidores de enzima de conversão res-salta a secreção de renina no rim afetado.
A medida da atividade da renina periférica pode ser umimportante parâmetro para avaliação da participação doSRA em determinadas situações fisiopatológicas. Uma vezque a secreção de renina é altamente influenciada pela in-gesta de sódio na dieta e pelo estado do volume extracelu-lar, a determinação de renina plasmática deve estar corre-lacionada ao balanço de sódio.
ENZIMA CONVERSORA DEANGIOTENSINA (ECA)
Propriedades Bioquímicas
A enzima conversora de angiotensina (ECA) é uma car- boxipeptidase com peso molecular de 120-180 kDa que

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 106/121
74 Peptídeos Vasoativos e o Rim
converte Ang I para Ang II e, adicionalmente, inativa a bradicinina.
Distribuição TecidualA ECA encontra-se totalmente distribuída no organis-
mo e mais abundantemente no endotélio, mas também na borda em escova (p. ex.: rim, duodeno e íleo) e em órgãossólidos como útero e coração. Além disso a ECA tambémestá presente no sistema nervoso central e em células mo-nonucleares.17 Originalmente o endotélio pulmonar foiresponsabilizado como principal local da conversão deAng I para Ang II. No entanto, a formação de Ang II emtecidos periféricos pode ser igualmente importante. Norim, a ECA está localizada nas células endoteliais e na bor-da em escova do túbulo proximal.18 Uma vez que o túbuloproximal é capaz de produzir Ang II isoladamente,19 a ECA
produzida pelas células da borda em escova nestes túbu-los deve participar da ativação local do SRA, importantena regulação da reabsorção do fluido tubular proximal.
ANGIOTENSINA II
A Ang II é um peptídeo que tem um papel chave naregulação da pressão arterial e no balanço de sódio e águaem resposta a alterações do volume extracelular ou da pres-são sanguínea sistêmica. Estas ações são resultado de umaação direta no rim, na vasculatura extra-renal e nos túbu-los renais e indiretamente através dos efeitos na supra-re-nal e no sistema nervoso central.1,2,3
Ações na VasculaturaA Ang II é um potente vasoconstritor, fundamental para
manter a homeostase da pressão sanguínea. A infusão deAng II aumenta a resistência periférica total, principalmen-te na circulação renal, mesentérica e da pele, mas não emmúsculo esquelético.20 O sistema nervoso central respon-de à Ang II aumentando a descarga simpática e diminuin-do o tônus vagal.
Ações Renais
No rim existe um SRA completo que gera Ang II local-mente. Assim, as ações da Ang II no rim podem ser deri-
vadas de Ang II da circulação ou da produção local. A AngII tem ações importantes no rim que incluem modificaçõesda resistência vascular com conseqüente alteração da fun-ção glomerular, além de influir de maneira marcante nareabsorção de sódio.
A Ang II diminui o fluxo sanguíneo renal e a taxa defiltração glomerular devido ao aumento da resistência vas-cular.21 Tanto a arteríola aferente como a eferente contra-
em-se sob a ação da Ang II.22 Entretanto, existe uma maiorsensibilidade na arteríola eferente. O óxido nítrico podemodular a ação vasoconstritora da Ang II na arteríola afe-
rente mas não na eferente. Ang II induz ainda a contraçãode células mesangiais e com isso leva à redução da super-fície de filtração glomerular, reduzindo o coeficiente defiltração glomerular (Kf). A Ang II afeta ainda o tamanhodo poro da membrana basal glomerular e assim influen-cia a proteinúria: aumento do tamanho do poro induzidopor aumento dos níveis locais de Ang II resulta em protei-núria. Por outro lado, inibidores de Ang II diminuem a pro-teinúria na síndrome nefrótica. No túbulo proximal, a AngII estimula a reabsorção de sódio, água e bicarbonato.
Ações na Supra-renal
Ang II estimula a síntese de aldosterona na zona glome-
rulosa do córtex supra-renal. Desta maneira, o SRA man-tém a homeostase de sódio, água e potássio.
Ações no Sistema Nervoso Central
A Ang II age aumentando a sede e o apetite ao sal, con-tribuindo assim para o aumento do volume extracelular.
Ações da Ang II no Crescimento Celular
A Ang II pode promover crescimento e hipertrofia ce-lular. A Ang II induz uma resposta hipertrófica em célu-las mesangiais em cultura, além de induzir a produção defatores de crescimento, tais como fator de crescimento de-rivado de plaqueta (PDGF) e TGF-, levando ao aumentoda produção de matriz extracelular.23
A Ang II modula também o crescimento celular das cé-lulas da musculatura lisa dos vasos e dos miócitos cardía-cos e assim deve ter participação no desenvolvimento dahipertrofia cardíaca que acompanha algumas formas dehipertensão arterial.24
Pontos-chave:
• A angiotensina II mantém a volemiaconservando sal e água e promovendovasoconstrição periférica (ação sistêmica)
• A angiotensina II é um agente proliferativo
e fibrogênico (ação local)
Receptores para Ang II eMecanismo de Ação Celular
As células respondem à Ang II através de receptoresaltamente específicos presentes na membrana celular.25

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 107/121
capítulo 7 75
Duas classes principais de receptores para a Ang II foramidentificadas: AT1 e AT2. O receptor AT1 é o mediador dequase todas as funções fisiológicas conhecidas da Ang II
(vasoconstrição, secreção de aldosterona, sede, crescimentoe reabsorção tubular de sódio). O Losartan é um antago-nista do receptor AT1 e o PD-123177 é o antagonista do re-ceptor AT2.
Os receptores para Ang II, particularmente AT1, foramdemonstrados no sistema nervoso central, nos vasos, fíga-do, supra-renal, rim, ovário, baço, pulmão e coração. Osreceptores vasculares para Ang II se concentram nas célu-las da musculatura lisa dos vasos. No rim, receptores paraAng II estão localizados nos vasos, glomérulos, túbulosproximais e distais, mácula densa e na medula renal.26,27
Após a interação da Ang II com seus receptores ocorreativação de uma cascata de eventos regulada principalmen-te pelas proteínas G associadas ao receptor. Estas proteí-nas reguladoras ativam (fosfolipase C) ou inibem (adenil-
ciclase) enzimas presentes na membrana celular, levandoà alteração da concentração de componentes intracelula-res (“segundos mensageiros”) como o aumento do inosi-tol trifosfato (IP3) e diacilglicerol (DAG) e a diminuição dosníveis de AMP cíclico (AMPc). O IP3 liga-se a receptoresno retículo endoplasmático, liberando cálcio ionizado deorganelas não-mitocondriais e assim aumenta o cálciointracitoplasmático. O DAG, na presença do aumento docálcio livre citosólico, une-se e ativa a proteína cinase C.Este mecanismo de ativação promove a contração das cé-lulas da musculatura lisa dos vasos, como também a con-tração de células mesangiais, além de agir como estímulomitogênico.28
ANGIOTENSINASES
A inativação da Ang II e da Ang III ocorre por hidrólisecausada por angiotensinases não-específicas que estão pre-sentes no sangue e tecidos. A degradação da angiotensinaocorre nos diferentes órgãos, incluindo o rim.
SISTEMA RENINA-ANGIOTENSINA EM SITUAÇÕES
FISIOPATOLÓGICAS
Hipertensão Arterial
Não há dúvida de que a infusão de Ang II leva à hiper-tensão arterial por induzir vasoconstrição, além de aumen-tar o volume intravascular mediado pela aldosterona.
Existem formas de hipertensão arterial que são classi-camente renina-dependentes, como é o caso da estenose deartéria renal e tumor secretor de renina. No entanto, a hi-pertensão maligna também está associada com hiperati-
vidade do SRA. Já nas formas de hipertensão arterial es-sencial, os níveis de renina se encontram dentro da faixade normalidade em 60% dos casos, enquanto que em 15%
dos casos os níveis estão elevados.As principais formas de inibir o SRA são através da uti-lização de inibidores da ECA, que bloqueiam a formaçãode Ang II, e dos antagonistas dos receptores AT1, que im-pedem a ação da Ang II. São drogas usadas na terapêuticada hipertensão arterial e da insuficiência cardíaca conges-tiva, situações nas quais há excessiva retenção de sal.
A inibição da ECA está associada ao aumento das cini-nas, que também contribuem para o efeito terapêutico(como será discutido ainda neste capítulo). Por outro lado,são responsáveis pelo aparecimento da tosse seca comoefeito colateral. Os antagonistas dos receptores AT1 dimi-nuem a pressão arterial e inibem os efeitos mitogênicosmediados por Ang II.
Nefropatia Diabética
No diabetes, associado ou não com hipertensão arteri-al, os níveis de renina encontram-se diminuídos, provavel-mente como resultado da expansão de volume, funçãoanormal do sistema nervoso autônomo e baixa produçãorenal de PGI2. Como conseqüência, há baixa produção dealdosterona, levando à hiperpotassemia.
Curiosamente, apesar de a atividade do SRA ser baixano diabetes, a Ang II é um importante mediador das alte-rações fisiopatológicas da nefropatia diabética. A Ang IIleva ao aumento da pressão intraglomerular (por vasocons-trição da arteríola eferente) e induz hipertrofia da célulamesangial com aumento da produção de matriz. O uso de
inibidores da ECA (e mais recentemente antagonista doreceptor AT1) diminui a proteinúria assim como retarda aprogressão da nefropatia diabética, tanto em modelos ex-perimentais como em humanos.
Nefropatias Crônicas
Em diversas nefropatias foi demonstrada a atividade doSRA, que por mecanismos mediados via Ang II pode le-var ao aumento da pressão capilar intraglomerular e pro-teinúria. Assim, inibidores da ECA têm sido utilizados
Pontos-chave:
Patologias tratáveis pela inibição do sistemarenina-angiotensina
• Hipertensão arterial• Hipertrofia do ventrículo esquerdo• Insuficiência cardíaca congestiva• Nefropatia diabética• Insuficiência renal crônica (inicial)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 108/121
76 Peptídeos Vasoativos e o Rim
como agentes antiproteinúricos e nefroprotetores. No en-tanto, o efeito antiproteinúrico é variável: a resposta pare-ce melhor em pacientes normotensos e quando se associa
a restrição de sal na dieta. Além desses efeitos o bloqueiodo SRA pode ser nefroprotetor por inibir a formação de fi- brose intersticial.
PEPTÍDEOS DERIVADOS DASANGIOTENSINAS
Alguns peptídeos derivados da degradação das angio-tensinas não são biologicamente inertes. Os mais conheci-dos são Ang-(1-7), Ang-(2-8) ou Ang III e Ang-(3-8) ou AngIV. Estes peptídeos são gerados através da ação deangiotensinases, conforme mostrado na Fig. 7.4.
Ang-(1-7)A Ang-(1-7) é um derivado da angiotensina que tem
ações fisiológicas, na maior parte das vezes, antagônicasàs da Ang II. O heptapeptídeo é produzido pela ação depeptidases teciduais sobre a angiotensina e é rapidamentehidrolisado depois de formado, principalmente através daação da enzima conversora da angiotensina. Desta forma,com o uso dos inibidores da ECA ocorre acúmulo de Ang-(1-7). Neste caso, o aumento da concentração de Ang-(1-7)
deriva tanto do bloqueio da degradação do heptapeptídeoquanto do acúmulo de substrato (Ang I). Além disso, o usode bloqueador de receptor AT1 também pode levar ao acú-
mulo de Ang-(1-7).4A Ang-(1-7) potencializa os efeitos hipotensores da bra-dicinina. O mecanismo envolvido é complexo e inclui fa-cilitação da liberação de óxido nítrico, prostaglandinas,fator hiperpolarizante derivado do endotélio (EDHF) einibição da quebra de bradicinina (BK) via ECA. No entan-to, a Ang-(1-7) também pode agir tendo a BK como medi-ador, pois a Ang-(1-7) é capaz de se ligar ao receptor AT2e a ativação deste estimula a síntese de bradicinina.29
No rim a Ang-(1-7) produz diurese e natriurese. Estesefeitos são bloqueáveis por losartan, mas não são devidosà ação do receptor AT1 e sim a um outro receptor sensívela losartan, denominado receptor AT1-símile. A Ang-(1-7)também afeta o transporte de água no túbulo renal, e comoeste efeito é bloqueável por PD 123319, deve ser mediadopor AT2. Parte dos efeitos da Ang-(1-7) no rim tambémpode ser mediada pelo receptor da Ang IV, uma vez que aAng-(1-7) pode ser convertida em Ang-(3-7) que, por suavez, é capaz de ativar o receptor da Ang IV.
Ang III
A Ang III determina os mesmos efeitos da ativação doreceptor AT1 da Ang II, tendo já sido inclusive questiona-do se os efeitos fisiológicos da Ang II não seriam media-dos por Ang III. Destes efeitos, parece que a Ang III real-mente é a responsável pela liberação de vasopressina, umavez que quando se bloqueia a conversão Ang II–Ang III oefeito não é mais observável. Além disso, a Ang III pode
ter um papel importante na inflamação e fibrose glomeru-lares.30
Ang IV
A Ang IV está envolvida nos mecanismos de recorda-ção de memória, vasodilatação encefálica e crescimentocelular da adeno-hipófise. O receptor da Ang IV está dis-tribuído em vários sítios anatômicos do sistema nervosocentral, mas também está presente em outros órgãos e te-cidos como coração, supra-renais e músculo liso vascular.Sua ação nestes alvos ainda está sob investigação.31
Pontos-chave:
Efeitos dos metabólitos da angiotensina II
• Ang-(1-7): vasodilatação e antiproliferação• Ang III: vasoconstrição e proliferação
celular• Ang IV: efeitos no sistema nervoso centralFig. 7.4 Catabolismo das angiotensinas. § representa ACE, * en-
dopeptidases neutras, # aminopeptidases e † carboxipeptidases.
Angiotensinogênio
Ang I (1-10)
Ang II (1-8) Ang-(1-7)
Ang III (2-8) Ang IV (3-8)
Ang-(3-7)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 109/121
capítulo 7 77
SISTEMA RENALCALICREÍNA-CININA
As cininas são peptídeos vasodilatadores, sendo que omais conhecido, a bradicinina, foi descoberta por um cien-tista brasileiro, o Professor Rocha e Silva.32 A participaçãodo sistema calicreína-cinina na função renal, com açõesprimordialmente vasodilatadoras, continua pouco defini-da. No entanto, existem fortes evidências de que estas subs-tâncias atuem na regulação do fluxo sanguíneo renal e nocontrole da excreção renal de sódio e água.2,3,33
Componentes do Sistema RenalCalicreína-cinina
Assim como existe um sistema vasoconstritor (sistema
renina-angiotensina) cujo elemento ativo é um peptídeo(Ang II, com oito aminoácidos), existe um sistema vasodi-latador cujo agonista ativo mais comum é outro peptídeo(bradicinina, com nove aminoácidos). Da mesma formaque o SRA, o sistema calicreína-cinina tem um zimogênioprecursor (cininogênio) que é quebrado para gerar os pep-tídeos ativos através da ação de uma enzima ativadora (ca-licreína). Além disso, os peptídeos são degradados porenzimas proteolíticas (cininases).34 A bradicinina é geradana circulação, mas nos tecidos é produzido um decapeptí-deo chamado calidina, que é uma molécula de bradicini-na acrescida de uma lisina em sua porção amino-terminal,portanto é uma lisil-bradicinina. Uma pequena quantida-de da calidina pode ser convertida a bradicinina por umaaminopeptidase (Fig. 7.5).
CININOGÊNIOSOs cininogênios são glicoproteínas de cadeia simples
sintetizados primariamente no fígado e depois secretadose transportados no plasma. O gene do cininogênio huma-no (localizado no cromossoma 3q26) codifica a produçãode dois cininogênios: um cininogênio de alto peso mole-cular — HMW (88-120 kDa) — e outro cininogênio de bai-
xo peso molecular — LMW (50-68 kDa). Na circulação sis-têmica a calicreína quebra o cininogênio de alto peso mo-lecular e nos tecidos a calicreína age sobre os dois tipos de
cininogênio. Os cininogênios estão também presentes namembrana das plaquetas, nos neutrófilos e no endotéliovascular. No rim, LMW-cininogênio (o substrato preferi-do para calicreína tissular renal) é detectado tanto no cór-tex como na medula renal.
CALICREÍNASAs calicreínas são proteases que existem em duas gran-
des formas, a plasmática e a tissular, e que diferem entresi estrutural e funcionalmente. A calicreína plasmática (100kDa) participa da cascata de coagulação e libera cininas(principalmente bradicinina) do cininogênio de alto pesomolecular mas não do LMW-cininogênio. A calicreína plas-mática não é encontrada no rim e é pouco provável que
afete a função renal. No entanto, pela liberação de bradici-nina, um potente vasodilatador, podem ocorrer efeitosvasculares periféricos. A calicreína tissular (24-45 kDa),também chamada calicreína glandular, está presente emglândulas endócrinas e exócrinas e no rim. Diversasproteinases são capazes de ativar a PRÓ-calicreína. Uma vezativada, a calicreína renal quebra usualmente o LMW-ci-ninogênio liberando a lisil-bradicinina (calidina).
A atividade enzimática das calicreínas tissulares podeser inibida pela aprotinina (6,5 kDa). A aprotinina está dis-ponível comercialmente e é amplamente empregada comoinibidor tissular de calicreína, ainda que não seja específi-ca para este fim.
O gene humano da calicreína renal localiza-se no cro-mossoma 19 (q13.2-13.4) e é denominado hKLK1. Foi ob-
servado que existe homologia, tanto no nível genômicoDNA como no nível protéico, entre a calicreína tissular e ohKLK3, que codifica o antígeno prostático específico (PSA).O PSA está presente na próstata e é relevante na detecçãodo carcinoma de próstata. Na verdade, o PSA pertence, doponto de vista estrutural, à família das calicreínas, embo-ra não tenha função correlata às mesmas. Outras proteínastambém têm esta característica, como a tonina, que, emboraseja assemelhada às cininas, gera angiotensina II a partirde angiotensinogênio.
CININASAs cininas têm uma meia-vida extremamente curta de 10
a 30 segundos, o que dificulta e limita o estudo destas subs-tâncias. A concentração de cininas em fluidos biológicostambém é muito baixa, da ordem de pg/ml. A cinina for-mada no rim é detectada na urina, no fluido intersticial re-nal e, em algumas circunstâncias, no sangue venoso renal.
CININASESAs cininas são rapidamente inativadas por cininases
(cininases I e II) e pelas endopeptidases neutras (EPN),Fig. 7.5 Componentes do sistema calicreínas-cininas. * represen-ta calicreína tissular, # calicreína plasmática e § aminopeptidase.
Cininogênio de baixopeso molecular
Cininogênio de altopeso molecular
Calidina Bradicinina

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 110/121
78 Peptídeos Vasoativos e o Rim
todas presentes no sangue e nos tecidos.35 A cininase I éuma carboxipeptidase específica que remove o aminoáci-do carboxiterminal das cininas (arginina). A cininase II e a
endopeptidase neutra EPN 24-11 quebram a ligação Pro-Phe da bradicinina.36 A cininase II também quebra a liga-ção His-Leu da Ang I, levando à formação da Ang II, sen-do também conhecida como enzima conversora da angio-tensina I. Os rins são muito ativos em degradar cininas, pois90% do hormônio é inativado e 1% é excretado na urina.37
Receptores para Cininas
As cininas agem nas células-alvo através de receptores,denominados BK1 e BK2.38 Os receptores BK2 são os prin-cipais mediadores das cininas. Os receptores BK1 são me-nos proeminentes que os BK2 e exercem efeitos quandoinduzidos por inflamação, como no choque endotóxico
induzido por endotoxina de E. coli, situação na qual ocor-re uma marcante vasodilatação e hipotensão.
Pontos-chave:
Componentes do sistema calicreína-cininas
• Cininogênio (tissular ou plasmático)• Calicreína• Bradicinina ou calidina• Receptores BK1 e BK2• Cininase II (ECA)
Localização Renal dos Componentes doSistema Calicreína-cinina
Utilizando técnicas de imuno-histoquímica39 e de hibri-dização in situ,40 foi possível localizar os componentes dosistema calicreína-cinina ao longo do néfron. LMW-cinino-gênio foi identificado no néfron distal, particularmente nostúbulos distais medulares e corticais e nos ductos coleto-res.39
Imunorreatividade para calicreína e atividade enzimá-tica foram demonstradas predominantemente no túbulo deconexão no córtex renal.39,41 Técnicas de imunocitoquími-ca sugerem que a calicreína é encontrada predominante-mente na membrana plasmática e sua reatividade é maiordo lado luminal. A calicreína é secretada para a luz do tú-
bulo distal, e parte vai para o espaço peritubular, ondepode agir no cininogênio plasmático e formar bradicinina.A calicreína urinária consiste predominantemente em ca-licreína secretada pelo rim, embora uma pequena partepossa ser resultado de calicreína filtrada.
Tanto a calidina como a bradicinina são excretadas naurina. É provável que os túbulos coletores sejam o princi-pal sítio de produção de cinina e de sua ação. A bradicini-
na extra-renal filtrada não alcança este local porque ocor-re degradação no néfron proximal.
A cininase II está localizada nas células endoteliais, cé-
lulas epiteliais do túbulo proximal (borda em escova) e dotúbulo distal e no glomérulo. A endopeptidase neutra estálocalizada na borda em escova do túbulo proximal. Assim,os rins são muito ativos em degradar. As cininas circulan-tes filtradas pelo rim são degradadas rapidamente pelacininase II na borda em escova do túbulo proximal, 33 e seelas entram na circulação pós-glomerular são inativadaspela cininase II das células endoteliais ou então degrada-das no pulmão. Uma vez que a atividade das cininases étão abundante no túbulo proximal e na vasculatura, é pou-co provável que cininas circulantes sejam capazes de mo-dular a função renal. Por isso, a geração endógena pareceser necessária para a ativação da cinina in vivo.33
Receptores para bradicinina estão presentes em altadensidade nos túbulos coletores corticais e medulares e em
células intersticiais, e mais recentemente foram detectadostambém em células mesangiais em cultura.42
Em resumo, a localização de cininogênio, calicreína elocais específicos de ligação para cininas no néfron distalsugere que esta região seja o principal local de produção ede ação das cininas renais.
Ações do SistemaCalicreína-cinina no Rim
Desde 1909, quando foram constatadas as propriedadeshipotensoras da urina (agora reconhecidas como sendo de-vido à excreção renal de calicreína), discute-se o papel dosistema calicreína-cinina na função renal, no controle da
pressão arterial e na regulação da excreção de sódio e água.No entanto, até hoje a contribuição isolada do sistema cali-creína-cinina isoladamente ainda não foi estabelecida. Osdados demonstrando um alto turnover diário da taxa decalicreína renal em humanos, além da presença de altas con-centrações de cininases renais e da identificação de recep-tores para cininas em túbulos, células intersticiais e célulasmesangiais, falam a favor da ação local deste sistema. Poroutro lado, estudos mais recentes indicam que o sistemacalicreína-cinina parece agir como parte de um complexosistema de regulação que envolve também as prostaglandi-nas, a renina, a Ang II e outros peptídeos vasoativos.
EFEITOS NA HEMODINÂMICA RENAL
Os primeiros estudos sobre a ação das cininas na fun-ção renal foram realizados em 1964 por Webster43 e cols eem 1965 por Gill e cols.44 Foi demonstrado que a infusãoaguda de doses farmacologicamente ativas de cininas in-duzia um importante efeito de vasodilatação renal, comaumento do fluxo sanguíneo renal e aumento da excreçãode sódio e água. Estes resultados sugerem que as cininassejam fatores capazes de regular o fluxo sanguíneo renal.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 111/121
capítulo 7 79
Já a administração crônica de bradicinina na artéria re-nal mostrou que, embora a vasodilatação renal permane-ça, o aumento na excreção de sódio e água não se mantém
ao longo do tempo.45 Estes achados falam contra a possi- bilidade de as cininas terem um papel natriurético croni-camente. Todos estes resultados devem ser analisados cri-teriosamente, pois infusões destes peptídeos podem nãorefletir verdadeiramente os efeitos da bradicinina geradaendogenamente. É pouco provável que in vivo as cininascirculantes modulem a função renal, pois, como já discuti-do acima, devido à abundante atividade das cininases nosvasos e no túbulo proximal, estas substâncias são rapida-mente degradadas. Assim, a geração endógena parece sernecessária para a ação de cinina in vivo.
A bradicinina tem um potente efeito relaxante na arte-ríola glomerular, promovendo vasodilatação tanto da ar-teríola aferente como da eferente.46 Entretanto, a arteríolaeferente dilata em resposta à bradicinina de uma maneiradose-dependente.46
EFEITOS NA EXCREÇÃO DE SÓDIO E ÁGUAAs cininas induzem o aumento da excreção de sódio e
água, porém o exato mecanismo deste efeito continua con-troverso. Muitos autores correlacionam a natriurese aosefeitos vasodilatadores das cininas. Desde os experimen-tos de Webster e Granger tem sido documentado que in-fusão de bradicinina na artéria renal induz natriurese semalterar a taxa de filtração glomerular,43,45 sugerindo efeitodireto no transporte tubular de sódio.
A favor desta possibilidade estão os estudos de micro-perfusão de porções distais do túbulo proximal com solu-ções suprafisiológicas de bradicinina, nos quais há aumen-
to da excreção de sódio, provavelmente por ação direta noepitélio urinário.35,47
A inibição do eixo calicreína-bradicinina endógenousando anticorpos específicos antibradicinina ou utilizan-do-se aprotinina acaba com o efeito natriurético e diuréti-co da infusão salina.35 Estes resultados indicam que as ci-ninas endógenas podem agir como substâncias natriuréti-cas.
A cinina produzida endogenamente, quer seja pelo usode inibidores de cininases renais (captopril) ou pelo trata-mento crônico com desoxicorticosterona (que aumenta osníveis de cininas endógenas), aumenta o fluxo sanguíneo
papilar, a diurese e a excreção de sódio. Neste modelo, ouso de bloqueadores específicos de receptores de cininasBK2 consegue atenuar ou mesmo abolir estes efeitos. As-
sim, fica claro que cinina produzida endogenamente afetade forma significativa a hemodinâmica renal e a funçãoexcretora.33
Inter-relações entre SistemaCalicreína-cinina e outros Sistemas
É provável que as cininas exerçam seus efeitos modula-dores na função renal por interação com outros hormôni-os vasoativos, incluindo o sistema renina-angiotensina,prostaglandinas e vasopressina.33
SISTEMA RENINA-ANGIOTENSINAA inter-relação entre o sistema calicreína-cinina e o sis-
tema renina-angiotensina é complexa e não compreendi-da completamente. No entanto, alguns aspectos têm sido bem reconhecidos. Há evidências de que a bradicininapode estimular diretamente a liberação de renina no glo-mérulo.48 Por outro lado, a enzima conversora de angioten-sina é eficiente em inativar cininas. Por este motivo, partedo efeito anti-hipertensivo dos inibidores da ECA pode seruma conseqüência da diminuição da destruição de cinina,mantendo os seus efeitos vasodilatadores. É possível queo uso de bloqueadores do receptor AT1 da Ang II condu-za a efeitos semelhantes, pois com o bloqueio destes recep-tores há aumento da concentração de Ang II e conseqüen-temente maior ativação dos receptores AT2. Existe a sus-peita de que os receptores AT2 possam ativar o sistemacalicreína-cininas.38
É interessante que em algumas situações os dois siste-mas parecem ter comportamentos opostos e paradoxais.Por exemplo, dietas pobres em sódio e outras alteraçõessistêmicas, como depleção de volume, estimulam a sínte-se de renina e aumentam os níveis de Ang II e aldostero-na, com objetivo de reter sódio e água e aumentar a pres-são arterial em defesa da homeostase circulatória. Por ou-tro lado, estes mesmos estímulos aumentam a calicreínarenal e a produção de cinina, que têm ações vasodilatado-ras e objetivam uma defesa local do fluxo sanguíneo renale da taxa de filtração glomerular.
EICOSANÓIDESO sistema calicreína-cinina ativa a síntese de eicosanói-
des no rim. Diversos estudos demonstram que a cininaestimula a liberação do ácido araquidônico e subseqüentesíntese de eicosanóides na vasculatura renal, células inters-ticiais e células epiteliais. Isto tudo parece ocorrer via ati-vação de receptor BK2 e liberação de ácido araquidônicomediado por fosfolipase A2.
A estimulação da produção de PGE2 ocorre em respos-ta à cinina em células dos ductos coletores assim como em
Pontos-chave:
Principais efeitos das cininas
• Diminuição da pressão arterial• Vasodilatação sistêmica• Vasodilatação renal• Natriurese• Diurese

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 112/121
80 Peptídeos Vasoativos e o Rim
arteríolas glomerulares e células mesangiais. A síntese dePGI2 vascular é poderosamente estimulada pela cinina,assim como é a síntese de tromboxane A2.
A vasopressina estimula a liberação de calicreína renale a produção de cininas, mas as cininas inibem a reabsor-ção de sódio e água induzida por vasopressina nos ductoscoletores, provavelmente via produção de PGE2 neste lo-cal. Assim, é possível que existam alças de feedback negati-vo local entre cininas, eicosanóides e vasopressina no né-fron distal.
PEPTÍDEO NATRIURÉTICO ATRIAL (ANP)A calicreína tissular é capaz de formar ANP a partir de
seu precursor e catabolizar a atividade dos eupeptídeos in
vitro. A administração de ANP afeta a excreção urinária decalicreína. A endopeptidase neutra NEP 24.11 degrada tan-to as cininas quanto os peptídeos natriuréticos, assim os
efeitos farmacológicos de sua ação são inespecíficos. Fatosemelhante ocorre com o bloqueio da ECA, que tanto im-pede a formação de Ang II quanto impede a degradaçãodas cininas.
ÓXIDO NÍTRICOO óxido nítrico é um mediador implicado na vasodila-
tação induzida por cinina. A vasodilatação produzida pelaadministração de bradicinina é significativamente, mas nãototalmente, dependente da síntese de óxido nítrico e podeser marcadamente atenuada por inibidores da óxido nítri-co sintetase.
SISTEMA CALICREÍNA-CININAEM SITUAÇÕES
FISIOPATOLÓGICAS
Cirrose Hepática
Pacientes com cirrose hepática apresentam aumento daexcreção urinária de calicreína. Quando estes pacientesevoluem para a síndrome hepatorrenal ocorre uma dimi-nuição drástica desta excreção, sugerindo que a incapaci-dade de produzir cininas possa contribuir para a gêneseda insuficiência renal aguda na síndrome hepatorrenal.
Hipertensão Arterial
Em hipertensão arterial clínica e experimental foi de-monstrada uma diminuição da excreção urinária de cali-creína e que esta redução correlaciona-se com a gravidadeda hipertensão. A diminuição da atividade do sistema ca-licreína-cinina renal pode ser responsável, em parte, pelaretenção de sódio e assim participar da fisiopatologia dahipertensão arterial. Além disso, a terapêutica com inibi-
dores da enzima de conversão diminui a destruição dascininas, o que provavelmente desempenha papel signifi-cante na sua eficiência anti-hipertensiva.
Fibrose Miointimal
Os inibidores da ECA são capazes de prevenir a fibrosemiointimal que ocorre em artérias lesadas. Estudos maisaprofundados mostraram que tanto a Ang II quanto as ci-ninas desempenham um papel na formação/prevenção dafibrose miointimal. O efeito das cininas é, provavelmente,mediado pela geração de óxido nítrico.41
Infarto do Miocárdio
Os inibidores da ECA, quando usados no infarto agu-do do miocárdio, são capazes de melhorar a função cardí-
aca, diminuir a mortalidade e a taxa de reinfarto. A maiorquantidade de cininas presentes na circulação coronáriapoderia, neste caso, gerar maior quantidade de PGI2 e NO,que são importantes inibidores da agregação plaquetária.Além disso, as cininas são estimuladores potentes da libe-ração de tPA (ativador tissular de plasminogênio), quepode, por sua vez, ativar a plasmina e induzir fibrinólise.41
Pontos-chave:
Patologias associadas à depleção de cininas
(ou que poderiam se beneficiar do aumento
de cininas)
• Síndrome hepatorrenal
• Fibrose miointimal• Infarto do miocárdio• Hipertensão arterial
PEPTÍDEOS NATRIURÉTICOS
Existe uma família de peptídeos natriuréticos conheci-dos, todos com fórmula estrutural semelhante, consistin-do em um anel de 17 aminoácidos ligados por uma pontede dissulfeto. No momento, quatro destes peptídeos foramrazoavelmente estudados, ANP, BNP, CNP e urodilatina.O ANP e a urodilatina são codificados pelo mesmo gene eo BNP e o CNP são codificados cada um por seu próprio
gene.
Peptídeo Natriurético Atrial (ANP)
O peptídeo natriurético atrial (ANP), que foi o primei-ro peptídeo natriurético descrito, é um potente hormônionatriurético produzido principalmente pelos miócitos do

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 113/121
capítulo 7 81
átrio cardíaco em resposta à distensão local da parede atri-al. Desta maneira, é uma substância que tem uma partici-pação importante na regulação da homeostase de volume
do organismo. DeBold e cols49 foram os primeiros a de-monstrar que a injeção intravenosa de extratos de átrio emratos produzia um efeito potente e imediato de aumentara excreção renal de sódio e de água. Sua infusão leva con-comitantemente a uma diminuição da pressão arterial. Sub-seqüentemente, foi demonstrado que os grânulos localiza-dos no átrio armazenavam uma substância natriurética.Este peptídeo foi inicialmente chamado de fator natriuré-tico atrial.
O ANP age principalmente nos rins, na vasculatura e nasglândulas supra-renais através de receptores específicos.Os principais efeitos do ANP incluem inibição do transpor-te de sódio e supressão da liberação de renina e aldostero-na, além do relaxamento da musculatura lisa dos vasos. Porestes mecanismos leva à natriurese, diurese e diminuiçãoda pressão arterial, com conseqüente diminuição do volu-me extravascular.50,51
SÍNTESE E ESTRUTURAO gene humano que codifica o PRÉ-PRÓ-ANP está loca-
lizado no braço curto do cromossoma 1 e contém 3 exons(Fig. 7.4). O produto do gene é oPRÉ-PRÓ-ANP formado por151 aminoácidos. A quebra da molécula resulta em PRÓ-ANP, que é a principal forma de armazenamento comogrânulos nos miócitos cardíacos. No momento da libera-ção do átrio cardíaco, o PRÓ-ANP é convertido no peptí-deo ativo ANP (composto por 28 aminoácidos), que é aforma do hormônio que se encontra na circulação comimportantes funções fisiológicas.50,51
Utilizando-se anticorpos específicos anti-PRÉ-PRÓ-ANPfoi possível localizar imunorreatividade nos grânulos se-cretórios do átrio cardíaco.52 Uma pequena quantidade dopró-hormônio também é produzida no ventrículo esquer-do e esta produção está muito aumentada na hipertrofiado ventrículo esquerdo. A expressão de mRNA para ANPtambém foi encontrada em cérebro, pituitária, hipotálamo,arco aórtico, pulmão, medula adrenal e rim, confirmandoa produção deste peptídeo em outros tecidos. No entan-to, a quantidade de ANP sintetizada nestes locais é bemmenor que a encontrada em átrio e aparentemente nãocontribui de maneira significativa para o hormônio circu-lante.
REGULAÇÃO DA SECREÇÃO DO ANPO estímulo mais importante para liberação de ANP é oestiramento cardíaco,53 que pode ser resultado de váriosfatores, dentre eles: sobrecarga de sal, sobrecarga aguda ecrônica de volume, estados clínicos associados com o au-mento da pressão intra-atrial (insuficiência cardíaca con-gestiva, insuficiência renal aguda e insuficiência renal crô-nica), aldosteronismo primário e síndrome da produção
inapropriada do hormônio antidiurético. Finalmente, en-dotelina, acetilcolina, epinefrina, vasopressina e glicocor-ticóides aumentam a secreção de ANP50,51,53 (Quadro 7.2).
A secreção do ANP pode ser controlada basicamentepelos seguintes mecanismos: primeiro, a conversão de PRÓ-ANP em ANP (e liberação de ANP armazenado) e aumentoda síntese de mRNA (levando ao aumento de PRÓ-ANP eANP). O primeiro mecanismo está principalmente envol-vido na liberação de ANP quando ocorrem estímulos agu-dos de aumento da pressão intra-atrial. Já estímulos crô-nicos promovem a secreção de ANP, via aumento da sín-tese.2,54
RECEPTORES PARA ANPPara exercer seus efeitos fisiológicos, o ANP tem que se
ligar a receptores específicos presentes na membrana dascélulas alvo.51 Os receptores estão presentes no rim, supra-renal, cérebro e vasculatura. No rim, localizam-se princi-palmente nos vasos renais, no glomérulo e na medula epapila renal. Há três tipos de receptores para ANP: GC-A(guanilato-ciclase A), GC-B (guanilato-ciclase B) e CR (cle-arance receptor, ou receptor catabólico).
Os receptores GC-A e GC-B são biologicamente ativos,são os mediadores das ações do ANP e dos outros peptí-deos natriuréticos e apresentam um domínio citosólicocom atividade enzimática associada à proteína G. O ANPage via receptor GC-A. Quando o hormônio se liga ao re-ceptor na superfície da membrana celular, a atividade da
guanilato ciclase é estimulada e desta maneira é sinteti-zado cGMP (que é o mediador dos efeitos do ANP). Osreceptores CR não apresentam atividade enzimática e sãoreceptores de clearance que servem para depurar o ANPda circulação e regular desta maneira o nível circulante deANP.2 O ANP tem uma outra importante via de catabo-lismo que ocorre pela ação da endopeptidase neutra NEP24.11.
Quadro 7.2 Situações que estimulam a secreçãode ANP
• estiramento cardíaco e aumento da pressão intra-atrial— sobrecarga de sal— sobrecarga aguda e crônica de volume— insuficiência cardíaca congestiva— insuficiência renal aguda— insuficiência renal crônica— aldosteronismo primário— síndrome da produção inapropriada do hormônioantidiurético
• endotelina• acetilcolina• epinefrina• vasopressina• glicocorticóides

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 114/121
82 Peptídeos Vasoativos e o Rim
AÇÕES DO ANPA ação do ANP é imediata e de curta duração. A meia-
vida do ANP exógeno injetado endovenoso é de 2 a 4 mi-
nutos.55,56 A administração endovenosa de ANP ou a inje-ção em artéria renal produz imediata diurese e natriurese,porém de duração muito curta.49 O aumento da excreçãode sódio e água é acompanhado por aumento importanteda excreção de cálcio, magnésio e cloro.2
EFEITOS NA HEMODINÂMICA RENAL EEFEITOS NA INDUÇÃO DE NATRIURESE
O exato mecanismo pelo qual o ANP induz natriureseé multifatorial. No rim, o ANP aumenta a taxa de filtraçãoglomerular.57 Estudos de micropunção demonstraram queo ANP dilata a arteríola aferente e leva à vasoconstriçãoda arteríola eferente, resultando em aumento da pressãohidráulica do capilar glomerular. Outros estudos mostra-
ram que o Kf aumenta significantemente após infusão deANP.58 Efetivamente, o aumento da filtração glomerularpode levar ao aumento da natriurese.50,51 A diminuição dahipertonicidade medular também contribui para o efeitonatriurético.50,51
Apesar de estas ações hemodinâmicas serem importan-tes, aparentemente a principal ação natriurética do ANP éo efeito direto nos túbulos, inibindo o transporte de sódionos ductos coletores.50,51,59 O principal local de ação do ANPsão as células do ducto coletor da medula interna. O ANPestimula a produção de cGMP nestas células e estudos demicroperfusão in vitro demonstraram que a reabsorção desódio é inibida pelo ANP neste segmento.
EFEITOS NO SISTEMA RENINA-
ANGIOTENSINA-ALDOSTERONA,VASOPRESSINA E ENDOTELINA
A infusão de ANP diminui significativamente a secre-ção de renina e aldosterona.60,61 A inibição da secreção derenina pode ser devida ao aumento da carga filtrada decloreto de sódio que alcança a mácula densa, que respon-de diminuindo a secreção de renina.62 Existe também umefeito inibitório direto do ANP na secreção de renina.63 OANP inibe a secreção de aldosterona, indiretamente peladiminuição da secreção de renina e diretamente afetandoas células da camada glomerulosa do córtex adrenal, quesecretam a aldosterona.64 ANP inibe a liberação de vaso-pressina,50,51 levando ao aumento do clearance de água li-vre. O ANP também diminui a liberação de endotelina, um
peptídeo vasoconstritor.65
EFEITOS NA VASCULATURAANP causa relaxamento da musculatura lisa dos vasos
levando à vasodilatação. Infusão de doses farmacológicasde ANP em indivíduos normais e hipertensos induz umarápida e mantida diminuição da PA média. Os mecanismospelos quais o ANP diminui a pressão sanguínea incluem
diminuição do débito cardíaco, redução da resistência vas-cular periférica e diminuição do volume intravascular, o queocorre tanto devido à diurese/natriurese como por transfe-
rência de líquido do interior dos vasos para o interstício.
Pontos-chave:
Principal estímulo para secreção de peptídeo
natriurético atrial
• Aumento da volemia
Principais efeitos do peptídeo natriurético
atrial
• Diurese• Natriurese• Vasodilatação sistêmica• Diminuição da produção de
vasoconstritores (Ang II, endotelina evasopressina)
ANP em Situações Fisiopatológicas
INSUFICIÊNCIA CARDÍACA CONGESTIVAUma vez que ANP é secretado em resposta ao estira-
mento atrial, não causam surpresa os achados de níveisplasmáticos elevados de ANP em pacientes com insufici-ência cardíaca congestiva. Na insuficiência cardíaca con-gestiva com hipertrofia ventricular foi detectada síntese au-mentada de ANP também pelos ventrículos.50
Há correlação entre os níveis plasmáticos de ANP com
a gravidade da insuficiência cardíaca congestiva, e o tra-tamento da insuficiência cardíaca se acompanha por dimi-nuição de ANP. No entanto, em fases avançadas de des-compensação cardíaca, os efeitos do sistema nervoso sim-pático e do sistema renina-angiotensina-aldosterona pas-sam a dominar, levando a um estado refratário ao ANP,contribuindo com a retenção de sal e água.
O PRÓ-ANP está presente no sangue de pacientes cominsuficiência cardíaca congestiva classe I (assintomática),e o peptídeo natriurético cerebral (BNP) (ver adiante) secorrelaciona com o grau de disfunção miocárdica avalia-do pelo ecocardiograma. Assim, os peptídeos natriuréticossão marcadores da disfunção cardíaca.54,65
TAQUICARDIA SUPRAVENTRICULARNa taquicardia supraventricular pode haver diurese/
natriurese induzidas pela liberação anômala de ANP de-sencadeada pela estimulação elétrica das fibras atriais.65
HIPERTENSÃO ARTERIALO ANP reduz a pressão arterial porque, além dos efei-
tos natriurético, diurético e vasodilatador, suprime a libe-

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 115/121
capítulo 7 83
ração de renina e aldosterona. Os níveis de ANP em paci-entes portadores de hipertensão arterial essencial são muitovariáveis.
A administração de ANP em pacientes hipertensos re-sulta em diminuição da pressão arterial. No entanto, o usode ANP como um agente anti-hipertensivo é limitado de-vido à falta de apresentação via oral desta substância. Ouso de preparações de ANP intranasal mostrou ser de di-fícil manuseio: pequenas doses têm pouco efeito e dosesum pouco maiores causam hipotensão grave.66,67
CIRROSE HEPÁTICANa cirrose hepática avançada ocorre retenção de sódio
e água, situação na qual existe a possibilidade do envolvi-mento do ANP. Em pacientes com ascite, os níveis plas-máticos de ANP encontram-se elevados. Entretanto, comona insuficiência cardíaca congestiva, parece existir umafalta de resposta do rim ao ANP, provavelmente por pre-domínio do estado de vasoconstrição induzido por ativa-ção do sistema renina-angiotensina-aldosterona e do sis-tema nervoso simpático, impedindo a ação do ANP eminduzir diurese e natriurese.
A infusão de ANP em pacientes cirróticos com asciteresultou em modesta e transiente natriurese e diurese.Adicionalmente causou como efeito colateral hipotensãoarterial grave.
DOENÇA RENALOs níveis de ANP na síndrome nefrótica estão geralmente
diminuídos, apesar do aumento do sódio e da água corpo-ral total, sugerindo que nesta situação existe uma diminui-ção do volume sanguíneo efetivo, o que realmente acontece
em alguns tipos de síndrome nefrótica. A administração deANP não se acompanha por aumento da excreção renal desódio e água, demonstrando um estado de falta de respostaao ANP na síndrome nefrótica.68 No entanto, este estado derefratariedade ao ANP não foi observado em humanos.69
Na insuficiência renal crônica, os níveis de ANP estãodiretamente relacionados ao grau de expansão do volume.
Em pacientes em hemodiálise, os níveis de ANP pré-diáli-se encontram-se muito elevados, sendo que após a diálisehá redução de ANP circulante.
Finalmente, em casos de obstrução ureteral bilateral, oANP pode estar envolvido na resposta diurética e natriuré-tica que ocorre após a desobstrução.
PEPTÍDEO NATRIURÉTICOCEREBRAL (BNP)
Este peptídeo foi inicialmente detectado em cérebro deporco (daí o nome em inglês, brain natriuretic peptide). Noentanto, seu principal sítio de produção é no ventrículo car-díaco, sendo secretado de forma constitutiva e não regu-lada.
O BNP tem 32 aminoácidos e é estruturalmente seme-
lhante ao ANP, pois também tem um anel de 17 aminoáci-dos ligados por ponte de dissulfeto. A seqüência está lo-calizada na parte carboxiterminal de um transcrito de 134aminoácidos (PRÉ-PRÓ-BNP). Após a remoção de 26 ami-noácidos obtém-se uma molécula de 108 aminoácidos, oPRÓ-BNP, e deste é formado o peptídeo ativo, após cliva-gem proteolítica.
A secreção de BNP aumenta enormemente na hipertro-fia do ventrículo esquerdo. Nesta condição, o pró-hormô-nio também é liberado na circulação, mas não está prova-do que este pró-hormônio circulante possa funcionar comouma reserva periférica de BNP.
Os efeitos biológicos do BNP são os mesmos do ANP:natriurese e diurese, hipotensão arterial e diminuição dovolume intravascular e diminuição de renina e aldostero-
na. Tal panorama é esperável, uma vez que os dois peptí-deos agem através do mesmo receptor, isto é, via GC-A.
Como descrito anteriormente, o ANP não tem um efei-to diurético importante na insuficiência cardíaca. Entretan-to, o BNP mantém seu efeito natriurético mesmo na insu-ficiência cardíaca congestiva. Isto se deve ao fato de que oBNP tem menos afinidade pelas vias de degradação dospeptídeos natriuréticos, seja pelos CR, seja pela endopep-tidase neutra NEP 24.11, e portanto tem uma meia-vidamaior. A meia-vida do BNP é de 8 a 22 minutos, compara-da aos 1 a 4 minutos do ANP.54,65
Peptídeo Natriurético do Tipo C (CNP)
Este peptídeo tem 22 aminoácidos e foi inicialmente iso-lado de cérebro de porco. Parece estar restrito ao cérebro,mais exatamente ao tálamo, cerebelo e hipotálamo. O CNPse liga ao receptor GC-B, cuja sinalização intracelular e efei-tos biológicos desencadeados são diferentes do receptorque liga os peptídeos ANP e BNP.
A injeção sistêmica de CNP provoca hipotensão arteriale diminuição do débito cardíaco; no entretanto o peptídeo
Pontos-chave:
Patologias que cursam com aumento de ANP
• Insuficiência renal com hipervolemia• Ascite• Insuficiência cardíaca
• Obstrução ureteral• Taquicardia supraventricular
Patologias que cursam com diminuição de
ANP
• Síndrome nefrótica (com diminuição dovolume efetivo)

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 116/121
84 Peptídeos Vasoativos e o Rim
é completamente desprovido de efeitos renais. Além disso,o CNP é antimitogênico para vasos estimulados por diver-sos fatores de crescimento (FGF, PDGF, EGF) e pode estar
envolvido no controle da fibrose vascular induzida por hi-pertensão arterial. Uma analogia pode ser traçada aqui como sistema renina-angiotensina. Um dos motivos pelos quaiso bloqueio do SRA se mostrou superior terapeuticamentequando comparado, por exemplo, com a hidralazina é o fatode que aquele tratamento inibe a proliferação e a fibroseinduzidas por Ang II. De modo semelhante, o CNP (ou pos-síveis agonistas do receptor GC-B) tem vantagem terapêu-tica teórica, ainda não comprovada, sobre os peptídeos queagem no receptor GC-A,65 pois inibiriam a proliferação in-duzida por fatores de crescimento (FGF, PDGF, EGF).
URODILATINA
Em 1988 foi identificado um peptídeo natriurético naurina humana que era praticamente idêntico ao ANP, ex-ceto pela adição de quatro aminoácidos suplementares àextremidade amino-terminal. Na verdade este peptídeo écodificado pelo mesmo gene do ANP e é produzido atra-vés da clivagem da molécula precursora em um sítio dife-rente do sítio de clivagem do ANP. Tal peptídeo foi deno-minado urodilatina (Fig. 7.6).70
A urodilatina é produzida no córtex renal no néfron dis-tal (túbulo contorcido distal, túbulo de conexão e túbulocoletor cortical) e age preferencialmente no ducto coletor
medular interno, local onde se dá a regulação fina da ex-creção de sódio. Deve ser ressaltado que muito pouco doque é filtrado de peptídeos natriuréticos alcança o ducto
coletor medular interno pela luz tubular, uma vez que háuma grande quantidade de endopeptidases neutras notúbulo contorcido proximal. Estas endopeptidases virtu-almente extinguem qualquer traço de peptídeos natriuré-ticos no fluido tubular. Assim, a urodilatina é o peptídeocapaz de impedir a absorção de sódio agindo na superfí-cie luminal do ducto coletor medular interno, onde há defato receptores para o peptídeo. A urodilatina age nestesítio, bloqueando a absorção de sódio via canal de sódiosensível ao amiloride.71 Entretanto, os outros peptídeos na-triuréticos também podem estimular as células do ductocoletor medular interno, mas atingindo o rim por via sis-têmica, ativando receptores na superfície basolateral.
A urodilatina tem variação circadiana concomitante-mente à excreção de sódio, donde se infere sua importân-cia na regulação fisiológica na excreção deste íon. Outropapel importante da urodilatina é o fato de que ela é o prin-cipal modulador da natriurese que ocorre após infusãosalina. Estas funções permanecem ativas mesmo na dener-vação do coração e podem ocorrer independentemente dasecreção de ANP.
Os efeitos biológicos da urodilatina, assim como os doANP, são mediados por receptores GC-A, mas a urodila-tina é um natriurético mais potente que o ANP, mesmoquando os dois peptídeos agem sob uma mesma pressãode perfusão.
Fig. 7.6 Estrutura dos peptídeos natriuréticos. Os aminoácidos conservados estão em azul-escuro e os variantes estão em amarelo.Observar que a urodilatina é idêntica ao ANP exceto pela adição de quatro aminoácidos à extremidade aminoterminal (parte supe-rior dos desenhos).
URODILATINACNP
ANP BNP

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 117/121
capítulo 7 85
Embora a urodilatina não esteja aumentada na insufici-ência cardíaca congestiva, ela pode ser benéfica nesta pa-tologia, uma vez que sua infusão em pacientes com esta
síndrome leva à natriurese e diurese, efeitos estes que nãosão vistos na infusão de ANP. O mesmo pode ocorrer nainsuficiência renal aguda.70,71
Pontos-chave:
Efeitos do BNP
• Natriurese e diurese• Vasodilatação e hipotensão• Diminuição da secreção de vasoconstritores
Efeitos do CNP
• Vasodilatação e hipotensão• Antimitogênese
Efeitos da urodilatina
• Natriurese e diurese
ENDOTELINA
A endotelina (ET) é uma substância produzida peloendotélio vascular que apresenta um potente efeito vaso-constritor.72 A família das endotelinas é composta por trêspeptídeos de 21 AA: endotelina-1 (ET-1), endotelina-2 (ET-2) e endotelina-3 (ET-3). A expressão das três endotelinasé diferente nos diversos tecidos. A ET-1 é a única endoteli-na expressa pelas células do endotélio vascular e foi tam-
bém detectada em cérebro, rim e pulmão. ET-1 é a formaclássica de endotelina e é a única forma do peptídeo iden-tificada na circulação humana. ET2 e ET3 são produzidosno cérebro, rim, supra-renal e intestino. Todas as isoformasde endotelina são potentes vasoconstritores.
Biossíntese e Estrutura
Três genes para endotelina humana foram identificados,cada um deles codificando um produto diferente.73 A ET-1 humana é derivada de PRÉ-PRÓ-endotelina, um precur-sor de 210 resíduos.74 A PRÉ-PRÓ-endotelina-1 é convertidaem pré-endotelina intermediária, também chamada de“ET-1 grande”. A “ET-1 grande” é convertida em ET-1 ati-
va através da enzima conversora de endotelina.A secreção de ET-1 por células endoteliais é controladaem nível transcripcional, e estas células não armazenamET-1 para liberação. A expressão de mRNA da PRÉ-PRÓ-endotelina aumenta induzida por trombina, epinefrina,estresse na parede do vaso, TGF-b, IL-1 e AII.75,76 No rim, aendotelina é produzida pelas células endoteliais renais,glomérulo e também pelas células tubulares renais.75
Receptores para Endotelina eMecanismo de Ação Celular
Receptores específicos para endotelina foram identifi-cados em diversos tecidos.77 Existem dois subtipos de re-ceptores para endotelina, receptor tipo A (ET-RA) e recep-tor tipo B (ET-RB), que apresentam afinidades diferentespara as várias isoformas de endotelina. ET-1 age principal-mente no ET-RA. A afinidade para ET-RA das diferentesendotelinas é: ET-1ET-2ET-3. Já o ET-RB tem afinida-de semelhante para as três isoformas.
O ET-RA está expresso nas células da musculatura lisados vasos. O ET-RB está presente nas células endoteliais eparece ser o responsável pela liberação de prostaciclina eóxido nítrico. A este respeito é curioso notar que na infu-são de endotelina inicialmente ocorre uma vasodilataçãofugaz seguida de vasoconstrição sustentada.
A ligação da ET com seu receptor leva à ativação da viado fosfatidil inositol,77 com estimulação da fosfolipase C,que aumenta o cálcio intracelular pelo aumento do influ-xo de cálcio através dos canais de cálcio. A ET leva à libe-ração de ácido araquidônico por ativação da fosfolipase A2.Além disso, a ET age despolarizando o potencial de mem- brana e aumentando a bomba de Na/H, e assim alcali-niza o interior da célula e inibe a Na+/K+-ATPase.
No rim, receptores para endotelina foram identificadosprincipalmente na vasculatura renal, incluindo as alças ca-pilares glomerulares, capilares peritubulares, vasa recta, eno endotélio das artérias e veias arqueadas, assim como nasarteríolas renais. Em menor intensidade nas células mesan-giais, nos túbulos proximais e nos ductos coletores.76,77
Ações da Endotelina no RimA ET é um vasoconstritor renal potente, 30 vezes mais
potente que a AII.76 A injeção de endotelina endovenosaleva a uma resposta pressora bifásica caracterizada inici-almente por uma resposta vasodilatadora periférica e di-minuição da pressão sanguínea de curta duração, prova-velmente devido à liberação de óxido nítrico e prostacicli-na e diminuição de liberação de norepinefrina. Em segui-da, ocorre uma resposta de vasoconstrição que leva aoaumento da pressão arterial sistêmica.
No rim, ET-1 produz vasoconstrição renal também pre-cedida de uma resposta vasodilatadora de curta duração.Em resposta ao ET-1, ocorre vasoconstrição tanto nas arte-
ríolas aferentes como nas eferentes. Infusões de ET direta-mente na artéria renal causam primariamente constriçãoda arteríola aferente, enquanto a administração endoveno-sa de doses maiores causa um maior efeito no vaso eferen-te.76 A infusão endovenosa de ET-1 em humanos resulta emdiminuição do fluxo sanguíneo renal (dose-dependente)com diminuição da taxa de filtração glomerular. A ET cau-sa uma diminuição do Kf, o qual pode ser explicado, pelo

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 118/121
86 Peptídeos Vasoativos e o Rim
menos em parte, pela contração da célula mesangial indu-zida pela ET-1.
Em doses que não diminuem a taxa de filtração glome-rular, o ET é natriurético.74 Este efeito é abolido se a pres-são de perfusão renal for mantida constante, o que sugereque a natriurese é pelo menos em parte relacionada à pres-são. Entretanto, também há evidências de um efeito tubu-lar direto.74 É discutível se a liberação de ANP induzida porET participe da resposta natriurética.
A ET tem outros efeitos que podem ser importantes nahomeostase do fluido e de eletrólitos. ET-1 estimula a li- beração adrenal de catecolaminas e aldosterona. Apesar deinibir a liberação de renina das células justaglomerularesin vitro, a ET, quando administrada por via sistêmica, au-menta a atividade da renina plasmática.74,78 Os níveis plas-máticos de ANP aumentam com a infusão de ET-1.
A ET pode ter um papel na inflamação glomerular atra-
vés do efeito mitogênico. A ET tem ação mitogênica nascélulas da musculatura lisa dos vasos, células mesangiaise fibroblastos. As células mesangiais humanas em culturaexpressam mRNA para ET-1 e a expressão do gene de ET-1 é aumentada com a incubação com certos mediadores in-flamatórios.79
Pontos-chave:
Efeitos da endotelina
• Vasoconstrição sistêmica• Aumento da pressão arterial• Mitogênese
Endotelina em Situações Fisiopatológicas
INSUFICIÊNCIA RENAL AGUDAA ET regula a hemodinâmica sistêmica e renal e tem
papel na reperfusão após a injúria induzida por isquemiarenal. Cinco minutos após a isquemia, os níveis de ET-1estão significativamente elevados. Utilizando-se hibridiza-
ção in situ, foi possível detectar aumento do mRNA paraET-1 no glomérulo e nos ductos coletores 12 horas apósisquemia com normalização 48 horas após. Concomitan-
temente, o mRNApara ET-RB está aumentado nestas regi-ões. O uso de antagonista de receptor de endotelina, assimcomo o uso de anticorpos antiendotelina, tem efeito pro-tetor.78
HIPERTENSÃO ARTERIALO papel da ET na hipertensão arterial tem sido ampla-
mente discutido, uma vez que esta substância causa poten-te vasoconstrição. Em ratos hipertensos, a administraçãode fosfaramidone, que bloqueia a ação de enzima conver-sora da endotelina, reduz a pressão arterial.
Os níveis circulantes de endotelina não estão aumenta-dos em diversas formas de hipertensão arterial. No entan-to, pacientes hipertensos com insuficiência renal crônica
apresentam níveis de ET maiores que os de indivíduosnormotensos.
CICLOSPORINA AA ciclosporina leva a alterações da hemodinâmica renal
caracterizadas por vasoconstrição com aumento da resis-tência vascular renal e diminuição da taxa de filtração glo-merular. Foi demonstrado que a ciclosporina aumenta aliberação de ET de células endoteliais em cultura. Alémdisso, pacientes em uso de ciclosporina apresentam aumen-to dos níveis sanguíneos de ET. Anticorpos antiendotelinae antagonistas do receptor de endotelina abolem a vaso-constrição induzida por ciclosporina. Assim, a endotelinapode ser a responsável pela diminuição do fluxo sanguí-neo renal durante a administração de ciclosporina e possi-
velmente por sua nefrotoxicidade.
DOENÇAS GLOMERULARESÉ sabido que a ET é um potente mitógeno e que pode
mediar os efeitos proliferativos de várias citocinas e que,portanto, pode participar da patogênese de doenças infla-matórias. A produção renal de ET está aumentada na glo-merulonefrite humana e experimental e há maior expres-são do receptor ETB na glomerulonefrite (GN) em ratos.Além disso, a ET está aumentada no plasma de pacientescom doenças glomerulares. O uso de antagonista de recep-tor de ET diminui a proliferação mesangial na glomerulo-nefrite experimental e diminuiu a lesão renal no lúpusmurino.
FIBROSE RENAL E INSUFICIÊNCIARENAL CRÔNICA
A ET-1 induz acúmulo de matriz e conseqüentementeleva à fibrose. A ET age na produção de componentes damatriz (fibronectina e colágeno), na inibição de sua degra-dação (metaloproteases) e na liberação de citocinas fibro-gênicas (TGF-). O uso de um antagonista de receptor de
Fig. 7.7 Biossíntese do ANP.
Met
EXON 1
COOH
EXON 2 EXON 3
Pré-Pró-ANP
Pró-ANP
ANP
cDNA
151 AA
126 AA
28 AA

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 119/121
capítulo 7 87
angiotensin system in human tissues. Quantitative analysis by thepolymerase chain reaction. J. Clin. Invest., 91:2058-2064, 1993.
6. LARAGH, J.H.; SEALEY, J.E. The renin-angiotensin-aldosteronesystem for normal regulation of blood pressure and sodium and
potassium homeostasis. In: Hypertension: Pathophysiology, Diagnosisand Management, 2nd edition, edited by J.H. Laragh and B.M. Brenner.Raven Press, New York, pp. 1763, 1995.
7. CELIO, M.R. Angiotensin II immuno-reactivity coexisting with reninin the human juxtaglomerular epithelioid cells. Kidney Int., 22(sup-pl. 12):S30-32, 1982.
8. CANTIN, M.; GUTKOWSKA, J.; LACASSE, J. Ultrastructural im-munocytochemical localization of renin and angiotensin II in the juxtaglomerular cells of the ischaemic kidney in experimental renalhypertension. Am. J. Pathol., 115:212, 1984.
9. TAUGNER, R.; KIM, S.J.; MURAKAMI, K.; WALDHERR, R. The fateof prorenin during granulopoiesis in epithelioid cells. Histochemistry,86:249, 1987.
10. HACKENTHAL, E.; PAUL, M.; GANTEN, D.; TAUGNER, R.Morphology, physiology, and molecular biology of renin secretion.Physiol. Rev., 70:1067, 1990.
11. BRIGGS, J.P.; SCHNERMANN, J. Control of renin release and glo-merular vascular tone by the juxtaglomerular apparatus. In: Hyper-
tension: Pathophysiology, Diagnosis and Management, 2nd
edition, edited by J.H. Laragh and B.M. Brenner. Raven Press, New York, pp. 1359,1995.
12. DiBONA, G.F. Neural control of renal function: Cardiovascularimplications. Hypertension, 13:539, 1989.
13. TIDGREN, B.; HJEMDAHL, P. Renal responses to mental stress andepinephrine in humans. Am. J. Physiol., 257:F682, 1989.
14. BEIERWALTES, W.H. Possible endothelial modulation of prosta-glandin-stimulated renin release. Am. J. Physiol., 258:F1363, 1990.
15. BACHMANN, S.; OBERBÄUMER, I. Structural and molecular dis-section of the juxtaglomerular apparatus: new aspects for the roleof nitric oxide. Kidney Int., 54(Suppl. 67):S29-S33, 1998.
16. LORENZ, J.N.; WEINPRECHT, H.; SCHNERMANN, J.; SK∅TT, O.,BRIGGS, J.P. Characterization of the macula densa for renin secre-tion. Am. J. Physiol., 259:F186-F193, 1990.
17. CHAI, S.Y.; JOHNSTON, C.I. Tissue distribution of angiotensin-con-verting enzyme. In: Hypertension: Pathophysiology, Diagnosis and Management, 2nd edition, edited by J.H. Laragh and B.M. Brenner.Raven Press, New York, pp. 1683, 1995.
18. METZGER, R.; BOHLE, R.M.; KATHARINA, P. Angiotensin-con-verting enzyme in non-neoplastic kidney diseases. Kidney Int.,56:1442-1454, 1999.
19. BRAAM, B.; MITCHELL, K.D.; FOX, J.; NAVAR, L.G. Proximal tu- bular secretion of angiotensin II in rats. Am. J. Physiol., 264:F891-F898,1993.
20. FORSYTH, R.P.; HOFFBRAND, B.I.; MELMON, K.L. Hemodyna-mic effects of angiotensin in normal and environmentally stressedmonkeys. Circulation, 44:119, 1971.
21. ROSIVALL, L.; NAVAR, G. Effects on renal hemodynamics of in-tra-arterial infusions of angiotensins I and II. Am. J. Physiol., 245:F181,1983.
22. ICHIKAWA, I.; HARRIS, R.C. Angiotensin actions in the kidney:Renewed insight into the old hormone. Kidney. Int., 40:583, 1991.
23. MEZZANO, S.A.; RUIZ-ORTEGA, M.; EGIDO, J. Angiotensin II andrenal fibrosis. Hypertension, 38[part 2]:635-638, 2001.
24. MORGAN, H.E.; BAKER, K.M. Cardiac hypertrophy: Mechanical,neural, and endocrine dependence. Circulation, 83:13, 1991.
25. GUNTHER, S.; ALEXANDER, R.W.; ATKINSON, W.J.; GIMBRO-NE, M.A. Jr. Functional angiotensin II receptors in cultured vascu-lar smooth muscle cells. J. Cell. Biol., 92:289, 1982.
26. MIYATA, N.; PARK, F.; LI, X.F.; COWLEY, A.W. Distribution of AT1
and AT2
receptors subtypes in the rat kidney. Am. J. Physiol.,277:F437-F446, 1999.
27. HARRISON-BERNARD, L.M.; GABRIEL NAVAR, L.; HO, M.M.;VINSON, G.P.; EL-DAHR, S.S. Immunohistochemical localizationof ANG II AT1 receptor in adult rat using a monoclonal antibody. Am. J. Physiol., 273:F170-F177, 1997.
ET diminui o acúmulo de matriz em modelos experimen-tais de glomerulonefrite. Interessantemente, o uso destes bloqueadores de ET promove diminuição da proteinúria,
hipertensão e elevação da creatinina no modelo de abla-ção renal.
Pontos-chave:
Patologias potencialmente tratáveis pela
inibição da endotelina
• Insuficiência renal aguda• Hipertensão arterial• Toxicidade por ciclosporina• Glomerulonefrites• Insuficiência renal crônica
ADRENOMEDULINA
A adrenomedulina é um peptídeo vasodilatador de 52aminoácidos, inicialmente isolado de amostras de feocromo-citoma, mas que está presente na medula adrenal normal.Ela também é encontrada em outros tecidos como cérebro,coração e pulmões. No rim, o peptídeo é expresso no glo-mérulo e túbulos coletores, corticais e medulares. A adre-nomedulina é derivada de um PRÉ-PRÓ-hormônio de 185aminoácidos, codificado por um gene no cromossoma 11.Inicialmente esta molécula precursora é convertida em PRÓ-hormônio de 164 aminoácidos e então na molécula ativa.80
Seus efeitos renais incluem natriurese e diurese e sãodevidos a aumento da taxa de filtração glomerular e dimi-
nuição da reabsorção de sódio. A concentração plasmáti-ca da adrenomedulina está aumentada na hipertensão ar-terial, insuficiência renal crônica e insuficiência cardíacacongestiva, possivelmente como mecanismo compensadorà vasoconstrição e à retenção de sal e água que ocorre nes-tas doenças.
REFERÊNCIAS BIBLIOGRÁFICAS
1. HSUEH, W.A.; ANTONIPILLAI, I. Renin-angiotensin system. In:Textbook of Nephrology, 3.ª edição, editado por Massry, S.G. e Glas-sock, R.J. Baltimore, Williams & Wilkins, pp. 197, 1995.
2. BALLERMANN, B.J.; ZEIDEL, M.L.; GUNNING, M.E.; BRENNER,B.M. Vasoactive peptides and the kidney. In: The Kidney, 5.ª edição,
volume 1, editado por Brenner, B.M. Philadelphia, WB Saunders Co,pp. 510, 1986.3. RABKIN, R.; DAHL, D.C. Hormones and the kidney. In: Diseases of
the Kidney. 5.ª edição, editado por Schrier, R.W. e Gottschalk, C.W.Little, Brown and Company, pp. 283, 1993.
4. SANTOS, R.A.S.; PASSAGLIO, K.T.; PESQUERO, J.B.; BADER, M.;SIMÕES E SILVA, A.C. Interactions between angiotensin-(1-7),kinins, and angiotensin II in kidneys and blood vessels. Hypertensi-on, 38[part II]:660-664, 2001.
5. PAUL, M.; WAGNER, J.; DZAU, V.J. Gene expression of the renin-

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 120/121
88 Peptídeos Vasoativos e o Rim
28. SIRAGY, H.M. AT1 and AT2 receptors in the kidney: role in disea-se and treatment. Am. J. Kidney Dis., 36 (Suppl. 1):S4-S9, 2000.
29. TSUTSUMI, Y.; MATSUBARA, H.; MUSAKI, H. Angiotensin II type2 receptor expression activates the vascular kinin system and cau-
ses vasodilation. J. Clin. Invest., 104:925-935, 1999.30. RUIZ-ORTEGA, M.; LORENZO, O.; EGIDO, J. Angiotensin III up-
regulates genes involved in kidney damage in mesangial cells andrenal interstitial fibroblasts. Kidney Int., 54 (Suppl. 68):S41-S45, 1998.
31. MOELLER, I.; ALLEN, A.M.; CHAI, S-Y.; MENDELSON, F.A.O.Bioactive angiotensin peptides. J. Human Hypertens., 12:289-293, 1998.
32. ROCHA E SILVA, M.; BERALDO, W.T.; ROSENFELD, G. Bradyki-nin, a hypotensive and smooth muscle stimulating factor releasedfrom plasma globin by snake venoms and by trypsin. Am. J. Physi-ol., 156:261-273, 1949.
33. MARGOLIUS, H.S. Kallikrein-kinin system. In: Textbook of Nephro-logy, 3.ª edição, editado por Massry, S.G. e Glassock, R.J . Baltimore,Williams & Wilkins, pp. 203, 1995.
34. BHOOLA, K.D.; FIGUEROA, C.D.; WORTHY, K. Bioregulation ofkinins: kallikreins, kininogens, and kininase. Pharmacol. Rev., 44:1-80, 1992.
35. SCICLI, A.G.; CARRETERO, O.A. Renal kallikrein-kinin system.Kidney Int., 29:120, 1986.
36. URA, N.; CARRETERO, O.A.; ERDOS, E.G. Role of renal endopep-tidase 24.11 in kinin metabolism in vitro and in vivo. Kidney Int.,32:507, 1987.
37. NASJLETTI, A.; COLESSA-CHORERIO, J.; McGIFF, J.C. Disappea-rance of bradykinin in the renal circulation of dogs: Effects of kini-nase inhibition. Cir. Res., 37:59, 1975.
38. CAMPBELL, D.J. Towards understanding the kallikrein-kinin sys-tem: insights from measurements of kinin peptides. Braz. J. Med. Biol.Res., 33:665-677, 2000.
39. FIGUEROA, C.D.; MACLVER, A.G.; MACKENZIE, J.C.; BHOOLA,K.D. Localization of immunoreactive kininogen and tissue kallikreinin the human nephron. Histochemistry, 89:437, 1988.
40. XIONG, W.; CHAO, L.; CHAO, J. Renal kallikrein mRNA localiza-tion by in situ hybridization. Kidney Int., 35:1324, 1989.
41. CARRETERO, O.A.; SCICLI, A.G. The kallikrein-kinin system as aregulator of cardiovascular and renal function. In: Hypertension:Pathophysiology, Diagnosis and Management, 2nd edition, edited by J.H.Laragh and B.M. Brenner. Raven Press, New York, pp. 983, 1995.
42. BASCANDS, J.L.; PECHER, C.; ROUAUD, S.; EDMOND, C.; TACK, J.L.; BASTIE, M.J.; BURCH, R.; REGOLI, D.; GIROLAMI, J.P. Evi-dence for existence of two distinct bradykinin receptors on rat me-sangial cells. Am. J. Physiol., 264:F548, 1993.
43. WEBSTER, M.E.; GILMORE, J.P. Influence of kallidin-10 on renalfunction. Am. J. Physiol., 206:714, 1964.
44. GILL, J.R.; MELMON, K.L.; GILLESPIE, L.; BARTTER, F.C. Bradyki-nin and renal function in normal man: Ef fects of adrenergic blocka-de. Am. J. Physiol., 209:844, 1965.
45. GRANGER, J.P.; HALL, J.E. Acute and chronic actions of bradykininon renal function and arterial pressure. Am. J. Physiol., 248 :F87, 1985.
46. EDWARDS, R.M. Response of isolated renal arterioles to acetylco-line, dopamine and bradykinin. Am. J. Physiol., 248:F183, 1985.
47. KAUKER, M.L. Bradykinin action on the efflux of luminal 22Na inthe rat nephron. J. Pharmacol. Exp. Ther., 214:119, 1980.
48. BEIERWALTES, W.H.; SCHRYVER, S.; SANDERS, E. Renin releaseselectively stimulated by prostaglandin PGI2 in isolated rat glome-ruli. Am. J. Physiol., 243:F276, 1982.
49. DeBOLD, A.J.; BORENSTEIN, H.B.; VERESS, A.T.; SONENBERG,A. A rapid and potent natriuretic response to intravenous injectionof atrial myocardial extract in rats. Life Sci., 28:89, 1981.
50. BRENNER, B.M.; BALLERMANN, B.J.; GUNNING, M.E.; ZEIDEL,M.L. Diverse biological actions of atrial natriuretic peptide. Physiol.Rev., 70:665, 1990.
51. COGAN, M.G. Atrial natriuretic peptide.Kidney Int., 37:1148, 1990.52. TANG, J.; FEI, H.; XIE, C.W. Characterization and localization of
atriopeptin in rat atrium. Peptides, 5:1173, 1984.53. INAGAMI, T. Atrial natriuretic factor. J. Biol. Chem., 264 :3043, 1989.
54. SUZUKI, T.; YAMAZAKI, T.; YAKAZI, Y. The role of the natriure-tic peptides in the cardiovascular system. Cardiovasc. Res., 51:489-494,2001.
55. LUFT, F.C.; LANG, R.E.; ARONOFF, G.R. Atriopeptin III kinetics
and pharmacodynamics in normal and anephric rats. J. Pharmacol.Exp. Ther., 236:416, 1986.
56. YANDLE, T.G.; RICHARDS, A.M.; NICHOLLS, M.G. Metabolic cle-arance rate and plasma half life of alpha human atrial natriureticpeptide in man. Life Science, 38:1827, 1986.
57. YUKIMURA, T.; ITO, K.; TAKENAGA, T. Renal effects of synthetichuman atrial natriuretic polypeptide in anesthetized dogs. Eur. J.Pharmacol., 103:363, 1984.
58. FRIED, T.A.; McCOY, R.N.; OSGOOD, R.W.; STEIN, J.H. Effect ofatriopeptin II on determinants of glomerular filtration rate in the invitro perfused dog glomerulus. Am. J. Physiol., 250:F 1119, 1986.
59. LIGHT, D.B.; SCHWIEBERT, E.M.; KARLSON, K.H.; STANTON,B.A. Atrial natriuretic peptide inhibits a caption channel in renalinner medullary collecting duct cells. Science, 243:383, 1989.
60. OELKERS, W.; KLEINER, S.; BAHR, V. Effects of incrementalinfusions of atrial natriuretic factor on aldosterone, renin, and bloodpressure in humans. Hypertension, 12:462, 1988.
61. MAACK, T.; MARION, D.N.; CAMARGO, M.J. Effects of auriculin
on blood pressure, renal function, and the renin-aldosterone systemin dogs. Am. J. Medicine, 77:1069, 1984.62. OPGENORTH, T.J.; BURNETT, J.C. Jr.; GRANGER, J.P.; SCRIVEN,
T.A. Effects of atrial natriuretic peptide on renin secretion innonfiltering kidney. Am. J. Physiol., 250:F798, 1986.
63. KURTZ, A.; DELLA BRUNA, R.D.; PFEILSCHIFTER, J. Atrial na-triuretic peptide inhibits renin release from juxtaglomerular cells by a cGMP-mediated process. Proc. Natl. Acad. Sci. USA, 83:4769,1986.
64. KUDO, T.; BAIRD, A. Inhibition of aldosterone production in theadrenal glomerulosa by atrial natriuretic factor. Nature, 312:756, 1984.
65. LEWICKI, J.A.; PROTTER, A.A. Physiological studies of the natriu-retic peptide family. In: Hypertension: Pathophysiology, Diagnosis and Management, 2nd edition, edited by J.H. Laragh and B.M. Brenner.Raven Press, New York, pp. 1029-1053, 1995.
66. JANSSEN, W.M.T.; deJONG, P.E.; VAN DER HEM, G.K.; deZEEUW, D. Effect of human atrial natriuretic peptide on blood pres-sure after sodium depletion in essential hypertension. Br. Med. J.,
293:351, 1986.67. RICHARDS, A.M.; NICHOLLS, M.G.; ESPINER, E.A. Effects ofalpha-human atrial natriuretic peptide in essential hypertension. Hypertension, 7:812, 1985.
68. HILDEBRANDT, D.A.; BANKS, R.O. Effects of atrial natriureticfactor on renal function in rats with nephrotic syndrome. Am. J. Phy-siol., 254:F210,1988.
69. ZIETSE, R.; SCHALEKAMP, M.A. Effect of synthetic human atrialnatriuretic peptide (102-126) in nephrotic syndrome. Kidney Int.,34:717, 1988.
70. FORSSMANN, W.G.; MEYER, M.; FORSSMANN, K. The renalurodilatin system: clinical implications. Cardiovasc. Res., 51:450-462,2001.
71. GUNNING, M.; BRENNER, B.M. Urodilatin. In: Hypertension: Pa-thophysiology, Diagnosis and Management, 2nd edition, edited by J.H.Laragh and B.M. Brenner. Raven Press, New York, pp. 1021-1027,1995.
72. YANAGISAWA, M.; KURIHARA, H.; KIMURA, S. A novel potent
vasoconstrictor peptide produced by vascular endothelial cells. Na-ture, 332:411, 1988.73. RUBANYI, G.M.; BOTELHO, L.H.P. Endothelins. FASEB J., 5:2713,
1991.74. MARSDEN, P.A.; GOLIGORSKY, M.S.; BRENNER, B.M. Endothe-
lial cell biology in relation to current concepts of vessel wall struc-ture and function. J. Am. Soc. Nephrol., 1:931, 1991.
75. LUSCHER, T.F.; BOCK, H.A.; YANG, Z.; DIEDERICH, D. Endothe-lin-derived relaxing and contracting factors: Perspectives in nephro-logy. Kidney Int., 39:575, 1991.

5/10/2018 Riella - Seção 01 - Estrutura e Função Renal - slidepdf.com
http://slidepdf.com/reader/full/riella-secao-01-estrutura-e-funcao-renal 121/121
capítulo 7 89
76. KING, A.J.; BRENNER, B.M. Endothelium-derived vasoactive fac-tors and the renal vasculature. Am. J. Physiol., 260:R653, 1991.
77. SIMONSON, M.S.; DUNN, M.J. Cellular signaling by peptides of theendothelin gene family. FASEB J., 4:2989, 1990.
78. NAICKER, S.; BHOOLA, K.D. Endothelins: vasoactive modulatorsof renal function in health and disease. Pharmacology & Therapeutics,90:61-88, 2001.
79. SIMONSON, M.S.; DUNN, M.J. Endothelin peptides: A possible rolein glomerular inflammation. Lab. Invest., 64:1, 1991.
80. JOUGASAKI, M.; BURNETT, J.C. Adrenomedullin: potential inphysiology and pathophysiology. Life Sci., 66:855-872, 2000.
ENDEREÇOS RELEVANTES NA INTERNET
Brazilian Journal of Medical and Biological Researchhttp://www.scielo.br/scielo.php?scriptsci-serial&pid0100-879X&lngen&nrmiso
American Heart Associationhttp://www.americanheart.com
American Heart Association (journals)http://www.ahajournals.org
American Society of Physiologyhttp://www.the-aps.com
American Society of Physiology (journals)http://intl-ajpcon.physiology.org
Nomenclatura de peptídeos (IUPAC)http://www.chem.qmw.ac.uk
Grupos de discussão sobre peptídeoshttp://www.bio.net/hypermail/PEPTIDES
Federation of American Societies for Experimental Biology(FASEB)http://www.faseb.org





























