Embed Size (px)
Citation preview

Plant Science 44 (1986) 89--96 89 Elsevier Scientific Publishers Ireland Ltd.
PROPERTIES OF GLUTAMINE SYNTHETASE OF BACTEROIDS FROM ROOT NODULES OF GL YCINE M A X
BASANT BHANDARI* and D.J.D. NICHOLAS
Department of Agricultural Biochemistry, Waite Agricultural Research Institute, University of Adelaide, Glen Osmond, S.A. 5064 (Australia)
(Received July 31st, 1985) (Revision received February 3rd, 1986) (Accepted February 17th, 1986)
Glutamine synthetase (GS) (EC 6.3.1.2) Isozyme I has been purified from bacteroids of root nodules of Glycine max. The purified GSI which had a molecular weight of 760 000 contained 12 subunits each of about 63 000 daltons. The GS had Kin-values of 3.3 mM for hydroxylamine and 11.1 mM for L-glutamine for the ~-glutamyl transferase reaction. The Kin-values for the biosynthetic reaction were 12.9 and 14.3 mM for L- glutamine and ATP, respectively. The Kin-values of 2.8 and 8.9 mM were determined for NH4C1 from a biphasic plot. The inhibition of 7-glutamyl transferase activity by L-glutamate and NH 4 Cl was competitive for L-glutamine and non-competitive for hydroxylamine. When hydroxylamine and L-glutamine were the substrates, NH, C! resulted in Ki-values of 18 and 10 mM, respectively. Triphosphate nucleotides viz. ITP, UTP, CTP and ATP (48--92%) and diphosphate nucleotides (20--85%) viz. ADP, IDP, UDP, CDP and ODP, restricted either trans- ferase or biosynthetic activities. Alanine, serine and glycine, respectively, inhibited GS activity by 78, 63 and 50%. Different pairs of amino acids in various combinations resulted in a cumulative inhibition of the enzyme.
Key words: adenylylation~ ammonium assimilation; bacteriods; Glycine max; glutamine synthetase
Introduction
GS (L-glutamate ammonia ligase, EC 6.3.1.2) occupies a key position in the meta- bolism of ammonia by bacteria [1--3], algae [4,5] higher plants [6] and animals [7,8]. Brown and Dilworth [9] have shown that the glutamine synthetase/glutamate synthase or the glutamate dehydrogenase pathways are important for the assimilation of ammonia depending on the type of nitrogen supply in the culture medium for growth of rhizobia strains. However, some workers do not
*Present address: Department of Medicine, Meta- bolism Section, Veterans Administration Medical Center, 10701 East Boulevard, Cleveland, OH 44106, U.S.A. Abbreviations: ~-GH, 7-glutamylhydroxamate; GOGAT, glutamate synthase; GS, glutamine synthetase; SDS-PAGE, sodium dodecyl sulfate- polyacrylamide gel electrophoresis.
consider that glutamate dehydrogenase is a key enzyme in ammonia assimilation in bacteria and higher plants [6,10].
Upchurch and Elkan [11] have shown that ammonia is transported from bacteroids to the nodule cytosol where it is assimilated via the glutamine synthetase/glutamate synthase pathway [12]. Based on 15N studies, we have confirmed that this pathway is present in rhizobia grown in culture solutions as well as in bacteroids of root nodules of G. max [13]. Glutamine synthetase activity is regulated by ammonia in Klebsiella pneumoniae by an adenylylation/deadenylylation mechanism [14]. The enzyme is also regulated similarly in rhizobia grown in culture solutions con- taining either L-glutamate [ 15] or ammonium [16,17] as a nitrogen source.
In free-living rhizobia, glutamine synthetase is present in two forms: GSI and GSII [17-- 19]. Glutamine synthetase has been purified
0168-9452/86/$03.50 © Elsevier Scientific Publishers Ireland Ltd. Printed and Published in Ireland

90
and characterized from strains of R. japonicu m grown in culture solutions [20] and in cytosol fractions of root nodules of soybean [21] but no information is available for glutamine synthetase from bacteroids. Two forms of GS have been purified from the plant fraction of Phaseolus vulgaris root nodules and one of these forms appears identical to the form of the enzyme found in roots but the other is probably associated with the nodule [22] . The assimilation of ammonia produced by N2 fixation in the bacteroids is primarily via the GS/glutamate synthase (GOGAT) pathway in the nodule cytosol [9 ,21,22] . We have recently purified glutamine synthetase (GSI) of bacteroids from root nodules of G. max and some of the kinetic properties were deter- mined [ 23 ]. The aim of the present s tudy was to consider the regulation of the enzyme in bacteroids from root nodules of G. max. Thus, some of the properties including the effects of Mg 2÷ and various mono-, di- and tri- phosphate nucleotides, organic acids on enzyme activity have been investigated.
Materials and methods
Chemicals Various amino acids, nucleotides, ~/-gluta-
mylhydroxamate (~/-GH) and bovine serum albumin were purchased from Sigma Chemical Co., St. Louis, MO, U.S.A. and Blue Sepharose CL-6B, thyroglobulin, ferritin0 catalase and aldolase were obtained from Pharmacia Fine Chemicals, Uppsala, Sweden. All the other chemicals used were of the best analytical grade available.
Bacteria, growth conditions and preparation o f bacteroids
R. japonicum CC705 was kindly supplied by Dr. F.J. Bergersen, CSIRO Division of Plant Industry, Canberra. This was maintained on yeast extract mannitol slopes at 30°C [24]. The liquid cultures were grown aerobically with 10 mM L-glutamate (11 in 2-1 Erlen- meyer flasks} in shake cultures as described previously [23] . Seeds of G. max (L.) Merr.
(cv. Clark 63) were inoculated with R. japoni- cum CC705 [23] and bacteroids were prepared as described previously [ 13 ].
Purification of GS
A 104-fold purified GS was prepared from bacteroids with a 37% recovery [23]. Washed bacteroids disrupted with an ultrasonic probe [20] , were then centrifuged for 20 min at 1000 × g at 4°C in a RC-2B refrigerated centrifuge (Sorvall, Norwalk, CO, U.S.A.). The supernatant fraction was centrifuged for 10 min at 40 000 X g, the resultant supernatant fraction $40 was heated at 55°C for 15 min, and after cooling it was centrifuged at 40 000 X g for 10 min. The supernatant fraction was loaded onto a Blue Sepharose CL-6B column (1.3 X 10 cm). The absorbed GS was then eluted with 3 mM ATP in 50 mM Tris--HC1 buffer containing 1 mM MnC12 (pH 7.0). The enzyme was desalted by passing through a Sephadex G-10 column (13 X 10 cm) pre- equilibrated with 50 mM Tris--HC1 buffer (pH 7.0} [23] . The purified GS which had a transferase activity of 44 gmol ~-GH produced (min)- ' (mg protein)- ' and biosynthetic activity of 6 tlmol phosphate (min) -1 (rag protein}-' at 37°C was used in the present study.
Enzyme assays and protein determination GS activity was determined by the biosyn-
thetic reaction measuring the release of phosphate and by the 7-glutamyltransferase assay using the methods of Shapiro and Stadtman [25] . For the biosynthetic system the assay mixture in a final volume of 0.3 ml contained in mM: imadazole--HC1 buffer (pH 7.0), 40; L-glutamate, 40; MgC12, 8, NH4C1, 40; ATP, 12 and enzyme protein, 50 tLg. L-Glutamate was omitted from the control tubes. The results in Table I are based on normal enzyme assays with saturating sub- strates.
For the transferase assay the reaction mixture in a final volume of 1 ml contained in mM: imidazole--HC1 buffer (pH 7.5),

Table I. Effects of Mg 2+ on GS activity and its adenylylation in bacteroids from G. max. Assay of ~- glutamyl transferase activity and determination of adenylylation as described in Materials and methods, except that Mg 2+ concentrations were varied as shown. Enzyme activity is ~mol ~-GH produced (min) -1 (mg protein) -1 at 37°C.
Mg ~÷ 0 5 10 15 (mM) 30 60
Enzyme 48 42 38 31 9 5 activity
% adenyly- -- 12 20 35 81 90 lation
40; L-glutamine, 30; hydroxylamine hydro- chloride (neutralized with NaOH), 30; MnC12, 0.3; sodium arsenate, 2.0; ADP, 0.4 and an appropriate amount of the enzyme. Control tubes without L-glutamine were included.
For each batch of purified GS, the amount of enzyme for the biosynthetic and 7-gluta- myltransferase activities was determined so that under conditions of the assay, the activity was linear over a period of 45 rain. Protein was determined by the method of Sedmak and Grossberg [26], using bovine serum albumin as a standard protein.
Determination of the adenylylation state of GS
To determine the extent of adenylylation, GS was assayed with and without MgCl2 • 6H20 {0--60 mM). The values obtained were then used in the following formula proposed by Shapiro and Stadtman [25] :
n = 1 2 - 12 0.3 mM Mn 2÷ + 60 mM Mg 2÷
0.3 mM Mn 2+
Percent adenylylation as determined by divi- ding n by 12, then multiplying by 100.
Polyacrylamide gel electrophoresis Dialysed preparations of the purified en-
zyme (25 ~g protein) were run in polyacryl- amide gel electrophoresis [27] , using 12.5
91
mM Tris buffer containing 96 mM glycine (pH 8.3). Gels were stained with Coomassie brilliant blue. There were one band of protein associated with the enzyme. GS activity was detected in the gels, washed once in cold 50 mM Tris--HC1 (pH 7.2) and then incubated at 37°C in the presence of the transferase assay mixture. The activity band for the enzyme was detected by adding FeC13 reagent (0.4 g FeCl3, 0.24 g trichloroacetic acid and 0.25 ml, 12 N--HC1). The gels were photo- graphed immediately. In another procedure the gels were cut into 2 mm slices after electro- phoresis. Each slice was then individually checked for transferase activity. When this purified enzyme was separated by electro- phoresis in SDS polyacrylamide gels [28] then only one subunit was detected.
Molecular weight and subunits of GS The molecular weight of the purified GS
from bacteroids was determined by gel filtration on a Sepharose 6B column (1.6 × 99 cm) equilibrated with 50 mM Tris-HC1 buffer (pH 7.5) according to the method of Andrews [29]. The column was calibrated with thyroglobulin (669 000), ferritin (440 000), catalase (232 000) and aldolase (158 000) as markers. The molecular weight of the enzyme subunit was determined by sodium dodecyl sulfate-polyacrylamide gel electro- phoresis (SDS-PAGE) [28] as described pre- viously [20]. The following molecular mark- ers were used in the SDS gels: phosphorl- lase b (94 000); albumin (67 000); carbonic anhydrase (30 000); trypsin inhibitor (20 000) and a-lactalbumin (14 000}.
Results
Molecular weight of the purified glutamine synthetase and its subunits
The 104-fold purified enzyme prepared from bacteroids as described in Materials and methods had a molecular weight of approx. 760 0 0 0 as determined by gel filtration in a Sepharose 6B column followed by a discon- tinuous, non-denaturing PAGE electrophoresis
92
in 7% (w/v) polyacrylamide gel tubes. The stacking gel was 3% (w/v) polyacrylamide in 125 mM Tris--HC1 buffer (pH 6.8) and the running gel 7% (w/v) polyacrylamide in 375 mM Tris--HC1 buffer (pH 8.8).
Effects o f concentration o[ su bstrates The GS activity determined by transferase
assay had a specific requirement for Mn 2*. Little activity was observed when either of Mn ~*, L-glutamine, hydroxylamine, ADP or arsentate was omit ted from the assay mixture. The biosynthetic reaction was dependent on Mg 2÷, L-glutamate, NH4C1 as well as on ATP for activity (data not shown).
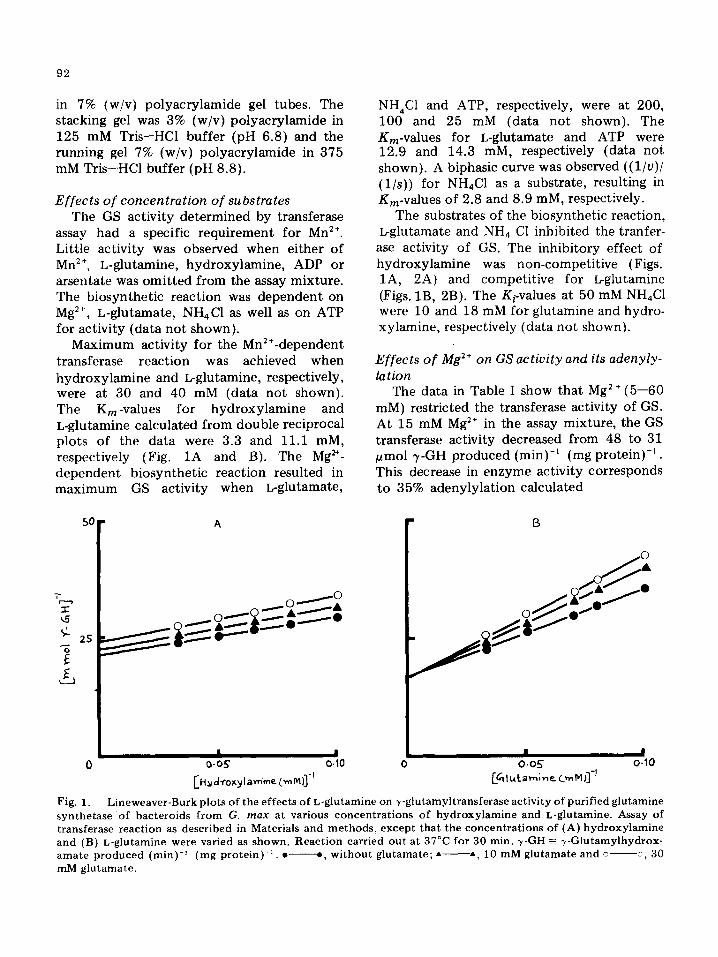
Maximum activity for the Mn:÷-dependent transferase reaction was achieved when hydroxylamine and L~glutamine, respectively, were at 30 and 40 mM (data not shown). The Kin-values for hydroxylamine and L-glutamine calculated from double reciprocal plots of the data were 3.3 and 11.1 raM, respectively (Fig. 1A and B). The Mg ~*- dependent biosynthetic reaction resulted in maximum GS activity when L-glutamate,
NH4C1 and ATP, respectively, were at 200, 100 and 25 mM (data not shown). The Kin-values for L-glutamate and ATP were 12.9 and 14.3 mM, respectively (data not shown). A biphasic curve was observed ((1/v)/ ( l /s)) for NH4C1 as a substrate, resulting in Kin-values of 2.8 and 8.9 mM, respectively.
The substrates of the biosynthetic reaction, L-glutamate and NH4 C1 inhibited the tranfer-
ase activity of GS. The inhibitory effect of hydroxylamine was non-competitive (Figs. 1A, 2A) and competitive for I~glutamine {Figs. 1B, 2B). The Ki-values at 50 mM NH,C1 were 10 and 18 mM for glutamine and hydro- xylamine, respectively (data not shown).
Effects o f Mg 2÷ on GS activity and its adenyly- lation
The data in Table I show that Mg 2÷ (5--60 mM) restricted the transferase activity of GS. At 15 mM Mg 2÷ in the assay mixture, the GS transferase activity decreased from 48 to 31 umol 7-GH produced (min) -1 (mg protein) -1 . This decrease in enzyme activity corresponds to 35% adenylylation calculated
50 A
+ A
0 0 . 0 5 0,10 0 0 . 0 ~ 0-10
[-H~ =l+'0x~l am'me (~M)] "1 ~ rut ami~e. (.m M)] -J
Pig. t . Lineweaver-Burk plots of the effects of L-gtutamine on +-glutamyltransferase act iv i ty o£ pur i f ied glutamine synthetase of bacteroids Prom G. m a x at various concentrat ions of hydroxy lamine and L-g]uta_mine. Assay of transferase reaction as described in Materials and methods, except that the concentrations of (A) hydroxy lamine and (B) L-glutamine were varied as shown. Reaction carried out at 37°C for 30 rain. +y-GH -= 7-G|utarnylhydrox- amate produced (rain) -~ (rag p r o t e i n ) -~ . • e, w i thou t glutamate; • • , 10 mM glutamate and c +, 30 mM glutamate.

93
Table II. Effec ts o f nucleot ides of GS activi ty o f bac tero ids f rom G. max. Assay o f 7-glutamyl t ransferase and were used at 2.5 mM final concen t ra t ion . The results are expressed as % inhib i t ion of cont ro l (wi thou t nucleot ides) . were used at 2.5 mM final concen t ra t ion . The results are expressed as % inhib i t ion of con t ro l (w i thou t nucleot ide) . The transferase activity of the cont ro l , 44 umol ~-GH p ro d u ced (min) -~ (rag p ro te in ) -z at 37°C. The b iosyn the t ic act ivi ty of the con t ro l was 6 umol phospha t e p roduced (min) -~ (rag pro te in) -~ at 37°C.
% inhibi t ion
Nucleot ide ITP UTP CTP ATP ADP IDP UDP GDP CDP IMP UMP GMP
(2.5 mM)
Transferase 90 92 75 92 - - 25 32 20 50 8 4 0 activity
Biosynthe t ic 48 50 62 - - 85 30 15 12 45 15 20 10 activity
in Materials and methods. Higher concentra- tions of Mg 2÷ (60 mM) resulted in 90% adenylylation form of GS (Table I).
Effect of various nucleotides and organic acids The transferase or biosynthetic reaction for
GS assay was inhibited by triphosphate and diphosphate nucleotides. ITP, UTP, CTP and ATP restricted GS activity by 48--92%, whereas diphosphate nucleotides, viz. IDP, UDP, GDP and CDP inhibited GS activity by
20--50% and ADP markedly reduced biosyn- thetic activity (85%). However, monophos- phate nucleotides, viz. IMP, UMP and GMP had little or no effect on GS activity (Table II). At 1.25 mM both CDP and ATP resulted in a decrease in transferase reaction by 25 and 70%, respectively, and at 2.5 mM these effects were 50 and 90%. Succinate and acetate at 2.5 mM restricted the transferase and bio- synthetic activities by only 10--20% and pyruvate, malate and oxaloacetate each at 2.5 mM were without effect {data not shown).
~}0 [ A I~ 0/0/0
I ~ o ~°'~°''~°~° ¢ 4o~
~ ~ ' ~ " ~ qj,......., o,_--- o--- • ~ °
0 ~.o5 O.~0 0 0.0S 0.10
Fig. 2. Lineweaver-Burk plots of the effects of NH4C! on 7-g]utamy]-transferase activity of purified g|utamin~ synthetase of hacteroids from G. max at various concentrations of hydroxylamine and L-g|utamine. Enzym~ assay as described in Materials and methods, except that the concentrations of (A) hydroxy|amine and (B) L g |u tamine were varied as shown. Reac t ion carried out at 37°C for 30 rain. 7-OH ~- -~-glutamylhydroxamat{ p roduced (rain) -~ (rag p ro te in ) -1 . • • , w i t h o u t NH4C1; a A 10 mM NH4C1 ; a n d - - ~ , 30 mM NH4C1

94
Table III. Effects of various combinations of amino acids on GS activity of bacteroids from G. max. Assay of -t-glutamyl transferase activity as described in Materials and methods, except that amino acids were used at 2.5 mM final concentration. Cumulative inhb bition was calculated by the method of Woolfolk and Stadtman [34]. The results are expressed as % inhibi- tion of control (without amino acids). The transferase activity of the control was 44 t~mol ~-GH produced (min)- ~ (mg protein)- l at 37°C.
Amino acid % inhibition (2.5 mM)
Observed Cumulative Additive
Alanine 78 -- -- Serine 63 -- -- Glycine 51 -- -- Arginine 10 -- -- Aspartate 0 -- -- Alanine + serine 83 92 141 Alanine + glycine 83 89 128 Serine + glycine 65 82 113
E f f e c t s o f amino acids The GS ac t iv i ty was m a r k e d l y inh ib i t ed by
e i ther 2.5 mM alanine, serine and glycine by 78, 76 and 50%, respec t ive ly (Table I I I ) , and to a lesser e x t e n t o r n o t a t all by arginine and aspar ta te . The c o m b i n e d inh ib i to ry e f fec t s o f var ious a m i n o acids were lower t han the to ta l addi t ive inh ib i t ions resul t ing f r o m an indivi- dual a m i n o acid. The cumula t ive inh ib i to ry e f fec t s o f a m i n o acids ca lcu la ted b y the m e t h o d of Wool fo lk and S t a d t m a n [34] were close to the obse rved inh ib i t ion of the trans- ferase assay {Table I I I ) .
Discussion
GS t o g e t h e r wi th g l u t a m a t e syn thase are key e n z y m e s for the ass imi la t ion of a m m o n i a in to organic n i t rogen c o m p o u n d s in micro- o rgan isms [ 1 , 3 0 ] , including rh i zob ia [ 1 9 ] . Based on 15N studies , we have f o u n d tha t the GS-g lu t ama te syn thase p a t h w a y is i m p o r t a n t in rh izob ia g rown in cul tures , as well as in bac te ro ids f r o m r o o t nodules of G. max [ 1 3 ] . The resul ts p r e sen t ed herein indica te tha t pur i f i ed GSI of bac te ro ids f r o m G. m a x has
similar physica l and b iochemica l p rope r t i e s to those r epo r t ed fo r pur i f ied GS f r o m cells of R. j a p o n i c u m grown in cu l tu re so lu t ion [20] bu t d i f fe ren t f r o m those r e p o r t e d for pur i f ied GS f r o m cy toso l f r ac t ion of r o o t nodules f r o m G. max [ 21 ].
An es t ima ted mo lecu l a r weight of 760 000 for GS of bac te ro ids f r o m G. max is c o m p a r - able wi th 720 000 r e p o r t e d for R. j apon icum grown in cu l ture so lu t ions [ 2 0 ] , bu t is m u c h higher than t ha t for GS of the cy toso l f rac t ion of r o o t nodules f r o m G. max (376 000) [21] or for ra t l iver GS [352 000] [25 ] . The GS o f bac te ro ids f r o m G. max con ta in 12 subuni ts each with a mo lecu l a r weight o f a round 63 000, being similar (60 000) to pur i f ied GS f r o m R. j apon icum grown in cu l tu re solut ions [ 2 0 ] . These results are c o m p a r a b l e wi th a subun i t value of 59 000 da l tons for GSI f rom R. j a p o n i c u m 6 1 A 7 6 [ 1 9 ] . On the o the r hand , the GS o f cy toso l f rac t ion f rom G. max con- tains 8 subuni t s wi th a subun i t mo lecu la r weight o f a b o u t 47 300 [21 ] .
The r e q u i r e m e n t for Mn 2÷ and arsenate indica te t ha t GS o f bac te ro ids f r o m G. m a x ca ta lyses a 7 -g lu tamyl t ransferase t y p e reac t ion as fo r GS f r o m R. j a p o n i c u m strains grown in cu l tu re so lu t ions [20] r a the r than by e i ther a m i n o h y d r o l a s e [31] or g lu taminase [32 ] . The Kin-values for h y d r o x y l a m i n e (3.3 mM) and L-g lu tamine (11.1 mM) were higher than those for GS f r o m R. j a p o n i c u m strains grown in cu l tu re solut ions , viz. h y d r o x y l a m i n e 1.2 mM, L-glu tamine , 7.7 mM for t ransferase reac t ion . For the b iosyn the t i c react ion, K m- values r e p o r t e d herein for g l u t a m a t e are m u c h higher for L-g lu tamate (12 .9 mM) and for NH4C1 (2.8 and 8.9 mM) than those r e p o r t e d fo r pur i f ied G8 f r o m N i t r o s o m o n a s europaea (1.4 and 5.4 mM) (2) and are also d i f fe ren t f r o m those for Chlorob ium v ibr io forme f. t h iosu l fa toph i lum (2.2 and 0.2 mM) [3 ] . The ef fec ts o f the subs t ra tes of the b iosyn the t i c r eac t ion (g lu tamate , a m m o n i a ) on t ransferase ac t iv i ty wi th L-g lu tamine are compe t i t i ve , whereas wi th h y d r o x y l a m i n e t hey are non- compe t i t i ve . These p rope r t i e s are similar to those r e p o r t e d fo r o the r bac te r ia [2 ,3] and

supports the postulated model for the mecha- nism of interaction of glutamine by GS [33].
The data reported here show that, as for GS from strains of R. japonicum grown in cultures [2] and other sources [24,31] the GS of bacteroids from G. max was also sensitive to various feedback inhibitors, viz. amino acids, nucleotides and organic acids. Thus the transferase activity of GS was markedly inhibited (> 50%) by alanine, serine and glycine. The inhibition of GS activity by alanine, glycine and serine was cumulative, as with the GS of R. japonicum strains grown in culture solutions [20] and of Escherichia coli [34,35]. These results contrast with non- cumulative effects of these compounds for rat liver GS [7].
Our results show that the GS of bacteroids from G. max was markedly inhibited (48-- 92%) by triphosphate nucleotides (ITP, UTP, CTP and ATP), to a lesser extent (20--50%) by diphosphate nucleotides (IDP, UDP, GDP and CDP) except for ADP where the bio- synthetic activity was reduced by 85% and with little or no effect by monophosphate nucleotides {IMP, UMP and GMP). These results are similar to those reported for GS from R. japonicum strains grown in culture solutions except that the extent of inhibition of transferase activity by triphosphate nucleo- tides was 40--68% [20]. The GS activity of the cytosol fraction of G. max measured by biosynthetic reaction was restricted by ADP (67%} and CTP (43%) [21].
The regulation of GS in microorganisms involves covalent alteration of the enzyme by adenylylation-deadenylylation and uridylyl- ation-deuridylylation [36--38]. The results presented herein show that GS of bacteroids from G. max is highly adenylylated {90%). Adenylylat ion results in an inactive form of the GS due to at tachment of AMP [25,36]. ~4C-AMP-labelled GS (GSI) from strains of R. japonicum grown in culture solutions was deadenylylated after t reatment with snake venom phosphodiesterase [13]. Our studies also show that snake venom phosphodiesterase t reatment deadenylylated the GS in extracts
95
of bacteroids but not the enzyme of the cytosol fraction of the nodules of G. max [13]. The GSII from strains of R. japonicum grown in culture solutions is not regulated by adenylylation [ 18].
It is shown in the present work that GSI of bacteroids from root nodules of G. max has similar properties to those of the enzyme from R. japonicum grown in culture solutions [20]. Thus, bacteroid GSI is similar to bacterial GS of R. japonicum in a free-living state but is different from the GS in the cytosol fraction obtained from plant sources [21]. GS activities have been determined in both the bacterial and cytosol fractions of root nodules from G. max [13], and these data demonstrate that there was a greater propor- tion of enzyme activity in the cytosol fraction than in bacteroids. Nevertheless, bacteroids contain GS which produces glutamine [39]. The GS of bacteroids from G. max is regulated by a number of amino acids, nucleotides, organic acids, as well as by an adenylylation- deadenylylation mechanism.
Acknowledgements
This work was supported by a grant to D.J.D.N. from the Australian Research Grants Scheme. B.B. held a Charles Everard post- doctoral fellowship of the University of Adelaide.
References
1 H. Nagantani, M. Shimizu and R.C. Valentine, Arch. Microbiol., 79 (1971) 164.
2 B. Bhandari and D.J.D. Nicholas, Aust. J. Biol. Sci., 34 (1981) 527.
3 S. Khanna and D.J.D. Nicholas, Arch. Microbiol., 134 (1983) 98.
4 S.K. Sawhney and D.J.D. Nicholas, Planta, 139 (1978a) 289.
5 S.K. Sawhney and D.J.D. Nicholas, Biochim. Biophys. Acta, 472 (1978b) 485.
6 B.J. Miflin and P.J. Lea, Annu. Rev. Plant. Physiol., 28 (1977) 299.
7 S.S. Tate and A. Meister, Proc. Natl. Acad. Sci. U.S.A., 68 (1971) 781.
8 S.S. Tate and A. Meister, Glutamine synthetase of mammalian liver and brain, in: S. Prusiner and

96
E.R. Stadtman (Eds.), The enzymes of Glutamine Metabolism, Academic Press, New York, London 1973, pp. 77--127.
9 C.M. Brown and M.J. Dilworth, J. Gen. Microbiol., 86 (1975) 39.
10 D.B. Scott, Ammonia assimilation in N~-fixing systems, in: J. I~obereiner, R.H. Burris and A. Hollaender (Eds.), Limitations and Potentials for Biological Nitrogen Fixation in the Tropics, Plenum Press, New York, 1978, pp. 223--236.
11 R.G. Upchurch and G.H. Elkan, J. Gen Microbiol., 104 (1978) 210.
12 T. Ohyama and K. Kumazawa, Soil Sci. Plant Nutr. (Tokyo) 26 {1980) 205.
13 F. Vairinhos, B. Bhandari and D.J.D. Nicholas, Planta, 159 {1983) 207.
14 S.L. Streicher, K.T. Shanmugam, F. Ausubel, C Morandi and R.B. Goldberg, J. Bacteriol., 120 (1974) 815.
15 P.E. Bishop, J.G. Guevara, J.A. Engelke and H.J. Evans, Plant Physiol., 57 {1976) 542.
16 R.A. Ludwig and E.R. Signer, Nature (London) 267 (1977) 245.
17 R.A. Ludwig, J. Bacteriol., 141 {1980) 1209. 18 R.A. Darrow and R.R. Knotts, Biochem. Biophys.
Res. Commun., 78 {1977) 554. 19 R.A. Darrow, Role of glutamine synthetase in
nitrogen fixation, in: J. Mora and R. Palacios (Eds.), Glutamine Metabolism Enzymology and Regulation, Academic Press, N.Y., London, 1980, pp. 139--166.
20 B. Bhandari, F. Vairinhos and D.J.D. Nicholas, Arch. Microbiol., 36 (1983) 84.
21 R.H. McParland, J.G. Guevara, R.R. Becker and H.J. Evans, Biochem. J., 153 {1976) 597.
22 J.V. Cullimore, M. Lara, P.J. Lea and B.J. Miflin, Planta, 157 (1983) 245.
23 B. Bhandari and D.J.D. Nicholas, FEMS Micro- biol. Letts., 25 (1984) 129.
24 H. Dalton, The cultivation of diazotropic micro- organisms, in: F.J. Bergersen (Ed.), Methods for Evaluating Biological Nitrogen Fixation, Wiley, Chichester, 1980, pp. 13--64.
25 B.M. Shapiro and E.R. Stadtman, Glutamine synthetase (Escherichia coli), in: S.P. Colowick and N.O. Kaplan (Eds.), Methods in Enzymology, Academic Press, New York, London, 1970, Vol. XVIIA, pp. 910--922.
26 J.J. Sedmak and S.E. Grossberg, Anal. Biochem., 79 (1977) 544.
27 U.K. Laemmli, Nature (London) 227 (1970) 680.
28 K. Weber, J.R. Pringie and M. Osborn, Measure- ment of molecular weights by electrophoresis on SDS acrylamide gel, in: S.P. Colowick and N.O. Kaplan (Eds.), Methods in Enzymology, Academic Press, New York, London, 1972, Vol. XXVI, pp. 3--27.
29 P. Andrews, Methods Biochem. Anal., 18 (1970) 1.
30 B. Ely, A.B.C. Amarasinghe and R.A. Bender, J. Bacteriol., 133 (1978) 225.
31 J.S. Hubbard and E.R. Stadtman, J. Bacteriol., 94 (1967) 1016.
32 A. Meister, Glutamine synthetase of mammals, in: P.D. Boyer (Ed.), The Enzymes, 3rd edn, Academic Press, New York, London, 1974, Vol. 10, pp. 669--754.
33 J.D. Gass and A. Meister, Biochemistry, 9 (1970) 1380.
34 C.A. Woolfolk and E.R. Stadtman, Arch. Biochem. Biophys., 118 (1967) 736.
35 C.A. Woolfolk, B.M. Shapiro and E.R. Stadtman, Arch Biochem. Biophys., 116 (1966) 177.
36 B.M. Shapiro, H.S. Kingdon and E.R. Stadtman, Proc. Natl. Acad. Sci. U.S.A., 58 {1967) 642.
37 R.A. Bender, K.A. Janseen, A.D. Resnick, M. Blumenberg, F. Foot and B. Magasanik, J. Bac- teriol., 129 {1977) 1001.
38 P.B. Chock, S.G. Rhee and E.R. Stadtman, Annu. Rev. Biochem., 49 (1980) 813.
39 E.R. Stadtman and A. Ginsburg (1974) Glutamine synthetase in Escherichia coli: structure and control, in: P.D. Boyer (Ed.), The Enzymes, 3rd edn., Academic Press, New York, London, 1974, Vol. 10, pp. 755--807.





























