Embed Size (px)
Citation preview

Assimilation of ammonia
glutamine synthetase (GS)
Glutamine synthetase of Salmonella thyphymurium (a bacterium closely related to E. coli)

Ciclo del nitrógeno
fig 22.1 Lehninger

fig 22.2 Lehninger

fig 22.3 Lehninger
All nitrogenases have an iron- and sulfur-containing cofactor that includes heterometal atom in the active site (e.g. FeMoCo). In most, this heterometal is molybdenum, though in some species it is replaced by vanadium or iron.

Lehninger Figure 22-04a

Reaction in assimilation of ammonia and major fates of the nitrogen atoms

Glucose (outside)
Glucose-6-P (inside)
Fructose-6-P
Fructose-1,6-diP
PEP
Pyruvate
acetyl~CoA
Glyceraldehyde-3-P
Glycerate-1,3-diP
Glycerate-3-P
Glycerate-2-P
DHAP
acetyl~P
acetate
EI
Hpr
IIAglc
IIB-IIC
glucose(outside)
glucose-6-P(inside)
EI~P
Hpr~P
IIAglc~P
IIB-IIC~P
Oxaloacetate
Malate
Succinate
Citrate
Isocitrate
2-ketoglutarate
Succinyl~CoA
Glyoxylate
Fumarate Glutamate
Glutamine
NH3
NH3acetyl~CoA
aspartate
CO2
CO2
CO2CO2
Inducer Exclusion
Activate AC
Fructose
Fructose-1-P
-
-
-
-
-
+++
+
+
ODH
PDH
GDHGOGAT
GS
ICD
Ammonia assimilation is tied tothe flux of carbon through centralmetabolic pathways
The 2-ketoglutarate/glutamine ratiois a signal of the cellular nitrogen status

Some of the reactions involving glutamine
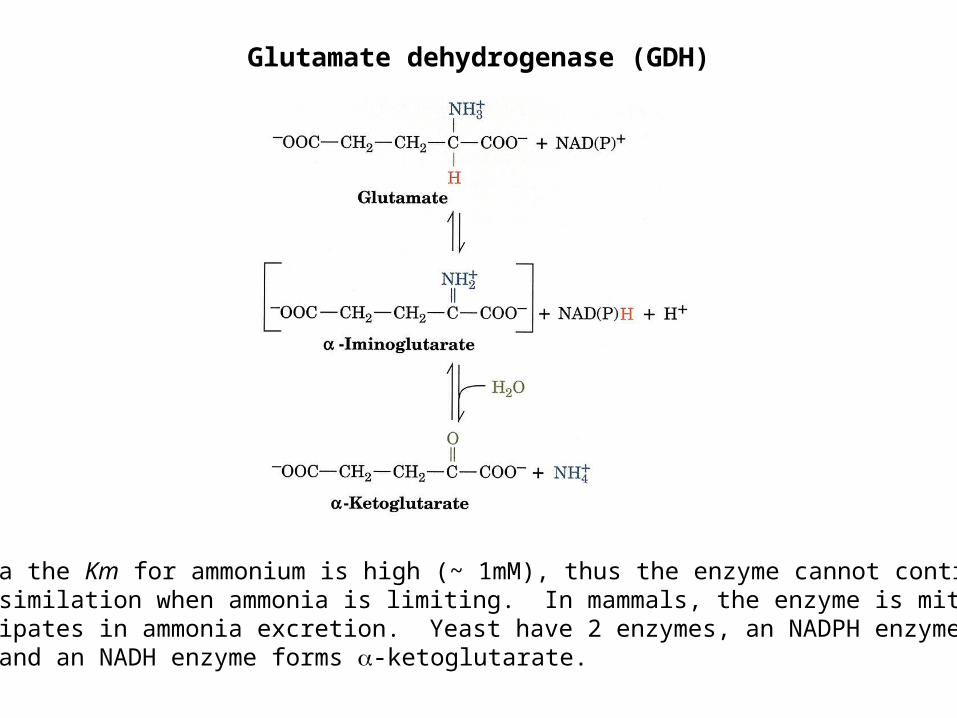
Glutamate dehydrogenase (GDH)
In bacteria the Km for ammonium is high (~ 1mM), thus the enzyme cannot contribute toammonia assimilation when ammonia is limiting. In mammals, the enzyme is mitochondrialand participates in ammonia excretion. Yeast have 2 enzymes, an NADPH enzyme formsglutamate and an NADH enzyme forms -ketoglutarate.

The glutamate synthase (GOGAT) reaction

Under conditions of ammonia limitation, the GS-GOGAT cycle is used for ammonia assimilation in bacteria and plants
2-ketoglutarate + NH3 + NAD(P)H + H+ glutamate + NAD(P)+
GDH
glutamate + ATP + NH3 glutamine + ADP + Pi GS
glutamine + 2-ketoglutarate + NADPH + H+ 2 glutamate + NADP+
glutamate synthase (GOGAT)
Sum of GS + GOGAT:
2-ketoglutarate + NH3 + ATP + NADPH + H+ glutamate + ADP + Pi + NADP+

Salmonella thyphimurium GS
top view showing ADP and 2 Mn
Adjacent subunits form the active sites

Glutamine synthetase reaction mechanism
ATP binds to GS
glutamate binds to (E.ATP)
E.ATP.glu ----> E.ADP.glutamyl--P
conformational change favors NH4+ binding
deprotonation of NH4+ by an Asp causes a flap (324-328)
to close over active site
ammonia attacks glutamyl--P forming tetrahedral intermediate
Pi and a proton are lost
The flap opens and glutamine leaves

Regulation of E. coli glutamine synthetase
E. coli is reported to be regulated in three distinct ways:
1. Cumulative feedback inhibition
2. Reversible covalent modification (adenylylation)
3. Regulation of enzyme synthesis

Cumulative feedback inhibition of GS
The enzyme is inhibited by the following compounds:
alanine, glycine, tryptophan, histidine, carbamyl phosphate,glucosamine-6-phosphate, CTP, and AMP
Each of the inhibitors provides only partial inhibition, completeinhibition requires all of the inhibitors.
Kinetic studies suggested that none of the inhibitors was competitivewith substrates.
BUT-Structural studies show a different picture:
AMP binds at the ATP substrate siteGly, ala, and ser bind at the glu sitecarbamyl phosphate binds overlapping the glu and Pi sitesthe binding of carbamyl phosphate prevents the binding of ala, gly, and ser.

GS is regulated by reversible covalent adenylylation
GS(active)
GS~AMP(inactive)
ATase
ATase
ATP PPi
ADP Pi

The activity and level of Glutamine Synthetase (GS) are regulated by theratio of carbon to nitrogen
Nutrient broth culture(N>C) C>N N>C
level of GS is low level of GS is high level of GS is low
GS mostly adenylylated GS mostly unadenylylated GS mostly adenylylated
add glucoseto 1%
add glutamineto 0.2%

GS is regulated at both the transcriptional and post-transcriptional levels
Ammonia scarce Ammonia plentiful
GS not adenylylated GS adenylylated
glnA gene highly expressed glnA gene not highly expressed
A large amount of very A small amount of enzyme thatactive enzyme is mostly inactive

Two bicyclic cascades control GS synthesis and activity
PII
PII-UMP
GS
GS-AMP
UTase/UR UTURgln gln glngln
ATaseAR AT
ketoglutarate
UTase/UR/PII monocycle ATase/GS monocycle
PII
PII-UMP
UTase/UR UTURgln gln
NRI~P
NRI
NRIIATP
ADP
ketoglutarate
NRII~P
NRII
UTase/UR/PII monocycle

Uridylyltransferase/uridylyl-removing enzyme measures glutamineand controls the activity of PII
PII PII~UMPUTase
UR
glutamine
glutamine
(N-rich) (N-poor)
KNTase 13-RMKIVHEIKERILDKYGDDVKAIGVYGSLGRQTDGPYSDIEMMCVMSTEE-(2)-FSHEWIT * * * **** ** * ****
DNA POL 154-MLQMQDIVLNEVKKL-DPEY-IATVCGSFRRGAES-SGDMDVLLTHPNFT-(31)-TKFMGVC * * * **** ** * ****E. c. UTase/UR 68-IDQLLQRLWIEAGFSQIADL-ALVAVGGYGRGELHPLSDVDLLILSRKKL-(6)-KVGELLT
AA A N N Figure 10. Alignment of the known active sites from kanamycin nucleotidyl transferase and rat DNA polymerase with theN-terminal part of the UTase/UR. The structures of KNTase and Pol are known. Below the UTase/UR sequence, the locations of the G93A, G94A, G98A, D105N, and D107N mutations in glnD are shown.

Structure of the unliganded form of PII
E. coli PII (top view) E. coli PII (side view)
Cyanobacterial PII (top view)E. coli PII subunit
T-Loop
B-Loop
C-Loop

Biphasic response of GS adenylylation reaction to 2-KG
GS + ATP GS~AMP + PPi gln
ATase + PII

Binding of 2-KG to PII (30 M) when ATP is present in excess
PII contains non-equivalent 2-KG binding sites

[Uridylylation reduces negativecooperativity in 2KGbinding]
Kd~ 5 M
Kd~ 150 M
No interaction withATase or NRII
Interacts with ATaseand NRII
No interaction withATase or NRII
UMP
UMPUMP
UM
P
2-ketoglutarate
low Gln
high Gln
low Gln
high Gln
No interaction withATase of NRII
high Gln low Gln
PII protein integrates antagonistic signals

NRII(NRI kinase)
NRII::PII(NRI~P phosphatase)
ATase ATase::PII(AT activity)
ATase::PII-UMP(AR activity)
UTase/UR(UT activity)
UTase/UR(UR activity)
glutamine
-ketoglutarate
uridylyl group
PII
PII-UMP
-k
eto
glu
tara
te
glutamine
PII protein integrates antagonistic signals

Reconstitution of the UTase/UR-PII monocycle
At physiological concentration of 2-KG and gln, only gln regulatesPII uridylylation state.

Reconstitution of the UTase/UR-PII-ATase-GS bicycle
both gln and 2KGregulate the bicycle
Only gln regulatesthe UTase/UR-PIImonocycle

Glutamine regulates the phosphorylation state of NRI by acting on UTase/UR

Response of the bicyclic system to glutamine addition

2-Ketoglutarate regulates NRI phosphorylation state, but not PIIuridylylation state in the bicyclic system

The two bicycles respond differently to glutamine

Gene cascade controlling nitrogen assimilation and fixation
glnALG
GS NRI NRII
NRI~P
NRIIPII
glnK
GlnK
nifLA
NifA NifL
nif genes
Ntr genes
Signals
“Level 1”
“Level 2”
“Level 3”
nac
activation andrepression of genes
Nac





























