Embed Size (px)
Citation preview

nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology 189
review articlePuBLIsHed onLIne: 12 feBruary 2010 | doI: 10.1038/nCHeMBIo.326
Proteins constitute vital components of life, and protein synthesis represents one of the most fundamental biochemical functions. Errors in post-transcriptional processing and translation have
deleterious consequences. En route from transcription to transla-tion, (pre-)mRNAs meet two of the largest macromolecular assem-blies in the cell: the spliceosome and the ribosome. Owing to the availability of antibacterial antibiotics, the prokaryotic translation machinery had been fairly well understood, even before high-reso-lution crystal structures of the ribosome became available. Progress on the eukaryotic counterpart, however, has been hampered by the absence of known specific inhibitors. As for splicing, until recently inhibitors were lacking completely. In this review we summarize recent advances in the identification of small molecules (Fig. 1) tar-geting the eukaryotic splicing and translation machinery and their utility as biochemical probes, as well as their clinical promise.
small molecules targeting pre-mrna splicingThe spliceosome consists of five small nuclear ribonucleopro-tein particles (snRNPs)—U1, U2, U4, U5 and U6—and many non-snRNP proteins. Each snRNP contains a small nuclear RNA (snRNA) core and several proteins that bind the snRNA to form a mature snRNP (refs. 1,2). The spliceosome assembles on a substrate pre-mRNA through the binding of individual snRNPs to their target sites. The sequences essential for recognition by snRNPs and other splicing factors are positioned around the intron-exon borders (Fig. 2a). The branch point sequence (BPS) lies at approximately 18–40 nucleotides upstream of the 3′ splice site (3′ss), and a stretch of pyri-midine residues (polypyrimidine tract, PPT) is positioned between the BPS and the 3′ss. Assembly of the spliceosome starts with the binding of the U1 snRNP to the 5′ splice site (5′ss).
Concurrently, the factor SF1 binds to the BPS, while U2AF65, a subunit of U2AF, attaches to the PPT, and U2AF35, the other U2AF subunit, adheres to the 3′ss, resulting in the formation of the E complex. In this complex, base pairing between the 5′ss and the U1 snRNA is important for splice site selection. For the formation of the A complex, SF1 is replaced by the U2 snRNP, which base pairs with the BPS. The U4/U6-U5 tri-snRNP complex joins this assem-bly to form the B complex before the U1 and U4 snRNPs dissociate, which results in the C complex, the catalytically active spliceosome.
garbled messages and corrupted translationstilman schneider-Poetsch1, takeo usui2, daisuke Kaida3 & Minoru yoshida1,4*
Following transcription, genomic information begins a long journey toward translation of its nucleotide sequence into the amino acids of a protein. In eukaryotes, synthesized pre-mRNAs become processed to mature mRNAs by 5′-end capping, splicing, 3′-end cleavage and polyadenylation in the nucleus, before being scrutinized for premature stop codons. Each step requires high precision and control to ensure that an intact and readable message is exported to the cytoplasm before finally becoming translated. Two important aspects of these processes are accurately managed by ribonucleoprotein machineries—the spliceo-some and the ribosome. Recently, several natural products targeting these macromolecular assemblies have been reported. For the first time in eukaryotes, these molecules allow chemical disruption and dissection of the sophisticated machinery that regulates post-transcriptional events. Beyond their great potential as bioprobes for investigating mRNA regulation and protein synthesis, these compounds also show promise in opening new therapeutic approaches.
In this active complex, two sequential transesterification reactions occur. First, a nucleophilic attack of the 2′ hydroxyl group of the branchpoint adenosine on the 5′ splice junction forms a branched linkage and produces a lariat structure. In the second step, the newly released 3′ hydroxyl of the 5′ exon attacks the 3′ splice junction, thereby forming a new phosphodiester bond between the 5′ and 3′ exons. In addition to the transesterification reactions, each recruit-ment or dissociation process presents a potential target for small-molecule inhibitors (Fig. 2b).
The snRNPs and some non-snRNP splicing factors are local-ized in the nuclear speckles, which are 20–50 intensely stained and irregularly shaped regions of the nucleus. An early study showed that microinjection of oligonucleotides targeted to snRNA or anti-bodies that inhibit splicing in vitro resulted in reorganization of the nuclear speckles. The irregularly shaped speckles became larger and rounded, and decreased in number, suggesting that in vivo splicing inhibition causes reorganization of splicing factors in the nucleus. Undoubtedly, the dissection of splicing itself and its relation to nuclear speckle organization would greatly benefit from specific small-molecule inhibitors.
Direct spliceosome inhibitors. Natural products FR901463, FR901464 and FR901465 were identified in a Pseudomonas sp. No. 2663 fermentation broth as enhancers of SV40 promoter transcrip-tion3–5. All three compounds showed activity, though FR901464 proved the most active. Subsequently, FR901464 was shown to potently inhibit the growth of a number of different human solid tumor cell lines at low-nanomolar half-maximal inhibitory concen-trations (IC50) and to prolong the life of tumor-bearing mice. Studies on the activity of FR901464 on the cell cycle revealed arrest in G1 and G2/M phases as well as the accumulation of a truncated form of the cyclin-dependent kinase (CDK) inhibitor p27 called p27*; how-ever, the direct target of these compounds remained unknown.
Chemical derivatization of FR901464 yielded a methyl ketal derivative, dubbed spliceostatin A (SSA), which proved as active as the parent compound but chemically more stable, as well as a bioti-nylated conjugate that retained sufficient biological activity to allow isolation of putative target proteins by affinity pulldown. These experiments revealed that SSA strongly binds the SF3b subcomplex,
1Chemical Genetics Laboratory/Chemical Genomics Research Group, RIKEN Advanced Science Institute, Wako, Saitama, Japan. 2Graduate School of Life and Environmental Sciences, University of Tsukuba, Tsukuba, Ibaraki, Japan. 3University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania, USA. 4Centre for Renewable Energy Systems Technology (CREST) Research Project, Japan Science and Technology Corporation, Saitama, Japan. *e-mail: [email protected]

190 nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology
review article NATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
which forms part of the U2 snRNP and is essential for BPS recogni-tion6. Indeed, SSA inhibited splicing before the first esterification step in vitro at low-nanomolar concentration, and pre-mRNAs of all tested genes accumulated in SSA-treated cells. Consistent with the observation that nuclear speckles change their form in splicing-inhibited cells, the number of nuclear speckles decreased under SSA treatment, though the remaining speckles increased in size. In addi-tion, the majority of poly(A) RNAs relocalized to these enlarged nuclear speckles in SSA-treated cells, suggesting that pre-mRNAs are captured by splicing factors when splicing is inhibited.
Subsequently, several groups succeeded in the total synthesis of FR901464 (refs. 7–9), and analogs such as meayamycin, which lack the hemiketal functionality, were shown to have more potent activity than the original natural product due to their increased stability. Meayamycin did indeed display splicing inhibition both in vivo and in vitro10.
Pladienolide is an antitumor macrolide that was isolated by a group at Eisai from a fermentation broth of Streptomyces platenensis Mer-11107 and that contains a 12-membered ring11,12. Pladienolide
was identified as an inhibitor of hypoxia-induced vascular endothe-lial growth factor (VEGF) expression. This compound inhibits the proliferation of several cancer cell lines and leads to complete regres-sion of tumors in mouse xenograft models. Due to its surprisingly strong antitumor activity in vivo, clinical trials using a pladienolide derivative have commenced. A mode of action study13 revealed that fluorescently tagged pladienolide localizes to nuclear speckles where splicing and transcription factors accumulate. Importantly, using 3H-labeled and photoaffinity/biotin-tagged pladienolide, they determined that pladienolide targeted the SAP130 component of the SF3b complex, suggesting that pladienolide is another specific inhibitor of SF3b. Indeed, pladienolide treatment leads to accumu-lation of pre-mRNA and changes the morphology of nuclear speck-les, although it is still unclear whether pladienolide inhibits in vitro splicing at the same step as the spliceostatins. Thus, SSA, pladieno-lide and related derivatives are the first identified small molecules that directly bind and inhibit the spliceosome6,13.
The third class of new splicing inhibitors centers on the bifla-vonoid isoginkgetin, a natural product found in a variety of plant
O
O
HN
OOH
O
O
OH
O
O
HOOC
O
OH
COOHO
H
H
O COOH
O
N
S
N OO
OO
O
O
HO
H
HOH
HOH
H HH
HN
O
O
O
O O
OH
HN O
O
OH
Pederin
Reveromycin A
Pateamine A Hippuristanol
Lactimidomycin
Cycloheximide
Translation inhibitors
OOH
OH
OOMe
OH
O
O
O O
13-Deoxytedanolide
HN
O
O
O
O O
Isomigrastatin
MeO
HO
Mycalamide A
O
O
HN
OOH
O
O
O
OH
OH
O
H2N
O
O
N
O
H
O
O
OHO
O
OH
O
O
O
O NHO
N
O
N
OO
O NH
HO
NH
O
O
N
O
N
O
HO
Homoharringtonine Didemnin B
O
NH
O O
O
OH
HO
O
NH
O O
HO
OH
HO
Cl
O
NH
O O
O
HO OH
O
O
O
O
O
OH
O
O
O
O
O
OH
OH
OOH
O
O
O
OH
OH
O
HO
OH O
O
FR901463
FR901464
FR901465
Isoginkgetin
Splicing inhibitors
Pladienolide B
O
NH
O O
O
O
HO
O
NH
O O
O
HO
O
O
O
O
Spliceostatin A
Meayamycin
O
figure 1 | overview of eukaryotic splicing and translation inhibitors.

nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology 191
review articleNATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
species, including Ginkgo biloba. Unlike FR901464 and pladieno-lide, which were discovered by their ability to modulate transcrip-tional activity, isoginkgetin was identified through screens designed for a general inhibitor of splicing using a cell-based reporter assay14. Isoginkgetin inhibited pre-mRNA splicing both in vivo and in vitro at similar micromolar concentrations. A detailed study on the mode of action indicated that isoginkgetin prevents the recruitment of the U4/U5/U6 tri-snRNP, resulting in accumulation of the spliceosomal A complex. Although it is still unclear which spliceosomal compo-nent the molecule targets, it does appear to act by a mechanism distinct from that of FR901464 or meyamamycin, both of which prevent A complex formation in the first place.
Inhibitors of splicing regulatory pathways. Indole derivatives that target SR proteins and cardiotonic steroids were identified in screens as modulators of alternative splicing15,16. SR proteins are serine- and arginine-rich molecules that become highly phosphorylated in liv-ing cells. They play a crucial role in spliceosome formation and in the regulation of alternative splicing. A benzothiazole compound called TG003, which inhibits the SR protein kinases Clk1 and Clk4, alters the splicing pattern of several genes. It also suppresses the morphological changes in nuclear speckles induced by SR kinase overexpression17.
In addition to phosphorylation, acetylation of proteins may regu-late the splicing reactions. SAP130 has been shown to associate with STAGA, a mammalian SAGA-like histone acetyltransferase (HAT) complex18. A recent global analysis of cellular lysine-acetylated proteins identified more than 100 splicing factors with more than 200 acetyla-tion sites19. Furthermore, Kuhn et al.20 examined the effects of various HAT and histone deacetylase (HDAC) inhibitors on in vitro splicing and showed that two HAT inhibitors—anacardic acid from cashew nut shell liquid21 and garcinol from Garcinia indica fruit rind22—inhibited the splicing cycle before the formation of B complex at concentrations similar to those necessary for HAT inhibition. Further careful analysis of their target specificity is necessary, but these small-molecule splic-ing inhibitors will be powerful tools in unraveling the network of post-translational modifications that regulate splicing activity and function. These inhibitors will also help elucidate the regulation of gene expres-sion between transcription and translation.
Collectively, the inhibitors described above constitute the first molecules reported to block spliceosomal function (Fig. 2b). Thus, although their exact binding sites and mechanisms still await dis-covery, the compounds are expected to provide much-needed insights into the relatively poorly understood yet critically impor-tant mechanisms and regulation of mRNA splicing.
AG GURAGU
5’ SS(U1)
a
b
CURACU
BPS(SF1 and U2)
(Yn)
PPT(U2AF)
YAG G
3’ SS(U2AF)
5’ exon
U1 U1
U1
?
SF1
SF1
E complex
U2AF
U2AF
3’ exon
U1
U1
FR901464SSA
MeayamycinPladienolide
Isoginkgetin
U2
U2
U2
U4
A complex
B complex
U2AF
U4/6U5
U4/6U5
U6
U2
C complex
+
U5
U6
U6
U2
U2
U5
U5
figure 2 | recognition of sequences around the intron-exon borders by splicing factors. (a) Summary of splice complex formation and the splice sequence of events. SS, BPS and PPT denote the splice site, branch point sequence and polypyrimidine tract, respectively. (b) The spliceostatin group of molecules interferes with splicing between formation of E and A complexes, before any RNA cleavage, whereas isoginkgetin prevents the U4/U6/U5 tri-snRNP from joining.

192 nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology
review article NATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
eukaryotic translationHaving cleared splicing and nonsense-mediated mRNA decay, the mature message finally reaches the ribosome. Thanks to small-mol-ecule antibiotics that target the bacterial ribosome, the workings of bacterial translation have been well established. The elucidation of crystal structures of prokaryotic ribosomes with bound antibi-otics yielded unprecedented insights into drug mechanisms and ribosome function at atomic resolution23–26. The fact that only a few molecules equally inhibit bacterial and eukaryotic translation may indicate that the eukaryotic ribosome does differ significantly from its bacterial counterpart. Unfortunately, for the eukaryotic ribos-ome, neither sufficiently specific inhibitors nor high-resolution structures are available. Even though several eukaryote-specific translation inhibitors could be modeled on the archaeal ribosome, a true eukaryotic structure remains desirable27.
Compared to prokaryotes, the mass of small and large ribosomal subunits combined increased from 2.5 to over 3 MDa in eukaryo-tes. The small bacterial subunit (30S) contains one molecule of rRNA (16S) and 21 proteins, whereas its eukaryotic counterpart (40S) typi-cally contains some 30 proteins and a longer (18S) rRNA molecule.The large bacterial subunit is composed of 5S and 23S rRNA and 34 proteins, which expanded to three species of rRNA (5S, 5.8S and 28S) and over 40 ribosomal proteins in the eukaryotic 60S subunit28.
The small subunit is responsible for decoding the mRNA message, whereas the large subunit contains the peptidyl transfer center (PTC).tRNAs form a bridge between the functions of the two ribosomal subunits. Their anticodon loop contacts the small ribosomal subunit, thereby allowing the decoding of the mRNA message, while the 3′ end, which carries the corresponding amino acid, provides the large subunit’s PTC with substrate. The ribosome contains three distinct binding sites for tRNAs, which span both subunits: the A-site accepts the incoming aminoacyl-tRNA and matches the mRNA codon and tRNA anticodon with one another; the P-site holds the peptidyl-tRNA and catalyzes the formation of new peptide bonds; and finally the E-site holds the deacylated tRNA after peptidyl transfer. For proper function the ribosome relies on a multitude of cytosolic factors that regulate translation initiation, elongation and termination.
Because of its fundamental importance in life, protein synthe-sis presents an excellent target for antimicrobial therapy, and many antibiotics block bacterial translation29,30. Most clinical therapeutics directly bind the ribosome, with many different classes of compounds converging on three sites of action. Aminoglycosides and tetracyclines bind into the small subunit’s A-site, interfering with proper decod-ing and tRNA binding, respectively31,32. Chloramphenicol and spar-somycin bind to the large subunit’s P-site and prevent peptide bond formation33,34. Macrolide antibiotics such as erythromycin also bind proximal to the P-site, but instead of interfering with peptidyl transfer they block the passage of the nascent peptide chain out of the ribos-ome35. A few antibiotics instead inhibit accessory factors, such as the thiazolyl family of compounds, which contains molecules that inter-fere with the function of the elongation factors, or pseudomonic acid, which targets isoleucil tRNA synthetase36–38. In summary, bacterial translation antibiotics work either via the A-site and the P-site of the ribosome, or less commonly by interfering with accessory proteins.
Because most prokaryotic translation inhibitors do not act on eukaryotic ribosomes, their utility in investigating eukaryotic trans-lation has been limited. A few compounds do show activity against both prokaryotic and eukaryotic translation, such as the aminogly-cosides G418 and hygromycin B, which find extensive use in the selection and maintenance of transfected cell lines39. Puromycin, which mimics the amino-acyl end of tRNAs, also inhibits both eukaryotic and prokaryotic protein synthesis24,40. It forms a peptide bond with the amino acid on the P-site tRNA, thereby terminat-ing and releasing the nascent peptide chain. Additionally, sordarin and didemnin B act on the eukaryotic translation elongation fac-tors41,42. Sordarin locks the translocase eEF2 in a ribosome-bound
conformation, whereas didemnin B arrests the ternary complex of eEF1A, GTP and the amino-acyl tRNA in the A-site of the ribos-ome, thereby preventing eEF2 from binding43,44. Still, molecules spe-cifically targeting eukaryotic translation have been rare, and their mechanisms were usually not well defined.
the discovery of inhibitors of eukaryotic translationMost known compounds were identified in the early 1970s before the introduction of biochemical tools that would have allowed a more detailed dissection45. Over the past years, several eukaryote-specific inhibitors have been reported that act by new mechanisms. These compounds fall into three categories: (i) molecules that deplete the translational apparatus of substrate, (ii) inhibitors that interfere with the function of translation factors, and (iii) agents that directly target the ribosome.
Depleting the substrates. Reveromycin A (RM-A) presents a good example of this mechanism46. As with many natural products, it was isolated from a strain of the actinobacterium Streptomyces. Though RM-A was originally identified as an inhibitor of EGF mitogenic activity, a genetic screen revealed that RM-A specifically inhibits isoleucyl-tRNA synthetase but does not display activity against the other tRNA charging enzymes46,47. Hence it stalls protein synthesis by slowly depleting the translational apparatus of one important substrate. The detailed mechanism of how RM-A achieves its inhibi-tion remains to be determined, though mutational analysis suggests that the molecule directly binds to its target47.
Interfering with translation factors. Undoubtedly translational maintenance is critical for all cellular function, and its loss affects many biological systems simultaneously. This may explain why many translation inhibitors were originally identified by their secondary effects before the discovery of their true mechanism of action48,49. Various elongation inhibitors were originally identified because they displayed similar effects across different cell lines, including the activation of stress-activated protein kinases50. Their effect appears reproducible enough to allow identification of new elongation inhibitors based on cytotoxicity profiles, as has been done with nagilactone C and phyllantoside51. Unfortunately, their mechanisms of action have not been elucidated in sufficient detail. Though it appears that both compounds interfere with A-site func-tion, it remains unclear whether they exert their effects via the ribosome, the elongation factors or both.
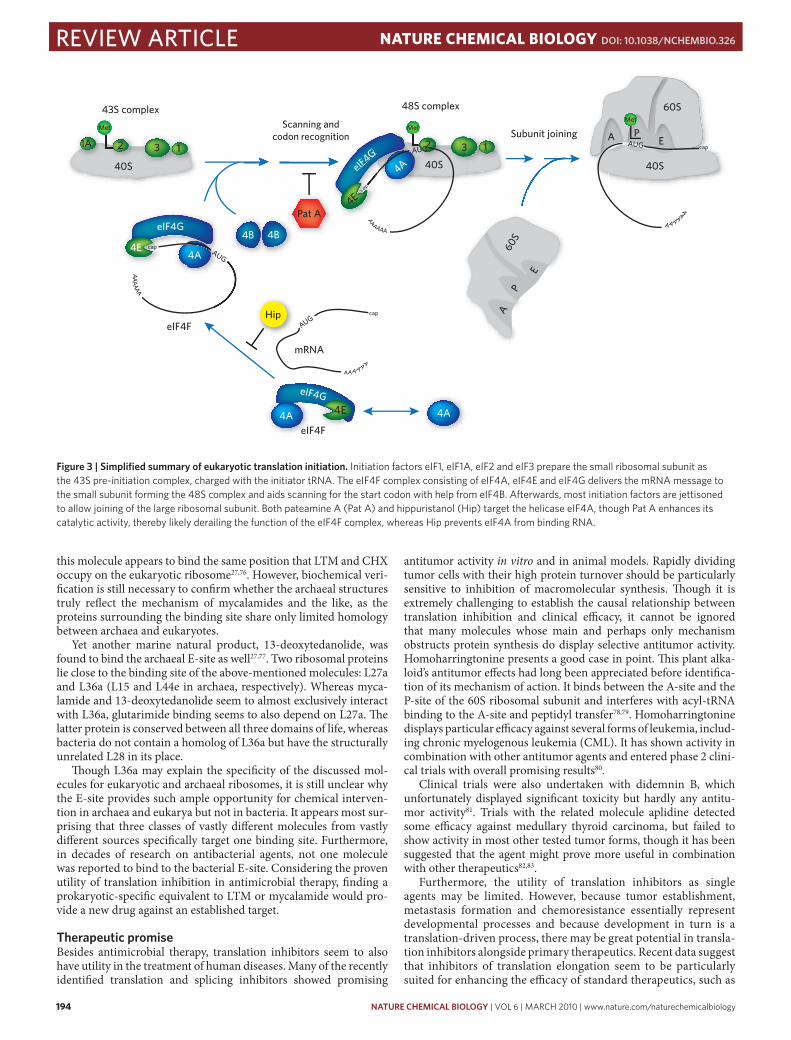
More recently, two molecules attracted attention that did not affect translation elongation but initiation. The initiation of eukaryotic pro-tein synthesis proceeds in a highly regulated fashion and involves at least 12 separate initiation factors. When grossly simplified, this proc-ess can be broken down into three stages: (i) the formation of the 43S pre-initiation complex, in which the initiator tRNA is positioned in the P-site of the small ribosomal subunit; (ii) 48S complex formation, in which the eIF4F complex delivers the mRNA to the 43S complex; and (iii) joining of the small and large subunits (Fig. 3)52.
The eIF4F complex, which consists of the scaffold protein eIF4G, the cap-binding protein eIF4E and the helicase eIF4A, plays a crucial role in coordinating mRNA binding to the small ribosomal subunit and hence in formation of the 48S complex. It is thought to deliver capped transcripts to the ribosome and aid the unwinding of mRNA structures to render the message more accessible. Although it has weak activity by itself, the helicase eIF4A is enhanced by eIF4G and the ancillary factor eIF4B, which itself is not part of the eIF4F complex53. The eIF4A protein has three isoforms. Of these, eIF4AI and eIF4AII are closely related and appear to fulfill functionally redundant roles in translation initiation. eIF4AIII represents a more distantly related isoform that constitutes part of the exon junction complex and has been linked to nonsense-mediated mRNA decay (NMD)54.

nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology 193
review articleNATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
The marine natural product pateamine A (Pat A) was originally purified from the marine sponge Mycale hentscheli, although it remains unclear whether the molecule originates from the sponge itself or from a symbiotic microorganism48,55. Besides this natural source, Pat A is available in synthetic form, which allowed the gen-eration of functional analogs and made structure-activity relationship studies possible56. Pat A was initially identified as a strong immuno-suppressant and cytotoxic agent, but it recently became clear that the molecule disrupts the function of the eIF4F complex48,57,58 by targeting eIF4A and preventing translation initiation from comple-tion. Curiously, it does not inhibit but rather enhances eIF4A’s cata-lytic activities, making it a better helicase and ATPase. The enzyme’s activity depends on the presence of RNA, and it appears that Pat A locks eIF4A in an RNA-bound conformation59. In this state Pat A weakens the interaction between eIF4A and eIF4G, thereby reduc-ing the stability of the eIF4F complex57,59. In contrast, Pat A appears to enhance the interaction between eIF4A and eIF4B, though this might be a result of eIF4A’s increased RNA binding under Pat A, as eIF4B has RNA binding activity by itself. It remains unclear at which stage this derails the translation initiation process, as two independ-ent research groups reported slightly contradictory observations. One team of researchers saw the formation of 48S complexes in vitro, which suggests that the eIF4F complex is halted while bound to an mRNA-ribosome complex57. The other group communicated the loss of 48S complexes and suggested that Pat A stalls initiation before eIF4F binds the ribosome58,59. It remains furthermore puzzling how a small molecule can achieve translational inhibition at nanomolar doses, when it targets a protein present at micromolar levels within the cell. The matter remains hard to resolve as Pat A initiates a cellular stress response in vivo with concomitant formation of stress granules, which in itself decreases translational activity and results in a re-shuffling of cellular components60,61. These aggregates contain among other ingredients mRNA, small ribosomal subunits and translation initiation factors and are believed to sequester mRNAs during times of stress and to prioritize expression of chaperones and other stress-relevant genes62. Because Pat A localizes to stress granules—as do 40S ribosomal subunits, eIF4E and eIF4A, all of which are components of the 48S complex—it seems plausible that Pat A indeed stalls transla-tion at the 48S stage. More importantly, it was initially thought that stress granule formation required the phosphorylation of the initia-tion factor eIF2α. However, Pat A treatment achieved stress granule assembly without eIF2α phosphorylation. It has since been proposed that any process preventing formation of 80S complexes may induce stress granule production, whereas compounds such as cycloheximide that stabilize 80S ribosomes and polysomes appear to protect against stress granule production60,61. Pat A’s effect does not seem entirely lim-ited to translation inhibition, as it also targets eIF4AIII and inhibits nonsense-mediated mRNA decay independently of its role in trans-lation63. Like many translation inhibitors, Pat A displays discernible antitumor activity and proved particularly efficacious against MDA MB435 melanoma xenografts in vivo64.
Pat A is not the only molecule acting on eIF4A. Another marine natural product, hippuristanol, exhibits similar cellular effects, albeit by a different mechanism. In contrast to Pat A, hippuristanol inhib-its eIF4A’s catalytic activities and weakens the enzyme’s RNA binding capability65. Though the Pat A binding site remains unknown, it seems to differ from that of hippuristanol, which is located on the C-terminal domain of eIF4A66. Furthermore, hippuristanol is specific for eIF4AI and eIF4AII, but does not appear to interfere with eIF4AIII function. Nevertheless, both molecules exhibit essentially the same cellular effect, as exemplified by their induction of eIF2α-phosphorylation-independent stress granules61.
Directly targeting the ribosome. Besides targeting translation initiation factors or substrates, binding the ribosome itself pro-vides an effective way of bringing protein synthesis to a halt.
The translation elongation cycle proceeds through three main steps (Fig. 4a). First, the ternary complex of eEF1A, GTP and amino-acyl tRNA binds to the acceptor (A) site of the 80S ribos-ome. If the tRNA anticodon matches the mRNA, eEF1A hydro-lyzes GTP, dissociates from the ribosome and leaves its tRNA cargo behind. In the second step, the ribosome catalyzes peptide bond formation, thereby transferring the nascent peptide chain onto the A-site tRNA, and consequently leaving the P-site tRNA deacylated. Finally the translocase eEF2 aids the movement of the A-site tRNA with the peptide chain into the P-site, as well as the transition of the deacylated tRNA into the E-site. Consequently the A-site becomes available for the next amino-acyl tRNA.
As described above, clinically used antibiotics target the A-site and the P-site of the bacterial ribosome. Surprisingly, the recently characterized eukaryote-specific elongation inhibitors bind a differ-ent ribosomal target altogether. Even more surprisingly, members of three distinct and structurally unrelated groups converge on the same binding pocket.
The most prominent eukaryotic translation inhibitor is undoubtedly cycloheximide (CHX). It has found application in the laboratory for decades and for some time was even used as a pesti-cide. Yet its mechanism remained unclear. For over 30 years CHX was known to interfere with eEF2-mediated translocation, but the details of how it accomplished this feat remained unresolved67. Curiously, CHX seemed to allow one complete cycle of transloca-tion to proceed and only stopped translation once the ribosome had reached the second codon68.
Its mechanism was serendipitously revealed in a study concern-ing a different class of glutarimide molecules69. The cell migration inhibitor migrastatin garnered some interest in the past years for its antitumor effect70–73. Migrastatin itself is a shunt metabolite of Streptomyces platensis74. The bacterium produces an isomeric form called isomigrastatin that contains a 12-membered macrocycle. This compound can undergo a water-mediated ring expansion to form the 14-membered cycle of migrastatin. Initially, the parent molecule isomigrastatin and the structurally related compound lactimidomycin (LTM) were also described as inhibitors of cell migration75. However, both molecules proved to potently inhibit protein synthesis, blocking eEF2-mediated translocation. In con-trast to CHX, LTM arrests protein synthesis during the first cycle of elongation, such that the ribosome remains stuck on the start codon. Furthermore, LTM treatment leads to an accumulation of stalled 80S complexes in vivo, suggesting inhibition of transla-tion at an early stage. This concentration of 80S ribosomes is not observed in CHX-treated cells. Neither CHX nor LTM interferes with eEF1A-mediated tRNA binding or peptidyl transfer. Instead, both molecules bind into the E-site of the large ribosomal sub-unit, close to the position where the 3′ hydroxyl group of the deacylated tRNA would normally bind. Nevertheless, CHX and LTM differ in their mode of action. CHX has only about half the mass of LTM and appears to bind together with the E-site tRNA, while the larger LTM excludes E-site access altogether. This would explain why CHX permits the ribosome to leave the start codon, whereas LTM prevents the first elongation cycle from comple-tion, as CHX acts once an E-site tRNA has bound, whereas LTM occludes E-site access in the first place (Fig. 4b). This means that CHX can inhibit translation at any stage in elongation and would explain its property of stabilizing polysomes. In contrast, LTM preferentially acts during the first elongation cycle, leading to an accumulation of 80S complexes and polysome depletion.
Surprisingly, the glutarimides LTM and CHX are not the only molecules acting on the eukaryotic E-site. Mycalamide A, which is named after the same genus of marine sponge that yielded Pat A, as well as molecules related to the beetle-derived natural product pederin, inhibit translation elongation. From recent structural data of mycalamide A bound to the archaeal large ribosomal subunit,
194 nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology
review article NATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
this molecule appears to bind the same position that LTM and CHX occupy on the eukaryotic ribosome27,76. However, biochemical veri-fication is still necessary to confirm whether the archaeal structures truly reflect the mechanism of mycalamides and the like, as the proteins surrounding the binding site share only limited homology between archaea and eukaryotes.
Yet another marine natural product, 13-deoxytedanolide, was found to bind the archaeal E-site as well27,77. Two ribosomal proteins lie close to the binding site of the above-mentioned molecules: L27a and L36a (L15 and L44e in archaea, respectively). Whereas myca-lamide and 13-deoxytedanolide seem to almost exclusively interact with L36a, glutarimide binding seems to also depend on L27a. The latter protein is conserved between all three domains of life, whereas bacteria do not contain a homolog of L36a but have the structurally unrelated L28 in its place.
Though L36a may explain the specificity of the discussed mol-ecules for eukaryotic and archaeal ribosomes, it is still unclear why the E-site provides such ample opportunity for chemical interven-tion in archaea and eukarya but not in bacteria. It appears most sur-prising that three classes of vastly different molecules from vastly different sources specifically target one binding site. Furthermore, in decades of research on antibacterial agents, not one molecule was reported to bind to the bacterial E-site. Considering the proven utility of translation inhibition in antimicrobial therapy, finding a prokaryotic-specific equivalent to LTM or mycalamide would pro-vide a new drug against an established target.
therapeutic promiseBesides antimicrobial therapy, translation inhibitors seem to also have utility in the treatment of human diseases. Many of the recently identified translation and splicing inhibitors showed promising
antitumor activity in vitro and in animal models. Rapidly dividing tumor cells with their high protein turnover should be particularly sensitive to inhibition of macromolecular synthesis. Though it is extremely challenging to establish the causal relationship between translation inhibition and clinical efficacy, it cannot be ignored that many molecules whose main and perhaps only mechanism obstructs protein synthesis do display selective antitumor activity. Homoharringtonine presents a good case in point. This plant alka-loid’s antitumor effects had long been appreciated before identifica-tion of its mechanism of action. It binds between the A-site and the P-site of the 60S ribosomal subunit and interferes with acyl-tRNA binding to the A-site and peptidyl transfer78,79. Homoharringtonine displays particular efficacy against several forms of leukemia, includ-ing chronic myelogenous leukemia (CML). It has shown activity in combination with other antitumor agents and entered phase 2 clini-cal trials with overall promising results80.
Clinical trials were also undertaken with didemnin B, which unfortunately displayed significant toxicity but hardly any antitu-mor activity81. Trials with the related molecule aplidine detected some efficacy against medullary thyroid carcinoma, but failed to show activity in most other tested tumor forms, though it has been suggested that the agent might prove more useful in combination with other therapeutics82,83.
Furthermore, the utility of translation inhibitors as single agents may be limited. However, because tumor establishment, metastasis formation and chemoresistance essentially represent developmental processes and because development in turn is a translation-driven process, there may be great potential in transla-tion inhibitors alongside primary therapeutics. Recent data suggest that inhibitors of translation elongation seem to be particularly suited for enhancing the efficacy of standard therapeutics, such as
eiF4F
1A
AA
AA
AA
eiF4G AA
G
cap
aUG
AAAAAA
4E
eiF4G
cap c
AAAA AAAA
AAA
4B 4B
4a
43S complex
40S
Met
1a 2 3 1
48S complex
80S complex
Pat a
Scanning andcodon recognition
4e
4A
eiF4G
4e 4a aUG
cap
40S
2 3 1 Subunit joining a P
e
60S
40S
cap aUG
AAAAAA
a P
e
60S
Hip cap
aUG
aaaaaa
mrNa
eiF4F
eiF4G
4a 4e
Met Met
figure 3 | simplified summary of eukaryotic translation initiation. Initiation factors eIF1, eIF1A, eIF2 and eIF3 prepare the small ribosomal subunit as the 43S pre-initiation complex, charged with the initiator tRNA. The eIF4F complex consisting of eIF4A, eIF4E and eIF4G delivers the mRNA message to the small subunit forming the 48S complex and aids scanning for the start codon with help from eIF4B. Afterwards, most initiation factors are jettisoned to allow joining of the large ribosomal subunit. Both pateamine A (Pat A) and hippuristanol (Hip) target the helicase eIF4A, though Pat A enhances its catalytic activity, thereby likely derailing the function of the eIF4F complex, whereas Hip prevents eIF4A from binding RNA.

nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology 195
review articleNATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
doxorubicin, in at least a subset of tumors84. The authors suggested that depletion of short-lived pro-survival proteins could promote induction of apoptosis via the primary therapeutic. Considering that even a fairly weak inhibitor such as CHX displayed synergy with doxorubicin, one could speculate that the results may have been even better if a more potent molecule such as LTM or myca-lamide had been used.
Recently, a lot of attention has been focused on mTOR signaling and the promise of its inhibition by rapamycin in chemotherapy. In particular, the downregulation of cap-dependent translation by rapamycin has garnered interest, even though it only lowers trans-lational activity by some 15–40%85,86. Perhaps it would be even more promising to consider completely abolishing cap-dependent trans-lation with a small dose of Pat A or hippuristanol.
Among the reviewed molecules, reveromycin A is no excep-tion in terms of clinical promise. RM-A contains three carboxylic acid moieties and is negatively charged at neutral pH; hence, it is incapable of penetrating the cell membrane. At a pH of around 4.5, the molecule becomes sufficiently neutral to permeate cells. This pH dependence provides RM-A with unique pharmacological spe-cificity for functional osteoclasts. In order to break down the bone matrix, active osteoclasts generate an acidic environment, which predisposes them for RM-A uptake. In vivo experiments have dem-onstrated RM-A’s efficacy in preventing bone demineralization and formation of bone metastases from lung cancer87,88.
It has been estimated that about 15% of all mutations that cause human genetic diseases are associated with a failure in splicing. In particular, dysregulation of alternative splicing constitutes a
eeF1
2
Codon recognition
GDP1a
Translocation
2
3
2
1rec
eeF1
1n rCCodon
GDP1a
cog
2
1c
1ongnitio
aPe
60S
40SaUG UUGUUG Uaa
aPe
60S
40SaUG UUG UUG Uaa
APe
60S
40SaUG UUG UUG Uaa
aPe
60S
40SaUG UUG UUG
Uaa
Met
MetMet
Phe Met
Phe
1a
0SAAPe
S
aUGaUGaUGGaUGG UUGUUGUUGUUUUGG UUGUUUUUU
Uaa
ttttM
et
Phe
2
60S
0S
o
33lTransl
3333aPe
S
aUGaUGaUGaUGaUGGaUG UUGUUUUGGUUGUUGGUUG UUGUUGUUGUUGUUGGUGUaaUaUaUaUU
ettM
et
Phe
2
aPe60S
40S
1a
2
aUG UUG UUG Uaa
aPe60S
40SaUG UUG UUG Uaa
aPE
60S
40SaUG UUG UUG Uaa
ltM
aPe60S
40SaUG UUG UUG
Uaa
1a
aPe60S
40SaUG UUG UUG
Uaa
cHX
Phe
Met Met
Phe
Phe
Phe
Phe
Phe
Met
Phe
Met
Phe
Met
Phe
a
b
GDP
GDP
GtP
GtP
GtP
1a
GtP
GtP
GtP
GtP
GtP
GDP
GDP
GDP
GDP
Peptidyl transfer
lactimidomycin
cycloheximide
figure 4 | the eukaryotic translation elongation cycle. (a) Elongation factor eEF1A delivers aminoacyl-tRNA into the A-site of the ribosome, which then catalyzes peptide bond formation, thereby transferring the nascent peptide onto the A-site tRNA. The translocase eEF2 pushes the peptidyl-tRNA into the P-site and the deacylated tRNA into the E-site, freeing the A-site for another round of elongation. (b) Model of elongation inhibition by lactimidomycin (LTM) and cycloheximide (CHX). LTM and CHX bind to the same spot in the E-site of the large ribosomal subunit. While LTM occludes tRNA access to the E-site, CHX allows one tRNA to bind before halting further elongation.

196 nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology
review article NATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
common cause for disease. As errors in pre-mRNA splicing are involved in autoimmune disorders, neurodegenerative diseases, cystic fibrosis, growth hormone deficiency, muscular dystrophy, and cancer, the splicing machinery is currently regarded as an important potential target for drug development89–91. The potent antitumor activity of FR901464 and pladienolide prompted the extensive mode-of-action studies that eventually led to the iden-tification of SF3b as the cellular target. For drug development, a challenging but important step is to elucidate the molecular mechanism underlying the anticancer activity of general splicing inhibitors. First, splicing in tumors might be altered, and alter-natively spliced variants may play a role in cancer development92. In support of this hypothesis, some cancer-specific splicing vari-ants of splicing factors have been observed93. This suggests that gene expression of splicing factors may be affected, thus weaken-ing splicing activity in cancer cells. Second, the inhibitor-induced accumulation of unspliced pre-mRNA may allow their leakage into the cytoplasm and translation into proteins. Because essen-tially every intron has in-frame stop codons, these prematurely translated proteins should be truncated. Indeed, FR901464 and spliceostatin A induce the production of a truncated form of the CDK inhibitor p27 (p27*), which retains its inhibitory activity but lacks the Thr187 residue that is subject to the CDK-mediated phosphorylation that is essential for ubiquitination and subse-quent proteasomal degradation. It is likely that accumulation of p27* triggers a circuit of events that arrests the cell cycle by reduc-ing CDK activity in cancer cells, which may lead to stabilization of endogenous p27 and a further decrease of CDK activity. However, the production of an ‘active’ truncated protein is rare, because most first exons do not encode a long polypeptide. This, too, might constitute a reason for the selective toxicity of FR901464 and SSA. Finally, it is also possible that the antitumor activity of splicing inhibitors is associated with other nuclear events such as splicing factor–mediated gene silencing94 and noncoding RNA processing, including intron-encoded miRNA and snoRNA93,95,96.
It should be noted that all molecules discussed herein were iso-lated from natural sources. A quick glance at Figure 1 makes a con-vincing case for the unprecedented structural diversity afforded by naturally occurring compounds. From an academic perspective it seems remarkable how so many structurally unrelated molecules converge on the same targets. Pladienolide and spliceostatins both inhibit the SF3b splicing factor, whereas pateamine A and hippur-istanol target eIF4A, and mycalamide A binds the same ribosomal site as lactimidomycin. On the other hand, structurally closely related molecules may have utterly different biological activity. Though the polyketides pladienolide and isomigrastatin were iso-lated from two closely related strains of the same species, their targets could not differ more. In terms of the translation inhibitor isomigrastatin, even its own isomer migrastatin inhibits a com-pletely different process. In this light it would make sense to spe-cifically screen isomers and congeners of molecules with known activity for new applications. In particular, molecules related to those with established clinical use might make good candidates, as their physicochemical properties and hence bioavailability should be similar to those of the known compound.
ConclusionThe molecules discussed in this article provide four new strategies for interrupting gene expression after transcription: halting post-transciptional events by obstructing splicing, depleting the transla-tion apparatus of substrate, interfering with translation initiation and disabling the ribosome itself. Not only do the reviewed compounds enable insights into enzymatic mechanisms of complex macromo-lecular assemblies, but they can also help dissect the cellular response to the breakdown of key biological processes. For instance, 13-deoxy-tedanolide induces a ribotoxic stress response, whereas pateamine
and hippuristanol bring about the formation of stress granules60,61,97. Hence the use of natural products in probing post-transcriptional events does not cease with the identification of their direct targets, but extends to more detailed mechanistic investigations of their impact on cellular function and their potential as new therapeutics.Published online 12 February 2010; corrected after print 18 March 2010
references1. Staley, J.P. & Woolford, J.L. Jr. Assembly of ribosomes and spliceosomes: complex
ribonucleoprotein machines. Curr. Opin. Cell Biol. 21, 109–118 (2009).2. Wahl, M.C., Will, C.L. & Lührmann, R. The spliceosome: design principles of a
dynamic RNP machine. Cell 136, 701–718 (2009).3. Nakajima, H. et al. New antitumor substances, FR901463, FR901464 and
FR901465. I. Taxonomy, fermentation, isolation, physico-chemical properties and biological activities. J. Antibiot. (Tokyo) 49, 1196–1203 (1996).
4. Nakajima, H. et al. New antitumor substances, FR901463, FR901464 and FR901465. II. Activities against experimental tumors in mice and mechanism of action. J. Antibiot. (Tokyo) 49, 1204–1211 (1996).
5. Nakajima, H., Kim, Y.B., Terano, H., Yoshida, M. & Horinouchi, S. FR901228, a potent antitumor antibiotic, is a novel histone deacetylase inhibitor. Exp. Cell Res. 241, 126–133 (1998).
6. Kaida, D. et al. Spliceostatin A targets SF3b and inhibits both splicing and nuclear retention of pre-mRNA. Nat. Chem. Biol. 3, 576–583 (2007).
7. Horigome, M., Motoyoshi, H., Watanabe, H. & Kitahara, T. A synthesis of FR901464. Tetrahedr. Lett. 42, 8207–8210 (2001).
8. Thompson, C.F., Jamison, T.F. & Jacobsen, E.N. FR901464: total synthesis, proof of structure, and evaluation of synthetic analogues. J. Am. Chem. Soc. 123, 9974–9983 (2001).
9. Albert, B.J., Sivaramakrishnan, A., Naka, T., Czaicki, N.L. & Koide, K. Total syntheses, fragmentation studies, and antitumor/antiproliferative activities of FR901464 and its low picomolar analogue. J. Am. Chem. Soc. 129, 2648–2659 (2007).
10. Albert, B.J. et al. Meayamycin inhibits pre-messenger RNA splicing and exhibits picomolar activity against multidrug-resistant cells. Mol. Cancer Ther. 8, 2308–2318 (2009).
11. Sakai, T. et al. Pladienolides, new substances from culture of Streptomyces platensis Mer-11107. I. Taxonomy, fermentation, isolation and screening. J. Antibiot. (Tokyo) 57, 173–179 (2004).
12. Sakai, T., Asai, N., Okuda, A., Kawamura, N. & Mizui, Y. Pladienolides, new substances from culture of Streptomyces platensis Mer-11107. II. Physico-chemical properties and structure elucidation. J. Antibiot. (Tokyo) 57, 180–187 (2004).
13. Kotake, Y. et al. Splicing factor SF3b as a target of the antitumor natural product pladienolide. Nat. Chem. Biol. 3, 570–575 (2007).
14. O’Brien, K., Matlin, A.J., Lowell, A.M. & Moore, M.J. The biflavonoid isoginkgetin is a general inhibitor of Pre-mRNA splicing. J. Biol. Chem. 283, 33147–33154 (2008).
15. Soret, J. et al. Selective modification of alternative splicing by indole derivatives that target serine-arginine-rich protein splicing factors. Proc. Natl. Acad. Sci. USA 102, 8764–8769 (2005).
16. Stoilov, P., Lin, C.H., Damoiseaux, R., Nikolic, J. & Black, D.L. A high-throughput screening strategy identifies cardiotonic steroids as alternative splicing modulators. Proc. Natl. Acad. Sci. USA 105, 11218–11223 (2008).
17. Hagiwara, M. Alternative splicing: a new drug target of the post-genome era. Biochim. Biophys. Acta 1754, 324–331 (2005).
18. Martinez, E. et al. Human STAGA complex is a chromatin-acetylating transcription coactivator that interacts with pre-mRNA splicing and DNA damage-binding factors in vivo. Mol. Cell. Biol. 21, 6782–6795 (2001).
19. Choudhary, C. et al. Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science 325, 834–840 (2009).
20. Kuhn, A.N., van Santen, M.A., Schwienhorst, A., Urlaub, H. & Luhrmann, R. Stalling of spliceosome assembly at distinct stages by small-molecule inhibitors of protein acetylation and deacetylation. RNA 15, 153–175 (2009).
21. Balasubramanyam, K., Swaminathan, V., Ranganathan, A. & Kundu, T.K. Small molecule modulators of histone acetyltransferase p300. J. Biol. Chem. 278, 19134–19140 (2003).
22. Balasubramanyam, K. et al. Polyisoprenylated benzophenone, garcinol, a natural histone acetyltransferase inhibitor, represses chromatin transcription and alters global gene expression. J. Biol. Chem. 279, 33716–33726 (2004).
23. Ban, N., Nissen, P., Hansen, J., Moore, P.B. & Steitz, T.A. The complete atomic structure of the large ribosomal subunit at 2.4 A resolution. Science 289, 905–920 (2000).
24. Brodersen, D.E. et al. The structural basis for the action of the antibiotics tetracycline, pactamycin, and hygromycin B on the 30S ribosomal subunit. Cell 103, 1143–1154 (2000).
25. Wimberly, B.T. et al. Structure of the 30S ribosomal subunit. Nature 407, 327–339 (2000).
26. Tu, D., Blaha, G., Moore, P.B. & Steitz, T.A. Structures of MLSBK antibiotics bound to mutated large ribosomal subunits provide a structural explanation for resistance. Cell 121, 257–270 (2005).

nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology 197
review articleNATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
27. Schroeder, S.J., Blaha, G., Tirado-Rives, J., Steitz, T.A. & Moore, P.B. The structures of antibiotics bound to the E site region of the 50 S ribosomal subunit of Haloarcula marismortui: 13-deoxytedanolide and girodazole. J. Mol. Biol. 367, 1471–1479 (2007).
28. Lecompte, O., Ripp, R., Thierry, J.C., Moras, D. & Poch, O. Comparative analysis of ribosomal proteins in complete genomes: an example of reductive evolution at the domain scale. Nucleic Acids Res. 30, 5382–5390 (2002).
29. Hermann, T. Drugs targeting the ribosome. Curr. Opin. Struct. Biol. 15, 355–366 (2005).
30. Sutcliffe, J.A. Improving on nature: antibiotics that target the ribosome. Curr. Opin. Microbiol. 8, 534–542 (2005).
31. Schroeder, R., Waldsich, C. & Wank, H. Modulation of RNA function by aminoglycoside antibiotics. EMBO J. 19, 1–9 (2000).
32. Chopra, I. & Roberts, M. Tetracycline antibiotics: mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 65, 232–260 (2001).
33. Nierhaus, D. & Nierhaus, K.H. Identification of the chloramphenicol-binding protein in Escherichia coli ribosomes by partial reconstitution. Proc. Natl. Acad. Sci. USA 70, 2224–2228 (1973).
34. Bashan, A. et al. Structural basis of the ribosomal machinery for peptide bond formation, translocation, and nascent chain progression. Mol. Cell 11, 91–102 (2003).
35. Tenson, T., Lovmar, M. & Ehrenberg, M. The mechanism of action of macrolides, lincosamides and streptogramin B reveals the nascent peptide exit path in the ribosome. J. Mol. Biol. 330, 1005–1014 (2003).
36. Hurdle, J.G., O'Neill, A.J. & Chopra, I. Prospects for aminoacyl-tRNA synthetase inhibitors as new antimicrobial agents. Antimicrob. Agents Chemother. 49, 4821–4833 (2005).
37. Rodnina, M.V. et al. Thiostrepton inhibits the turnover but not the GTPase of elongation factor G on the ribosome. Proc. Natl. Acad. Sci. USA 96, 9586–9590 (1999).
38. Heffron, S.E. & Jurnak, F. Structure of an EF-Tu complex with a thiazolyl peptide antibiotic determined at 2.35 A resolution: atomic basis for GE2270A inhibition of EF-Tu. Biochemistry 39, 37–45 (2000).
39. Eustice, D.C. & Wilhelm, J.M. Mechanisms of action of aminoglycoside antibiotics in eucaryotic protein synthesis. Antimicrob. Agents Chemother. 26, 53–60 (1984).
40. Nathans, D. Puromycin inhibition of protein synthesis: incorporation of puromycin into peptide chains. Proc. Natl. Acad. Sci. USA 51, 585–592 (1964).
41. Justice, M.C. et al. Elongation factor 2 as a novel target for selective inhibition of fungal protein synthesis. J. Biol. Chem. 273, 3148–3151 (1998).
42. Ahuja, D. et al. Inhibition of protein synthesis by didemnin B: how EF-1alpha mediates inhibition of translocation. Biochemistry 39, 4339–4346 (2000).
43. Gomez-Lorenzo, M.G. et al. Three-dimensional cryo-electron microscopy localization of EF2 in the Saccharomyces cerevisiae 80S ribosome at 17.5 A resolution. EMBO J. 19, 2710–2718 (2000).
44. Jorgensen, R. et al. Two crystal structures demonstrate large conformational changes in the eukaryotic ribosomal translocase. Nat. Struct. Biol. 10, 379–385 (2003).
45. Vazquez, D. Inhibitors of protein biosynthesis. Mol. Biol. Biochem. Biophys. 30, i–x, 1–312 (1979).
46. Takahashi, H. et al. Reveromycins, new inhibitors of eukaryotic cell growth. II. Biological activities. J. Antibiot. (Tokyo) 45, 1414–1419 (1992).
47. Miyamoto, Y. et al. Identification of Saccharomyces cerevisiae isoleucyl-tRNA synthetase as a target of the G1-specific inhibitor Reveromycin A. J. Biol. Chem. 277, 28810–28814 (2002).
48. Hood, K.A., West, L.M., Northcote, P.T., Berridge, M.V. & Miller, J.H. Induction of apoptosis by the marine sponge (Mycale) metabolites, mycalamide A and pateamine. Apoptosis 6, 207–219 (2001).
49. Sugawara, K. et al. Lactimidomycin, a new glutarimide group antibiotic. Production, isolation, structure and biological activity. J. Antibiot. (Tokyo) 45, 1433–1441 (1992).
50. Lee, K.H. et al. Inhibition of protein synthesis and activation of stress-activated protein kinases by onnamide A and theopederin B, antitumor marine natural products. Cancer Sci. 96, 357–364 (2005).
51. Chan, J., Khan, S.N., Harvey, I., Merrick, W. & Pelletier, J. Eukaryotic protein synthesis inhibitors identified by comparison of cytotoxicity profiles. RNA 10, 528–543 (2004).
52. Kapp, L.D. & Lorsch, J.R. The molecular mechanics of eukaryotic translation. Annu. Rev. Biochem. 73, 657–704 (2004).
53. Rogers, G.W. Jr., Richter, N.J., Lima, W.F. & Merrick, W.C. Modulation of the helicase activity of eIF4A by eIF4B, eIF4H, and eIF4F. J. Biol. Chem. 276, 30914–30922 (2001).
54. Palacios, I.M., Gatfield, D., St Johnston, D. & Izaurralde, E. An eIF4AIII-containing complex required for mRNA localization and nonsense-mediated mRNA decay. Nature 427, 753–757 (2004).
55. Northcote, P.T., Blunt, J.W. & Munro, M.H.G. Pateamine: a potent cytotoxin from the New Zealand marine sponge Mycale sp. Tetrahedr. Lett. 32, 6411–6414 (1991).
56. Rzasa, R.M., Shea, H.A. & Romo, D. Total synthesis of the novel, immunosuppressive agent (-)-pateamine A from Mycale sp. employing a β-lactam-based macrocyclization. J. Am. Chem. Soc. 120, 591–592 (1998).
57. Low, W.K. et al. Inhibition of eukaryotic translation initiation by the marine natural product pateamine A. Mol. Cell 20, 709–722 (2005).
58. Bordeleau, M.E. et al. Stimulation of mammalian translation initiation factor eIF4A activity by a small molecule inhibitor of eukaryotic translation. Proc. Natl. Acad. Sci. USA 102, 10460–10465 (2005).
59. Bordeleau, M.E. et al. RNA-mediated sequestration of the RNA helicase eIF4A by pateamine A inhibits translation initiation. Chem. Biol. 13, 1287–1295 (2006).
60. Dang, Y. et al. Eukaryotic initiation factor 2α-independent pathway of stress granule induction by the natural product pateamine A. J. Biol. Chem. 281, 32870–32878 (2006).
61. Mazroui, R. et al. Inhibition of ribosome recruitment induces stress granule formation independently of eukaryotic initiation factor 2α phosphorylation. Mol. Biol. Cell 17, 4212–4219 (2006).
62. Kedersha, N. & Anderson, P. Stress granules: sites of mRNA triage that regulate mRNA stability and translatability. Biochem. Soc. Trans. 30, 963–969 (2002).
63. Dang, Y. et al. Inhibition of nonsense-mediated mRNA decay by the natural product pateamine A through eukaryotic initiation factor 4AIII. J. Biol. Chem. 284, 23613–23621 (2009).
64. Kuznetsov, G. et al. Potent in vitro and in vivo anticancer activities of des-methyl, des-amino pateamine A, a synthetic analogue of marine natural product pateamine A. Mol. Cancer Ther. 8, 1250–1260 (2009).
65. Bordeleau, M.E. et al. Functional characterization of IRESes by an inhibitor of the RNA helicase eIF4A. Nat. Chem. Biol. 2, 213–220 (2006).
66. Lindqvist, L. et al. Selective pharmacological targeting of a DEAD box RNA helicase. PLoS One 3, e1583 (2008).
67. Obrig, T.G., Culp, W.J., McKeehan, W.L. & Hardesty, B. The mechanism by which cycloheximide and related glutarimide antibiotics inhibit peptide synthesis on reticulocyte ribosomes. J. Biol. Chem. 246, 174–181 (1971).
68. Pestova, T.V. & Hellen, C.U. Translation elongation after assembly of ribosomes on the Cricket paralysis virus internal ribosomal entry site without initiation factors or initiator tRNA. Genes Dev. 17, 181–186 (2003).
69. Schneider-Poetsch, T. et al. Inhibition of translation elongation by cycloheximide and lactimidomycin. Nat. Chem. Biol. 6, 209–217 (2010).
70. Nakae, K. et al. Migrastatin, a new inhibitor of tumor cell migration from Streptomyces sp. MK929–43F1. Taxonomy, fermentation, isolation and biological activities. J. Antibiot. (Tokyo) 53, 1130–1136 (2000).
71. Woo, E.J. et al. Migrastatin and a new compound, isomigrastatin, from Streptomyces platensis. J. Antibiot. (Tokyo) 55, 141–146 (2002).
72. Takemoto, Y., Tashiro, E. & Imoto, M. Suppression of multidrug resistance by migrastatin. J. Antibiot. (Tokyo) 59, 435–438 (2006).
73. Shan, D. et al. Synthetic analogues of migrastatin that inhibit mammary tumor metastasis in mice. Proc. Natl. Acad. Sci. USA 102, 3772–3776 (2005).
74. Ju, J., Lim, S.K., Jiang, H. & Shen, B. Migrastatin and dorrigocins are shunt metabolites of iso-migrastatin. J. Am. Chem. Soc. 127, 1622–1623 (2005).
75. Ju, J. et al. Lactimidomycin, iso-migrastatin and related glutarimide-containing 12-membered macrolides are extremely potent inhibitors of cell migration. J. Am. Chem. Soc. 131, 1370–1371 (2009).
76. Gurel, G., Blaha, G., Steitz, T.A. & Moore, P.B. The structures of triacetyloleandomycin and mycalamide A bound to the large ribosomal subunit of Haloarcula marismortui. Antimicrob. Agents Chemother. 53, 5010–5014 (2009).
77. Nishimura, S. et al. 13-Deoxytedanolide, a marine sponge-derived antitumor macrolide, binds to the 60S large ribosomal subunit. Bioorg. Med. Chem. 13, 449–454 (2005).
78. Fresno, M., Jimenez, A. & Vazquez, D. Inhibition of translation in eukaryotic systems by harringtonine. Eur. J. Biochem. 72, 323–330 (1977).
79. Gurel, G., Blaha, G., Moore, P.B. & Steitz, T.A. U2504 determines the species specificity of the A-site cleft antibiotics: the structures of tiamulin, homoharringtonine, and bruceantin bound to the ribosome. J. Mol. Biol. 389, 146–156 (2009).
80. Quintas-Cardama, A., Kantarjian, H. & Cortes, J. Homoharringtonine, omacetaxine mepesuccinate, and chronic myeloid leukemia circa 2009. Cancer 115, 5382–5393 (2009).
81. Kucuk, O. et al. Phase II trail of didemnin B in previously treated non-Hodgkin’s lymphoma: an Eastern Cooperative Oncology Group (ECOG) study. Am. J. Clin. Oncol. 23, 273–277 (2000).
82. Le Tourneau, C. et al. Reports of clinical benefit of plitidepsin (Aplidine), a new marine-derived anticancer agent, in patients with advanced medullary thyroid carcinoma. Am. J. Clin. Oncol. (2009).
83. Ocio, E.M., Mateos, M.V., Maiso, P., Pandiella, A. & San-Miguel, J.F. New drugs in multiple myeloma: mechanisms of action and phase I/II clinical findings. Lancet Oncol. 9, 1157–1165 (2008).
84. Robert, F. et al. Altering chemosensitivity by modulating translation elongation. PLoS One 4, e5428 (2009).

198 nature CHeMICaL BIoLogy | VOL 6 | MARCH 2010 | www.nature.com/naturechemicalbiology
review article NATuRE CHEMICAL BIoLogy doI: 10.1038/nCHeMBIo.326
85. Beretta, L., Gingras, A.C., Svitkin, Y.V., Hall, M.N. & Sonenberg, N. Rapamycin blocks the phosphorylation of 4E–BP1 and inhibits cap-dependent initiation of translation. EMBO J. 15, 658–664 (1996).
86. Averous, J. & Proud, C.G. When translation meets transformation: the mTOR story. Oncogene 25, 6423–6435 (2006).
87. Woo, J.T. et al. Reveromycin A, an agent for osteoporosis, inhibits bone resorption by inducing apoptosis specifically in osteoclasts. Proc. Natl. Acad. Sci. USA 103, 4729–4734 (2006).
88. Hiraoka, K. et al. Inhibition of bone and muscle metastases of lung cancer cells by a decrease in the number of monocytes/macrophages. Cancer Sci. 99, 1595–1602 (2008).
89. Faustino, N.A. & Cooper, T.A. Pre-mRNA splicing and human disease. Genes Dev. 17, 419–437 (2003).
90. Ng, B. et al. Increased noncanonical splicing of autoantigen transcripts provides the structural basis for expression of untolerized epitopes. J. Allergy Clin. Immunol. 114, 1463–1470 (2004).
91. Wang, G.S. & Cooper, T.A. Splicing in disease: disruption of the splicing code and the decoding machinery. Nat. Rev. Genet. 8, 749–761 (2007).
92. Blencowe, B.J. Alternative splicing: new insights from global analyses. Cell 126, 37–47 (2006).
93. Kim, Y.K. & Kim, V.N. Processing of intronic microRNAs. EMBO J. 26, 775–783 (2007).
94. Bayne, E.H. et al. Splicing factors facilitate RNAi-directed silencing in fission yeast. Science 322, 602–606 (2008).
95. Hirose, T., Shu, M.D. & Steitz, J.A. Splicing-dependent and -independent modes of assembly for intron-encoded box C/D snoRNPs in mammalian cells. Mol. Cell 12, 113–123 (2003).
96. Kataoka, N., Fujita, M. & Ohno, M. Functional association of the Microprocessor complex with the spliceosome. Mol. Cell. Biol. 29, 3243–3254 (2009).
97. Lee, K.H. et al. Induction of a ribotoxic stress response that stimulates stress-activated protein kinases by 13-deoxytedanolide, an antitumor marine macrolide. Biosci. Biotechnol. Biochem. 70, 161–171 (2006).
acknowledgmentsWe thank J.O. Liu of the Johns Hopkins University School of Medicine for his support and for making an unpublished manuscript available to us.
Competing interests statementThe authors declare no competing financial interests.
additional informationReprints and permissions information is available online at http://npg.nature.com/reprintsandpermissions/. Correspondence should be addressed to M.Y.

correctionsNature chemical biology
Garbled messages and corrupted translationsTilman Schneider-Poetsch, Takeo Usui, Daisuke Kaida & Minoru Yoshida
Nat. Chem. Biol. 6, 189–198 (2010); published online 12 February 2010; corrected after print 18 March 2010
In the version of this article initially published, the page numbers in reference 69 were incorrect. The error has been corrected in the HTML and PDF versions of the article.
erratum





























