Embed Size (px)
Citation preview

i
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DE ÁGUA DOCE E PESCA
INTERIOR - BADPI
Metazoários parasitos de um peixe detritívoro de lagos de várzea do rio
Solimões, Pterygoplichthys pardalis (Castelnau, 1855) (Siluriformes:
Loricariidae) e uma avaliação de seu uso como espécies bioindicadoras de
efeito em mudanças ambientais na Amazônia
DANIEL BRITO PORTO
Manaus, Amazonas
Novembro, 2017

ii
DANIEL BRITO PORTO
Metazoários parasitos de um peixe detritívoro de lagos de várzea do rio
Solimões, Pterygoplichthys pardalis (Castelnau, 1855) (Siluriformes:
Loricariidae) e uma avaliação de seu uso como espécies bioindicadoras de
efeito em mudanças ambientais na Amazônia
ORIENTADOR: JOSÉ CELSO DE OLIVEIRA MALTA Dr.
Manaus, Amazonas
Novembro, 2017
Tese apresentada ao Instituto
Nacional de Pesquisas da
Amazônia como parte dos
requisitos para obtenção do
título de DOUTOR em
CIÊNCIAS BIOLÓGICAS

iii

iv
P 853 Porto, Daniel Brito
Metazoários parasitos de um peixe detritívoro de lagos de várzea
do rio Solimões, Pterygoplichthys pardalis (Castelnau, 1855)
(Siluriformes: Loricariidae) e uma avaliação de seu uso como
espécies bioindicadoras de efeito em mudanças ambientais na
Amazônia / Daniel Brito Porto. --- Manaus: [s.n.], 2017.
viii, 84 f.: il.
Tese (Doutorado) --- INPA, Manaus, 2017.
Orientador: José Celso de Oliveira Malta
Área de concentração: Biologia de Água Doce e Pesca Interior
1. Acarí bodó. 2. Parasitas de peixes. 3. Bioindicadora ambiental. I. Título.
CDD 597.49
SINOPSE
Este trabalho faz parte do projeto PIATAM (Inteligência Socioambiental Estratégica da Indústria do Petróleo na
Amazônia), que estuda os efeitos da mineração do petróleo e seu transporte entre os municípios de Manaus e
Coari no Estado do Amazonas. Este estudo se insere na sub-área de Ictioparasitologia. As coletas dos peixes
foram realizadas nos meses de março, maio, setembro e dezembro de 2012. Foram coletados e identificados 1.635
parasitos de cinco grupos taxonômicos. Foram encontradas 12 espécies de parasitos. Das doze espécies
encontradas na comunidade componente: Diplostomum sp. e U. brevispinus foram as espécies que parasitaram o
maior número de hospedeiros e tiveram os maiores índices parasitológicos. Unilatus brevispinus foi a espécie
mais abundante. Diplostomum sp. foi a espécie que preencheu os requisitos propostos por Overstreet (1997) para
ser uma espécie bioindicadora de efeito.
Palavras chaves: Parasitos de peixes; Monogenoidea; Digenea; Acanthocephala; Cestoda; Copepoda; espécie
bioindicadora ambiental.

v
Aos meus Amores
Amanda
Thierry
José Marcos
Eliana
Maria Clara
Meus maiores incentivos

vi
AGRADECIMENTOS
A Deus por tudo que permite acontecer nesta trajetória.
Ao meu grande orientador José Celso de Oliveira Malta que sempre me incentivou e
sempre teve MUITA paciência em me orientar durante esta longa jornada.
E um agradecimento especial à Bióloga Amanda Karen Silva de Souza fundamental na
realização das necropsias e por desenhar todas as pranchas desta tese muito obrigado pela
paciência e seu amor.
Aos meus pais Almir e Eliana e meus irmãos Charles e Israel pelo apoio e carinho.
Aos meus filhos Thierry, José Marcos, Eliana e Maria Clara que sempre me motivam e dão
força nesta jornada.
Um agradecimento especial para o técnico do laboratório Edilson de Araújo Silva por todo
o trabalho realizado em nosso laboratório e amizade em todos estes anos.
Aos Professores do curso de Biologia de água doce pela transmissão do conhecimento e
atenção ao longo do curso.
Ao INPA por todos estes anos onde pude aperfeiçoar todo o trabalho que venho realizando.
Ao projeto PIATAM e todas equipes em especial a de Ictiologia e ao pescadores Luís,
Fonseca e China que fizeram parte deste projeto por toda logística na realização do trabalho
de campo.
A Coordenação de Aperfeiçoamento de Nível Superior CAPES pela concessão da bolsa.
A todos que contribuíram direta e indiretamente neste trabalho.
UM SINCERO AGRADECIMENTO!!!

vii
“Quem quer faz, quem não quer inventa uma história”
Malta, JCO

viii
RESUMO
Foram analisados 248 indivíduos de Pterygoplichthys pardalis capturados em cinco lagos
de várzea da Amazônia. O comprimento médio dos peixes foi 24,5cm ± 8,8 e o peso médio
213,9g ± 102,6. Os lagos amostrados foram: Baixio; Preto; Ananá e Araçá no rio Solimões
e São Tomé no rio Purus, localizados entre as cidades de Manaus e Coari no estado do
Amazonas. As necropsias foram feitas em campo e os órgãos foram fixados. Os descritores
qualitativos das populações de parasitos foram: número de indivíduos, estágio de
maturação e dominância de cada espécie e sua importância. Foram coletados 1.635
espécimes parasitos de cinco táxons: Monogenoidea (785); Digenea (795), Cestoda (1),
Acanthocephala (50) e Copepoda (4). Quatro espécies parasitavam as brânquias: três de
Monogenoidea: Unilatus brevispinus, Heteropriapulus heterotylus e Trinigyrus mourei e
uma de Copepoda, Therodamas elongatus. Seis espécies de Digenea ocorreram:
metacercárias de Diplostomum sp., Austrodiplostomum compactum, Odhineriotrema
microcephala e adultos de Megacoelium spinispecum, M. spinicavum e Kalitrema
kalitrema. As metacercárias parasitavam os olhos, estômago, gônadas e superfície dos
órgãos internos. Uma larva de Proteocephalidea foi encontrada na superfície do intestino.
Uma espécie de Acanthocephala Gorytocephalus elongorchis dentro do intestino. Digenea
foi o táxon com maior diversidade com 50% das espécies, seguido por Monogenoidea 25%.
Dentre os componentes das infracomunidade de P. pardalis, duas espécies foram as mais
abundantes durante os quatro períodos hidrológicos: o Monogenoidea, U. brevispinus na
enchente (56,1 %) e na vazante (81,6%) e o Digenea, Diplostomum sp. na cheia (73,2 %)
e na seca (93%). Das doze espécies encontradas na comunidade componente: Diplostomum
sp. e U. brevispinus foram as espécies que parasitaram o maior número de hospedeiros e
tiveram os maiores índices parasitológicos. Unilatus brevispinus foi a espécie mais
abundante. Diplostomum sp. foi a espécie que preencheu os requisitos propostos por
Overstreet (1997) para ser uma espécie bioindicadora ambiental.
Palavras-chave: Parasitos de peixes; Monogenoidea; Digenea; Acanthocephala; Cestoda;
Copepoda; espécie bioindicadora ambiental.

ix
ABSTRACT
Were analyzed 248 individuals of Pterygoplichthys pardalis caught in five lowland lakes
of the Amazon. The mean fish length was 24.5cm ± 8.8 and the mean weight was 213.9g
± 102.6. The sampled lakes were: Baixio; Black; Ananá and Araçá on the river Solimões
and São Tomé on the Purus river, located between the cities of Manaus and Coari in the
state of Amazonas. Necropsies were performed in the field and the organs were fixed. The
qualitative descriptors of the parasite populations were: number of individuals, stage of
maturation and dominance of each species and their importance. A total of 1,635 parasite
specimens from five taxa were collected: Monogenoidea (785); Digenea (795), Cestoda
(1), Acanthocephala (50) and Copepoda (4). Four species parasitized the gills: three of
Monogenoidea: Unilatus brevispinus, Heteropriapulus heterotylus and Trinigyrus mourei
and one of Copepoda, Therodamas elongatus. Six species of Digenea occurred:
metacercariae of Diplostomum sp., Austrodiplostomum compactum, Odhineriotrema
microcephala and adults of Megacoelium spinispecum, M. spinicavum and Kalitrema
kalitrema. The metacercariae parasited the eyes, stomach, gonads and surface of internal
organs. A larva of Proteocephalidea was found on the surface of the intestine. A species of
Acanthocephala Gorytocephalus elongorchis inside the intestine. Digenea was the most
diverse taxa with 50% of the species, followed by Monogenoidea 25%. Among the
components of the P. pardalis infracommunity, two species were the most abundant during
the four hydrological periods: Monogenoidea, U. brevispinus in flood (56.1%) and ebb
(81.6), and Digenea, Diplostomum sp . (73.2%) and dry (93%). Of the twelve species found
in the community component: Diplostomum sp. and U. brevispinus were the species that
parasitized the largest number of hosts and had the highest parasitological indexes. Unilatus
brevispinus was the most abundant species. Diplostomum sp. was the species that fulfilled
the requirements proposed by Overstreet (1997) to be an environmental bioindicator
species.
Key-words: Fish parasites; Monogenoidea; Digenea; Acanthocephala; Cestoda;
Copepoda; environmental bioindicator species.

x
SÚMÁRIO
INTRODUÇÃO...................................................................................................................1
Bioindicadores ambientais.......................................................................................2
Espécies parasitas como bioindicadoras ambientais ...............................................4
Hospedeiro: Pterygoplichthys pardalis (Castelnau,1855) .....................................10
OBJETIVOS..................................................................................................................... 14
MATERIAL E MÉTODOS................................................................................................15
Área de estudo....................................................................................................... 16
Coleta dos peixes....................................................................................................18
Coleta e processamento das espécies parasitas ......................................................19
Análises das comunidades componente das espécies parasitas..................................22
RESULTADOS.................................................................................................................25
Taxonomia das espécies de metazoários parasitas..................................................25
Biodiversidade das espécies de metazoários parasitas (Comunidade componente)43
DISCUSSÃO.....................................................................................................................59
BIBLIOGRAFIA CITADA...............................................................................................74

xi
LISTA DAS TABELAS
Tabela 01 - Índices parasitários, número de peixes parasitados, estágio de desenvolvimento
e órgãos parasitados pelas espécies parasitas de Pterygoplichthys pardalis (Castelnau,
1855) capturados em lagos de várzea do rio Solimões........................................................45
Tabela 02 - Prevalência (P), Intensidade média (IM), Abundancia média (AM) e Status
comunitário das espécies parasitas de Pterygoplichthys pardalis (Castelnau, 1855)
capturados em cinco lagos, 2012........................................................................................47
Tabela 03 - Prevalência (P), Intensidade media (IM) e abundância média (AM) de
Diplostomum sp. espécie parasita do Pterygoplichthys pardalis (Castelnau, 1855) por
órgão parasitado e lago onde foi capturado pardalis (Castelnau, 1855) capturados em cinco
lagos, .................................................................................................................................47
Tabela 04 - Estrutura da comunidade componente de Pterygoplichthys pardalis (Castelnau,
1855) entre os lagos de várzea na enchente......................................................................50
Tabela 05 - Estrutura da comunidade componente de Pterygoplichthys pardalis (Castelnau,
1855) entre os lagos de várzea na cheia............................................................................50
Tabela 06 - Estrutura da comunidade componente de Pterygoplichthys pardalis (Castelnau,
1855) entre os lagos de várzea na vazante.........................................................................51
Tabela 07 - Estrutura da comunidade componente de Pterygoplichthys pardalis (Castelnau,
1855) entre os lagos de várzea na seca................................................................................51
Tabela 08 - Índice de Dominância (D) em % das espécies parasitas de Pterygoplichthys
pardalis (Castelnau, 1855) durante as fases do período hidrológico...................................52
Tabela 09 - Índice de dispersão (ID) e índice de Green das espécies parasitas de
Pterygoplichthys pardalis durante as fases do período hidrológico....................................53
Tabela 10 - Coeficiente de Jaccard entre os lagos de coleta de Pterygoplichthys pardalis
durante as fases do período hidrológico..............................................................................54

xii
LISTA DAS FIGURAS
Figura 01 - Espécime adulto de Pterygoplichthys pardalis (Castelnau, 1855) ...................13
Figura 02 - Vista lateral da cabeça de P. pardalis (Castelnau, 1855)................................14
Figura 03 - Vista superior da cabeça de P. pardalis (Castelnau, 1855) .............................14
Figura 04 - Boca ventral e dentes de P. pardalis (Castelnau, 1855) .................................14
Figura 05 - Vista lateral dos raios dorsais de P. pardalis (Castelnau, 1855) .......................14
Figura 06 - Padrão de coloração vista superior de P. pardalis (Castelnau, 1855) ...............14
Figura 07- Padrão de coloração vista ventral de P. pardalis (Castelnau, 1855) .................14
Figura 08 - Imagem de satélite e localização geográfica dos lagos do rio
Solimões.............................................................................................................................15
Figura 09 - Desenho de Unilatus brevispinus Suriano, 1985..............................................26
Figura 10 - Fotomicrografia de Unilatus brevispinus Suriano, 1985..................................26
Figura 11 - Desenho de Heteropriapulus heterotylus, 2006. 1 - corpo inteiro, 2 – barra
dorsal, 3 - âncora, 4 - Cirrus (MCO), 5 – barra ventral, 6- gancho, 7 - âncora ventral.........28
Figura 12 - Desenho de Trinigyrus mourei Boeger & Jegú, 1994 ......................................29
Figura 13 - Fotomicrografia em contraste de fase de Trinigyrus mourei Boeger & Jegú,
1994....................................................................................................................................29
Figura 14 - Desenho de Diplostomum sp............................................................................31
Figura 15 - Fotomicrografia de Diplostomum sp................................................................31
Figura 16 - Fotomicrografia da metacercária de Austrodiplostomum compactum (Lutz,
1928) parasito dos olhos.....................................................................................................32
Figura 17 - Desenho da metacercária de Austrodiplostomum compactum (Lutz, 1928)
parasito dos olhos...............................................................................................................32
Figura 18 - Desenho de Megacoelium spinicavum Thatcher e Varella, 1981......................33
Figura 19 - Desenho de Megacoelium spinispecum Thatcher e Varella, 1981....................34
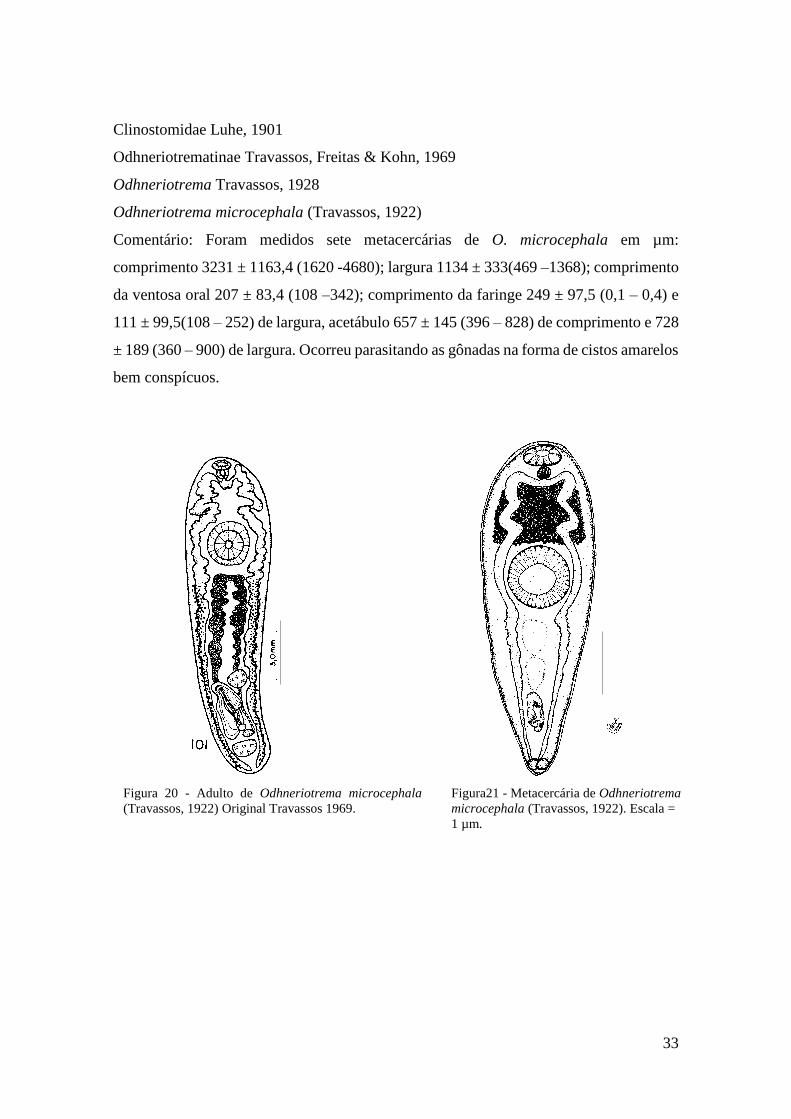
Figura 20 - Adulto de Odhneriotrema microcephala (Travassos, 1922) Original Travassos
1969....................................................................................................................................36
Figura 21 - Metacercária de Odhneriotrema microcephala (Travassos, 1922) ..................36
Figura 22 - Fotomicrografia da metacercária de Odhneriotrema microcephala (Travassos,
1922)..................................................................................................................................36
Figura 23 - Órgãos internos do adulto de Kalitrema kalitrema Travassos, 1933................37

xiii
Figura 24 - Desenho do adulto de Kalitrema kalitrema Travassos, 1933..........................38
Figura 25 - Ventosas e órgão apical do escólex da larva de Proteocephalidea....................39
Figura 26 - Escólex da larva de Proteocephalidea..............................................................39
Figura 28 - A) Adulto macho de Gorytocephalus elongorchis Thatcher, 1981
B) Detalhe probóscide. (Desenho original Thatcher, 1981)................................................41
Figura 29 - Parte anterior fêmea adulta de Gorytocephalus elongorchis...........................41
Figura 30 - A) Therodamas elongatus; B) primeira antena e C) segunda antena. Desenhos
originais de Thatcher, 1986................................................................................................43
Figura 31 - Variação da prevalência de Diplostomum sp. e Unilatus brevispinus parasitas
de Pterygoplichthys pardalis durante o período hidrológico.............................................54
Figura 32 - Variação da Intensidade média de Diplostomum sp. e Unilatus brevispinus
parasitas de Pterygoplichthys pardalis durante o período hidrológico...............................55
Figura 33 - Variação da abundância de Diplostomum sp. e Unilatus brevispinus parasitas
de Pterygoplichthys pardalis por fase do período hidrológico............................................55
Figura 34 - Variação da prevalência de Diplostomum sp. de Pterygoplichthys pardalis entre
os lagos durante as fases do período hidrológico. B – Baixio, P – Preto, ST – São Tomé,
AN – Ananá e AR – Araçá..................................................................................................56
Figura 35 - Variação da intensidade média de Diplostomum sp. de Pterygoplichthys
pardalis entre os lagos durante as fases do período hidrológico. B – Baixio, P – Preto, ST
– São Tomé, AN – Ananá e AR – Araçá.............................................................................56
Figura 36 - Variação da abundância de Diplostomum sp. entre os lagos durante as fases do
período hidrológico. B – Baixio, P – Preto, ST – São Tomé, AN – Ananá e AR – Araçá....57
Figura 37 - Variação da prevalência de Unilatus brevispinus entre os lagos durante as fases
do período hidrológico. B – Baixio, P – Preto, ST – São Tomé, AN – Ananá e AR –
Araçá..................................................................................................................................57
Figura 38 - Variação da intensidade média de Unilatus brevispinus de Pterygoplichthys
pardalis entre os lagos durante as fases do período hidrológico. B – Baixio, P – Preto, ST
– São Tomé, AN – Ananá e AR – Araçá.............................................................................58
Figura 39 - Variação da abundância de Unilatus brevispinus de Pterygoplichthys pardalis
entre os lagos durante as fases do período hidrológico. B – Baixio, P – Preto, ST – São
Tomé, AN – Ananá e AR – Araçá.......................................................................................58

1
INTRODUÇÃO
Os sete milhões de km2 da bacia Amazônica, com seus inúmeros cursos d’água
fluindo através de uma grande variedade de solos e comunidade vegetais, oferecem uma
abundância de nichos aquáticos e terrestres. O Amazonas, o maior rio tropical tanto em
descarga como em área, acolhe mais espécies de peixes que qualquer outro sistema fluvial
(Smith 1979).
A água é um fator preponderante na paisagem amazônica. Isto se pode dizer tanto
para os majestosos rios e igarapés, que contribuem para a formação dos rios gigantes. A
rede de igarapés na Amazônia é tão densa como em quase nenhuma região do mundo.
Porém, olhando-se de perto verifica-se que os corpos de água não são uniformes.
Encontram-se diferenças consideráveis tanto em relação à morfologia de seus leitos
quanto as suas características químicas e biológicas e seus efeitos na comunidade de
animais e plantas (Junk 1983; Junk et al. 1989).
Os lagos que acompanham os grandes rios e que são típicos para as áreas alagadas
(várzea e igapó) faltam nas áreas não inundáveis (terra firme) onde igarapés e pequenos
rios caracterizam a paisagem. Rios com água barrenta ocorrem tanto quanto rios de água
preta ou cristalina. Enquanto alguns rios quase não têm correnteza, outros passam por
corredeiras e cachoeiras (Junk 1983).
Na Amazônia a distribuição da precipitação durante o ano não é homogênea e
difere também nas diferentes áreas da vasta bacia. Em consequência disso, o nível dos
rios está submetido a fortes oscilações. Em dependência da distribuição irregular da
precipitação, os grandes afluentes do rio Amazonas atingem seu nível máximo em
diferentes épocas do ano (Salati 1983).
Por meio de um lento processo de interações, as forças da natureza definem os
ecossistemas naturais. Há uma caracterização do solo, do clima e das espécies do reino
vegetal e animal. O ecossistema atinge um estado de equilíbrio dinâmico e assim
permanece até que novas alterações sejam introduzidas no ambiente (Salati 1983).
Nas oscilações do equilíbrio dos ecossistemas, têm-se que levar em consideração
as decorrentes de fenômenos naturais que ocorrem e que determinam grandes
modificações climáticas. Como consequências das variações do clima, adaptações
ocorrem na flora e na fauna que impõe à biosfera diferentes condições de equilíbrio (Salati
1983).
De um modo geral, o ambiente aquático amazônico fornece uma grande variedade
de itens alimentares para os peixes. Estes exploram todas as fontes disponíveis, desde

2
invertebrados como esponjas e briozoários até peixes e frutos (Santos e Ferreira 1999).
As águas doces fornecem habitat para uma variedade de organismos: bactérias,
protozoários, fungos, esponjas, celenterados, platelmintos, nematódeos, rotíferos,
briozoários, moluscos, crustáceos, aracnídeos e vários grupos de insetos. Se o
conhecimento sobre a biodiversidade em águas doces é incompleto para vertebrados o
quadro se agrava mais ainda para micro-organismos e invertebrados (Rocha 2002).
A bacia Amazônica abriga a maior e mais diversa ictiofauna de água doce do
mundo, com estimativas que variam de 1.500 a 6.000 espécies (Reis et al. 2003). A pesca
na região Amazônica se destaca em relação às demais regiões brasileiras, tanto de áreas
costeiras quanto de águas interiores, pela riqueza de espécies exploradas, pela quantidade
de pescado capturado e pela dependência das populações tradicionais a esta atividade
(Barthem e Fabré 2003).
As espécies de peixes amazônicas apresentam estratégias notáveis para se
adaptarem às mudanças sazonais nos diversos ambientes que ocupam. A compreensão
destas adaptações é de fundamental relevância para o entendimento da abundância e da
composição dos recursos pesqueiros. Consequentemente para a definição de políticas de
manejo da pesca (Barthem e Fabré 2003).
Os lagos de várzea possuem a maior diversidade e abundância íctica da bacia
Amazônica (Handerson e Crampton 1997). Esta ictiofauna apresenta-se basicamente
constituída por grupos recentes, sendo 43% de Characiformes, 39% de Siluriformes e 3%
de Gymnotiformes (Santos e Ferreira 1999).
Na Amazônia não se sabe ao certo quantas espécies de peixes existem. Muito
menos quantas espécies de parasitas utilizam o peixe como hospedeiro intermediário ou
definitivo. Mais informações sobre essa fauna aumentariam o conhecimento sobre a
biodiversidade amazônica bem como subsídios para sua utilização como marcadores
biológicos e bioindicadores ambientais (Malta e Varella 2009).
Assim os peixes servem de substrato para uma grande variedade de espécies
parasitas. Estes que possuem uma distribuição mundial afetando a grande maioria das
espécies, das águas tropicais às polares e qualquer nicho ecológico e habitat do hospedeiro
(Roberts 1978; Eiras 1994; Thatcher 2006).
Bioindicadores ambientais
As profundas mudanças causadas nos ecossistemas aquáticos no decorrer dos
últimos anos têm causado uma preocupação crescente na população e nas autoridades

3
ambientais (Queiroz et al. 2000). Problemas que trazem consequências ao homem, como
alterações nos regimes hidrológicos, aumento das doenças de veiculação hídrica,
contaminação química, erosão e assoreamento dos corpos d’água (Pompeu et al. 2004).
Os impactos ambientais nos ecossistemas aquáticos têm diferentes origens e
formas. Muitas vezes, o lançamento de compostos químicos nas águas resulta em
concentrações muito superiores àquelas encontradas naturalmente. Como consequência
observa-se modificações no curso e composição físico-química natural dos rios, na
cobertura vegetal, nas margens, na cor da água e na biota existente (Callisto et al. 2001).
O uso de parâmetros biológicos para medir a qualidade da água se baseia nas
respostas dos organismos em relação ao meio onde vivem. Como os rios estão sujeitos a
inúmeras perturbações, a biota aquática reage a esses estímulos, sejam eles naturais ou
antropogênicos (Cairns Jr. et al. 1993). Assim suas características ambientais
especialmente as comunidades biológicas fornecem informações sobre as consequências
das ações do homem (Callisto et al. 2001).
A biota aquática está constantemente exposta a um grande número de substâncias
tóxicas lançadas no ambiente, oriundas de diversas fontes de emissão. A descarga de lixos
tóxicos provenientes de efluentes industriais, os processos de drenagem agrícola, os
derrames acidentais de lixos químicos e os esgotos domésticos lançados em rios e mares
contribuem para a contaminação dos ecossistemas aquáticos (Rashed 2001).
A definição de biomonitoramento mais aceita é o uso sistemático das respostas de
organismos vivos para avaliar as mudanças ocorridas no ambiente, geralmente causadas
por ações antropogênicas (Matthews et al. 1982 apud Buss et al. 2003). Bioindicadores
são espécies escolhidas por sua sensibilidade ou tolerância a vários parâmetros, como
poluição orgânica ou outros tipos de poluentes (Washington 1984 apud Buss et al. 2003).
A utilização dos bioindicadores é extremamente útil, especialmente para a
avaliação de impactos ambientais decorrentes de descargas pontuais de esgotos
domésticos e efluentes industriais. Monitorando-se estações de amostragem a montante,
no local de lançamento e a jusante da fonte poluidora, pode-se identificar as
consequências ambientais para a qualidade da água e saúde do ecossistema aquático
(Callisto et al. 2001)
Programas de biomonitoramento utilizam os mais variados tipos de
bioindicadores. Entre os desenvolvidos para ecossistemas aquáticos podemos citar:
macroinvertebrados bentônicos (Goulart e Callisto 2003; Queiroz et al. 2000); algas
(Favero et al. 1996; Jonsson et al. 2009); sanguessugas (Metcalf et al. 1988); protozoários

4
de vida livre (Boikova, 1990); crustáceos (Farkas et al. 2003); peixes (Belmejo e Martos
2008; Ramsdorf 2007; Bell e Burt 1991) e parasitos de peixes (Madi 2005; Madi e Ueta
2009; Morais 2011; Lacerda et al. 2017)
O grau de diversidade biológica é frequentemente utilizado como um indicador da
saúde do sistema ecológico. É de consenso geral que qualquer distúrbio no habitat indica
uma alteração na diversidade. Essa visão pode ser aplicada para analisar a biodiversidade
de uma comunidade parasitária. A diversidade parasitária em peixes hospedeiros é por
esta razão diretamente dependente de grau de diversidade do habitat e pode ser empregado
como um monitor altamente sensível para detectar as alterações na biodiversidade, que
caracterizam os habitats afetados por poluição (D’amelio e Gerasi 1997; Ponce de Leon
e Prieto 2002).
Espécies parasitas como bioindicadoras
Uma variedade de organismos tem sido investigada para avaliar seu potencial
como indicador biológico de diferentes tipos de poluição no ambiente aquático. Certas
espécies têm sido identificadas como altamente sensíveis à resposta fisiológica gerada
por estes poluentes (Sures et al. 1997). Entre estes as espécies parasitas têm demonstrado
que são uma ferramenta bastante eficiente como bioindicadores (Vidal-Martinez et al.
2009).
Uma vez que a poluição e outros estressores podem exercer impactos sobre as
populações e as comunidades de organismos e sobre a estrutura da rede alimentar. Então
as espécies parasitas podem também ser usadas como indicadoras biológicas naturais de
alterações na “saúde” do ecossistema (Marcogliese 2005).
As espécies parasitas são indicativas de muitos aspectos da biologia de seus
hospedeiros: dieta; migração; “recrutamento” e filogenia. Espécies parasitas com ciclos
de vida heteroxeno, a sua transmissão depende da presença de uma variedade de
hospedeiros intermediários invertebrados ou vertebrados, hospedeiros paratênicos e
definitivos dentro do ecossistema (Anderson et al. 1988; Williams et al. 1992).
As interações entre as populações de parasitos também podem influenciar a
composição, abundância e distribuição das populações de parasitos de peixes (Rhode et
al. 1995). A comunidade de espécies parasitas de peixes pode mostrar uma considerável
variação de acordo com as condições ambientais onde ela vive (Hossain et al. 2008).
O monitoramento ambiental tem como objetivo detectar mudanças, em especial,
a causada pela ação do homem aparte daquelas que ocorrem naturalmente. Com ele

5
permite localizar e avaliar as condições dos sistemas e as consequências das atividades
humanas (Silva-Souza et al. 2006). Alterações significativas na biodiversidade e no
número de indivíduos das populações são utilizadas como um alerta das condições que
deterioram o ambiente (MacKenzie et al. 1995).
Uma vez que a presença de espécies parasitas em hospedeiros vertebrados é
diretamente dependente do grau de diversidade do habitat. A análise da comunidade
parasitária dos peixes de um local pode ser um bom demonstrativo de estresse ambiental
e biodiversidade (Overstreet 1997).
Espécies parasitas são geralmente negligenciadas no estudo, manejo e
conservação dos ecossistemas (Marcogliese 2004). Como são encontradas em quase
todos os ambientes, desde locais extremamente frios como regiões polares até climas
quentes como os trópicos. O conhecimento sobre elas é uma ferramenta importante e útil
em avaliações ambientais (Lewis e Hoole 2003).
Tendo os parasitos como indicadores da biologia do hospedeiro, de contaminantes
ambientais e estrutura da cadeia alimentar, a comunidade parasitaria de peixes pode ser
um bom demonstrativo de estresse ambiental e da biodiversidade (Chubb 1979; 1980;
Overstreet 1997). Pois eles podem prover uma grande quantidade de informações a
respeito de seu hospedeiro e o ambiente ao seu redor (Marcogliese 2004).
Existem vários tipos de poluentes encontrados nos ambientes aquáticos mais os
principais são hidrocarbonetos, metais pesados e poluição termal(usinas termoelétricas,
nucleares) (Mackenzi et al. 1995). O efeito de cada tipo de poluente pode gerar uma
resposta diferente que depende do táxon que estiver sendo estudado e pode aumentar ou
diminuir a abundância das espécies parasitas em seu hospedeiro (Galli et al. 2001; Madi
e Ueta 2009).
As espécies parasitas são uma parte essencial do ambiente aquático. A presença
destes organismos se torna evidente quando ocorre uma massiva infestação manifestando
sinais clínicos em seu hospedeiro, levando a sua morte. Uma situação que pode ser
ocasionada por mudanças ambientais que ocorrem nos fatores bióticos ou abióticos do
ambiente (Moller 1987).
O conhecimento da biologia da espécie parasita e seu hospedeiro, as relações
hospedeiro-parasito e a situação ambiental podem ajudar na detecção de mudanças no
ambiente. O uso de parasitos de peixes como indicadores biológicos é um estudo de
extrema importância não somente para estudos de impactos nos ambientes aquáticos, mas
também para estudos ecológicos e biodiversidade (Palm e Dobberstein 1999).

6
As espécies parasitas podem ser divididas de maneira geral em dois tipos:
ectoparasitas que utilizam como substrato para sua sobrevivência a parte externa de seu
hospedeiro e endoparasitas que utilizam as partes internas de seus hospedeiros (Thatcher
2006). O efeito decorrente do agente poluidor é diferente para cada um dos tipos de
parasitos (Mackenzie et al. 1995).
Dentre todas as espécies parasitas, as ectoparasitas são as mais utilizadas na
bioindicação por responderem diretamente às alterações ambientais (Lafferty 1997).
Porém no âmbito geral as espécies de Cestoda e Nematoda, presentes nos peixes, são
utilizadas principalmente para efeito nos cálculos dos índices de diversidade (Schmidt et
al. 2003).
Uma vez que muitos destas espécies parasitas envolvem em seu ciclo de vida,
vários hospedeiros que são encontrados nos diferentes níveis tróficos. Os parasitos
refletem os hábitos de vida dos peixes e suas interações com as comunidades bentônicas,
planctônicas e ícticas (Silva-Souza et al. 2006).
As espécies parasitas com ciclo de vida heteroxênico são potenciais indicadoras
ambientais. Isto porque, em alguma parte do seu ciclo de vida tem estágios de vida livre
que dependem de uma série de organismos pequenos, delicados, que são seus hospedeiros
intermediários que são muito sensíveis às variações ambientais (Mackenzie et al. 1995).
Espécies de diversos táxons de parasitas têm sido utilizadas para avaliar os efeitos
de poluentes nos ambientes aquáticos (Overstreet 1988; Galli et al. 2001; Marcogliese
2004; 2005; Vidal-Martinez et al. 2009.). Os principais táxons utilizados: Trichodina spp.
(Palm e Dobberstein 1999; Ogut e Palm 2005); Monogenoidea (Madi e Ueta 2009);
Digenea (Anderson 1988; Galli et al. 2001); Cestoda (Sures et al. 1997; Giese 2010);
Nematoda (Morais 2011); Acanthocephala (Sures 1997; Zimmermman et al. 1999)
Crustacea (Galli 2001).
As espécies de Monogenoidea são ectoparasitas, monoxenos e vivem em contato
direto com o seu hospedeiro e o ambiente aquático. Parasitam principalmente as
brânquias, têm um curto ciclo de vida e reagem imediatamente a qualquer mudança
causada no ambiente. E dependendo do agente a população pode diminuir ou aumentar
(Sures 2001)
As espécies de Digenea são endoparasitas e heteroxênicas. Elas dependem de
espécies de moluscos, peixes, anfíbios, répteis, aves e mamíferos que vivem no ambiente
aquático para completar seu ciclo de vida (Thatcher 2006). De acordo com o tipo de
poluente este pode agir sobre qualquer um dos elos da sua cadeia de transmissão

7
acarretando a diminuição da população ou a extinção naquele ambiente (Valtonen et al.
2003).
As espécies de Acanthocephala são endoparasitas frequentes de peixes ocorrem
na maioria das vezes no intestino. Este táxon tem sido muito utilizado em estudos de
acumulo de metais pesados, pois acantocéfalos concentram mais metais pesados em seus
tecidos que seus hospedeiros (Sures et al. 1997; 2001).
O sistema de monitoramento através das espécies parasitas de peixes possui um
grande valor. Este responde a mudanças que ocorrem em muitas partes do ecossistema
aquático. Estas comunidades parasitas refletem mudanças na dinâmica populacional dos
hospedeiros definitivos e intermediários que estão em contato direto com os poluentes
(Landsberg et al. 1998; Diamant et al. 1999; Valtonen et al. 2003).
Conhecendo melhor essa fauna e suas relações com os peixes teremos um
indicador da situação natural desses ambientes. É ampliado o conhecimento sobre a
biodiversidade de parasitas de peixes amazônicos e as inter-relações com os hospedeiros
em diferentes níveis tróficos, identificando ainda espécies de parasitas com potencial
bioindicador (Morais 2011).
Para utilizar as espécies parasitas de peixes como bioindicadores foram
estabelecidos critérios que abrangem tanto o parasito como o hospedeiro indicador
(Overstreet 1997).
Critérios para selecionar uma espécie de peixe como hospedeira de espécies
parasitas indicadoras da qualidade ambiental (Overstreet 1997):
1. A área onde vive o peixe deve ser restrita. Devem residir em uma área pequena
ou bem definida onde passem a maior parte do tempo. Os peixes que fazem
grandes migrações e têm o habitat menos definido fica muito difícil avaliar a saúde
do ambiente, pois não se sabe em que local os parasitas foram adquiridos;
2. O peixe deverá ser capaz de servir como hospedeiro para um número grande de
espécies parasitas. Preferivelmente, pelo menos várias dessas espécies, devem
incorporar uma grande variedade de hospedeiros adicionais (intermediários,
paratênicos definitivos) com os seus ciclos de vida;
3. O peixe deve ser comum, de fácil captura e identificação. Uma vantagem
adicional é ser pequeno. Que facilitaria os processos de captura, exame, necropsia
e coleta dos parasitas. Peixes grandes são mais difíceis de serem capturados, se
gasta mais tempo para serem examinados e demandam mais trabalho para obter

8
dados histológicos.
Critérios para selecionar espécies parasitas como bioindicadoras ambientais
(Overstreet 1997):
1. Riqueza de espécies parasitas heteroxênicas, ou daquelas que têm mais de um
hospedeiro em seu ciclo de vida. Espécies parasitas que incluem mais de um
hospedeiro em seu ciclo de vida geralmente ocorrem em, relativamente, baixos
números em ambientes contaminados. Estes resultados podem ocorrer porque os
contaminantes reduzem ou eliminam os outros hospedeiros, intermediários ou
definitivos, nos ciclos desses parasitas ou porque o contaminante tem um efeito
tóxico diretamente em cima do parasita;
2. Informações biológicas e epidemiológicas sobre o ciclo de vida das espécies
parasitas para indicar a biodiversidade. O comportamento racional dessa
ferramenta é similar ao critério 1 (um), que é restrito ao número de espécies. Essa
ferramenta requer conhecimento adicional sobre o a história de vida das espécies
parasitas. Valores para a prevalência e intensidade média de uma espécie
específica de parasita em diferentes localidades demonstram a diferente natureza
dos habitats em termos das associações de invertebrados de vida livre e
hospedeiros vertebrados;
3. Informações epidemiológicas sobre as espécies parasitas que reproduzem dentro
(internamente) ou sobre (externamente) o hospedeiro. Essa ferramenta e a do
critério 4 (abaixo) consistem em valores de dados, onde os hospedeiros
apresentaram índices parasitários muito maiores nos ambientes altamente
contaminados do que naqueles sem contaminantes. A razão dessas altíssimas
infestações envolve um decréscimo da resistência ou imunidade do hospedeiro,
um aumento na fonte de nutrição da espécie parasita ou em uma interação com
outra infecção por um agente microbiano;
4. Alterações histológicas relativas à infestação por espécies parasitas e o estresse
causado pela substância contaminante. Como indicado acima no critério #3 o
contaminante pode reduzir a resistência do hospedeiro permitindo ao parasito
reproduzir e desenvolver. Isto pode causar necroses ou alguma outra condição
patológica no tecido do hospedeiro, os quais podem tornar-se uma fonte adicional
de nutrientes para a espécie parasita reproduzir ainda mais. Alguns desses
parasitos são externos, mas muitos são protozoários histozóicos e outros parasitos

9
internos.
Em adição ao uso das quatro abordagens acima, para amostragens simples e com
somente um ponto de amostra da saúde ambiental. O monitoramento “stricto senso” deve
ser conduzido em áreas cuidadosamente selecionadas, onde se tenha conhecimento dos
dados da rotina básica do local para servir como referência, para poder comparar com as
mudanças do parasitismo, da qualidade da água ou efluentes e também as mudanças no
tempo (Overstreet 1997).
Para essas amostragens, também seriam especialmente úteis, que os peixes
hospedeiros capturados tivessem amostras de indivíduos adultos e juvenis. E, que ao
menos periodicamente, deveriam ser examinadas para poder estimar quando uma espécie
específica de parasita foi ou não foi adquirida (Overstreet 1997).
Fatores que podem complicar os resultados na utilização das espécies parasitas
como indicadoras de qualidade ambiental (bioindicadores) segundo Overstreet
(1997).
1. Utilizar um ou mais locais de referência apropriados para comparação, utilizar o
mesmo peixe hospedeiro e se possível, os mesmos fatores para comparação
(mesma área geral, salinidade, profundidade da água, vegetação) e outras
características comparáveis. Os locais devem ser amostrados ao mesmo tempo ou
tão próximo quanto possível que os pressupostos de estarem contaminados.
2. Flutuações sazonais normais da espécie parasita podem produzir dados
enganosos, em especial quando o parasito possui ciclo de vida mais curto que a
biota está sendo avaliada. Se apenas uma coleta for planejada é ideal que seja no
período sazonal em que a abundância da espécie parasita esteja alta no local
escolhido. Além disso, fatores como efluentes antropogênicamente aquecidos
podem resultar no aumento de estágios infecciosos para o peixe, quando
comparados com uma área normalmente não aquecida.
3. Condições ambientais extremas ou atípicas, tais como inundações, furacões,
congelamento atípico, prolongados períodos de chuvas ou seca podem produzir
resultados enganosos, especialmente quando comparados com dados obtidos em
longo prazo ou ano - a - ano.
4. O efeito de algumas toxinas em situações específicas sobre um peixe pode
aumentar, ou influenciar na presença de um parasita específico ou vice-versa,
como indicado acima. A interação normal das condições ambientais para este

10
relacionamento também pode influenciar grandemente a saúde do hospedeiro e do
parasita.
5. A inter-relação entre as espécies parasitas pode produzir um aumento ou
diminuição do número de indivíduos ou espécies.
6. O modelo deverá ser adequado para acomodar uma grande riqueza de espécies
parasitas, levando em conta se os dados epidemiológicos devem ser considerados
como indicadores.
7. Se o hospedeiro possui uma vasta dispersão, poderia adquirir ou perder alguns
parasitas distante dos locais amostrais
Hospedeiro: Pterygoplichthys pardalis (Castelnau, 1855)
Pterygoplichthys pardalis (Castelnau, 1855) pertence à ordem Siluriformes, a
família Loricariidae e a subfamília Hypostominae. Estes peixes têm o corpo roliço ou
achatado em seção transversal e corpo coberto por placas ósseas e formam de cinco a seis
séries sobre o tronco. Cada placa com pequenas estruturas ósseas, bastante ásperas ao tato
ou mesmo perfurantes, os odontódios (Reis et al. 2003; Santos et al. 2006; Soares et al.
2007). Para este gênero são conhecidas 16 espécies (Froese e Pauly 2017).
Pterygoplichthys pardalis ocorrem em áreas de várzea, lagos e margem de rios de
águas brancas. Nos locais onde a concentração de oxigênio é alta sua respiração é
totalmente aquática. Mas sua estratégia para explorar os ambientes pobres em oxigênio é
a utilização da respiração aérea acessória (Santos et al. 2006).
Assim como os demais loricariídeos, P. pardalis tem o estômago extremamente
vascularizado que é utilizado como órgão respiratório acessório que auxilia nas trocas
gasosas, quando há deficiência de oxigênio na água. Por causa desta função, o alimento
não fica retido no estômago e vai direto para o intestino (Brito, 1981; Almeida-Val et al.
1999).
Pterygoplichthys pardalis é endêmico a bacia Amazônica e ocorre ao longo do rio
Amazonas/Solimões (Reis et al. 2003). É um peixe detritívoro que se alimenta de matéria
orgânica particulada e micro-organismos associados, como protozoários fungos e
bactérias. Detrito é a mais importante fonte de alimentos da planície inundada da
Amazônia e cerca de 40% da ictiomassa utiliza esse recurso (Yossa e Araújo-Lima 1998;
Santos et al. 2006).
Muitas espécies de peixes consomem detrito na região Neotropical. Algumas
ocasionalmente, outras, como as espécies da família Prochilodontidae, Curimatidae e

11
Loricariidae são altamente especializados no consumo deste recurso. Elas alimentam-se
exclusivamente de detrito (Weber 1992).
Espécies detritívoras são dominantes nas assembleias de peixes na planície de
inundação dos rios da América do Sul. Elas representam mais de 40% da ictiomassa
(Araújo-Lima et al. 1995). É uma parte importante da biomassa dos peixes da bacia
amazônica (Araújo-Lima et al. 1986).
As espécies detritívoras mais importantes são: Prochilodus nigricans Agassiz,
1829; P. pardalis; Semaprochilosdus insignis (Jardine & Schomburgk, 1841); S.
taeniurus (Vallenciennes, 1817); Pothamorhina altamazonica (Cope, 1878); P.
pristigaster (Steindachner, 1876); P. latior (Spix & Agassiz, 1829); Psectrogaster
amazonica Eigenmann & Eigenmann, 1889; Curimata inornata Vari, 1989 (Araújo-Lima
et al. 1986; Yossa e Araújo-Lima, 1998; Santos et al. 2006).
Pterygoplichthys pardalis tem o intestino longo e enovelado, com cerca de 18
vezes o seu comprimento total, que auxilia na digestão e absorção da matéria orgânica
ingerida (Brito 1981). É considerado de grande porte, pode chegar a 50 cm. É a espécie
da família Loricariidae mais importante comercialmente na calha do rio
Solimões/Amazonas devido à sua abundância e aceitação no mercado (Batista e Petrere
2003).
Existem poucas informações sobre as espécies de parasitas que utilizam os peixes
detritívoros como hospedeiros intermediários e definitivos. Entre os trabalhos que
focaram seus estudos neste grupo podem ser citados Thatcher (2006) que estudou várias
espécies, mas sem um aprofundamento e mais recente Porto (2009) onde foi estudada a
fauna componente de parasitas de P. pardalis de lagos de várzea.
São citados para P. pardalis sete espécies parasitas de quatro grupos. Parasitando
as brânquias as espécies de Monogenoidea: Unilatus sp. e Heteropriapulus sp.; e larvas
de Nematoda. O Branchiura parasito da cavidade branquial Dolops geayi (Bouvier, 1897).
Parasitos dos olhos, estômago e gônadas, metacercárias livres de Austrodiplostomum
compactum (Lutz, 1928). Parasito do estômago Megacoelium spinicavum Thatcher &
Varella, 1981 adultos. Parasitos do intestino Gorytocephalus sp. (Thatcher 1981; 2006;
Thatcher e Varella 1981; Jogunoori et al. 2004; Kritsky 2007; Eiras et al. 2010; Porto et
al. 2012).
Pterygoplichthys pardalis é uma espécie que atende a todos os critérios propostos
por Overstreet (1997) para que uma espécie de peixe possa ser usada como hospedeira de
espécies parasitas indicadores da qualidade ambiental:

12
1. A área onde P. pardalis vive é restrita, vive nos lagos de várzea da bacia Amazônica,
onde passa a maior parte do tempo;
2. É hospedeira de um número razoável de espécies parasitas. Essas espécies
incorporaram diferentes hospedeiros adicionais em seus ciclos de vida;
3. É um peixe muito comum, de fácil captura e identificação.
Figura 01 – Espécime adulto de Pterygoplichthys pardalis (Castelnau, 1855)
(Escala=10cm).

13
OBJETIVOS
Figura 02 - Vista lateral da cabeça de Pterygoplichthys pardalis
(Castelnau, 1855) (Escala=1cm).
Figura 03 - Vista superior da cabeça de Pterygoplichthys
pardalis (Castelnau, 1855) (Escala=1cm).
Figura 04 – Boca ventral e dentes de Pterygoplichthys
pardalis (Castelnau, 1855) (Escala=1cm). Figura 05 - vista lateral dos raios dorsais de Pterygoplichthys
pardalis (Castelnau, 1855) (Escala=1cm).
Figura 07- padrão de coloração vista ventral de
Pterygoplichthys pardalis (Castelnau, 1855) (Escala=1cm).
Figura 06- padrão de coloração vista superior de
Pterygoplichthys pardalis (Castelnau, 1855) (Escala=1cm).

14
OBJETIVOS
Objetivo geral
Conhecer as espécies de metazoários parasitas de um peixe detritívoro de
lagos de várzea do rio Solimões, P. pardalis e avaliar seu uso como
espécies bioindicadoras de efeito de mudanças ambientais na Amazônia
Central.
Objetivos específicos:
Identificar as espécies de parasitas metazoários de P. pardalis dos lagos
várzea estudados;
Determinar os índices parasitários de cada espécie parasita;
Detectar padrões temporais nas infestações pelas espécies de parasitas;
Avaliar e selecionar as espécies parasitas que preencham os critérios para
serem utilizadas indicadoras ambientais.

15
MATERIAIS E MÉTODOS
Este trabalho foi desenvolvido em colaboração com o projeto “Inteligência
Socioambiental Estratégica da Indústria do Petróleo na Amazônia – “PIATAM”, um
projeto interinstitucional e interdisciplinar que envolveu instituições de ensino e pesquisa
e teve como principal meta a caracterização socioambiental da área de atuação da
Petrobrás no estado do Amazonas. O presente trabalho se insere na subárea de
Ictioparasitologia no projeto “Estudo da fauna de parasitas de peixes e sua avaliação como
bioindicadores ambientais, na Amazônia Central”.
Área de estudo
Caracterização da área de estudo e coleta dos dados
Os lagos amostrados são lagos de várzea localizados ao longo das margens do rio
Solimões. Áreas essas denominadas de várzeas baixas, influenciadas pelo pulso de
inundação na época de cheia, sendo abastecido por lençol freático ou igarapés oriundos
da floresta circundante no período da seca. Os lagos amostrados foram: Baixio; Preto;
São Tomé (rio Purus); Ananá; Araçá localizados no trecho compreendido entre as cidades
de Manaus e Coari no estado do Amazonas, em um trecho de aproximadamente 400 km.
As coordenadas geográficas e características físicas de cada lago estão descritas abaixo.
Figura 8 - Imagem de satélite e localização geográfica dos lagos ao longo do rio Solimões
(Fonte: Google Earth).

16
Lago do Baixio (S 03°17’27,2”/ W 60°04’29,6”) localizado no município de
Iranduba é elíptico com área de 79,776 ha e água de cor branca. Próximo às margens
ocorrência de macrófitas aquáticas: Eichhornia crassipes (Mart.) Solms; Paspalum
repens Berg.; Pistia stratiotes Linné, 1753 entre outras. Profundidade máxima de 8,50 m,
na cheia e mínima de 0,85 m na seca.
Lago Preto (S03°21’17,1”/ W60°37’28,6”) localizado no município de
Manacapuru é mais redondo que alongado com área de 221,203 ha. Água de cor de chá,
no período da cheia, com a influência das águas do rio Solimões, ela torna-se mais turva.
Profundidade máxima 7,16 m na cheia e mínima de 0,15 m na seca.
Lago São Tomé (S03˚50'32,4"/ W61˚25'45,4") localizado no rio Purus, um rio de
águas brancas que representa 5% (375.000 km2) da área da bacia do rio Amazonas e
possui aproximadamente 21,000.00 ha de planícies de inundação em suas margens,
constituindo a maior área de planície inundada dentre os afluentes do rio Amazonas
(Goulding 1980). Destaca-se por possuir uma ictiofauna rica com mais de 180 espécies
de peixes, que constitui um grande estoque pesqueiro.
Lago Ananá (S03°53’54,8”/ W61°40’18,4”) localizado no município de Anori,
com área de 551,43 ha e água de cor branca. Lago com um volume muito grande de água
na cheia e na vazante/seca reduziu-se a um canal estreito. Profundidade máxima de 9,37
m na cheia e mínima de 1,08 m na seca.
Lago Araçá (S03°45’04,3”/ W62°21’25,9”) localizado na margem esquerda do
rio Solimões com área é de 188,89 ha e água de cor branca. Lago bastante largo na cheia
e na seca tem uma grande área recoberta por macrófitas aquáticas: Utricular ia foliosa L.
1753; Salvina agriculta Abule; Protenderia sp.; P. repensa Berg. (Prado 2005).
Profundidade máxima de 9,76 m na cheia e mínima de 1,30 m na seca.
Coleta, identificação e transporte dos peixes.
Foram realizadas quatro excursões, no ano de 2012, para cobrir os períodos de
cheia, seca, enchente e vazante. Cada viagem de campo durou cerca de sete dias corridos
sendo as coletas bimensais. As capturas dos peixes, para as análises parasitológicas,
foram realizadas em parceria com o grupo de pesquisa da ictiofauna do projeto PIATAM.
O esforço de pesca foi padronizado, em todas as estações e amostragens, por meio do uso
de redes de espera, dispostas aleatoriamente nos lagos, não obedecendo a um padrão

17
quanto ao local amostrado (margens, água aberta, pausadas, vegetação flutuante).
As redes tiveram dimensões de 20 m de comprimento por 2 m de altura e os
tamanhos das malhas foram de 30, 40, 50, 60, 70, 80, 90 e 100 mm entre nós adjacentes.
O tempo de permanência das redes na água foi de aproximadamente de 10 horas por lago,
período diurno, com duas despescas ao longo deste tempo.
Todos os pontos de coleta foram fixados com a utilização de um GPS (Sistema de
Posicionamento Global) Garbin e-Trecho, para diminuir a margem de erro na recolocação
das redes. Os peixes capturados foram triados e identificados em campo com o auxílio de
chaves e de especialistas. Dentre os peixes coletados, alguns foram selecionados para
serem depositados na Coleção de Peixes Piatam, localizada na Universidade Federal do
Amazonas.
Nos peixes identificados em campo foram examinados imediatamente: a
superfície do corpo; a base das nadadeiras; a cavidade branquial; a cavidade bucal; a
cavidade anal e brânquias a procura de ectoparasitos. Quando encontrados imediatamente
foram coletados, fixados e conservados de acordo com o grupo pertencente para
identificação no laboratório de Parasitologia de Peixes (LPP) do Instituto Nacional de
Pesquisas do Amazonas (INPA).
Posteriormente os peixes foram pesados, medidos e necropsiados, registrados em
fichas de campo, e seus órgãos acondicionados em frascos e registrados de acordo com o
local de coleta e guardados em caixas de isopor na embarcação. Ao chegar a Manaus as
caixas de isopor de 120 litros foram transferidas para o LPP do INPA, onde foram
organizadas até o momento da triagem.
Tamanho das amostras
Como se trata de exemplares de populações naturais, o tamanho efetivo
populacional é desconhecido e, consequentemente não é estatisticamente possível
estabelecer um grau de confiança para detectar pelo menos um exemplar parasitado para
um determinado grau de prevalência. Dessa forma, o tamanho da amostra foi o maior
possível levando em conta as possibilidades de coleta, posterior armazenamento e
processamento do material (Eiras et al. 2006).
Necropsia dos peixes
No laboratório os frascos com os órgãos dos peixes fixados foram retirados das

18
caixas de isopor. Cada exemplar teve uma ficha onde todos os dados referentes ao
hospedeiro, local coletado, condições do peixe, dia, hora, local, coletor, necropsiador,
parasitos, aparência externa, sexo e estágio gonadal (quando possível), número de
parasitas, local de fixação foram registrados. A necropsia foi realizada seguindo o roteiro
do Laboratório de Parasitologia de Peixes do Instituto Nacional de Pesquisas da
Amazônia (LPP/INPA) descrito resumidamente abaixo:
Roteiro de necropsia
Para cada peixe uma ficha de necropsia foi aberta onde todos os dados referentes
ao hospedeiro, local coletado, condições do peixe, dia, hora, local, coletor, necropsiador,
parasitos, aparência externa, sexo e estágio gonadal (quando possível), número de
parasitos, local de fixação foram registrados. As necropsias foram feitas em campo
seguindo um roteiro adaptado do Laboratório de Parasitologia de Peixes do Instituto
Nacional de Pesquisas da Amazônia (LPP-INPA).
Coleta e fixação das espécies parasitas
Todos os parasitos foram fixados em campo juntamente com o órgão que estavam
parasitando até o momento da necropsia no “Laboratório de Parasitologia de Peixes do
INPA. Exceto os espécimes de Branchiura, Isopoda, Annelida e larvas encistadas na pele.
Se encontrados, os indivíduos foram coletados com finos: pinceis; estiletes e pinças. A
metodologia de coleta sofreu algumas adaptações.
Monogenoidea: as brânquias foram retiradas e colocadas em frascos de tamanho
proporcional ao tamanho das brânquias contendo formol 1:4.000; agitadas algumas vezes;
e adicionando formol puro até obter a concentração de 5% (Amato et al. 1991). Para a
coleta de Monogenoidea parasitos da superfície do corpo, os hospedeiros foram
acondicionados em frascos com água quente (60 - 70ᵒC), agitados vigorosamente,
coletando-se posteriormente os parasitos foram conservados em formol 5% (Kritsky e
Stockwell 2005).
Para o exame das fossas nasais a cavidade nasal foi lavada com água destilada.
A roseta foi retirada, colocada em uma placa de Petri com água destilada e lavada várias
vezes com auxílio de uma pisseta. Cada dobra foi examinada, utilizando finos estiletes.
Todos os exames foram feitos sob microscópio estereoscópio. Também foram colocadas
inteiras em recipiente de vidro com tampa, com formol 1:4000 como no processo com a
s brânquias (Varella 1992; 1994; Varella e Malta 1995; 2001).

19
Digenea: foram examinados cuidadosamente o tegumento, escamas, nadadeiras,
a olho nu e com o auxílio de estereomicroscópio pontos brancos, amarelos ou pretos a
procura de metacercárias encistadas ou no líquido dos olhos. Verificado a presença, os
cistos foram rompidos para a liberação e coleta dos parasitas (Thatcher 1993).
Cada órgão interno foi aberto em uma placa de Petri individual. O conteúdo de
cada órgão foi passado por uma peneira de coleta com malha de 154 µm de abertura.
Após lavagem do conteúdo e da parede do órgão todo o material foi transferido para uma
placa de Petri com água da torneira. O órgão foi transferido para outra placa de Petri com
água de torneira. Com pinceis finos foram transferidos os helmintos para placas de Petri
com
Os digenéticos encontrados foram fixados com e sem compressão em A.F.A. (95
partes de etanol 70º GL, 3 partes de formalina comercial (37-40%) e 2 partes de ácido
acético glacial), o tempo de compressão variou de 30 minutos a algumas horas. Estas
preparações foram feitas dentro de placas de Petri fundas, onde o fixador frio foi colocado
com cuidado (Amato et al. 1991; Eiras et al. 2006). Para os helmintos pequenos a
compressão foi feita entre lâmina e lamínula e os extremamente pequenos não foram
comprimidos. Depois de terminada a fixação os digenéticos foram retirados com auxílio
de finos pincéis e conservados em A.F.A. (Amato et al. 1991; Thatcher, 1993; Morais et
al. 2011).
Cestoda: o trato digestivo foi aberto e os cestoides (adultos e larvas) foram
coletados com pincéis, finos estiletes e pinças e transferidos para água destilada. Todos
os Cestoda quando pequeno (metacestódeos) ou seções contendo o escólex, porção
imatura, madura e grávida foram levemente comprimidos entre lâminas e colocados no
fixador A.F.A. em temperatura ambiente (Kennedy e Andersen 1982; (Knoff com.
pessoal). Alguns helmintos foram levados ao refrigerador em água destilada por no
mínimo 24 horas, para possibilitar o relaxamento dos escólices. Larvas foram tratadas
como digenéticos pequenos. Sempre que estavam encistadas foram desencistadas (Amato
et al. 1991).
Acanthocephala: o trato digestivo foi aberto e os acantocéfalos coletados com
pinceis, finos estiletes e pinças. Evitando com que a probóscide se rompesse e ficasse
presa na parede intestinal, em seguida foram resfriados em refrigerador por
aproximadamente 24 horas em solução salina a 0,85% para que a probóscide ficasse
totalmente para fora do corpo, em seguida foram fixados em A.F.A. e posteriormente

20
armazenados em álcool 70º. As larvas foram tratadas como adultos, as não encistadas,
acantelas e as encistadas, cistacantos antes foram removidas do cisto (Amato et al. 1991).
Preparações das espécies parasitas
Monogenoidea: para o estudo dos órgãos internos os parasitos foram corados
com tricrômico de Gomori. Para o estudo morfométrico e das estruturas esclerotizadas,
espécimes foram montados em lâminas permanentes em meio de Hoyer’s modificado
para helmintologia e em Gray e Wess (Amato et al. 1991; Kritsky et al. 1995).
Digenea: para os estudos morfológicos os digenéticos foram desidratados pela
série alcoólica, corados com Carmim Alcoólico Clorídrico de Langeron (Langeron 1949),
diafanizados em óleo de imersão e montados em lâmina e lamínula em bálsamo do
Canadá (Amato et al. 1991; Eiras et al. 2006; Morais 2011).
Cestoda: foram corados pelo carmim de Langeron e Hematoxilina de Delafield,
diferenciados em álcool clorídrico a 0,5%, desidratados em série alcoólica crescente,
clarificados em creosoto de Faia e montados entre lâmina e lamínula com bálsamo do
Canadá ou preservadas em álcool 70 ° (Eiras et al. 2006; Morais 2011).
A terminologia das larvas de Cestoda seguiu a estabelecida por Chervy (2002),
baseada principalmente nas características morfológicas determináveis da larva de
Cestoda totalmente desenvolvida, ou a existência de uma lacuna principal e a retração ou
invaginação do escólex. As medidas foram dadas em mm.
Acanthocephala: os estudos anatômicos foram realizados de acordo com Amin
(1969). Para os estudos morfológicos os acantocéfalos foram corados com Carmim
Alcoólico Clorídrico e diafanizados em óleo de imersão (Eiras et al. 2006) e montados
em bálsamo do Canadá.
Análise dos dados
Descritores qualitativos das populações de parasitos foram: número de indivíduos,
estágio de maturação, sexo e dominância de cada espécie e sua importância. A descrição
quantitativa das infrapopulações de parasitas foi conforme os conceitos de Bush et al.
1997.
Índices parasitários
Os índices parasitários da prevalência (P), Intensidade (I); Intensidade média (IM)
e a abundância (A) foram calculados acordo com Bush et al. 1997:
1. Prevalência (%)

21
𝑷 = 𝑷𝑷
𝑷𝑬 𝑿 𝟏𝟎𝟎
PP-peixes parasitados, PE – peixes examinados
2. Intensidade média
𝑰𝑴 = 𝑵𝒔𝒑𝟏
𝑻𝑷𝑷𝒔𝒑𝟏
Nsp1 – total de espécimes da espécie 1, TPP sp1 – total de peixes parasitados por esta espécie 1
3. Abundância média:
𝑨𝑴 =𝑵𝑻𝑷𝒔𝒑𝟏
𝑵𝑻𝑷𝑬
NTPsp1 – número total de espécimes da espécie 1, NTPE – número total de peixes examinados.
Diversidade da comunidade componente
A diversidade parasitária de cada infracomunidade foi calculada através do índice de
Shannon (H'). O índice de Shannon se eleva conforme aumenta o número de espécies
presentes na amostra e a distribuição das espécies se torna uniforme. H'= 0 se apenas uma
espécie ocorre na amostra e H' é máximo quando todas as espécies são representadas pelo
mesmo número de indivíduos, isto é, em perfeita distribuição uniforme de abundância. É
baseado na abundância proporcional das espécies contidas na amostra.
𝑯′ = − ∑ (𝑷𝒆)(𝑳𝒏𝑷𝒆)𝑺𝒊=𝟏 𝑷𝒆 =
𝒏𝒆
𝑵
Onde:
S = número de espécies; Pe = abundância relativa da espécie; ne = número de indivíduos
da espécie e; N = número total de indivíduos; Ln = logaritimo neperiano.
Para se estimar a riqueza de parasitos em cada ponto por fase do período
hidrológico foi utilizado o índice de diversidade de Margalef (DMg) que analisa a relação
entre o número total de espécimes e o número total de indivíduos observados
𝑫𝑴𝒈=
𝑺−𝟏
𝑳𝒏(𝒏)
Onde:
S = Número total de espécies na amostra; n = Número total de indivíduos na amostra.
O cálculo da diversidade com base na dominância de uma ou mais espécies

22
parasitas na estrutura da comunidade componente foi realizado por meio do índice de
Berger-Parker que expressa a importância proporcional da espécie mais abundante.
𝒅 =𝒏𝑴𝒂𝒙
𝑵
Onde:
nmax = número de indivíduos da espécie mais abundante; N = número de indivíduos
presentes na amostra.
A equitatividade das espécies que equivale a proporção entre a diversidade
observada e a máxima foi calculada pela equitatividade de Pielou (J) baseado no índice
de Shannon-Wiener. A equitatividade mede o quanto as proporções das espécies estão
igualmente distribuídas. Pode assumir valores de 0 (dominância total de uma espécie) a
1 (todas as espécies com a mesma proporção).
𝑱 = 𝑯′
𝑳𝒏𝑺
Onde:
H’= é o índice de diversidade de Shannon; S = é o número de espécie presentes na
amostra.
O índice de Simpson (C) foi calculado para determinar a concentração para
dominância entre espécies parasitas, sendo que a dominância aceita quando C≥0,25.
𝑪 = 𝚺 (𝒏𝒊(𝒏𝒊 − 𝟏)
𝑵(𝑵 − 𝟏))
Onde:
ni = número de exemplares da espécie parasita i e N = número total de exemplares de
todas as espécies parasitas.
Foi calculado o índice de dominância (DA) para verificar o grau de dominância de
cada componente nas infracomunidade de parasitas entre os lagos estudados. É calculado
a partir:
𝑫𝑨 = 𝑵𝑨
𝑵𝑨+𝑵𝑩+𝑵𝑪…𝑵𝒏 X 100
Onde:
NA = dominância da espécie A; NA+NB+NC+……..NN = número de indivíduos das

23
espécies A,B,C......N.
O índice de dispersão (padrão de distribuição em relação à população hospedeira)
foi calculado através do quociente entre a variância e a abundância parasitária média de
cada espécie de parasita, sendo a distribuição considerada aleatória quando = 1, uniforme
quando < 1 e agregada quando > 1 (Zar, 1996).
𝑰𝑫 = (𝑺𝟐
𝒙)
Onde:
s2 = variância amostral; x = abundância parasitária média.
O grau de agregação foi obtido através do cálculo do índice de Green (Krebs,
1999). O índice de Green baseia-se na relação variância pela média e mostra quão
agrupados os indivíduos se encontram na população. Varia de 0 para distribuições
aleatórias até 1 para a máxima agregação.
𝑰𝑮 = (
𝑺𝟐
�̅� ) − 𝟏
𝚺𝒙 − 𝟏
Onde:
s2 = variância amostral; x = média amostral; x = somatória do número de indivíduos na
amostra.
Foi aplicado o coeficiente de similaridade de Jaccard, para verificar o grau de
similaridade entre as espécies-alvo nos lagos amostrados (Magurran 1983).
𝑪𝒋 = 𝒂
𝒂 + 𝒃 + 𝒄
Onde:
Cj = coeficiente de Jaccard; a = número de espécies comuns às duas amostras; b = número
de espécies presentes somente na amostra B; c = número de espécies que ocorrem somente
na amostra A.
Os componentes das infracomunidades parasitárias foram classificados de acordo
com Bush e Holmes (1986) em espécies centrais (presentes em mais de 2/3 (66,66%) dos

24
hospedeiros), espécies secundárias (presentes em 1/3 a 2/3 dos hospedeiros) e espécies
satélites (presentes em menos de 1/3(33,33%) dos hospedeiros).
Os índices parasitários e ecológicos foram avaliados para definir parâmetros da
comunidade de parasitas que poderão ser utilizados como bioindicadores de mudanças
ambientais.
Imagens, desenhos e medidas taxonômicas
Os desenhos foram feitos a partir de montagens totais de exemplares em lâminas
permanentes e provisórias em câmara clara acoplada a microscópio de luz com contraste
de fase Olympus BH. As fotomicrografias dos espécimes menores foram feitas em um
microscópio óptico Zeiss com câmera digital acoplada. Todos os desenhos são originais,
exceto os que a autoria é indicada na legenda da figura. Todas as medidas e escalas foram
feitas com o auxílio de uma ocular micrométrica e as medidas apresentam-se em
micrômetros ou milímetros com médias entre parênteses. Quando isso não ocorrer foi
indicada a unidade correspondente.
O material testemunho desse trabalho será depositado na coleção de invertebrados
não-insecta do INPA em Manaus.

25
RESULTADOS
Os resultados foram divididos em duas partes: taxonômica onde foi observada a
diversidade da comunidade componente e a estrutura da comunidade dos parasitas P.
pardalis. Foram analisados 248 indivíduos de P. pardalis capturados em cinco lagos de
várzea da Amazônia. A média do comprimento padrão dos peixes foi 24,5cm ± 8,8 e o
peso médio foi 213,9g ± 102,6. Foram coletados 1.635 espécimens parasitas incluídos em
quatro táxons: Monogenoidea (785); Digenea (795), Cestoda (1), Acanthocephala (50) e
Copepoda (4).
Taxonomia dos metazoários parasitas (Comunidade componente)
Monogenoidea
Foram encontradas três espécies de Monogenoidea: Unilatus brevispinus Suriano,
1985; Heteropriapulus heterotylus Jogunoori, Kritsky & Venkatanarasaiah, 2006;
Trinigyrus mourei Boeger e Jegú, 1989. Das espécies registradas a mais representativa na
infracomunidade de Monogenoidea foi U. brevispinus.
Platyhelminthes Gegenbaur, 1859
Monogenoidea Bychowsky, 1933
Dactylogyridae Bychowsky, 1933
Unilatus Mizelle e Kritsky, 1967
Unilatus brevispinus Suriano, 1985
Comentários: foram medidos 10 espécimes. Corpo longo 544 (408 – 629), fusiforme,
largura 88,6 (61,2 – 119), lobos cefálicos bem desenvolvidos, contendo dois órgãos
cefálicos. Faringe 22,7 (20 – 47). Haptor comprimento 87,1 (51 – 110), largura 71,5 (57,8
– 83). Barra dorsal comprimento 8,1 (5,2 – 11); ventral 10,6 (7,4 – 11). Cirrus
comprimento 88, 6 (72 – 100) e Peça acessória 72,2 (66 – 80). As medidas estão de acordo
com a descrição original da espécie. Foi a espécie com maior prevalência e intensidade
entre as espécies de Monogenoidea.

26
Heteropriapulus heterotylus Jogunoori, Kritsky & Venkatanarasaiah, 2006
Comentários: foram medidos 8 espécimes. Corpo longo 258 (187 – 340), fusiforme,
largura 73,1 (51 – 102), lobos cefálicos bem desenvolvidos, contendo dois órgãos
cefálicos. Faringe não observado. Haptor comprimento 40,3 (34 – 56,1), largura 55,4
(39,1 – 73,1). Barra dorsal comprimento 3,7 (2 – 5); ventral 5,2 (3,4 – 3). Cirrus
comprimento 88,6 (72 – 100) e Peça acessória 72,2 (66 – 80). Espécie menos
representativa do grupo. Menor em comparação as outras espécies. Descrito em peixes
exóticos de aquários na Índia da mesma família de P. pardalis. O cirrus desta espécie é
bastante complexo na sua morfologia, e bastante distinto das demais espécies. As
características morfológicas encontradas estão na média para esta espécie de parasito.
Figura 9 – Desenho de Unilatus brevispinus Suriano, 1985. Escala = 1µm
Figura 10 – Fotomicrografia de Unilatus brevispinus
Suriano, 1985. Escala = 1µm.
27
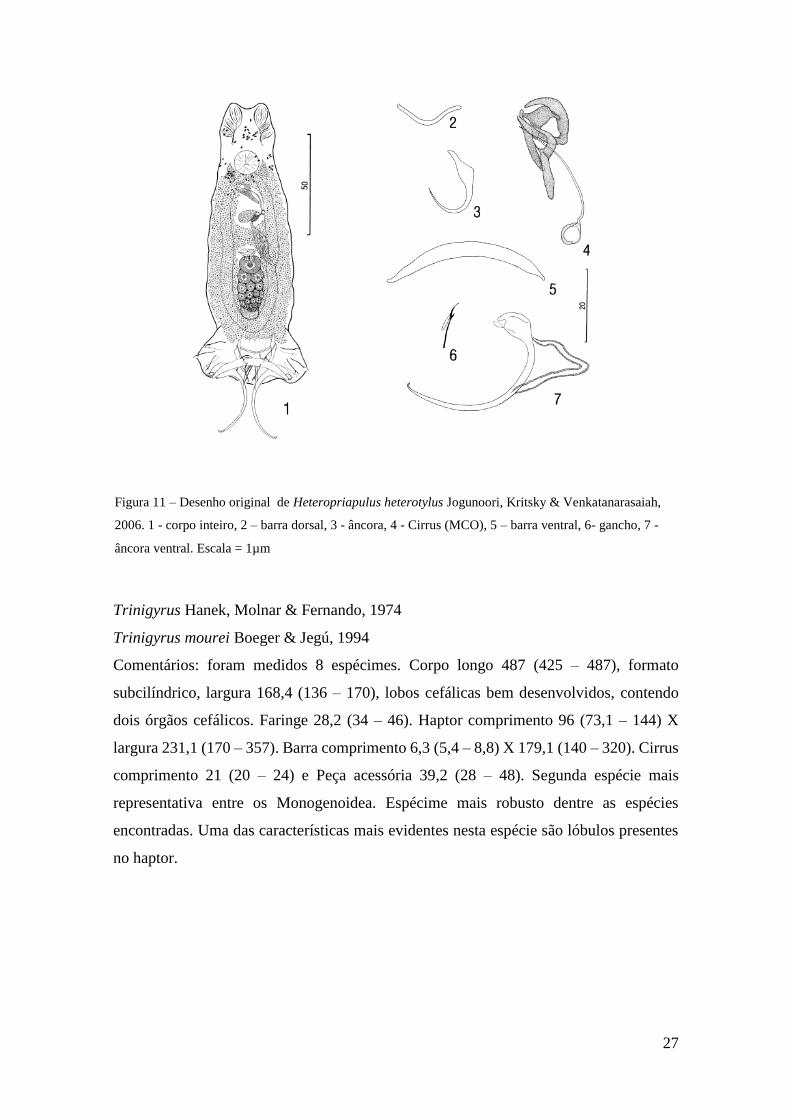
Trinigyrus Hanek, Molnar & Fernando, 1974
Trinigyrus mourei Boeger & Jegú, 1994
Comentários: foram medidos 8 espécimes. Corpo longo 487 (425 – 487), formato
subcilíndrico, largura 168,4 (136 – 170), lobos cefálicas bem desenvolvidos, contendo
dois órgãos cefálicos. Faringe 28,2 (34 – 46). Haptor comprimento 96 (73,1 – 144) X
largura 231,1 (170 – 357). Barra comprimento 6,3 (5,4 – 8,8) X 179,1 (140 – 320). Cirrus
comprimento 21 (20 – 24) e Peça acessória 39,2 (28 – 48). Segunda espécie mais
representativa entre os Monogenoidea. Espécime mais robusto dentre as espécies
encontradas. Uma das características mais evidentes nesta espécie são lóbulos presentes
no haptor.
Figura 11 – Desenho original de Heteropriapulus heterotylus Jogunoori, Kritsky & Venkatanarasaiah,
2006. 1 - corpo inteiro, 2 – barra dorsal, 3 - âncora, 4 - Cirrus (MCO), 5 – barra ventral, 6- gancho, 7 -
âncora ventral. Escala = 1µm

28
DIGENEA
Foram encontradas 06 espécies de Digenea: três em fase de metacercárias e três
adultos. As metacercárias parasitavam os olhos, estômago, gônadas e superfície dos
órgãos internos. As espécies de metacercárias foram: Diplostomum sp.,
Austrodiplostomum compactum, Odhineriotrema microcephala e adultos de
Megacoelium spinispecum, M. spinicavum e Kalitrema kalitrema. Foi o grupo mais
abundante da comunidade componente de P. pardalis.
Trematoda Rudolphi, 1808
Digenea Carus, 1863
Strigeformes Travassos, Freitas & Kohn, 1969
Diplostomidae Poirier, 1886
Diplostominae Monticelli, 1892
Diplostomum Nordmann, 1832
Figura 12 - Desenho de Trinigyrus mourei
Boeger & Jegú, 1994. Escala = 1µm.
Figura 13 - Fotomicrografia em contraste de
fase de Trinigyrus mourei Boeger & Jegú,
1994. Escala = 1 µm.
29
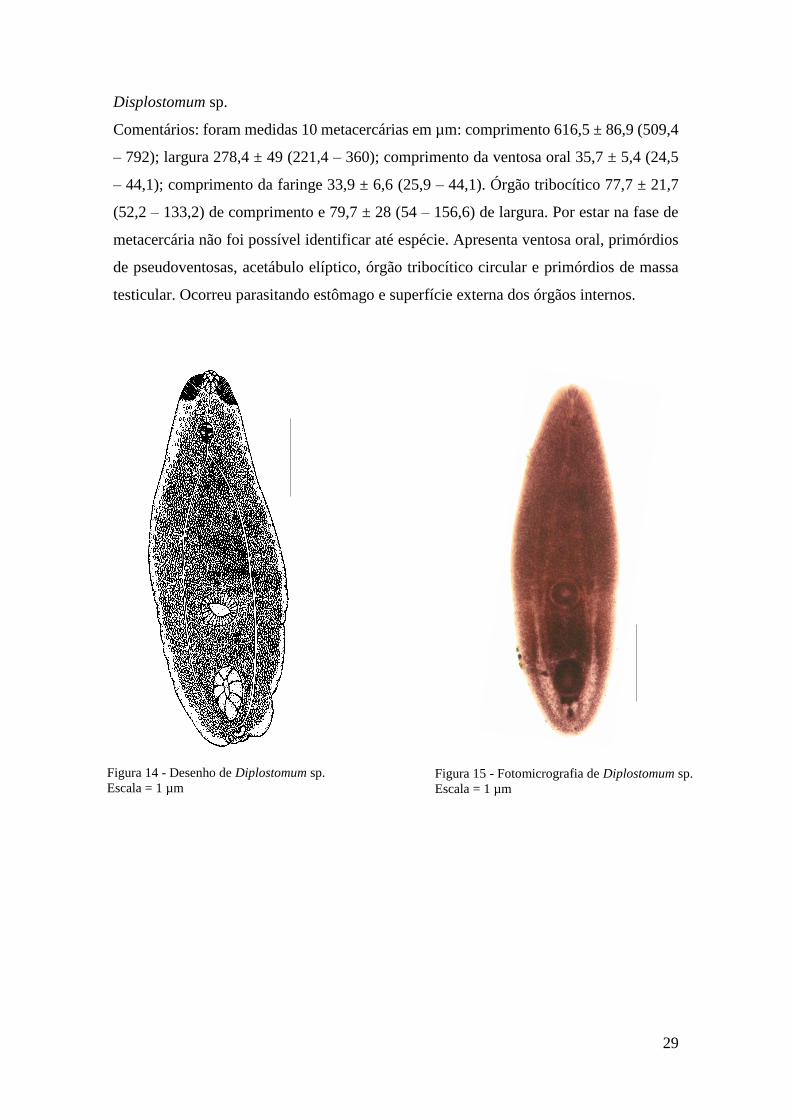
Displostomum sp.
Comentários: foram medidas 10 metacercárias em µm: comprimento 616,5 ± 86,9 (509,4
– 792); largura 278,4 ± 49 (221,4 – 360); comprimento da ventosa oral 35,7 ± 5,4 (24,5
– 44,1); comprimento da faringe 33,9 ± 6,6 (25,9 – 44,1). Órgão tribocítico 77,7 ± 21,7
(52,2 – 133,2) de comprimento e 79,7 ± 28 (54 – 156,6) de largura. Por estar na fase de
metacercária não foi possível identificar até espécie. Apresenta ventosa oral, primórdios
de pseudoventosas, acetábulo elíptico, órgão tribocítico circular e primórdios de massa
testicular. Ocorreu parasitando estômago e superfície externa dos órgãos internos.
Figura 15 - Fotomicrografia de Diplostomum sp.
Escala = 1 µm
Figura 14 - Desenho de Diplostomum sp.
Escala = 1 µm
30
Austrodiplostomum Szidat & Nani, 1951
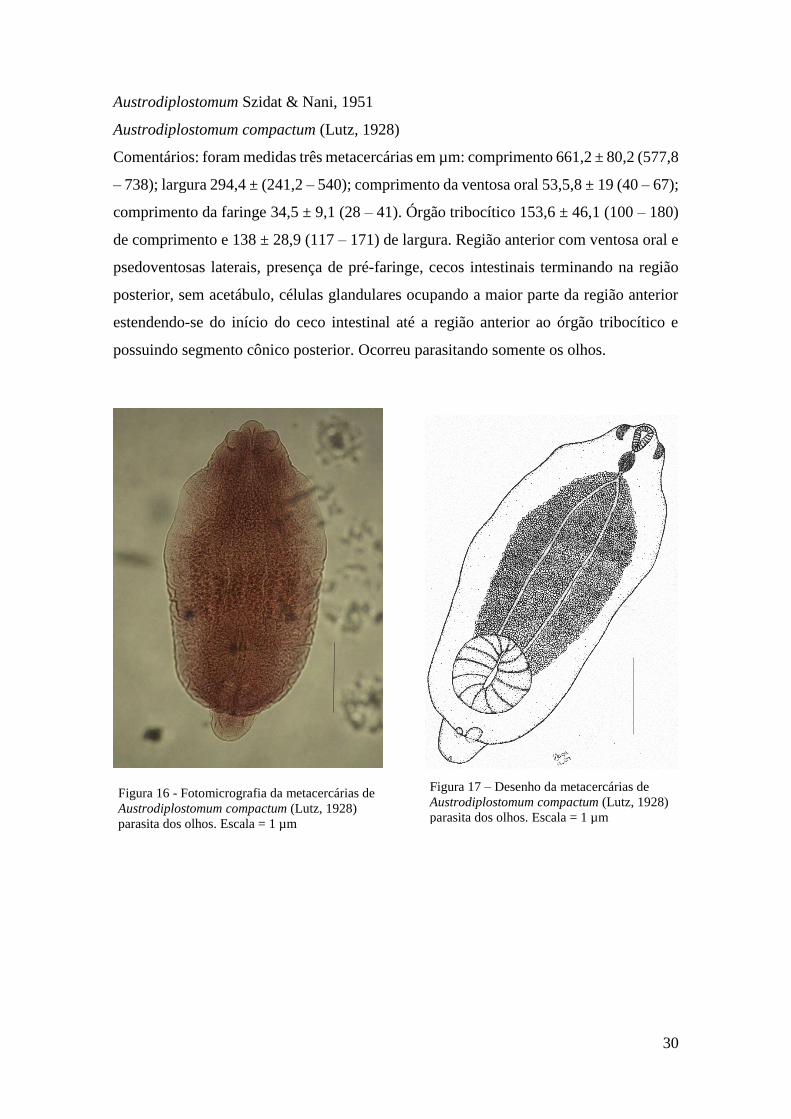
Austrodiplostomum compactum (Lutz, 1928)
Comentários: foram medidas três metacercárias em µm: comprimento 661,2 ± 80,2 (577,8
– 738); largura 294,4 ± (241,2 – 540); comprimento da ventosa oral 53,5,8 ± 19 (40 – 67);
comprimento da faringe 34,5 ± 9,1 (28 – 41). Órgão tribocítico 153,6 ± 46,1 (100 – 180)
de comprimento e 138 ± 28,9 (117 – 171) de largura. Região anterior com ventosa oral e
psedoventosas laterais, presença de pré-faringe, cecos intestinais terminando na região
posterior, sem acetábulo, células glandulares ocupando a maior parte da região anterior
estendendo-se do início do ceco intestinal até a região anterior ao órgão tribocítico e
possuindo segmento cônico posterior. Ocorreu parasitando somente os olhos.
Figura 16 - Fotomicrografia da metacercárias de
Austrodiplostomum compactum (Lutz, 1928) parasita dos olhos. Escala = 1 µm
Figura 17 – Desenho da metacercárias de
Austrodiplostomum compactum (Lutz, 1928)
parasita dos olhos. Escala = 1 µm
31
Haploporidae Nicoll, 1914
Chalcinotrematinae Overstreet & Curran, 2005
Megacoelium Szidat, 1954
Megacoelium spinicavum Thatcher &Varella, 1981
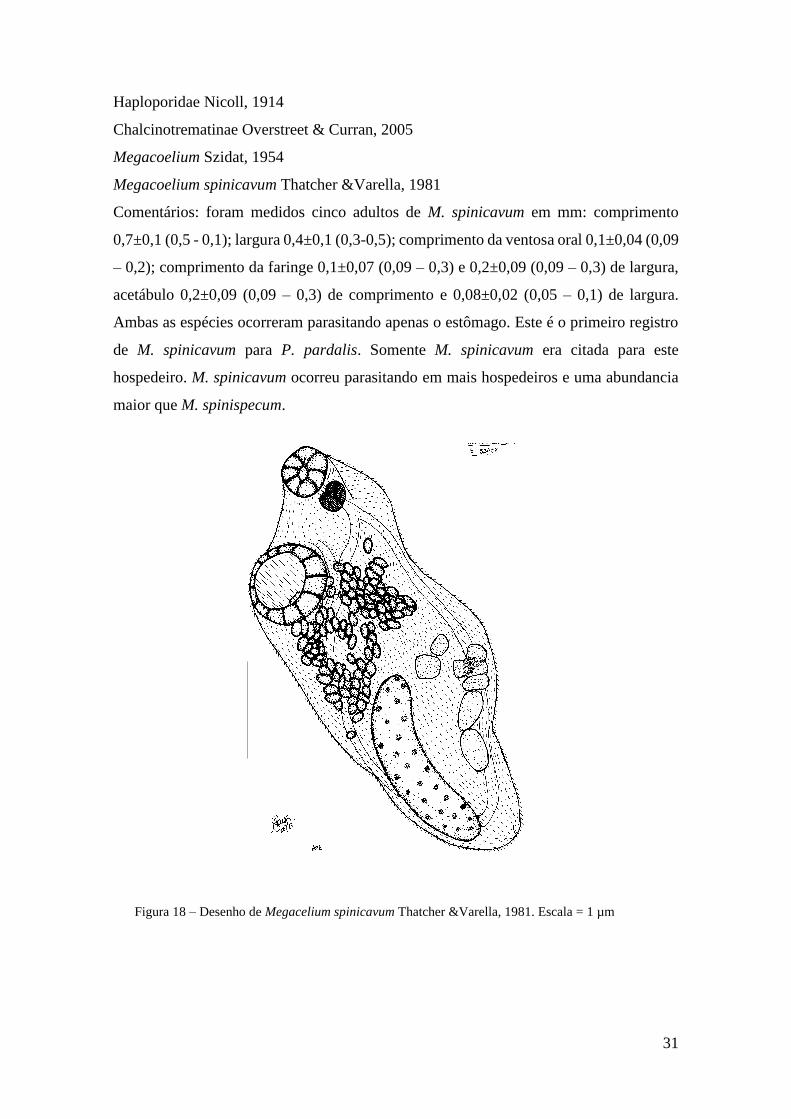
Comentários: foram medidos cinco adultos de M. spinicavum em mm: comprimento
0,7±0,1 (0,5 - 0,1); largura 0,4±0,1 (0,3-0,5); comprimento da ventosa oral 0,1±0,04 (0,09
– 0,2); comprimento da faringe 0,1±0,07 (0,09 – 0,3) e 0,2±0,09 (0,09 – 0,3) de largura,
acetábulo 0,2±0,09 (0,09 – 0,3) de comprimento e 0,08±0,02 (0,05 – 0,1) de largura.
Ambas as espécies ocorreram parasitando apenas o estômago. Este é o primeiro registro
de M. spinicavum para P. pardalis. Somente M. spinicavum era citada para este
hospedeiro. M. spinicavum ocorreu parasitando em mais hospedeiros e uma abundancia
maior que M. spinispecum.
Figura 18 – Desenho de Megacelium spinicavum Thatcher &Varella, 1981. Escala = 1 µm

32
Megacoelium spinispecum Thatcher & Varella, 1981
Comentários: foram medidos sete adultos de M. spinispecum em mm: comprimento 4,1±
1,7 (2,3 - 6); largura 1,7±0,7(0,9 – 2,5); comprimento da ventosa oral 0,3±0,2 (0,1 – 0,6);
comprimento da faringe 0,5±0,2(0,1 – 0,4) e 0,6±0,2(0,1 – 0,4) de largura, acetábulo
0,6±0,2 (0,3 – 0,9) de comprimento e 0,2±0,1 (0,3 – 0,9) de largura.
Figura 19 – Desenho de Megacelium spinispecum Thatcher & Varella, 1981. Escala = 1 µm
33
Figura 20 - Adulto de Odhneriotrema microcephala
(Travassos, 1922) Original Travassos 1969.
Figura21 - Metacercária de Odhneriotrema
microcephala (Travassos, 1922). Escala =
1 µm.
Clinostomidae Luhe, 1901
Odhneriotrematinae Travassos, Freitas & Kohn, 1969
Odhneriotrema Travassos, 1928
Odhneriotrema microcephala (Travassos, 1922)
Comentário: Foram medidos sete metacercárias de O. microcephala em µm:
comprimento 3231 ± 1163,4 (1620 -4680); largura 1134 ± 333(469 –1368); comprimento
da ventosa oral 207 ± 83,4 (108 –342); comprimento da faringe 249 ± 97,5 (0,1 – 0,4) e
111 ± 99,5(108 – 252) de largura, acetábulo 657 ± 145 (396 – 828) de comprimento e 728
± 189 (360 – 900) de largura. Ocorreu parasitando as gônadas na forma de cistos amarelos
bem conspícuos.
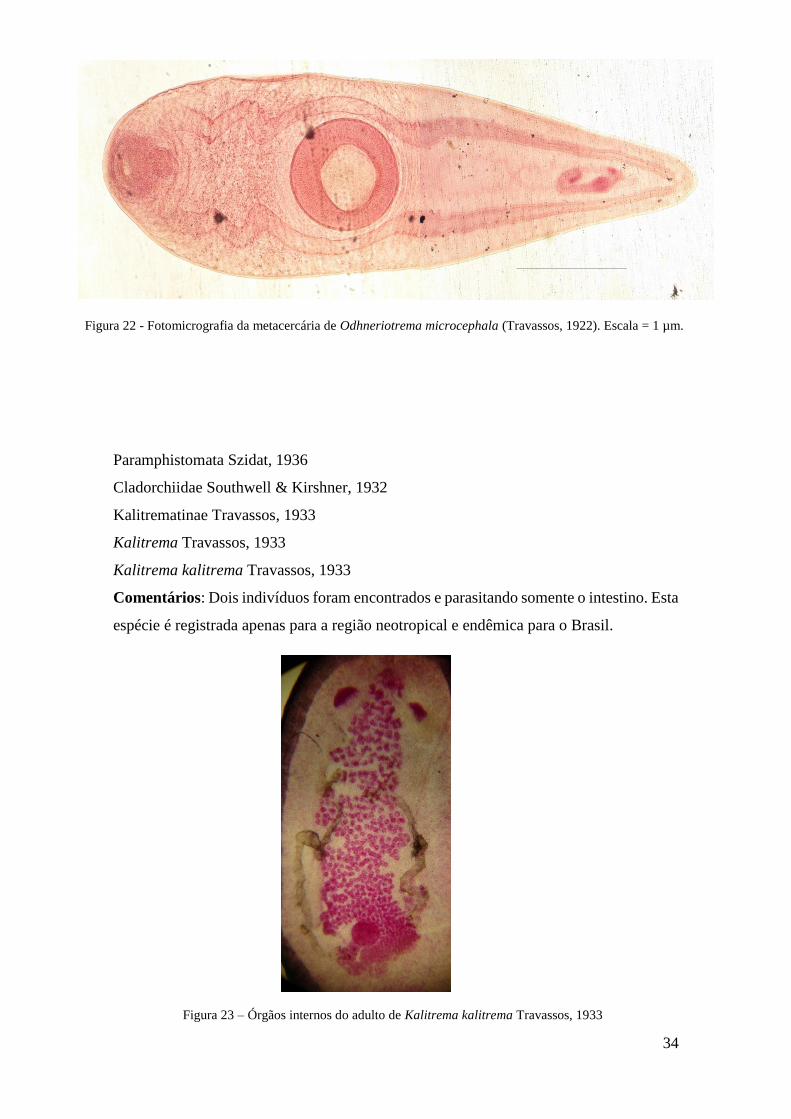
34
Figura 22 - Fotomicrografia da metacercária de Odhneriotrema microcephala (Travassos, 1922). Escala = 1 µm.
Paramphistomata Szidat, 1936
Cladorchiidae Southwell & Kirshner, 1932
Kalitrematinae Travassos, 1933
Kalitrema Travassos, 1933
Kalitrema kalitrema Travassos, 1933
Comentários: Dois indivíduos foram encontrados e parasitando somente o intestino. Esta
espécie é registrada apenas para a região neotropical e endêmica para o Brasil.
Figura 23 – Órgãos internos do adulto de Kalitrema kalitrema Travassos, 1933

35
Figura 24 – Desenho do Adulto de Kalitrema kalitrema Travassos, 1933. Escala = 1 µm.
CESTODA
Cestoda Van Beneden, 1849
Proteocephalidea Mola, 1928
Proteocephalidae La Rue, 1911
Comentário: Os espécimes não foram medidos e nem foi possível desenhá-las pois não
possuíam nenhum órgão interno evidente. Este é o primeiro registro de uma espécie de
Cestoda para P. pardalis. Sendo está espécie fixada na superfície do intestino na forma
de um cisto. Puderam ser observados vários cistos e seu aspecto era semelhantes a
gordura.
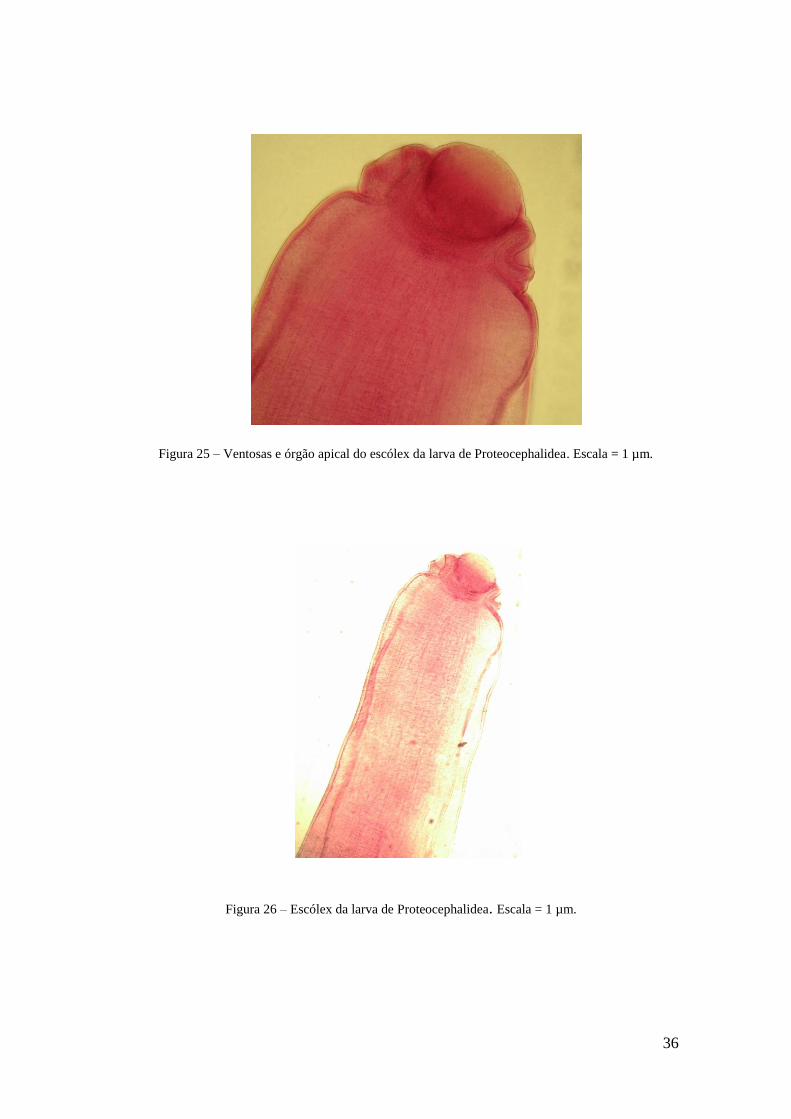
36
Figura 25 – Ventosas e órgão apical do escólex da larva de Proteocephalidea. Escala = 1 µm.
Figura 26 – Escólex da larva de Proteocephalidea. Escala = 1 µm.

37
ACANTHOCEPHALA
Acanthocephala Rudolphi, 1801
Neoechinorhynchidae Van Cleave, 1929
Neoechinorhynchinae Travassos, 1926
Gorytocephalus Nickol e Thatcher, 1971
Gorytocephalus elongorchis Thatcher, 1981
Comentário: Foram medidos oito adultos, um macho e sete fêmeas em mm: fêmeas
comprimento 18,5 ± 5,6 (11,8 – 29,4); largura 1,6 ± 1,2 (0,8 – 4,5); largura da crista 0,08
(0,05 – 0,1); comprimento do lemnisco maior 1,8 ± 1,7 (2,6 – 3,8), lemnisco menor 1,4 ±
1,3 (2,3 – 2,7) de largura; área reprodutiva 0,3±0,06 (0,3 – 0,5). Macho comprimento
11,5; largura 1,2; largura da crista 0,05; comprimento do lemnisco maior 2,8; lemnisco
menor 1,8 de largura; testículo anterior 1,6 x 0,3 e posterior 0,9 x 0,2. Área reprodutiva
0,4. Ocorreu parasitando o intestino, nenhum espécime foi encontrado fixado na parede
do intestino.
A
B
Figura 28– A) Adulto macho de Gorytocephalus elongorchis Thatcher, 1981, B) Detalhe probóscide.
(Desenho original Thatcher, 1981). Escala = 1mm.
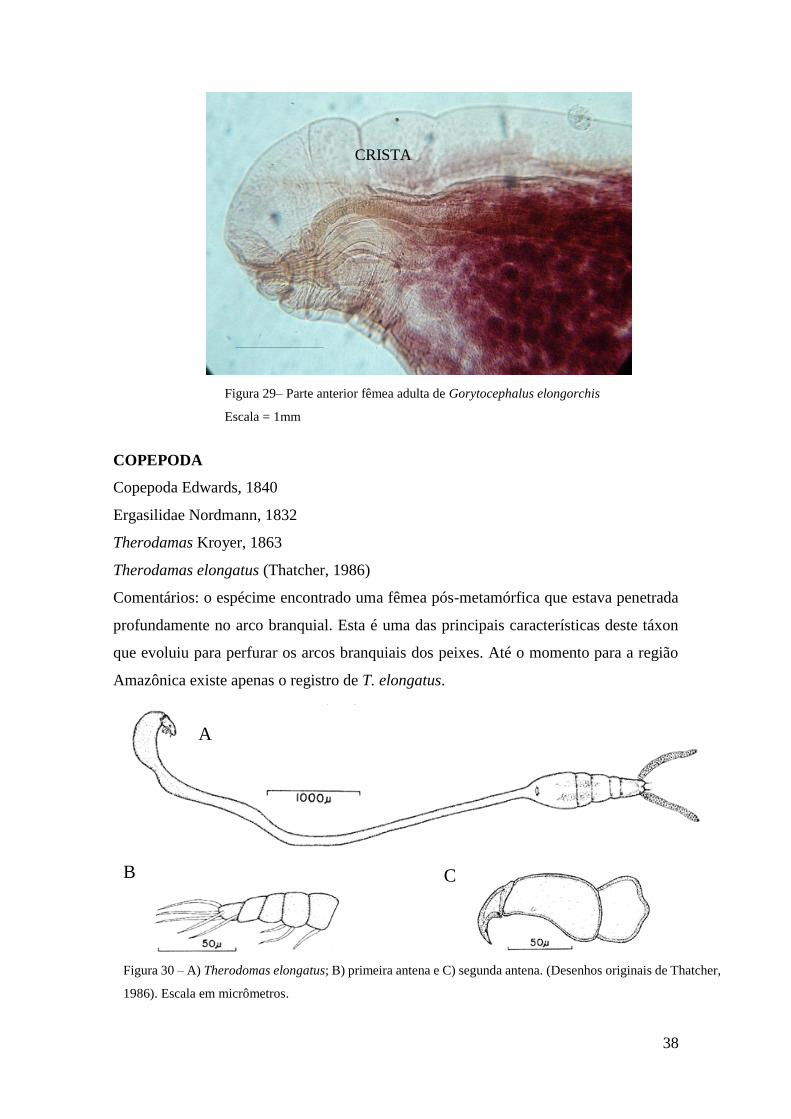
38
COPEPODA
Copepoda Edwards, 1840
Ergasilidae Nordmann, 1832
Therodamas Kroyer, 1863
Therodamas elongatus (Thatcher, 1986)
Comentários: o espécime encontrado uma fêmea pós-metamórfica que estava penetrada
profundamente no arco branquial. Esta é uma das principais características deste táxon
que evoluiu para perfurar os arcos branquiais dos peixes. Até o momento para a região
Amazônica existe apenas o registro de T. elongatus.
B C
CRISTA
Figura 30 – A) Therodomas elongatus; B) primeira antena e C) segunda antena. (Desenhos originais de Thatcher,
1986). Escala em micrômetros.
Figura 29– Parte anterior fêmea adulta de Gorytocephalus elongorchis
Escala = 1mm
A

39
Biodiversidade dos metazoários parasitas (Comunidade componente)
Foram analisados 248 espécimes de P. pardalis coletados em cinco lagos de
várzea ao longo do rio Solimões. Espécies de cinco táxons foram encontradas parasitando
P. pardalis: Monogenoidea, Digenea, Cestoda, Acanthocephala e Copepoda. Digenea foi
o táxon com maior diversidade com 50% das espécies (6 espécies), seguido por
Monogenoidea 25% (3 espécies), Cestoda (1), Acanthocephala (1) e Copepoda (1) com
8, 33% (Tabela 01).
Dos doze taxons parasitas de P. pardalis 8 espécies eram adultas, 3 na fase de
metacercárias e uma na fase de larva. Três espécies tiveram os maiores valores de
prevalência (acima de 10%) e intensidade média: de Monogenoidea: U. brevispinus;
Digenea: Diplostomum sp. e o Acanthocephala: G. elongorchis (Tabela 01).
Entre os lagos amostrados U. brevispinus e Diplostomum sp. foram as espécies
parasitas que apresentaram os maiores índices parasitários. Unilatus brevispinus
apresentou sua maior prevalência no lago Baixio, a maior intensidade média no lago
Ananá e a maior abundância média no lago Preto. Diplostomum sp. apresentou sua maior
valor de prevalência no lago Baixio, a maior intensidade média no lago Preto e a maior
abundância média no lago Ananá (Tabela 02).
Das espécies identificadas, nenhuma foi considerada central em nenhum dos
lagos. As espécies apresentaram uma classificação de importância como Secundaria e
Satélite entre os lagos (Tabela 02).
Diplostomum sp. foi a única espécie de parasita que ocorreu parasitando três
órgãos: estômago, gônadas e superfície dos órgãos internos (cavidade interna) (Tabela
03). As brânquias foram os órgãos mais parasitados (4 espécies), estômago (três
espécies), intestino (duas espécies) e gônadas duas espécies. Nos olhos ocorreu uma
espécie A. compactum (Tabela 01).
Três espécies parasitas ocorreram somente em peixes de um lago: K. kalitrema no
lago Preto, Proteocephalus sp. no lago Ananá e T. elongatus no lago Araçá. Todas as
demais ocorreram nos cinco lagos amostrados. Este é o primeiro registro destas espécies
para P. pardalis.
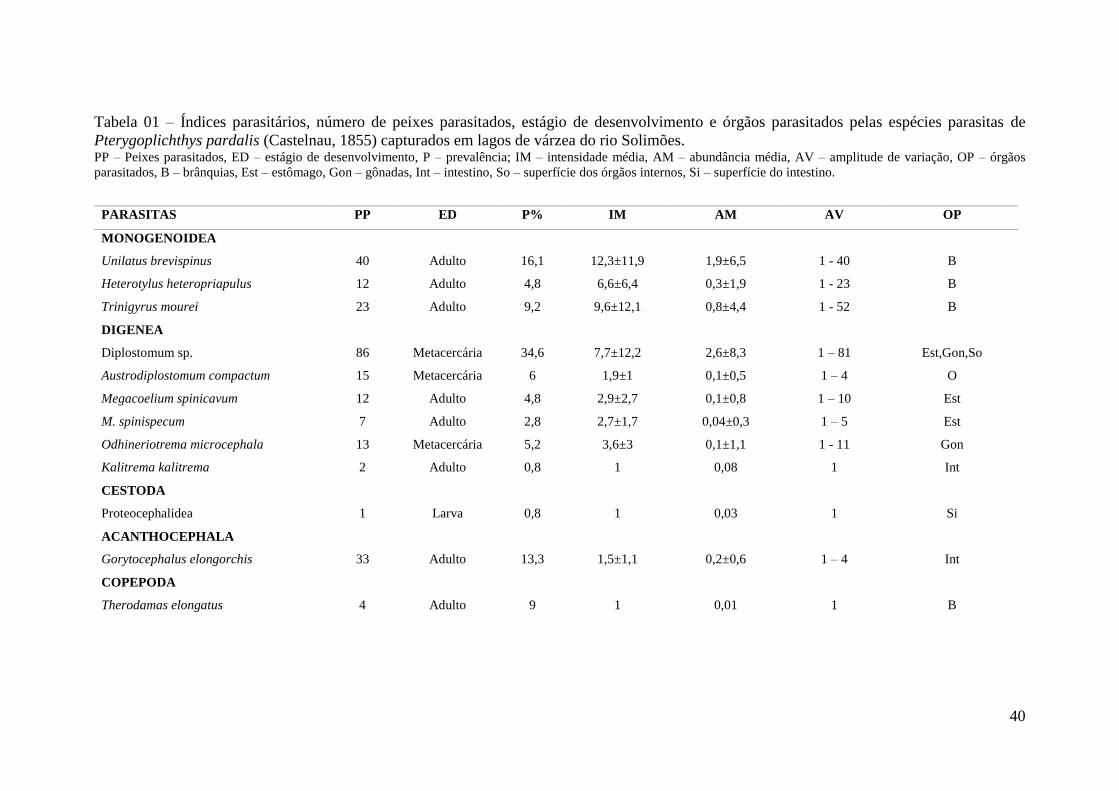
40
Tabela 01 – Índices parasitários, número de peixes parasitados, estágio de desenvolvimento e órgãos parasitados pelas espécies parasitas de
Pterygoplichthys pardalis (Castelnau, 1855) capturados em lagos de várzea do rio Solimões. PP – Peixes parasitados, ED – estágio de desenvolvimento, P – prevalência; IM – intensidade média, AM – abundância média, AV – amplitude de variação, OP – órgãos
parasitados, B – brânquias, Est – estômago, Gon – gônadas, Int – intestino, So – superfície dos órgãos internos, Si – superfície do intestino.
PARASITAS PP ED P% IM AM AV OP
MONOGENOIDEA
Unilatus brevispinus 40 Adulto 16,1 12,3±11,9 1,9±6,5 1 - 40 B
Heterotylus heteropriapulus 12 Adulto 4,8 6,6±6,4 0,3±1,9 1 - 23 B
Trinigyrus mourei 23 Adulto 9,2 9,6±12,1 0,8±4,4 1 - 52 B
DIGENEA
Diplostomum sp. 86 Metacercária 34,6 7,7±12,2 2,6±8,3 1 – 81 Est,Gon,So
Austrodiplostomum compactum 15 Metacercária 6 1,9±1 0,1±0,5 1 – 4 O
Megacoelium spinicavum 12 Adulto 4,8 2,9±2,7 0,1±0,8 1 – 10 Est
M. spinispecum 7 Adulto 2,8 2,7±1,7 0,04±0,3 1 – 5 Est
Odhineriotrema microcephala 13 Metacercária 5,2 3,6±3 0,1±1,1 1 - 11 Gon
Kalitrema kalitrema 2 Adulto 0,8 1 0,08 1 Int
CESTODA
Proteocephalidea 1 Larva 0,8 1 0,03 1 Si
ACANTHOCEPHALA
Gorytocephalus elongorchis 33 Adulto 13,3 1,5±1,1 0,2±0,6 1 – 4 Int
COPEPODA
Therodamas elongatus 4 Adulto 9 1 0,01 1 B

41
No geral os descritores ecológicos não variaram consideravelmente entre os lagos.
Pequenas variações foram detectadas quando comparados os índices entre os mesmos
lagos durante as fases do período hidrológico (Tabela 05, 06, 07 e 08).
A maior riqueza de espécies parasitas ocorreu no período da enchente no lago São
Tomé, com 9 espécies. A menor ocorreu também no período da enchente, no lago Baixio
com apenas uma espécie, Diplostomum sp. (Tabela 05).
O valor estimado pelo índice de Shannon com dados de abundância foi maior no
período da vazante no lago Baixio (H’=1,67). O menor valor foi no período de seca no
lago Araçá (H’= 0,09) (Tabela 07 e 08).
O maior valor de riqueza específica estimado pelo índice de Margalef foi de (DMg
= 1,54) ocorreu no lago Araçá no período da enchente. O menor (DMg = 0,36) no lago
Ananá no período da seca (Tabela 05 e 08).
A dominância estimada pelo índice de Berger-Parker (d) foi maior no lago Araçá
no período da seca d = 0,98. O menor foi também no lago Araçá, no período da vazante
d = 0,27 (Tabela 07 e 08) ambos para Diplostomum sp.
O maior valor de equitatividade foi registrado no período da enchente no lago
Araçá J = 0,95. O menor valor registrado para este índice foi no lago Ananá no período
da seca J = 0,14. (Tabela 05 e 08).
A comunidade parasitária de P. pardalis apresentou concentração para a
dominância nas fases do período hidrológico avaliada por meio do índice de Simpson (C)
com valores de C = 25 na enchente, C = 0,53 no período da cheia, C = 0,31 no período
da vazante e C = 0, 88 no período da seca para Diplostomum sp.
Dentre os componentes das infracomunidade de P. pardalis houve a dominância
por duas espécies durante as fases do período hidrológico. A primeira o monogenóideo
U. brevispinus na fase enchente (56,1 %) e vazante (81,6). A segunda o digenético
Diplostomum sp. na fase de cheia (73,2 %) e seca (93%) (Tabela 09).
O resultado do índice de dispersão (ID) mostrou que a maioria das espécies (7)
apresentou uma dispersão agregada nas fases do período hidrológico. O índice de Green
(IG) mostrou que todas as espécies apresentaram uma dispersão aleatória.
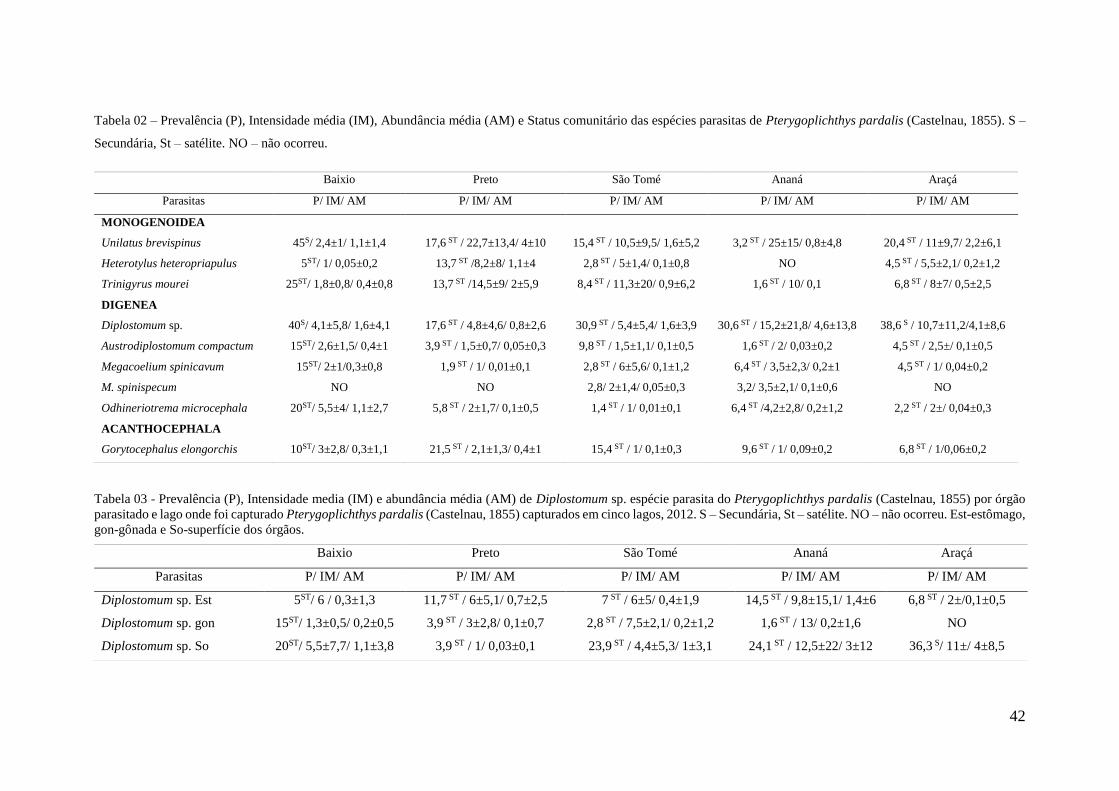
42
Tabela 02 – Prevalência (P), Intensidade média (IM), Abundância média (AM) e Status comunitário das espécies parasitas de Pterygoplichthys pardalis (Castelnau, 1855). S –
Secundária, St – satélite. NO – não ocorreu.
Tabela 03 - Prevalência (P), Intensidade media (IM) e abundância média (AM) de Diplostomum sp. espécie parasita do Pterygoplichthys pardalis (Castelnau, 1855) por órgão
parasitado e lago onde foi capturado Pterygoplichthys pardalis (Castelnau, 1855) capturados em cinco lagos, 2012. S – Secundária, St – satélite. NO – não ocorreu. Est-estômago,
gon-gônada e So-superfície dos órgãos.
Baixio Preto São Tomé Ananá Araçá
Parasitas P/ IM/ AM P/ IM/ AM P/ IM/ AM P/ IM/ AM P/ IM/ AM
MONOGENOIDEA
Unilatus brevispinus 45S/ 2,4±1/ 1,1±1,4 17,6 ST / 22,7±13,4/ 4±10 15,4 ST / 10,5±9,5/ 1,6±5,2 3,2 ST / 25±15/ 0,8±4,8 20,4 ST / 11±9,7/ 2,2±6,1
Heterotylus heteropriapulus 5ST/ 1/ 0,05±0,2 13,7 ST /8,2±8/ 1,1±4 2,8 ST / 5±1,4/ 0,1±0,8 NO 4,5 ST / 5,5±2,1/ 0,2±1,2
Trinigyrus mourei 25ST/ 1,8±0,8/ 0,4±0,8 13,7 ST /14,5±9/ 2±5,9 8,4 ST / 11,3±20/ 0,9±6,2 1,6 ST / 10/ 0,1 6,8 ST / 8±7/ 0,5±2,5
DIGENEA
Diplostomum sp. 40S/ 4,1±5,8/ 1,6±4,1 17,6 ST / 4,8±4,6/ 0,8±2,6 30,9 ST / 5,4±5,4/ 1,6±3,9 30,6 ST / 15,2±21,8/ 4,6±13,8 38,6 S / 10,7±11,2/4,1±8,6
Austrodiplostomum compactum 15ST/ 2,6±1,5/ 0,4±1 3,9 ST / 1,5±0,7/ 0,05±0,3 9,8 ST / 1,5±1,1/ 0,1±0,5 1,6 ST / 2/ 0,03±0,2 4,5 ST / 2,5±/ 0,1±0,5
Megacoelium spinicavum 15ST/ 2±1/0,3±0,8 1,9 ST / 1/ 0,01±0,1 2,8 ST / 6±5,6/ 0,1±1,2 6,4 ST / 3,5±2,3/ 0,2±1 4,5 ST / 1/ 0,04±0,2
M. spinispecum NO NO 2,8/ 2±1,4/ 0,05±0,3 3,2/ 3,5±2,1/ 0,1±0,6 NO
Odhineriotrema microcephala 20ST/ 5,5±4/ 1,1±2,7 5,8 ST / 2±1,7/ 0,1±0,5 1,4 ST / 1/ 0,01±0,1 6,4 ST /4,2±2,8/ 0,2±1,2 2,2 ST / 2±/ 0,04±0,3
ACANTHOCEPHALA
Gorytocephalus elongorchis 10ST/ 3±2,8/ 0,3±1,1 21,5 ST / 2,1±1,3/ 0,4±1 15,4 ST / 1/ 0,1±0,3 9,6 ST / 1/ 0,09±0,2 6,8 ST / 1/0,06±0,2
Baixio Preto São Tomé Ananá Araçá
Parasitas P/ IM/ AM P/ IM/ AM P/ IM/ AM P/ IM/ AM P/ IM/ AM
Diplostomum sp. Est 5ST/ 6 / 0,3±1,3 11,7 ST / 6±5,1/ 0,7±2,5 7 ST / 6±5/ 0,4±1,9 14,5 ST / 9,8±15,1/ 1,4±6 6,8 ST / 2±/0,1±0,5
Diplostomum sp. gon 15ST/ 1,3±0,5/ 0,2±0,5 3,9 ST / 3±2,8/ 0,1±0,7 2,8 ST / 7,5±2,1/ 0,2±1,2 1,6 ST / 13/ 0,2±1,6 NO
Diplostomum sp. So 20ST/ 5,5±7,7/ 1,1±3,8 3,9 ST / 1/ 0,03±0,1 23,9 ST / 4,4±5,3/ 1±3,1 24,1 ST / 12,5±22/ 3±12 36,3 S/ 11±/ 4±8,5

43
Os maiores valores do índice de dispersão foram para: U. brevispinus na fase da seca (ID
= 45), H. heteropriapulus na fase da cheia (ID = 32, 9) e Diplostomum sp. na fase da seca (ID =
30,6) (tabela 10).
O coeficiente de associação de Jaccard calculado para verificar o grau de similaridade das
espécies entre os pontos de coleta mostrou uma baixa similaridade entre os lagos durante as fases
do período hidrológico (Tabela 11).
Das doze espécies encontradas na comunidade componente, duas espécies destacaram.
Elas ocorreram parasitando o maior número de hospedeiros e apresentaram as maiores
prevalências e intensidade médias: Diplostomum sp. e U. brevispinus.
Destas duas espécies, apenas Diplostomum sp. se adequa aos requisitos propostos por
(Overstreet 1997) para espécies parasitas serem usadas bioindicadoras de mudanças ambientais.
Porém como as espécies ectoparasitas são boas indicadoras de hidrocarboneto (Khan e Thulin,
1991) pois em contato com estes compostos sua fisiologia é estimulada a reprodução. Como estas
duas espécies destacarem-se das demais, a estrutura de suas populações foram avaliadas mais
detalhadamente.
A variação da prevalência durante o período hidrológico indicou que Diplostomum sp.
apresentou as maiores prevalências, em P. pardalis, na fase da cheia e seca. Unilatus brevispinus
as menores prevalências, em P. pardalis, no período da seca (Figura 30).
A variação da intensidade média durante o período hidrológico indicou que Diplostomum
sp. apresentou as maiores intensidades médias, em P. pardalis, na fase da cheia e seca. Unilatus
brevispinus teve as maiores intensidades médias, em P. pardalis, no período de enchente e cheia
(Figura 31).
A variação da abundância durante o período hidrológico indicou que Diplostomum sp.
apresentou as maiores abundâncias, em P. pardalis, na fase da seca. Unilatus brevispinus teve as
maiores abundâncias, em P. pardalis, no período da enchente (Figura 32).
Analisando os resultados da variação da prevalência de Diplostomum sp., durante as fases
do período hidrológico para cada lago, Diplostomum sp. apresentou baixa prevalência indicando
não ocorrer em muitos exemplares de P. pardalis (Figura 33). A intensidade média foi maior em
P. pardalis de três lagos: São Tomé, Ananá e Araçá, nos quatro períodos hidrológicos (Figura
34). A maior abundância ocorreu em P. pardalis do lago Ananá no período da seca (Figura 35).
Analisando os resultados da variação da prevalência de U. brevispinus, durante as fases
do período hidrológico, para cada lago. Unilatus brevispinus teve baixa prevalência indicando
não ocorrer em muitos P. pardalis, mas teve alta prevalência em P. pardalis de dois lagos o
Ananá e o Araçá, no período da cheia (Figura 36). A intensidade média foi maior em P. pardalis

44
de três lagos, o Preto, o Ananá e o Araçá, nos períodos da enchente e cheia (Figura 37). A maior
abundância ocorreu em P. pardalis do lago Preto, no período da enchente. Nos demais lagos
abundância foi baixa (Figura 38).
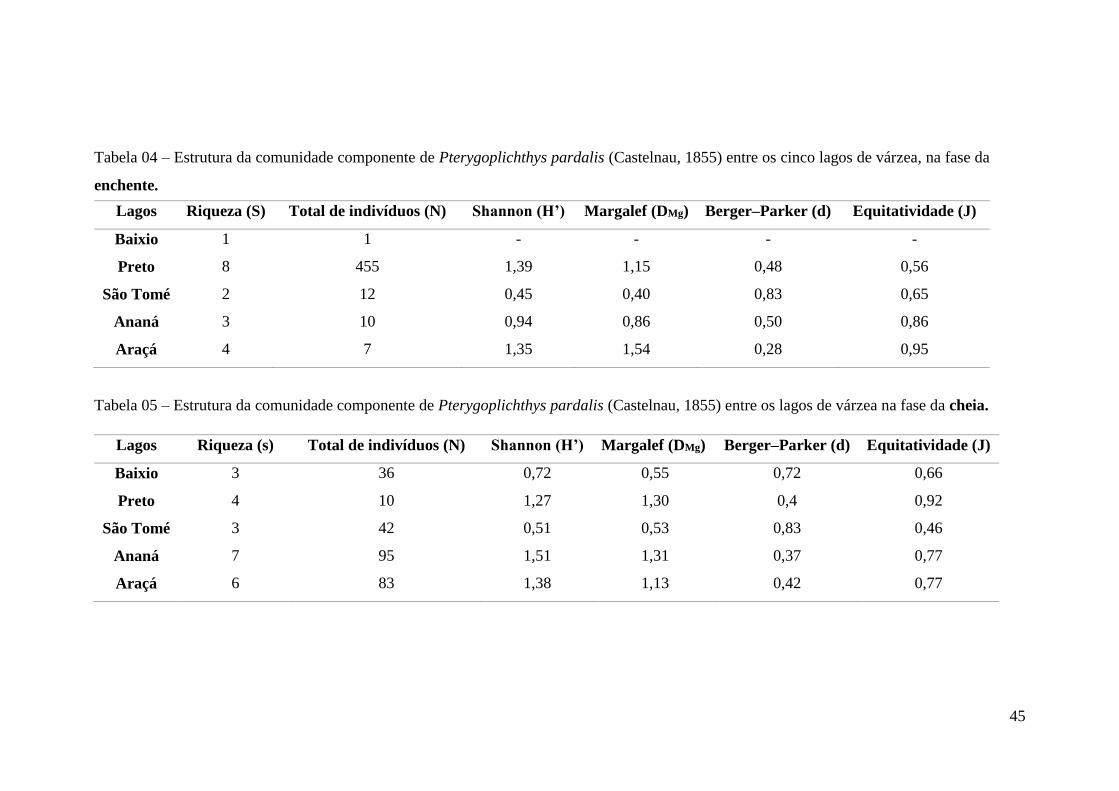
45
Tabela 04 – Estrutura da comunidade componente de Pterygoplichthys pardalis (Castelnau, 1855) entre os cinco lagos de várzea, na fase da
enchente.
Tabela 05 – Estrutura da comunidade componente de Pterygoplichthys pardalis (Castelnau, 1855) entre os lagos de várzea na fase da cheia.
Lagos Riqueza (S) Total de indivíduos (N) Shannon (H’) Margalef (DMg) Berger–Parker (d) Equitatividade (J)
Baixio 1 1 - - - -
Preto 8 455 1,39 1,15 0,48 0,56
São Tomé 2 12 0,45 0,40 0,83 0,65
Ananá 3 10 0,94 0,86 0,50 0,86
Araçá 4 7 1,35 1,54 0,28 0,95
Lagos Riqueza (s) Total de indivíduos (N) Shannon (H’) Margalef (DMg) Berger–Parker (d) Equitatividade (J)
Baixio 3 36 0,72 0,55 0,72 0,66
Preto 4 10 1,27 1,30 0,4 0,92
São Tomé 3 42 0,51 0,53 0,83 0,46
Ananá 7 95 1,51 1,31 0,37 0,77
Araçá 6 83 1,38 1,13 0,42 0,77
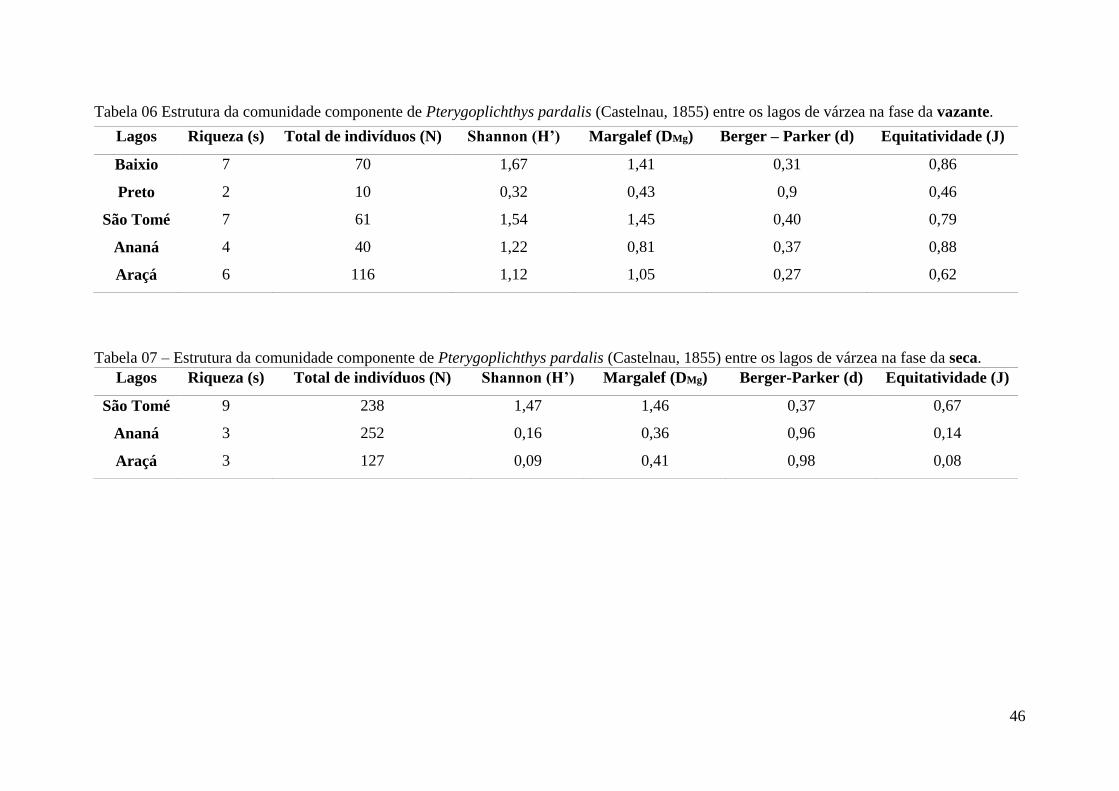
46
Tabela 06 Estrutura da comunidade componente de Pterygoplichthys pardalis (Castelnau, 1855) entre os lagos de várzea na fase da vazante.
Tabela 07 – Estrutura da comunidade componente de Pterygoplichthys pardalis (Castelnau, 1855) entre os lagos de várzea na fase da seca.
Lagos Riqueza (s) Total de indivíduos (N) Shannon (H’) Margalef (DMg) Berger – Parker (d) Equitatividade (J)
Baixio 7 70 1,67 1,41 0,31 0,86
Preto 2 10 0,32 0,43 0,9 0,46
São Tomé 7 61 1,54 1,45 0,40 0,79
Ananá 4 40 1,22 0,81 0,37 0,88
Araçá 6 116 1,12 1,05 0,27 0,62
Lagos Riqueza (s) Total de indivíduos (N) Shannon (H’) Margalef (DMg) Berger-Parker (d) Equitatividade (J)
São Tomé 9 238 1,47 1,46 0,37 0,67
Ananá 3 252 0,16 0,36 0,96 0,14
Araçá 3 127 0,09 0,41 0,98 0,08
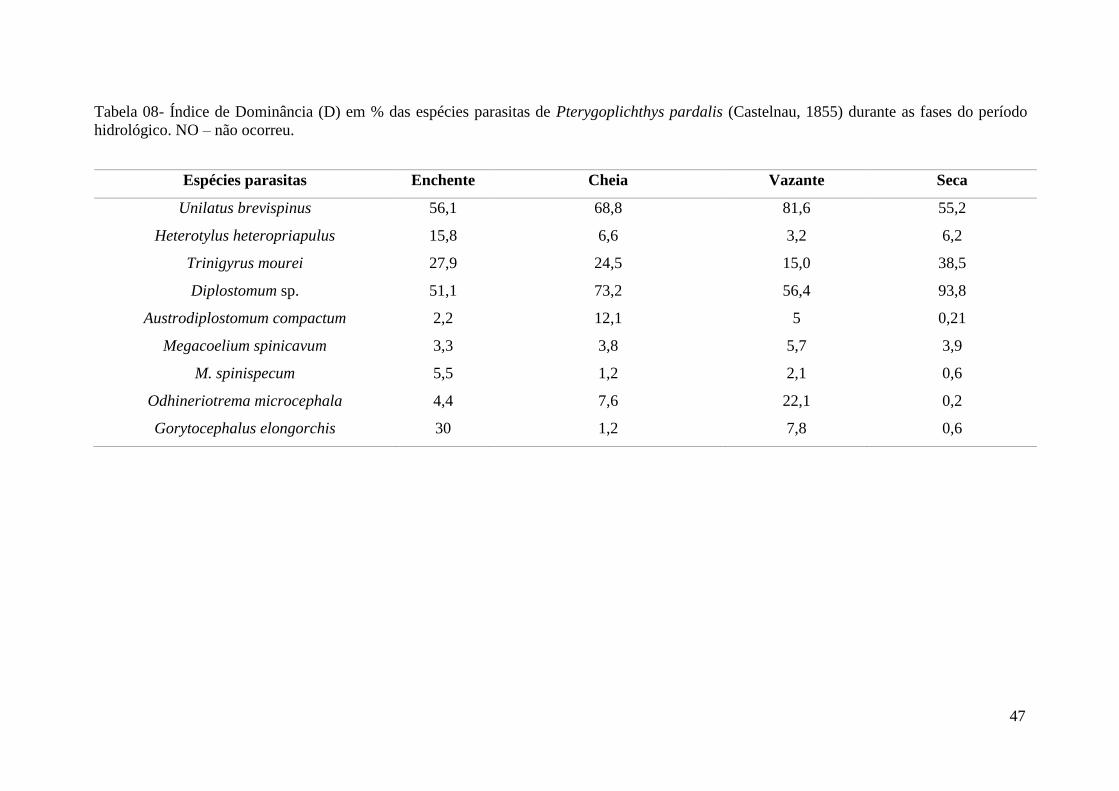
47
Tabela 08- Índice de Dominância (D) em % das espécies parasitas de Pterygoplichthys pardalis (Castelnau, 1855) durante as fases do período
hidrológico. NO – não ocorreu.
Espécies parasitas Enchente Cheia Vazante Seca
Unilatus brevispinus 56,1 68,8 81,6 55,2
Heterotylus heteropriapulus 15,8 6,6 3,2 6,2
Trinigyrus mourei 27,9 24,5 15,0 38,5
Diplostomum sp. 51,1 73,2 56,4 93,8
Austrodiplostomum compactum 2,2 12,1 5 0,21
Megacoelium spinicavum 3,3 3,8 5,7 3,9
M. spinispecum 5,5 1,2 2,1 0,6
Odhineriotrema microcephala 4,4 7,6 22,1 0,2
Gorytocephalus elongorchis 30 1,2 7,8 0,6
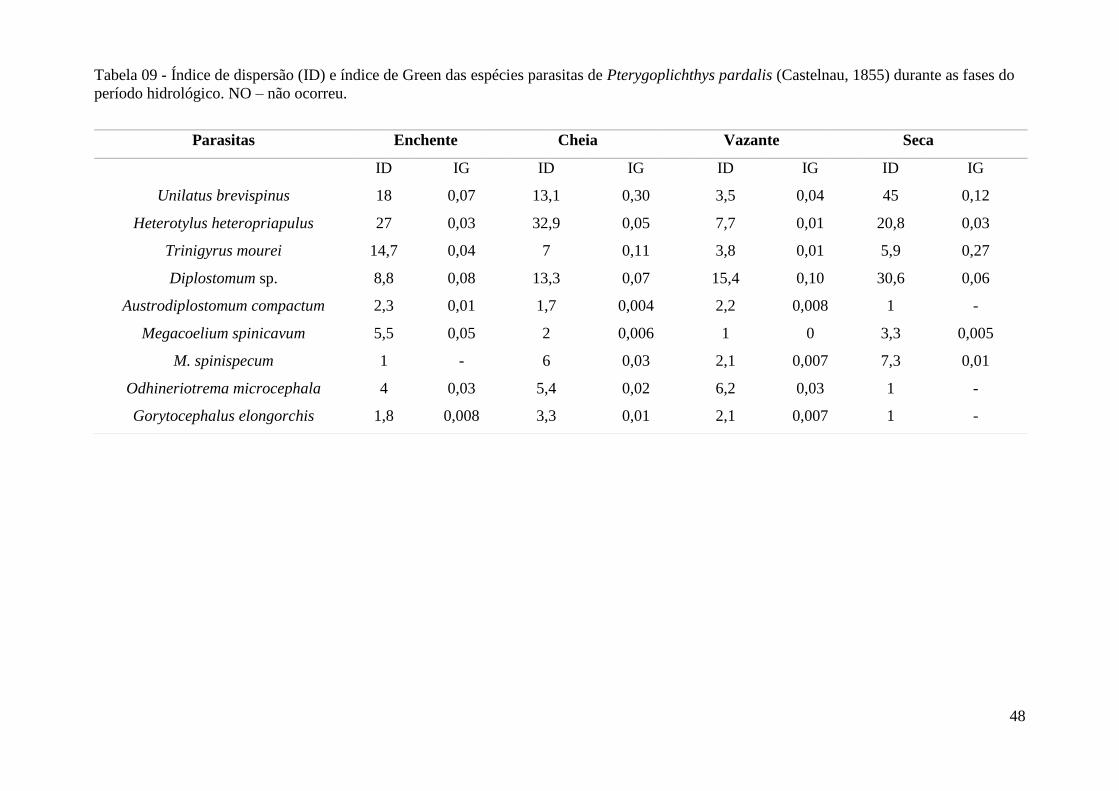
48
Tabela 09 - Índice de dispersão (ID) e índice de Green das espécies parasitas de Pterygoplichthys pardalis (Castelnau, 1855) durante as fases do
período hidrológico. NO – não ocorreu.
Parasitas Enchente Cheia Vazante Seca
ID IG ID IG ID IG ID IG
Unilatus brevispinus 18 0,07 13,1 0,30 3,5 0,04 45 0,12
Heterotylus heteropriapulus 27 0,03 32,9 0,05 7,7 0,01 20,8 0,03
Trinigyrus mourei 14,7 0,04 7 0,11 3,8 0,01 5,9 0,27
Diplostomum sp. 8,8 0,08 13,3 0,07 15,4 0,10 30,6 0,06
Austrodiplostomum compactum 2,3 0,01 1,7 0,004 2,2 0,008 1 -
Megacoelium spinicavum 5,5 0,05 2 0,006 1 0 3,3 0,005
M. spinispecum 1 - 6 0,03 2,1 0,007 7,3 0,01
Odhineriotrema microcephala 4 0,03 5,4 0,02 6,2 0,03 1 -
Gorytocephalus elongorchis 1,8 0,008 3,3 0,01 2,1 0,007 1 -
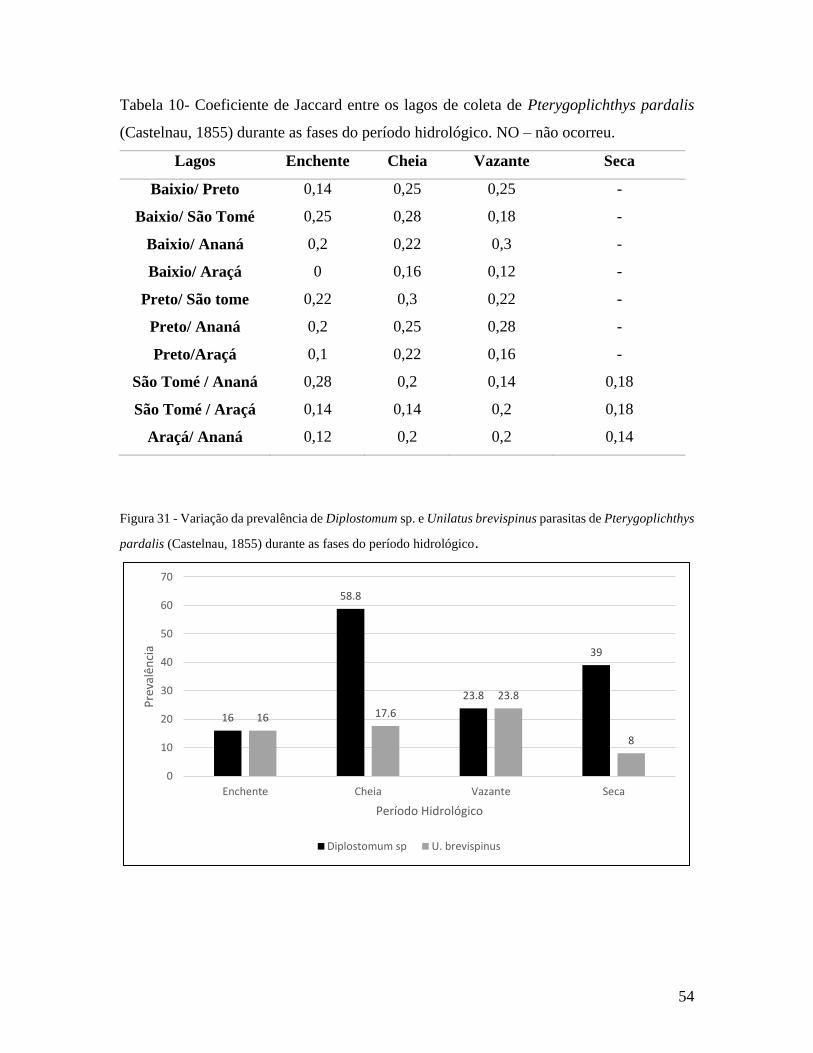
54
Tabela 10- Coeficiente de Jaccard entre os lagos de coleta de Pterygoplichthys pardalis
(Castelnau, 1855) durante as fases do período hidrológico. NO – não ocorreu.
Lagos Enchente Cheia Vazante Seca
Baixio/ Preto 0,14 0,25 0,25 -
Baixio/ São Tomé 0,25 0,28 0,18 -
Baixio/ Ananá 0,2 0,22 0,3 -
Baixio/ Araçá 0 0,16 0,12 -
Preto/ São tome 0,22 0,3 0,22 -
Preto/ Ananá 0,2 0,25 0,28 -
Preto/Araçá 0,1 0,22 0,16 -
São Tomé / Ananá 0,28 0,2 0,14 0,18
São Tomé / Araçá 0,14 0,14 0,2 0,18
Araçá/ Ananá 0,12 0,2 0,2 0,14
Figura 31 - Variação da prevalência de Diplostomum sp. e Unilatus brevispinus parasitas de Pterygoplichthys
pardalis (Castelnau, 1855) durante as fases do período hidrológico.
16
58.8
23.8
39
16 17.6
23.8
8
0
10
20
30
40
50
60
70
Enchente Cheia Vazante Seca
Pre
valê
nci
a
Período Hidrológico
Diplostomum sp U. brevispinus
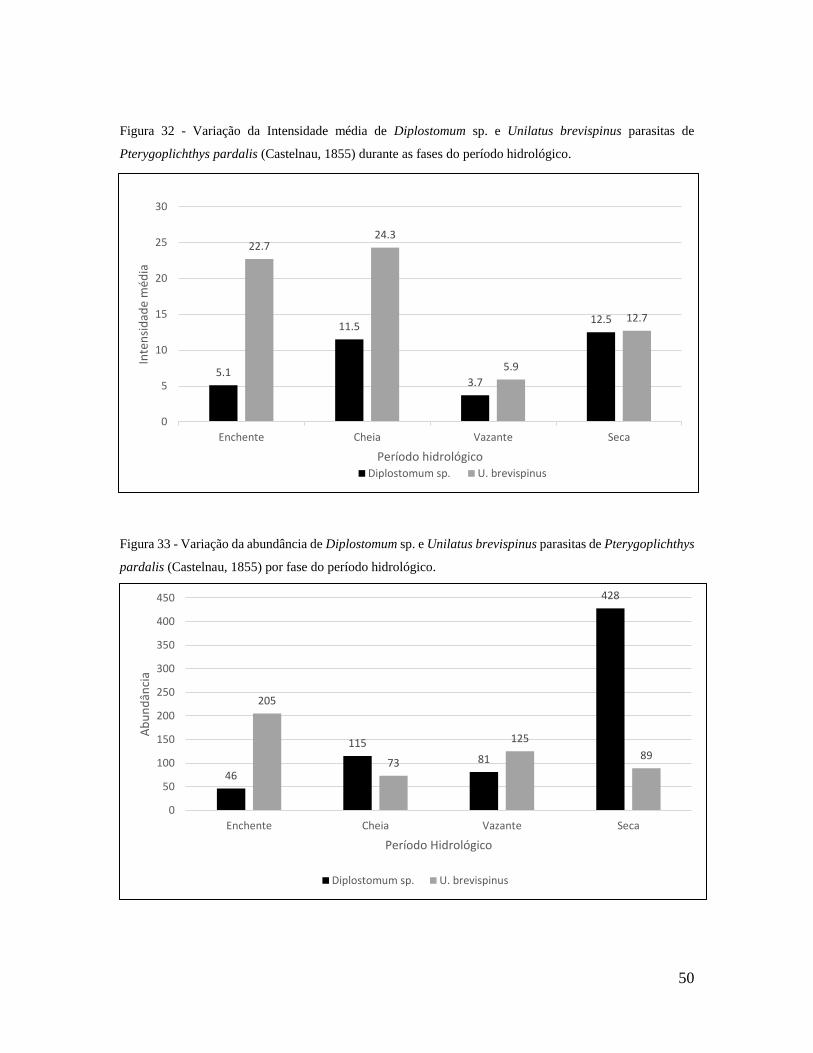
50
Figura 32 - Variação da Intensidade média de Diplostomum sp. e Unilatus brevispinus parasitas de
Pterygoplichthys pardalis (Castelnau, 1855) durante as fases do período hidrológico.
Figura 33 - Variação da abundância de Diplostomum sp. e Unilatus brevispinus parasitas de Pterygoplichthys
pardalis (Castelnau, 1855) por fase do período hidrológico.
5.1
11.5
3.7
12.5
22.724.3
5.9
12.7
0
5
10
15
20
25
30
Enchente Cheia Vazante Seca
Inte
nsi
dad
e m
édia
Período hidrológicoDiplostomum sp. U. brevispinus
46
115
81
428
205
73
125
89
0
50
100
150
200
250
300
350
400
450
Enchente Cheia Vazante Seca
Ab
un
dân
cia
Período Hidrológico
Diplostomum sp. U. brevispinus
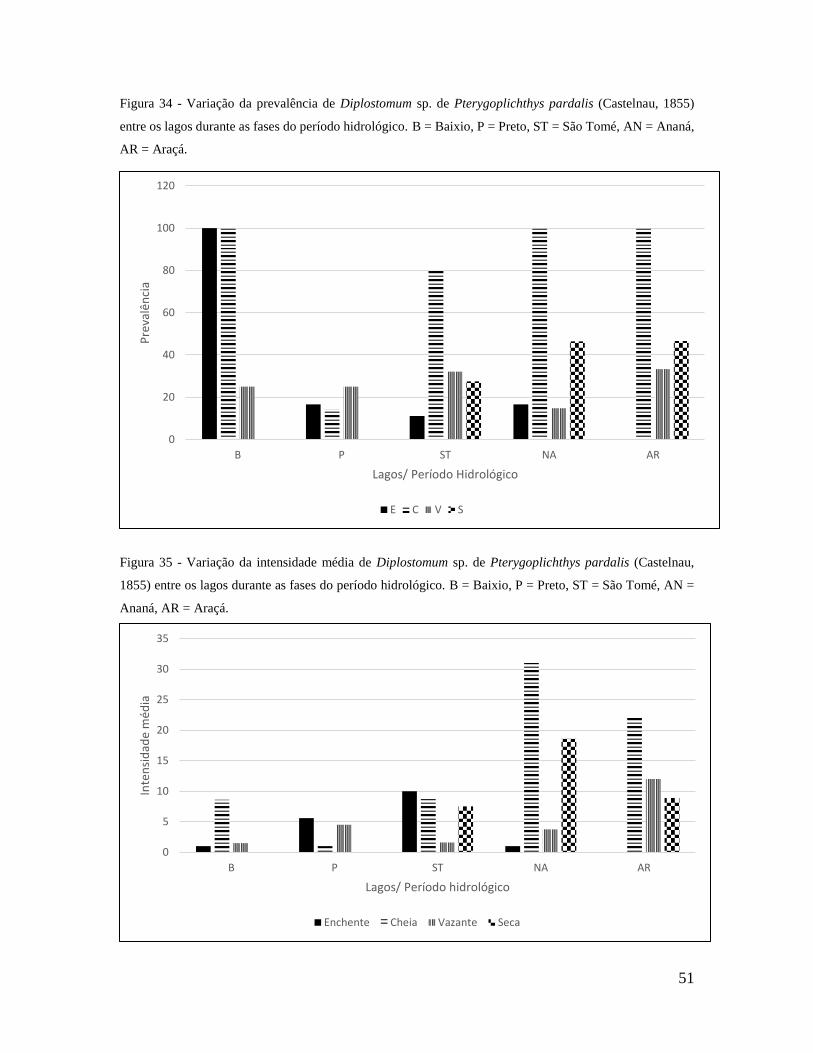
51
Figura 34 - Variação da prevalência de Diplostomum sp. de Pterygoplichthys pardalis (Castelnau, 1855)
entre os lagos durante as fases do período hidrológico. B = Baixio, P = Preto, ST = São Tomé, AN = Ananá,
AR = Araçá.
Figura 35 - Variação da intensidade média de Diplostomum sp. de Pterygoplichthys pardalis (Castelnau,
1855) entre os lagos durante as fases do período hidrológico. B = Baixio, P = Preto, ST = São Tomé, AN =
Ananá, AR = Araçá.
0
5
10
15
20
25
30
35
B P ST NA AR
Inte
nsi
dad
e m
édia
Lagos/ Período hidrológico
Enchente Cheia Vazante Seca
0
20
40
60
80
100
120
B P ST NA AR
Pre
valê
nci
a
Lagos/ Período Hidrológico
E C V S

52
Figura 36 - Variação da abundância de Diplostomum sp. entre os lagos durante as fases do período
hidrológico. B – Baixio, P – Preto, ST – São Tomé, AN – Ananá e AR – Araçá.
Figura 37 - Variação da prevalência de Unilatus brevispinus de Pterygoplichthys pardalis (Castelnau, 1855)
entre os lagos durante as fases do período hidrológico. B = Baixio, P = Preto, ST = São Tomé, AN = Ananá,
AR = Araçá.
0
50
100
150
200
250
300
B P ST NA AR
Ab
un
dân
cia
Lago/ Período Hidrológico
E C V S
0
20
40
60
80
100
120
B P ST NA AR
Pre
valê
nci
a
E C V S
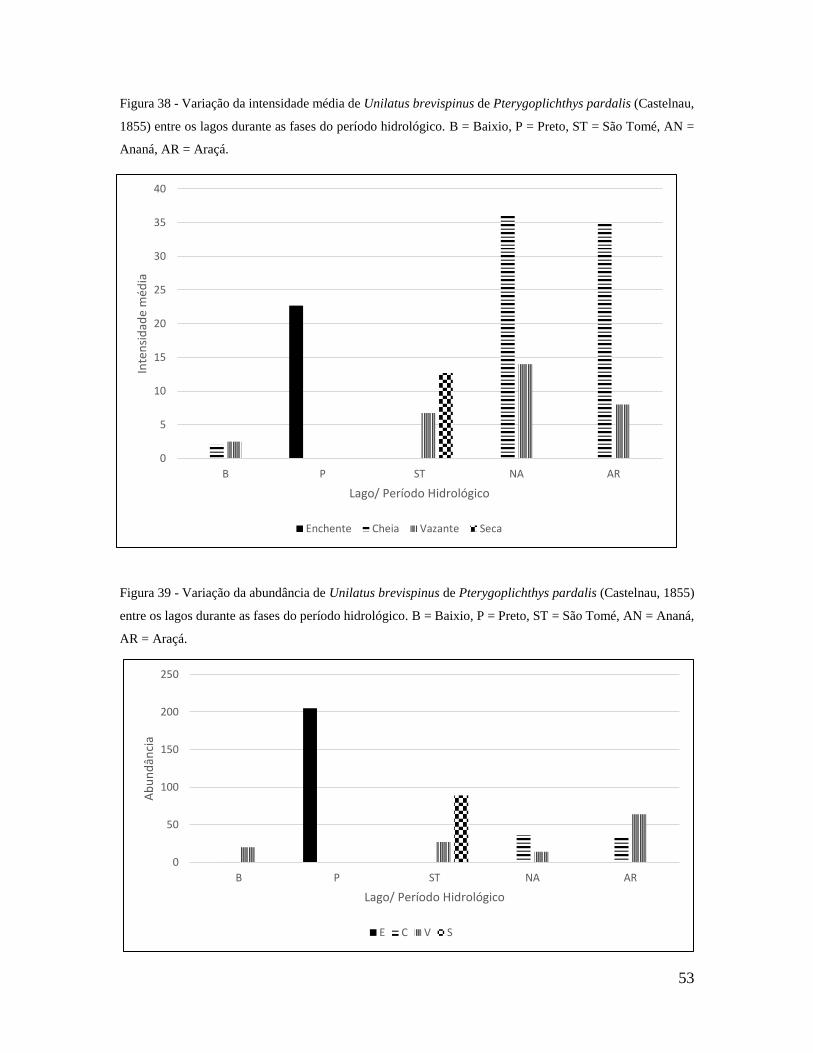
53
Figura 38 - Variação da intensidade média de Unilatus brevispinus de Pterygoplichthys pardalis (Castelnau,
1855) entre os lagos durante as fases do período hidrológico. B = Baixio, P = Preto, ST = São Tomé, AN =
Ananá, AR = Araçá.
Figura 39 - Variação da abundância de Unilatus brevispinus de Pterygoplichthys pardalis (Castelnau, 1855)
entre os lagos durante as fases do período hidrológico. B = Baixio, P = Preto, ST = São Tomé, AN = Ananá,
AR = Araçá.
0
5
10
15
20
25
30
35
40
B P ST NA AR
Inte
nsi
dad
e m
édia
Lago/ Período Hidrológico
Enchente Cheia Vazante Seca
0
50
100
150
200
250
B P ST NA AR
Ab
un
dân
cia
Lago/ Período Hidrológico
E C V S

54
DISCUSSÃO
A composição da comunidade de parasitas é o resultado, entre outros fatores, de
interações entre a história evolucionária e características ecológicas dos hospedeiros
(Poulin 1995). O nível da cadeia trófica em que o hospedeiro se enquadra influencia a
composição da sua fauna parasitária (Takemoto et al. 2004). Para peixes piscívoros é
esperada uma alta riqueza de endohelmintos, porém para peixes que se alimentam de outras
fontes, que não sejam de origem animal (plantas ou detritos), uma menor riqueza é esperada
(Choudhury e Dick 2000).
Nas águas interiores a cadeia alimentar pode começar dos detritos do fundo,
passando por micro-organismos, invertebrados ou peixes detritívoros até os diversos níveis
de piscivoria (Lowe-McConnell 1999). Peixes detritívoros são membros dominantes nas
assembleias de peixes nas várzeas dos rios da América do Sul, onde correspondem
geralmente em mais de 40% da ictiomassa (Araújo-Lima et al. 1995). As espécies de peixes
de Curimatidae, Prochilodontidae e Loricariidae são os maiores consumidores de detritos
das várzeas da bacia Amazônica (Lowe-McConnell 1999; Santos et al. 2006; Soares et al.
2011).
O hábito alimentar do hospedeiro é uma característica a ser considerada, sob o
aspecto epidemiológico, pois tem forte relação com a incidência de endoparasitos (Dogiel
1961; Marcogliese 1995). Pterygoplichthys pardalis por ser detritívoro ocupa uma posição
intermediária na cadeia trófica e é presa para vários vertebrados como outros peixes,
jacarés, lontras, ariranhas e aves. A posição de P. pardalis, na cadeia trófica, faz com que
ele seja hospedeiro intermediário ou definitivo de várias espécies parasitas, viabilizando a
complementação de seus ciclos de vida. Os dados indicam que P. pardalis é parasitado por
espécies parasitas adultas e formas larvais de vários táxons.
Em estudos anteriores Pterygoplichthys pardalis capturados em quatro lagos de
várzea do rio Solimões estavam parasitados com espécies de Monogenoidea da família
Dactylogyridae não identificados. Duas espécies de Digenea, Diplostomum sp. e
Megacelium spinicavum. Uma de Acanthocephala, Gorytocephalus sp. Uma de espécie
Nematoda não identificada e uma de Branchiura, Dolops geayi (Bouvier, 1899) (Porto
2009).

55
A fauna de parasitas de P. pardalis capturados no rio Solimões e de feiras da cidade
de Manaus era composta por cinco espécies parasitas. Duas de Monogenoidea Unilatus sp.
e Heteropriapulus sp. Duas de Digenea, Austrodiplostomum compactum e Megacoelium
spinicavum e uma de Acanthocephala Gorytocephalus sp. (Porto et al. 2012).
A fauna de metazoários parasitas de P. pardalis, do igarapé Fortaleza, no estado do
Amapá, era composta por três espécies parasitas. Uma espécie de Monogenoidea, Unilatus
unilatus, uma espécie de Digenea, Acanthostomum gneri e uma de Acantocephala,
Gorytocephalus elongorchis (Cardoso et al. 2017).
Neste trabalho a fauna de metazoários parasitas de P. pardalis foi composta por
doze espécies. A segunda maior riqueza de espécies foi no trabalho de Porto (2009) seis,
metade do número de espécies encontradas neste trabalho. No terceiro trabalho, o de Porto
et al. (2012), com peixes das feiras de Manaus foi encontrado cinco espécies parasitas. E
com o menor número de espécies, três, um quarto do encontrado neste trabalho, foi o
encontrado em P. pardalis capturados no igarapé Fortaleza, no estado do Amapá (Cardoso
et al. 2017). O maior número de espécies encontrado neste trabalho é devido ao número
maior de coletas, o número maior de peixes examinados e uma extensão maior da área de
coleta.
Para os peixes da família Loricariidae, na bacia Amazônica, existem o registro de
espécies de Monogenoidea de duas famílias Gyrodactylidae (Boeger e Vianna, 2006;
Kritsky et al. 2007) e Dactylogyridae (Suriano, 1967; Boeger e Vianna, 2006). Os
monogenóideos possuem um elevado grau de especificidade parasitária, ocorrendo em um
hospedeiro, ou em hospedeiros filogeneticamente muito próximos (Cone e Kurt 1982). Em
P. pardalis coletados em um igarapé no estado do Amapá foi coletada uma espécie de
Monogenoidea U. unilatus (Cardoso et al. 2017). Não foram encontradas espécies da
família Gyrodactylidae. Pterygoplichthys pardalis coletados em lagos de várzea do rio
Solimões no Amazonas estavam parasitados por duas espécies de Dactylogyridae, Unilatus
sp. e Heteropriapulus sp. (Porto et al. 2012).
Neste trabalho foram encontradas três espécies de Monogenoidea da família
Dactylogyridae: U. brevispinus, T. mourei e H. heterotylus. Todas as três específicas a
peixes da família Loricariidae. Pterygoplichthys pardalis é um novo hospedeiro para estas
espécies. Nenhuma espécie de Gyrodactylidae foi encontrada.

56
As espécies de Digenea são de grande importância tanto por sua diversidade quanto
seu impacto na saúde animal e humana (Thatcher 1993; Bullard e Overstreet 2008;
Fernandes et al. 2015). Elas são heteroxenas, com ciclos de vida que incluem todos os
táxons de vertebrados (Olsen 1978; Núñez e Pertierra 2004).
As espécies de Digenea do gênero Megacoelium Szidat, 1954 são parasitas do
estômago, de peixes da família Loricariidae, na América do Sul. Duas espécies foram
citadas para loricariídeos capturados no lago Janauacá, no rio Solimões, M. sipinicavum
parasita de P. pardalis e M. spinispecum de Pterygoplichthys sp. (Thatcher e Varella 1981)
Pterygoplichthys pardalis capturados em quatro lagos de várzea: Baixio, Preto,
Iauara e Ananá, do rio Solimões estavam parasitados por M. spiniscavum no estômago
(Porto 2009). Neste trabalho as duas espécies de Megacoelium parasitavam
simultaneamente o estômago de P. pardalis.
Austrodiplostomum compactum suas metacercárias parasitam um grande número
de espécies de peixes (Ramos et al. 2013). O primeiro registro desta espécie foi em
Plagioscion squamosissimus (Heckel, 1840) coletados no reservatório de Itaipú (Kohn et
al. 1994). No complexo de lagos Catalão, estado do Amazonas, próximo a Manaus, A.
compactum foi citado parasitando os olhos de peixes das famílias Curimatidae, Cichlidae,
Sciaenidae, Pristigasteridae, Erythrinidae, Characidae, Loricariidae, Pimelodidae e
Serrasalmidae (Souza et al. 2017).
As metacercárias de A. compactum, apesar de não ter especificidade parasitária, têm
uma certa “afinidade” com peixes da família Loricariidae, parasitando várias espécies (Zica
et al. 2010; Ramos et al. 2013). Austrodiplostomum compactum foi citado parasitando P.
pardalis capturados nos lagos de várzea do rio Solimões: Baixio, Preto, Iauara e Ananá,
(prevalência de 22,2 %) (Porto 2009). Pterygoplichthys pardalis adquiridos em feiras de
Manaus, estavam com os olhos parasitados por A. compactum (prevalência de 26, 3 %)
(Porto et al. 2012).
Neste trabalho A. compactum (prevalência de 6,0%) também ocorreu nos P.
pardalis capturados nos cinco lagos de várzea do rio Solimões amostrados (Baixio; Preto;
São Tomé (rio Purus); Ananá; Araçá) parasitando os olhos. Estes dados corroboram com
os de Porto (2009) indicando que A. compactum é uma espécie amplamente distribuída nos
lagos de várzea do rio Solimões.

57
São citadas para o Brasil duas espécies do gênero Diplostomum Nordmann, 1832,
D. alarioides Dubois, 1937 e D. meduse Dubois, 1936 parasitas do intestino de Pteronura
brasiliensis e Caiman crocodilus (Travassos et al. 1969). Posteriormente, em uma listagem
de hospedeiros vertebrados de helmintos do Brasil, apenas D. alarioides foi citado
parasitando P. brasiliensis (Muniz-Pereira et al. 2009). Em um outro trabalho com os
digenéticos da América do Sul, parasitas de aves e mamíferos, somente D. alarioides foi
citado (Fernandes et al. 2015).
Loricariichthys anus (Valenciennes, 1840), um peixe da família Loricariidae,
capturado em lagoas do Rio Grande do Sul estavam parasitados com metacercárias livres
de Diplostomidae nos rins, cavidade do corpo, peritônio ao redor das vísceras e no cérebro.
A espécie não foi identificada por estar em um estágio de desenvolvimento muito inicial,
com apenas rudimentos de órgãos internos (Amato et al. 2001).
Pimelodus maculatus Lacepede, 1803 dos rios São Francisco e Paraná, o
parênquima renal cefálico, a musculatura associada a bexiga gasosa e placas faringeanas
estavam parasitados por metacercárias de Diplostomidae. A espécie foi identificada como
Diplostomum sp. As maiores prevalências foram dos peixes do rio São Francisco (Brasil-
Sato e Pavanelli, 2004).
Neste trabalho foram encontradas metacercárias de Diplostomum sp. não
encistadas, no estômago, gônadas e na superfície dos órgãos internos de P. pardalis. Não
foi possível a identificação da espécie por estarem em um estágio de desenvolvimento
muito inicial, apenas com rudimentos de órgãos internos. Os dados encontrados foram
semelhantes aos de Amato et al. (2001) e Brasil-Sato e Pavanelli (2004). Neste trabalho
Diplostomum sp. foi a espécie mais abundante e o P. pardalis sendo um peixe de fundo e
sedentário, as cercárias de Diplostomum sp. encontraram nele os requisitos ideais para ser
um de seus melhores hospedeiro intermediário.
Entre as espécies de digenéticos registradas para a família Clinostomidae
Odhineriotrema microcephala é única citada para o Brasil (Hughs et al. 1942; Travassos
et al. 1969; Thatcher 1981; Fernandes e Kohn 2014). O. microcephala parasita a cavidade
bucal e o esôfago de jacarés Caiman crocodilos. Ele é o hospedeiros definitivo, e os peixes
são os hospedeiros intermediários (Travassos et al. 1969; Kanev et al. 2002).

58
Cistos com metacercárias grandes, de 6 a 8 mm de comprimento, de O.
microcephala parasitavam as gônadas de Pterygoplichthys sp. na Amazônia Central.
Causando o que é chamado de a “doença das manchas amarelas” (yellow spot diseases).
Estas metacercárias, como parasitam as gônadas, provocam hipoplasia ou reabsorção dos
tecidos reduzindo a capacidade reprodutiva do peixe (Thatcher 1981).
Neste trabalho foram encontrados os mesmos cistos de metacercárias de O.
microcephala parasitando as gônadas de P. pardalis. Os cistos também eram de cor
amarela, uma das características mais evidentes para esta espécie de parasita. Através do
exame visual não foi evidenciado nenhuma anomalia causada por esta espécie de parasita.
As gônadas parasitadas apresentavam aspecto normal em sua morfologia.
Kalitrema é uma espécie que foi descrita parasitando o intestino de Hypostomus
punctatus (Valenciennes, 1840) (Travassos et al. 1969; Thatcher 1993). Esta é a única
espécie descrita para este gênero e é endêmica ao Brasil (Kohn et al. 2007). Neste trabalho
K. kalitrema ocorreu parasitando o intestino de P. pardalis.
Muitos Siluriformes neotropicais de água doce, em particular os da família
Pimelodidae, são parasitados por espécies de Cestoda da família Proteocefalidae
(Chambrier e Rego 1994). Estas espécies de Cestoda têm uma grande afinidade com os
pimelodídeos, porém, muitos peixes amazônicos, de outras famílias, foram citados como
hospedeiros de espécies de Proteocephalidae (Rego 1994; Rego 2003; Thatcher 2006). Para
os peixes da família Loricariidae, da região Amazônica, não há nenhuma citação de
espécies de proteocefalídeos os parasitando (Thatcher 1981; Chambrier et al. 2006;
Chambrier et al. 2015).
Um grande número de pequenos cistos, com larvas encapsuladas de Cestoda foram
encontrados no peritônio, intestino e fígado de Loricariichthys platymetopon Isbrucker &
Nijssen, 1979 coletados no pantanal, no estado do Mato Grosso e no rio Paraná no estado
do Paraná. Porém não foi possível identificar até espécie devido o estágio de
desenvolvimento das larvas ser muito inicial (Schaffer et al. 1992). Estes pequenos cistos,
com larvas encapsuladas eram formas jovens de espécimes da família Proteocephalidae
(Rego 1989).
Algumas espécies de peixes têm pequenos cistos, com larvas encapsuladas de
Cestoda, sobre a parede intestinal, Enquanto em outros peixes são encontradas sobre o

59
mesentério ou superfície das vísceras. Larvas crescidas removidas da cápsula dos tecidos
do hospedeiro, têm escólex de Proteocephalidae (Rego et al. 1999).
Neste trabalho foram encontrados em P. pardalis diversos cistos sobre a superfície
do intestino. Somente um indivíduo estava desenvolvido o suficiente para se observar
algumas das características de Proteocephalidea, Este resultado corrobora com Rego
(1999) que em alguns hospedeiros larvas de proteocefalídeos podem estar fixadas na
superfície das vísceras ou órgãos.
Gorytocephalus elongorchis Thatcher 1979 (Acancthocephala,
Neoechinorhynchidae) foi descrita do intestino de Hypostomus carinatus (Steindachner,
1881) um Loricariidae capturado no lago Janauacá, rio Solimões, no estado do Amazonas
(Thatcher 1979). Gorytocephalus elongorchis aparentemente não apresenta estruturas de
fixação que possam causar grande patogenicidade em seus hospedeiros. Ele tem uma
probóscide muito pequena e baixa intensidade de infestação. No máximo dez indivíduos
foram encontrados no intestino de H. carinatus. Como o hospedeiro é detritívoro, possui
um intestino longo e delicado com uma movimentação lenta, G. elongorchis não precisa
de órgão de fixação forte. Estas características de seu hospedeiro tornaram possível sua
presença no intestino de H. carinatus com o mínimo de danos teciduais e sem sua
eliminação (Thatcher 1981).
Gorytocephalus elongorchis foi a única espécie de Acanthocephala encontrada
parasitando P. pardalis nos lagos de várzea do rio Solimões no estado do Amazonas,
parasitavam o intestino (Porto 2009; Porto et al. 2012). Pterygoplichthys pardalis
capturados no igarapé Fortaleza, no estado do Amapá, também estavam parasitados por G.
elongorchis e parasitavam a cavidade abdominal (Cardoso et al. 2015).
Neste trabalho G. elongorchis foi encontrado parasitando o intestino de P.
pardalis. Poucos indivíduos os parasitavam, a maior intensidade foram quatro indivíduos
no intestino. Nenhum dos G. elongorchis coletados estavam fixados na parede do intestino;
corroborando com os dados de Thatcher (1981) que citou que esta espécie apresenta poucas
indicações de patogenicidade por possuir probóscide muito pequena, baixa intensidade de
infestação e não se fixar nas finas paredes do intestino de P. pardalis. Gorytocephalus
elongorchis é específico de peixes da família Loricariidae que são seus hospedeiros
definitivos.

60
Para a Amazônia são citadas 47 espécies de Copepoda parasitas de peixes, 43 são
da família Ergasilidae e estão incluídas em 16 gêneros (Varella e Malta
2009). Therodamas Krøyer, 1863 é um destes gêneros com seis espécies: T.
serrani Krøyer, 1863, T. sphyricephalus Thomsen, 1949, T. dawsoni Cressey, 1972, T.
fluviatilis, T. elongatus (Thatcher, 1986) e T. frontalis El-Rashidy e Boxshall 2001 (El-
Rashidy e Boxshall 2001).
As espécies de Therodamas ocorrem somente na região Neotropical e foram citadas
para peixes de seis famílias: T. fluviatilis para uma espécie de Characidae da Argentina
(Paggi 1976); T. dawsoni de um Dactyloscopidae do Panamá (Cressey 1972); T.
elongatus de um Sciaenidae do Brasil (Thatcher 1986; Amado e Rocha 1996); T.
sphyricephalus de um Carangidae do Uruguai (Thomsen 1949); T. serrani de um
Serranidae do Caribe (Krøyer, 1863); T. frontalis de um Carangidae do Uruguai (Thomsen,
1949 como T. serrani) e um Mugilidae do Brasil (Carvalho 1955, como T. serrani). A
utilização dos hospedeiros representa seis diferentes famílias indicando o baixo nível de
especificidade parasitária ao nível genérico (El-Rashidy e Boxshall 2001).
Therodamas elongatus é a única espécie do gênero citada para a Amazônia e foi
encontrada parasitando P. squamosissimus da família Scianidae (Thatcher 2006) e A.
crassipinis (Heckel, 1840) da Cichlidae (Atroch 2016). Neste trabalho T. elongatus foi
encontrada parasitando as brânquias e perfurando o raio do arco branquial de P. pardalis,
e quatro espécimes foram coletados.
Diferenças podem ser encontradas nas comunidades componentes de uma mesma
espécie de hospedeiro em diferentes locais onde possa ser coletada, por estar sendo
influenciada por fatores locais (Luque et al. 2004). Neste trabalho os descritores da
comunidade componente: riqueza, diversidade, dominância e equitatividade das espécies
parasitas de P. pardalis capturadas nos cinco lagos apresentaram variações. Os índices
parasitários variaram ao longo do período hidrológico, mas as diferenças foram
influenciadas pela abundancia das espécies de Diplostomum sp. e U. brevispinus.
As espécies parasitas de P. pardalis apresentaram algumas diferenças em seus
descritores e índices epidemiológicos entre os lagos. Mas houve um alto grau de
semelhança entre as infrapopulações de parasitas indicando que, apesar das distâncias entre
os lagos, as espécies co-ocorreram em diferentes populações de P. pardalis.

61
O caráter agregado de distribuição é considerado comum em parasitas de peixes
marinhos e de água doce, a causa primária deste tipo de distribuição, dentro de uma
população de hospedeiros, está associada principalmente a fatores estocásticos (Zuben
1997). O tipo de dispersão e agregação encontrado para a comunidade componente de P.
pardalis foi agregado que é o padrão para as espécies parasitas de peixes, corroborando
com Zuben (1997). Este tipo de distribuição aumenta a regulação dependente da densidade
e abundância, tanto nos hospedeiros como nas espécies parasitas, além de reduzir a
competição interespecífica entre os parasitas.
Os peixes desempenham papel primordial nos ecossistemas amazônicos,
principalmente em razão da excepcional e extensa rede hidrográfica da bacia. Ela permite
interagir em um vasto espaço geográfico e em uma grande variedade de ambientes e nichos
ecológicos. Essas características induzem a coexistência de complexas cadeias tróficas,
com vários níveis e grande diversidade (Isaac et al. 2012).
Qual a função dos parasitas? Os humanos responderão que eles não têm função
nenhuma para a humanidade. Que eles são apenas responsáveis por doenças e/ou qualquer
coisa ruim que se possa imaginar com relação a esta palavra que designa estes animais
(Overstreet 1997; Lafferty 1997). As espécies parasitas são um importante, mais
geralmente negligenciado, componente de qualquer ecossistema (Marcogliese 1995). Em
muitos estudos ecológicos eles são deixados de lado por serem considerados irrelevantes
(Loreau et al. 2005).
Neste trabalho foi avaliado, entre as espécies parasitas de P. pardalis, qual delas
poderia ser utilizada como bioindicadora ambiental, segundo os requisitos propostos por
Overstreet (1997). E para mostrar a importância dos parasitas de peixes como uma
ferramenta útil, com uma importante função no ecossistema, para monitorar as mudanças
no meio aquático. Esta avaliação foi realizada analisando-se qualitativa e quantitativamente
as espécies de metazoários.
Muitos trabalhos, utilizando espécies parasitas de peixes como bioindicadoras,
foram realizados para verificar qual a melhor espécie para indicar determinado efeito no
ambiente aquático (Mackenzie et al. 1995; Marcogliese e Cone, 1997; Sures 2004; Madi
2005; Vidal-Martinez et al. 2009). As alterações ambientais causam efeitos diferenciados,
ema cada grupo de parasitas, dentro da comunidade componente. Avaliando a comunidade

62
componente de P. pardalis foi possível verificar que entre as espécies que fazem parte da
sua diversidade algumas duas podem ser bem promissoras como bioindicadoras:
Diplostomum sp e Unilatus brevispinus, porém, mais estudos precisam ser realizados para
efetivar este processo.
Neste trabalho duas espécies parasitas foram predominantes em P. pardalis. O
Monogenoidea ectoparasita U. brevispinus e o Digenea endoparasita Diplostomum sp.
Estas espécies também foram encontradas por Porto (2009) como as mais representativas
para P. pardalis de lagos de várzea da Amazônia.
A grande maioria dos trabalhos estudando a estrutura da comunidade componente
de metazoários parasitas apresentaram pelo menos uma espécie de parasita com status
central (Alves et al. 2003; Guidelli et al. 2003; Lopes 2009, Morais 2011). Em um estudo
sobre a fauna de metazoários de Hoplosternum litoralle (Hancock, 1828) (Callichthyidae)
foi encontrado um baixa prevalência e diversidade, provavelmente pelas condições do rio
Guandu que recebe efluentes domésticos (Abdallah et al. 2006). Neste trabalho as
prevalências foram baixas para as espécies de metazoários parasitas de P. pardalis
coletados em lagos de várzea, porém, este deve ser o padrão deste grupo pois, os lagos
onde foram coletados não possuíam indícios de alterações antrópicas.
Para uma comunidade de espécies parasitas se encontrar em equilíbrio a mesma
deve ser formada por um núcleo de espécies dominantes (Bush e Holmes 1986). A
comunidade componente de P. nattereri, de lagos de várzea da Amazônia e nos diferentes
períodos hidrológicos, apresentaram um núcleo de espécies dominantes e constantes,
compostas por espécies de Monogenoidea e Nematoda como espécies centrais. As demais
espécies parasitas foram consideradas comuns ou raras. Por esta razão a comunidade
componente de P. nattereri foi considerada em equilíbrio (Morais 2011).
Da fauna de A. microlepis, de lagos de várzea do rio Solimões, as espécies de
Digenea foram as mais importantes. Porém não foi encontrada nenhuma espécie central
(Dumbo 2014). Neste trabalho com as infracomunidades de P. pardalis não foi encontrada
nenhuma espécie central. Nenhuma das espécies parasitas apresentou uma prevalência que
indicasse este status comunitário. Isto é um indicativo de que as infracomunidades das
espécies parasitas de P. pardalis, dos lagos de várzea estudados, ainda não atingiram o

63
equilíbrio, devido à ausência de um táxon dominantes. Mesmo com a grande abundância
de uma espécie de Monogenoidea e uma Digenea.
As espécies de Monogenoidea possuem alta especificidade parasitária, por
parasitarem hospedeiros de uma família ou espécie de peixe (Boeger e Viana 2006; Viana
2011; Kritsky 2000). Devido a este alto grau de especificidade, as espécies de
Monogenoidea são limitadas ao seu hospedeiro definitivo. Como não têm hospedeiros
intermediários elas serão diretamente afetadas, caso o hospedeiro entrar em contato com
algum poluente ou o ambiente sofrer alguma mudança (Mackenzi et al. 1995; Khan e
Kiceniuk 1983).
Em um estudo da fauna de espécies parasitas de P. nattereri, como bioindicadora
ambiental, as espécies mais abundantes foram as de Monogenoidea. Foram encontradas 16
espécies de um total de 2.271 espécimes coletados. Destas espécies três, Amphitecium junki
Boeger e Kritsky, 1988, A. brachycirrum Boeger e Kritsky, 1988 e A. falcatum Boeger e
Kritsky, 1988 apresentaram correlação entre a abundância com as variáveis limnológicas
(Morais 2011).
Avaliando as espécies parasitas de peixes como bioindicadoras, as espécies de
Ancyrocephalinae (não identificadas) parasitas das brânquias de G. brasiliensis mostraram
uma resposta as alterações de turbidez nos reservatórios Juqueri e Jaguari (Madi 2009).
Neste trabalho três espécies de Monogenoidea Heteropriapulus heterotylus, Unilatus
brevispinus e Trinigyrus mourei foram encontradas parasitando P. pardalis, mas somente
U. brevispinus apresentou os maiores valores de abundância. Monogenoidea foi o segundo
táxon que apresentou a maior abundância e nenhuma espécie apresentou alguma correlação
com as variáveis limnológicas.
Espécies de Monogenoidea em contato com hidrocarbonetos ou derivados do
petróleo são estimuladas a reproduzir. Os locais com altos níveis destes contaminantes
espera-se maior abundância de Monogenoidea em seus hospedeiros (Galli et al. 2001). Em
locais onde o ambiente aquático possuía altos níveis de contaminação (coliformes fecais,
nitrito, nitrato e chumbo) as espécies de Monogenoidea apresentaram altas taxas de
infestação nos peixes (Galli et al. 2001).
Neste trabalho U. brevispinus apresentou os maiores índices parasitológicos entre
as espécies ectoparasitas. Porém os lagos de várzea estudados não são antropizados e

64
também não apresentaram qualquer alteração na qualidade da água. Os maiores índices
parasitológicos de U. brevispinus em P. pardalis deve ser o resultado da especificidade
parasitária ou outros fatores que esta relação hospedeiro-parasita.
As espécies de metazoários de parasitas de Hoplias malabaricus (Bloch, 1794)
foram avaliadas para o seu uso como bioindicadoras ambientais. O Nematoda
Contracaecum sp. e espécies de Monogenoidea, não identificadas, foram as que
apresentaram as maiores prevalências e intensidade. Porém elas não apresentaram nenhum
resultado positivo para poderem serem usadas como bioindicadoras ambientais (Corrêa et
al. 2009).
Em um trabalho avaliando a viabilidade do uso de espécies de Ancyrocephalinae
parasitas de G. proximus como indicadoras de mudanças no ambiente aquático, também
não foi encontrado um resultado positivo na correlação entre prevalência e intensidade
media e parâmetros limnológicos. Porém, apesar deste resultado, este grupo de parasitos
foi recomendado como bioindicador ambiental (Madi 2009)
Neste trabalho as espécies de Monogenoidea foram o segundo táxon que apresentou
a maior abundância e riqueza. Entre as espécies de Monogenoidea, U. brevispinus foi a
espécie de maior prevalência e intensidade média. Ela mostrou pequenas diferenças entre
a prevalência, intensidade média e abundância entre os períodos hidrológicos, mas
nenhuma correlação com os parâmetros limnológicos.
As espécies de Digenea são as mais conhecidas entre os grupos parasitas, sua
principal característica é o seu ciclo de vida que exige mais de um hospedeiro para chegar
à fase adulta e o primeiro hospedeiro é uma espécie de Mollusca (Thatcher 1993; Morley
e Lewis 2001). Estas características são fundamentais para o estudo das espécies de
Digenea, que podem ser utilizados como indicadores de mudanças aquáticas, além de
serem utilizadas em testes de acúmulo de metais pesados (Evans et al. 2001).
O potencial do uso de espécies parasitas de peixes marinhos, como indicadoras de
mudanças ambientais, foi avaliado em vários tipos de ciclo de vida de espécies de Digenea,
para elucidar suas rotas de transmissão entre seus hospedeiros. Entre os grupos
encontrados, os digenéticos estavam entre as melhores espécies para indicar as alterações
ambientais. Como precisam de mais de um hospedeiro para completarem seu ciclo de vida,

65
as alterações no ambiente aquático podem desequilibrar as populações de seus hospedeiros
intermediários, levando a diminuição de sua população (Makcenzie et al. 1995).
As espécies parasitas avaliadas como indicadoras da saúde do ecossistema,
mostraram que as perturbações na estrutura e funcionamento do ecossistema afetaram a
topologia da teia trófica e também a transmissão de espécies parasitas. Estas perturbações
afetaram a abundância e composição destas espécies. Comunidades e populações de
espécies parasitas são úteis como indicadoras de estresse ambiental, estrutura da teia trófica
e biodiversidade. Ficou evidente que espécies parasitas com ciclo de vida heteroxeno são
muito úteis em estudos de bioindicadoras ambientais (Marcogliese 2005).
Foi avaliada a qualidade da água utilizado quatro espécies de metacercárias,
Bucephalus polymorphus Baer, 1827, Asymphylodora tincae (Modeer, 1790),
Diplostomum spathaceum (Rudolphi, 1819) e Tylodelphys clavata (Von Nordmann, 1832)
em quatro pontos de coleta. Local não poluído, moderadamente poluído, poluído e
extremamente poluído. No local não poluído e moderadamente poluído as espécies de D.
spathacecum eram abundantes. Nos locais poluídos e extremamente poluídos eram
ausentes (Galli et al. 2001). Neste trabalho as metacercárias de Diplostomum sp. foram a
espécie mais abundante. Ela ocorreu parasitando P. pardalis em todos os lagos estudados
e nas quatro fases do período hidrológico. Os dados indicaram que os lagos de várzea
estudados não apresentaram alterações ambientais.
O efeito antropogênico nas comunidades de Mollusca e Digenea, no canal
Basingstoke no Reino Unido foi avaliado. A pressão antropogênica causou a diminuição
das espécies de moluscos hospedeiras intermediárias de digenéticos. Consequentemente as
populações de espécies parasitas diminuíram. Mesmo alguns anos depois quando um
programa de recuperação, para esta área foi colocado em prática, as populações de espécies
parasitas não conseguiram se restabelecer (Morley e Lewis 2001).
Neste trabalho as infrapopulações de metacercárias de Diplostomum sp. parasitas
de P. pardalis foram as que apresentaram a maior abundância. Os dados indicaram que os
lagos de várzea onde os P. pardalis foram capturados, não apresentam modificações
antropogênicas, apesar de estarem próximos ao canal principal do rio Solimões, que é rota
de navegação de muitos barcos.

66
Para avaliar o efeito do petróleo bruto em espécies de parasitas gastrointestinais do
Pseudopleuronectes americanos (Walbaum, 1972), naturalmente infectado com o Digenea
Steringophorus furciger (Olsson, 1867). Os peixes foram expostos a sedimentos
contaminados com petróleo, ou a frações de petróleo solúveis em água do petróleo
venezuelano por 34 e 160 dias, respectivamente. Em todos os casos, a prevalência e a
intensidade das infecções parasitárias foram menores nos peixes submetidos ao petróleo
do sedimento. O efeito pareceu ser mais pronunciado nos peixes expostos a extratos
solúveis em água do que ao sedimento contaminado com petróleo. Menos parasitos
presentes nos peixes expostos ao petróleo podem ser atribuídos à toxicidade direta,
induzida por frações solúveis em água potável e/ou modificação do ambiente intestinal,
provocada por alterações na fisiologia do hospedeiro (Khan e Kiceniuk 1983).
Na Finlândia os efeitos da poluição e eutrofização em lagos foi estudado. Os dados
indicaram que a poluição afetava a abundância das espécies de Digenea, isto devido à
escassez das espécies de Mollusca, os hospedeiros intermediários, eliminados pela
poluição (Valtonen et al. 1997). As variações da prevalência e da intensidade média de
metacercárias de Clinostomum sp., parasitas de G. brasiliensis, foram avaliadas, como
indicadoras ambientais, em dois reservatórios no estado de São Paulo. Nos reservatórios
do Juqueri (eutrófico) as metacercárias de Clinostomum sp. apresentaram prevalência
maior indicando ser um ambiente eutrofizado e com elevada produção primária líquida. As
altas intensidades médias ocorreram em áreas de remanso e desague de tributários com alta
quantidade de dejetos orgânicos. A elevada quantidade de nutrientes, nestas áreas,
promoveu o desenvolvimento de macrófitas. Locais ideais para o desenvolvimento de
invertebrados e moluscos, estes o primeiro hospedeiro intermediário de espécies de
Digenea (Madi 2005).
No reservatório do Jaguari (Oligomesotrófico) que não era poluído, não tinha áreas
de remanso ou desague de tributários, semelhantes ao Juqueri. Possivelmente por isso a
prevalência e a intensidade média não apresentaram diferenças significativas entre os
pontos de coleta e sempre seus valores foram mais baixos que no reservatório do Juqueri
(Madi 2005).
Neste trabalho as metacercárias de Diplostomum sp. que parasitavam os órgãos
internos de P. pardalis ocorreram em todos os lagos de várzea, sua prevalência e a

67
intensidade média não apresentaram diferenças durante as fases do período hidrológico,
indicando não existir fatores antrópicos ou indicativo de alguma alteração no ambiente. Ao
contrário do ambiente artificial dos reservatórios de Juqueri (Madi 2005). Os lagos
amazônicos não apresentam nenhuma ou muito pouca ação antrópica ao longo do ano
(Petesse et al. 2016). E a grande abundância de Diplostomum sp. indicou que esta espécie
parasita está completando o seu ciclo de vida plenamente nestes lagos amazônicos. Pois
todos os seus hospedeiros estão presentes no meio ambiente.
A grande abundância de Diplostomum sp. parasitando P. pardalis, durante a estação
de cheia, deve ser devido à grande inundação das terras da várzea, dos furos, dos canais e
dos pequenos lagos formando imensas áreas alagadas. Neste grande ambiente aquático é
viabilizado o encontro dos miracídios eclodidos dos ovos, com o seu primeiro hospedeiro
intermediário, as espécies de Mollusca. Depois da reprodução assexuada dos miracídeos
dentro dos moluscos, as cercárias são liberadas e nadam ativamente a procura de seu
segundo hospedeiro intermediário e penetram ativamente através da pele. No caso as
cercárias de Diplostomum sp. penetraram nos corpos de P. pardalis, o seu segundo
hospedeiro intermediário.
Os indicadores ecotoxicológicos foram avaliados, pelo método de alterações na
enzima GST (glutationa s-transferase) e metais pesados em P. pardalis de lagos de várzea
do rio Solimões. Não foram encontradas diferenças significativas, para a enzima GST entre
os P. pardalis dos lagos amostrados. Esta enzima atua no metabolismo antioxidante e no
metabolismo de xenobióticos (substâncias tóxicas). Para metais pesados (Cádmio [Cd] e
Cobre [Cu]), em P. pardalis, as concentrações estavam abaixo dos limites recomendados
pela Anvisa, durante todo o período hidrológico (Val et al. 2011). Metais pesados podem
influenciar no tempo de vida de cercárias (Pietrock et al. 2002a, 2002b).
Neste trabalho não foram feitos testes de metais pesados e ecotoxicológicos. Mas
os dados corroboram com os de Val et al. (2011). Pterygoplichthys pardalis não esteve em
contato nem com a enzima GST (glutationa s-transferase metais pesados (Cádmio [Cd] e
Cobre [Cu]). Como as infracomunidades de espécies parasitas de P. pardalis não tiveram
contato com estes contaminantes, estas substâncias tóxicas não interferiram nos seus ciclos
de vida, não alterando as suas taxas populacionais nos lagos estudados.

68
Foram avaliados os lagos: Baixio, Preto, Araçá, Ananá, Maracá e Poraquê
utilizando o “Índice de Gradiente Ambiental” (EGI), para saber se os lagos sofreram ação
antrópica. O EGI do lago Baixio foi classificado na categoria “média”, ele apresentou sinais
da pressão antrópica e os efeitos negativos já começam a aparecer. Os lagos Araçá e Ananá
obtiveram as mais altas classificações, eles foram colocados na categoria de “excelentes”.
Que corresponde a uma muito boa condição natural com nenhum ou o mínimo de sinais de
efeitos antrópicos, permitindo estes lagos serem chamados de inalterados. Os lagos
Poraquê, Maracá e Preto foram classificados na categoria de “bom”. Que corresponde a
situação em que os sinais das alterações aparecem, mas onde o uso antrópico é sustentável
e os impactos humanos não são fortes para danificar o meio ambiente (Petesse et al. 2016).
Neste trabalho, quatro dos cinco lagos estudados por Petesse et al. (2016): Baixio,
Preto, Araçá e Ananá, foram estudadas as comunidades de espécies parasitas de P.
pardalis. Dez espécies parasitas de P. pardalis ocorreram em peixes capturados em todos
os lagos. Exceto duas espécies: M. spinispecum encontrado em dois lagos São Tomé e
Araçá e H. heteropriapulus que só não ocorreu no lago Ananá. Todas as espécies parasitas
heteroxenas e monoxenas ocorreram nos lagos, indicando que os seus ciclos de vida
estavam completando-se. E, que as ações antrópicas nos lagos eram mínimas e que os sinais
das alterações não apareciam. Mas quando apareciam, o uso antrópico foi sustentável e os
impactos humanos não foram fortes para danificar o meio ambiente.
A diversidade parasitária constituída por 12 espécies parasitas, em diferentes fases
do ciclo de vida, indicara que P. pardalis é um elo importante na teia de transmissão destes
parasitas, nos lagos amostrados e em todos os ambientes em que esta espécie está
distribuída. Com este trabalho aumentou o conhecimento sobre a diversidade das espécies
parasitas de P. pardalis. Aumentaram as informações sobre as espécies parasitas que
poderão ser usadas em programas de monitoramento, como uma ferramenta a mais para o
estudo de modificações, que eventualmente possam vir a acontecer no ambiente aquático
da Amazônia.
Duas espécies parasitas destacaram-se na comunidade componente de P. pardalis:
Diplostomum sp. e U. brevispinus. Elas são indicadas para um estudo mais detalhado para
serem usadas como bioindicadoras ambientais. A comunidade componente de metazoários
parasitas de P. pardalis indicou estar desempenhando suas funções dentro dos lagos

69
estudados. O ciclo biológico está se completando com todos os hospedeiros exigidos
presentes no ecossistema. São necessários mais estudos, para uma melhor compreensão de
como as espécies parasitas de peixes, da região Amazônica, podem ser usados como
ferramentas para monitorar os ecossistemas aquáticos amazônicos.
BIBLIOGRAFIA CITADA
Abdallah, V.D.; Azevedo, R.K.; Luque, J.L. 2006. Ecologia da comunidade de metazoários
parasitos do Tamboatá Hoplosternum littorale (Hancock, 1828) (Siluriformes:
Callichthyidae) do rio Guandu, estado do Rio de Janeiro, Brasil. Acta Scientiarum
Biological Science, 28: 413-419.
Amato, J.F.R.; Boeger, W.A.; Amato, S. B. 1991. Protocolos para laboratório-coleta e
processamento de parasitas do pescado. Imprensa Universitária, Universidade Federal
do Rio de Janeiro Rio de Janeiro, Brasil. 81p.
Amato, S.B; Amato, J.F.R.; e Albrecht, M. 2001. Metacercárias livres de diplostomídeos
(Digenea, Diplostomidae) em Loricariichthys anus (Val, 1840) (Siluriformes,
Loricariidae) do estado do Rio Grande do Sul, Brasil. Parasitologia al dia, 25(1): 1-7.
Almeida-Val, V.M.F; Val, A.L.; Walker, I. 1999. Long and short-term adaptation of
Amazon fishes to varying oxygen levels: intra-specific phenotypic plasticity and
interspecific variation. In: Val, A.L.; Almeida-Val, V.M.F (Eds.). Biology of Tropical
Fishes. INPA, Manaus, Brasil. p.185-206.
Alves, D.R.; Luque. J.L. 2003. Ecologia das comunidades de metazoários parasitos de
cinco espécies de escombrídeos (Perciformes: Scombridae) do litoral do estado do Rio
de Janeiro, Brasil. Revista Brasileira de Parasitologia Veterinária, 15: 167-181.
Amado, M.A.P.M; Rocha, C.E.F. 1996. Therodamas tamarae, a new species of copepod
(Poecilostomatoidea: Ergasilidae) parasitic on Plagioscion squamosissimus (Heckel)
from the Araguai river, Brazil; with a key to the species of the genus. Hydrobiologia,
325: 77-82.
Amin, O. 1969. Helminthes fauna of suckers (Catostomidae) of the Gila river system,
Arizona. II. Five parasites from Catostomus spp. American Midland Naturalist, 82:
429-443.
Anderson, T.; Forlin, L.; Hardig, J.; Larsson, A. 1988. Physiological disturbances in fish

70
living in coastal coastal polluted with blanched kraft pulp mill effluents. Canadian.
Journal Fish Aquatic. Science, 45: 1525-1536.
Araújo-Lima, C.A.R.M.; Forsberg, B.R.; Victoria, R.; Martinelli, L. 1986. Energy source
for detritivorous fishes in the Amazon. Science, 234: 1256-1258.
Araújo-Lima, C.A.R.M.; Agostinho, A.A.; Fabré, N.N. 1995. Trophic aspects of fish
communities in Brazilian rivers and reservoirs. In: Tundisi, J.G.; Bicudo, C.E.; Tundisi,
T.M. (Eds.) Limnology in Brazil. Academia Brasileira de Ciências, Sociedade
Brasileira de Limnologia, p.1-32.
Atroch, F.M.P.B. 2016. Metazoários parasitas de Astronotus ocellatus (Agassiz, 1831) e
Astronotus crassipinis (Heckel, 1840) em lagos de várzea na Amazônia, Amazonas,
Brasil. Dissertação de Mestrado, Instituto Nacional de Pesquisas da Amazônia – INPA.
80p.
Barthem, R.B.; Fabré. N.N. 2003. Biologia e diversidade dos recursos pesqueiro da
Amazônia. In: Rufino, M.L. A pesca e os recursos pesqueiros da Amazônia. Provárzea,
Manaus, Brasil, p.11-55.
Batista, V.S.; Petrere Jr. M. 2003. Characterization of the commercial fish production
landed at Manaus, Amazonas State, Brazil. Acta Amazonica, 33: 53-66.
Bell, G.; Burt, A.1991. The comparative biology of parasite species diversity: internal
helmints of freshwater fish. Journal Animal Ecology, 60: 1047-1063.
Belmejo, L.; Martos, H.L. 2008. Utilização de Xiphophorus hellericomo como
bioindicador de poluição hídrica de derivados de petróleo em condições tropicais.
Revísta Eletrônica de Biologia, 1: 1-17.
Boikova, E. 1990. Protozoans as a component of biological monitoring of the Baltic Sea.
Limnologica, 20: 119-125.
Boeger, W.A.; Kritsky, D.C. 1988. Neotropical Monogenea 12. Dactylogyridae from
Serrasalmus nattereri (Kner) (Cypriniformes, Serrasalmidae) and aspects of their
morphologic and distribution variation in the Brazilian amazon. Proceedings of the
Helminthological Society of Washington, 55: 188-213.
Boeger, W.A.; Viana, R.T. 2006. Monogenoidea. In: Thatcher, V.E. Amazon fish parasites.
2a Ed., Editora Aquatic Biodiversity in Latin America, Sofia, Moscow, p.42-116.

71
Brito, A.L. 1981. Aspectos anatômicos e considerações sobre os hábitos de
Pterygoplichtys multiradiatus Hancock, 1828 do bolsão do Janauacá – Amazonas,
Brasil (Osteichthyes, Siluriformes, Loricariidae). Dissertação de Mestrado, Instituto
Nacional de Pesquisas da Amazônia/Fundação Universidade do Amazonas, Manaus,
Amazonas, Brasil. 102p.
Bullard, S.A.; Overstreet, R.M. 2008. Chapter 14 Digeneans as enemies of fishes. In: Eiras
et al (Edt). Fish Disease Volume 2. Science publishers. p.817-976.
Bush, A.O. Holmes, J.C. 1986. Intestinal helminthes of lesser scaup ducks: an interactive
community. Canadian Journal of Zoology, 64: 142-154.
Bush, A.O.; Lafferty, K.D.; Lotz, M.J.; Shotask, A.W. 1997. Parasitalogy meets ecology
on its own terms: Margolis et al. Revised. Journal Parasitalogy, 83: 575-583.
Buss, D.F.; Baptista, D.F.; Nessimian, J.L. 2003. Bases conceituais para a aplicação de
biomonitoramento em programas de avaliação da qualidade da água de rios. Cadernos
de Saúde Pública, 19: 465-473
Cairns, Jr.J.; Mccormick, P.V.; Niederlehner, B.R. 1993. A proposal framework for
developing indicators of ecosystem health. Hydrobiologia, 263: 1-44.
Callisto, M.; Moretti, M.; Goulart, M.D.C. 2001. Macroinvertebrados bentônicos como
ferramenta para avaliar a saúde de riachos. Revista Brasileira de Recursos Hídricos,
6: 71-82.
Cardoso, A.C.F; Oliveira, M.S.B.; Neves, L.R.; Tavares-Dias, M. 2017. Metazoan fauna
parasitizing Peckoltia braueri and Pterygoplichthys pardalis (Loricariidae) catfishes
from the northeastern Brazilian Amazon. Acta Amazonica, 47: 147-154.
Chambrier, A.; Rego, A.A. 1994. Proteocephalus sophiae n. sp. (Cestoda:
Proteocephalidea) a parasite of the siluroid fish Paulicea lutkeni (Pisces: Pimelodidae)
from the brazilian Amazon. Revue Suisse de Zoologie, 101: 361-368.
Chambrier, A.; Scholz, T. Kuchta, R.; Posel, P.; Mortenthaler, M.; Guardia, C.C. 2006.
Tapeworms (Cestoda: Proteocephalidea) of fishes from the Amazon river in Peru.
Comparative parasitology, 73: 111-120.
Chambrier, A.; Kuchta, R.; Scholz, T. 2015. Tapeworms (Cestoda: Proteocephalidea) of
teleost fishes from the Amazon river in Peru: additional records as an evidence of
unexplored species diversity. Revue Suisse de Zoologie, 122: 149-163.

72
Chervy, L. 2002. The terminology of larval cestodes or metacestodes. Systematic
parasitology, 52: 1-33.
Chubb, J.C. 1979. Seasonal occurrence of helminthes in freshwater fishes. Part II.
Trematoda. Advances in Parasitology, 17: 141-313.
Chubb, J.C. 1980. Seasonal occurrence of helminthes in freshwater fishes. Part III. Larval
Cestoda and Nematoda. Advances in Parasitology, 18: 1-120.
Cone, D.K.; Kurt, M.D.B. 1982. The host especifity of Urocleidus adspectus (Muller,
1938) (Monogenea: Ancyrocephalinae). Journal of Parasitology, 75: 702-706.
Corrêa, L.L. 2009. Avaliação de metazoários parasitos de Hoplias malabaricus (Bloch,
1794) (Pisces: Erythrynidae) como potenciais indicadores de qualidade ambiental.
Dissertação de Mestrado Universidade Estadual de Campinas, São Paulo. 87p.
D’Amelio, S.; Gerasi, L. 1997. Evaluation of environmental deterioration by analyzing fish
parasite biodiversity and community structure. Parassitologia, 39: 237-241.
Diamant, A.; Banet, A.; Paperna, I.; Westernhagen, H.V.; Broeg, K.; Krüner, G. Körting,
W.; Zander, S. 1999. The use of fish metabolic, parasitological indices in pollution
monitoring. II. The Red Sea and Mediterranean. Helgoland Marine Research, 53: 195-
208.
Dogiel, V.A. 1961. Ecology of the parasites of freshwater fishes. In: Dogiel, V.A.
Parasitology of fishes. Translated by Z. Kabata. 1st ed. Edinburgh; London: Oliver
and Boyd. p. 147-148
Dumbo, J.C. 2014. Espécies de metazoários parasitos do Acestrorhyncus falcirostris
(Cuvier, 1819) (Characiformes: Acestrorhynchidae) de lagos de várzea da Amazônia
central. Dissertação de Mestrado, Instituto Nacional de Pesquisas da Amazônia –
INPA, Manaus, Amazonas. 134p.
Eiras, J.C. 1994. Elementos de ictioparasitologia. Porto, Fundação Engenheiro Antônio de
Almeida, 339p.
Eiras. J.C.; Takemoto R.M; Pavanelli G.C. 2006. Métodos de estudos e técnicas
laboratoriais em parasitalogia de peixes. 2ª Ed., Eduem, Universidade Estadual de
Maringá, Maringá, Brasil. 199p.
Eiras. J.C.; Takemoto R.M.; Pavanelli G.C. 2010. Diversidade dos parasitas de peixes de
água doce do Brasil. Ed. Clichetec, Maringá, Brasil. 333p.

73
El-Rashid, H.; Boxshall, G. A. 2001. The mesoparasitic genera of the Ergasilidae
(Copepoda) with descriptions of new species of Paeonodes Wilson and Therodamas
Kroyer. Systematic Parasitology, 50: 199-217.
Evans, D.W.; Irwin, S.W.B.; Fitzpatrick, S. 2001. The effect of digenean (Platyhelminthes)
infections on heavy metal concentrations in Littorina littorea. Journal of Marine
Biological Association of the United Kingdom, 81: 349-350.
Farkas, A.; Salánki, J.; Varanka, I. 2003. Crustaceans as biological indicators of heavy
metal pollution in Lake Balaton (Hungary). Hydrobiologia, 509: 359-364.
Favero, N.; Cattalini, F.; Bertaggia, D.; Albergoni, V. 1996. Metal accumulation in a
biological indicator (Ulva rigida) from the lagoon of Venice (Italy). Archives of
Environmental Contamination and Ecotoxicology, 31: 9-18.
Fernandes, M.M.; Kohn, A. 2014. South American trematodes parasites of amphibians and
reptiles. 1 ed. Rio de Janeiro: Oficina de Livros. 226p.
Fernandes, M.M; Justo, M.C.N.; Cárdenas, M.Q.; Cohen, S.C. 2015. South American
trematodes parasites of birds and Mammals. 1 ed. Rio de Janeiro: Oficina de Livros.
549p.
Froese, R.; Pauly, D. 2017. FishBase. World Wide Web electronic publication.
www.fishbase.org, version (06/2017). Acesso 20/06/2017.
Galli, P.; Crosa. L.; Mariniello, M.; Ortis, M.; D´amelio, D. 2001. Water quality as a
determinant of the composition of fish parasite communities. Hydrobiologia, 452:
173-179.
Giese, E.G. 2010. Diversidade de helmintos de Ageneiosus ucayalensis Castelnau 1855
(Pisces: Siluriformes) da foz do rio Guamá e baía do Guajará, Belém, Pará. Tese de
Doutorado. Universidade Federal do Pará, Belém, Pará. 157p.
Goulding, M. 1980. The fishes and the forest, explorations in Amazonian natural history.
University of California Press, Berkeley. USA. 280p.
Goulart, M.; Callisto, M. 2003. Bioindicadores de qualidade de água como ferramenta em
estudos de impacto ambiental. Revista da FAPAM, 2: 32-43.
Guidelli, G.; Tavechio, W.L.G.; Takemoto, R.M.; Pavanelli, G.C. 2003. Fauna parasitária
de Leporinus lacustris e Leporinus friderici (Characiformes, Anostomidae) da planície

74
de inundação do alto rio Paraná, Brasil. Acta Scientiarum Biological Science, 28: 281-
290.
Handerson, P.A.; Crampton. W.G.R. 1997. A comparison of fish diversity and abundance of
between nutrient-rich and nutrient lakes in the Upper Amazon. Journal of Tropical
Ecology, 13: 173-198.
Hossain, M.D.; Hossain, M.K.; Rahman, M.H.A.; Akter, A.; Khanom, D.A. 2008.
Prevalence of ectoparasites of carpfingerlings at Santaher, Bogra. University Journal of
Zoology Rajshahi University, 27: 17-19.
Hughs, R.C.; Higginbotham, J.W.; Clary, J.W. 1942. The trematodes of reptiles –
Systematic section part I. The American Midland Naturalist, 20: 109-134.
Isaac, V.J.; Fabré, N.N.; Silva, C.O.; Ruffino, M.L.; Saint-Paul, U. 2012. Ecologia da fauna
íctica. In: Vandick et al. (Org). Peixes e pesca no rio Solimões-Amazonas: uma
avaliação integrada. p.207-249.
Jogunoori, W.; Kritsky, D.C.; Venkatanarasaiah, J. 2004. Neotropical Monogenoidea. 46.
Three new species from the gills of introduced aquarium fishes in Indian, the proposal
of Heterotylus n. g. and Diaphorocleidus n. g., and the reassignment of some previously
described species of Urocleidoides Mizelle & Price, 1964 (Polyonchoinea:
Dactylogyridae). Systematic Parasitology, 58: 115-124.
Jonsson, C.M.; Paraíba, L.C.; Aoyama, H. 2009. Algae acid phosphatase affected by ten
metals and enzymatic parameters determination in presence of mercury and copper.
Pesticidas: Revista de Ecotoxicologia e Meio Ambiente, 19: 1-10.
Junk. W.J. 1983 As águas da região Amazônica. In: Salati, E.; Junk, W.J.; Shubart, H.O.R.;
Oliveira, A.E. (Eds.). Amazonia: desenvolvimento, integração e ecologia. Conselho
nacional de desenvolvimento científico e tecnológico. Ed Brasiliense, Brasília, Brasil.
p.45-100.
Junk, W.J.; Bayley, P.B.; Sparks, R.E. 1989. The flood pulse concept in river-floodplain
systems. Canadian Journal of Fishers and Aquatic, 106: 110-127.
Kanev, I.; Radev, V.; Fried, B. 2002. Superfamily Clinostomoidea. In: Gibson, D.I.; Jones,
A.; Bray, R.A (Edt). Keys to trematodes, CABI, The Natural History Museum, London,
UK. p.111-125.

75
Kennedy, C.R.; Andersen, K. 1982. Redescription of Acinstrocephalus microcephalus
(Ruldophi) (Cestoda, Pseudophyllidea) from sunfish Mola mola. Zoologica Scripta,
11: 101-105.
Khan, R.A.; Kiceniuk, J. 1983. Effects of crude oils on the gastrointestinal parasites of two
species of marine fish. Journal of Wildlife Disease, 19: 253-258.
Kohn, A.; Fernandes, B.M.M.; Baptista-Farias, F.D. 1994. Metacercariae of Diplostomum
(Austrodiplostomum) compactum (Trematoda, Diplostomidae) in the eyes of
Plagioscion squamosissimus (Teleostei, Sciaenidae) from the Reservoir of the
Hydroelectric Power Station of Itaipu, Brazil. Memórias do Instituto Oswaldo Cruz,
90: 341-344.
Kohn, A.; Fernandes, M.M.; Cohen, S.C. 2007. South American trematodes parasites of
Fishes. 1 ed. Fiocruz, Rio de Janeiro. 318p.
Krebs, C.J. 1999. Ecological methodology. 2. ed., Editora Benjamin Cummings, Menlo
Park, CA. 250p.
Kritsky, D. C.; Boeger, W.A.; Popaoglo, F. 1995. Neotropical Monogenoidea. 22.
Variation in Scleroductus species (Gyrodactylidea, Gyrodactylidae) from Siluriformes
fishes of sutheastern Brazil. Journal of Helminthology of Washigton, 62: 53-56.
Kritsky, D. C.; Stockwell, C. A. 2005. New species of Gyrodactylus (Monogenoidea,
Gyrodactylidae) from the white sands pupfish, Cyprinodon tularosa, in New Mexico.
The Southwestern Naturalist, 50: 312-317.
Kritsky, D.C. 2007. Heteropriapulus nom. nov. (Monogenoidea: Dactylogyridae) for
Heterotylos Jogunoori, Kritsky & Venkatanarasiah, 2004, a junior homonym of
Heterotylus Kirsch & Reitter, 1913 (Coleoptera: Curculionidae). Systematyc
Parasitology, 68: 233.
Lacerda, A.C.F.; Roumbedakis, K.; Bereta Junior, J.G.S.; Nuñer, A.P.O.; Petrucio, M.M.;
Martins, M.L. 2017. Fish parasites as indicators organic pollution in southern Brazil.
Journal of Helminthology, June p.1-10.
Lafferty, K.D. 1997. Environmental parasitalogy: what can parasites tell us about human
impacts on the environment? Parasitalogy Today, 13: 251-255.

76
Landsberg, J.H.; Blakesley, B.A.; Reese, R.O.; Mcrae, G.; Forstchen, P.R. 1998. Parasites
of fish as indicators of environmental stress. Environmental Monitoring and
Assessment, 51: 211-232.
Lewis, J.; Hoole, D. 2003. Parasitism and environmental pollution: parasites and hosts as
indicators of water quality. Parasitology, 126: S1-S3.
Lima, C.A.; Goulding, M. 1998. Os frutos do tambaqui ecologia, conservação e cultivo na
Amazônia. Sociedade Civil Mamirauá, MCT-CNPq. 186p.
Lopes, L.P.C.; Varella, A.M.B.; Malta, J.C.O. 2009. Metazoan parasites of
Pseudoplatystoma punctifer (Linneus, 1766) and Pseudoplatystoma tigrinum (Spix &
Agassiz, 1829) (Siluriformes: Pimelodidae) of the central Amazon Basin, Brazil.
Biologia Geral e Experimental, 9: 3-15.
Loreau, M.; Roy, J.; Tilman, D. 2005. Liking ecosystem and parasite ecology. In: Thomas,
F.; Renaud, F.; Guégan, J.F. (Edt). Parasitism and Ecosystem. Oxford University
Press. p.13-21.
Luque, J.L.; Mouillot, D.; Poulin R. 2004. Parasite biodiversity and its determinants in
coastal marine teleost fishes of Brazil. Parasitology, 128: 671-682.
Mackenzie, K.; Williams, H.H.; Williams, B.; McVicar, A.H.; Siddal, R. 1995. Parasites
as indicators of water quality and the potential use of helminth transmission in marine
pollution studies. Advanced Parasitalology, 35: 85-144.
Madi, R.R. 2005. Utilização dos helmintos parasitas de Geophagus brasiliensis (Quoy &
Gaimard, 1824) (Cichlidae: Perciformes) como indicadores ambientais. Tese de
Doutorado, UNICAMP, São Paulo, Brasil. 110p.
Madi, R.R.; Ueta, M.T. 2009.O papel de Ancyrocephalinae (Monogenea: Dactylogyridae)
parasito de Geophagus brasiliensis (Pisces: Cichlidae) como indicador ambiental.
Revista Brasileira de Parasitologia Veterinária, 18(2): 38-41.
Magurran, A.E. 1983. Diversidad ecológica y su medición. Croom Helm, Barcelona,
Espanha. 200p.
Malta, J.C.O.; Varella, A.M.B. 2009. Os crustáceos branquiúros parasitas de peixes
(Argulidae: Maxillopoda). In: Fonseca, C.R.V.; Magalhães, C.; Rafael, J.A.; Franklin,
E. (Eds.). A Fauna de Artrópodos da Reserva Florestal Adolpho Ducke. Estado Atual
do Conhecimento Taxonômico e Biológico. Instituto Nacional de Pesquisas da

77
Amazônia, Manaus, Amazonas. p. 17-20.
Marcogliese, D.J. 1995. The role of zooplankton in the transmission of helminth parasites
to fish. Reviews in Fish Biology and Fisheries, 5: 336-371.
Marcogliese, D.J. 2004. Parasites: small players with crucial roles in the ecological
theather. Ecohealth, 2: 151-161.
Marcogliese, D.J. 2005. Parasites of the superorganism: are they indicators of ecosystem
health? International Journal of Parasitalology, 35: 705-716.
Matthews, R. A.; Buikema, A. L.; Cairns Jr., J., 1982. Biological monitoring part II A:
Receiving system functional methods relationships, and indices. Water Research,
16:129-139.
Moller, H. 1987. Pollution and parasitism in the aquatic environment. International
Journal of Parasitology, 17: 353-361.
Morais, A.M. 2011. Biodiversidade de parasitos da piranha vermelha Pygocentrus
nattereri (Kner, 1858) (Characiformes: Serrasalmidae) e sua avaliação como
bioindicadores na Amazônia central. Tese de Doutorado. Instituto Nacional de
Pesquisas da Amazônia. Manaus, Amazonas. 234p.
Morley, N.J.; Crane, M.; Lewis, J.W. 2001. Toxicity of cadmium and zinc to Diplostomum
sphateceum (Trematoda: Diplostomidae) cercarial survival. International Journal for
Parasitology, 31: 1211-1217.
Muniz-Pereira, L.C.; Vieira, F.M.; Luque, J.L. 2009. Checklist of helminth parasites of
threatened vertebrate from Brazil. Zootaxa, 2123: 1-45.
Olsen, O.W. 1978. Animal parasites their life cycles and ecology. University Park press
Baltimore, London and Tokyo. 562p.
Ogut, H,; Palm, H.W. 2005. Seasonal dynamics of Trichodina spp. on whiting (Merlangius
merlangus) in relation to organic pollution on the eastern Black Sea coast of Turkey.
Parasitology Reserch, 96:149-153
Overstreet, R.M. 1988. Aquatic pollution problems, Southeastern U.S. coast:
histophatological indicators. Aquatic toxicology, 11: 213-239.
Overstreet, R.M. 1997. Parasitological data as monitors of environmental health.
Parassitologia, 39: 169-175.
Paggi, J.C. 1976. Una nueva especie de Therodamas (Therodamasidae; Cyclopoida)

78
copepod parasite de peces de agua dulce de la república Argentina. Physis, 91: 93-102.
Palm, H.W.; Dobberstein, R.C.1999. Occurrence of trichodinid ciliates (Peritricha:
Urceolariidae) in the Kiel Fjord, Baltic Sea, and its possible use as a biological
indicator. Parasitology Research, 85:726-732.
Petesse, M.L.; Siqueira-Souza, F.K.; Freitas, C.E.C.; Petrere Jr, M. 2016. Selection of
reference lakes and adaption of a fish multimetric index of biotic integrity to six
amazon floodplain lakes. Ecological Engineering, 97: 535-544.
Pompeu, P. P.; Alves, M. C. B.; Callisto, M. 2004. The effects of urbanization on
biodiversity and water quality in the Rio das Velhas basin, Brazil. American Fisheries
Society, 8:243-251.
Ponce de León, G.P; Prieto, L.G. 2001. Los parásitos en el contexto de la biodiversidad y
la conservación. Conabio. Biodiversitas, 34: 11-15.
Porto, D.B. 2009. Metazoários parasitas de Pterygoplichthys pardalis (Castelnau, 1855)
de lagos de várzea do rio Solimões entre Manaus e Coari, amazonas. Dissertação de
mestrado, Instituto Nacional de Pesquisas da Amazônia, Manaus, Amazonas. 34p.
Porto, D.B.; Vital, J.F.; Santos, A.K.S.; Morais, A.M.; Varella, A.M.B.; Malta, J.C.O.
2012. Metazoários parasitos de Pterygoplichthys pardalis (Castelnau, 1855)
(Siluriformes: Loricariidae) da Amazônia central. Revista Brasileira de Zoociências,
14: 35-40.
Poulin, R. 1995. Phylogeny, Ecology, and the richness of parasite communities in
vertebrates. Ecological Monographs, 65: 283-302.
Prado, K.L.L. 2005. Assembleias de peixes associadas as macrófitas aquáticas em lagos
de várzea do baixo rio Solimões. Dissertação de mestrado, Instituto Nacional de
Pesquisas da Amazônia/ Universidade Federal do Amazonas, Manaus-Amazonas, 66p.
Queiroz, H.L.; Sardinha, A.D. 1999. A preservação e o uso sustentado dos pirarucus em
Mamirauá. In: Estratégias para manejo de recursos pesqueiros em Mamirauá. Queiroz,
H.L.; Crampton, W.G.R. (Edt) Sociedade Civil Mamirauá, MCT-Cnpq. p.108-141.
Queiroz, H.L. 2000. Natural history and conservation of pirarucu, Arapaima gigas at the
Amazonian Várzea: Red giants in muddy water. Tese de Doutorado/ University of Saint
Andrews, 226p.

79
Queiroz, J.F.; Trivinho-Strixino, S.; Nascimento, V.M.C. 2000. Organismos bentônicos
bioindicadores da qualidade das águas da bacia do médio São Francisco. Novembro,
Comunicado técnico Embrapa meio ambiente, 4p.
Ramos, I.P.; Franceschine, L.; Zago, A.C.; Zica, E.O.P.; Wunderlich, A.C.; Carvalho, E.D.;
Silva, R.J. 2013. New host records and a checklist of fishes infected with
Austrodiplostomum compactum (Digenea: Diplostomidae) in Brazil. Revista
Brasileira de Parasitologia Veterinária, 22: 511-518.
Rashed, M.N. 2001. Monitoring of environmental heavy metals in fish from Nasser Lake.
Environment International. 27: 27-33.
Ramsdorf, W. 2007. Utilização de duas espécies de Astyanax (Astyanax sp. B e A.
altiparana) com bioindicadores de região contaminada por agrotóxico (Fazenda
Cangüiri- UFPR). Dissertação de Mestrado. Departamento de Genética, Universidade
Federal do Paraná, Curitiba, 127p.
Rego, A.A. 1994. Order Proteocephalidea. In: Khalil, L.F.; Jones, A.; Bray, R.A. (Eds.)
Keys to the cestodes parasites of vertebrates. University press, Cambridge. p.257-293.
Rego, A.A.; Chubb, J.C.; Pavanelli, G.C. 1999. Cestodes in South American freshwater
teleosts fishes: Keys to genera and brief description of species. Revista Brasileira de
Zoologia, 16: 299-367.
Rego, A.A. 2003. Problems of classification South American proteocephalids (Cestoda).
On a new classification of the group. Acta Scientiarum: Biological Sciences, 25: 15-
22.
Reis, R.E.; Kullander, S.O.; Ferraris, C.J.Jr. 2003. Check list of the freshwater fishes of
South and Central America. Edipucrs, Porto Alegre, Brasil. 742p.
Roberts, R.J. 1978. Fish Pathology. University Press, Aberdeen, Scotland. 318p.
Rocha, O. 2002. Perfil do conhecimento de biodiversidade em águas doces do Brasil. In:
Lewinsohn, T.M; Prado, P.I. (Eds.). Biodiversidade brasileira: síntese do estado atual
do conhecimento. Contexto, São Paulo, Brasil. p. 165-169.
Rhode, K.; Hayward, C.; Heap, M. 1995. Aspects of the ecology of metazoan ectoparasites
of marine fishes. International Journal for Parasitology, 25: 945-970.
Salati, E. 1983. O clima atual depende da floresta. In: Salati, E.; Junk, W.J.; Shubart,
H.O.R.; Oliveira, A.E. (Eds.). Amazônia: desenvolvimento, integração e ecologia.

80
Conselho nacional de desenvolvimento científico e tecnológico. Brasília, Brasil. p.45 -
100.
Santos, G.M.; Ferreira, E.J.G. 1999. Peixes da bacia Amazônica. In: Lowe-McConnell,
R.H. Estudos ecológicos de peixes tropicais. Editora da Universidade de São Paulo. p.
345-354.
Santos, G.; Ferreira, E.; Zuanon, J. 2006. Peixes comerciais de Manaus. Ibama, Manaus,
Brasil. 144p.
Schaffer, G.D.; Rego, A.A.; Pavanelli, G.C. 1992. Memórias do Instituto Oswaldo Cruz,
87: 257-258.
Schmidt, V.; Zander, S.; Körting, W., Steinhagen, D. 2003. Parasites of flounder
Platichthys flesus (L.) From the german Bight, Nort Sea, and their potential use in
ecosystem monitoring. Community structure and fish parasite biodiversity. Helgoland
Marine Research, 57: 252-261.
Silva, R.E. 2010. Ecologia alimentar da ariranha (Pteronura brasiliensis) e da lontra
Neotropical (Lontra longicaudis) no parque nacional do Jaú, Amazonas, Brasil.
Dissertação de Mestrado, Instituto Nacional de Pesquisas da Amazônia – INPA. 48p.
Silva-Souza, A.T.; Shibatta, O.A.; Matsumura-Tundisi, T.; Tundisi, J.G.; Dupas, F.A.
2006. Parasitas de peixes como indicadores de estresse ambiental e eutrofização. In:
Tundisi, J.G; Tundisi, T.M.; Galli, C.S. (Org.). Eutrofização na América do Sul:
causas, consequências e tecnologias para gerenciamento e controle. 1 ed. São Carlos:
Instituto Internacional de Ecologia. p.373-386.
Smith, N.J.H. 1979. A pesca no rio Amazonas. Conselho Nacional de Desenvolvimento
Científico/ Instituto Nacional de Pesquisas da Amazônia. Falangola, Belém, Pará,
Brasil. 154p.
Soares, M.G.M; Costa, L.E.; Siqueira-Souza, K.F.; Anjos, B.D.H.; Yamamoto, C.K.;
Freitas, C.E.C. 2007. Peixes de lagos do médio rio Solimões. EDUA, Manaus, Brasil.
172p.
Souza, A.K.S.; Porto, D.B.; Malta, J.C.O. 2017. Diplostomum (Austrodiplostomum)
compactum (Lutz, 1928) (Platyhelminthes, Digenea) free Metacercariae in the eyes of
fishes from the floodplain lakes to the lower Solimões river, Brazil. Neotropical
Helminthology, 11:156-163.

81
Sures, B.; Taraschewski, H.; Rydlo, M. 1997. Intestinal Fish Parasites as Heavy Metal
Bioindicators: A Comparison between Acanthocephalus lucii (Palaeacanthocephala)
and the Zebra Mussel, Dreissena polymorpha. Bulletin of Environmental
Contamination and Toxicology, 59: 14-21.
Sures, B. 2001. The use of fish parasites as bioindicators of heavy metals in aquatic
ecosystem: a review. Aquatic Ecology, 35: 245-255.
Takemoto, R.M.; Lizama, M. de los A.P, Guidelli, G.M., Pavanelli, G.C. 2004. Parasitas
de peixes de águas continentais. In: Sanidade de Organismos Aquáticos.1 ed. São
Paulo: Editora Varela. p.179-197.
Thatcher, V.E. 1979. Uma nova espécie de Gorytocephalus Nickol e Thatcher, 1971
(Acanthocephala: Neoechinorhynchidae) do acari bodó (Pisces: Loricariidae) da
Amazônia, Brasil. Acta Amazonica, 9: 199-202.
Thatcher, V.E. 1981. Patologia de peixes da Amazônia brasileira, 1. Aspectos gerais. Acta
Amazonica, 11: 125-140.
Thatcher, V.E. 1986. The parasitic Crustaceans of fishes from the Brazilian Amazon, 16,
Amazonicopeus elongatus gen. et. sp. nov. (Copepoda: Poecilostomatoida) with the
proposal of Amazonicopeidae fam. nov. and remarks on its pathogenicity. Amazoniana,
10: 49-56.
Thatcher, V.E. 1993. Trematódeos Neotropicais. Instituto Nacional de Pesquisas da
Amazônia (INPA), Manaus, Amazonas, Brasil. 553p.
Thatcher, V.E. 2006. Amazon fish parasites. 2a Ed., Editora Aquatic Biodiversity in Latin
America, Sofia, Moscow. 508p.
Thatcher, V.E.; Varella, A.B. 1981. Duas novas espécies de Megacoelium Szidat, 1954
(Trematoda: Heploporidae), parasitas estomacais de peixes da Amazônia Brasileira,
com uma redefinição do gênero. Acta Amazonica, 11: 285-289.
Travassos, L.; Freitas, J.F.; Kohn, A. 1969. Trematódeos do Brasil. Memórias do Instituto
Oswaldo Cruz, 67: 1-886.
Val, V.M.F.A.; Silva, M.N.P.; Cardoso, S.S.O.; Val, A.L. 2011. Indicadores em
ecotoxicologia. In: Cavalcante, K.V.; Rivas, A.A.; Freitas, C.E.C.; Mourão, R.R.
(Org.). Indicadores Socioambientais e atributos de referência para o trecho Urucu-
Coari-Manaus, rio Solimões, Amazônia Ocidental. p.113-118.

82
Valtonen, E.T.; Holmes, J.C.; Koskivaara, M. 1997. Eutrophication, pollution,
fragmentation: effects on parasites communites in roach (Rutilus rutilus) and perch
(Perca fluviatilis) in four lakes in central Finland. Canadian Journal Fisheries and
Aquatic Science, 54: 572-585.
Valtonen, E.T.; Holmes, J.C.; Aronen, J.; Rautalahti, I. 2003. Parasites communities as
indicator of recovery from pollution: parasites of roach (Rutilus rutilus) and perch
(Perca fluviatilis) in central Finland. Parasitology, 126: S43-S52.
Varella, A.M.B. 1992. Copépodos (Crustacea) parasitas das fossas nasais de peixes,
coletados na região de Rondônia, Brasil. Tese de Doutoramento, Instituto de
Biociências do Campus de Rio Claro, Universidade Estadual Paulista “Júlio de
Mesquita Filho”, Rio Claro, São Paulo. 105p.
Varella, A. 1994. Gamidactylus bryconis sp.n. (Copepoda, Poecilostomatoida,
Vaigamidae) das fossas nasais de peixes, Brycon pellegrini Holly, 1929 e B.
melanopterus (Cope, 1872) da Amazônia Brasileira. Acta Amazonica, 24: 145-152.
Varella, A.; Malta, J.C.O. 1995. Gamidactylus hoplius sp.n. (Copepoda,
Poecilostomatoida, Vaigamidae) das fossas nasais de Hoplias malabaricus (Bloch,
1794) (Characiformes, Erythrynidae) da Amazônia Brasileira. Acta Amazonica, 25:
281-288.
Varella, A.M.B.; Malta, J.C.O. 2001. Brasergasilus mamorensis sp. n. (Copepoda:
Ergasilidae) from the nasal cavities of Hydrolycus pectoralis (Guenther, 1866)
(Characiformes: Cynodontidae) from the Brasilian Amazon, and considerations about
Abergasilinae. Acta Amazonica, 30: 323-330.
Varella, A.M.B.; Malta, J.C.O. 2009. Copepoda Cyclopoida e Poecilostomatoida. In:
Fonseca, C.R.V.; Magalhães, C.; Rafael, J.A.; Franklin, E. (Eds.). A fauna de
artrópodes da Reserva Florestal Adolpho Ducke. Estado atual do conhecimento
taxonômico e biológico. Instituto Nacional de Pesquisas da Amazônia, Manaus,
Amazonas, p. 21-24.
Viana, R.T. 2011. Filogenia e biogeografia histórica dos Gyrodactylidae (Monogenoidea):
morfologia, moléculas e evidência total. Tese de Doutorado, Universidade Federal do
Paraná, Paraná. 295p.

83
Vidal-Martínez, V.M.; Daniel Pech, D.; Sures, B.; S. Purucker, T.; Poulin, R. 2009. Can
parasites really reveal environmental impact? Trends in parasitology, 26:44-51.
Washington, H. G., 1984. Diversity, biotic and similarity indices. A review with special
relevance to aquatic ecosystems. Water Research, 18: 653-694.
Weber, C. 1992. Revision du genre Pterygoplichthys sensu lato (Pisces, Siluriformes,
Loricariidae). Revue Française Aquariologie, 19: 1–36.
Williams, H.H.; Mackenzie, K.; Mccarthy, A.M. 1992. Parasites as biological indicators of
the population biology, migrations, diet, and phylogenetics of fish. Reviews in Fish
Biology and Fisheries, 2: 144-176.
Yossa, M.I.; Araújo-Lima, C.A.R.M. 1998. Detritivory in two Amazonian fish species.
Journal of Fish Biology, 52: 1141-1153.
Zar, J.H. 1996. Biostatistical analysis. 3Ed. Prentice-Hall, Upper Saddle River, New
Jersey, USA. 662p.
Zica, E.O.P.; Brandão, H.; Zawadzki, C.H.; Nobile, AB.; Carvalho, E.D.; Silva, R.J. 2010.
The occurrence of Austrodiplostomum compactum (Lutz, 1928) (Digenea:
Diplostomidae) metacercariae in the eye of Loricariidae fish (Siluriformes:
Osteichthyes: Loricariidae) from Brazil. Journal of Helminthology, 85: 73-79.
Zimmermann, S.; Sures, B.; Taraschewski, H. 1999. Experimental Studies on Lead
Accumulation in the Eel-Specific Endoparasites Anguillicola crassus (Nematoda) and
Paratenuisentis ambiguous (Acanthocephala) as Compared with Their Host, Anguilla
anguilla. Archives of Environmental Contamination and Toxicology, 37: 190-195.
Zuben, C.J.V. 1997. Implicações da agregação espacial de parasitas para a dinâmica
populacional na interação hospedeiro-parasita. Revista de Saúde Pública, 31: 523-530.



![ORNAMENTAL AQUACULTURE [objectives]science.kln.ac.lk/.../images/stories/Lecture_Materials/ornamental.pdf · annual fish, meaning killifish ... (Pterygoplichthys, Loricaria), Discus,](https://img.dokumen.tips/doc/110x75/5b31a0c57f8b9aa0238b6b89/ornamental-aquaculture-objectives-annual-fish-meaning-killifish-pterygoplichthys.jpg)


![ANTICORPOS - MEUS SOLDADINHOS, MEUS PROTETORES · 2020. 8. 10. · anticorpos - meus soldadinhos, meus protedores, por joão josé da costa [ 2] anticorpos - meus soldadinhos, meus](https://img.dokumen.tips/doc/110x75/607bc6fd4f65ba4ee03efbb8/anticorpos-meus-soldadinhos-meus-protetores-2020-8-10-anticorpos-meus.jpg)























![[Recensão a:] Emmanuel Faye , Heidegger 1'introduction du ......Heidegger, Carl Schmitt, Erik Wolf, Oskar Becker ou Ernst Jünger. Atese fundamental acerca do nazismo intrínseco](https://img.dokumen.tips/doc/110x75/610a46134e88b17c133fc66f/recenso-a-emmanuel-faye-heidegger-1introduction-du-heidegger-carl.jpg)