Embed Size (px)
Citation preview

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2019
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 1528
Prostasomes as Diagnostic,Prognostic and TherapeuticVesicles
LOUISE DUBOIS
ISSN 1651-6206ISBN 978-91-513-0541-7urn:nbn:se:uu:diva-369166

Dissertation presented at Uppsala University to be publicly examined in Universitetshuset, SalIX, Biskopsgatan 3, Uppsala, Saturday, 16 February 2019 at 13:15 for the degree of Doctorof Philosophy (Faculty of Medicine). The examination will be conducted in Swedish. Facultyexaminer: Senior professor Lucia Mincheva-Nilsson (Umeå Universitet, Institutionen förklinisk mikrobiologi).
AbstractDubois, L. 2019. Prostasomes as Diagnostic, Prognostic and Therapeutic Vesicles. DigitalComprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 1528.56 pp. Uppsala: Acta Universitatis Upsaliensis. ISBN 978-91-513-0541-7.
This thesis explores prostasomes and their ability to be used as a new diagnostic tool for prostatecancer. Alongside diagnosis, this thesis also suggests prostasomes as a tool for prognosis andtherapeutic treatment in patients with prostate cancer. By further characterizing prostasomeswe can identify a biomarker and also a method of visualizing and interpreting the informationprovided in order to conduct a correct and fast diagnosis for prostate cancer.
In Paper I, we show that the prostasomal bilayered membrane consists of lipid rafts, clustersthat holds cholesterol, sphingolipids and gives receptors a rigid platform upon which towork. We compare the proteomic content of prostasome lipid rafts with the entire prostasomemembrane in the search for a specific biomarker.
In Paper II, we show that purified lipid rafts from the prostasome membrane can re-vesiculateand create new bioengineered vesicles. These new vesicles can carry different agents insidethem and we find that the method is also applicable to blood cells. This suggests a new methodfor cell-specific delivery of drugs and cancer therapy.
In Paper III, we further characterize the prostasome membrane, this time mapping purinergicreceptors. This could be used in the development of prostate cancer treatment and to gain betterunderstanding of how prostasomes interact with surrounding cells in their ambient environment.
In Paper IV, we investigate the difference in thymidine kinase 1 (TK1) enzyme activitybetween prostasomes and malignant exosomes. TK1 is considered to be a biomarker of cellproliferation and could therefore be used as a biomarker for prostate cancer diagnosis andprogression.
In summary, this thesis contributes to the puzzle of how to better diagnose, prognose and treatprostate cancer. Although it is mainly pre-clinical research it opens up new possibilities for thediagnosis and treatment of prostate cancer.
Keywords: Exosomes, Prostasomes, Lipid rafts, Bioengineered vesicles, Prostate cancer,Purinergic receptors, Thymidine kinase 1
Louise Dubois, Department of Medical Sciences, Clinical Chemistry, Akademiska sjukhuset,ingång 40, 5tr., Uppsala University, SE-75185 Uppsala, Sweden.
© Louise Dubois 2019
ISSN 1651-6206ISBN 978-91-513-0541-7urn:nbn:se:uu:diva-369166 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-369166)

Till min familj

“Everyone is a genius. But if you judge a fish by its ability to climb a tree, it will live its whole life believing that it is stupid.” -Albert Einstein

List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Dubois, L., Ronquist, GK., Ek, B., Ronquist, G., Larsson, A.
(2015) Proteomic profiling of detergent resistant membranes (li-pid rafts) of prostasomes. Molecular and Cellular Proteomics, 14(11):3015-22
II Dubois, L*., Löf, L*., Larsson, A., Hultenby, K., Waldenström, A., Kamali-Moghaddam, M., Ronquist, G., Ronquist, GK. (2018) Human erythrocyte-derived nanovesicles can readily be loaded with doxorubicin and act as anticancer agents. Cancer Re-search Frontiers, 4(1): 13-26
III Dubois, L., Larsson, A. (2018) Occurrence of purinergic recep-tors in the prostasome (prostate epithelial cell-derived exo-somes). Purinergic Signalling, Submitted
IV Dubois, L., Löf., L., Kamali-Moghaddam, M., Larsson, A^., Stålhandske P^ (2018) Increased levels of thymidine kinase 1 in malignant cell-derived exosomes. In manuscript
*Indicates shared first author ^Indicates shared senior author
Reprints were made with permission from the respective publishers.

List of papers not included in this thesis
I Dubois L, Stridsberg M, Kharaziha P, Chioureas D, Meersman N, Panaretakis T, Ronquist KG. (2015) Malignant cell-derived extracellular vesicles express different chromogranin epitopes compared to prostasomes. Prostate, 75: 1063-73
II Ronquist KG, Sanchez C, Dubois L, Chioureas D, Fonseca P, Larsson A, Ullén A, Yachnin J, Ronquist G, Panaretakis T. (2016) Energy-requiring uptake of prostasomes and PC3 cell-derived exosomes into non-malignant and malignant cells. Journal of Extracellular Vesicles, 5: 29877
III Carlsson L, Ronquist G, Elisasson R, Dubois L, Ronquist KG, Larsson A. (2016) High concentrations of the angiogenic peptide VEGF-A in seminal fluid and its association to prostasomes. Clinical laboratory, 62(8):1515-20
IV Löf L, Ebai T, Dubois L, Wik L, Ronquist KG, Nolander O, Lundin E, Söderberg O, Landegren U, Kamali-Moghaddam M. (2016) Detecting individual extracellular vesicles using a multicolor in situ proximity ligation assay with flow cytometric readout. Scientific Reports, 6:34358
V Larssen P, Wik L, Czarnewski P, Eldh M, Lof L, Ronquist KG, Dubois L, Freyhult E, Gallant CJ, Oelrich J, Larsson A, Ronquist G, Villablanca, EJ, Landegren U, Gabrielsson S, Kamali-Moghaddam M. (2017) Tracing cellular origin of human exosomes using multiplex proximity extension assays. Molecular and Cellular Proteomics, 16(3):502-11

Contents
Introduction ................................................................................................... 11
The prostate ................................................................................................... 13
Seminal plasma ............................................................................................. 14
Prostate cancer .............................................................................................. 15 Diagnostics, Prognostics and Therapeutics .............................................. 15
Extracellular vesicles .................................................................................... 17 Exosomes, microvesicles and apoptotic bodies ....................................... 17
Prostasomes .................................................................................................. 20
Lipid rafts ...................................................................................................... 23
Purinergic receptors ...................................................................................... 25
Thymidine kinase 1 ....................................................................................... 27
Methods ........................................................................................................ 29 Purification of prostasomes ...................................................................... 29 Purification of lipid rafts .......................................................................... 30 SDS-PAGE ............................................................................................... 31 Immunoblotting ........................................................................................ 32 Mass spectrometry .................................................................................... 33 Electron microscopy ................................................................................. 34
Scanning electron microscopy ............................................................. 34 Transmission electron microscopy ...................................................... 34
TK1 real-time assay ................................................................................. 34
Perspectives on Paper I: Proteomic profiling of detergent resistant membranes (lipid rafts) of prostasomes ........................................................ 36
Introduction .............................................................................................. 36 Aim and procedure ................................................................................... 36 Summary .................................................................................................. 36
Perspectives on Paper II: Human erythrocyte-derived nanovesicles can readily be loaded with doxorubicin and act as anticancer agents ................. 38
Introduction .............................................................................................. 38

Aim and procedure ................................................................................... 38 Summary .................................................................................................. 39
Perspectives on Paper III: Occurrence of purinergic receptors in human prostasomes (prostate epithelial cell-derived exosomes) .............................. 40
Introduction .............................................................................................. 40 Aim and procedure ................................................................................... 40 Summary .................................................................................................. 40
Perspectives on Paper IV: Increased levels of thymidine kinase 1 in malignant cell-derived exosomes .................................................................. 42
Introduction .............................................................................................. 42 Aim and procedure ................................................................................... 42 Summary .................................................................................................. 42
Conclusions and future perspectives ............................................................. 43
Populärvetenskaplig sammanfattning på svenska ......................................... 45
Acknowledgements ....................................................................................... 48
References ..................................................................................................... 49

Abbreviations
ADT Androgen deprivation therapy ALL Acute lymphoblastic leukemia ALIX Programmed cell death 6-interacting protein APL Acute promyelocytic leukemia ATP Adenosine triphosphate BPH Benign prostate hyperplasia CRPC Castration-resistant prostate cancer DRM Detergent-resistant membrane EMT Epithelial-to-mesenchymal transition ESCRT Endosomal sorting complex required for transport EVs Extracellular vesicles GnRH Gonadotropin-releasing hormone GPI Glycosylphosphatidylinositol ILVs Intraluminal vesicles MAC Membrane attack complex MS Mass spectrometry MVBs Multivesicular bodies PBS Phosphate-buffered saline PM Plasma membrane PSA Prostate-specific antigen ROS Reactive oxygen species RP Radical prostatectomy RT Room temperature SP Seminal plasma TEM Transmission electron microscope TK Thymidine kinase TDP (deoxy)Thymidine diphosphate TMP (deoxy)Thymidine monophosphate TTP (deoxy)Thymidine triphosphate TSG 101 Tumor susceptibility gene 101 protein


11
Introduction
When I was a little girl my father told me that we are all made of stardust and this really sparked my interest in science. The fact that my grandfather was a professor at Chalmers University of Technology helped a bit too, of course.
I wanted to know more about the human body, what it is made of, how it works and what we can do when it doesn’t work. I started my studies in Bio-medicine at Uppsala University in 2008 and I loved it from the start! This was what I wanted to do, although I’m not saying it was always easy. A few “omtentor” later and I was finished with my degree. So, what to do now? I knew I wanted to be a scientist; being at the forefront of knowledge intrigued me and in 2014 I contacted a group that was conducting research about pros-tasomes. I’d never heard about it, but it sounded interesting! This opened up the world of exosomes to me, how they act as nano-messengers in the body and how cancer can both spread and be contained with the help of exosomes.
Every type of cancer has six specific hallmarks: sustaining proliferative sig-naling, evading growth suppressors, activation of invasion and metastasis, en-abling replicative immortality, inducing angiogenesis and resisting cell death [1]. Normal cells control their cell-cycle and a homeostasis of the number of cells is achieved. Cancer cells bind growth factors which help them sustain proliferative signaling [2]. Cancer cells are not only capable of upregulating growth factors, they can also circumvent the powerful tumor suppressor genes, such as the retinoblastoma gene and the p53 gene [3].
When prostate epithelial cells lose their polarity and adherence to each other, they undergo an epithelial-to-mesenchymal transition (EMT) and this helps the cancer cells migrate and invade other parts of the body. Thus, me-tastases are formed [4]. Unlike other cells, including stem cells, cancer cells have the ability to replicate forever and evidence points to the telomers which protect the ends of the chromosomes, as they are involved in unlimited prolif-eration [5]. Tumors need oxygen and nutrients just like normal cells and tissue, but in tumor progression, angiogenic sprouting is activated so that new blood vessels are formed as the tumor grows [6].
Programmed cell death, apoptosis, is something all cells holds as a natural way of resisting cancer development. Unfortunately cancer cells develop a variety of ways of limiting and avoiding apoptosis so they can continue to proliferate [7]. A proposal for another hallmark is evading immune destruc-tion, meaning tumors somehow manage to go “under the radar” and avoid

12
detection from the body’s immune system, thereby evading eradication [8]. A further emerging hallmark is the reprogramming of energy metabolism. In the 1930s [9, 10], Otto Warburg discovered what is now known as the “Warburg effect”, where cancer cells produce their energy through aerobic glycolysis [11].
Being able to solve the puzzle of cancer, or at least help find one piece of the puzzle, has been the motivation for the duration of my PhD. Now, I get to present my thesis to you all. I am so happy and proud of what I have achieved. I hope whoever reads this thesis will find it inspiring.
13
The prostate
The male accessory glands consist of the seminal vesicles, the bulbourethral glands (also known as Cowper’s glands) and the prostate gland. When ejacu-lation occurs, sperm from the epididymis is transported by the vas deferens to the ejaculatory ducts within the prostate, where it mixes with fluids from the male accessory glands and forms semen [12]. The prostate is divided into two lobes (left and right) and three zones: the central zone, transition zone and peripheral zone. A large portion of the gland is also encapsulated [13].
The prostate gland continues to grow throughout a man’s life, from about 20 g in his thirties to about 30 g in his eighties [14]. If the prostate increases in size the urethra is obstructed and urinating becomes increasingly harder. This is not normally noticed until a tumor occurs or there is a benign hyper-plasia in the prostate.
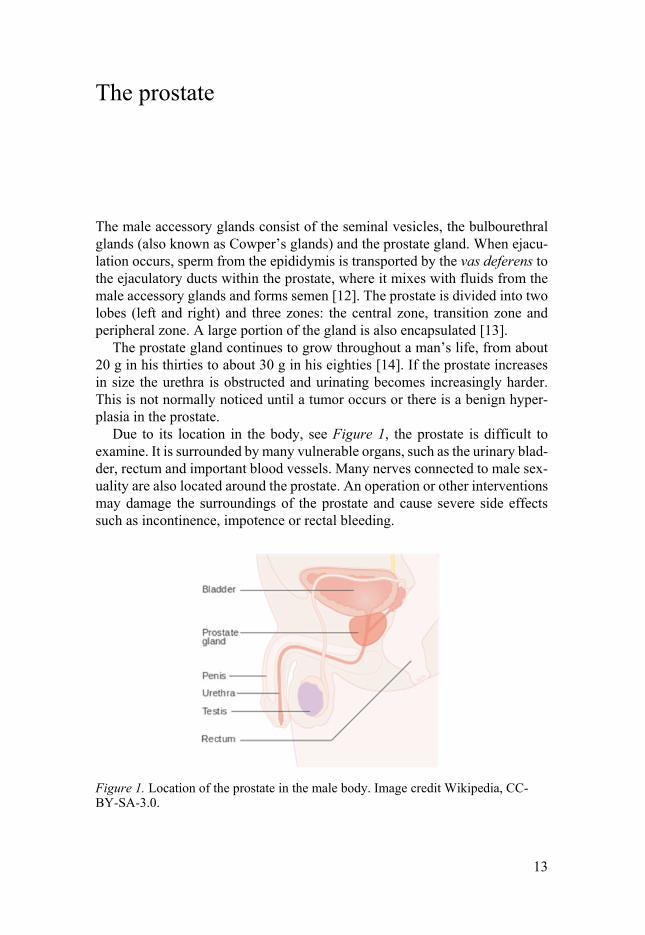
Due to its location in the body, see Figure 1, the prostate is difficult to examine. It is surrounded by many vulnerable organs, such as the urinary blad-der, rectum and important blood vessels. Many nerves connected to male sex-uality are also located around the prostate. An operation or other interventions may damage the surroundings of the prostate and cause severe side effects such as incontinence, impotence or rectal bleeding.
Figure 1. Location of the prostate in the male body. Image credit Wikipedia, CC-BY-SA-3.0.

14
Seminal plasma
Seminal plasma (SP) is ejaculate fluid, minus spermatozoa, which together constitute the semen. SP consists of a mix of fluids from the prostate and the male sex accessory glands. At each ejaculation about 3-4 mL is released and has a pH of 7.5-8 [15], which means the fluid is slightly alkaline. The largest amount of seminal plasma comes from the seminal vesicles (65-75%), the prostate contributes with around one half of this amount (25-30%) and the remainder comes from the bulbourethral glands and the epididymis (<1%)
Before 1935, SP was only regarded as a transportation medium for sperma-tozoa. However, SP is actually essential to the normal function of male fertil-ity [16]. Seminal plasma comprises many proteins, sugars, ions and extracel-lular vesicles (prostasomes), all necessary for the correct function of sperma-tozoa [15].
Semenogelin and fibronectin are proteins that help create a coagulum which then traps the spermatozoa [17, 18]. Liquefaction starts when the se-menogelins are degraded with the help of prostate-specific antigen (PSA), re-leasing spermatozoa [19, 20]. A vaginal pH level of around 4 cleaves the pro-tein progastricsin into the active enzyme gastricsin, which degrades other pro-teins, including PSA. This degradation process may occur in order to prevent immune infertility from forming [21].
Seminal plasma acts as a buffer for the spermatozoa in the acidic environ-ment of the vagina [22] and ions such as calcium, sodium, potassium, magne-sium and zinc help the immunosuppressive and antibacterial functions [23, 24]. The monosaccharide fructose provides energy and nutrition for the sper-matozoa [25] and the movement, acrosome reaction and capacitation are or-chestrated by the influence of prostasomes and are necessary for the sperma-tozoa to mature and fertilize the ovum [26].

15
Prostate cancer
Diagnostics, Prognostics and Therapeutics Prostate cancer is the most frequent type of cancer among men in the Western world [27]. The prognosis is often good and most men die of other causes than prostate cancer. However, it is still the third leading cause of death among men.
Prostate-specific antigen (PSA), which is widely used today for diagnosis, has clear limitations as a cancer biomarker. It is very prostate specific, but not very cancer specific, and a positive PSA test must always be followed by a prostate biopsy, which is an invasive operation that causes much pain for the patient. Also, PSA does not differentiate between aggressive tumors and less aggressive tumors, which means that many men undergo unnecessary prosta-tectomies and therefore have reduced quality of life due to the side effects associated with the surgical procedure, such as impotence and incontinence.
In Sweden alone, more than 10,000 new cases of prostate cancer are dis-covered every year (10,473 cases, National Board of Health and Welfare/So-cialstyrelsen, 2016). With increasing age, the incidence of prostate cancer rises, but the mortality stays the same throughout. In fact, most prostate cancer patients die of other causes than cancer. This is because prostate cancer has many forms, ranging from highly aggressive to non-aggressive. The aggres-sive forms often cause metastases in either the skeleton or the brain, while slow, non-aggressive cancers can remain in the prostate for many years or be held back by hormone treatment. There is a clear genetic component in pros-tate cancer and the risk of being diagnosed with prostate cancer is three times higher if you have a first degree relative who had prostate cancer compared to same age controls [28, 29].
To diagnose prostate cancer today, a blood sample is taken and the level of PSA is measured. PSA is a small protein of 34kD, which means it can easily leak from the prostate cell into the blood stream. It is an exocrine serine pro-tease and is important for seminal fluid where it cleaves the prostate derived protein seminogelin, which helps with semen liquefaction [30]. PSA is an or-gan-specific biomarker but is not a cancer-specific biomarker and an elevated level of PSA does not indicate cancer, but requires further investigation. High level PSA could for instance be due to benign prostate hyperplasia (BPH) [31]. The incidence and prevalence of BPH and prostate cancer both increase with age [14].

16
STHLM3 is a new model for the diagnosis of prostate cancer and Grönberg et al. have shown that it reduces the number of biopsies needed by one third as well as maintaining sensitivity to the Gleason Score in comparison with PSA tests [32]. However, STHLM3 has its flaws: it is very expensive com-pared to PSA tests and is primarily a risk predictor rather than a true cancer diagnosis test.
Commonly used forms of therapy for prostate cancer are drugs, hormones and cytostatics, radiation, prostatectomy or expectancy. Androgen deprivation therapy (ADT) is a form of treatment that uses hormones to reduce androgen levels. The male hormone group androgens can stimulate cancer growth. ADT aims to slow down or even stop cancer cell proliferation, and PSA also de-creases [33]. To achieve this deprivation, there are two solutions: antiandro-gen monotherapy or castration. Castration can then be achieved either surgi-cally, by removing the testicles, or medically by estrogen, which lowers tes-tosterone levels by affecting the hypothalamus. Gonadotropin-releasing hor-mone analog (GnRH) antagonists works by blocking androgen receptors [34]. The patient will need long-term hormone therapy that can cause side effects such as gynecomastia, anemia, hot flashes, fatigue, osteoporosis and de-creased sexual function [35-38]. Unfortunately, over time castration-resistant prostate cancer (CRPC) will develop despite low levels of testosterone [39, 40].
If a patient show signs of an aggressive form of PC, a radical prostatectomy (RP) must be performed. Complications for the patient include incontinence and sexual dysfunction but can be reduced by nerve-sparing surgery. Nerve-sparing surgery cannot be used in all cases [41]. Often, RP is combined with radiation treatment, hormone treatment, or cytostatic drug treatment.

17
Extracellular vesicles
Exosomes, microvesicles and apoptotic bodies Extracellular vesicles (EVs) are nanosized particles released by virtually every cell in the body [42]. Cells have always communicated with each other by either cell-cell contact or by transferring molecules secreted by them. Initially, researchers believed the vesicles exocytosed by cells were just cell debris, or garbage, but over the last 40 years these vesicles have emerged as a very im-portant means of intercellular communication, the extracellular vesicles.
The vesicles that are shed from the cells originate either from the plasma membrane (PM) or they first fuse with the PM and are then exocytosed. The different particles have been given names such as microvesicles, microparti-cles, ectosomes, apoptotic bodies, shedding vesicles or exosomes [43, 44].
Exosomes are part of the extracellular vesicle family. They are formed in-side the mother cell, which could be any cell from prostate cells, to cardiac cells to immunological cells etc., thus creating subtypes of different exosomes [45-47]. Each EV carries proteins, DNA and RNA representative of the mother cell and EVs can exchange proteins, lipids and nucleic acids with other healthy and diseased cells, as well as promote metastasis in malignant cells [48, 49].
Exosome biogenesis starts in the endosomal pathway [50], see Figure 2. Late endosomes are formed by acidification and changes in protein content [51]. After the formation of late endosomes they start inward budding, creat-ing intraluminal vesicles (ILVs) and the late endosome forms into a mul-tivesicular body (MVB) [52]. The MVB then fuses with the PM to release the exosomes into the extracellular space. The fusion of the MVB to the PM is mostly driven by Rab GTPases, myosins and SNAREs [53]. The sorting of proteins and budding of ILVs into MVB and subsequently exosomes is not yet fully understood, but the endosomal sorting complex required for transport (ESCRT) is thought to play an active role in the budding [54]. Membrane mi-cro domains such as lipid rafts have been discovered to be a passive role of sorting [55].

18
Figure 2. The biogenesis of exosomes starts with the invagination of the plasma membrane at the location of a lipid raft area. This creates an early endosome, which starts to invaginate, creating intraluminal vesicles (ILVs) and a multivesicular body (MVB) is thus formed. The MVB migrates towards the plasma membrane (PM) and fuses with the PM to exocytose the exosomes into the extracellular space. Image from Waldenström, A. and Ronquist, G. [56].
The extracellular vesicle family sub-types are classified based on size, origin and biomarker proteins. Exosomes are normally classified as double mem-brane vesicles that contain the proteins ALIX, TSG101 and proteins from the tetraspanin family such as CD9, CD63, CD81 and CD82 [57]. They have a diameter of 30-100 nm, although the upper limit is debated and tends to in-crease [58].
Microvesicles (MVs), are vesicles released directly from the PM and are up to 1000 nm in diameter [48, 59]. The sorting of proteins into MVs and forming of MVs are still not very well defined, but the oligomerization of cy-toplasmic proteins as well as myristoylation and palmitoylation appears to form part of both [60].
Exosomes have a bilayered membrane, as does the cell, but exosome mem-branes are enriched with cholesterol and sphingomyelin. They do not have an abundance of phosphatidylcholine and phosphatidylethanolamine. The fatty acids in exosome membranes are saturated or monosaturated. Exosomes also have a high ratio of cholesterol and this makes their PM more rigid and they can withstand mechanical force. The cell PM has a ratio of cholesterol to phos-pholipids close to 1:1, but the exosome (prostasome) membrane has a ratio of 2:1 [61, 62].

19
Malignant cells also release exosomes into the tumor microenvironment and evidence has emerged that these can contribute to tumor progression and metastasis [63]. The exosomal protein profile reflects the cells content [64], thus making the exosome a potentially perfect biomarker for liquid biopsies, helping to diagnose many diseases [65, 66].

20
Prostasomes
Prostasomes were first reported in prostatic fluid in the 1970s by Ronquist and Hedström [67] and were later observed as extracellular organellar structures in vivo [68, 69], see Figures 3 and 4. Prostasomes are a sub class of exosomes which are derived and secreted from the acinar, epithelial cells of the prostate gland [70]. They are secreted by both benign and malignant cells, and in healthy males prostasomes can be found in both prostatic and seminal fluids, although in the case of cancer, they can also be found in increased amounts of blood because of the epithelial-to-mesenchymal transition that promotes leak-age into bodily fluids [71].
Figure 3. Scanning electron microscopy image of prostasomes. The bar size indi-cates 100 nm. Image taken by Kjell Hultenby, Karolinska Institute, Huddinge.
Prostasomes play a pivotal physiological role by interacting with sperm cells and enabling the transfer of molecules and proteins essential to fertilization [72, 73]. Prostasomes were first observed to fuse hydrophobically with sper-matozoa in 1990 by Ronquist et al. [72] and the interaction was later discov-ered to be pH dependent, where a more acidic pH is to prefer [74]. Sperm cells gain important abilities from prostasomes so they can survive the hostile en-vironment of the female genital tract in order to reach and penetrate the oocyte

21
for fertilization [72]. Prostasomes promote sperm motility, capacitation and an acrosome reaction on their way to the oocyte [75, 76]. The motility promo-tion effect may come from the transfer of calcium signaling proteins from the prostasome to the sperm [77]. Sperm cells become less active in acidic sur-roundings, but since seminal plasma and prostasomes are slightly alkaline, they appear to have a protective effect in the vaginal area [78], and sperm motility is crucial to natural fertility [79]. The capacitation of the sperm cell occurs when the glycoprotein coat is removed. The capacitation process must be complete before the acrosome reaction takes place [80]. The acrosome re-action permits sperm cells to fuse with the egg but it cannot happen prema-turely and it is believed that the cholesterol in the prostasome stabilizes the acrosomal cap [75].
Figure 4. Transmission electron microscope image of prostasomes, where the bi-layered membrane is clearly visible. The bar size indicates 200 nm. Image taken by Kjell Hultenby, Karolinska Institute, Huddinge.
Furthermore prostasomes show a protective ability against reactive oxygen species (ROS) and against bacteria [81, 82]. When prostasomes were incu-bated with Bacillus megaterium it was observed that the bacteria decreased or experienced a total withdrawal. Thus prostasomes are considered to be anti-bacterial agents [81]. When spermatozoa enters the female genital tract they are considered to be intruders and prostasomes act as immunosuppressants to protect the sperms [83].
Prostasomes make an important contribution to the normal fertilization pro-cess because of their antibiotic, antioxidant and immunosuppressive proper-

22
ties, as mentioned above. This leads us to believe that disruption to the pros-tasome function could cause infertility. Kravets et al. [84] suggested that me-dium containing prostasomes could be effective in assisted reproductive tech-nologies.
The prostasomal membrane has a unique composition with a 2:1 ratio of cholesterol to phospholipids [85], where sphingomyelin is the most abundant. The high cholesterol makes the prostasomal membrane stable and resistant to detergents [86]. The composition is similar to that of lipid rafts and it is be-lieved that the biogenesis of prostasomes (and exosomes) starts with these mi-cro domains [55, 87].
DNA has been found inside prostasomes [88, 89] and was proved to be transferred into sperm cells [90]. This could be important for the normal de-velopment of the pregnancy [91].
The membrane attack complex (MAC) inhibitory protein CD59 is localized on the prostasomal surface, attached via a glycosylphosphatidylinositol (GPI) anchor [92]. Because of this the spermatozoa are saved from the female com-plement system. CD59 from prostasomes can be transferred to blood cells lacking this protein and this results in preserved function [93]. Prostasomes have a high coagulant activity and this is due to the protein tissue factor (CD142), which is located on the surface of the prostasomal membrane [94].
Tissue factor is known to promote cancer growth [95] and because pros-tasomes have such high levels they can be active players in the proliferation of cancer cells. Both prostasomes and malignant cell-derived exosomes are capable of producing adenosine triphosphate (ATP) but prostasomes have a higher ATPase activity compared to malignant exosomes, thus creating a higher net value for ATP production [96], which is essential for an active up-take into recipient cells, and in turn, making them active messengers in the spread of cancer.

23
Lipid rafts
In 1988 van Meer and Simons [97] put forward the first hypothesis of lipid rafts, domains in the membrane enriched with glycosphingolipids, proteins and cholesterol, see Figure 5. Glycosphingolipid clusters were insoluble in non-ionic detergents at 4°C, thus the term detergent-resistant membranes (DRMs) was created [98]. These domains play an important role in cell endo-cytosis and cell exocytosis, as well as cell signaling. Lipid rafts and DRMs have been designated as platforms for cellular signaling [99, 100] because of the many proteins localized to these domains, as well as for signal transduction [101] and membrane trafficking [102].
There are primarily two types of lipid rafts: caveolae, with the primary bi-omarker caveolin [103] and non-caveolae (planar), which is recognized by the protein flotillin instead [104, 105]. GPI-anchored proteins (glycosylphospha-tidylinositol) have been shown to be associated with DRMs and primarily when it comes to signaling [106, 107].
Figure 5. Image of a planar lipid raft situated in the bilayered plasma membrane. Cholesterol and glycosphingolipids make the lipid raft more rigid compared to the surrounding membrane and hold transmembrane proteins and receptors. Image credit Wikipedia.

24
Plasma membranes are ordered structures where the DRMs are regions of high cholesterol content. The phospholipids in the lipid rafts fatty acid side chains have high saturation and this results in a denser packing together with the acyl chains of the sphingolipids. This leads to phase separation and a more rigid domain compared to the normal plasma membrane [108].
Exosomes could play an important role in biomarker studies for certain di-agnoses due to their isolation from bodily fluids and the fact that they can provide a more specific insight into the protein composition of the micro do-mains derived from the respective host cells [109].

25
Purinergic receptors
Purinergic receptors comprise of three classes: P1 receptors (adenosine), P2X receptors and P2Y receptors. Each class then has subtype receptors; A1-A3, P2X1-P2X7 and P2Y1-P2Y14 (see Figure 6) and together they are involved in many different biological processes [110]. Purinergic receptors are all membrane bound [111] and P1 is a G-protein coupled receptor, as well as P2Y. The P2X receptor belongs to the ligand gated ion-channel family.
Figure 6. A schematic diagram of how purinergic receptors are divided into three classes: P1 receptors, P2X receptors and P2Y receptors as well as further division into subtype receptors. Image adapted from the manuscript Occurrence of purinergic receptors in human prostasomes (prostate epithelial cell-derived exosomes) by Du-bois et.al., 2018.
There is an abundance of adenosine in seminal plasma [112] and previous work shows the production of ATP by human prostasomes [113]. Adenosine receptors are involved, for example, in cardiac rhythm, circulation [114, 115], the immune system [116], neurogenerative diseases [117] and angiogenesis [118]. Due to their wide range of biological functions, previous studies have pointed to P1 receptors as therapeutic targets [119, 120]. However, the only

26
drug on the market is an antagonist to the A2a receptor, Regadenoson (Lex-iscan; Astellas Pharma) [121]. Caffeine is also an antagonist to the adenosine receptor [122].
Among the most investigated receptors in the P2X subgroup is the P2X7 receptor [123]. Studies show that the P2X7 receptor is a mediator of invasive-ness and metastasis in PC [124, 125]. P2X7 has been shown to regulate the release of microvesicles [126-128] as it effects the composition of lipids and proteins in the PM. The receptor mediates trafficking in the PM as well as intracellular organelles [129].
The drug Clopidogrel, which reduces the risk of blood clots forming, is an antagonist to the receptor P2Y12 [130] and has antiplatelet activity. Thus it is often used in patients with cardiovascular diseases. The receptor P2Y12 is in-volved in platelet aggregation [131] and a way of reducing a tumor and stop-ping metastasis from forming is to inhibit platelet activity [132, 133]. This has been shown by Clopidogrel in both prostate and colon cancer [134].

27
Thymidine kinase 1
Thymidine kinase (TK) consists of two isoenzymes: TK1 (cytosolic) and TK2 (mitochondrial). Thymidine is salvaged from extracellular catabolic activity and TK converts salvaged thymidine from extracellular activities into its monophosphate counterpart (TMP) by the coupling of a phosphate group from ATP. After two additional phosphorylation steps, TMP is converted into de-oxythymidine triphosphate (dTTP), which is subsequently incorporated into DNA [135].
The TK1 enzyme activity is upregulated during cell proliferation, through the S-G2 phase of the cell cycle, see Figure 7. During the G1 phase the levels of TK1 activity are low and activity peaks during the S phase. The levels de-crease during the G2 phase and for the M phase TK1 can hardly be detected [136].
Figure 7. An image of the cell cycle where thymidine kinase expression is at its highest level during the S phase. Picture from Biovica International AB, Uppsala, Sweden.
Tumor aggressiveness have been shown to correlate with high levels of TK1 activity in serum. It has been suggested that this is indicative of the early events of carcinogenesis [137, 138]. Using both liquid and solid biopsies, TK1 activity can be detected [139]. TK1 has mostly been applied on hematological

28
malignancies as a tumor marker in routine health care and to a lesser extent applied to solid tumor diagnostic applications, such as PC [140, 141]. How-ever, if we combine prostasome detection with a TK1 assay, the specific signal from TK1 could be detected more easily and help in the diagnosis of prostate cancer.

29
Methods
Purification of prostasomes Seminal fluid was collected from the Reproduction center at Akademiska Hos-pital, Uppsala, Sweden. The pooled seminal fluid was then subjected to a cen-trifugation at 3000 g for 12 min. The supernatant was transferred to ultracen-trifugation tubes and spun at 10,000 g for 30 min at 4°C using a 90Ti rotor (Beckman Coulter, Brea, CA, USA).
The supernatant was transferred again and another ultracentrifugation took place, at 100,000 g for 2h at 4°C. The pellet was resuspended in phosphate buffered saline (PBS) over night at 2-8°C. The suspension was then loaded on to a XK16/70 Superdex 200 gel column (GE Healthcare, Uppsala, Sweden) and equilibrated with PBS in order to exclude amorphous material [45].
At a flow rate of 5 mL/h fractions were collected in volumes of 1.3 mL. Peaks with elevated absorbance at 260 nm (nucleic acid) and 280 nm (pro-teins) were measured and the fractions containing prostasomes were pooled and ultracentrifuged at 100,000 g for 2 h at 4°C. The pellet was resuspended in PBS overnight at 2-8°C before loaded on to a sucrose density gradient of 1M, 1.5M and 2M sucrose. The gradient was ultracentrifuged at 85,000 g for 21 h at 4°C using a SW28.1 rotor (Beckman Coulter), see Figure 8.
The fraction at 1.5M contains prostasomes and is considered the main frac-tion (density range 1.13-1.19 g/ml). This fraction was ultracentrifuged at 100,000 g for 2 h at 4°C and then solved in PBS. The solution was adjusted to 2 mg/mL in accordance with a BCA protein assay kit (Merck, Darmstadt, Ger-many) and stored at -70°C until use.

30
Figure 8. Schematic image of prostasome purification. Differential centrifugations followed by column chromatography and a sucrose gradient.
Purification of lipid rafts Prostasomes, purified as stated above, were pooled to contain approximately 8 mg and then top loaded on to a 1% Triton X-100 sucrose density gradient at 0.15 M, 0.8 M and 1.5 M. The gradient was ultracentrifuged at 256,000 g for 28 h at 4°C with a SW40Ti rotor (Beckman Coulter), see Figure 9.
The fraction at 0.8M (density of 1.10 g/mL) was collected and ultracentri-fuged at 100,000 g for 2 h at 4°C. The pellet was resuspended in PBS and stored at -70°C until use.
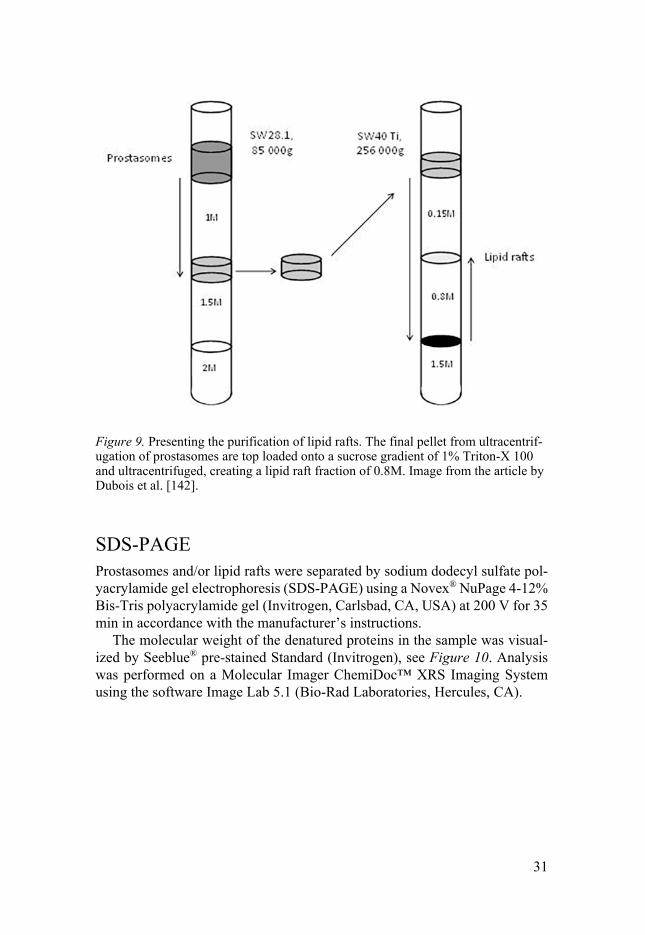
31
Figure 9. Presenting the purification of lipid rafts. The final pellet from ultracentrif-ugation of prostasomes are top loaded onto a sucrose gradient of 1% Triton-X 100 and ultracentrifuged, creating a lipid raft fraction of 0.8M. Image from the article by Dubois et al. [142].
SDS-PAGE Prostasomes and/or lipid rafts were separated by sodium dodecyl sulfate pol-yacrylamide gel electrophoresis (SDS-PAGE) using a Novex® NuPage 4-12% Bis-Tris polyacrylamide gel (Invitrogen, Carlsbad, CA, USA) at 200 V for 35 min in accordance with the manufacturer’s instructions.
The molecular weight of the denatured proteins in the sample was visual-ized by Seeblue® pre-stained Standard (Invitrogen), see Figure 10. Analysis was performed on a Molecular Imager ChemiDoc™ XRS Imaging System using the software Image Lab 5.1 (Bio-Rad Laboratories, Hercules, CA).

32
Figure 10. SDS-PAGE gel with standard (ladder) to the far left and the following three lanes depicting lipid rafts with varying protein concentration, that have been separated according to molecular weight.
Immunoblotting The proteins from the SDS-PAGE gel were subsequently transferred to a 0.2 µm Novex® Nitrocellulose membrane (Invitrogen) using 25 V for 1 h. Bovine serum albumin (BSA 1%) in PBS was used to block any unspecific binding.
Primary antibodies (1:1000) directed against receptors and proteins of in-terest were incubated with the nitrocellulose membrane at room temperature (RT) for 1 h and then washed in PBS. The secondary biotin conjugated anti-body (1:1000) was added and reacted with the membrane in RT for 1 h and washed in PBS. Streptavidin conjugated to alkaline phosphatase was added to the membrane and incubated at RT for 1 h. The binding of biotin and strep-tavidin is a very strong non-covalent interaction.
The nitrocellulose membrane was washed in PBS and the proteins were visualized using a BCIP/NBT kit (Invitrogen), see Figure 11, and analysis was then performed on a Molecular Imager ChemiDoc™ XRS Imaging System us-ing the software Image Lab 5.1 (Bio-Rad Laboratories, Hercules, CA, USA).

33
Figure 11. A primary antibody binds to its antigen, and a secondary biotinylated an-tibody binds to the primary antibody, making it possible to detect the protein using streptavidin-alkaline phosphatase.
Mass spectrometry Prostasomal lipid rafts (20 µg) were denatured and reduced in SDS (1%), ammonium bicarbonate (0.1 M) and dithiothreitol (DTT) (10 mM) by heat-ing for 5 min at 98 °C, and subsequent alkylation by iodoacetamide (20 mM) followed by incubation for 15–30 min at 20 °C in the dark. The sample was thereafter transferred to a spintube, 10 kDa cutoff (Pall Corporation, Port Washington, NY) [143].
Urea (8 M) and ammonium bicarbonate (50 mM) were added four times (4 x 450 µL) and centrifuged for 15 min at 14,000 g each time as previously described (40). Before the fifth centrifugation only 2 M urea and 50 mM am-monium bicarbonate were added and spun after which the solution was diluted to a final concentration of 1 M urea and 50 mM ammonium bicarbonate. The digestion was executed for 16 h at 20 °C by 0.5 µg trypsin, and stopped by acetic acid.
UV-absorbance was measured in a NanoDrop 2000 (Thermo Fischer Sci-entific Inc., Waltham, MA) to establish protein concentration. A C18 filter absorbed the digest for 24 h [144] before mass analysis was performed on a Thermo Velos (Thermo Fischer Scientific Inc.). The column was 100 mm long with an internal diameter of 75 µm, packed with 3 µm C18-Aq particles. The elution was carried out by a constructed gradient from 5–65% over 60 min starting with two solutions: 0.1% formic acid and 100% acetonitrile with 0.1% formic acid, respectively.
A maximum of 10 fragmentations were made for each MS-spectrum. Frag-mentation was carried out by CID using standard settings. The list of peaks obtained was used to search the X! Tandem website.

34
Electron microscopy
Scanning electron microscopy The prostasome pellet received after the final 100,000 g ultracentrifugation was fixated with PBS containing 2% glutaraldehyde. The fixated pellet was briefly rinsed in distilled water and dehydrated with 70% ethanol for 10 min, 95% ethanol for 10 min and 99.9% ethanol for 15 min.
All steps were conducted at 4 °C and then transferred to acetone. The pellet was dried in a critical point dryer (Balzer, CPD 010, Liechtenstein) using car-bon dioxide. After drying, the pellet was mounted on an aluminum stub and coated with carbon (Bal-Tec MED 010, Liechtenstein).
Analysis was performed with an Ultra 55 field emission scanning electron microscope (Zeiss, Oberkochen, Germany) at 5 kV, see Figure 3. Transmission electron microscopy The prostasome pellet received after the final 100,000 g ultracentrifugation was treated with 2% glutaraldehyde for fixation. The fixated pellet was washed once in PBS and once in PBS containing 2% osmium tetroxide at 4 °C for 2 h. The pellet was then transferred into ethanol for dehydration fol-lowed by acetone and subsequently embedded in LX-112 (Ladd, Burlington, VT, USA). The embedded pellet was then cut into sections of around 50-60 nm using a Leica Ultracut UCT (Leica, Wien, Austria).
The sections were then contrast colored using uranyl acetate and lead cit-rate, then analyzed in a Tecnai 12 Spirit Bio TWIN transmission electron mi-croscope (FEI Company, Eindhoven, The Netherlands) at 100 kV and a Veleta camera (Olympus Soft Imaging Solutions, Münster, Germany) was used for preparing the digital images, see Figure 4.
TK1 real-time assay Prostasomes and malignant cell derived exosome samples, respectively, were mixed 1:1 with lysis buffer containing a protease inhibitor (Complete Lysis ®, Roche GmbH), incubated at RT for 10 min and vortexed for 2 sec. Samples and TK1 activity calibrator samples (to create an activity standard curve) respectively were mixed with a reagent mix (Biovica International AB, Uppsala, Sweden) to a final sample concentration of 10 %.
Calibrator materials were from Diasorin S.r.1. Catalytic activity concentra-tion values are expressed as TK-REA Units per liter. One TK-REA Unit is 1.2x10-12 mole TMP converted per sec. A 20 µL reagent sample mix was transferred into 100 µL PCR tubes (Qiagen, Hilden, Germany) and loaded into

35
a Rotor-Gene Q (Qiagen) real-time PCR instrument. All samples were run in triplicate.
The incubation profile was 30 min at 37 °C followed by fluorescence cap-ture every minute for a total of 60 min at 37 °C. A runfile was created from the RotorGene Q Series (Qiagen) software. From this file, fluorescence pat-tern images of developing normalized fluorescence as well as final activity data, based on the assigned values for the calibrators, were subsequently downloaded as an editable Excel file for further analysis.

36
Perspectives on Paper I: Proteomic profiling of detergent resistant membranes (lipid rafts) of prostasomes
Introduction Prostasomes consist of a bilayered membrane similar to normal cells, but with a higher composition of sphingomyelin and cholesterol. Lipid rafts are do-mains within the bilayered cell membrane that are more densely packed and it has been proposed that they act as a signaling center for active receptors.
We asked whether prostasomes had these lipid rafts in their membrane and, if so, what kind of proteins and receptors they hold as a way of further char-acterizing the prostasome in the search for a specific biomarker.
Aim and procedure The aim of this paper was to characterize the proteins in the prostasomal mem-brane using mass spectrometry. By targeting proteins in the DRMs we hoped to identify proteins that could be good candidates for specific prostasome bi-omarkers, but also potential proteins responsible for interactions with the sur-rounding cells.
By treating the prostasomes with 1% Triton X-100 we were able to separate the lipid rafts, or DRMs, from the rest of the membrane using a density gradi-ent. The DRMs were then subjected to liquid chromatography-tandem mass spectrometry (MS).
Summary By examining the DRMs from prostasomes we can highlight some of the in-teresting proteins and biomarkers, many of which would be lost in the “back-ground noise” of all the other proteins if whole cells, or just whole pros-tasomes, were subjected to homogenization.
Almost 400 specific proteins were identified as being part of the prostaso-mal lipid raft, many of them involved in intraluminal vesicle formation. The proteins with the highest Mascot score were CD13, CD26 and CD10, which

37
have long been associated with prostasomes. Both CD26 and CD10 associate with tetraspanins (CD9), which are used as a marker for exosomes.
The general markers for exosomes CD81 and CD82 were not found in pros-tasomal DRMs and this could mean they are not associated with lipid rafts in the prostasomal bilayered membrane.

38
Perspectives on Paper II: Human erythrocyte-derived nanovesicles can readily be loaded with doxorubicin and act as anticancer agents
Introduction Lipid rafts, or DRMs, from prostasomes were found to re-vesiculate when subjected to osmotic pressure in a physiological buffer. From one vesicle we could engineer another, which could now be loaded with the desired cargo. Due to the immunologic response of the prostasomes if administered intrave-nously, we used erythrocytes in this method instead, as erythrocyte transfu-sions have been used for many years with few adverse events.
When the loaded vesicles were subjected to cells, they showed greater up-take into the cells, thus delivering the cargo (in this case, doxorubicin) into the target cells, and killing them.
Aim and procedure The aim of this paper was to investigate if we could use these engineered vesicles to deliver cytostatic drugs more efficiently compared to cytostatic drugs free in solution the way they are administered today, in the blood of cancer patients.
Erythrocytes were emptied of hemoglobin, turning them into erythrocyte ghosts which were subjected to Triton X-100 and ultracentrifugation. The re-vesiculated lipid rafts from the erythrocyte ghosts were loaded with the de-sired cargo (i.e. hemoglobin, gold colloidal particles and cytostatic drugs) by osmotic pressure and administered into the media of PC3 cells.

39
Summary This is a new approach for the on-point delivery of pharmaceuticals. Many cancer drugs have high pH values, making them weak bases. The microenvi-ronment is acidic and thus the weak bases are protonated and neutralized when entering cells.
By using a biologically engineered vesicle for delivery, this problem is cir-cumvented and, as shown in the outcome of this article, doxorubicin encapsu-lated in erythrocyte DRM vesicles can kill malignant cells more efficiently than when the doxorubicin is administered in the cell medium.

40
Perspectives on Paper III: Occurrence of purinergic receptors in human prostasomes (prostate epithelial cell-derived exosomes)
Introduction Purine and pyrimidine receptor subtypes were cloned and characterized during the 1990s. P1, the adenosine receptor, includes four subtypes. P2X, the ion channel, has seven subtypes and P2Y, the G-protein coupled receptor, has eight subtypes.
Purinergic receptors are involved in many biological functions in the body and the idea of nucleotides as signaling molecules had already been proposed as early as 1972 [145]. The rapid action of purinergic signaling can be seen in secretion, neurotransmission and neuromodulation. The long-term action can be seen in, for example, proliferation, differentiation and migration, as well as regeneration [146].
Aim and procedure In this project we investigated what kind of purinergic receptors are present in the prostasomal membrane.
Prostasomes were purified from human seminal plasma and then subjected to SDS-PAGE and immunoblotting using an array of antibodies directed against purinergic receptors.
Summary We found that almost all purinergic receptors are represented in pros-tasomes.
Adenosine has been used clinically since the mid-1900s and prostate tumor cell proliferation is inhibited by adenosine and the drug Regadenoson is an A2a receptor antagonist [121]. P2X7 has been suggested to be an early marker for prostate cancer [147] and the receptor effects both the lipid and the protein

41
content of the plasma membrane [129]. P2Y12 receptor inhibitors, such as Clopidogrel, are used by many patients as an antiplatelet therapy [148, 149] and the drug was found to be able to reduce the risk of metastasis in PC [150].

42
Perspectives on Paper IV: Increased levels of thymidine kinase 1 in malignant cell-derived exosomes
Introduction Thymidine kinase 1 has long been regarded as a marker of cancer. TK1 and TK2 are isoenzymes, where TK1 is found in the cytoplasm and is cell cycle dependent, whilst TK2 is cell cycle independent and is present in the mito-chondria [151]. TK1 phosphorylates nucleosides into nucleotides which are needed for DNA synthesis during cell division.
Weagel et al. [152] showed that in Burkitt’s lymphoma, acute promyelocy-tic leukemia (APL), T cell leukemia and acute lymphoblastic leukemia (ALL), TK1 is upregulated and localized to the plasma membrane.
Aim and procedure In this project, we compared non-malignant prostasomes with malignant exo-somes derived from different PC cell lines. We aimed to investigate the rela-tionship between TK1 and malignant exosomes from prostate cancer cell lines (PC3, LNCaP and Du145) and non-malignant prostasomes from healthy do-nors.
Prostasomes and exosomes were purified and subjected to the TK1 activity enzyme assay based on real-time determination of TK1 activity from Biovica International AB (Uppsala, Sweden). TK1 enzyme activity was measured over time tracing a fluorescence pattern.
Summary We observed a difference in TK1 enzyme activity between malignant exo-somes and prostasomes, where malignant exosomes showed higher enzyme activity. Thus, TK1 could prove useful for monitoring and prognosis of pros-tate cancer.

43
Conclusions and future perspectives
The work in this thesis has contributed to the pre-clinical research that is greatly needed to solve the prostate cancer puzzle.
• In Paper I we were able to purify lipid rafts from the prostasome
membrane and through a proteomic analysis highlight some of the more abundant proteins that gathered in these microdomains.
• In Paper II we allowed the purified lipid rafts from prostasomes to re-vesiculate and create bioengineered vesicles capable of loading agents of choice. We used this method on erythrocytes and loaded them with doxorubicin. Malignant cells were subjected to these vesicles and the result was a better uptake of doxorubicin compared to free drug.
• In Paper III we mapped the existence of purinergic receptors on prostasomes in order to understand how current and future drugs might affect patients with prostate cancer.
• In Paper IV we looked at benign human prostasomes and malignant cell-derived exosomes and how TK1 enzyme activity varies using a real-time TK1 assay developed by Biovica International AB.
Further studies could be directed toward looking more closely at (malignant) prostasomes from prostate cancer patients. I would also suggest pursuing the creation of specific antibodies for prostasomes as a continuance of Paper I. This could be used to develop an assay for capturing prostasomes from blood samples.
In further studies for Paper II we could let prostasomes donate receptors, as in the papers by Babiker et al. [93, 153], to erythrocytes to be able to target recipient cells more specifically. This would give the bioengineered vesicles and address tag to better find their way to the correct recipient cells as a way for a new drug treatment for prostate cancer.
For Paper III it would be interesting to verify what the effects of drugs targeted to purinergic receptors have on PC patients and other diseases. Also I would suggest to repeat the Western blot experiments, this time using pros-tate cancer cell-derived exosomes.
For Paper IV, as a minimum, the next project would be to look at more exosomes from other type of cells and the TK1 enzyme activity they hold.

44
TK1 is mostly measured in plasma, but is it free or is it bound to other exo-somes? By further investigating TK1 enzyme activity in exosomes a liquid biopsy assay could form, making it easier to monitor, or even diagnose pros-tate cancer.
In conclusion, this thesis provides new knowledge to the fight against pros-tate cancer and how to better diagnose and treat it more efficiently.

45
Populärvetenskaplig sammanfattning på svenska
I Sverige diagnosticeras varje år ca 10 000 män med prostatacancer (Social-styrelsen, 2016). Prostatacancer är den vanligaste cancerformen hos män och drabbar främst män över 50 år. Idag används prostataspecifikt antigen, PSA, som blodprovsbiomarkör för att upptäcka prostatacancer [154]. Halten av PSA kan vara förhöjd vid t.ex. prostatit och prostatahyperplasi. Detta gör att PSA inte är specifikt för just prostatacancer. Att använda sig av PSA-värden vid en storskalig screening skulle innebära att man får alldeles för många falskt positiva svar, men även falskt negativa svar kan förekomma.
Förutom PSA består seminalvätskan av en mängd olika proteiner som har specifika roller för spermiernas funktion. Vår forskargrupp var den första att visa att det sker en aktiv utsöndring av små vesikler omgärdade av ett dubbelt lipidmembran till prostatavätskan, så kallade prostasomer [70].
Alla kroppens celler kommunicerar med varandra på olika sätt. Ibland ge-nom cell-cell kontakt, och ibland genom att skicka ut molekyler eller intercel-lulära budbärare mellan varandra, som till exempel prostasomer (exosomer). Prostasomer är membranomgärdade extracellulära vesikler som utsöndras av prostatans acinära epitelceller [68, 69]. Prostasomerna bildas då den sena en-dosomen börjar invaginera och bilda små vesikler inuti sig själv. Då bildas en multivesikulär kropp, MVB, som kan fusera med cellens plasmamembran och då frigöra prostasomerna genom exocytos [70, 155]. De flesta prostasomer är i storleksordningen 30-200 nm i diameter. Ett fåtal kan vara så stora som upp till 500 nm, men medeldiametern är 150 nm [156].
Huvuduppgiften för prostasomer är att skydda spermierna på deras väg upp till ägget genom att tillföra proteiner som stöd vid migrationen [74, 157].
Prostasomernas membran har ett högt kolesterol/fosfolipid förhållande; 2:1 [85]. Membranets lipidsammansättning visar att sfingomyelin är det domine-rande fettslaget, till skillnad från cellmembran där fosfatidylkolin är den do-minerande fosfolipiden [62]. Dessutom bär prostasomerna med sig ett innehåll av bland annat cytosolproteiner, DNA och RNA [88].
Vi har kunnat visa att prostataceller som omvandlats till cancerceller be-håller förmågan att utsöndra prostasomer till omgivningen [158]. Funktionen hos prostasomen, att primärt skydda spermier, kommer då att flyttas till att gälla skyddande av prostatacancerceller, så tack vare egentillverkade prostasomer får cancercellen en överlevnadsfördel.

46
Den första hypotesen om lipidflottar (specialiserade membrandomäner, beri-kade med glykosfingolipider, proteiner och kolesterol) kom 1988 från Simons och van Meer [97]. Den vidareutvecklades sedan 1997 av Simons och Ikonen [159]. Lipidflottar blev definierade som subdomäner av plasmamembranet med låg densitet och som är resistenta mot oladdade detergenter vid låga tem-peraturer [160, 161]. Man har funnit att många cellsignaleringsvägar är asso-cierade med just lipidflottar eller detergentresistanta membran, DRM [162].
För att få fram lipidflottar används detergenten Triton X-100 i den lösning som man vid 4 °C låter provet (i detta fall prostasomer) centrifugera i vid väl-digt högt g-tal, ca 256 000 g, under 28 h mha en swing out rotor (SW40) [163]. Vi har funnit att lipidflottar finns i prostasomernas membran och vi har gjort proteomik på dessa i jakten efter nya intressanta biomarkörer.
Efter att ha tagit fram lipidflottar ur prostasomens membran såg vi att vi kunde skapa helt nya vesikler genom osmotiskt tryck. Dessa vesikler kunde vi då ladda med vald agent. Eftersom dessa nya vesikler gjorda av lipidflottar från prostasomer skulle kunna störa immunsystemet hos en potentiell patient beslöt vi oss för att använda metoden på röda blodkroppar (erytrocyter).
Vi laddade dessa med cytostatika och utsatte prostatacancerceller för dessa nya vesikler. Vi såg då att våra nya bio-vesikler upptogs av cellerna i större utsträckning än om man gav cytostatiska blandat i cellernas medium. Detta gör att våra nya bio-vesikler skulle kunna användas som terapi mot prosta-tacancer i framtiden.
Det finns tre typer av purinerga receptorer, P1, P2Y och P2X. P1 receptorn, som också kallas adenosinreceptor, delas in i fyra subtyper: A1, A2a, A2b, samt A3. Receptorn P2X har sju subtyper; P2X1-7 och denna receptor är en jonkanalreceptor, medan de två andra typerna (P1 och P2Y) är GPCR-kopp-lade receptorer. Slutligen har vi P2Y receptorn som består av åtta subtyper; P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13 och P2Y14 [164].
P2 receptorn uttrycks av alla tumörer. Genom att inhibera specifika subty-per av P2 receptorn kan man framkalla antingen apoptos/nekros eller reglering av celltillväxten. P1 receptorn är inblandad i inhibering av aktiverade immun-celler, vilket gör att cancercellerna kan växa och bli resistenta mot kemoterapi [165, 166]. Mikromiljön hos tumörer är hypoxisk vilket inducerar ackumule-ringen av adenosin pga den accelererande hydrolysen av ATP [167]. Under många år har man sett att adenosin varit inblandad i tumörassocierad im-munsuppression [168].
Vi har genom Western blot kunnat se att det finns flera subtyper av P1, P2Y och P2X receptorer på prostasomerna, vilket innebär en ökad förståelse hur prostasomer påverkas av olika läkemedel vid exempelvis terapi av prostata cancer.

47
Thymidinkinas 1 är en biomarkör för cancer [151]. Då en cell ska dela sig ökar enzymet TK1 för att kunna fosforylera thymidinmonofosfat (TMP) till först thymidindifosfat (TDP) och sedan till thymidintrifosfat (TTP), som då kan inkorporeras in i DNA [135].
Vi ville ta reda på om det finns en skillnad i thymidinkinas 1 aktivitet hos prostasomer från friska män jämfört med exosomer från maligna celler, och vi fann att det var en högre aktivitet hos de maligna exosomerna. Detta kan i förlängningen möjligtvis användas för att ställa diagnos vid misstänkt prosta-tacancer.

48
Acknowledgements
The work presented in this thesis was performed at the Department of Medical Sciences at Uppsala University, Uppsala, Sweden. Financial support was pro-vided by Lions Cancer Research Fund, AIMDay, Selanders Fund and SciL-ifeLab.
Jag skulle vilja börja med att tacka min huvudhandledare Anders Larsson för att du kämpade för min doktorandregistrering och för att jag har fått jobba så fritt. Tack för bra vetenskaplig input och din aldrig sinande optimism!
Tack till mina bihandledare Kim Kultima och Masood Kamali-Moghad-dam, för att ni tog er tid att hjälpa mig. Tack Masood för vetenskaplig väg-ledning och hjälp med problem på vägen.
Jag skulle även vilja tacka Karin Eriksson för att du varit min vän och lyssnat på mig, i både bra och dåliga dagar. Utan dig hade det varit mycket jobbigare.
Tack till Lena Kask och Åsa Thulin för mycket bra information om barn, barnafödande och ammande. Nu kommer nästa kapitel förhoppningsvis inte som en chock!
Tack Lena Moberg för hjälp med hur allt fungerar, från att beställa kaffe till metoder i labbet.
Tack Desireé Edén för att jag fått kräkas över problem med antagning. Det har varit skönt att ha en annan doktorand i korridoren.
Tack Marie Carlson för glada tillrop när jag träffat på dig i korridoren eller i fikarummet. Det har gjort mig väldigt glad.
Tack till alla på Klinisk kemi för goda fredagskakor och trevliga fikastun-der. Jag kommer sakna det!
Tack Liza Löf för att du gjort många av mina projekt roligare. Tack för din entusiasm och kompetens, men framför allt tack för att du blev min vän.
Tack Per Stålhandske för trevligt samarbete på projektet om TK1 och våra långa vetenskapliga diskussioner som gör att vi båda förstår mer och mer hela tiden.
Tack till mina vänner för att ni tycker att jag är bra, att ni lyssnat när jag behöver prata av mig, att ni ger råd när det varit tufft och kämpigt och att ni ger mig glädje när jag behöver tänka på annat.
Tack till min familj för att ni pushat och peppat under åren! Utan det stödet hade det inte gått.
Största tacket går till min man Erik Dubois. Tack för din kärlek. Med dig vid min sida är livet både enklare och roligare! Snart är vi tre ♥

49
References
1. Hanahan, D. and R.A. Weinberg, The hallmarks of cancer. Cell, 2000. 100(1): p. 57-70.
2. Witsch, E., M. Sela, and Y. Yarden, Roles for growth factors in cancer progres-sion. Physiology (Bethesda), 2010. 25(2): p. 85-101.
3. Sherr, C.J. and F. McCormick, The RB and p53 pathways in cancer. Cancer Cell, 2002. 2(2): p. 103-12.
4. Thiery, J.P., [Epithelial-mesenchymal transitions in cancer onset and progres-sion]. Bull Acad Natl Med, 2009. 193(9): p. 1969-78; discussion 1978-9.
5. Blasco, M.A., Telomeres and human disease: ageing, cancer and beyond. Nat Rev Genet, 2005. 6(8): p. 611-22.
6. Hanahan, D. and J. Folkman, Patterns and emerging mechanisms of the angio-genic switch during tumorigenesis. Cell, 1996. 86(3): p. 353-64.
7. Elmore, S., Apoptosis: a review of programmed cell death. Toxicol Pathol, 2007. 35(4): p. 495-516.
8. Hanahan, D. and R.A. Weinberg, Hallmarks of cancer: the next generation. Cell, 2011. 144(5): p. 646-74.
9. Warburg, O., On respiratory impairment in cancer cells. Science, 1956. 124(3215): p. 269-70.
10. Warburg, O., On the origin of cancer cells. Science, 1956. 123(3191): p. 309-14.
11. Vander Heiden, M.G., L.C. Cantley, and C.B. Thompson, Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science, 2009. 324(5930): p. 1029-33.
12. Jodar, M., et al., Semen proteomics and male infertility. J Proteomics, 2017. 162: p. 125-134.
13. Myers, R.P., Structure of the adult prostate from a clinician's standpoint. Clin Anat, 2000. 13(3): p. 214-5.
14. Berry, S.J., et al., The development of human benign prostatic hyperplasia with age. J Urol, 1984. 132(3): p. 474-9.
15. Owen, D.H. and D.F. Katz, A review of the physical and chemical properties of human semen and the formulation of a semen simulant. J Androl, 2005. 26(4): p. 459-69.
16. Mann, T., Experimental approach to the study of semen and male reproductive function. Int J Fertil, 1978. 23(2): p. 133-7.
17. Lilja, H. and C.B. Laurell, The predominant protein in human seminal coagu-late. Scand J Clin Lab Invest, 1985. 45(7): p. 635-41.
18. Malm, J., et al., Isolation and characterization of the major gel proteins in hu-man semen, semenogelin I and semenogelin II. Eur J Biochem, 1996. 238(1): p. 48-53.
19. Lilja, H., Structure and function of prostatic- and seminal vesicle-secreted pro-teins involved in the gelation and liquefaction of human semen. Scand J Clin Lab Invest Suppl, 1988. 191: p. 13-20.

50
20. Lee, C., et al., Demonstration of the role of prostate-specific antigen in semen liquefaction by two-dimensional electrophoresis. J Androl, 1989. 10(6): p. 432-8.
21. Szecsi, P.B. and H. Lilja, Gastricsin-mediated proteolytic degradation of human seminal fluid proteins at pH levels found in the human vagina. J Androl, 1993. 14(5): p. 351-8.
22. Wolters-Everhardt, E., et al., Buffering capacity of human semen. Fertil Steril, 1986. 46(1): p. 114-9.
23. Kavanagh, J.P., Sodium, potassium, calcium, magnesium, zinc, citrate and chlo-ride content of human prostatic and seminal fluid. J Reprod Fertil, 1985. 75(1): p. 35-41.
24. Fair, W.R. and R.F. Parrish, Antibacterial substances in prostatic fluid. Prog Clin Biol Res, 1981. 75A: p. 247-64.
25. Lindholmer, C., The importance of seminal plasma for human sperm motility. Biol Reprod, 1974. 10(5): p. 533-42.
26. Burden, H.P., et al., Prostasomes--their effects on human male reproduction and fertility. Hum Reprod Update, 2006. 12(3): p. 283-92.
27. Boyle, P., P. Maisonneuve, and P. Napalkov, Geographical and temporal pat-terns of incidence and mortality from prostate cancer. Urology, 1995. 46(3 Suppl A): p. 47-55.
28. Steinberg, G.D., et al., Family history and the risk of prostate cancer. Prostate, 1990. 17(4): p. 337-47.
29. Spitz, M.R., et al., Familial patterns of prostate cancer: a case-control analysis. J Urol, 1991. 146(5): p. 1305-7.
30. Lin, D.W., et al., Serum percent free prostate-specific antigen in metastatic pros-tate cancer. Urology, 1998. 52(3): p. 366-71.
31. Armitage, T.G., et al., The value of the measurement of serum prostate specific antigen in patients with benign prostatic hyperplasia and untreated prostate cancer. Br J Urol, 1988. 62(6): p. 584-9.
32. Gronberg, H., et al., Prostate cancer screening in men aged 50-69 years (STHLM3): a prospective population-based diagnostic study. Lancet Oncol, 2015. 16(16): p. 1667-76.
33. Klotz, L., et al., Nadir testosterone within first year of androgen-deprivation therapy (ADT) predicts for time to castration-resistant progression: a secondary analysis of the PR-7 trial of intermittent versus continuous ADT. J Clin Oncol, 2015. 33(10): p. 1151-6.
34. Iversen, P., Antiandrogen monotherapy: indications and results. Urology, 2002. 60(3 Suppl 1): p. 64-71.
35. Schroder, F., et al., Androgen deprivation therapy: past, present and future. BJU Int, 2012. 109 Suppl 6: p. 1-12.
36. Nguyen, P.L., et al., Adverse effects of androgen deprivation therapy and strat-egies to mitigate them. Eur Urol, 2015. 67(5): p. 825-36.
37. Tuck, S.P., et al., Prostate cancer and osteoporosis. Curr Osteoporos Rep, 2013. 11(1): p. 11-20.
38. Thorstenson, A., et al., Incidence of fractures causing hospitalisation in prostate cancer patients: results from the population-based PCBaSe Sweden. Eur J Can-cer, 2012. 48(11): p. 1672-81.
39. Katsogiannou, M., et al., The hallmarks of castration-resistant prostate cancers. Cancer Treat Rev, 2015. 41(7): p. 588-97.
40. Ceder, Y., et al., The Molecular Evolution of Castration-resistant Prostate Can-cer. Eur Urol Focus, 2016. 2(5): p. 506-513.

51
41. Van Poppel, H., et al., Radical prostatectomy for locally advanced prostate can-cer: results of a feasibility study (EORTC 30001). Eur J Cancer, 2006. 42(8): p. 1062-7.
42. Cocucci, E. and J. Meldolesi, Ectosomes and exosomes: shedding the confusion between extracellular vesicles. Trends Cell Biol, 2015. 25(6): p. 364-72.
43. Cocucci, E., G. Racchetti, and J. Meldolesi, Shedding microvesicles: artefacts no more. Trends Cell Biol, 2009. 19(2): p. 43-51.
44. Gyorgy, B., et al., Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cell Mol Life Sci, 2011. 68(16): p. 2667-88.
45. Stegmayr, B. and G. Ronquist, Promotive effect on human sperm progressive motility by prostasomes. Urol Res, 1982. 10(5): p. 253-7.
46. Raposo, G., et al., B lymphocytes secrete antigen-presenting vesicles. J Exp Med, 1996. 183(3): p. 1161-72.
47. van Niel, G., et al., Intestinal epithelial cells secrete exosome-like vesicles. Gas-troenterology, 2001. 121(2): p. 337-49.
48. Raposo, G. and W. Stoorvogel, Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol, 2013. 200(4): p. 373-83.
49. Colombo, M., G. Raposo, and C. Thery, Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol, 2014. 30: p. 255-89.
50. Keller, S., et al., Exosomes: from biogenesis and secretion to biological function. Immunol Lett, 2006. 107(2): p. 102-8.
51. Stoorvogel, W., et al., Late endosomes derive from early endosomes by matura-tion. Cell, 1991. 65(3): p. 417-27.
52. Novikoff, A.B., E. Essner, and N. Quintana, Golgi Apparatus and Lysosomes. Fed Proc, 1964. 23: p. 1010-22.
53. Ostrowski, M., et al., Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol, 2010. 12(1): p. 19-30; sup pp 1-13.
54. Fisher, R.D., et al., Structure and ubiquitin binding of the ubiquitin-interacting motif. J Biol Chem, 2003. 278(31): p. 28976-84.
55. de Gassart, A., et al., Lipid raft-associated protein sorting in exosomes. Blood, 2003. 102(13): p. 4336-44.
56. Waldenstrom, A. and G. Ronquist, Role of exosomes in myocardial remodeling. Circ Res, 2014. 114(2): p. 315-24.
57. Escola, J.M., et al., Selective enrichment of tetraspan proteins on the internal vesicles of multivesicular endosomes and on exosomes secreted by human B-lymphocytes. J Biol Chem, 1998. 273(32): p. 20121-7.
58. van der Pol, E., et al., Classification, functions, and clinical relevance of extra-cellular vesicles. Pharmacol Rev, 2012. 64(3): p. 676-705.
59. Minciacchi, V.R., M.R. Freeman, and D. Di Vizio, Extracellular vesicles in can-cer: exosomes, microvesicles and the emerging role of large oncosomes. Semin Cell Dev Biol, 2015. 40: p. 41-51.
60. Shen, B., et al., Protein targeting to exosomes/microvesicles by plasma mem-brane anchors. J Biol Chem, 2011. 286(16): p. 14383-95.
61. Carlsson, L., et al., Characteristics of human prostasomes isolated from three different sources. Prostate, 2003. 54(4): p. 322-30.
62. Arienti, G., et al., Fatty acid pattern of human prostasome lipid. Arch Biochem Biophys, 1998. 358(2): p. 391-5.
63. Rak, J., Microparticles in cancer. Semin Thromb Hemost, 2010. 36(8): p. 888-906.
64. Johnstone, R.M., Revisiting the road to the discovery of exosomes. Blood Cells Mol Dis, 2005. 34(3): p. 214-9.

52
65. Tavoosidana, G., et al., Multiple recognition assay reveals prostasomes as prom-ising plasma biomarkers for prostate cancer. Proc Natl Acad Sci U S A, 2011. 108(21): p. 8809-14.
66. Minciacchi, V.R., et al., Extracellular vesicles for liquid biopsy in prostate can-cer: where are we and where are we headed? Prostate Cancer Prostatic Dis, 2017. 20(3): p. 251-258.
67. Ronquist, G. and M. Hedstrom, Restoration of detergent-inactivated adenosine triphosphatase activity of human prostatic fluid with concanavalin A. Biochim Biophys Acta, 1977. 483(2): p. 483-6.
68. Ronquist, G., et al., An Mg2+ and Ca2+-stimulated adenosine triphosphatase in human prostatic fluid: part I. Andrologia, 1978. 10(4): p. 261-72.
69. Ronquist, G., et al., An Mg2+ and Ca2+-stimulated adenosine triphosphatase in human prostatic fluid--part II. Andrologia, 1978. 10(6): p. 427-33.
70. Ronquist, G. and I. Brody, The prostasome: its secretion and function in man. Biochim Biophys Acta, 1985. 822(2): p. 203-18.
71. Pang, A., et al., Carcinosarcomas and Related Cancers: Tumors Caught in the Act of Epithelial-Mesenchymal Transition. J Clin Oncol, 2017: p. JCO2017749523.
72. Ronquist, G., B.O. Nilsson, and S. Hjerten, Interaction between prostasomes and spermatozoa from human semen. Arch Androl, 1990. 24(2): p. 147-57.
73. Carlini, E., et al., Fusion of sperm with prostasomes: effects on membrane fluid-ity. Arch Biochem Biophys, 1997. 343(1): p. 6-12.
74. Arienti, G., E. Carlini, and C.A. Palmerini, Fusion of human sperm to pros-tasomes at acidic pH. J Membr Biol, 1997. 155(1): p. 89-94.
75. Arienti, G., et al., Role of human prostasomes in the activation of spermatozoa. J Cell Mol Med, 2004. 8(1): p. 77-84.
76. Aalberts, M., T.A. Stout, and W. Stoorvogel, Prostasomes: extracellular vesi-cles from the prostate. Reproduction, 2014. 147(1): p. R1-14.
77. Fabiani, R., et al., Prolongation and improvement of prostasome promotive ef-fect on sperm forward motility. Eur J Obstet Gynecol Reprod Biol, 1995. 58(2): p. 191-8.
78. Arienti, G., et al., The motility of human spermatozoa as influenced by pros-tasomes at various pH levels. Biol Cell, 1999. 91(1): p. 51-4.
79. Saez, F., G. Frenette, and R. Sullivan, Epididymosomes and prostasomes: their roles in posttesticular maturation of the sperm cells. J Androl, 2003. 24(2): p. 149-54.
80. Breitbart, H., Signaling pathways in sperm capacitation and acrosome reaction. Cell Mol Biol (Noisy-le-grand), 2003. 49(3): p. 321-7.
81. Carlsson, L., et al., Antibacterial activity of human prostasomes. Prostate, 2000. 44(4): p. 279-86.
82. Skibinski, G., et al., Immunosuppression by human seminal plasma--extracellu-lar organelles (prostasomes) modulate activity of phagocytic cells. Am J Reprod Immunol, 1992. 28(2): p. 97-103.
83. Kelly, R.W., Seminal plasma immunosuppressive activity: the achilles heel of reproduction. Int J Androl, 1991. 14(4): p. 243-7.
84. Kravets, F.G., et al., Prostasomes: current concepts. Prostate, 2000. 43(3): p. 169-74.
85. Arvidson, G., et al., Human prostasome membranes exhibit very high choles-terol/phospholipid ratios yielding high molecular ordering. Biochim Biophys Acta, 1989. 984(2): p. 167-73.

53
86. Fabiani, R. and G. Ronquist, Association of some hydrolytic enzymes with the prostasome membrane and their differential responses to detergent and PIPLC treatment. Prostate, 1995. 27(2): p. 95-101.
87. Harder, T. and K. Simons, Caveolae, DIGs, and the dynamics of sphingolipid-cholesterol microdomains. Curr Opin Cell Biol, 1997. 9(4): p. 534-42.
88. Ronquist, K.G., et al., Human prostasomes contain chromosomal DNA. Prostate, 2009. 69(7): p. 737-43.
89. Olsson, I. and G. Ronquist, Nucleic acid association to human prostasomes. Arch Androl, 1990. 24(1): p. 1-10.
90. Ronquist, G.K., et al., Prostasomal DNA characterization and transfer into hu-man sperm. Mol Reprod Dev, 2011. 78(7): p. 467-76.
91. Carrell, D.T., et al., Sperm DNA fragmentation is increased in couples with un-explained recurrent pregnancy loss. Arch Androl, 2003. 49(1): p. 49-55.
92. Rooney, I.A., et al., Physiologic relevance of the membrane attack complex in-hibitory protein CD59 in human seminal plasma: CD59 is present on extracel-lular organelles (prostasomes), binds cell membranes, and inhibits complement-mediated lysis. J Exp Med, 1993. 177(5): p. 1409-20.
93. Babiker, A.A., et al., Transfer of prostasomal CD59 to CD59-deficient red blood cells results in protection against complement-mediated hemolysis. Am J Reprod Immunol, 2002. 47(3): p. 183-92.
94. Fernandez, J.A., et al., Potent blood coagulant activity of human semen due to prostasome-bound tissue factor. Biol Reprod, 1997. 56(3): p. 757-63.
95. Bromberg, M.E., et al., Tissue factor promotes melanoma metastasis by a path-way independent of blood coagulation. Proc Natl Acad Sci U S A, 1995. 92(18): p. 8205-9.
96. Ronquist, K.G., et al., Energy-requiring uptake of prostasomes and PC3 cell-derived exosomes into non-malignant and malignant cells. J Extracell Vesicles, 2016. 5: p. 29877.
97. van Meer, G. and K. Simons, Lipid polarity and sorting in epithelial cells. J Cell Biochem, 1988. 36(1): p. 51-8.
98. Helenius, A. and K. Simons, Solubilization of membranes by detergents. Bio-chim Biophys Acta, 1975. 415(1): p. 29-79.
99. Hancock, J.F., Lipid rafts: contentious only from simplistic standpoints. Nat Rev Mol Cell Biol, 2006. 7(6): p. 456-62.
100. Simons, K. and J. Gruenberg, Jamming the endosomal system: lipid rafts and lysosomal storage diseases. Trends Cell Biol, 2000. 10(11): p. 459-62.
101. Dykstra, M., et al., Location is everything: lipid rafts and immune cell signaling. Annu Rev Immunol, 2003. 21: p. 457-81.
102. Kirkham, M. and R.G. Parton, Clathrin-independent endocytosis: new insights into caveolae and non-caveolar lipid raft carriers. Biochim Biophys Acta, 2005. 1746(3): p. 349-63.
103. Rothberg, K.G., et al., Caveolin, a protein component of caveolae membrane coats. Cell, 1992. 68(4): p. 673-82.
104. Fernow, I., A. Icking, and R. Tikkanen, Reggie-1 and reggie-2 localize in non-caveolar rafts in epithelial cells: cellular localization is not dependent on the expression of caveolin proteins. Eur J Cell Biol, 2007. 86(6): p. 345-52.
105. Stuermer, C.A., et al., Glycosylphosphatidyl inositol-anchored proteins and fyn kinase assemble in noncaveolar plasma membrane microdomains defined by reggie-1 and -2. Mol Biol Cell, 2001. 12(10): p. 3031-45.
106. Zisch, A.H., et al., The glypiated neuronal cell adhesion molecule contactin/F11 complexes with src-family protein tyrosine kinase Fyn. Mol Cell Neurosci, 1995. 6(3): p. 263-79.

54
107. Romagnoli, P. and C. Bron, Phosphatidylinositol-based glycolipid-anchored proteins enhance proximal TCR signaling events. J Immunol, 1997. 158(12): p. 5757-64.
108. Brown, D.A. and E. London, Structure and function of sphingolipid- and cho-lesterol-rich membrane rafts. J Biol Chem, 2000. 275(23): p. 17221-4.
109. Simpson, R.J., et al., Exosomes: proteomic insights and diagnostic potential. Expert Rev Proteomics, 2009. 6(3): p. 267-83.
110. Ralevic, V. and G. Burnstock, Receptors for purines and pyrimidines. Pharmacol Rev, 1998. 50(3): p. 413-92.
111. North, R.A., Molecular physiology of P2X receptors. Physiol Rev, 2002. 82(4): p. 1013-67.
112. Fabiani, R. and G. Ronquist, Abundance of guanine, guanosine, inosine and adenosine in human seminal plasma. Int J Clin Lab Res, 1995. 25(1): p. 47-51.
113. Ronquist, K.G., et al., Human prostasomes express glycolytic enzymes with ca-pacity for ATP production. Am J Physiol Endocrinol Metab, 2013. 304(6): p. E576-82.
114. Eltzschig, H.K., M.V. Sitkovsky, and S.C. Robson, Purinergic signaling during inflammation. N Engl J Med, 2012. 367(24): p. 2322-33.
115. Eltzschig, H.K., Adenosine: an old drug newly discovered. Anesthesiology, 2009. 111(4): p. 904-15.
116. Rosenberger, P., et al., Hypoxia-inducible factor-dependent induction of netrin-1 dampens inflammation caused by hypoxia. Nat Immunol, 2009. 10(2): p. 195-202.
117. Fredholm, B.B., Adenosine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ, 2007. 14(7): p. 1315-23.
118. Liu, X.L., et al., Genetic inactivation of the adenosine A2A receptor attenuates pathologic but not developmental angiogenesis in the mouse retina. Invest Oph-thalmol Vis Sci, 2010. 51(12): p. 6625-32.
119. Jacobson, K.A. and Z.G. Gao, Adenosine receptors as therapeutic targets. Nat Rev Drug Discov, 2006. 5(3): p. 247-64.
120. Fredholm, B.B., Adenosine receptors as drug targets. Exp Cell Res, 2010. 316(8): p. 1284-8.
121. Chen, J.F., H.K. Eltzschig, and B.B. Fredholm, Adenosine receptors as drug tar-gets--what are the challenges? Nat Rev Drug Discov, 2013. 12(4): p. 265-86.
122. El Yacoubi, M., et al., The stimulant effects of caffeine on locomotor behaviour in mice are mediated through its blockade of adenosine A(2A) receptors. Br J Pharmacol, 2000. 129(7): p. 1465-73.
123. North, R.A., P2X receptors. Philos Trans R Soc Lond B Biol Sci, 2016. 371(1700).
124. Qiu, Y., et al., P2X7 mediates ATP-driven invasiveness in prostate cancer cells. PLoS One, 2014. 9(12): p. e114371.
125. Kunzli, B.M., et al., Impact of CD39 and purinergic signalling on the growth and metastasis of colorectal cancer. Purinergic Signal, 2011. 7(2): p. 231-41.
126. VanWijk, M.J., et al., Microparticles in cardiovascular diseases. Cardiovasc Res, 2003. 59(2): p. 277-87.
127. Hess, C., et al., Ectosomes released by human neutrophils are specialized func-tional units. J Immunol, 1999. 163(8): p. 4564-73.
128. Heijnen, H.F., et al., Activated platelets release two types of membrane vesicles: microvesicles by surface shedding and exosomes derived from exocytosis of mul-tivesicular bodies and alpha-granules. Blood, 1999. 94(11): p. 3791-9.

55
129. Qu, Y. and G.R. Dubyak, P2X7 receptors regulate multiple types of membrane trafficking responses and non-classical secretion pathways. Purinergic Signal, 2009. 5(2): p. 163-73.
130. Kunapuli, S.P., et al., ADP receptors--targets for developing antithrombotic agents. Curr Pharm Des, 2003. 9(28): p. 2303-16.
131. Hechler, B., M. Cattaneo, and C. Gachet, The P2 receptors in platelet function. Semin Thromb Hemost, 2005. 31(2): p. 150-61.
132. Battinelli, E.M., et al., Anticoagulation inhibits tumor cell-mediated release of platelet angiogenic proteins and diminishes platelet angiogenic response. Blood, 2014. 123(1): p. 101-12.
133. Dredge, K., et al., PG545, a dual heparanase and angiogenesis inhibitor, in-duces potent anti-tumour and anti-metastatic efficacy in preclinical models. Br J Cancer, 2011. 104(4): p. 635-42.
134. Symeonidis, D., et al., Impact of antiplatelet treatment on colorectal cancer staging characteristics. World J Gastrointest Endosc, 2012. 4(9): p. 409-13.
135. Munch-Petersen, B., Enzymatic regulation of cytosolic thymidine kinase 1 and mitochondrial thymidine kinase 2: a mini review. Nucleosides Nucleotides Nu-cleic Acids, 2010. 29(4-6): p. 363-9.
136. Sherley, J.L. and T.J. Kelly, Regulation of human thymidine kinase during the cell cycle. J Biol Chem, 1988. 263(17): p. 8350-8.
137. Alegre, M.M., et al., Serum detection of thymidine kinase 1 as a means of early detection of lung cancer. Anticancer Res, 2014. 34(5): p. 2145-51.
138. Gronowitz, J.S., et al., The use of serum deoxythymidine kinase as a prognostic marker, and in the monitoring of patients with non-Hodgkin's lymphoma. Br J Cancer, 1983. 47(4): p. 487-95.
139. Jagarlamudi, K.K. and M. Shaw, Thymidine kinase 1 as a tumor biomarker: technical advances offer new potential to an old biomarker. Biomark Med, 2018.
140. Zhou, J., E. He, and S. Skog, The proliferation marker thymidine kinase 1 in clinical use. Mol Clin Oncol, 2013. 1(1): p. 18-28.
141. Jagarlamudi, K.K., L.O. Hansson, and S. Eriksson, Breast and prostate cancer patients differ significantly in their serum Thymidine kinase 1 (TK1) specific ac-tivities compared with those hematological malignancies and blood donors: im-plications of using serum TK1 as a biomarker. BMC Cancer, 2015. 15: p. 66.
142. Dubois, L., et al., Proteomic Profiling of Detergent Resistant Membranes (Lipid Rafts) of Prostasomes. Mol Cell Proteomics, 2015. 14(11): p. 3015-22.
143. Wisniewski, J.R., et al., Universal sample preparation method for proteome analysis. Nat Methods, 2009. 6(5): p. 359-62.
144. Rappsilber, J., M. Mann, and Y. Ishihama, Protocol for micro-purification, en-richment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat Protoc, 2007. 2(8): p. 1896-906.
145. Burnstock, G., Purinergic nerves. Pharmacol Rev, 1972. 24(3): p. 509-81. 146. Burnstock, G., Short- and long-term (trophic) purinergic signalling. Philos
Trans R Soc Lond B Biol Sci, 2016. 371(1700). 147. Slater, M., et al., Early prostate cancer detected using expression of non-func-
tional cytolytic P2X7 receptors. Histopathology, 2004. 44(3): p. 206-15. 148. Huber, K., et al., Antiplatelet and anticoagulation agents in acute coronary syn-
dromes: what is the current status and what does the future hold? Am Heart J, 2014. 168(5): p. 611-21.
149. Yeung, J. and M. Holinstat, Newer agents in antiplatelet therapy: a review. J Blood Med, 2012. 3: p. 33-42.

56
150. Choe, K.S., et al., The use of anticoagulants improves biochemical control of localized prostate cancer treated with radiotherapy. Cancer, 2010. 116(7): p. 1820-6.
151. Aufderklamm, S., et al., Thymidine kinase and cancer monitoring. Cancer Lett, 2012. 316(1): p. 6-10.
152. Weagel, E.G., et al., Biomarker analysis and clinical relevance of TK1 on the cell membrane of Burkitt's lymphoma and acute lymphoblastic leukemia. Onco Targets Ther, 2017. 10: p. 4355-4367.
153. Babiker, A.A., et al., Transfer of functional prostasomal CD59 of metastatic prostatic cancer cell origin protects cells against complement attack. Prostate, 2005. 62(2): p. 105-14.
154. Balk, S.P., Y.J. Ko, and G.J. Bubley, Biology of prostate-specific antigen. J Clin Oncol, 2003. 21(2): p. 383-91.
155. Sahlen, G.E., et al., Ultrastructure of the secretion of prostasomes from benign and malignant epithelial cells in the prostate. Prostate, 2002. 53(3): p. 192-9.
156. Ronquist, G., Prostasomes are mediators of intercellular communication: from basic research to clinical implications. J Intern Med, 2012. 271(4): p. 400-13.
157. Pons-Rejraji, H., et al., Prostasomes: inhibitors of capacitation and modulators of cellular signalling in human sperm. Int J Androl, 2011. 34(6 Pt 1): p. 568-80.
158. Sahlen, G., et al., Prostasomes are secreted from poorly differentiated cells of prostate cancer metastases. Prostate, 2004. 61(3): p. 291-7.
159. Simons, K. and E. Ikonen, Functional rafts in cell membranes. Nature, 1997. 387(6633): p. 569-72.
160. Pike, L.J., Lipid rafts: bringing order to chaos. J Lipid Res, 2003. 44(4): p. 655-67.
161. Brown, D.A. and J.K. Rose, Sorting of GPI-anchored proteins to glycolipid-en-riched membrane subdomains during transport to the apical cell surface. Cell, 1992. 68(3): p. 533-44.
162. Anderson, R.G., The caveolae membrane system. Annu Rev Biochem, 1998. 67: p. 199-225.
163. Chamberlain, L.H., Detergents as tools for the purification and classification of lipid rafts. FEBS Lett, 2004. 559(1-3): p. 1-5.
164. Burnstock, G., Purine and pyrimidine receptors. Cell Mol Life Sci, 2007. 64(12): p. 1471-83.
165. Lukashev, D., A. Ohta, and M. Sitkovsky, Hypoxia-dependent anti-inflamma-tory pathways in protection of cancerous tissues. Cancer Metastasis Rev, 2007. 26(2): p. 273-9.
166. Stagg, J., et al., Anti-CD73 antibody therapy inhibits breast tumor growth and metastasis. Proc Natl Acad Sci U S A, 2010. 107(4): p. 1547-52.
167. Blay, J., T.D. White, and D.W. Hoskin, The extracellular fluid of solid carcino-mas contains immunosuppressive concentrations of adenosine. Cancer Res, 1997. 57(13): p. 2602-5.
168. MacKenzie, W.M., D.W. Hoskin, and J. Blay, Adenosine inhibits the adhesion of anti-CD3-activated killer lymphocytes to adenocarcinoma cells through an A3 receptor. Cancer Res, 1994. 54(13): p. 3521-6.


Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 1528
Editor: The Dean of the Faculty of Medicine
A doctoral dissertation from the Faculty of Medicine, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofMedicine. (Prior to January, 2005, the series was publishedunder the title “Comprehensive Summaries of UppsalaDissertations from the Faculty of Medicine”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-369166
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2019