Embed Size (px)
Citation preview

SPECIAL ARTICLE
Retinal amino acid neurochemistry in health and disease
Clin Exp Optom 2013; 96: 310–332 DOI:10.1111/cxo.12015
Michael Kalloniatis*†§� BSc (Optom) MSc(Optom) PhDChee Seang Loh� BOptomMonica L Acosta� BSc MSc PhDGuido Tomisich¶ BScYuan Zhu† BSc MSc PhDLisa Nivison-Smith† BSc (Hons) PhDErica L Fletcher§ BSc (Optom) MSc(Optom) PhDJacqueline Chua� BOptom PhDDaniel Sun� BOptom MOptom PhDNiru Arunthavasothy� BSc* Centre for Eye Health and † School of Optometryand Vision Science, University of New South Wales,Sydney, New South Wales, Australia§ Department of Anatomy and Neuroscience, TheUniversity of Melbourne, Parkville, Victoria, Australia� Department of Optometry and Vision Sciences, NewZealand National Eye Centre, University of Auckland,Auckland, New Zealand¶ Department of Optometry and Vision Science, TheUniversity of Melbourne, Parkville, Victoria, Australia.E-mail: [email protected]
Advances in basic retinal anatomy, genetics, biochemical pathways and neurochemistryhave not only provided a better understanding of retinal function but have also allowedus to link basic science to retinal disease. The link with disease allowed measures to bedeveloped that now provide an opportunity to intervene and slow down or even restoresight in previously ‘untreatable’ retinal diseases. One of the critical advances has been theunderstanding of the retinal amino acid neurotransmitters, related amino acids, theirmetabolites and functional receptors. This review provides an overview of amino acidlocalisation in the retina and examples of how retinal anatomy and amino acid neurochem-istry directly links to understanding retinal disease. Also, the implications of retinal remod-elling involving amino acid (glutamate) receptors are outlined in this review and insightsare presented on how understanding of detrimental and beneficial retinal remodellingwill provide better outcomes for patients using strategies for the preservation or restorationof vision. An internet-based database of retinal images of amino acid labelling patternsand other amino acid-related images in health and disease is located at http://www.aminoacidimmunoreactivity.com.
Submitted: 5 May 2012Revised: 1 July 2012Accepted for publication: 17 July 2012
Key words: amino acids, GABA, GAD, glutamate, glutamine, glutamine synthetase, glycine, immunoreactivity, remodelling, retina
The aim of this review is to provide an over-view of basic retinal neurochemistry, focus-ing on the amino acid neurotransmittersand metabolites (glutamate, g-amino butyricacid [GABA], glycine, aspartate, glutamine)and osmoregulators (taurine), synthesisingand degradation enzymes, (glutamic aciddecarboxylase [GAD], glutamine synthetase[GS]), glutamate receptor subunits andfunctional activation. The extensive litera-ture on this topic means that it is not possibleto do justice to all published manuscripts.The focus of this review is to highlight theconservation of core amino acid neuro-chemistry across different vertebrate retinaeand provide the foundations to understandretinal disease. This approach allows a holis-tic appreciation and an understanding ofthe vertebrate retina in health and disease,
based on core knowledge of retinal neuro-chemistry, anatomical and functional data.
The internet-based archive located athttp://www.aminoacidimmunoreactivity.com, provides an opportunity to undertakegeneral or specific searches on a varietyof retinal topics. Images can be identified,often with the corresponding figure legends,by undertaking a search using the followingparameters:1. author2. species classification or name3. amino acid4. retinal cell type or retinal layer5. retinal condition (retinal detachment;
retinal dystrophy; metabolic insult)6. labelling pattern (different amino acids,
manufacturing enzymes or cation probe-agmatine or AGB)
7. image analysis compilations (red-green-blue images; theme maps; mono-to-trivariate amino acid distributionpatterns).
Supplementary Figures 1 to 5 outlinethe phylogenetic classification for differentspecies available in this review and theinternet-based archive.
Basic neurochemistry andanatomy (a clinical perspective)The vertebrate retina has a common struc-ture that displays species-specific specialisa-tions. Photoreceptors capture light andthrough a series of interactions within theretina, the fundamental encoding of thespatio-temporal and chromatic informationis achieved and transmitted to other partsof the visual processing centres. The basic
C L I N I C A L A N D E X P E R I M E N T A L
OPTOMETRY
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
310 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

retinal architecture involves photorecep-tors-bipolar cells-ganglion cells forming the‘through’ retinal pathway associated with‘lateral’ elements (horizontal cells, ama-crine cells and interplexiform cells) interact-ing at the two retinal synaptic layers.1–3
The three spectral classes of cone pho-toreceptors in the primate retina synapseonto two distinct sets of bipolar cells: thedepolarising bipolar cells (ON bipolar cells)and the hyperpolarising bipolar cells (OFFbipolar cells4,5). The two bipolar cell typessubsequently synapse with different types ofganglion cells to form two major streams ofvisual information flow: ON and OFF gan-glion cells (Figure 1). Although the termi-nology of ‘ON’ and ‘OFF’ is commonly usedto describe bipolar cell and amacrine cellresponses, it was originally reserved for gan-
glion cell responses. Turning the light ‘ON’or ‘OFF’ led to an increase in spike firingrate in the corresponding type of ganglioncells.6
The encoding of visual informationoccurs through synaptic interactions at eachof the two synaptic layers: the outer plexi-form and inner plexiform layers (Figure 1).It is at these synaptic layers that the lateralelements (horizontal cells in the outerplexiform layer and amacrine cells in theinner plexiform layer), interact with thethrough pathway providing the feedback toencode spatial, chromatic and temporalinformation.1,7–10
The mammalian scotopic system has onlyone type of bipolar cell: a depolarising orON bipolar cell. Rod bipolar cells synapseon a unique type of amacrine cell (the AII
amacrine cell) that subsequently conveys thescotopic information to the ON and OFFcone system11 (Figure 1). Thus, AII ama-crine cells subserve a similar function tobipolar cells as part of the through pathway.There are no unique rod ganglion cells andthus, the mammalian rod circuitry ‘piggybacks’ onto cone circuitry to transmitscotopic information.11
Clinical manifestations provide evidenceof a single bipolar cell type in the scotopicsystem as well as its dependence upon conecircuitry to put out its signal. In the paraneo-plastic syndrome, melanoma associatedretinopathy (MAR), photoreceptors areintact yet nyctalopia results due to systemicantibodies destroying rod and cone ONbipolar cells by targeting the unique cationchannel (transient receptor potential cationchannel subfamily M member 1 or TRPM1)located on these cells.12–14 Although theOFF cone pathway remains functional, thepatients are afflicted with night-blindnessas the rod photoreceptors lose their soleconnection with the inner retina.
When ganglion cells are damaged in glau-coma or non-glaucomatous optic nervedamage, dark adaptation is affected, result-ing in greater threshold elevation for the rodpathway, as well as elevated cone thresh-olds.15 In the clinical cone adaptation recov-ery test (photostress test), patients withglaucoma displayed a significantly longerrecovery time (approximately 70 ms) com-pared to controls (approximately 42 ms).16
Thus, any disease where ganglion cell dys-function occurs will also result in rod andcone visual dysfunction.
Non-mammalian retinae display similarcellular physiological responses to those ofmammalian retinae, although one uniquefeature is that bipolar cells carry mixedrod/cone signals and synapse directly ontoganglion cells.1,3,17 Subsequent to retinalprocessing, ganglion cells project to a multi-tude of locations within the central nervoussystem.
It is estimated that there are approxi-mately 60 different cell types in the mamma-lian retina.18,19 The classical studies usingGolgi impregnation by Ramón y Cajal20 pro-vided the first retinal morphological andmorphometric information, which has beenexpanded with the application of specificcell markers, where whole populations ofneurons are labelled.21–23 New analyticalmethods of cell imaging and visualisationusing self-reporting methods (for example,green fluorescent protein), have further
Figure 1. Schematic representation of the mammalian retina showing the two majorpathways operating under photopic ‘cones’ or scotopic ‘rods’ conditions. Cone photore-ceptors are presynaptic to ‘ON’ or ‘OFF’ cone bipolar cells (BCs) and horizontal cellbodies (HC-CB). The cone-driven bipolar cells synapse onto ganglion cells (GC) andamacrine cells (AC) in the inner plexiform layer and form the through pathway (colour-coded green). The lateral elements, horizontal cells, amacrine cells and interplexiformcells (IPC) are colour-coded red. All elements in the rod pathway are shaded grey. Rodphotoreceptors synapse onto horizontal cell axon terminals (HC-AT) and one type of ONbipolar cell. The rod BC synapses with the AII amacrine cells in the inner plexiform layer.The AII amacrine cell sends the signal via heterocellular gap junctions (gj) to ON conebipolar cells and synapses directly onto OFF ganglion cells. The Müller cell extendsthroughout the whole retina with endfeet in the inner retina and microvillous processesin the outer retina.GCL: ganglion cell layer, INL: inner nuclear layer, IPL: inner plexiform layer, NFL: nervefiber layer, OPL: outer plexiform layer, RL: receptor layer, RPE: retinal pigmentepithelium. Figure modified from Kalloniatis and Tomisich.31
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 311

transformed our understanding of retinalstructure/function relationships.24,25
The early anatomical work has beencombined with cell marking techniquesand physiology to identify adult cellulararchitecture and retinal developmental
characteristics.21,23,26–30 Despite the diversemorphology of individual cells in the verte-brate retina, there is an overall conservationof core neurochemical architecture, particu-larly in the amino acid neurochemistry. Thisis not surprising, given that amino acid
manufacturing and degradation pathwaysare preserved across different vertebratespecies. The study of amino acids formsa core foundation in our knowledge baseto complement that of retinal structure,metabolism and neurotransmission.31–40
The perplexing outcome—asuccessful operation but poorrecovery of visionIn rhematogenous retinal detachment, theretina is deprived of one of its major bloodsupplies, the choriocapillaris, as well as dis-ruption of other metabolic and cellular func-tions (Figures 2A, B and C). Various surgicaltechniques are available to treat retinaldetachment with a high rate of success.Thelen and colleagues41 reported a surgicalsuccess rate of over 83 per cent for the com-bined group (macula on and macula off).Pastor and colleagues42 achieved an surgicalreattachment success rate of approximately95 per cent in a study of 546 patients. In thisstudy, the majority of patients with reat-tached retinas at three months (n = 517) hada poor visual outcome: 21.5 per cent worsethan 6/30, 36 per cent between 6/15 and6/30 and only 42.7 per cent had visual acui-ties of 6/12 or better.42 The best predictor offinal visual outcome appears to be present-ing visual acuity before surgery.43 Thus,despite a high rate of surgical success,visual outcomes appear to be poor, evenwhen surgery was performed very soon afterdetachment.43
Although initial retinal hypoxia anddeath of some photoreceptors occur beforesurgery, the sequelae of anatomical andfunctional detrimental retinal remodell-ing are a possible reason for the poorvisual outcome following surgical reattach-ment.44–48 The concept of retinal remodel-ling explains not only the poor outcomeswhen some vision preservation/restora-tion therapies are attempted in patientswith inherited retinal dystrophies but isalso applicable to other forms of retinaldegeneration.49
The need to understandretinal remodellingRetinal neuronal cell death and remodel-ling occur in a variety of ocular condi-tions, leading to irreversible visual loss inconditions such as age-related maculardegeneration, optic nerve disease includingglaucoma, and retinopathies including dia-betic retinopathy.47,50–65 Some conditions
Figure 2. Clinical examples of metabolic insult caused by retinal detachment (A, B andC) and retinal changes in retinitis pigmentosa (D and E). A 54-year-old patient with visualacuity of 6/6 in both eyes and minor complaints of recent onset vitreous floaters in the lefteye (A and B). A red-free wide-field retinal image shows a horseshoe tear inferiorly in theleft eye (circle in A). An optical coherent tomographic image through the lesion (circled)shows it to be a full-thickness retinal tear and retinal detachment (asterisk in B). Thesequelae of such retinal tears are shown in panel C with a large inferior, infero-nasal andinfero-temporal retinal detachment and a shallow superior retinal detachment in the lefteye (VA 6/95) in a 79-year-old patient (C). A tear that may have caused the detachment inthis patient is circled in the superior temporal region (C). A 48-year-old patient with visualacuities 6/7.5 in both eyes displays restricted visual field and fundus images consistentwith retinitis pigmentosa (D). The typical pigmented bone spicule-like changes areevident in this wide-field fundus image. The optical coherent tomographic image acrossthe fovea and optic nerve head shows the loss of the photoreceptor layer (loss ofphotoreceptor integrity line is identified by the large arrows in E). The three small arrowsidentify a retinal area with disrupted outer retina and the two arrowheads identify anepiretinal membrane.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
312 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

result in long-term visual affliction (suchas inherited retinal dystrophies, includingretinitis pigmentosa and cone-rod dystro-phies such as Stargardt’s disease [Figures 2Dand E]).66–69
Currently, there are some exciting devel-opments in treatment options for inheritedretinal dystrophies. Although associatedwith controversy,70 nutritional supplementsmay slow the progression of some forms ofinherited retinal dystrophies (that is, vita-min A for retinitis pigmentosa). Throughbasic scientific research, we are also aware ofthe potential dangers of ingesting vitamin Ain other retinal conditions, that is, in Star-gardt’s disease and related cone-rod dystro-phies and in age-related maculopathy.71 Theuse of gene therapy,72–78 stem cells79–82 andnutritional supplements or pharmacologicalagents74,83–86 is restoring or maintaining sightin both humans and non-human models ofretinal disease.
Retinal remodelling may restore visualfunction (beneficial remodelling). In ani-mal models, beneficial anatomical remodel-ling occurs with photoreceptor migration,particularly when low-energy laser photoco-agulation is used.87,88 The photoreceptormigration to fill in the damaged areaappears to be dependent upon retinalpigment epithelial viability, whereby theirmigration into damaged areas allows subse-quent photoreceptor filling in and presum-ably a reduced area of visual dysfunction. Inhuman studies, the use of small, short (20milliseconds) pulse laser lead to restorationof anatomical photoreceptor characteristicsand overall sustained healing response oversix months.89
The future is promising for many retinalconditions that were thought to be ‘untreat-able’ a decade or so ago; however, under-standing detrimental retinal remodelling iscrucial. This is highlighted in the disparitybetween high surgical success for retinalreattachment versus poor visual functionpost-surgery, most likely due to detrimentalretinal remodelling.41–43,46–48
Some intervention measures assume thatneurons and neuronal circuitry ‘down-stream’ of the disease locus are functionallynormal. Yet, a wealth of evidence indicatesthat neurons and glia undergo anatomicaland functional remodelling secondary toretinal disease that may impede interventionsuccess.47–49,56,57,60,90–97 The recent successfultransplantation of photoreceptors in theadult retina82 occurred in an animal modelof congenital stationary night blindness,
where photoreceptors are intact but dys-functional. In dystrophies where photore-ceptors degenerate, the anatomical and glialremodelling, including the formation of aglial seal,97 would make it difficult for trans-planted cells to make functional contacts.Conversely, it is also critical to understandbeneficial remodelling87–89 and use it tomaximise visual outcomes for patients.
Changes associated with anatomicalremodelling in the retina can be identifiedusing a range of cell markers or using clinicalimaging modalities. This knowledge is usefulto characterise cell death or cell anatomicalchanges during disease. Functional (neuro-chemical) remodelling may alter neuro-transmitter content, recycling and release,neurotransmitter receptor disposition orneurotransmitter receptor functionality.Functional remodelling may result in ana-tomically intact neural tissue displaying poorfunction, thereby creating a quandary inunderstanding structure/function relation-ships. If retinal circuitry is altered, any newsignal from implanted cells/devices may be‘jumbled’ and thus higher-order neuronsare unable to decode, or worse, the neuronsmay not be able to send a signal, that is, thecollision circuit theory of Marc and col-leagues.56 Functional remodelling is a rela-tively new discovery and occurs in the earlystages of the disease process. By identifyingthe time course, the specific neurotransmit-ter receptors affected and modifying thetime over which the retina is pliable, it will bepossible to improve the success of interven-tion measures. For example, the suitabletime of intervention for procedures such ascell transplantation or retinal prostheticimplants will result in the maximum possibil-ity of success.
THE VISUAL PROCESSAND NEUROTRANSMISSION
Localisation of glutamate,GABA and glycine
GLUTAMATEThe mammalian retina is a complex array ofglial and neuronal cells that convert andencode light energy into electrical impulsesto begin the visual process. Glutamate is themajor excitatory neurotransmitter in theretina (Figures 3A, 4 and 5).37,98–102 Figure 3Adisplays a schematic of mammalian retinalstructure (for simplicity only the cone path-way is shown) and the concurrent gluta-
mate labelling pattern (rabbit retina). Thephotoreceptor layer is typically glutamateimmunoreactive as are bipolar and ganglioncells. Glutamate is a metabolite and precur-sor for GABA103,104 and therefore, is alsolocalised in GABAergic and other amacrinecells. Müller cells are virtually devoid ofglutamate (arrows in Figure 3A) and in thehigh cellular density of the central primateretina, the light band of immunoreactivitywithin the middle of the inner nuclear layerreflects the location of Müller cell somata(Figure 4D).
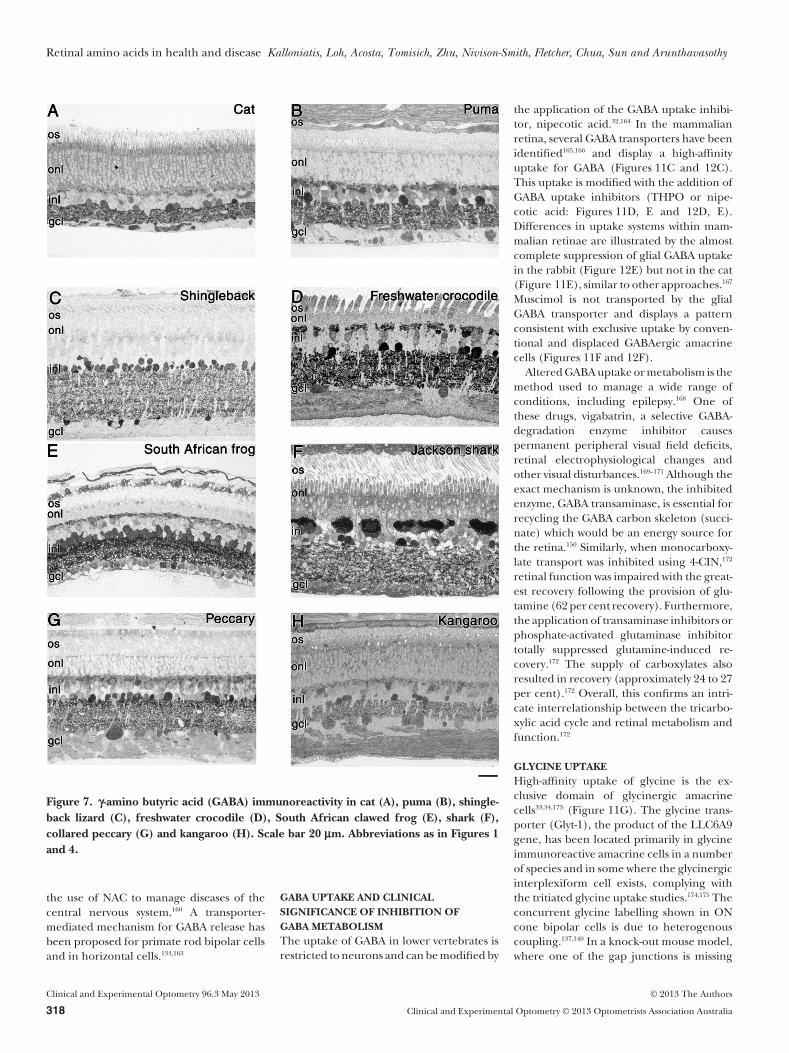
Examples of amino acid labelling from avariety of species (Figures 4, 5 and 7–11)illustrate the conservation of amino acidlabelling patterns but also reveal some inter-esting species differences. Figure 4 is theprimate retina (monkey—Macaca fascicula-ris) and includes a Nissl stained central area(Figure 4A), and a sample away from thefovea (the low ganglion cell density but largenerve fibre layer indicates that it is close tothe optic nerve head, Figure 4B), followedby immunostaining for a range of aminoacids in the central retina (Figures 4C–H).Other species presented include the domes-tic cat (Felis catus), cougar (Puma concolour),Shingleback or stumpy tail lizard (Tiliquarugosa), freshwater crocodile (Crocodylus john-stoni), South African clawed frog (Xenopuslaevis), Port Jackson shark (Heterodontusportusjacksoni), collared peccary (Pecaritajacu) and eastern grey kangaroo (Macropusgiganteus).
The cat visual system has excellent binocu-lar vision and the physiology and neuro-anatomy are well characterised.105–108 Beinga predator, the cougar has keen vision,high visual sensitivity in poor light, a rod-dominated retina, large pupil, tapetum,reduced visual acuity during day light, gooddepth perception and a broad field ofview.109 The blue-tongue lizards have welldeveloped colour vision, reaction to preyand predator and also contain oil dropletsin their photoreceptors.110 Crocodiles arethought to possess good eyesight that sensesmotion, have a wide sight range, good colourvision above water but poor vision underwater, as they use their nictitating mem-brane.111 The Xenopus visual system is wellstudied with colour discrimination possible,good motion detection and good vision atlow light levels.112,113 Sharks have low packingdensity, large diameter rod photoreceptorsand presumed low resolution.114,115 The col-lared peccary is thought to have poor visionin daylight and poor movement percep-
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 313

tion.116 The eastern grey kangaroo feedsduring both daylight and nighttime, dif-ferent from other kangaroos that havedaytime eating habits.117 Although there isstrong evidence that glutamate is used byphotoreceptors,35–37 considerable variationsin glutamate immunoreactivity have beenreported in primate tissue.118–121 Post-mortem changes, alterations due to ex-perimental manipulations or even lightadaptation changes are some of the reasonsfor these differences within a single species.In monkey retina, Figure 4D shows strongglutamate immunoreactivity in most cellswith the ganglion cells and their axons dis-playing the highest level of labelling. Pho-toreceptors are also strongly labelled, as aretheir axons (Henle’s fibres, the thick bandbelow the outer nuclear layer in Figure 4D).In contrast, Figure 5 illustrates considerablediversity in glutamate labelling of photore-ceptors, from being strong in the cat, shin-gleback, crocodile, frog, shark and kangaroo(Figures 5A, C–F and H) to weak in the col-lared peccary and puma (Figures 5B and G),that likely reflects real differences in gluta-mate content.
Bipolar cells often display the highest levelof immunoreactivity while ganglion cellsmay be extremely large and highly glutamateimmunoreactive (for example, Figure 5G).When evident, the nerve fibre layer isalso strongly glutamate immunoreactive(Figures 4D, 5A, B, D, F and H). Lateralelements most likely use glutamate as a pre-cursor and display distinct morphology(large layered horizontal cells in the shark,Figure 5F) or smaller highly immunoreac-tive somata in the crocodile (Figure 5D).Müller cells display very low levels of gluta-mate immunoreactivity under physiologicalconditions.
Both ionotropic and metabotropic gluta-mate receptors are dispersed throughoutthe retina.122 In the outer retinal synapticlayer (outer plexiform layer; OPL), ONbipolar cells display functional metabo-tropic glutamate receptor 6 (mGluR6123),while OFF bipolar cells and horizontal cellsdisplay functional kainate or 2-amino-3-(5-methyl-3-oxo-1,2- oxazol-4-yl) propanoicacid (AMPA) receptors.124 Within the innerretina, kainate/AMPA and n-methyl-D-aspartate (NMDA) receptors are located onamacrine and ganglion cells.4,22,31,122,125–128
Distribution of glutamate receptors129 can beobserved in Figure 6, where the small focalpuncta identify the location of receptorsubunits. Receptors are composed of numer-
Figure 3. Schematic of the cone pathway associated and the amino acid immunocyto-chemistry for glutamate (A) or g-amino butyric acid (GABA) and glycine (B). Forsimplicity only the through cells are shown (photoreceptors, bipolar cells and ganglioncells). Glutamate-labelled bipolar cells (B), amacrine cells (A) and ganglion cells (G) areshown with Müller cell processes (white arrows) displaying very low levels of glutamateimmunoreactivity. The schematic in B includes the lateral elements that display GABAor glycine immunoreactivity. Abbreviations as in Figure 1.Visualisation of amino acid content in the retinal tissue shown in this figure andsubsequent figures was achieved using post-embedding silver-intensified immunocyto-chemistry.94,95,99,100,207 Retinae were fixed in 2.5 per cent glutaraldehyde/one per centparaformaldehyde in phosphate buffer, dehydrated, resin impregnated and polymer-ised. Glutaraldehyde tissue fixation is critical to preserve amino acid levels. There wasvirtually no delay in fixation for retinae collected as part of laboratory experiments andminimal post-mortem delay for samples collected from our other sources. Resin blockscontaining retinal samples were serially sectioned using an ultramicrotome, typicallysectioned at approximately 500 nm. The immunocytochemical and immunoglobulin G(IgG) production procedures for the primary antibodies were as described previ-ously.99,190 IgGs were raised in rabbits and confirmed to be selective for proteinglutaraldehyde-linked agmatine (AGB) or amino acid antigens using dot immu-noassays.99,190,197 The primary rabbit polyclonal antibodies used for post-embeddingimmunocytochemistry were diluted in one per cent goat serum in phosphate bufferedsaline at the following working concentrations: AGB (1:800), glutamate (1:5,000), GABA(1:4,500), glycine (1:4,500), aspartate (1:200), glutamine (1:500) and taurine (1:1,000).The primary antibodies were detected using goat anti-rabbit IgGs adsorbed to 1 nm goldparticles and visualised with silver intensification.100
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
314 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

ous subunits that combine to impart thephysiological characteristics for variousneurotransmitter receptors.
GABAThe two major inhibitory neurotransmitters,GABA and glycine, are located in the innerretina (amacrine cells and interplexiformcells, although in some species GABA isalso found in horizontal cells: Figure 3B).The volatility of GABA localisation is wellrecognised130 with examples of the primateretina showing very low GABA labelling(Figure 4C) in contrast to other stronglylabelled examples.121,131 In the rabbit retina,GABA is predominantly localised in conven-tional and occasionally displaced amacrinecells and their processes within the innerplexiform layer (Figure 3B). GABA horizon-tal cell labelling is evident in five of the eightspecies illustrated in Figure 7, with the shin-gleback lizard devoid of horizontal cellGABA immunoreactivity. Amacrine cells arethe dominant cell class displaying GABAimmunoreactivity in all vertebrates.32,40,132
Stronger GABA labelling in centralbipolar cells allowed the classification of aunique neurochemical class reflective of rodbipolar cells.121,133 Recent evidence suggeststhat these bipolar cells may tonically releaseGABA through reversal of a GABA trans-porter.134,135 The GABA immunoreactivitywithin ganglion cells and their axons,136
evident particularly in the crocodile andkangaroo, is most likely from leakagethrough gap junctions.137–139 A solitary GABAimmunoreactive ganglion cell is evident inthe central ganglion cell layer of the primate(Figure 4C).
GLYCINEGlycine is predominantly localised in ama-crine cells and ON bipolar cells (Figures 3B,4F and 8). Localisation in bipolar cellsand ganglion cells again is likely due to het-erologous gap junctions.137,140 Also, a sub-population of interplexiform cells in theteleost fish, amphibian and avian retinaeare glycine immunoreactive.33,34,100,141 Thecorresponding GABA or glycine receptorshave been identified on bipolar cell termi-nals, amacrine cells and ganglion celldendrites.31,126,142,143
AMINO ACIDS AS PRECURSORS ANDMETABOLITES (GLUTAMINEAND ASPARTATE)Glutamate is the immediate precursor toGABA as well as a cellular metabolite103,104
Figure 4. Nissl staining (A and B) and various amino acid-labelling patterns of themonkey retina (C to H). A. Central Nissl-stained primate retina showing the densephotoreceptors and thick ganglion cell layer. B. A Nissl-stained section away from thecentral foveal region showing the distinct anatomy of rod and cone photoreceptors anda thick nerve fibre layer indicating that the sample is close to the optic nerve head.C. g-amino butyric acid (GABA), D. Glutamate, E. Glutamine, F. Glycine, G. Taurine andH- aspartate immunoreactivity. The ‘gcl’ label identifies both the ganglion cell and nervefibre layers and in some panels the label is located within the nerve fibre layer. Scale bar20 mm. OS: outer segment, ONL: outer nuclear layer. Other abbreviations as in Figure 1.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 315

and therefore, is localised throughout differ-ent cell classes in the retina. Although notneurotransmitters, glutamine and aspartateare amino acids critical for both neurotrans-mission and metabolism. Glutamine is part
of the glutamate-glutamine cycle and aspar-tate is involved in transamination reactionswith glutamate31 (Figures 4E, H, 9 and 10).In both cat and rat retinas, the carbon skel-eton of glutamine has been localised as
GABA, glutamine, glutamate and to a lesserextent, aspartate,144 highlighting the key rolethis amino acid plays in amino acid neuro-transmitter homeostasis.
Glutamine and aspartate are localisedthroughout the retina with ganglion cells inparticular displaying intense labelling145
(Figures 4E, H, 9 and 10). Glutamine is alsolocalised within Müller cells and this isparticularly evident in the primate, collaredpeccary and shark retinae. The Müller cellprocesses are visible in the middle of theinner nuclear layer and strong endfeetlabelling (Figures 4E, 9G and F).
Anaplerotic biochemical reactions arerequired to replenish the carbon skeleton tothe tri-carboxylic acid (TCA) cycle, whenamino acid production consumes TCA cycleintermediaries.103,104,146 Inhibition of mono-carboxylates resulted in dramatic reductionin retinal function (assessed using the elec-troretinogram) and also dramatic changesin amino acid immunoreactivity.147 Virtuallyall retinal cell types showed reduced gluta-mate and glutamine and amacrine cellsshowed reduced GABA and glycine and aconcurrent increase of aspartate.147 Aspar-tate levels are also influenced by physiologi-cal changes in retinal metabolic activity andcan be dramatically altered in more extremeinsult, when the retina is incubated underanoxic conditions.148
Preventing glutamine productionthrough enzymatic inhibition of glutaminesynthetase resulted in markedly alteredamino acid levels and retinal dysfunc-tion.101,149,150 The provision of glutaminecompletely restored function secondaryto glutamine synthetase enzyme inhibi-tion,149,150 whereas the provision of other sub-strates such as a-ketoglutarate, succinate,lactate or pyruvate only partially restoredfunction (16 to 20 per cent).150 A key findingby Bui and colleagues150 was the dramaticchange in precursor amino acids in otherretinal neurons consistent with a role of pro-vision of metabolic substrates by Müller cells.Also, several studies have shown that theretina uses amino acids or keto-acids tomaintain retinal function.31,131,151
Transport (uptake) pathways forglutamate, glutamine, GABAand glycine
GLUTAMATE AND GLUTAMINE UPTAKENeurotransmitter systems require mecha-nisms to rapidly deactivate active chemicals.Furthermore, compounds required for cel-
Figure 5. Glutamate immunoreactivity in cat (A), puma (B), shingleback lizard (C),freshwater crocodile (D), South African clawed frog (E), shark (F), collared peccary (G)and kangaroo (H). Scale bar 20 mm. Abbreviations as in Figures 1 and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
316 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

lular function must be selectively trans-ported into or out of a cell through amechanism that allows the maintenance ofcellular integrity. One of the ways thesecriteria are achieved is through the expres-sion of unique transport proteins thatundertake either low-affinity or high-affinitytransport of compounds into or out of acell.31,33,34,38,152–155 Figures 11 (cat) and 12(rabbit) illustrate the high-affinity uptake ofvarious amino acids (autoradiographic dis-tribution of tritiated amino acid uptake) andhow the uptake can be modified using avariety of analogues, such as the GABA ana-logue (muscimol) with GABA transpor-ter inhibitors, 4,5,6,7-tetrahydroisoxazolo[4,5-c] pyridin-3-ol (THPO) or nipecoticacid.
Both neurons and glia have uptakesystems for glutamate and glutamine illus-trating a diffuse labelling pattern in both catand rabbit retinae (Figures 11A, B and 12A,B). Glutamate uptake is not restricted to
neurons for use as a neurotransmitter butalso involves glial cells and other neuronswhich use glutamate as a precursor ormetabolite.31,156,157 In the rat retina, Rauenand colleagues158 established that glial high-affinity uptake (Km = 2.1 � 0.4 mmol/l)is the predominant deactivation pathwayfor glutamate. Glutamate uptake by photo-receptors through high-affinity uptakeappears to be the predominant means ofdeactivating the neurotransmitter in theouter plexiform layer.22,159
There is a group of systemic drugs thatincludes, N-acetylcysteine (NAC: producescystine), d-cycloserine (binds to glycinebinding site of the NMDA receptor),memantine (NMDA receptor antagonist)and topiramate (an AMPA receptor antago-nist) that acts on the glutamatergic system,particularly useful in managing depressivedisorders, drug addictions and potentially inthe treatment of dementia.160,161 With theexception of d-cycloserine, the other neuro-
chemicals affect multiple neurotransmittersystems in addition to the glutamate systemand therefore, the direct link to clinical con-ditions is not established.160 Laboratory worksupports the role of glutamate neurotrans-mission in drug reward, reinforcement andrelapse.160 There was considerable interest inmemantine as a neuroprotective agent inthe treatment of glaucoma; however, thereis little support that the use of neuro-protective agents, including memantine,preserves retinal ganglion cells and thus pre-serves vision in patients with open angleglaucoma.162
Our concept of neurotransmitter releaseand deactivation is changing with the discov-ery of a glutamate-cystine transporter (Xc-)in photoreceptor terminals and evidencethat supports the role of this transporter intonic release of glutamate.153 This discoveryof a function for a glutamate-cystine trans-porter in glutamate neurotransmission, mayexplain the pharmacological mechanism for
Figure 6. Confocal fluorescence photomicrographs of vertical frozen sections through mouse retinas that are immunolabelled forglutamate receptors (GluR). The 2-amino-3-(5-methyl-3-oxo-1,2-oxazol-4-yl) propanoic acid (AMPA) receptor subunits, GluR1 andGluR4, are present in both the outer plexiform layer (OPL) and inner plexiform layer (IPL). The GluR1-immunoreactive puncta in theIPL have a stratified distribution and the GluR4-immunoreactive puncta appear as a broad band in the centre of the IPL. The GluR1subunit is also found extra-synaptically in some amacrine cells and ganglion cells.129 The NR1C2' antibody, identifying the splice variantsof the NR1 subunit of the n-methyl-D-aspartate (NMDA) receptor, appears to be clustered in synaptic hotspots in the IPL and also labelscone pedicles in the OPL.128 Isolated retinal samples were fixed in four per cent paraformaldehyde, 0.01 per cent gluteraldehyde for30 minutes. Samples were cryoprotected in 30 per cent sucrose solution overnight before being sectioned vertically at 16 mm thicknesson a cryostat and immunocytochemistry was performed as described previously.208 Briefly, retinal sections were incubated in a blockingbuffer before primary antibodies were applied overnight. Rabbit polyclonal antibodies of GluR1 (Millipore; AB1504), GluR4(Millipore; AB1508) and NR1C2' (Millipore; AB5050P) were diluted in phosphate buffer containing three per cent goat serum, one percent bovine serum albumin and 0.5 per cent Triton X-100 at concentrations of 1:1,000, 1:200 and 1:500, respectively. To visualise thelabelling, sections were incubated in the dark with secondary antibody conjugated to Alexa TM488 for two hours. Specific dyes of4�,6-diamino-2-phenylindole (DAPI) or bisBenzimide were used to label nuclei (blue). Scale bar = 20 mm.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 317
the use of NAC to manage diseases of thecentral nervous system.160 A transporter-mediated mechanism for GABA release hasbeen proposed for primate rod bipolar cellsand in horizontal cells.134,163
GABA UPTAKE AND CLINICALSIGNIFICANCE OF INHIBITION OFGABA METABOLISMThe uptake of GABA in lower vertebrates isrestricted to neurons and can be modified by
the application of the GABA uptake inhibi-tor, nipecotic acid.32,164 In the mammalianretina, several GABA transporters have beenidentified165,166 and display a high-affinityuptake for GABA (Figures 11C and 12C).This uptake is modified with the addition ofGABA uptake inhibitors (THPO or nipe-cotic acid: Figures 11D, E and 12D, E).Differences in uptake systems within mam-malian retinae are illustrated by the almostcomplete suppression of glial GABA uptakein the rabbit (Figure 12E) but not in the cat(Figure 11E), similar to other approaches.167
Muscimol is not transported by the glialGABA transporter and displays a patternconsistent with exclusive uptake by conven-tional and displaced GABAergic amacrinecells (Figures 11F and 12F).
Altered GABA uptake or metabolism is themethod used to manage a wide range ofconditions, including epilepsy.168 One ofthese drugs, vigabatrin, a selective GABA-degradation enzyme inhibitor causespermanent peripheral visual field deficits,retinal electrophysiological changes andother visual disturbances.169–171 Although theexact mechanism is unknown, the inhibitedenzyme, GABA transaminase, is essential forrecycling the GABA carbon skeleton (succi-nate) which would be an energy source forthe retina.150 Similarly, when monocarboxy-late transport was inhibited using 4-CIN,172
retinal function was impaired with the great-est recovery following the provision of glu-tamine (62 per cent recovery). Furthermore,the application of transaminase inhibitors orphosphate-activated glutaminase inhibitortotally suppressed glutamine-induced re-covery.172 The supply of carboxylates alsoresulted in recovery (approximately 24 to 27per cent).172 Overall, this confirms an intri-cate interrelationship between the tricarbo-xylic acid cycle and retinal metabolism andfunction.172
GLYCINE UPTAKEHigh-affinity uptake of glycine is the ex-clusive domain of glycinergic amacrinecells33,34,173 (Figure 11G). The glycine trans-porter (Glyt-1), the product of the LLC6A9gene, has been located primarily in glycineimmunoreactive amacrine cells in a numberof species and in some where the glycinergicinterplexiform cell exists, complying withthe tritiated glycine uptake studies.174,175 Theconcurrent glycine labelling shown in ONcone bipolar cells is due to heterogenouscoupling.137,140 In a knock-out mouse model,where one of the gap junctions is missing
Figure 7. g-amino butyric acid (GABA) immunoreactivity in cat (A), puma (B), shingle-back lizard (C), freshwater crocodile (D), South African clawed frog (E), shark (F),collared peccary (G) and kangaroo (H). Scale bar 20 mm. Abbreviations as in Figures 1and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
318 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

(connexin45 knock out), scotopic functionwas impaired and glycine immunoreactivitywas eliminated in ON cone bipolar cells.176
These results176 confirm the importance of
coupling in retinal function and that coneON bipolar cells receive their glycinethrough heterologous coupling of retinalneurons.
Alterations in amino acid uptakeMüller cell function is altered in retinal dis-ease.177 Altered amino acid uptake or degra-dation has been reported in glia during earlyretinal degeneration,102 diabetes178 or sec-ondary to metabolic insult.179–182 Figure 13illustrates two examples of insult that alteredMüller cell function, ultimately leading tomodification of retinal function. Glutaminesynthetase is a useful marker of retinalMüller cells and is localised throughout thesoma (Figure 13A). Secondary to severeischaemia, the retinal architecture is severelydisrupted (Figure 13B), glutamine syn-thetase immunoreactivity is reduced andglutamate content redistributed.183,184 Inaddition, once Müller cells become reactivein retinal dystrophy and thus presumablyalter their function, they display increasedimmunoreactivity to glial fibrillary acid pro-tein183 (GFAP, Figures 13C and D).
Taurine localisationTaurine is a sulphated amino acid, the func-tion of which in the retina is not fully under-stood. Taurine is involved in cell volumeregulation, neuromodulation, antioxidantdefense, protein stabilisation, stress res-ponses and via formation of taurine-chloramine in immunomodulation.185 A lackof taurine leads to severe photoreceptordegeneration.185,186 Taurine is also involvedin rod photoreceptor production duringdevelopment through interactions with aglycine receptor and a GABA receptor.187
Taurine levels are altered in retinal detach-ment,58 retinal degeneration102 or secondaryto prolonged anaesthesia in mammals.31
Figures 4G and 14 show the distribution oftaurine in the nine species. Almost all cellsare labelled, particularly photoreceptorsand retinal Müller cells. A striking speciesdifference exists, where the South Africanclawed frog displays little taurine labellingwithin the inner retina and Müller cells(Figure 14E).
PATTERN RECOGNITION—A POWERFUL IMAGEANALYSIS TECHNIQUE
Although most current neuroanatomicalapproaches provide analytical tools, wherethe integrity and density characteristics ofspecific sub-populations of neurons can beinvestigated, the sheer volume and complex-ity of retinal cell architecture does not allowa uniform analysis to occur of all retinal cells.
Figure 8. Glycine immunoreactivity in cat (A), puma (B), shingleback lizard (C), fresh-water crocodile (D), South African clawed frog (E), shark (F), collared peccary (G) andkangaroo (H). Scale bar 20 mm. Abbreviations as in Figures 1 and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 319

Pattern recognition applied to serial-sectionimmunocytochemistry employs computa-tional methods identical to those applied inthe analysis of gene arrays.188,189 The volumeand complexity of information generated bygene arrays necessitates the use of clusteringalgorithms such as k-means and isodataapproaches followed by statistical testingsuch as transformed divergence. Similarly,the reason for applying this approach to theretina was due to the volume of data andcomplexity of cell marking patterns.190
Pattern recognition using small moleculesas cell markers is the only method that pro-vides a complete structural map of the verte-brate retina or brain and can classify allcells.58,62,94,95,121,138,190–193 Pattern recognitionusing serial-section silver-intensified immu-nocytochemistry aims to classify data setsusing rigid statistical paradigms and sepa-rates cells into distinct populations sharing acommon signature (that is, a common label-ling pattern). Virtually all retinal cells canbe classified into taurine-rich, glutamate-rich, GABA-rich or glycine-rich cell classes,groups of cells or ‘theme classes’.
The classification uses signal correlationsfrom n-dimensional space (‘n’ dependsupon the number of small molecule labelsthat are used, for example, the number ofamino acids). Theme classes reflect theresult of the classification approach identi-fying pixels that have the same labellingpattern (for example, metabolic signaturedue to similar amino acid labelling pat-terns). Theme classes arise exclusivelyfrom their labelling intensity signals anddo not depend on morphological identi-ties. The anatomical location allows thecell identity to be determined throughcorrelation of cellular position and mor-phometrics (the ‘truth point’ principleoutlined by Sun, Vingrys and Kalloniatis95).A ‘truth point’ is defined from satelliteimaging analysis defined as the observa-tion, measurement, and collection ofinformation about the actual conditionson the ground to determine the relation-ship between remote sensing data and theobserved objects.194
Pattern recognition only classifies pixelsbased upon their unique signal correlationswith no spatial information used in the clas-sification. Once the classification is made,the addition of spatial information usingknown retinal anatomy (applying the ana-tomical truth point principle), allows forretinal cell types to be identified at theindividual neuron level, ‘horizontal cells’
Figure 9. Glutamine immunoreactivity in cat (A), puma (B), shingleback lizard (C),freshwater crocodile (D), South African clawed frog (E), shark (F), collared peccary (G)and kangaroo (H). Scale bar 20 mm. Abbreviations as in Figures 1 and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
320 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

or groups of cells, ‘amacrine cells’. Forexample, the spatial patterns and neuro-chemical labelling of sub-populations ofamacrine cells195,196 allow for the identifica-tion of these cells during functionalmapping.95,197 Marc197 identified the highsensitivity of the cholinergic amacrine cellsto kainate activation due to their knowninner plexiform stratification pattern andsomata location. Sun, Vingrys and Kalloni-atis95 also used anatomical truth points tolink theme maps and extract out the aminoacid signature patterns for the displacedcholinergic amacrine cell population.
Once signal correlations are determinedand theme maps completed, statistical analy-sis is required. The statistical analysis tests forseparability of the different clusters and isnot based upon differences in the ‘mean’labelling intensity but rather in the overlapof labelled distributions. The approachis analogous to d’ and associated valuesderived from ‘signal detection theory’. Oncethe clusters are identified into their differentclasses, the degree of overlap in their prob-ability distributions (that is, separationindex) is tested using transformed diver-gence DT. The criterion routinely employedis DT = 1.9 that translates to less then one percent error in errors of classification. Thus,the probability of correct classification is 99per cent. This criterion in ‘signal detectiontheory’ is analogous to having an overlap ofless than one per cent between the ‘noise’distribution to the ‘signal plus noise’ distri-bution in ‘signal detection theory’. As such,one can predict that out of 100 cells thathave the same classification, 99 will be cor-rectly classified, with only one cell beingmisclassified.
The number of theme classes that havebeen separable has varied among speciesand the number of reporting molecules thathave been used. Marc, Murry and Basinger190
reported a total of nine theme classes using asix amino acid set in the goldfish and 14 cellclasses using a seven amino acid set in the catretina.191 Kalloniatis, Marc and Murry121
found 16 separable classes in the monkeyretina with a six amino acid set.
The theme class in the adult mouse retinaderived from the analysis of amino acidlabelling is shown in Figure 15. Photorecep-tors are separated into various theme classesreflective of outer segment, inner segmentand nuclear layer. The different pseudocol-ours indicate that the amino acid labellingpattern is distinctly different as determinedby the separability index (transform diver-
Figure 10. Aspartate immunoreactivity in cat (A), puma (B), shingleback lizard (C),freshwater crocodile (D), South African clawed frog (E), shark (F), collared peccary (G)and kangaroo (H). Scale bar 20 mm. Abbreviations as in Figures 1 and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 321

gence). The multitude of colours within theinner nuclear and ganglion cell layers illus-trates the unique amino acid labellingpattern of inner retinal neurons allowing fordiscrete signature patterns to be identified.Müller cell endfeet form a distinct signature(black) within the inner-most retinal layers
(Müller cell somata within the inner nuclearlayer were not classified). Using thisapproach, new insights have been providedin retinal disease.62,94,95,97,138,193
In both ischaemia-reperfusion93–95 andretinopathy of prematurity,198 the results ofpattern recognition allowed the investiga-
tion of specific cell classes (using cellmarkers) or retinal function (using elec-troretinography). Downie and colleagues198
were able to show a specific loss of AIIamacrine cells, indicating the potentialloss of rod function in this disease. Sunand colleagues93–95 were able to identify a
Figure 11. Autoradiographs showinguptake in the cat retina of (A) 3H-glutamate(Glu), (B) 3H-glutamine (Gln), (C)3H-GABA, (D) 3H-GABA with 5 mmol/l4,5,6,7-tetrahydroisoxazolo [4,5-c] pyridin-3-ol (THPO) and (E) 3H-GABA with1 mmol/l nipecotic acid. A g-amino butyricacid (GABA) analogue, tritiated muscimolis shown in panel F and tritiated glycine inpanel G. A. Müller cells show intense label-ling in their somata (arrow with M) andendfeet (arrowhead). Note the high graindensity in the outer nuclear layer confirm-ing high affinity uptake by photoreceptors.B. Glutamine is a precursor to glutamateand a metabolite and displays uptakethroughout the retina. C. Müller cells showintense labelling in their somata (arrowwith M) and endfeet (arrowhead), but notethe extremely low labelling in other cells inthe inner and outer nuclear layer. D. THPOand E. nipecotic acid are GABA agonistsand are taken up by the high-affinity GABAtransporter on Müller cells (M and arrow-head identifies endfeet) allowing 3H-GABAto be taken up by GABAergic neurons (Ac).F. Muscimol is a GABA analogue that is nottaken up by glia leading to selective label-ling by conventional (Ac) or displaced(dAc) GABAergic amacrine cells. G.Glycine uptake is exclusively located in gly-cinergic amacrine cells (Ac) with lightbipolar cell labelling (Bc) also present.Scale bar 20 mm. Abbreviations as inFigures 1 and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
322 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

dramatic loss of cone bipolar cells and dys-functional neurotransmission between rodphotoreceptors and rod bipolar cells.Further, it was only through the unequivocalidentification of the amino acid signaturepatterns that the different stages in anatomi-cal remodelling revealed by theme mapsduring retinal degeneration were character-ised.97,193 The concept of a glial seal hasevolved because of these studies,97,193 allow-ing us to understand why in moderate-to-advanced stages of retinal dystrophy somevision restoration strategies will encountermajor obstacles.
Theme maps are important in that theynot only provide a statistically rigid classifica-tion for the unequivocal identification ofcell types but can be used to extract theamino acid distribution patterns of indi-vidual classes of neurons. The extraction ofunivariate, bivariate or trivariate amino aciddistributions allows for the individual aminoacid distribution within classes of cells to bedetermined but also establishes differencesdue to the disease process.62,94,95,121,190,193
FUNCTIONAL MAPPING USING ANORGANIC CATION (AGMATINE—AGB)
The activity of glutamate receptors canbe probed by using an organic cation,agmatine199 (AGB). Agmatine is a divalentpolyamine (de-carboxylated arginine) thatpenetrates acetylcholine-activated ion chan-nels in sympathetic neurons and frog end-plate, as well as indicating physiologicalactivity of invertebrate visual and vestibularneurons.199–201 Agmatine or related mol-ecules (guanidinium compounds) entercells via ionic channels in a Nernstianfashion200,201 with AGB tracking allowingfor excitatory mapping of neuronal tis-sue.93–95,197,202–209 The amount of AGB enter-ing a cell is proportional to the channelpermeability.197,202,203,207 Agmatine concentra-tion is a key driving force with AGB reportingcurrent flow over time.202
Agmatine entry into different sub-populations of neurons will be distinctly dif-ferent due to two factors.128,197,202,203 First,basal AGB entry is dependent upon theoverall ion channel activity in the ‘resting’state and may be further modified if L-2-amino-4-phosphono butyric acid (L-APB orL-AP4), a specific metabotropic glutamatereceptor agonist, is used to modify AGBentry into ON bipolar cells.93,128,197,202 Thesecond reason relates to the diverse dis-position of glutamate receptors within the
Figure 12. Autoradiographs showing uptake in the rabbit retina of (A) 3H-glutamate(Glu), (B) 3H-glutamine (Gln), (C) 3H-GABA (D) 3H-GABA with 5 mmol/l 4,5,6,7-tetrahydroisoxazolo [4,5-c] pyridin-3-ol (THPO) and (E) 3H-GABA with 1 mmol/l nipe-cotic acid. A GABA analogue, tritiated muscimol is shown in panel F. A. Müller cells showintense labelling in their somata (arrow with M) and endfeet (arrowhead). Note the highgrain density in the outer nuclear layer confirming high affinity uptake by photoreceptors.B. Glutamine is a precursor to glutamate and a metabolite and displays uptake throughoutthe retina but is more localised in glia compared to the cat retina (Figure 11B). C. Müllercells show intense labelling in their somata (arrow with M) and endfeet traversing thenerve fibre layer (arrowhead). Note the extremely low labelling in the outer nuclear layerbut clear labelling of GABAergic amacrine cells (Ac) in the inner nuclear layer. D. THPOand E. nipecotic acid are g-amino butyric acid (GABA) agonists and are taken up by thehigh-affinity GABA transporter on Müller cells (M and arrowheads identifies endfeet)allowing 3H-GABA to be taken up by GABAergic neurons (Ac). F. Muscimol is a GABAanalogue that is not taken up by glia leading to selective labelling by conventional (Ac) ordisplaced (dAc) GABAergic amacrine cells. Scale bar 20 mm. Abbreviations as in Figures 1and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 323

vertebrate retina. The activation of thesereceptors using AMPA, kainate and NMDAproduces distinctly different AGB labellingpatterns that can be combined with aminoacid labelling or cell markers to either clas-sify the retina or provide cell identificationof activated neurons (Figures 16 and1747,48,197,203,208,209).
The core principle of AGB labelling isillustrated in Figure 16. Basal AGB entry isshown in Figure 16A, where AGB labellingis diffusely located throughout the retinaindicative of basal AGB entry. Cone photore-ceptors are evident as more darkly stained
nuclei in the inner segment region. Inanimal models of retinal dystrophy or sec-ondary to light damage, AGB labelling ofphotoreceptors appears to precede photore-ceptor loss.210–212 Functional mapping of theretina is reflected by altered AGB labellingwhen glutamate analogues are added inthe incubation medium. The inclusion ofNMDA in the incubation medium resultedin labelling of processes within the innerplexiform layer and somata in the amacrinecell and ganglion cell layer (Figure 16B).
The inclusion of AGB labelling assists increating greater diversity in the results pro-
vided by image analysis. Pattern recognitionanalysis of overlapping amino acid and func-tional profile using the cation channelprobe (AGB) has provided 28 separable‘theme classes’ within the rat retina.95 Theinclusion of AGB labelling (functional pro-filing) almost doubled the number of sepa-rable theme classes. The 14 retinal ganglioncell classes reported by Marc and Jones138
were only possible when AMPA-activatedAGB signal was added to the metabolicprofiling.
The introduction of techniques to com-bine cell markers with functional tracking208
has allowed the identification of selectiveneurochemically identified neurons thatdisplay functional glutamate receptors innormal retinal development and in animalmodels of retinal disease.47,48,93–96,208,209,213–215
Using a glutamate agonist (kainate), it ispossible to combine labelling with a cellmarker for the cholinergic amacrine cells toconfirm that this population of neuronscontain kainate receptors (Figures 17A–C).Similarly, the synthesising enzyme forGABA, glutamic acid decarboxylase (GAD),can be co-localised with activated neurons todisplay localisation patterns (Figures 17D–F). Using this approach, the development offunctional glutamate receptors in the retinahas been characterised209 and this neuro-chemical development has been associatedwith changes in retinal cell spatial localisa-tion during development.213
INSIGHTS INTO RETINAL DISEASEREVEALED BY BASIC ANATOMICAL,NEUROCHEMICAL ANDFUNCTIONAL STUDIES
Remodelling triggered aftermetabolic insultMetabolic insult triggers a range of cellulardamaging mechanisms including excessiveglutamate release (excitotoxicity).54,216,217
Altered vascularisation of the vertebrateretina not only results in anomalous vesselformation and disposition but dramaticchanges in amino acid distribution pat-terns.62,64,198 In more severe metabolic insult,ischaemia-reperfusion deprives the vasculartissues of oxygen, metabolites and removalof waste products.217 There is evidence thatanatomical changes may begin within hoursof ischaemic insult, particularly ischaemiadue to elevated intraocular pressure (IOP)that reaches 50 mmHg or more.218–220 Gan-glion cell survival after transient intervals of
Figure 13. Confocal photomicrographs of glutamine synthetase (GS) immunoreactivityunder normal (A) or ischaemic conditions (B) in the rat retina and of glial fibrillary acidprotein (GFAP) immunoreactivity in retina from a normal rat (C) or a transgenic rat (D)model of retinitis pigmentosa (P23H). Glutamine synthetase immunoreactivity, as anestablished marker for Müller cells, was localised within the Müller cell somata (arrow-head in A), endfeet (arrow in A) and processes spanning throughout the entire retina. Inrats subjected to acute metabolic insult (ischaemia), intense labelling of GS was observedin discrete clusters with the feature of Müller cell shape being lost. GFAP-positive stainingwas mainly localised in the astrocytes in the ganglion cell layer and nerve fibre layer in thenormal retina. In the degenerative retina, GFAP immunoexpression was detected inMüller cell processes (arrows in D). Ocular ischaemia was induced by elevating intra-ocular pressure through a cannula inserted into the anterior chamber, as described inprevious studies.93,94 Isolated retinal samples were processed for immunocytochemistryas described in Figure 6 using primary antibodies for GS (Millipore; MAB302) and GFAP(Sigma; G3893) at dilutions of 1:1,000.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
324 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

retinal ischaemia proceeds for differentlengths of time and the severity and durationis related to the length of the ischaemicinterval.221 Amacrine cell death is also wellestablished although possible further deathas a function of time after reperfusion is notfully understood.222,223 Overall, there is littlechange in overall retinal amino acid levelsfrom two to 14 days after ischaemic insult94
but a dramatic alteration in ganglion cellnumbers with continuous ganglion celldeath for months after 90 minutes of ocularischaemia.221 The factors causing this pro-longed ganglion cell death are unknown butthis model may provide a unique opportu-nity to study cell death post-insult. Suchstudies would provide useful insights as tocell death and function in common clinicalconditions such as retinal detachment andthus provide a better understanding as to thereason(s) for poor recovery of vision second-ary to reattachment surgery.
Functional profiling of the ischaemic-reperfused rat retina was also dramaticallyaltered.94,95 Altered glutamate release byphotoreceptors was observed throughalterations in AGB permeation in theischaemic-reperfused retina. In addition, asub-population of amacrine cells loses itsexquisite sensitivity to activation by theglutamate analogue kainite.93 Followingischaemia-reperfusion, the remaining rodbipolar cells are dysfunctional due to a lackof glutamate release by photoreceptors aswell as altered metabolic profiles.93
Remodelling in retinal dystrophyAnatomical reshaping of inner retinalneurons occurs during retinal degenera-tion, including loss of bipolar cell dendrites,neurite sprouting and soma displace-ment.90–92,224 Inner retinal neurons alsodisplay neurochemical and functionalchanges.56,102,225–228 At late stages of retinaldegeneration (after complete loss of pho-toreceptors), rod bipolar cells lose theirresponsiveness to glutamate.228
An emerging concept from models ofretinal degeneration is alterations in gluta-mate receptor function. For example, in theretinal degeneration animal model, the rd1mouse, ON cone bipolar cells show anaberrant expression of functional AMPA/kainate receptors, with the apparent con-tinual preservation of receptors on OFFcone bipolar cells during the early stages ofretinal degeneration.56,96 ON cone bipolarcells with the normal metabotropic recep-tors and aberrant functional ionotropic
Figure 14. Taurine immunoreactivity in cat (A), puma (B), shingleback lizard (C), fresh-water crocodile (D), South African clawed frog (E), shark (F), collared peccary (G) andkangaroo (H). Scale bar 20 mm. Abbreviations as in Figures 1 and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 325

glutamate receptors would be unable torespond to glutamate activation, reflecting adetrimental effect of remodelling.
During later stages of inherited formsof retinal degeneration where there is pho-toreceptor loss, ON and OFF cone bipolarcells lose both mGluR6 and ionotropic gluta-mate receptor expression, respectively.56,91
mGluR6 function (as assessed via AGBgating) was not present in the rd1 mouseretina.96
The response to NMDA of amacrine cellsis also gradually altered during degenera-tion in the rd1 mouse retina. There is agradual loss of functional NMDA receptorresponse in immunocytochemically identi-fied amacrine cells that increased with agewith under 10 per cent of cells being acti-vated in the adult rd1 retina.96
These results also demonstrate that innerretinal glutamatergic drive is altered from anearly age in the rd1 mouse retina. The mor-phology and dendritic arbour of ganglioncells appear to be normal in both the rd1 andrd10 models.229–230 Stasheff231 has shown ahigh sustained spontaneous hyperactivity inOFF responses from ganglion cells in the rd1mouse early in the degenerative process(P14) with rhythmic (10 Hz) firing present
when photoreceptors have degenerated.229
Although the intrinsic physiology of gan-glion cells in the rd1 mouse is normal,229 theneurotransmitter receptors on ganglioncells may be abnormal.
The altered physiology of ganglion cellsconfirms that the input is abnormal and anyattempts to target ganglion cells as outputcells in intervention studies must take intoconsideration the altered input. Possiblecauses for the altered ganglion cell physiol-ogy could include changes in retinal cir-cuitry,56,229,231 including alterations in NMDAreceptors in the inner retina.
Afferent input to the NMDA receptor aswell as complex protein interactions areresponsible for receptor clustering at thesynapse.232 De-afferentation could stimu-late a down regulation and/or a change inthe subunit composition of glutamatereceptors.223,233
Functional changes at theneurotransmitter receptor levelGlutamate receptors are altered in rodentmodels of inherited retinal dystrophy. Twomajor changes have been reported: first,the down regulation of the metabotropic(mGluR6) receptors on the depolarising
bipolar cells; and second, a concurrentexpression of aberrant functional iono-tropic receptors on depolarising bipolarcells.56,90–92,96,234 Puthussery and colleagues234
propose that the down regulation of first themGluR6 receptor on rod bipolar cells, fol-lowed by the cone bipolar cells while largelyretaining ionotropic receptor function onOFF cone bipolar cells, signifies a need forthe mGluR6 pathway to receive glutamateactivation. Altered inner retinal function(presumed altered NMDA receptor), follow-ing known cone activity in the rd1 mouseretina,96 also suggests an activity-dependentalteration in function.
Further evidence of ‘plasticity’ in theretina includes glutamate receptor changessecondary to altered gene expression(CPG15) activated by a range of stimulants,including light. CPG15 gene expressionleads to an increased growth rate of retinalaxons and promoted synaptic maturation byrecruitment of synaptic AMPA receptors.235
Trafficking of AMPA receptors also occurswithin the retina through an activity-dependent plasticity that may be regulatedin the course of a normal light/dark cycle.236
Guenther and colleagues237 showed lightand development influence expression ofNMDA receptors. Kamphuis, Dijk andO’Brien238 found the gene expression of anaberrant glutamate receptor in rod bipolarcells under different physiological lightlevels. Whether these receptor subunitscombined to form functional ion channels isunknown. Rapid aberrant expression ofAMPA receptor subunit (GluR2) occurssecondary to light damage.49
CONCLUDING REMARKS
Advances in the basic sciences over the past60 years have lead to the discovery andcharacterisation of the biochemical path-ways involved in the visual cycle and pho-totransduction followed by gene expressionand associated mutations that lead toocular disease. The development of genetictechniques that allow the insertion of thecorrect genetic code into cells, has led toviable treatment options. For example,gene therapy is now possible with the resto-ration of sight in patients affected with theRPE65 mutation causing Leber’s congeni-tal amaurosis.72–78 Similarly, exciting devel-opments with nutritional supplements (forexample, using the retinoid QLT091001)has the potential to restore sight in muta-tions associated with lecithin:retinol acyl-
Figure 15. Pseudocolour theme map developed after applyingpattern recognition to serial section amino acid labelling in adultmouse retina. Each unique colour reflects cells with distinct aminoacid neurochemical signatures.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
326 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

transferase (LRAT) or RPE65 that causesLeber’s congenital amaurosis or retinitispigmentosa.86
Similar advances are required in under-standing the poor visual outcome secondaryto, for example, retinal detachment surgery.Recent studies providing an explanation forthis poor outcome have come about throughapplication of basic scientific techniques,such as amino acid localisation in the verte-brate retina, followed by understanding andlocalisation of the amino acid receptors, andmore recently, development of techniquesto functionally map responses from retinal
neurons.47,56,96,97,193 These and other methodshave revealed detrimental remodellinginvolving specific subclasses of glutamatereceptors that occur over a precise timeframe. Further advances in fully characteris-ing the time course, the receptors involvedand the physiology of the aberrant expres-sion of glutamate receptors will provide thefoundations to target therapies for the resto-ration of vision before detrimental remodel-ling occurs.
Finally, other parts of the eye are beingexplored to better understand both normaland abnormal amino acid neurochemistry.
Recently, work has characterised amino acidlabelling and amino acid transporters in theanterior eye.154,239–242 Changes to amino acidand glutamate metabolic enzyme are notrestricted to the retina secondary to meta-bolic insult: altered enzymatic expression inthe ciliary processes and dramatic altera-tions in amino acid labelling patterns werefound in the ciliary processes and lens.154,242
Despite these dramatic amino acid changesin the ciliary processes and in particular thelens, aqueous amino acid levels remainedrelatively unchanged.242 We have proposedthat the lens is a reservoir for amino acidsand may provide a mechanism of amino acidhomeostasis for the eye.242 As we furtherexplore this possibility, such interesting find-ings highlight the need to better understandthe intricate interrelationships betweendifferent ocular structures,243 therebyexpanding our understanding and providefurther pathways for therapeutic interven-tion leading to better management of oculardisease.
ACKNOWLEDGEMENTSThis work was supported, in part, by grantsfrom the National Health and MedicalResearch Council of Australia (1009342and 1021042) and Optometrists AssociationAustralia.
We are grateful to Dr Robert Marc(University of Utah) for the kind gift of theAGB and amino acid antibodies and for hisguidance.
Ethics approval was obtained for the col-lection of eyes from university laboratoriesand from other institutions. Samples of dif-ferent animal species not previously pub-lished were obtained either as part ofexperiments within the Retinal NetworksLaboratory at the University of Melbourne,the University of New South Wales orUniversity of Auckland or post-mortemfrom animals at the Melbourne Zoo, Queen-scliff Marine Station, Healesville bird sanc-tuary or from Northern Territory crocodilefarms.
We are grateful for access to the eyes pro-vided to us by:
The Melbourne Zoo, Parkville Victoria3052, Australia
Queenscliff Marine Station, QueenscliffVictoria 3225, Australia
Healesville Sanctuary, Badger CreekRoad, Healesville Victoria 3777, Australia
Northern Territory Crocodile Farms(Crocodylus Park), Karama NT 0812,Australia (Grahame Webb)
Figure 16. Schematic of a glutamate ionotropic receptor showing basal agmatine (AGB)labelling (A) and secondary to 1 mmol/l n-methyl-D-aspartate (NMDA) activation (B). A.Schematic of a receptor including the different ions or charged compounds (K+, Na+,Ca2+, AGB—a divalent polyamine) that move extra- or intra-cellular once the receptor isactivated. Receptor activation occurs when a glutamate agonist (green object) activatesthe receptor (grey rectangle). The right hand panels display AGB immunoreactivity usingsilver-intensified immunogold detection of the antigen (details in Figure 3 caption). BasalAGB labelling is evident in panel A, with little inner retinal labelling. When 1 mmol/lNMDA is included in the incubation medium, AGB entry is evident in small amacrine cellsomata and a number of intensely labelled large ganglion cells are evident within theganglion cell layer. Isolated rabbit retina was incubated for six minutes in physiologicalbuffer containing 25 mmol/l exogenous AGB with or without 1 mmol/l NMDA (Sun, Raitand Kalloniatis207). Abbreviations as in Figures 1 and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 327

REFERENCES1. Dowling JE, Werblin FS. Organization of retina
of the mudpuppy, Necturus maculosus. I.Synaptic structure. J Neurophysiol 1969; 32: 315–338.
2. Wässle H, Boycott BB. Functional architecture ofthe mammalian retina. Physiol Rev 1991; 71: 447–480.
3. Werblin FS, Dowling JE. Organization of theretina of the mudpuppy, Necturus maculosus. II.Intracellular recording. J Neurophysiol 1969; 32:339–355.
4. DeVries SH. Bipolar cells use kainate and AMPAreceptors to filter visual information into separatechannels. Neuron 2000; 28: 847–856.
5. Euler T, Schneider H, Wässle H. Glutamateresponses of bipolar cells in a slice preparationof the rat retina. J Neurosci 1996; 16: 2934–2944.
6. Kuffler SW. Discharge patterns and functionalorganization of mammalian retina. J Neurophysiol1953; 16: 37–68.
7. Stell WK. The structure and relationships of hori-zontal cells and photoreceptor-bipolar synapticcomplexes in goldfish retina. Am J Anat 1967; 121:401–423.
8. Stell WK, Lightfoot DO. Color-specific intercon-nections of cones and horizontal cells in the retinaof the goldfish. J Comp Neurol 1975; 159: 473–502.
9. Nelson R, Kolb H. Synaptic patterns and responseproperties of bipolar and ganglion cells in the catretina. Vision Res 1983; 23: 1183–1195.
10. Kolb H. The organization of the outer plexiformlayer in the retina of the cat: electron
microscopic observations. J Neurocytol 1977; 6:131–153.
11. Kolb H, Famiglietti EV. Rod and cone pathways inthe inner plexiform layer of cat retina. Science1974; 186: 47–49.
12. Lin P, Mruthyunjaya P. Retinal manifestations ofoncologic and hematologic conditions. Int Oph-thalmol Clin 2012; 52: 67–91.
13. Milam AH, Saari JC, Jacobson SG, Lubinski WP,Feun LG, Alexander KR. Autoantibodies againstretinal bipolar cells in cutaneous melanoma-associated retinopathy. Invest Ophthalmol Vis Sci1993; 34: 91–100.
14. Dhingra A, Fina ME, Neinstein A, Ramsey DJ, XuY, Fishman GA, Alexander KR et al. Autoantibod-ies in melanoma-associated retinopathy targetTRPM1 cation channels of retinal ON bipolarcells. J Neurosci 2011; 31: 3962–3967.
15. Jonas JB, Zach FM, Naumann GO. Dark adapta-tion in glaucomatous and nonglaucomatous opticnerve atrophy. Graefes Arch Clin Exp Ophthalmol1990; 228: 321–325.
16. Sherman MD, Henkind P. Photostress recovery inchronic open angle glaucoma. Br J Ophthalmol1988; 72: 641–645.
17. Tomita T. Retrospective review of retinal circuitry.Vision Res 1986; 26: 1339–1350.
18. Masland RH. The fundamental plan of the retina.Nat Neurosci 2001; 4: 877–886.
19. Masland RH. Neuronal cell types. Curr Biol 2004;14: R497–R500.
20. Ramón Y, Cajal S. The vertebrate retina. In:Rodieck RW ed. The Vertebrate Retina; Principlesof Structure and Function. San Francisco:Freeman, 1973. p 733–904.
21. Vaney DI. Retinal neurons: cell types and couplednetworks. Prog Brain Res 2002; 136: 239–254.
22. Haverkamp S, Grünert U, Wässle H. The conepedicle, a complex synapse in the retina. Neuron2000; 27: 85–95.
23. MacNeil MA, Masland RH. Extreme diversityamong amacrine cells: implications for function.Neuron 1998; 20: 971–982.
24. Meyer JS, Katz ML, Kirk MD. Stem cells for retinaldegenerative disorders. Ann N Y Acad Sci 2005;1049: 135–145.
25. Harvey AR, Hellstrom M, Rodger J. Gene therapyand transplantation in the retinofugal pathway.Prog Brain Res 2009; 175: 151–161.
26. Djamgoz MB, Stell WK, Chin CA, Lam DM. Anopiate system in the goldfish retina. Nature 1981;292: 620–623.
27. Stell WK. Putative peptide transmitters, amacrinecell diversity and function in the inner plexiformlayer. In: Gallego A, Gouras P eds. Neurocircuitryof the Retina: a Cajal Memorial. New York: Else-vier, 1985. p 171–187.
28. Greferath U, Grünert U, Wässle H. Rod bipolarcells in the mammalian retina show protein kinaseC-like immunoreactivity. J Comp Neurol 1990; 301:433–442.
29. Bagnoli P, Dal Monte M, Casini G. Expression ofneuropeptides and their receptors in the develop-ing retina of mammals. Histol Histopathol 2003; 18:1219–1242.
30. Marshak DW. Peptidergic neurons of teleostretinas. Vis Neurosci 1992; 8: 137–144.
31. Kalloniatis M, Tomisich G. Amino acid neuro-chemistry of the vertebrate retina. Prog Retin EyeRes 1999; 18: 811–866.
Figure 17. Rat retina agmatine (AGB) activated using 20 mmol/l kainate combined with cell marker (choline acetyl transferase [ChAT]immunoreactivity) in A-C and glutamic acid decarboxylase isoform 67 (GAD 67) in D to F. White arrowheads indicate amacrine cellsco-localised with cell markers, black arrowheads indicate amacrine cells positive for a cell marker but not activated by kainate and whitearrows indicate kainite-activated amacrine cells with no cell marker. Isolated retinal samples were processed for immunocytochemistryas described in Figure 6 using primary antibodies for AGB (Millipore; AB1568), ChAT (Millipore; MAB5270) and GAD67 (Millipore;MAB5406) at dilutions of 1:100, 1:500 and 1:500, respectively. Scale bar 20 mm. Abbreviations as in Figures 1 and 4.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
328 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

32. Marc RE. Structural organization of GABAergiccircuitry in ectotherm retinas. Prog Brain Res 1992;90: 61–92.
33. Marc RE. The role of glycine in retinal circuitry.In: Morgan WW ed. Retinal Transmitters andModulators: Models for the Brain. Boca Raton:CRC Press, 1985. p 119–158.
34. Marc RE. The role of glycine in the mammalianretina. In: Osborne NN, Chader GJ eds. Progressin Retinal Research. Oxford: Pergamon Press,1989. p 67–107.
35. Marc RE. Retinal neurotransmitters. In: ChalupaLM, Werner J eds. The Visual Neurosciences.Cambridge: MIT Press, 2004. p 315–330.
36. Marc RE. The synaptic organization of the retina.In: Levin LA, Nilsson SFE, Ver Hoeve J, Wu S,Kaufman PL eds. Adler’s Physiology of theEye, 11th ed. Saunders: Elsevier, 2011. p 443–458.
37. Massey SC, Miller RF. N-methyl-D-aspartate recep-tors of ganglion cells in rabbit retina. J Neurophysiol1990; 63: 16–30.
38. Pow DV. Amino acids and their transporters in theretina. Neurochem Int 2001; 38: 463–484.
39. Thoreson WB, Witkovsky P. Glutamate receptorsand circuits in the vertebrate retina. Prog Retin EyeRes 1999; 18: 765–810.
40. Yazulla S. GABAergic mechanisms in the retina.In: Osborne NN, Chader GJ eds. Progress inRetinal Research. New York: Pergamon Press,1986. p 1–51.
41. Thelen U, Amler S, Osada N, Gerding H.Outcome of surgery after macula-off retinaldetachment—results from MUSTARD, one of thelargest databases on buckling surgery in Europe.Acta Ophthalmol 2012; 90: 481–486.
42. Pastor JC, Fernandez I, Rodriguez de la Rua E,Coco R, Sanabria-Ruiz Colmenares MR, Sanchez-Chicharro D, Martinho R et al. Surgical outcomesfor primary rhegmatogenous retinal detachmentsin phakic and pseudophakic patients: the Retina 1Project–report 2. Br J Ophthalmol 2008; 92: 378–382.
43. Wykoff CC, Smiddy WE, Mathen T, Schwartz SG,Flynn HW Jr, Shi W. Fovea-sparing retinal detach-ments: time to surgery and visual outcomes. Am JOphthalmol 2010; 150: 205–210.
44. Mervin K, Valter K, Maslim J, Lewis G, Fisher S,Stone J. Limiting photoreceptor death and decon-struction during experimental retinal detach-ment: the value of oxygen supplementation. Am JOphthalmol 1999; 128: 155–164.
45. Cook B, Lewis GP, Fisher SK, Adler R. Apoptoticphotoreceptor degeneration in experimentalretinal detachment. Invest Ophthalmol Vis Sci 1995;36: 990–996.
46. de Souza CF, Kalloniatis M, Christie D, Polking-horne P, McGhee CN, Acosta ML. Creatine Trans-porter Immunolocalization in aged Human andDetached Retinas. Invest Ophthalmol Vis Sci 2012;53: 1936–1945
47. de Souza CF, Kalloniatis M, Polkinghorne PJ,McGhee CN, Acosta ML. Functional and anatomi-cal remodeling in human retinal detachment. ExpEye Res 2012; 97: 73–89.
48. de Souza CF, Kalloniatis M, Polkinghorne PJ,McGhee CN, Acosta ML. Functional activa-tion of glutamate ionotropic receptors in thehuman peripheral retina. Exp Eye Res 2012; 94:71–84.
49. Lin Y, Jones BW, Liu A, Vazquez-Chona FR, Lau-ritzen JS, Ferrell WD, Marc RE. Rapid glutamate
receptor 2 trafficking during retinal degenera-tion. Mol Neurodegener 2012; 7: 7.
50. Sarks SH, Arnold JJ, Killingsworth MC, Sarks JP.Early drusen formation in the normal and agingeye and their relation to age related maculopathy:a clinicopathological study. Br J Ophthalmol 1999;83: 358–368.
51. Taylor HR, Keeffe JE, Vu HT, Wang JJ, RochtchinaE, Pezzullo ML, Mitchell P. Vision loss in Australia.Med J Aust 2005; 182: 565–568.
52. Wickham L, Lewis GP, Charteris DG, Fisher SK,Da Cruz L. Histological analysis of retinas sampledduring translocation surgery: a comparison withnormal and transplantation retinas. Br J Ophthal-mol 2009; 93: 969–973.
53. Oshitari T, Yamamoto S, Hata N, Roy S.Mitochondria- and caspase-dependent cell deathpathway involved in neuronal degeneration indiabetic retinopathy. Br J Ophthalmol 2008; 92:552–556.
54. Bessero AC, Clarke PG. Neuroprotection for opticnerve disorders. Curr Opin Neurol 2010; 23: 10–15.
55. Kern TS, Barber AJ. Retinal ganglion cells in dia-betes. J Physiol 2008; 586: 4401–4408.
56. Marc RE, Jones BW, Anderson JR, Kinard K,Marshak DW, Wilson JH, Wensel T et al. Neuralreprogramming in retinal degeneration. InvestOphthalmol Vis Sci 2007; 48: 3364–3371.
57. Marc RE, Jones BW, Watt CB, Vazquez-Chona F,Vaughan DK, Organisciak DT. Extreme retinalremodeling triggered by light damage: implica-tions for age related macular degeneration.Mol Vis 2008; 14: 782–806.
58. Marc RE, Murry RF, Fisher SK, Linberg KA, LewisGP. Amino acid signatures in the detached catretina. Invest Ophthalmol Vis Sci 1998; 39: 1694–1702.
59. Sullivan R, Penfold P, Pow DV. Neuronal migra-tion and glial remodeling in degenerating retinasof aged rats and in nonneovascular AMD. InvestOphthalmol Vis Sci 2003; 44: 856–865.
60. Sullivan RK, Woldemussie E, Pow DV. Dendriticand synaptic plasticity of neurons in the humanage-related macular degeneration retina. InvestOphthalmol Vis Sci 2007; 48: 2782–2791.
61. Pow DV, Sullivan RK. Nuclear kinesis, neuritesprouting and abnormal axonal projections ofcone photoreceptors in the aged and AMD-afflicted human retina. Exp Eye Res 2007; 84: 850–857.
62. Downie LE, Hatzopoulos KM, Pianta MJ, VingrysAJ, Wilkinson-Berka JL, Kalloniatis M, FletcherEL. Angiotensin type-1 receptor inhibition is neu-roprotective to amacrine cells in a rat model ofretinopathy of prematurity. J Comp Neurol 2010;518: 41–63.
63. Wong VH, Bui BV, Vingrys AJ. Clinical and experi-mental links between diabetes and glaucoma. ClinExp Optom 2011; 94: 4–23.
64. Fletcher EL, Downie LE, Hatzopoulos K, VesseyKA, Ward MM, Chow CL, Pianta MJ et al. Thesignificance of neuronal and glial cell changes inthe rat retina during oxygen-induced retinopathy.Doc Ophthalmol 2010; 120: 67–86.
65. Batcha AH, Greferath U, Jobling AI, Vessey KA,Ward MM, Nithianantharajah J, Hannan AJ et al.Retinal dysfunction, photoreceptor protein dys-regulation and neuronal remodelling in the R6/1mouse model of Huntington’s disease. NeurobiolDis 2012; 45: 887–896.
66. Kalloniatis M, Fletcher EL. Retinitis pigmentosa:understanding the clinical presentation,
mechanisms and treatment options. Clin ExpOptom 2004; 87: 65–80.
67. Hartong DT, Berson EL, Dryja TP. Retinitis pig-mentosa. Lancet 2006; 368: 1795–1809.
68. Goodwin P. Hereditary retinal disease. Curr OpinOphthalmol 2008; 19: 255–262.
69. Cideciyan AV, Swider M, Aleman TS, Tsybovsky Y,Schwartz SB, Windsor EA, Roman AJ et al. ABCA4disease progression and a proposed strategyfor gene therapy. Hum Mol Genet 2009; 18: 931–941.
70. Massof RW, Fishman GA. How strong is theevidence that nutritional supplements slow theprogression of retinitis pigmentosa? Arch Ophthal-mol 2010; 128: 493–495.
71. Maeda T, Golczak M, Maeda A. RetinalPhotodamage mediated by all-trans-retinal.Photochem Photobiol 2012; 88: 1309–1319.
72. Jacobson SG, Cideciyan AV, Ratnakaram R, HeonE, Schwartz SB, Roman AJ, Peden MC et al. Genetherapy for leber congenital amaurosis caused byRPE65 mutations: safety and efficacy in 15 chil-dren and adults followed up to 3 years. Arch Oph-thalmol 2012; 130: 9–24.
73. Cideciyan AV. Leber congenital amaurosis due toRPE65 mutations and its treatment with genetherapy. Prog Retin Eye Res 2010; 29: 398–427.
74. Sahni JN, Angi M, Irigoyen C, Semeraro F,Romano MR, Parmeggiani F. Therapeuticchallenges to retinitis pigmentosa: from neuro-protection to gene therapy. Curr Genomics 2011;12: 276–284.
75. Beltran WA, Cideciyan AV, Lewin AS, Iwabe S,Khanna H, Sumaroka A, Chiodo VA et al. Genetherapy rescues photoreceptor blindness in dogsand paves the way for treating human X-linkedretinitis pigmentosa. Proc Natl Acad Sci USA 2012;109: 2132–2137.
76. Bainbridge JW, Smith AJ, Barker SS, Robbie S,Henderson R, Balaggan K, Viswanathan A et al.Effect of gene therapy on visual function inLeber’s congenital amaurosis. N Engl J Med 2008;358: 2231–2239.
77. Maguire AM, Simonelli F, Pierce EA, Pugh EN Jr,Mingozzi F, Bennicelli J, Banfi S et al. Safetyand efficacy of gene transfer for Leber’s con-genital amaurosis. N Engl J Med 2008; 358: 2240–2248.
78. Cideciyan AV, Aleman TS, Boye SL, Schwartz SB,Kaushal S, Roman AJ, Pang JJ et al. Human genetherapy for RPE65 isomerase deficiency activatesthe retinoid cycle of vision but with slow rodkinetics. Proc Natl Acad Sci USA 2008; 105: 15112–15117.
79. MacLaren RE, Pearson RA, MacNeil A, DouglasRH, Salt TE, Akimoto M, Swaroop A et al. Retinalrepair by transplantation of photoreceptor pre-cursors. Nature 2006; 444: 203–207.
80. Pearson RA, Barber AC, West EL, MacLaren RE,Duran Y, Bainbridge JW, Sowden JC et al. Tar-geted disruption of outer limiting membranejunctional proteins (Crb1 and ZO-1) increasesintegration of transplanted photoreceptor pre-cursors into the adult wild-type and degeneratingretina. Cell Transplant 2010; 19: 487–503.
81. Tibbetts MD, Samuel MA, Chang TS, Ho AC. Stemcell therapy for retinal disease. Curr Opin Ophthal-mol 2012; 23: 226–234.
82. Pearson RA, Barber AC, Rizzi M, Hippert C, XueT, West EL, Duran Y et al. Restoration of visionafter transplantation of photoreceptors. Nature2012; 485 (7396): 99–103.
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 329

83. Ma L, Kaufman Y, Zhang J, Washington I. C20-D3-vitamin A slows lipofuscin accumulation and elec-trophysiological retinal degeneration in a mousemodel of Stargardt disease. J Biol Chem 2011; 286:7966–7974.
84. Maeda T, Cideciyan AV, Maeda A, Golczak M,Aleman TS, Jacobson SG, Palczewski K. Loss ofcone photoreceptors caused by chromophoredepletion is partially prevented by the artificialchromophore pro-drug, 9-cis-retinyl acetate. HumMol Genet 2009; 18: 2277–2287.
85. Berson EL, Rosner B, Sandberg MA, Weigel-Difranco C, Willett WC. Omega-3 intake andvisual acuity in patients with retinitis pigmentosareceiving vitamin A. Arch Ophthalmol 2012; 130:707–711.
86. Clinicaltrials.gov/ct2/show/NCT01014052:Safety/Proof of Concept Study of OralQLT091001 in Subjects With Leber CongenitalAmaurosis (LCA) or Retinitis Pigmentosa (RP)Due to Retinal Pigment Epithelial 65 Protein(RPE65) or Lecithin:Retinol Acyltransferase(LRAT) Mutations. Clinicaltrials.gov/ct2/show/NCT01543906: Oral QLT091001 in Retinitis Pig-mentosa (RP) Subjects With an AutosomalDominant Mutation in Retinal Pigment Epithelial65 Protein (RPE65).
87. Busch EM, Gorgels TG, Van Norren D. Filling-inafter focal loss of photoreceptors in rat retina. ExpEye Res 1999; 68: 485–492.
88. Paulus YM, Jain A, Gariano RF, Stanzel BV,Marmor M, Blumenkranz MS, Palanker D.Healing of retinal photocoagulation lesions.Invest Ophthalmol Vis Sci 2008; 49: 5540–5545.
89. Muqit MM, Denniss J, Nourrit V, Marcellino GR,Henson DB, Schiessl I, Stanga PE. Spatial andspectral imaging of retinal laser photocoagulationburns. Invest Ophthalmol Vis Sci 2011; 52: 994–1002.
90. Pignatelli V, Cepko CL, Strettoi E. Inner retinalabnormalities in a mouse model of Leber’s con-genital amaurosis. J Comp Neurol 2004; 469: 351–359.
91. Strettoi E, Pignatelli V. Modifications of retinalneurons in a mouse model of retinitis pigmen-tosa. Proc Natl Acad Sci USA 2000; 97: 11020–11025.
92. Strettoi E, Pignatelli V, Rossi C, Porciatti V, FalsiniB. Remodeling of second-order neurons in theretina of rd/rd mutant mice. Vision Res 2003; 43:867–877.
93. Sun D, Bui BV, Vingrys AJ, Kalloniatis M. Altera-tions in photoreceptor-bipolar cell signaling fol-lowing ischemia/reperfusion in the rat retina.J Comp Neurol 2007; 505: 131–146.
94. Sun D, Vingrys AJ, Kalloniatis M. Metabolic andfunctional profiling of the ischemic/reperfusedrat retina. J Comp Neurol 2007; 505: 114–130.
95. Sun D, Vingrys AJ, Kalloniatis M. Metabolic andfunctional profiling of the normal rat retina.J Comp Neurol 2007; 505: 92–113.
96. Chua J, Fletcher EL, Kalloniatis M. Functionalremodeling of glutamate receptors by inner re-tinal neurons occurs from an early stage of retinaldegeneration. J Comp Neurol 2009; 514: 473–491.
97. Marc RE, Jones BW, Watt CB, Strettoi E. Neuralremodeling in retinal degeneration. Prog Retin EyeRes 2003; 22: 607–655.
98. Ehinger B, Ottersen OP, Storm-Mathisen J,Dowling JE. Bipolar cells in the turtle retina arestrongly immunoreactive for glutamate. Proc NatlAcad Sci USA 1988; 85: 8321–8325.
99. Marc RE, Liu WL, Kalloniatis M, Raiguel SF,van Haesendonck E. Patterns of glutamate
immunoreactivity in the goldfish retina. J Neurosci1990; 10: 4006–4034.
100. Kalloniatis M, Fletcher EL. Immunocytochemicallocalization of the amino acid neurotransmittersin the chicken retina. J Comp Neurol 1993; 336:174–193.
101. Pow DV, Robinson SR. Glutamate in some retinalneurons is derived solely from glia. Neuroscience1994; 60: 355–366.
102. Fletcher EL, Kalloniatis M. Neurochemical archi-tecture of the normal and degenerating rat retina.J Comp Neurol 1996; 376: 343–360.
103. Bender DA. Amino Acid Metabolism, 2nd ed.Chichester, New York: Wiley, 1985.
104. Erecinska M, Silver IA. Metabolism and role ofglutamate in mammalian brain. Prog Neurobiol1990; 35: 245–296.
105. Barlow HB, Blakemore C, Pettigrew JD. Theneural mechanism of binocular depth discrimina-tion. J Physiol 1967; 193: 327–342.
106. Kolb H. Amacrine cells of the mammalian retina:neurocircuitry and functional roles. Eye (Lond)1997; 11: 904–923.
107. Cleland BG, Levick WR. Physiology of cat retinalganglion cells. Invest Ophthalmol 1972; 11: 285–291.
108. Stone J, Fukuda Y. Properties of cat retinal gan-glion cells: a comparison of W-cells with X- andY-cells. J Neurophysiol 1974; 37: 722–748.
109. Ewer RF. The Carnivores. Ithaca, N.Y.: CornellUniversity Press, 1973.
110. Hall MI. Comparative analysis of the size andshape of the lizard eye. Zoology (Jena) 2008; 111:62–75.
111. Fleishman LJ, Howland HC, Howland MJ, RandAS, Davenport ML. Crocodiles don’t focus under-water. J Comp Physiol A 1988; 163: 441–443.
112. Lettvin JY, Maturana HR, Pitts WH, McCullochWS. Two remarks on the visual system of the frog.AFOSR TR United States Air Force Off Sci Res 1960;60–77: 1–25.
113. Stirling RV, Merrill EG. Functional morphologyof frog retinal ganglion cells and their centralprojections: the dimming detectors. J Comp Neurol1987; 258: 477–495.
114. Schieber NL, Collin SP, Hart NS. Comparativeretinal anatomy in four species of elasmobranch.J Morphol 2012; 273: 423–440.
115. Litherland L, Collin SP. Comparative visual func-tion in elasmobranchs: spatial arrangement andecological correlates of photoreceptor and gan-glion cell distributions. Vis Neurosci 2008; 25: 549–561.
116. MacDonald D. The Encyclopedia of Mammals.New York: Facts on File, 1984.
117. Dawson TJ. Kangaroos: Biology of the LargestMarsupial. Sydney: University of New South WalesPress, 1995.
118. Davanger S, Ottersen OP, Storm-Mathisen J.Glutamate, GABA and glycine in the humanretina: an immunocytochemical investigation.J Comp Neurol 1991; 311: 483–494.
119. Crooks J, Kolb H. Localization of GABA, glycine,glutamate and tyrosine hydroxylase in the humanretina. J Comp Neurol 1992; 315: 287–302.
120. Martin PR, Grünert U. Spatial density and immu-noreactivity of bipolar cells in the macaquemonkey retina. J Comp Neurol 1992; 323: 269–287.
121. Kalloniatis M, Marc RE, Murry RF. Amino acidsignatures in the primate retina. J Neurosci 1996;16: 6807–6829.
122. Brandstätter JH, Koulen P, Wässle H. Diversity ofglutamate receptors in the mammalian retina.Vision Res 1998; 38: 1385–1397.
123. Vardi N, Duvoisin R, Wu G, Sterling P. Localiza-tion of mGluR6 to dendrites of ON bipolar cells inprimate retina. J Comp Neurol 2000; 423: 402–412.
124. Hack I, Frech M, Dick O, Peichl L, BrandstätterJH. Heterogeneous distribution of AMPA gluta-mate receptor subunits at the photoreceptor syn-apses of rodent retina. Eur J Neurosci 2001; 13:15–24.
125. Grünert U, Lin B, Martin PR. Glutamate receptorsat bipolar synapses in the inner plexiform layer ofprimate retina: light microscopic analysis. J CompNeurol 2003; 466: 136–147.
126. Miller RF. Cell communication mechanisms inthe vertebrate retina the proctor lecture. InvestOphthalmol Vis Sci 2008; 49: 5184–5198.
127. Fletcher EL, Hack I, Brandstätter JH, Wässle H.Synaptic localization of NMDA receptor subunitsin the rat retina. J Comp Neurol 2000; 420: 98–112.
128. Kalloniatis M, Sun D, Foster L, Haverkamp S,Wässle H. Localization of NMDA receptor subu-nits and mapping NMDA drive within the mam-malian retina. Vis Neurosci 2004; 21: 587–597.
129. Haverkamp S, Wässle H. Immunocytochemicalanalysis of the mouse retina. J Comp Neurol 2000;424: 1–23.
130. Deniz S, Wersinger E, Schwab Y, Mura C, ErdelyiF, Szabo G, Rendon A, Sahel JA, Picaud S, RouxMJ. Mammalian retinal horizontal cells areunconventional GABAergic neurons. J Neurochem2011; 116: 350–362.
131. Robin LN, Kalloniatis M. Interrelationshipbetween retinal ischaemic damage and turnoverand metabolism of putative amino acid neuro-transmitters, glutamate and GABA. Doc Ophthal-mol 1992; 80: 273–300.
132. Mosinger JL, Yazulla S, Studholme KM. GABA-likeimmunoreactivity in the vertebrate retina: aspecies comparison. Exp Eye Res 1986; 42: 631–644.
133. Grünert U, Wässle H. GABA-like immunoreactiv-ity in the macaque monkey retina: a light andelectron microscopic study. J Comp Neurol 1990;297: 509–524.
134. Lassova L, Fina M, Sulaiman P, Vardi N. Immuno-cytochemical evidence that monkey rod bipolarcells use GABA. Eur J Neurosci 2010; 31: 685–696.
135. Kao YH, Lassova L, Bar-Yehuda T, Edwards RH,Sterling P, Vardi N. Evidence that certain retinalbipolar cells use both glutamate and GABA.J Comp Neurol 2004; 478: 207–218.
136. Rogers PC, Pow DV. Immunocytochemical evi-dence for an axonal localization of GABA in theoptic nerves of rabbits, rats, and cats. Vis Neurosci1995; 12: 1143–1149.
137. Vaney DI, Nelson JC, Pow DV. Neurotransmittercoupling through gap junctions in the retina.J Neurosci 1998; 18: 10594–10602.
138. Marc RE, Jones BW. Molecular phenotyping ofretinal ganglion cells. J Neurosci 2002; 22: 413–427.
139. Mills SL, Massey SC. A series of biotinylated tracersdistinguishes three types of gap junction in retina.J Neurosci 2000; 20: 8629–8636.
140. Trexler EB, Li W, Mills SL, Massey SC. Couplingfrom AII amacrine cells to ON cone bipolar cells isbidirectional. J Comp Neurol 2001; 437: 408–422.
141. Kalloniatis M, Marc RE. Interplexiform cells ofthe goldfish retina. J Comp Neurol 1990; 297: 340–358.
142. Wässle H, Koulen P, Brandstätter JH, Fletcher EL,Becker CM. Glycine and GABA receptors in the
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
330 Clinical and Experimental Optometry © 2013 Optometrists Association Australia

mammalian retina. Vision Res 1998; 38: 1411–1430.
143. Grünert U. Distribution of GABA and glycine re-ceptors on bipolar and ganglion cells in the mam-malian retina. Microsc Res Tech 2000; 50: 130–140.
144. Voaden M, J. The localization and metabolism ofneuroactive amino acids in the retina. In:Fonnum F ed. Amino Acids as Chemical Transmit-ters. New York: Plenum Press, 1978. p 257–274.
145. Kalloniatis M, Tomisich G, Marc RE. Neurochemi-cal signatures revealed by glutamine labeling inthe chicken retina. Vis Neurosci 1994; 11: 793–804.
146. Berg JM, Tymoczko JL, Stryer L. Biochemistry, 6thed. New York: WH Freeman, 2007.
147. Bui BV, Vingrys AJ, Wellard JW, Kalloniatis M.Monocarboxylate transport inhibition altersretinal function and cellular amino acid levels.Eur J Neurosci 2004; 20: 1525–1537.
148. Kalloniatis M, Napper GA. Glutamate metabolicpathways in displaced ganglion cells of thechicken retina. J Comp Neurol 1996; 367: 518–536.
149. Barnett NL, Pow DV, Robinson SR. Inhibition ofMüller cell glutamine synthetase rapidly impairsthe retinal response to light. Glia 2000; 30: 64–73.
150. Bui BV, Hu RG, Acosta ML, Donaldson P, VingrysAJ, Kalloniatis M. Glutamate metabolic pathwaysand retinal function. J Neurochem 2009; 111: 589–599.
151. Kalloniatis M, Napper GA. Retinal neurochemicalchanges following application of glutamate as ametablolic substrate. Clin Exp Optom 2002; 85:27–36.
152. Chin CA, Lam DM. The uptake and release of[3H]glycine in the goldfish retina. J Physiol 1980;308: 185–195.
153. Hu RG, Lim J, Donaldson PJ, Kalloniatis M. Char-acterization of the cystine/glutamate transporterin the outer plexiform layer of the vertebrateretina. Eur J Neurosci 2008; 28: 1491–1502.
154. Hu RG, Lim JC, Kalloniatis M, Donaldson PJ.Cellular localization of glutamate and glutaminemetabolism and transport pathways in the ratciliary epithelium. Invest Ophthalmol Vis Sci 2011;52: 3345–3353.
155. Reye P, Sullivan R, Fletcher EL, Pow DV. Distribu-tion of two splice variants of the glutamate trans-porter GLT1 in the retinas of humans, monkeys,rabbits, rats, cats, and chickens. J Comp Neurol2002; 445: 1–12.
156. Eliasof S, Arriza JL, Leighton BH, Amara SG,Kavanaugh MP. Localization and function of fiveglutamate transporters cloned from the salaman-der retina. Vision Res 1998; 38: 1443–1454.
157. Bringmann A, Pannicke T, Biedermann B,Francke M, Iandiev I, Grosche J, Wiedemann Pet al. Role of retinal glial cells in neurotransmitteruptake and metabolism. Neurochem Int 2009; 54:143–160.
158. Rauen T, Taylor WR, Kuhlbrodt K, Wiessner M.High-affinity glutamate transporters in the ratretina: a major role of the glial glutamate trans-porter GLAST-1 in transmitter clearance. CellTissue Res 1998; 291: 19–31.
159. Marc RE, Lam DM. Uptake of aspartic andglutamic acid by photoreceptors in goldfishretina. Proc Natl Acad Sci USA 1981; 78: 7185–7189.
160. Olive MF, Cleva RM, Kalivas PW, Malcolm RJ.Glutamatergic medications for the treatment ofdrug and behavioral addictions. Pharmacol BiochemBehav 2012; 100: 801–810.
161. Hashimoto K. Emerging role of glutamatein the pathophysiology of major depressive
disorder. Brain Res Rev 2009; 61: 105–123.
162. Sena DF, Ramchand K, Lindsley K. Neuroprotec-tion for treatment of glaucoma in adults. CochraneDatabase Syst Rev 2010: CD006539.
163. Calaza KC, Gardino PF, de Mello FG. Transportermediated GABA release in the retina: role of exci-tatory amino acids and dopamine. Neurochem Int2006; 49: 769–777.
164. Muller JF, Marc RE. GABA-ergic and glycinergicpathways in the inner plexiform layer of the gold-fish retina. J Comp Neurol 1990; 291: 281–304.
165. Brecha NC, Weigmann C. Expression of GAT-1, ahigh-affinity gamma-aminobutyric acid plasmamembrane transporter in the rat retina. J CompNeurol 1994; 345: 602–611.
166. Johnson J, Chen TK, Rickman DW, Evans C,Brecha NC. Multiple gamma-Aminobutyric acidplasma membrane transporters (GAT-1, GAT-2,GAT-3) in the rat retina. J Comp Neurol 1996; 375:212–224.
167. Pow DV, Baldridge W, Crook DK. Activity-dependent transport of GABA analogues into spe-cific cell types demonstrated at high resolutionusing a novel immunocytochemical strategy. Neu-roscience 1996; 73: 1129–1143.
168. Howard P, Twycross R, Shuster J, Mihalyo M, RemiJ, Wilcock A. Anti-epileptic drugs. J Pain SymptomManage 2011; 42: 788–804.
169. Sergott RC. Recommendations for visual evalua-tions of patients treated with vigabatrin. Curr OpinOphthalmol 2010; 21: 442–446.
170. Wild JM, Robson CR, Jones AL, Cunliffe IA, SmithPE. Detecting vigabatrin toxicity by imaging of theretinal nerve fiber layer. Invest Ophthalmol Vis Sci2006; 47: 917–924.
171. Hosking SL, Hilton EJ. Neurotoxic effects ofGABA-transaminase inhibitors in the treatment ofepilepsy: ocular perfusion and visual perform-ance. Ophthalmic Physiol Opt 2002; 22: 440–447.
172. Bui BV, Kalloniatis M, Vingrys AJ. Retinal functionloss after monocarboxylate transport inhibition.Invest Ophthalmol Vis Sci 2004; 45: 584–593.
173. Hendrickson AE, Koontz MA, Pourcho RG, SarthyPV, Goebel DJ. Localization of glycine-containingneurons in the Macaca monkey retina. J CompNeurol 1988; 273: 473–487.
174. Menger N, Pow DV, Wässle H. Glycinergic ama-crine cells of the rat retina. J Comp Neurol 1998;401: 34–46.
175. Pow DV, Hendrickson AE. Distribution of theglycine transporter glyt-1 in mammalian andnonmammalian retinae. Vis Neurosci 1999; 16:231–239.
176. Maxeiner S, Dedek K, Janssen-Bienhold U,Ammermuller J, Brune H, Kirsch T, Pieper Met al. Deletion of connexin45 in mouse retinalneurons disrupts the rod/cone signaling pathwaybetween AII amacrine and ON cone bipolar cellsand leads to impaired visual transmission. J Neuro-sci 2005; 25: 566–576.
177. Bringmann A, Wiedemann P. Müller glial cells inretinal disease. Ophthalmologica 2012; 227: 1–19.
178. Ward MM, Jobling AI, Kalloniatis M, Fletcher EL.Glutamate uptake in retinal glial cells during dia-betes. Diabetologia 2005; 48: 351–360.
179. Holcombe DJ, Lengefeld N, Gole GA, Barnett NL.The effects of acute intraocular pressure elevationon rat retinal glutamate transport. Acta Ophthalmol2008; 86: 408–414.
180. Otori Y, Shimada S, Tanaka K, Ishimoto I, Tano Y,Tohyama M. Marked increase in glutamate-
aspartate transporter (GLAST/GluT-1) mRNAfollowing transient retinal ischemia. Brain Res MolBrain Res 1994; 27: 310–314.
181. Pow DV, Crook DK. Rapid postmortem changes inthe cellular localisation of amino acid transmittersin the retina as assessed by immunocytochemistry.Brain Res 1994; 653: 199–209.
182. Napper GA, Pianta MJ, Kalloniatis M. Reducedglutamate uptake by retinal glial cells underischemic/hypoxic conditions. Vis Neurosci 1999;16: 149–158.
183. Chen CT, Alyahya K, Gionfriddo JR, DubielzigRR, Madl JE. Loss of glutamine synthetase immu-noreactivity from the retina in canine primaryglaucoma. Vet Ophthalmol 2008; 11: 150–157.
184. Napper GA, Pianta MJ, Kalloniatis M. Localizationof amino acid neurotransmitters following in vitroischemia and anoxia in the rat retina. Vis Neurosci2001; 18: 413–427.
185. Lombardini JB. Taurine: retinal function. BrainRes Brain Res Rev 1991; 16: 151–169.
186. Warskulat U, Heller-Stilb B, Oermann E, Zilles K,Haas H, Lang F, Haussinger D. Phenotype of thetaurine transporter knockout mouse. MethodsEnzymol 2007; 428: 439–458.
187. Young TL, Cepko CL. A role for ligand-gated ionchannels in rod photoreceptor development.Neuron 2004; 41: 867–879.
188. Tavazoie S, Hughes JD, Campbell MJ, Cho RJ,Church GM. Systematic determination of geneticnetwork architecture. Nat Genet 1999; 22: 281–285.
189. D’Haeseleer P. How does gene expression cluster-ing work? Nat Biotechnol 2005; 23: 1499–1501.
190. Marc RE, Murry RF, Basinger SF. Pattern recogni-tion of amino acid signatures in retinal neurons.J Neurosci 1995; 15: 5106–5129.
191. Marc RE, Murry RF, Fisher SK, Linberg KA, LewisGP, Kalloniatis M. Amino acid signatures in thenormal cat retina. Invest Ophthalmol Vis Sci 1998;39: 1685–1693.
192. Hill E, Kalloniatis M, Tan SS. Cellular diversity inmouse neocortex revealed by multispectral analy-sis of amino acid immunoreactivity. Cereb Cortex2001; 11: 679–690.
193. Jones BW, Watt CB, Frederick JM, Baehr W, ChenCK, Levine EM, Milam AH et al. Retinal remod-eling triggered by photoreceptor degenerations.J Comp Neurol 2003; 464: 1–16.
194. Congalton RG. A review of assessing the accuracyof classifications of remotely sensed data. RemoteSens Environ 1991; 37: 35–46.
195. Vaney DI, Young HM. GABA-like immunoreactiv-ity in cholinergic amacrine cells of the rabbitretina. Brain Res 1988; 438: 369–373.
196. Whitney IE, Keeley PW, Raven MA, Reese BE.Spatial patterning of cholinergic amacrine cellsin the mouse retina. J Comp Neurol 2008; 508:1–12.
197. Marc RE. Mapping glutamatergic drive inthe vertebrate retina with a channel-permeantorganic cation. J Comp Neurol 1999; 407: 47–64.
198. Downie LE, Pianta MJ, Vingrys AJ, Wilkinson-Berka JL, Fletcher EL. Neuronal and glial cellchanges are determined by retinal vascularizationin retinopathy of prematurity. J Comp Neurol 2007;504: 404–417.
199. Yoshikami D. Transmitter sensitivity of neuronsassayed by autoradiography. Science 1981; 212:929–930.
200. Picco C, Menini A. The permeability of the cGMP-activated channel to organic cations in retinal
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
© 2013 The Authors Clinical and Experimental Optometry 96.3 May 2013
Clinical and Experimental Optometry © 2013 Optometrists Association Australia 331

rods of the tiger salamander. J Physiol 1993; 460:741–758.
201. Dwyer TM, Adams DJ, Hille B. The permeability ofthe endplate channel to organic cations in frogmuscle. J Gen Physiol 1980; 75: 469–492.
202. Marc RE, Kalloniatis M, Jones BW. Excitationmapping with the organic cation AGB2+. VisionRes 2005; 45: 3454–3468.
203. Marc RE. Kainate activation of horizontal,bipolar, amacrine, and ganglion cells in the rabbitretina. J Comp Neurol 1999; 407: 65–76.
204. Michel WC, Steullet P, Cate HS, Burns CJ, Zhain-azarov AB, Derby CD. High-resolution functionallabeling of vertebrate and invertebrate olfactoryreceptor neurons using agmatine, a channel-permeant cation. J Neurosci Methods 1999; 90: 143–156.
205. Edwards JG, Michel WC. Odor-stimulated gluta-matergic neurotransmission in the zebrafisholfactory bulb. J Comp Neurol 2002; 454: 294–309.
206. Edwards JG, Michel WC. Pharmacological charac-terization of ionotropic glutamate receptors inthe zebrafish olfactory bulb. Neuroscience 2003;122: 1037–1047.
207. Sun D, Rait JL, Kalloniatis M. Inner retinalneurons display differential responses toN-methyl-D-aspartate receptor activation. J CompNeurol 2003; 465: 38–56.
208. Sun D, Kalloniatis M. Mapping glutamateresponses in immunocytochemically identifiedneurons of the mouse retina. J Comp Neurol 2006;494: 686–703.
209. Acosta ML, Chua J, Kalloniatis M. Functional acti-vation of glutamate ionotropic receptors in thedeveloping mouse retina. J Comp Neurol 2007; 500:923–941.
210. Kalloniatis M, Tomisich G, Wellard JW, Foster LE.Mapping photoreceptor and postreceptorallabelling patterns using a channel permeableprobe (agmatine) during development in thenormal and RCS rat retina. Vis Neurosci 2002; 19:61–70.
211. Acosta ML, Fletcher EL, Azizoglu S, Foster LE,Farber DB, Kalloniatis M. Early markers of retinaldegeneration in rd/rd mice. Mol Vis 2005; 11:717–728.
212. Yu TY, Acosta ML, Ready S, Cheong YL, Kalloni-atis M. Light exposure causes functional changesin the retina: increased photoreceptor cationchannel permeability, photoreceptor apoptosis,and altered retinal metabolic function. J Neuro-chem 2007; 103: 714–724.
213. Acosta ML, Bumsted O’Brien KM, Tan SS,Kalloniatis M. Emergence of cellular markers andfunctional ionotropic glutamate receptors on tan-gentially dispersed cells in the developing mouseretina. J Comp Neurol 2008; 506: 506–523.
214. Chang YC, Chiao CC. Localization and functionalmapping of AMPA receptor subunits in the devel-oping rabbit retina. Invest Ophthalmol Vis Sci 2008;49: 5619–5628.
215. Chang YC, Chen CY, Chiao CC. Visual experience-independent functional expression of NMDAreceptors in the developing rabbit retina. InvestOphthalmol Vis Sci 2010; 51: 2744–2754.
216. Casson RJ. Possible role of excitotoxicity in thepathogenesis of glaucoma. Clin Experiment Oph-thalmol 2006; 34: 54–63.
217. Osborne NN, Casson RJ, Wood JP, Chidlow G,Graham M, Melena J. Retinal ischemia: mecha-nisms of damage and potential therapeutic strat-egies. Prog Retin Eye Res 2004; 23: 91–147.
218. Naskar R, Vorwerk CK, Dreyer EB. Concurrentdownregulation of a glutamate transporter andreceptor in glaucoma. Invest Ophthalmol Vis Sci2000; 41: 1940–1944.
219. Lai JS, Tham CC, Chan JC, Yip NK, Tang WW, LiPS, Yeung JC, Lam DS. Scanning laser polarimetryin patients with acute attack of primary angleclosure. Jpn J Ophthalmol 2003; 47: 543–547.
220. Resta V, Novelli E, Vozzi G, Scarpa C, Caleo M,Ahluwalia A, Solini A et al. Acute retinal ganglioncell injury caused by intraocular pressure spikes ismediated by endogenous extracellular ATP. Eur JNeurosci 2007; 25: 2741–2754.
221. Lafuente MP, Villegas-Perez MP, Selles-Navarro I,Mayor-Torroglosa S, Miralles de Imperial J, Vidal-Sanz M. Retinal ganglion cell death after acuteretinal ischemia is an ongoing process whoseseverity and duration depends on the duration ofthe insult. Neuroscience 2002; 109: 157–168.
222. Osborne NN, Larsen AK. Antigens associated withspecific retinal cells are affected by ischaemiacaused by raised intraocular pressure: effect ofglutamate antagonists. Neurochem Int 1996; 29:263–270.
223. Dijk F, Kamphuis W. Ischemia-induced altera-tions of AMPA-type glutamate receptor subunit.Expression patterns in the rat retina–an immuno-cytochemical study. Brain Res 2004; 997: 207–221.
224. Gargini C, Terzibasi E, Mazzoni F, Strettoi E.Retinal organization in the retinal degeneration10 (rd10) mutant mouse: a morphological andERG study. J Comp Neurol 2007; 500: 222–238.
225. Fletcher EL, Kalloniatis M. Neurochemical devel-opment of the degenerating rat retina. J CompNeurol 1997; 388: 1–22.
226. Fletcher EL. Alterations in neurochemistryduring retinal degeneration. Microsc Res Tech2000; 50: 89–102.
227. Barhoum R, Martinez-Navarrete G, CorrochanoS, Germain F, Fernandez-Sanchez L, de la Rosa EJ,de la Villa P et al. Functional and structural modi-fications during retinal degeneration in the rd10mouse. Neuroscience 2008; 155: 698–713.
228. Varela C, Igartua I, De la Rosa EJ, De la Villa P.Functional modifications in rod bipolar cells in amouse model of retinitis pigmentosa. Vision Res2003; 43: 879–885.
229. Margolis DJ, Newkirk G, Euler T, Detwiler PB.Functional stability of retinal ganglion cells afterdegeneration-induced changes in synaptic input.J Neurosci 2008; 28: 6526–6536.
230. Mazzoni F, Novelli E, Strettoi E. Retinal ganglioncells survive and maintain normal dendriticmorphology in a mouse model of inherited pho-toreceptor degeneration. J Neurosci 2008; 28:14282–14292.
231. Stasheff SF. Emergence of sustained spontaneoushyperactivity and temporary preservation of OFFresponses in ganglion cells of the retinal degen-eration (rd1) mouse. J Neurophysiol 2008; 99:1408–1421.
232. Kamboj S, Huganir RL. Receptor clustering: acti-vate to accumulate? Curr Biol 1998; 8: R719–721.
233. Goebel DJ, Poosch MS. Transient down-regulation of NMDA receptor subunit geneexpression in the rat retina following NMDA-induced neurotoxicity is attenuated in the pres-ence of the non-competitive NMDA receptorantagonist MK-801. Exp Eye Res 2001; 72: 547–558.
234. Puthussery T, Gayet-Primo J, Pandey S, DuvoisinRM, Taylor WR. Differential loss and preservationof glutamate receptor function in bipolar cells in
the rd10 mouse model of retinitis pigmentosa. EurJ Neurosci 2009; 29: 1533–1542.
235. Cantallops I, Haas K, Cline HT. PostsynapticCPG15 promotes synaptic maturation and presy-naptic axon arbor elaboration in vivo. Nat Neurosci2000; 3: 1004–1011.
236. Xia Y, Carroll RC, Nawy S. State-dependent AMPAreceptor trafficking in the mammalian retina.J Neurosci 2006; 26: 5028–5036.
237. Guenther E, Schmid S, Wheeler-Schilling T,Albach G, Grunder T, Fauser S, Kohler K. Devel-opmental plasticity of NMDA receptor function inthe retina and the influence of light. FASEB J 2004;18: 1433–1435.
238. Kamphuis W, Dijk F, O’Brien BJ. Gene expressionof AMPA-type glutamate receptor subunits in rod-type ON bipolar cells of rat retina. Eur J Neurosci2003; 18: 1085–1092.
239. Marc RE, Cameron D. A molecular phenotypeatlas of the zebrafish retina. J Neurocytol 2001; 30:593–654.
240. Lim J, Lam YC, Kistler J, Donaldson PJ. Molecularcharacterization of the cystine/glutamateexchanger and the excitatory amino acid trans-porters in the rat lens. Invest Ophthalmol Vis Sci2005; 46: 2869–2877.
241. Lim J, Lorentzen KA, Kistler J, Donaldson PJ.Molecular identification and characterisationof the glycine transporter (GLYT1) and theglutamine/glutamate transporter (ASCT2) inthe rat lens. Exp Eye Res 2006; 83: 447–455.
242. Hu RG, Zhu Y, Donaldson PJ, Kalloniatis M.Alterations of glutamate, glutamine and relatedamino acids in the anterior eye secondary toischaemia and reperfusion. Curr Eye Res 2012; 37:633–643.
243. Coca-Prados M, Escribano J. New perspectives inaqueous humor secretion and in glaucoma: theciliary body as a multifunctional neuroendocrinegland. Prog Retin Eye Res 2007; 26: 239–262.
SUPPORTING INFORMATION
Additional Supporting Information may befound in the online version of this article:
Supplementary Figure 1. Phylogenetic treeof neopterygii a subclass of highly successfulfishes that includes goldfish and catfish.Supplementary Figure 2. Phylogenetic treeof Lissamphibia a subclass that includesrecent amphibians. The subclass includestoads, frogs, tiger salamander and newts.Supplementary Figure 3. Phylogenetic treeof aves, jawless fish (cephalospidomorphi:sea lamprey) and the cartilaginous fishes(chondrichthyes: skates and sharks).Supplementary Figure 4. Phylogenetic treeof holocephali (a taxon of cartilaginousfish—ratfish), reptiles including lizards,snakes and turtle, sarcopterygii (lungfish),sauropsida (crocodiles) and an arthropod(barnacle).Supplementary Figure 5. Phylogenetic treefor mammals including primates, carni-vores, lagomorphs, rodents, pigs/antelopes,marsupials and the prototheria (echidna).
Retinal amino acids in health and disease Kalloniatis, Loh, Acosta, Tomisich, Zhu, Nivison-Smith, Fletcher, Chua, Sun and Arunthavasothy
Clinical and Experimental Optometry 96.3 May 2013 © 2013 The Authors
332 Clinical and Experimental Optometry © 2013 Optometrists Association Australia




















![[MDMA]MDMA Neurochemistry](https://img.dokumen.tips/doc/110x75/577dab601a28ab223f8c57f3/mdmamdma-neurochemistry.jpg)








