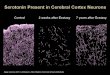
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
1/9
/_! Raven _ Ltd., New York1990 International Society for
Ncurochemistry
ShortReview
Neurochemistry and Neurotoxicityof
3,4-Methylenedioxymethamphetamine (MDMA, "Ecstasy")
,6 DennisJ. McKennaandStephenJ.Peroutka.xxt Department of
Neurology, Stanford University School of Medicine, Stanford,
California, U.S,A.
[_ 3,4-Methylenedioxymethamphetamine (MDMA; acutely, but may
have long-term consequences; others,' also known as "Ecstasy") is a
ting-substituted phenyl- such as the rapid depletion of brain
hydroxyindolesJ isopropylamine that is related to both amphetamines
within hours following MDMA, appear to be unrelated
and hallucinogens, such as mescaline (Fig. 1).Although to the
slower, more persistent decreases in brain hy-the drug was patented
in 1914, interest in the com. droxyindoles that are among the
long-term neuro-pound was minimal until the past decade. During
this chemical changes that are indicative of neurotoxicity.f'
period, MDMA began to be used as an adjunct to psy- Table 1
summarizes the acute and long-term effects ofchotherapy by certain
therapists due to its purported MDMA. "
q/_ ability to induce a state of reduced anxiety and
lowereddefensiveness (Downing, 1986; Greer and Tolbert,_4 1986). In
addition, the recreational use of MDMA, ACUTE NEUROCHEMICAL
EFFECTSparticularly on college campuses, appears to have in-[
creased significantly in recent years (Peroutka, 1987). Brain
hydroxyindolesThe human use of this agent is of concern due to
the
fact that MDMA and some of its congeners are selective One of
the most characteristic acute effects oferotonergic neurotoxins in
laboratory animals. MDMA is a rapid and pronounced decrease in
brainlevels o f 5-hydroxytryptam ine (5-H T) and 5-hydroxy-J Partly
as result of the recem interest in MDMA, nu- indoleacetic acid
(5-HIAA). This effect is thought tomerous investigators have begun
to explore the neu-
rochemical effects of MDMA. During the past 3 years, result from
the massive release of 5-HT from presyn-iB over 80 publications on
MDMA have appeared in the aptic vesicles. Reductions in brain
hydroxyindole con-scientific literature. The following review
attempts to centrations ofgreater than 80% by 3 h after
injectionofMDMAhavebeenreportedSchmidtetal.,1986provide a brief
summary of recent data on the neu-o_ rochemistry and neurotoxicity
of MDMA and its de- 1987; Stone et al., 1986, 1987b; Schmidt,
1987b). Res-ivatives, toration ofbrain concentrationsof
hydroxyindolesoAdministration of MDMA to animals elicits a char-
control levels after a single acute dose of MDMA is_q acteristic
biphasic response which, for convenience of rapid, usually
occurring within 24 h post injection
_; discussion, can be categorized into acute and long-term
(Schmidt, 1987b; Stone et al., 1987b; Schmidt and[ effects. Some of
the acute effects of MDMA appear to Taylor, 1989).be related more
directly to its behaviora l and psycho-l _ logical effects, whereas
the long-term effects have been TPH activityl'yJt_ correlated with
the development of serotonergic neu- The reduction in brain 5-HT
and 5-HIAA is accom-rotoxicity. It should be noted, however, that
the acute panied by a marked reduction in TPH activity. and
long-term effects cannot be considered in isolation Whereas the
hydroxyindoles are restored to controlfrom one another. Some of the
effects, e.g., reduction values within 24 h after a single
injection of MDMA.s_ in tryptophan hydroxylase (TPH) activity,
manifest TPH activity remains significantly reduced in the
stria-r-'/_ Address correspondence and reprint requests to Dr. S.
J. Peroutka hydroxyindoleacetic acid; 5-HT, 5-hydroxytryptamine;
MDA, 3,4-at Department of Neurology, Stanford University School of
Medicine, methylenedioxyamphetamine; MDE,
3,4-methylenedioxy-N-ethyl-! Stanford, CA 94305, U,S.A.
amphetamine; MDMA, 3,4-methylenedioxymethamphetamine; NE.
Abbreviations used: AMT, a-methyl-p-tyrosine; DA_, dopamine;
norepinephrine; PCA, p-chloroamphetamine; TH, tyrosine hydrox-DOM,
2,5-dimethoxy4-methylphenylisopropylamine; 5-H1AA, 5- ylase; TPH,
tryptophan hydroxylase.
c] 14
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
2/9
3,4.METHYLENEDIOXYMETHAMPHETAMINE 15
O_.__NHR' results partly from the ability of the drug to
modulate1_ _ 5-HT release and uptake. Differential effects of the
ste-reoisomersofMDA,MDMA,and someof their ana-logues on both
release and uptake of [3H]5-HT and
R R' [3H]DAhavebeen noted (Nicholsetal., 1982;Johnsonet al.,
1986; Schmidt, 1987a; Schmidt et at., 1987;MDMA CH3 CH3 Steele et
al., 1987). With respect to release from syn-MDA CH3 H
aptosomes(Nicholsetal., 1982)orsuperfusedbrainslices (Johnson et
al., 1986; Schmidt et al., 1987),MDE CH3 CH2CH3 MDMA and its
analogues are more potent releasers ofBDB CH2CH3 H [3H]5-HT than of
[3H]DA. Stereospecific effects onMBDB CH2CH 3 CH3 [3H]5-HT release
are generally not apparent, but dif-ferential effects on DA release
have been noted. For
FIG. 1. MDMA and its analogues.BDB, 1-(1,3-benzodioxol-f-yl)-
example, Johnson et al. (1986) reported that
racemic2-butanamine;MBDB,N-methyl-l-(1,3-benzod{oxol-S-yl)-2-butan-
MDA induced a significantly greater effect than
racemicamine;MOA,3,4-methylenedioxyamphetamine; MDE, 3,4-methyl-
MDMA, but no differences were found between
S(+)-enedioxy-N-ethylamphetamine;DMA,3,4-methylenedioxymeth- MDA
and S(+)-MDMA, or between R(-)-MDA andamphetamine, R(-)-MDMA. These
investigators speculated that thes,
existenceofcompetitivemechanismsfortheRandStum, hippocampus, and
cortex for 2 weeks or longer enantiomers of MDMA would account for
these re-
(Schmidt and Taylor, 1987; Stone et al., 1987b). This suits.
Ethylation of the side-chain nitrogen with ethyl- effect apparently
is not due to direct inhibition of the to give the analogue
3,4-methylenedioxy-N-ethylam-enzyme, because MDMA exhibits no
effect on TPH phetamine (MDE) results in a lowered potency for.
activity in vitro (Schmidt and Taylor, 1987). The Vms, [3H]DA
release compared to MDA and MDMA, butf ofTPH is reduced by 50% in
treated animals, whereas all three drugs are still potent releasers
of [3H]5-HT
the affinity for either of its substrates, tryptophan (Schmidt,
1987a). Pretreatment with the selective DAor the hydroxylase
cofactor, 6-methyl-5,6,7,8-tetra- uptake inhibitor amfonelic acid
or with the selectivehydopterine, is unchanged (Schmidt and Taylor,
1987, 5-HT uptake inhibitor, citalopram, blocks the MDMA-1989).
Simultaneous administration of the 5-HT up- induced release of
[3H]DA or [3H]5-HT in vitro.take inhibitor, citalopram, with MDMA
completely Methamphetamine and p-chloroamphetamine (PCA)prevents
the acute depletion of cortical hydroxyindoles, are significantly
more potent releasers of [3H]DA thanbut only slightly attenuates
the reduction in cortical MDMA, but MDMA and PCA are equipment and
sig-f TPH activity. However, fluoxetine, given 3 h post nificantly
more potent than methamphetamine asMDMA does facilitate the
recovery of TPH activity [3H]5-HT releasers (Schmidt et al.,
1987).
- levels to control values within I week (Schmidt and The
effects of the enantiomers of MDMA and itsTaylor, 1987, 1989).
analogues on the synaptosomal uptake of 3H-mono-- amines (5-HT, DA,
and NE) have been compared withChanges in dopamine and other
monoamines those of the isomers of amphetamine and the hallu-In
contrast to the rapid and marked reductions in cinogenic
phenylisopropylamine, 2,5-dimethoxy-4-
, 5-HT and its metabolite, 5-HIAA, similar changes in
methylphenylisopropylamine (DOM) (Steele et al.,levels or turnover
ofdopamine (DA) or norepinephrine 1987). Both enantiomers of MDA
and MDMA po-(NE) in response to acute treatment with MDMA haves not
been observed. Schmidt et al. (1986) reported aslight, but
significant, rise in neostriatal concentrations TABLE 1. Summary of
the neurochemicaleffects of MDMAin DA and its metabolite,
homovanillic acid, at 3 hfollowing administration of MDMA. Levels
of dihy- Acuteeffects(36h)elicits a dose-dependent release of DA in
the nucleus Persistentslowdecreasein 5-HT and
5-HIAAfollowingnitialaccumbens and caudate of awake-behaving rats,
mca- recovery
- sured by in vivo voltammetry and HPLC.
DepressedTPHactivitypersistsDecrease in 5-HT terminal densityl-
Effects on neurotransmitter release and uptake Marked species
variations (mice < rats < primates), Changes are prevented by
5-HT uptake blockersThe acute effects observed on brain monoamines
Slowrecovery(i.e. , months) of someparametersand their metabolites
following treatment with MDMA
J. Neurochern. Vol. 54.No. 1, 1990
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
3/9
_ 16 D. J. McKENNA AND $. J. PEROUTKA-_ tently inhibited the
uptake of [3H]5-HT, but only the TABLE 2. MDMA interactionswith--_
S(+) enantiomers inhibited [3H]DA uptake. By con-
neurotransmitterreceptorstrast, both enantiomers of amphetamine
were more Neurotransmitter _ v_ potent inhibitors of [3H]DA uptake
compared to that bindingsite Radioligand (gf [3H]5-HT. Th e
stereoisomers of th e M DMA-related
compounds and amphetamine were equipotent inhib-
Serotonergictors of [3H]NE uptake into hypothalamic synapto- Uptake
[3H]Paroxetine 0.6somes. Neither isomer of DOM inhibited
synaptosomal 5-HT2A [7?BrlDOB 1.5[ uptake of any of the monoamines
examined. 5-HT_A [3H]8-OH-DPAT 10 +Both Schmidt et al. (1987) and
Wang et al. (1987) 5-HTm [3H]5-HT 10 +
E found little evidence for carder-mediated uptake of 5-HT2
[_HlSpiperone 20 _[3H]MDMA into synaptosomes, suggesting that
Adrene_,ic
'-_ MDMA probably enters the neuron by passive mech- tl 2
[3H]Aminoclonidine 3.6 +anisms where it causes the displacement and
release of t_ [3H]Dihydroalprenolol 19 +al [_H]Prazosin 20 +c,x_
monoamines; this release, however, is carrier-mediated/" and
blocked by uptake inhibitors. DopamineL D2 {3H]Spiperone 95 +_
Receptor interactions D, [3HISCH23990 150+It is presently unclear
whether the psychoactive, as MuscariniccholinergicM_ [3H](-)-QNB
5.8 +opposed to the neurotoxic, effects of MDMA and its M2
[3H](-)-QNB 15_glil congeners result from the presynaptic release
of 5-HT_ from serotonergic terminals, or from interactions with
Opioidt9 postsynaptic target receptors, particularly 5-HT recep- _'
[3H]Dihydromorphine >5tors. Nichols (1986) has argued in favor
ofa presynaptic b [3HlD-Ala,D-Leu-enkephalin >5K
[3HlEthylketazocine > 5[' mechanism, based on the fact that the
S(+) enantiomer
ofMDMA ismore potent than the R(-) enantiomer, Othersites/_
which is the opposite of what would be expected if a HistamineH_
[3H]Mepyramine 6 -+Benzodiazepine 13H]Flunitrazepam
>50postsynaptic interaction similar to that involving DOM CRF
_2_I-CRF >5,q4 or other 5-HT2-selective hallucinogens were
respon- Ca:+ channels [3HlNitrendipine >5,_" sible for the
psychotropic effects. Conceivably, thenique psychotropic
effectsofMDMA may result from Dataare derived from Pierce and
Peroutka (1988) and Bat
a combination of pre- and postsynaptic actions. A et al.
(1988a). CRF, corticotropin releasing factor; DOB,
2,5-dthoxy-4-bromophenylisopropylamine; 8-OH-DPAT, 8-hydroxy-2umber
of investigators have examined the receptor n-propylamino)tetralin;
QNB, quinuclidinyl benzilate.
selectivities and affinities of MDMA and its derivatives *These
are I_ovalues in human cortex; KD values for these rusing
radioligand binding assays (see summary in Ta- ligandshavenot
beendeterminedin human tissue.ble 2).13 Lyon and co-workers (1986)
examined the affinities, ofracemic MDA and MDMA and their optical
isomers lng site labeled by R(-)-2,5-dimethoxy-4-[77Br]bromo-[ at
5-HT2, 5-HTr, and DA D2 sites. None of the deriv-
phenylisopropylamine (Pierce and Peroutka, 19tives showed
significant affinity for the DA D2 site. MDA displayed moderate
(< 10 taM) affinity for th4 The most potent compound was
R(-)-MDMA at the HT_A, 5-HTm, and a2-adrenergic sites. Affinity-HT2
site labeled with [3H]ketanserin (3.3 taM), al- MDA and the other
derivatives was greater than,_ though this compound was slightly
less potent (4.2 _1_/) tam at all other sites measured.
at 5-HT_ sites labeled by [3H]5-HT. Racemic MDMA Gehlert and
co-workers (1985) reported the spec[ and S(+)-MDMA were less potent
at both sites. The binding of [3H]MDMA to rat cortical membranes,
wpotency of MDA and its stereoisomers was comparable an apparent KD
of 99 nM and Bm_ of 30 fmol/mgl_ to that of MDMA at both sites,
protein. These findings were questioned, however,Battaglia et al.
(1988a) screened racemic MDMA at Wang et al. (1987) who reported on
"specific" bind. a variety ofneurotransmitter recognition sites and
up- of [3H]MDMA to glass fiber filters in the absence
take sites. MDMA showed the highest affinity overall brain
membranes. The artifactual binding yielded for the
[3H]paroxetine-labeled 5-HT uptake site (0.61 parent Kt>and B_
values similar to those determinet_ taM), followed by the
a2-adrenergic receptor (3.6 taM), in the presence of membrane
homogenates and
the 5-HT2 receptor (5.1 taM), the histamine H_ receptor played a
distinct pharmacological profile, with ri[ Mz receptor (5.8 #M).
substituted amphetamines and 5-HI uptake inhibit(5.7 taM), an d th
e muscarinicThe potency at all other sites examined was 10 tzM or
displacing apparent specific binding with the grea'_q greater,
potency. Pretreatment of the glass fiber filters with 0
Pierce and Peroutka (1988) reported that racemic
polyethylenimine completely eliminated "specifi,q.x MDA, MDMA, and
MDE displayed highest affinity [3H]MDMA binding in both the
presence and abseni (approximately I /_M) for the putative 5-HT2A
bind- ofbrain tissue.
J. Neurochern.,Vol. 54, NO. 1990I,
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
4/9
3,4-METHYLENEDIOXYMETHAMPHETAMINE 17Neuroendocrine responses
tential neurotoxicity of MDMA and its analogues.Nash et al. (1988)
reported that intraperitoneal ad- Schmidt and Taylor (1987) found
that cortical TPHministration of MDMA resulted in a rapid elevation
activity remained significantly depressed as long as 7of serum
corticosterone and prolactin levels, and sig- days after injection
of MDMA. These authors specu-nificantly elevated body temperature.
Both the eleva- lated that the relatively long period required for
recov-tion ofcorticosterone levels and the hyperthermia were ery of
TPH activity ind icated an irreversible inacti-5 blocked by the
5-HT2 antagonists, ketanserin and vation of the enzyme, requiring
synthesis of new en-' mianserin, but not by 5-HTr^ antagonists or
nonse- zyme for full restoration ofactivity. Stone et al.
(1987b)lective 5-HT antagonists. These data suggest that reported
significant and long-term reductions of TPHMDMA induces
hyperthermia and corticosterone se- activity in the neostriatum,
hippocampus, and hypo-cretion via a 5-HT2 receptor-mediated
mechanism (ei- thalamus, as well as the cortex.th er d irect o r
ind irect). Th e effect o n pro lactin secre-1 tion, however,
appears to be mediated by a 5-HT re- 5-HT uptake site densityceptor
different from both the 5-HTr^ and 5-HT2 In addition to long-term
changes in neurochemicalreceptors. Evidence that the effect is
mediated via a 5- parameters, M DMA also causes pronounced reduc-HT
receptor was provided by the fact that prior deple- tions in the
density of 5-HT uptake sites and histolog-tion of 5-HT by
p-chlorophenylalanine abolished the ical changes in the
distribution and density of seroto-effectof MDMA on prolactin
secretion. None of the nergic nerve fibers. Both of these
parameters are in-5-HT antagonists blocked the M DMA-induced ele-
dicative ofa neurodegenerative process. Battaglia et al.vation of
prolactin, indicating that neither the 5-HTr^ (1987) reported a
50-75% reduction in the density ofnor the 5-HT2 receptor mediated
the response. In a [3H]paroxetine-labeled 5-HT uptake sites in the
cortex,separate study , M erchant et al. (1 98 7) reported that h
ippocampus, striatum, hypothalamus, and midbrainmultiple doses of
MDA, MDMA, and methamphet- at 2weeks following repeated systemic
administrationamine significantly elevated neurotensin-like immu-
of MDMA and MDA. Scatchard analyses ofthese datanoreactivity 18h
after treatment in the substantia nigra, indicated a significant
reduction in the Bmax values instria tum, and nucleus accumbens of
rat brain , the treated an imals with no significant altera tion in
theKDvalues. In a subsequent study, in vitro autoradi-LONG-TERM
EFFECTS ography with [3H]paroxetine was used to determinefurther
the neuroanatomical localization of MDA-in-Brain 5-HT and 5-HIAA
levels duced lesions (De Souza and Kuyatt, 1987). They re-Long-term
reductions in brain levels of 5-HT and ported greater than 70%
reductions in [3H]paroxetinelia 5-HIAA induced by M DM A can be
read ily quantifiede- binding in the frontal cortex, pyriform
cortex, and ol-di- with HPLC and areamong the indices
ofneurotoxicity factory tubercles. Similar observations have been
madethat have received the most attention . Long-term de- when the
reduction in synaptosomal uptake of [3H]5-- pletions of 5-HT and
its metabolite by MDMA were HT in MDMA-treated animals is used as a
means offirst reported by Schmidt eta l. (1986), and Ricaurte et
quantifying the reduction of 5-HT uptake sites (Ri-al. (1985)had
noted a similar action of MDA. In sub- caurte et al., 1 98 5;
Commins et al., 1 98 7; Schmidt,sequent studies (Schmidt, 1987b;
Schmidt et al., 1987), 1987b).S(+)-MDMA was significantly more
potent than R(-)-. MDMA in producing long-term 5-HT depletion, but
Histopathology- the reverse relationship was found for the acute
deple-f tion. Since these initial reports, other investigators have
A variety of histological techniques have been usedconfirmed that
MDMA can induce significant, long- to visualize the
neurodegenerative changes induced byM DMA. Commins et al. (1 98 7)
and Ricaurte et al.term, and regionally specific reductions in 5-HT
and (1985) utilized the Fink-Heimer staining method toc 5-HIAA,
which in some instances can persist for Up to12 months following
repeated administration (Mokler localize degenerating nerve
terminals in the striatumand somatosensory cortex of rats following
acute or 'f et al., 1987; Stone et al., 1987b; Battaglia etal.,
1988b). chronic treatments with MDA or MDMA. O'Hearny Both Mokler
et al. (1987) and Stone et al. (1987b) re- and co-workers (1988)
utilized 5-HT specific immu-g ported that serotonergic neurons in
the frontal cortex, nocytochemistry to document the extensive loss
of 5-
f hippocampus, and striatum weremore sensitiveto the HT axons
2weeksafter systemictreatment withMDA- effect ofMDMA than those in
the hypothalamus. Theyd suggested that the h igher density o f
fibers of passage or MDMA. They reported a selective vulnerability
of- and cell bodies in the hypothalamus might account for 5-HT axon
terminals originating primarily in the dorsalraph e nucleus, wh
ereas beaded ax ons h aving large var-- its reduced sensitivity.rs
icositiesndoriginatingrimarilynthemedianapheest TPH activity
nucleiwerelargelyspared.Preterminalaxons,fibersofLong-term
depression of TPH activity typically ac- passage, and raphe cell
bodies were not damaged (Mol-c" companies the reduction in brain
hydroxyindoles fol- liver, 1987;O'Hearn et al., 1988). Wilson et
al. (1989)e lowing MDMA. Most investigators regard this as an
reported a similar differential vulnerability to MDMA
additional biochemical marker indicative of the po- of dorsal
raphe 5-HT axons in the macaque monkey.J. Neurochem., Vol. 54, No.
1, 1990
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
5/9
18 D. J. McKENNA AND S. J. PEROUTKA
q'_ FACTORS GOVERNING THE rats and mice to single and multiple
injections of_ DEVELOPMENT OF NEUROTOXICITY MDMA. They reported
that although a single 10 mg/kg dose in rats produced both an acute
and long-termDosage and treatment regimen depression of TPH
activity, 5-HT, and 5-HIAA, similarlong-term effects were not
observed in mice after a sin-oth the acute and long-term
neurochemical effects
of MDMA are influenced by the dosage used and by gte 15 mg/kg
dose. Brain hydroxyindoles and TPH ac-he treatment regimen (i.e.,
whether single or multiple tivity were significantly reduced,
however, in micedoses are administered). The acute neurochemical
el- given multiple injections of MDMA (6 X 15 mg/kg).fects of MDMA
(cf. Table 1) can result from a single These authors attributed the
greater sensitivity of ratsdose of approximately 5-10 mg/kg
(Schmidt and Tay- to metabolic differences between the two species.
In
[ lor, 1987; Battaglia et al., 1988b). By contrast, long- rats,
the primary metabolic route is via ring para-hy-erm serotonergic
deficits develop in response to either droxylafion, which could be
impaired due to the pres-a single administration of a comparatively
large dose(20 rog/kg) or repeated, frequent administration of ence
of the methylenedioxy substituent. In mice, how-ever, side-chain
deam ination and ring hydroxy lation
4 smaller doses (8 5 mg/kg) (Battaglia et al., 1988b, metabolic
and thus thear e equally important routes,r 1989). Therefore, the
degree of MDMA-induced neu- drug could be eliminated more quickly
from brain. In_. rotoxicity is a cumulative, dose-related
phenomenon, a similar study, Peroutka (1988) measured the acuteand
long-term effects of single (1, 3, 10, or 30 mg/kg)_ Route of
administration and multiple (8 1, 3, 10, or 30 mg/kg) injections
of
Oral ingestion is the most common route of admin- MDMA in mice
using [3H]paroxetine to label 5-HTi istration used by human
consumers of MDMA. Ani- uptake sites. No significant changes in
either the KD or,_ mai studies, by contrast, routinely employ
subcuta- Bmr, values of [3H]paroxetine binding compared toneous or
in trape ritoneal in jections. D ifferen tial effects contro ls
were found w ith any of the dose regimens used.o' of the route of
administration on MDMA-mediated By contrast, rats administered a
single injection ofneurotoxicity have been examined by several
investi- MDMA (30 mg/kg) showed a 44% reduction in B_,ators
(Finnegan et al., 1988; Ricaurte et al., 1988a; with respect to
controls, with no change in KD. These
q% Slikker et al., 1988). Finnegan et al. (1988) found no r esul
ts d emonst ra te d that mice ar e relatively
insensitivesignificant difference in the dose-dependent reduction
to the neurotoxic effects of MDMA.4 in 5-HT concentrations in rats
administered MDMA Primates are even more sensitive to MDMA than
orally compared to rats treated subcutaneously. Ri- rats, by
both the oral and subcutaneous routes of ad-caurte et al. (1988a),
however, reported that in the ministration (Ricaurte et al., 1988a;
Slikker et al.,squirrel monkey, oral administration of multiple
doses 1988). In one important study, Ricaurte and co-work-of MDMA
(8 5 mg/kg)was approximately one-third ers (1988b) measured 60%
lower levels of the 5'HTto one-half as effective as subcutaneous
administration, metabolite 5-HIAA in CSF of squirrel monkeys
treatedIn additional experiments, multiple oral doses pro- with
toxic doses of MDMA (8 5 mg/kg). Brain levelsuced significant
reductions in 5-HT levels in all brain of 5-HT and 5-HIAA in the
same animals were 94 toregions, but single oral doses resulted in
significant re- 73% lower than the saline controls. These results
in-3 ductions only in the hypothalamus and thalamus, dicate that
the quantitative assay of CSF5 -HIAA levels
[ Slikker et al. (I 988)assessed long-term neurochemical in
human users of MDMA might be used as a diag-nd histological changes
in the rat given large oral doses nostic tool to assess
serotonergic damage.of MDMA. Histological studies show a 90%
increase4 in silver-stained terminals (indicative of terminal de-
Stereochemical and structure/activity parameters, generation) at 18
h after a single 40 mg/kg dose of Stereochemical and structural
parameters appear to,xl, MDMA, and a 105% increase after a single
80 mg/kg be important determinants of neurotoxicity forose. In the
same study, these workers reported sig- MDMA and its analogues.
Schmidt (1987a) observed[ nificant (70-80%) reductions in 5-HT and
5-HIAA that both isomers of MDA produced long-term deple-
levels in the frontal cortex and hippocampus after tions of
cortical 5-HT. However, the same study re-l_ multiple oral doses of
MDMA (8 40 or 8 80 mg/ ported that although racemic MDA, MDMA, andi
_ kg over 4 days). Nonsignificant reductions were also MDE all
caused acute depletion of cortical 5-HT, onlydetected in the
caudate nucleus, racemic MDA and MDMA caused a long-term deple-t'
tion of 5-HT and loss of 5-HT uptake sites. Schmidtta Species
differences (1987b) demonstrated that both enantiomers of
Mammalian species 'vary in their sensitivity to the MDMA caused
an acute depletion of cortical 5-HT,[' neurotoxic effects of MDMA.
Differential species sen- but only S(+)-MDMA resulted in
significant depletions
sitivity has been noted in rats, guinea pigs, mice, rhesus of
5-HT and 5-HT uptake sites 7 days after injection,monkeys, and
squirrel monkeys (Stone et al., 1987c, Johnson et al. (1988b)
reported that the S(+) isomers_,,_ Battaglia et al., 1988b;
Finnegan et al., 1988; Peroutka, of both MDA and MDMA were more
potent than theq-_ 1988; Ricaurte et al., 1988a; Slikker et al.,
1988). Stone R(-) isomers in depleting 5-HT and 5-HIAA and in_, and
co-workers (1987c) compared the sensitivities of depressing TPH
activity in cortex, hippocampus, and
i'
J. Neurochern., Vol.34, No. I, 1990 _ '
__
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
6/9
J
3,4-METHYLENEDIOXYMETHAMPHETAMINE 19f neostriatum, in rats
killed 18 h following multiple raphe system represents one possible
mode of recovery.treatments (5 X 3.5, 5, or 10 mg/kg, s.c.). On the
other hand, MDMA is toxic to the terminals
Several independent studies (Johnson et al., 1987; of 5-HT
neurons while sparing the cell bodies, so re-r Ricaurte et al.,
1987; Stone et al., 1987a)haveprovided generation of the terminal
from the affected neuronsevidence that MDE, the N-ethyl congener of
MDMA, is also possible.- is significantly less toxic in terms of
its effects on long-term markers for neurotoxicity. The acute
effects of Prevention of neurotoxieity
. MDE are similar to those of MDMA and include tran- Both the
acute and long-term neurochemical deficitssient reductions of 5-HT,
5-HIAA, and TPH (Johnson produced by MDMA or MDA can be blocked
corn-et al., 1987; Stone et al., 1987a). However, recovery pletely
by co-administration of a selective 5-HT uptake- from MDE appears
to be more rapid than with blocker, such as fluoxetine or
citalopram (Schmidt,- MDMA, because neurochemical parameters in
MDE- 1987b; Schmidt and Taylor, 1987; Schmidt et al.,- treated
animals are not significantly different from 1987). For example,
rats given a single dose of fluox-those in control animals at 1-2
weeks following treat- etine (5 mg/kg) at 3 or 6 h after MDMA
displayed ament (Stone et al., 1987a). Ricaurte et al. (1987) found
reduction in the depletion of cortical 5-HT levelsmea-that
approximately fourfold higher doses of MDE than sured 7 days later
(Schmidt, 1987b). Fluoxetine corn-MDMA are required to produce
comparable long-term pletely antagonized 5-HT depletion in animals
treated) depletions of 5-HT. 3 h after MDMA. At 6 h after MDMA,
5-HT levels inf Neurotoxicity to monoaminergic systems is also a
fluoxetine-treated animals were 72% of controls, andproperty of
several other amphetamine derivatives at 12 h, 5-HT values were not
different from those ofr (Fuller et al., 1975; Sanders-Bush et al.,
1975; Sanders- animals given MDMA alone (39% of controls). This
Bushand Sterenka, 1978;Fuller and Hemrick-Luecke, antagonistic
effect of a 5-HT uptake inhibitor on the. 19824Peat et al., 1985;
Schmidt et al., 1985). MDMA, long-term 5-HT depletion, even up to 6
h post MDMA,f fenfluramine, and PCA are similar in that their neu-
suggests that the neurotoxic action of MDMA mightmtoxic action
primari y affects the serotonergic system, involve its
extracellular conversion to a toxic metab-whereas methamphetamine
and amphetamine neu- olite, which (unlike MDMA itself) has
significant af-mtoxicityischaracterized by long-term deficitsin
both finity for the 5-HT uptake carrier. Schmidt and Taylor5-HT and
DA neuronal systems (Clineschmidt et al., (1987) also found that
fluoxetine given 3 h after1978;Peat et al., 1985;Schmidt et al.,
1985, 1987; MDMA treatment facilitatedthe recoveryofTPH ac--
Schuster et al., 1986; Kleven et al., 1988; Wagner and tivity; 7
days after treatment, fiuoxetine-treated animals, Peroutka,
1988).As pointed out previously in the sec- given MDMA had
recovered 100% of TPH activity,- tion on acute neurochemieal
effects , the changes in whereas animals treated on ly with MDMA
remainedcatecholaminergic systems caused by MDMA and its at 54% of
controls.congeners are transient and relatively slight. No long-
Other pharmacological interventions have beens term deficits in DA,
NE, and their metabolites or in shown to have attenuating effects
on both the acutethe activity of TH following MDMA administration
and long-term neurochemical effects of MDMA- have been reported.
(Schmidt and Taylor, 1987; Johnson et al., 1988a;
ls Stoneetal.,1988).Forexample,SchmidtndTaylor- Recoveryof the
serotonerglc system (I 987)found that co-administration
of5-HT2selectiveRelatively few investigations have focused on the
antagonists, or the nonspecific 5-HT antagonist me-recovery of the
serotonergic system after exposure to tbiothepin, partially blocked
MDMA-induced reduc-neurotoxic doses of MDMA. Stone et al. (1987b)
re- tions in cortical TPH activity. By contrast, treatmentported
that, in rats, striatal TPH, 5-HT, and 5-HIAA with yohimbine,
a-methyl-p-tyrosine (AMT), or reser-r remainedsignificantly
depressed 110days after expo- pine had no effecton the acute
MDMA-induced lossed sureto multipledosesof MDMA.Battagliaet al.
ofTPH activity.- (1988b)found a slow regeneration of 5-HT uptake
site Stone et al. (1988) investigated the role ofDA in the ,-
densitiesover a 12-month period, with aconcomitant acute and
long-term serotonergic deficits induced by
recoveryincortical 5-HTlevels.At 6months, theden- MDMA. They
foundthat pretreatment withAMT,re-airyof 5-HT uptake sites was only
75% of controls, serpine, or reserpine plus AMT significantly
reduced- whereas by 12 months the densities had returned to the
acute loss of neostriatal TPH activity with respectt contro l
levels . This study d id not determine whether to MDMA alone.
Reserpine plus AMT did not affo rdf therecoveryresulted
fromregeneration of neurons that any additional protection over
reserpine alone. Con-, hadundergone terminal degeneration or from
an in- current treatment withAMT and MDMA significantlys
creasedcollateral sprouting of neurons unaffected by attenuated the
loss of TPH activity in animals killed. thetreatment. MDMA is
selectivelyneurotoxic to fine 18hafter treatment. Prior lesioning
of the neostriatum5-HT nerve terminals originating in the dorsal
raphe with local injections of 6-hydroxydopamine providedaxons,
while sparing the varicose terminals originating significant pro
tection against loss of neostriatal TPHin the median raphe nuclei
(Molliver, 1987; O'Hearn activity at 3 h after MDMA versus
sham-lesioned an-et al., 1988), so collateral sprouting from this
median imals receiving equivalent doses. TPH activity in the
J. Neurochern., Vol. 54, No. 1, 1990
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
7/9
20 D. J. McKENNA AND 5'. J. PEROUTKA,q._ frontal cortex and
hippocampus, which did not receive (1987) found no evidence of
neurotoxicity following' local 6-hydroxydopamine, was still
significantly de- systemic administration of the major metabolites
oL pressed 3 h after MDMA in both sham-operated and MDA or MDMA
(a-methyldopamine, 4-hydroxy-3-
lesioned animals. Prior depletion of DA and other
methoxyamphetamine, ot-methylnorepinephrine, andonoamines with
reserpine significantly attenuated the a-methylepinephfine).,_
long-term loss of TPH activity. A similar protectiveeffect was
found for pretreatment with AMT, which Possible formation of a
toxic DA metabolite
protected against loss of TPH activity and also pre- DA has been
implicated as a possible mediator o3] vented long-term reductions
in 5-HT, but not 5-HIAA. the neurotoxicity produced by both
methamphetamine(Schmidt et al., 1985) and MDMA (Stone et al.,
1988)GBR 12909, a DA-specific uptake inhibitor, provided
Interestingly, selective 5-HT uptake blockers blocka significant
protective effect with respect to all threeparameters. The authors
speculated that their results methamphetamine-induced changes in
the serotonergic
_ implied a role for endogenous DA in mediating system, but do
not alter its effects on dopaminergicystems (Hotchkiss and Gibb, 1
98 0 ; Bakhit et al., 1 98 1_ MDMA neurotoxicity. Hekmatpanah et
al. (1989), Schmidt and Gibb, 1985). By contrast, MDMA is se
x'xxar' however, reported that prior depletion of central
5-HTand DA with reserpine (5 mg/kg) did not prevent the lectively
neurotoxic to the serotonergic system withL decrease in density of
5-HT uptake sites measured by relatively little effect on markers
for dopaminergic[3H]paroxetine binding 2 weeks after a single
exposure neurotoxicity, such as brain catecholamine levels oTH
activity. Nevertheless, MDMA elicits DA release_i to MDMA (30
mg/kg, i.p.), in vitro (Schmidt et al., 1987)and in vivo (Yamamotoi
MECHANISMS OF NEUROTOXICITY and Spanos, 1988), and inhibition of DA
synthesis bypretreatment with AMT results in a significant atten-'_
Although the serotonergic neurotoxicity of MDMA uation of the
neurotoxic effects of MDMA (Stone ec, is now well established, the
underlying mechanisms of al., 1988)._[ action remain unknown.
Although various theories Gibb and colleagues (1989) have
speculated that in-ave been proposed, two major hypotheses have
been terruption of the normal route of DA catabolism req/,_
advanced to explain MDMA-induced neurotoxicity, suiting from
MDMA-induced blockade ofmonoamine,
oxidasecouldresultinhighconcentrationsfextr4 Possible formation of
a toxicMDMA metabolite cellular DA adjacent to 5-HT neurons,
enhancing the_'i The finding, previously mentioned above, that
carrier-mediated uptake of DA, an effect specificallyMDMA-induced
neurotoxicity can be specifically blocked by 5-HT uptake
inhibitors, such as fluoxetine,[ blocked by 5-HT uptake inhibitors
(Schmidt, 1987b) Oxidative processes inside the 5-HT terminal
could
illustrates the importance of the 5-HT uptake carrier result in
the generation ofcytotoxic metabolites capablein mediating the
neurotoxic effects of MDMA. How- of reacting covalently with
nucleophilic cell compo-
ever, MDMA itself does not appear to be transported nents, such
as enzymes and cell membranes. A similars) directly into 5-HT nerve
terminals (Schmidt et al., role for autooxidation products of
endogenous DA in1987; Wang et al., 1987). The fact that fluoxetine
can the neurotoxicity of amphetamine and methamphet-13 provide
significant protective effects up to 6 h post amine has also been
proposed (Fuller and Hemrick-
MDMA (Schmidt, 1987b) is evidence that the neuro- Luecke, 1982;
Schmidt et al., 1985). The differential%[ toxic agent may be a
metabolite of either MDMA or neurotoxicity of MDMA on serotonergic
terminalsn endogenous substance. Additional support for the versus
dopaminergic terminals could result from factorsexistence ofa
neurotoxic metabolite is the finding that such as drug affinity for
DA and 5-HT uptake carriers,_" direct intracerebral injection of
MDMA yielded no ev- drug metabolism and half-life in brain, and the
inherent._xS idence of a neurotoxic response, whereas systemic
ad-inistration produces the long-term serotonergic del- metabolic
differences which exist between catechol-x','
amineand5-HTneurons(Hillarpetal.,1966).Agaiq
icits associated with neurotOxicity (Molliver et al.,1986). the
DA theory of MDMA-induced neurotoxicity is thefact that 5-HT
terminal damage can be observed_ Schmidt (1987b) speculated that
demethylation of throughout the central nervous system, whereas
DAthe side-chain nitrogen followed by intramolecular cy- systems
are essentially localized to the basal ganglia.._lt clization could
result in formation of an indolic 5-HT and mesolimbic
system.neurotoxinstructurallyelatedto5,6-dihydroxytryp-
tamine. However, the possibility that N-demethylation
CONCLUSIONSt_ of MDMA to its more neurotoxic congener MDAmight
constitute an intermediate step in the formation MDMA causes an
acute, but reversible, depletion
_ { of a neurotoxic metabolite was rendered unlikely by of 5-HT,
an effect which may account for its psycho-Schmidt and Taylor
(1987), who established that pre- logical effects. In addition,
MDMA and some of it_'_ treatment with the selective N-demethylase
inhibitor, derivatives have long-term effects on the nervous sys,!
piperonyl butoxide, had no effect on the acute depletion tem which
are indicative ofserotonergic neurotoxicity:,,_ of cortical 5-HT or
on suppression of TPH activity long-term depletions of 5-HT,
5-HIAA, and TPH ac-''_ [ induced by MDMA. In a separate study, Yeh
and Hsu tivity; histopathologic changes in 5-HT-containing
J. Neurochern., Vol. 54, No. 1, 1990
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
8/9
i 3,4-METHYLENEDIOXYMETHAMPHETAMINE 21
nerve terminals; and ablation of 5-HT uptake sites, causes a
long-term depletion of serotonin in mt brain. Brainof The
neurochemical and neurotoxic properties of Res. 447, 141-144.uller
R. W. and Hemrick-Luecke S. K. (1982) Further studies onMDMA and
its derivatives are similar in many respects the long-term
depletion of striatal dopamine in iprindole-treatedto those of
other neurotoxic amphetamine derivatives, rats by amphetamine.
Neuropharmacology 21, 433--438.such as methamphetamine, PeA, and
fenfluramine, Fuller R. W., Perry K. W., and Molloy B.B . (1975)
Reversible andand it is likely that common mechanisms mediate both
irreversible phases of serotonin depletion by 4-chloroamphet-the
acute and long-term actions of some or all of these amine. Eur. J.
Pharmacol. 33, i 1 9-124.G eh lert D . R ., Sch mid t C . J., W u
L., an d L ove nbe rg W . (1 98 5) Ev-
of agents. Although enantiomers and structural analogues
idencefor specificmethylenedioxymethamphetamine (ecstasy)e of MDMA
exhibit clear differences in neurotoxic po- binding sites inthe rat
brain. Eur. J. Pharmacol. 119, 135-136.tential, a complete
understanding of the structural de- Gibb J. W., Stone D. M.,
Johnson M., and Hanson J. R. (1989)k terminants of neurotoxicity
within the methylene- Neurochemical effects of MDMA, in MDMA:
"Ecstasy" and/orHumanNeurotoxin(Peroutka S.J., ed),pp. 133-150.
Kluweric dioxyamphetamines is still lacking.
AcademicPublishers,Norwell,Massachusetts.The possible clinical
significance of the neurotoxic Greer G. and Toibert R. (1986)
Subjective reports of the effects of1; potential of MDMA in human
users of the drug re- MDMA in a clinical setting. J.
PsychoactiveDrugs 18, 319-328.e- mains unknown. The difficulty in
extrapolating the Hekmatpanah C. R., McKenna D. J., and Peroutka
S.J. (1989) Re-h data derived in laboratory animals to humans is
corn- serpine does not prevent
3,4-methylenedioxymethamphetamine-induced neurotoxicity. Neurosci.
Lett. (in press).
pounded by the lack of a reliable clinical index ofhu- Hillarp
N.-A., Fuxe K., and Dahlstrom A. B. (1966) Demonstrationor man
serotonergic function and the lack of a readily and mapping of
central neurons containing dopamine, nor-e recognizable
pathological syndrome resulting from se- adrenaline, and
5-hydroxytryptamine and their reactions to
rotonergic deficits. It is hoped that significant progress
psychopharmaca. Pharmacol. Rev. 18, 727-741.Hotchkiss A. and Gibb
J. W. (1980) Long-term effects of multipleby on both of these
fronts: will be achieved in the near d os es o f m etbam ph etam in
e o n try pto ph an h yd ro xy la se an d ty -future. In the
meantime, MDMA and its congeners rosine hydroxylase activity in rat
brain. J.Pharmacol.Exp. Ther.et co_titute useful tools for the
neurochemist and po- 214, 257-262.tenfial health hazards of
uncertain proportions for their Johnson M. P., Hoffman A. L, and
Nichols D. E.0986) Effectsofin- human users, the enantiomers of
MDA, MDMA, and related analogues on
re- [aH]serotonin and [3H]dopamine release from superfused
ratbrain slices.Eur. J. Pharmacol. 132,269-276.a- REFERENCES
Johnson M., Hanson G. R., and Gibb J. W. (1987) Effects of
N-ethyl-3,4-methylenedioxyamphetamine (MDE) on central se-e BakhitC
., Morgan M. E., and Gibb J. W. (1981) Long-term effects rotonergic
and dopaminergic systems of the rat. Biochem. Phar-
of methamphetamine on the synthesis and metabolism of 5- macol.
36, 4085--4093.hydroxytryptamine in various regions of the rat
brain. Neuro- Johnson M., Hanson G. R., and Oibb I. W. (1988a)
Effects ofd 'pharmacology 20, 1135- I 140. paminergic and
serotonevgicreceptor blockade on neurochemicald BattagliaG., Yeh S.
Y., O'Hearn E., Molliver M. E., Kuhar M.J.," changes induced by
acute administration ofmethamphetaminee and De Souza E. B. (1987)
3,4-Methylenedioxymethamphet- and
3,4-methylenedioxymethamphetamine. Neuropharrnacology- amine and
3,4-methylenedioxyamphetamine destroy serotonin 27,
1089-1096.terminals in rat brain: quantification of
neurodegeneration by Johnson M., Letter A. A., Merchant K., Hanson
G. R., and Gibbmeasurement of [3H]paroxetine-labeled serotonin
uptake sites. J.W. (1988b) Effects of 3,4-methylenedioxyamphetamine
andin J. Pharrnacol.Exp. Ther. 242, 911-916.
3,4-methylenedioxymethampbetamine isomers on central sero-t-
Battaglia G., Brooks B. P., Kulsakdinun C., and De Souza E.B.
tonergic, dopaminergic and nigral neurotensin systems of the-
(1988a) Pharmacologic profile ofMDMA (3,4-methylenedioxy- rat. J.
Pharmacol. Exp. Ther.244, 977-982.l methamphetamine) at various
brain recognition sites. Eur. J. Kleven M. S., Schuster C. R., and
Seiden L. S. (1988) Effect of de-PharmacoL 149, 159-163. pletion of
brain serotonin by repeated fenfluramine on neuro-BattagliaG., Yeh
S. Y., and DeSouza E. B.(1988b) MDMA-induced chemical and anorectic
effects of acute fenfiuramine. J. Phar-
neurotoxicity: parameters ofdegeneration and recovery of brain
macol. Exp. Ther.246, 822-828.s, serotonin neurons. Pharmacol.
Biochem. Behav. 29, 269-274. Lyon R. A., Glennon R.A., and Titeler
M. (1986) 3,4-Methylenedi-BattagliaG., Zaczek R., and De Souza E.
B. (1989) MDMA effects oxymethamphetamine (MDMA): stereoselective
interactions atl- in brain: pharmacologic profile and evidence of
neurotoxicity brain 5HT, and 5HT: receptors. Psychopharmacology 88,
525-st from neurochemical and autoradiographic studies, inMDMA:
526."Ecstasy" and/or Human Neurotoxin (Peroutka S. J, ed), pp.
Merchant K. M., Letter A. A., Johnson M., Stone D. M., Gibb /he
171_199.Kluwer Academic Publishers, Norweil, Massachusetts. J.W.,
and Hanson G. R. (1987) Effects of amphetamine ana-L-']inesehmidtB.
V., Zacchei A. G., Totaro J. A., Pfieuger A.B., !ogues on
neurotensin concentrations in rat brain. Eur. J. Phar-DA McGut_n J.
C., and Wishousky T. I. (1978) Fenfluramine and macoL 138,
151-154.
brain serotonin. Ann. NYAcad. Sci. 305, 222-241.ComminsD.
L.,VosmerG., VirnsR. M.,Woolverton W. L., Schuster Mokler D. J.,
Robinson S. E., and Rosecrans J. A. (1987) (+_)3,4-C. R ., and
Seiden L. S. (1 98 7) B iochemical and histo logical er- M
ethylenedioxymethamphetamine (M DM A) produces !ong-idence that
methylenedioxymethylamphetamine (MDMA) is term reductions in brain
5-hydroxytryptamine in rats.Eur. J.toxic to neurons in the rat
brain. J. Pharmacol.Exp. Ther.241, Pharmacol. 138, 265-268.338-345.
MolliverM.E.(1987)Serotonerg iceuronalsystems:whattheiran-
on DeSouzaE. B. and Kuyatt B. L. (1987) Autoradiographic
localization atomic organization tells us about function. J. Clin.
Psycho-o- of aH-paroxetine-labeled serotonin uptake siteS in rat
brain, pharmacol. 7, 3S--23S.Synapse 1, 488--496. Moll iver M.E.,
O'Hearn E., Battaglia G., and De Souza E. B.(1986)its Downing J.
(1986) The psychological and physiological effects of Direct
intracerebral administration of MDA and MDMA doessy_- MDMA on
normal volunteers. J. PsychoactiveDrugs 18, 335- not produce
serotonin neurotoxicity. Soc. Neurosci. Abst. 12,ty: 340. 1234.ac-
Finnegan K. T., Ricaurte G. A., Ritchie L. D., Irwin I., Peroutka
Nash J. F. ,Meltzer H. Y., and Gudelsky G. A. (1988) Elevation ofng
s.J., and Langston J. W. 0988) Orally administered MDMA serum
prolactin and corticosterone concentrations in the rat
J.Neurochem.,Vol.54,No. 1. 1990
-
8/3/2019 Dennis McKenna - Neurochemistry and Neurotoxicity of
MDMA
9/9
v
'_ 22 D. J. McKENNA AND S. J. PEROUTKA
'_ tophan hydroxylase activity following the acute
administratiafter the administration of
3,4-methylenedioxymethamphet-
/ amine. J. Pharmacol. Exp. Ther.245, 873-879. of
methylenedioxymcthamphetamine. Biochem.Pharmacol.l, Nichols D. E.
(1986) Differences between the mechanism of action
4095--4102.ofMDMA, MBDB, and the classic hallucinogens.
Identification Schmidt C. J. and Taylor V. L. (1989) Neurochemical
effectsof a new therapeutic class: entactogens. J.
PsychoactiveDrugs methylenedioxymethamphetamine in the rat: acute
versuslo18, 305-313. term changes, in MDMA: "Ecstasy" and/orHuman
Neuroto
Nichols D. E., Lloyd D. H., Hoffman A. J., Nichols M. B., and
Yim (Peroutka S.J., ed), pp. 151-169. Kluwer Academic PublishG. K.
(1982) Effects of certain hallucinogenic amphetamine an- Norwell,
Massachusetts.
i alogues on the release of [3H]serotonin from rat brain synap-
Schmidt C. J., Ritter J. K., SonsaI!a P.K., Hanson G. R., and
Gtosomes. J.Med. Chem. 25, 530-535. J.W. (1985) Role of dopamine in
the neurotoxic effects'Hearn E., Battaglia G., De Souza E. B.,Kuhar
M. J., and Molliver methamphetamine. J. Pharmacol. Exp. Ther.233,
539-544Schmidt C. J., Wu L., and Lovenberg W. (1986)
Mcthylenedioxo! M.E. (I 988) Methylenedioxyamphetamine (MDA) and
meth-
ylenedioxymethamphetamine (MDMA) cause selectiveablation
methamphetamine: a potentially neurotoxic amphetamineof
serotonergic axon terminals in forebrain: immunocytochem- alogue.
Eur. J. Pharmacol. 124, i75-178.q4[ ical evidence for
neurotoxicity. J. Neurosci. 8, 2788-2803. Schmidt C. J., Levin J.
A., and Lovenberg W. (1987) In vitroandPeat M. A., Warren P. F.,
Bakhit C., and Gibb J. W. (1985) The vivoneurochemical effects
ofmethylenedioxymethamphetamin
%X' acute effects ofmethamphetamine, amphetamine and p-chloro-
on striatal monoaminergic systems in the rat brain.
Biocheamphetamine on the cortical serotonergic system ofthe rat
brain: Pharmacol. 36, 747-755.evidence fordifferences in the
effects of methamphetamine and Schuster C. R., Lewis M., and Seiden
L. S. (1986) Fenfluramiamphetamine. Eur. J. Pharmacol. 116, 11-16.
neurotoxicity. Psychopharmacol.Bull. 22, 148- !51.
: Peroutka S.J. (1987) Incidence ofrecreational useof
3,4-methylene- Slikker W., Ali S. F., ScalletA.C., FrithC. H.,
Newport G. D., a'_ dioxymethamphetamine (MDMA, "Ecstasy") on an
under- Bailey J. R. (1988) Neurochemical and neurohistological
algraduate campus. N. Eng. J.Med. 317, 1542-1543. ations in the rat
and monkey produced by orally administer
PeroutkaS. J. (1988)Relative insensitivity ofmice to
3,4-mcthylene- methylenedioxymethamphetamine (MDMA). Toxicol.
Apdioxymethamphetamine neurotoxicity. Res. Commun. Subs. Pharmacol.
94, 448--457.Abuse 9, 193-206. Steele T. D., Nichols D. E., and Yim
G. K. W. (1987)StereochemiPierce P. A. and Peroutka S.J. (1988)
Ring-substituted amphetamine effects of
3,4-methylenedioxymethamphetamine (MDMA) ainteractions with
neurotransmitter receptor binding sites in hu- related amphetamine
derivatives on inhibition of uptake of [3man cortex. Neurosci.
Lett. 95, 208-212. monoamines into synaptosomes from different
regions ofr Pierce P. A. and Peroutka S. J. (1989)Evidence for
distinct 5- brain. Biochem. Pharmacol. 36,2297-2303.
hydroxytryptamine: binding site subtypes incortical membrane
Stone D. M., Stahl D. C., Hanson G. R., and Gibb J. W. (1986)
Tpreparations. J. Neurochem 52, 656-658. effects of
3,4-methylenedioxymethamphetamine (MDMA) a_ Ricaurte G., Bryan G.,
Strauss L, Soiden L, and Schuster C. (1985)
3,4-methylenedioxyamphetamine (MDA) on
monoaminevgnerveHallucingeniCterminals.amphetaminescience29,selectivelY986_988,estroys
brain serotonin StoneSyStemsD..,injohnsontheat
brain.M.,ansonEUr'.G.Pharmacl'R.,nd Gibb128,.4IW.--48.
: Ricaurte G. A., Finnigan K. F., Nichols D. E., DeLanney L. E.,
Irwin A comparison of the neurotoxic potential of methylenediox-[
I., and Langston J. W. (1987) 3,4-Methylenedioxyethylamphet-
amphetamine (MDA) and itsN-methylated and N-ethylatedamine (MDE), a
novel analogue of MDMA, produces long-last- rivatives. Eur. J.
Pharmacol. 134,245-248.ing depletion of serotonin in the rat brain.
Eur. J. Pharmacol. StoneD .M., Merchant K. M., Hanson G. R., and
Gibb J. W. (198
265-268. Immediate and long-term effects of
3,4.methylenedioxymet37 ,Ricaurte G. A., DeLanney L.E., Irwin I.,
and Langston J. W. (19881) amphetamine on serotonin pathways in
brain of rat. Neu_ Toxic effect ofMDMA on central serotonergic
neurons in the pharmacology 26, 1677-1683.primate: importance of
route and frequency of drug adminis- Stone D. M., Hanson G. R., and
Gibb J. W. (1987c) Differencestration. BrainRes. 4146,165-168. the
central serotonergic effects of methylenedioxymethamphe
B Ricaurte G. A., DeLanney L. E., Weiner S. G., Irwin 1., and
Langston amine (MDMA) in mice and rats.Neuropharmacology26, 16t
J.W. (1988b) 5-Hydroxyindoleacetic acid in cerebrospinal fluid
1661.f reflects serotonergic damage induced by 3,4-methylenedioxy-
Stone D. M., Johnson M., Hanson G. R., and Gibb J. W.
(19methamphetamine inCNS of non-human primates. BrainRes. Role of
endogenous dopamine in the central serotonergicdefiIn 474, 359-363.
induced by 3,4-methylenedioxymethamphetamine. J. Ph
Sanders-Bush E. and Sterenka L. R. (1978) Immediate and long-
macol. Exp. Ther.247, 79-87.term effects ofp-chloroamphetamine on
brain amines. Ann.NY Wagner J. A. and Peroutka S. J. (1988)
Comparative neurotoxic' Acad. Sci. 305, 208-221. of fenfiuramine
and 3,4-methylenedioxymethamphetaminSanders-Bush E., Bushing J. A.,
and Sulser F. (1975)Long-term effects (MDMA). Soc. Neurosci. Abst.
14, 327.a ng S . S ., R ica urte G . A ., a nd P ero utka S . J .
(1 98 7 ) [3H ]-3,4 -M et[. of p-chloroamphetamine and related
drugs on central seroto- ylenedioxymethamphetamine
(MDMA)interactions with brnergic mechanism. J. Pharmacol. Exp.
Ther. 192, 33-41. membranes and glass fiber filter paper. Eur. J.
PharmacoJ. 1Schmidt C.J. (19871) Acute administration of
methylenedioxyam- 439-443.phetamine: comparison with the
neurochemical effects ofits N- W ilson M . A ., Ricaurte G . A .,
and M olliver M . E. (1 98 9) D istidesmethyi and N-ethyl
analogues. Eur. J. Pharmacol. 136, 81- morphologic classes of
serotonergic axons in primates exh
_, 88. differential vulnerability to the psychotropic drug
3,4-methSchmidt C.J. (1987b)Neurotoxicity of the psychedelic
amphetamine, enedioxymethamphetamine. Neuroscience 28, 121-137.r'
methylenedioxYmethamphetamine. J. Pharmacol. Exp. Ther. Yamamoto
B.K. and SpanosL. J. (1988) The acute effects o f me240,1-7.
ylenedioxymethamphetaminendopamineeleasentheawSchmidt C. J. and
Gibb 1.W. (1985) Role of the dopamine uptake behaving rat. Eur. J.
Pharmacol. 148, 195-203.
[ carrier in the neurochemical response to methamphetamine: ef-
Yeh S. Y. and Hsu F.-L. (1987) Neurotoxicity of the metaboliteseets
of amfonelic acid. Eur. J. Pharmacol. 109, 73-80. MDA and MDMA
(Ecstasy) in the rat. Soc. Neurosci.Abstq/_ Schmidt C. J. and
Taylor V. L. (1987) Depression of rat brain tryp- 906.s4
J. Neurochem., VoL 54, No. 1, 1990















![[MDMA]MDMA Neurochemistry](https://img.dokumen.tips/doc/110x75/577dab601a28ab223f8c57f3/mdmamdma-neurochemistry.jpg)