Embed Size (px)
Citation preview

Vol. 152, No. 2JOURNAL OF BACTERIOLOGY, Nov. 1982, p. 626-6350021-9193/82/110626-10$02.00/0Copyright © 1982, American Society for Microbiology
Regulation of Glutamine Synthetase Activity and Synthesis inFree-Living and Symbiotic Anabaena spp.
JAMES ORRt AND ROBERT HASELKORN*Department ofBiophysics and Theoretical Biology, University of Chicago, Chicago, Illinois 60637
Received 24 May 1982/Accepted 4 August 1982
Regulation of the synthesis and activity of glutamine synthetase (GS) in thecyanobacterium Anabaena sp. strain 7120 was studied by determining GStransferase activity and GS antigen concentration under a variety of conditions.Extracts prepared from cells growing exponentially on a medium supplementedwith combined nitrogen had a GS activity of 17 ,umol of y-glutamyl transferaseactivity per min per mg of protein at 37°C. This activity doubled in 12 h aftertransfer of cells to a nitrogen-free medium, corresponding to the time required forheterocyst differentiation and the start of nitrogen fixation. Addition of NH3 to aculture 11 h after an inducing transfer immediately blocked the increase in GSactivity. In the Enterobacteriaceae, addition of NH3 after induction results in thecovalent modification of GS by adenylylation. The GS of Anabaena is notadenylylated by such a protocol, as shown by the resistance of the transferaseactivity of the enzyme to inhibition by Mg2e and by the failure of the enzyme toincorporate 32P after NH3 upshift. Methionine sulfoximine inhibited AnabaenaGS activity rapidly and irreversibly in vivo. After the addition of methioninesulfoximine to Anabaena, the level of GS antigen neither increased nor decreased,indicating that glutamine cannot be the only small molecule capable of regulatingGS synthesis. Methionine sulfoximine permitted heterocyst differentiation andnitrogenase induction to escape repression by NH3. Nitrogen-fixing culturestreated with methionine sulfoximine excreted NH3. The fern Azolla carolinianacontains an Anabaena species living in symbiotic association. The Anabaenaspecies carries out nitrogen fixation sufficient to satisfy all of the combinednitrogen requirements of the host fern. Experiments by other workers have shownthat the activity ofGS in the symbiont is significantly lower than the activity ofGSin free-living Anabaena. Using a sensitive radioimmune assay and a normalizationprocedure based on the content of diaminopimelic acid, a component unique tothe symbiont, we found that the level of GS antigen in the symbiont was about 5%of the level in free-living Anabaena cells. Thus, the host fern appears to represssynthesis of Anabaena GS in the symbiotic association.
In the Enterobacteriaceae, glutamine synthe-tase (GS) is regulated at levels of synthesis andactivity. Synthesis, which is controlled at thelevel of transcription of the glnA gene by othernitrogen-regulatory gene products, has been thesubject of considerable study (16). A majormechanism for the short-term regulation of GSactivity is adenylylation of the enzyme (covalentattachment of AMP) through a cascade of reac-tions controlled by the relative levels of gluta-mine and a-ketoglutarate; the enzyme graduallyloses activity as increasing numbers of subunitsare adenylyated. For practical purposes, thiscascade is activated most strongly by nitrogenupshift (4). The first question addressed in this
t Present address: University of Illinois at the MedicalCenter, Chicago, IL 60612.
paper is whether Anabaena GS is subject toregulation by adenylylation.We approached this question in the following
two ways: through direct examination of the GSprotein for covalent binding of 32P and throughinvestigation of the response of the enzyme tothe metal cofactors Mge' and Mn2 . In Esche-richia coli, the latter method is diagnostic foradenylylation, which renders the enzyme ex-tremely susceptible to inhibition by Mg2+ (29).We found no evidence for adenylylation witheither method, even under conditions where itwas to be expected most.
Examination of the protein for 32P binding wasdone by using anti-GS antiserum; characteriza-tion of this antiserum is described below. Wethen used the antiserum to explore the relation-ship among GS activity, GS synthesis, and nitro-
626
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

REGULATION AND SYNTHESIS OF GS IN ANABAENA
genase synthesis, both in free-living Anabaenaand in Anabaena living in association with thewater fern Azolla. In this symbiosis, Anabaenain leaf cavities provides all of the fixed nitrogenrequired by the host fern.
MATERIALS AND METHODSStrains and growth conditions. The cyanobacterium
Anabaena sp. strain 7120 was originally obtained fromthe Botany Department of Iowa State University,where it was catalogued as Nostoc muscorum. Themedium used for growth was Chu 10 medium (17) withthe Na2CO3 concentration raised fivefold to 1 mM andthe Ca(NO3)2 concentration lowered threefold to 0.33mM. The cyanobacteria Anabaena azollae strains laand 2a were supplied by Jack Newton of the UnitedStates Department of Agriculture Research Station,Peoria, Ill. (19), and were grown in the mediumdescribed above.The water fern Azolla caroliniana in native and
cyanobacterial symbiont-free forms was also suppliedby Jack Newton. This organism was grown floating on6 liters of medium in 15-liter carboys. Air filteredthrough 0.4S5-tm membrane filters was gently blownon the cultures. The media used were those describedby Peters and Mayne (23). The symbiotic cyanobacte-rium (as distinct from the free-living form) was isolatedfrom the water fern by a modification of the procedureof Peters and Mayne (23). The roots were removedfrom the fern with scissors, and the fern was thencrushed in a pasta machine (Schiller and Asmus, Inc.,Chicago, Ill.) whose rollers were adjusted to a 0.003-in. (0.076-mm) clearance. The crushed fern was thensuspended in Chu 10 medium to give a 10%o (wt/vol)solution and ifitered through two thicknesses of 100-,um nylon mesh. The filtrate, which contained mostlycyanobacterial cells and cellular debris, was washedseveral times with Chu 10 medium to remove thedebris and was pelleted by centrifugation for S min at1,000 x g between washings.
Purification and assay of GS. The enzyme waspurified and assayed exactly as described previously(20, 21). Protein determination and polyacrylamide gelelectrophoresis were also done as described previous-ly (21). A 200-pg amount of purified GS in buffercontaining 50 mM imidazole, 0.2 mM EDTA, 1 mMmercaptoethanol, and 1 mM MnCl2 (pH 7) (designatedDB/Mn) emulsified 1:1 with Freund complete adjuvantwas injected into two rabbits. Serum taken from therabbits 5 weeks after the initial injection showed goodresponse to purified GS on Ouchterlony plates. Subse-quently, rabbits were injected with approximately 80SLg of purified GS in DB/Mn, which was emulsified inFreund incomplete adjuvant, from 10 days to 2 weeksbefore a need for antiserum was anticipated.Immunological diffusion methods. Ouchterlony
plates for immunodiffusion were prepared by themethod of Garvey et al. (10), using DB/Mn as the gelbuffer. Immunoelectrophoresis was performed by themethod of Weeke (33), with the gel and the electrodechambers containing Svendsen buffer (sodium barbi-tal, 6.5 g/liter; barbital, 1.04 g/liter; glycine, 58.2 g/liter; and Tris, 52.7 g/liter). A 1% agarose gel withbuffer was poured 1.5 mm thick on a glass slide (3.25by 4 in. [8.26 by 10.16 cm]); sample wells (diameter, 3mm) were then cut centered 26.5 mm from a narrow
side of the gel. A 10-pl portion of sample was appliedto each well, and electrophoresis was carried out at 40V/cm in a DeSaga thin-layer electrophoresis chamberfor 1 h with water cooling. Slits 86 mm long were thencut between the sample wells and filled with antise-rum. The precipitin lines were allowed to develop atroom temperature for 24 h. The plates were deprotein-ized at room temperature in two changes of 2% NaCl-0.01% NaN3 for 2 days, dried, stained in 0.5% Coo-massie brilliant blue R-250-10% acetic acid-43% etha-nol for 10 min, destained in 10% acetic acid-45%ethanol until the background was colorless, and dried.
Radloimmunological methods. lodination of GS forradioimmune assays was performed by chloramine Toxidation or solid-phase glucose oxidase-lactoperoxi-dase oxidation. A typical chloramine T oxidation isdescribed below. Chloramine T (4 mg/ml in 0.1 Mpotassium phosphate-0.15 M NaCl, pH 7.5) and sodi-um metabisulfite (4 mg/ml in the same buffer) solutionswere freshly prepared. A 1-ml portion of an enzymefraction containing 0.5 mg of protein from a BlueSepharose column (21) was added to 1 ml of thephosphate-NaCl buffer described above. Carrier (non-radioactive) KI was added to the protein on an equi-molar basis (1 mol of KI per mol of GS subunits; 7 ,ulof 1 mM KI in water). Then 1 mCi of Na125I wasadded, followed by 50 p.g of chloramine T in solution(12.5 ILI, or 0.1 g of chloramine T per g of protein). Thereaction was allowed to proceed at room temperaturefor 5 min. The degree of iodination was checked induplicate runs. A 0.5-,lI portion of iodination solutionwas added to 1 ml of 10o rabbit serum and mixed.Then 1 ml of 20%o trichloroacetic acid was added andmixed. The solutions were centrifuged briefly, and thesupernatants were withdrawn. Both precipitates andsupernatants were assayed with a gamma ray counter;15% of the radioactivity was found in the precipitate ineach run. A 0.05-mg amount of chloramine T wasadded, and the solution was allowed to react for 5 minlonger. Another check of trichloroacetic acid-precipi-table radioactivity showed no further incorporation ofiodine into the precipitate. The reaction was stoppedby adding 30 .Ll of a metabisulfite solution (slightlymore than the total amount of the chloramine Tsolution used in the reaction). The reaction mixturewas then dialyzed against several changes ofDB/Mn inthe cold. Solid-phase iodinations were also performed,as described by Orr (Ph.D. thesis, University of Chica-go, 1981).Radioimmune assays were done by using Formalin-
fixed, heat-killed Staphylococcus aureus Cowan Istrain prepared by the method of Kessler (15) (the kindgift of Ralph Buttyan and Patricia Spear) to precipitatethe antigen-antibody complex (14, 15). All dilutionswere carried out with DB/Mn supplemented with 0.1%bovine serum albumin and 0.15% NaCl. In an individ-ual assay, antibody, labeled GS, and an unknownsample were added to a disposable plastic tube, mixed,and allowed to stand for 15 min at room temperature.S. aureus was then added with mixing, and the solu-tion was centrifuged at 12,000 x g for 1 min. Theradioactivity remaining in the supernatant was thenmeasured. Greater quantities of GS antigen in theunknown would have released more labeled GS intothe supernatant by displacement of labeled materialfrom the antigen-antibody complex.A convenient level of radioactivity was chosen for
VOL. 152, 1982 627
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

628 ORR AND HASELKORN
10 mp10 10c t
Samp e conc.n troation
I0 - -*0
FIG. 1. Radioimmune assay determination of GSantigen in a lysate ofAnabaena sp. strain 7120. A 190-,ul amount of an appropriate dilution of crude extract(O) or purified GS (0) was mixed with 10 ,ul ofantiserum (diluted 103) and 8 p.1 of 125I-labeled GS andallowed to stand for 15 min at room temperature. Then21 pLI of 2% S. aureus was added, the suspension wascentrifuged at 12,000 x g, and 150 ,ul of the resultingsupernatant was counted with a gamma ray counter.The unknown was 1.85 times less concentrated thanthe purified GS. Similarly, dilutions of extracts ofAnabaena azollae (*) and Azolla caroliniana (A)were assayed. The Anabaena azollae (free-living en-dosymbiont) extract was 63.6 times more concentratedthan the Azolla (symbiotic association) extract.
the individual assays (usually 1,000 cpm), and theremaining parameters were adjusted to that figure. S.aureus was always sufficient at a level of 21 p,l of a2.0% (vol/vol) solution per assay to precipitate all ofthe antigen-antibody complexes, since at maximumantibody concentrations 96% of the radiolabel was
precipitable. The amount of antibody needed to pre-cipitate 70% of the label (typically, 10 of a 10'-dilution) was chosen as the operating concentration forthe assay. An extract containing an unknown amountofGS antigen was assayed by adding serial dilutions totubes containing antibody, radiolabeled GS, and S.aureus. The concentration of GS antigen in the un-
known could then be read from the inflection point onthe competition curve, compared with the same pointon a standard curve generated with purified GS (Fig.1).
Nitrogenase assays. Nitrogenase was assayed bymeasuring the reduction of acetylene to ethylene (6,27). For Anabaena, this assay was performed instoppered 10-ml serum bottles containing 5 ml ofculture. A 1-ml portion of acetylene was injected intothe gas phase, and the bottle was illuminated withapproximately 4,000 lux of light with occasional agita-tion. The reaction was stopped after 1 h by injecting 1ml of 0.5 M H2SO4, and the concentration of ethylenein the gas phase was analyzed by chromatography on a
Porapak R column in a Perkin-Elmer model 3920B gaschromatograph equipped with a flame ionization de-tector. Under these conditions acetylene reductionwas linear for at least 1 h (Orr, Ph.D. thesis).For Azolla, the assay was performed in 25-ml serum
bottles containing approximately 1 g ofAzolla and 9 ml
ofAzolla growth medium; the other conditions were asdescribed above. However, no H2SO4 was added tostop the reaction. Instead, the exact time of samplewithdrawal was noted, and the acetylene reductionrate was calculated from the time of withdrawal andthe ethylene concentration measured.
Preparation of [rSJmethionine sulfoximine. Radiola-beled methionine sulfoximine was prepared by usingthe procedure of Bentley et al. (2), modified formicroscale quantities. Methionine was first oxidized tomethionine sulfoxide by using hydrogen peroxide in anacid solution. The product sulfoxide was then reactedwith HN3 in the presence of H2SO4. In a typicalpreparation, 1 mCi of L-[35S]methionine (600 to 1,300Ci/mmol) was placed in an 8-ml conical plastic centri-fuge tube with 10 pA of a 0.1% solution of unlabeledmethionine in water. Then 2 ,ul of concentrated HCIand 2 ,ul of H202 (30% in water) were added. Themixture was heated in a boiling water bath for 3 minand then cooled immediately. The reaction mixturewas frozen and lyophilized to remove the HCI andunreacted H202. The product was dissolved in 30 p.l ofwater and transferred to a 4-mm Pyrex tube, whichhad previously been silanated with 5% dimethyldich-lorosilane in CHCl3, washed with CHCI3 and water,and sealed at one end. Three successive 30-pl washesof the conical reaction tube were then added to thePyrex tube, which was then frozen in liquid N2 andlyophilized.A 30-plI amount of H2SO4-water (5:1) was then
added to the tube, which was briefly centrifuged, and75 plA of HN3 prepared by the method of Fieser andFieser (8) was added. The tube contents were frozen inliquid N2, evacuated under a vacuum (while remainingimmersed in liquid N2), and sealed at the open endwith a torch. This sealed tube was then immersed in anNa2CO3 solution (to prevent escape of HN3 in case oftube rupture) and kept at 43°C for 5 h. The tube wasthen centrifuged, frozen in liquid N2, and broken open.The product was isolated by chromatography on amicrocolumn (Pasteur pipette) containing 400-meshDowex-50 resin. Further purification was accom-plished by thin-layer chromatography on MN-300 cel-lulose plates in n-butanol-acetone-water-diethylamine(10:10:5:2) and thin-layer electrophoresis. Electropho-resis was performed at 20 V/cm for 1 h at right anglesto the first separation, using 7% HCOOH. Methylgreen, which was used as a marker, migrated about40%o of the distance available in this time.
Preparation of labeled extracts and Immunoprecipi-tates. A 250-ml Anabaena sp. strain 7120 log-phaseculture containing ammonia as the nitrogen source wascentrifuged and suspended in 100 ml of PB medium(34) containing MgSO4 at a concentration of 1 pFM.H235S04 (carrier-free) was added to the culture at aconcentration of 10 ,uCi/ml. The culture was thenaerated with 0.5% CO2 in air, and growth was allowedto continue for 2 days. The cells were harvested,suspended in NET buffer (150 mM NaCl, 5 mMEDTA, 50 mM Tris, 0.02% NaN3, 0.5% Nonidet P-40[NP-40], pH 7.4), and then lysed in a French pressurecell at 20,000 lb/in2. The lysate was centrifuged at7,700 x g for 15 min. The resulting pellet was washedwith 0.5 ml of NET buffer and centrifuged again, andthe washings were combined with the first superna-tant.Two 0.5-ml samples of this supernatant were taken.
J. BACTERIOL.
. -t9.
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

REGULATION AND SYNTHESIS OF GS IN ANABAENA 629
A B C, w.
*.:zai> .:.:
*:... ..
b. s|Er..
se; .. :.....FIG. 2. Autoradiogram of a polyacrylamide gel
electrophoresis analysis of immunoprecipitates of a"5SO4-grown culture ofAnabaena sp. strain 7120 withanti-GS antiserum. Lane A, Crude lysate, produced byFrench pressure cell disruption ofAnabaena at 20,000lb/in2 and removing debris by centrifugation for 1 minat 15,000 x g; lane B, immunoprecipitation of lysateusing anti-GS antiserum; lane C, immunoprecipitateusing preimmune serum.
To one of these samples we added 20 ,ul of antiserumraised against GS; the second, a control, was treatedidentically, except that preimmune serum was used.After 15 min of incubation at room temperature, 0.1 mlof a 10%o suspension of heat-killed, glutaraldehyde-fixed S. aureus was added (15). After 30 min ofincubation, 2.5 ml of NET buffer was added, and thesample was centrifuged for 6 min at 650 x g. The pelletwas washed twice with 1 ml of NET buffer andcentrifuged after each wash as described above. Thepellet was then suspended in 100 g1l of cracking buffer,heated, and subjected to electrophoresis and subse-quent fixing and staining.To prepare a 32P-labeled extract ofAnabaena, 10 ml
of stock culture of Anabaena sp. strain 7120 wasadded to 100 ml of Chu 10 medium lacking Ca(NO3)2but supplemented with 2.8 ml of 0.6 M HEPES (N-2-
hydroxyethylpiperazine-N'-2-ethanesulfonic acid)buffer (pH 7.7) and 1.4 mCi of H332PO4 (carrier-free).A 20-ml portion of the culture was harvested; to one-half of this sample we added 1 ml of 1.1% cetyltrimeth-ylammonium bromide (CTAB). This reagent has beenfound to stabilize the adenylylated form ofGS in somegram-negative bacteria (1, 13). The culture was centri-fuged for 5 min at 7,700 x g. The cells were transferredto smaller tubes, pelleted by centrifugation at 15,000 xg for 30 s, frozen in liquid N2, and stored at -20°C.Immediately after these samples were withdrawn fromthe culture, 0.9 ml of 100 mM NH4Cl was added to theremainder, which raised the NH4+ concentration from0 to 1.1 mM. The cells were harvested as describedabove 1.1 and 2.5 h after the ammonium upshift. Onthe next day, lysates were prepared from the frozencells; 0.5 ml ofDB/Mn supplemented with 0.5% NP-40was added to the cells which had been harvestedwithout CTAB. Similarly, DB/Mn supplemented with0.05% NP-40 and 0.1% CTAB was added to the cellswhich had been harvested with CTAB. The cells werelysed by sonic disruption, and 50 ,ul of a 10%6 suspen-sion of heat-killed, glutaraldehyde-fixed S. aureuscells was added to each tube to control any nonspecificprecipitation by S. aureus. After centrifugation 20 Plof anti-GS antiserum and 50 Pl of an S. aureussuspension were added to the supernatants. After 15min of incubation at room temperature, the sampleswere centrifuged for 2 min at 15,000 x g. The resultingpellets were washed twice with the same buffer usedfor lysis. To disperse the samples in the electrophore-sis cracking buffer, it was necessary to sonicate thembriefly. Electrophoresis was then performed as de-scribed above.Other procedures. Ammonia was determined by the
method of Chaney and Marbach (3). A sample (0.333ml) was mixed with 0.333 ml of a solution containing50 g of phenol per liter and 250 mg of Na2Fe(NO)(CN)sper liter. An oxidizing solution (2.1 g of NaOCl perliter, 0.625 M NaOH) was added with mixing, and theabsorption was read at 625 nm after 30 min at roomtemperature.
Diaminopimelic acid was determined by hydrolyz-ing samples in 6 N HCI for 24 h, followed by analysisusing automated single-column ion-exchange chroma-tography and a Durrum D-500 analyzer.
RESULTSCharacterization of antiserum raised against
GS. To characterize the antiserum to AnabaenaGS, we determined the specificity with whichthis antiserum precipitates proteins from Ana-baena crude extracts. To do this, we used 35S-labeled crude extracts and precipitated anti-body-antigen complexes with S. aureus proteinA. As described above, an Anabaena sp. strain7120 log-phase culture was labeled with 35 O4for 48 h. The cells were lysed, and two samplesof the centrifuged lysate were removed. To oneof these antiserum was added, and the other wasa control. Heat-killed, glutaraldehyde-fixed S.aureus was added and then removed by centrifu-gation; the resulting pellet of fixed cells withabsorbed antibody-antigen complexes was
VOL. 152, 1982
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
630 ORR AND HASELKORN
TIME (HOURS)
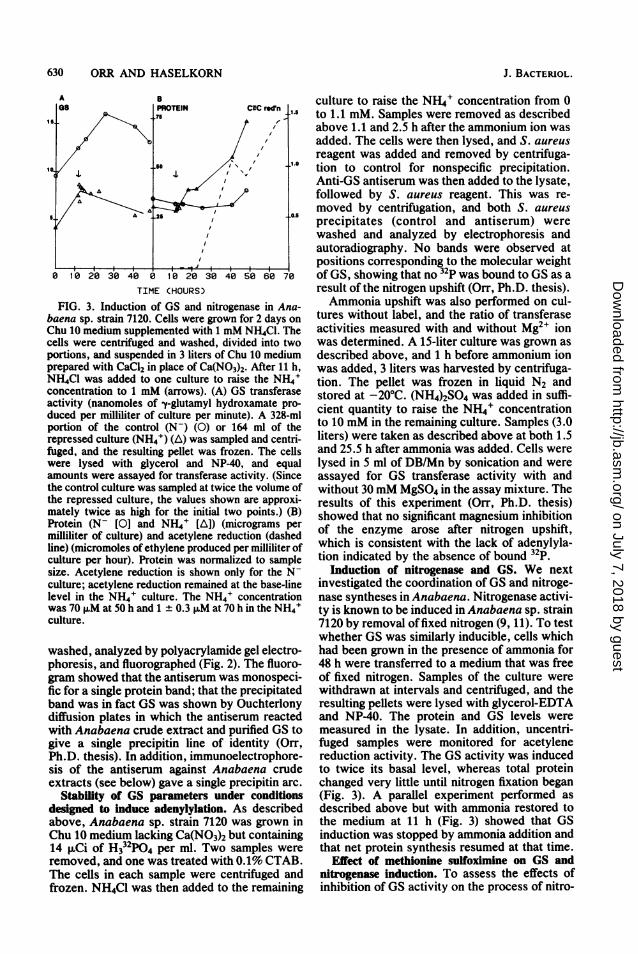
FIG. 3. Induction of GS and nitrogenase in Ana-baena sp. strain 7120. Cells were grown for 2 days onChu 10 medium supplemented with 1 mM NH4CI. Thecels were centrifuged and washed, divided into twoportions, and suspended in 3 liters of Chu 10 mediumprepared with CaCl2 in place of Ca(N03)2. After 11 h,NH4C1 was added to one culture to raise the NH4'concentration to 1 mM (arrows). (A) GS transferaseactivity (nanomoles of y-glutamyl hydroxamate pro-
duced per milliliter of culture per minute). A 328-mlportion of the control (N-) (0) or 164 ml of therepressed culture (NH4+) (A) was sampled and centri-fuged, and the resulting pellet was frozen. The cellswere lysed with glycerol and NP-40, and equalamounts were assayed for transferase activity. (Sincethe control culture was sampled at twice the volume ofthe repressed culture, the values shown are approxi-mately twice as high for the initial two points.) (B)Protein (N- [01 and NH4' [A]) (micrograms permilliliter of culture) and acetylene reduction (dashedline) (micromoles of ethylene produced per milliliter ofculture per hour). Protein was normalized to samplesize. Acetylene reduction is shown only for the N-culture; acetylene reduction remained at the base-linelevel in the NH4' culture. The NH4' concentrationwas 70 F.M at 50 h and 1 ± 0.3 ,uM at 70 h in the NH4'culture.
washed, analyzed by polyacrylamide gel electro-phoresis, and fluorographed (Fig. 2). The fluoro-gram showed that the antiserum was monospeci-fic for a single protein band; that the precipitatedband was in fact GS was shown by Ouchterlonydiffusion plates in which the antiserum reactedwith Anabaena crude extract and purified GS togive a single precipitin line of identity (Orr,Ph.D. thesis). In addition, immunoelectrophore-sis of the antiserum against Anabaena crudeextracts (see below) gave a single precipitin arc.
Stability of GS parameters under conditionsdesigned to induce adenylylation. As describedabove, Anabaena sp. strain 7120 was grown inChu 10 medium lacking Ca(NO3)2 but containing14 pCi of H332P04 per ml. Two samples wereremoved, and one was treated with 0.1% CTAB.The cells in each sample were centrifuged andfrozen. NH4Cl was then added to the remaining
culture to raise the NH4' concentration from 0to 1.1 mM. Samples were removed as describedabove 1.1 and 2.5 h after the ammonium ion wasadded. The cells were then lysed, and S. aureusreagent was added and removed by centrifuga-tion to control for nonspecific precipitation.Anti-GS antiserum was then added to the lysate,followed by S. aureus reagent. This was re-moved by centrifugation, and both S. aureusprecipitates (control and antiserum) werewashed and analyzed by electrophoresis andautoradiography. No bands were observed atpositions corresponding to the molecular weightof GS, showing that no 32P was bound to GS as aresult of the nitrogen upshift (Orr, Ph.D. thesis).Ammonia upshift was also performed on cul-
tures without label, and the ratio of transferaseactivities measured with and without Mg2+ ionwas determined. A 15-liter culture was grown asdescribed above, and 1 h before ammonium ionwas added, 3 liters was harvested by centrifuga-tion. The pellet was frozen in liquid N2 andstored at -20°C. (NH4)2SO4 was added in suffi-cient quantity to raise the NH4' concentrationto 10 mM in the remaining culture. Samples (3.0liters) were taken as described above at both 1.5and 25.5 h after ammonia was added. Cells werelysed in 5 ml of DB/Mn by sonication and wereassayed for GS transferase activity with andwithout 30 mM MgSO4 in the assay mixture. Theresults of this experiment (Orr, Ph.D. thesis)showed that no significant magnesium inhibitionof the enzyme arose after nitrogen upshift,which is consistent with the lack of adenylyla-tion indicated by the absence of bound 32p.
Induction of nitrogenase and GS. We nextinvestigated the coordination of GS and nitroge-nase syntheses in Anabaena. Nitrogenase activi-ty is known to be induced in Anabaena sp. strain7120 by removal of fixed nitrogen (9, 11). To testwhether GS was similarly inducible, cells whichhad been grown in the presence of ammonia for48 h were transferred to a medium that was freeof fixed nitrogen. Samples of the culture werewithdrawn at intervals and centrifuged, and theresulting pellets were lysed with glycerol-EDTAand NP-40. The protein and GS levels weremeasured in the lysate. In addition, uncentri-fuged samples were monitored for acetylenereduction activity. The GS activity was inducedto twice its basal level, whereas total proteinchanged very little until nitrogen fixation began(Fig. 3). A parallel experiment performed asdescribed above but with ammonia restored tothe medium at 11 h (Fig. 3) showed that GSinduction was stopped by ammonia addition andthat net protein synthesis resumed at that time.
Effect of methionine sulfoximine on GS andnitrogenase induction. To assess the effects ofinhibition of GS activity on the process of nitro-
J. BACTERIOL.
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
REGULATION AND SYNTHESIS OF GS IN ANABAENA 631
TIME (HOURS)
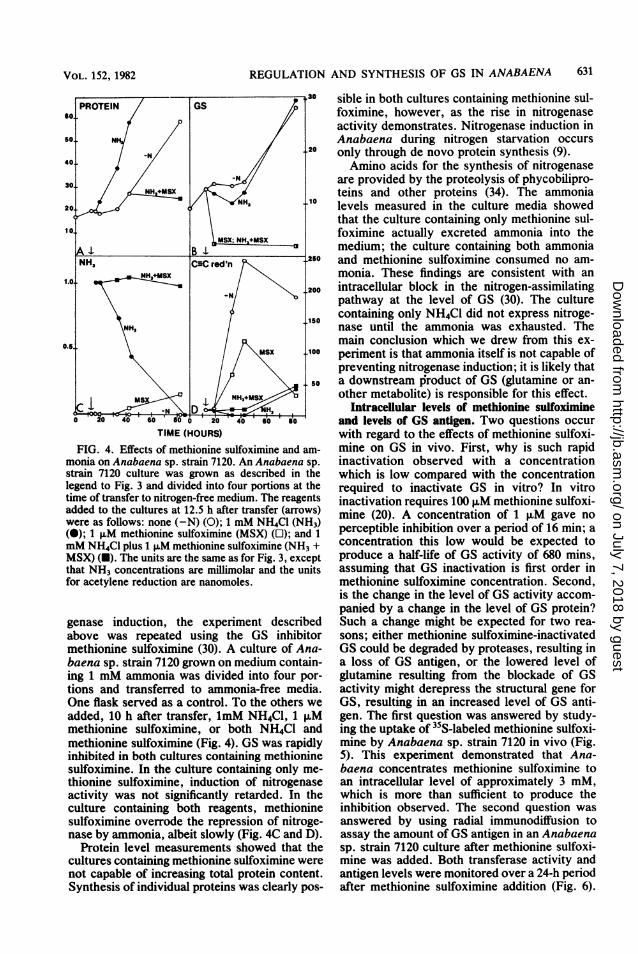
FIG. 4. Effects of methionine sulfoximine and am-monia on Anabaena sp. strain 7120. An Anabaena sp.strain 7120 culture was grown as described in thelegend to Fig. 3 and divided into four portions at thetime of transfer to nitrogen-free medium. The reagentsadded to the cultures at 12.5 h after transfer (arrows)were as follows: none (-N) (0); 1 mM NH4Cl (NH3)(e); 1 F.M methionine sulfoximine (MSX) (O); and 1mM NH4Cl plus 1 ,uM methionine sulfoximine (NH3 +MSX) (U). The units are the same as for Fig. 3, exceptthat NH3 concentrations are millimolar and the unitsfor acetylene reduction are nanomoles.
genase induction, the experiment describedabove was repeated using the GS inhibitormethionine sulfoximine (30). A culture of Ana-baena sp. strain 7120 grown on medium contain-ing 1 mM ammonia was divided into four por-tions and transferred to ammonia-free media.One flask served as a control. To the others weadded, 10 h after transfer, 1mM NH4Cl, 1 ,uMmethionine sulfoximine, or both NH4Cl andmethionine sulfoximine (Fig. 4). GS was rapidlyinhibited in both cultures containing methioninesulfoximine. In the culture containing only me-thionine sulfoximine, induction of nitrogenaseactivity was not significantly retarded. In theculture containing both reagents, methioninesulfoximine overrode the repression of nitroge-nase by ammonia, albeit slowly (Fig. 4C and D).
Protein level measurements showed that thecultures containing methionine sulfoximine werenot capable of increasing total protein content.Synthesis of individual proteins was clearly pos-
sible in both cultures containing methionine sul-foximine, however, as the rise in nitrogenaseactivity demonstrates. Nitrogenase induction inAnabaena during nitrogen starvation occursonly through de novo protein synthesis (9).Amino acids for the synthesis of nitrogenase
are provided by the proteolysis of phycobilipro-teins and other proteins (34). The ammonialevels measured in the culture media showedthat the culture containing only methionine sul-foximine actually excreted ammonia into themedium; the culture containing both ammoniaand methionine sulfoximine consumed no am-monia. These findings are consistent with anintracellular block in the nitrogen-assimilatingpathway at the level of GS (30). The culturecontaining only NH4Cl did not express nitroge-nase until the ammonia was exhausted. Themain conclusion which we drew from this ex-periment is that ammonia itself is not capable ofpreventing nitrogenase induction; it is likely thata downstream product of GS (glutamine or an-other metabolite) is responsible for this effect.
Intracellular levels of methionine sulfoximineand levels of GS antigen. Two questions occurwith regard to the effects of methionine sulfoxi-mine on GS in vivo. First, why is such rapidinactivation observed with a concentrationwhich is low compared with the concentrationrequired to inactivate GS in vitro? In vitroinactivation requires 100 ,uM methionine sulfoxi-mine (20). A concentration of 1 ,uM gave noperceptible inhibition over a period of 16 min; aconcentration this low would be expected toproduce a half-life of GS activity of 680 mins,assuming that GS inactivation is first order inmethionine sulfoximine concentration. Second,is the change in the level of GS activity accom-panied by a change in the level of GS protein?Such a change might be expected for two rea-sons; either methionine sulfoximine-inactivatedGS could be degraded by proteases, resulting ina loss of GS antigen, or the lowered level ofglutamine resulting from the blockade of GSactivity might derepress the structural gene forGS, resulting in an increased level of GS anti-gen. The first question was answered by study-ing the uptake of 35S-labeled methionine sulfoxi-mine by Anabaena sp. strain 7120 in vivo (Fig.5). This experiment demonstrated that Ana-baena concentrates methionine sulfoximine toan intracellular level of approximately 3 mM,which is more than sufficient to produce theinhibition observed. The second question wasanswered by using radial immunodiffusion toassay the amount of GS antigen in an Anabaenasp. strain 7120 culture after methionine sulfoxi-mine was added. Both transferase activity andantigen levels were monitored over a 24-h periodafter methionine sulfoximine addition (Fig. 6).
VOL. 152, 1982
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
632 ORR AND HASELKORN
1000
800
nu)
E0.
600
400
200
0 15 30 60 120Time (minutes)
FIG. 5. Uptake of methionine sulfoximine by Ana-baena sp. strain 7120. A culture was grown to aconcentration of 2 x 106 cells per ml. "S-labeledmethionine sulfoximine (specific activity, 1.66 x 106cpm/,umol) was added to a concentration of 1 FM. Atdifferent times after addition of the labeled compound,5-ml samples were filtered through 0.45-~Lm celluloseacetate filters, washed once with 5 ml of water, andcounted with a scintillation counter.
Although GS activity dropped sharply, the anti-gen level remained relatively constant. There-fore, we conclude that the inactivated GS is notdegraded and that glutamine is not a necessarycomponent of the regulatory circuit controllingexpression of the structural gene for GS inAnabaena.GS antigen levels in the Azolla-Anabaena sym-
biosis. We next examined the role of GS in thesymbiotic association of the water fern Azollacaroliniana and the cyanobacterium Anabaenaazollae. In this relationship, Anabaena is knownto fix N2 and to release nitrogen in the form ofammonia, which is then taken up by the fern. Inone model for the association, the fern producesa diffusible GS inhibitor which disables the GS inthe symbiont, causing the latter to excreteammonia for the use of the fern. This model isconsistent with the work of Ray et al. (24), whoshowed that GS activity in the cyanobiont wasdisproportionately low. We have shown that theGS antigen level does not change when methio-nine sulfoximine is added to Anabaena cultures,although the activity declines precipitously.Therefore, in a system structured like the model,we would expect that the comparative levels ofGS antigen would be much the same in symbiot-ic and free-living forms of Anabaena, eventhough the activity levels might be lower in thesymbiont.To be able to measure levels of GS antigen in
Anabaena azollae, we first needed to show thatour antiserum to Anabaena sp. strain 7120 GScross-reacts with the enzyme from Azolla azol-lae. This was demonstrated in two experiments,the radioimmune assay described above (Fig. 1)
and an immunoelectrophoresis experiment (Fig.7). The Anabaena azollae antigen cross-reactedsufficiently to form a precipitin line in the immu-noelectrophoresis experiment and to displaceAnabaena sp. strain 7120 GS from the antigen-antibody complex in the radioimmune assay. Wethen assayed for the presence of GS antigen inAnabaena azollae, both as an endosymbiont inAzolla and in the free-living form (19). To com-pare the antigen levels obtained in the twosituations, a normalization standard was need-ed. Protein did not suffice for this purpose; in thesymbiosis the amount of cyanobacterial proteinvaries from 12 to 22% of the total (24). In theirinvestigation of the partition of GS activity, Rayet al. normalized with respect to phycocyanin,which they measured by fluorescence. We pre-ferred not to use this protein; in Anabaenaphycocyanin is one of the first sources of intra-cellular nitrogen to be scavenged by the nitro-gen-deprived cyanobacterium (18, 32) and thusmight easily be altered under the conditions ofthe symbiosis. This is especially true consider-ing the low level of phycocyanin in the hetero-cysts (7) and the higher-than-normal heterocyst
C0- 1-.AiCaL0
-I.,
-W.
(1
>,
007 24
Time Chours)FIG. 6. GS activity and antigen level after addition
of methionine sulfoximine to Anabaena sp. strain7120. A 15-liter culture was grown on N-free mediumto a density of 5 x 106 cells per ml. A 1.5-liter samplewas centrifuged, and the cells were frozen. Methioninesulfoximine was then added to the remainder of theculture to a concentration of I FM. Additional sampleswere taken at 7 and 24 h after methionine sulfoximinewas added. The cyanobacterial samples were lysed in3 ml of DB/Mn, and serial twofold dilutions weresubjected to radial immunodiffusion against anti-GSantiserum. The zero-time samples were used to con-struct a calibration curve (log spot area versus loglysate concentration), and the dilutions of the 7- and24-h samples were read against this. The multiplepoints plotted represent values obtained from thedifferent dilutions (0). The solid line without symbolsrepresents the transferase activity of the samples.
J. BACTERIOL.
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

REGULATION AND SYNTHESIS OF GS IN ANABAENA 633
An. 7120
An. azollae
symn Az. carol.
Az. carol.
An. 7120 -
!mm
-imm
-pro
FIG. 7. Immunoelectrophoresis of extracts of Ana-baena sp. strain 7120, of Anabaena azollae, and ofAzolla caroliniana with and without the cyanobacter-ial symbiont. Crude extracts were placed in wells andsubjected to electrophoresis with the positive elec-trode at the lower end of the gel. Troughs were thencut and filled with either immune (anti-GS) serum(imm) or preimmune serum (pre).
frequency found in the endosymbiont population(12). Therefore, we performed the assay byusing the following two different normalizationstandards: acetylene reduction activity and theconcentration of diaminopimelic acid, a procary-otic cell wall constituent.The assays standardized to acetylene reduc-
tion were performed as follows. A quantity ofAzolla (approximately 5 g) was assayed foracetylene reduction activity, frozen in liquid N2,and ground in a mortar. This material was thensuspended in the buffer used for radioimmuneassays, and the concentration of GS antigen wasmeasured by a radioimmune assay. Similarly,Anabaena azollae was assayed for acetylenereduction, a known quantity was suspended inthe radioimmune assay buffer, and the cyano-bacterium was lysed by sonic disruption.For assays standardized to diaminopimelic
acid, Anabaena azollae was isolated from Azollaand sonicated in radioimmune assay buffer. Thelysate was divided equally into two samples, oneof which was analyzed for GS antigen and theother of which was analyzed for diaminopimelicacid. The results of these experiments are shownin Table 1. We found that GS antigen levels weresubstantially lower in Anabaena in symbioticassociation than in the free-living form, rangingfrom 5 to 10% in three determinations.The following objection might be raised to the
above-described use of diaminopimelic acid as anormalizing compound: diaminopimelic acid is acell wall constituent, and heterocyst frequency(and the amount of possibly variant cell wallmaterial) is greatly increased in the endosymbi-otic cyanobacterium compared with the free-living form (30 versus 12%) (12). To control forthis, we analyzed the diaminopimelic acid levelsin Anabaena azollae grown on NH4' (whichrepresses heterocysts) and on N2. The diamino-
pimelic acid and phenylalanine levels of thevarious samples analyzed are given in Table 2.The ratio of diaminopimelic acid to phenylala-nine was 0.054 for the undifferentiated culture,0.063 for the differentiated culture, and 0.073 forthe isolated symbiont. The last value is in goodagreement with the value of 0.076 that would beexpected for a sample containing 30% hetero-cysts, based on the values of 0.054 and 0.063obtained for samples containing 0 and 12% het-erocysts, respectively. Correction of thediaminopimelic acid content of the endosym-biont for the increase assumed to be due to theincreased heterocyst content yielded a correctedvalue of 4.1 nmol of diaminopimelic acid and acorrected GS antigen ratio of 15.6, which is stillwithin the range determined by using acetylenereduction activity as the basis for normalizingthe GS antigen level.
DISCUSSIONThe lack of adenylylation which we observed
in GS is not surprising in view of previousstudies in which Anabaena cylindrica GS (26)and Anabaena CA GS (28) were found to beunchanged in their properties after treatmentwith snake venom phosphodiesterase. Otherworkers (31) have shown that the properties ofAnabaena L-31 GS are the same whether the
TABLE 1. Relative levels of GS antigen insymbiotic and free-living Anabaena azollae
Symbiotic Free-living
Expt GS Normal- GS Normal- Ratioca ization .a izationantigena standardb antigen standardb
1d 8 18,040 135 14,080 21.62e 6 27,900 614 29,5000 9.73f 1 4.7 75 19.5 18.1
a GS antigen concentration was defined as unity forthe lysate of Azolla with symbiont; the level forAnabaena was then referred to that. Both were multi-plied by the lysate volume to obtain the total antigenamount, shown here. Azolla free of endosymbiontshowed no detectable level of GS antigen.
b For experiments 1 and 2, acetylene reduction(peak height on a gas chromatograph chart record) inthe total sample is given. For experiment 3, thediaminopimelic acid content (in nanomoles) of thetotal sample is given. Symbiont-free Azolla showed nodetectable diaminopimelic acid or acetylene reduction.
c Ratio of free-living (antigen/normalization stan-dard) to symbiotic (antigen/normalization standard).
d An old, slowly growing culture ofAzolla was used.A fresher, more rapidly growing culture of Azolla
was used.fA fresher, more rapidly growing culture of Azolla
was used. The endosymbiont was isolated from theassociation before its levels of GS antigen and diamin-opimelic were measured.
VOL. 152, 1982
w.
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

634 ORR AND HASELKORN
TABLE 2. Diaminopimelic acid and phenylalaninelevels in Anabaena azollae
Amt of Amt ofdiamin- Alao
Sample opimelic phenylala Ratio'acid rune
(nmol) (nmol)
Isolated symbiont 4.7 64.4 0.073Free-living, N2-grown 19.5 310.6 0.063NH4 -grownb 16.9 303.4 0.056
3.8 73.1 0.052a Ratio of diaminopimelic acid to phenylalanine.b The sample was analyzed twice, the second time
using a smaller quantity than the first time.
enzyme is prepared from ammonia- or N2-growncells.A comparison of the regulatory properties of
Anabaena sp. strain 7120 GS with the regulatoryproperties of some other species, at both syn-thetic and post-translational levels, shows someinteresting relationships. Escherichia coli GSshows the greatest degree of induction (23-fold)(35). This enzyme also is covalently modifiedand shows the weakest degree of feedback inhi-bition. In contrast, Anabaena sp. strain 7120 GSand Bacillus subtilis GS are neither so stronglyinducible nor covalently modifiable; also, theseenzymes show much stronger feedback inhibi-tion. Figure 3 shows a 3.4-fold change in GSspecific activity for Anabaena sp. strain 7120. B.subtilis shows an approximately eightfold spreadbetween maximum and minimum levels (22, 25).Deuel and Stadtman (5) have suggested that thestronger response shown to feedback inhibitorsby B. subtilis GS (in particular its strong re-sponse to glutamine) is necessary because of thelack of a cascade modification system in thisorganism. A similar argument may be made forAnabaena GS and its strong response to alanine,serine, and glycine. The values reported forintracellular amino acid concentrations in Ana-baena cylindrica (26) are in a range (approxi-mately 1 mM for alanine and glycine) whichwould give significant inhibition of GS activityand thus perform effective modulation.Glutamine is neither an inhibitor of Anabaena
GS activity nor a repressor of Anabaena GSsynthesis (Fig. 6). However, ammonia may wellbe a repressor ofGS synthesis in Anabaena. In anitrogen-fixing culture (such as that used for theexperiment shown in Fig. 6), ammonia would beexpected to accumulate rapidly after inhibitionof GS; this would explain the constancy in GSantigen levels after addition of the GS inhibitormethionine sulfoximine. Our exclusion of gluta-mine as a possible negative regulator of GSsynthesis is based on the failure of the GSantigen level to increase after addition of methi-onine sulfoximine.
The relative levels of GS antigen that wefound in free-living and symbiotic cyanobacteriaare consistent with the lowered GS activityfound by other workers in the endosymbiont(24). Specific activity ratios were twofold higherin the free-living form for biosynthetic activityand threefold higher for transferase activity.These ratios are not as high as we have observedfor antigen levels; however, if phycocyanin lev-els (used by Ray et al. for normalization) aredepressed in the endosymbiont, as the increasedheterocyst fraction and nitrogen-starved charac-ter of the endosymbiont lead us to expect, thesymbiont would then appear to be present inlower amounts than it actually is, and the specif-ic GS activity determined for it would be spuri-ously high.Our results suggest that the decreased level of
GS activity in the endosymbiotic cyanobacter-ium is due to control at the level of GS synthesisor degradation and need not be due to inactiva-tion of the enzyme by a diffusible inhibitor, suchas methionine sulfoximine. Whether such aninhibitor is also produced by the host fern can-not be ruled out on the basis of our work. Wecannot at this point state the exact means bywhich the regulation of GS protein levels iseffected. If it is through diffusion of a natural orartificial repressor of GS synthesis, that com-pound would have to be incapable of serving as anitrogen source for the cyanobacterium, lest thefern suffer a net loss by providing it.
ACKNOWLEDGMENTS
This research was supported by Public Health Service grantGM 21823 from the National Institutes of Health and by grant5901-0410 from the United States Department of AgricultureScience and Education Administration through the Competi-tive Grants Office.We are grateful to Pamela Keim for devising and performing
the diaminopimelic acid determination.
LITERATURE CITED
1. Bender, R. A., K. A. Janssen, A. D. Resnick, M. Blumen-berg, F. Foor, and B. Maganlk. 1977. Biochemical pa-rameters of glutamine synthetase from Klebsiella aero-genes. J. Bacteriol. 129:1001-1009.
2. Bentley, H. R., E. E. McDermott, and J. K. Whitehead.1951. Action of nitrogen trichloride on certain proteins. II.Synthesis of methionine sulphoximine and other sulphoxi-mines. Proc. R. Soc. London Ser. B 138:265-272.
3. Chaney, A. L., and E. P. Marbach. 1962. Modified re-agents for determination of urea and ammonia. Clin.Chem. (N.Y.) 8:130-132.
4. Chock, P. B., S. G. Rhee, and E. R. Stadtman. 1980.Interconvertible enzyme cascades in cellular regulation.Annu. Rev. Biochem. 49:813-843.
5. Denel, T. F., and E. R. Stadtman. 1970. Some kineticproperties of Bacillus subtilis glutamine synthetase. J.Biol. Chem. 245:5206-5213.
6. Dllworth, M. 1966. Acetylene reduction by nitrogen fixingpreparations from Clostridium pasteurianum. Biochim.Biophys. Acta 127:285-294.
7. Fay, P. 1969. Cell differentiation and pigment compositionin Anabaena cylindrica. Arch. Mikrobiol. 67:62-70.
8. Fieser, L., and M. Fleser. 1967. Reagents for organic
J. BACTERIOL.
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

REGULATION AND SYNTHESIS OF GS IN ANABAENA
synthesis, vol. 1. John Wiley & Sons, Inc., New York.9. Fleming, H., and R. Haselkorn. 1973. Differentiation in
Nostoc muscorum: nitrogenase is synthesized in hetero-cysts. Proc. Natl. Acad. Sci. U.S.A. 70:2727-2731.
10. Garvey, J., N. Cremer, and D. Sumsdorf. 1974. Methods inimmunology, 3rd ed. W. A. Benjamin, Reading, Mass.
11. laselkorn, R. 1978. Heterocysts. Annu. Rev. Plant Phy-siol. 29:319-344.
12. Hill, D. J. 1975. The pattern of development ofAnabaenain the Azolla-Anabaena symbiosis. Planta 122:179-184.
13. Johan, B. C., and H. Gest. 1977. Adenylylation/deadenylylation control of the glutamine synthetase ofRhodopseudomonas capsulata. Eur. J. Biochem. 81:365-371.
14. Jonsson, S., and G. Kronvall. 1974. The use of protein A-containing Staphylococcus aureus as a solid phase anti-IgG reagent in radioimmunoassays as exemplified in thequantitation of a-fetoprotein in normal human adult se-rum. Eur. J. Immunol. 4:29-33.
15. Kesler, S. W. 1975. Rapid isolation of antigens from cellswith a staphylococcal protein A-antibody absorbent: pa-rameters of the interaction of antibody-antigen complexeswith protein A. J. Immunol. 115:1617-1624.
16. Kustu, S., D. Burton, E. Ga, L. McCarter, andN. McFarland. 1979. Nitrogen control in Salmonella: reg-ulation by the gInR and glnF gene products. Proc. Natl.Acad. Sci. U.S.A. 76:4576-4580.
17. Lufttg, R., and R. Haselkorn. 1967. Morphology ofa virusof blue-green algae and properties of its deoxyribonucleicacid. J. Virol. 1:344-361.
18. Nelson, A., R. Rlp, and R. Kunlsawa. 1971. Hetero-cyst formation and nitrogenase synthesis in Anabaenasp.: a kinetic study. Arch. Mikrobiol. 76:139-150.
19. Newton, J. W., and A. I. Herman. 1979. Isolation ofcyanobacteria from the aquatic fern, Azolla. Arch. Micro-biol. 120:161-165.
20. Orr, J., and R. Huelkorn. 1981. Kinetic and inhibitionstudies of glutamine synthetase from the cyanobacteriumAnabaena 7120. J. Biol. Chem. 256:13099-13104.
21. Orr, J., L. M. Keefer, P. KeIm, T. D. Nguyen, T. Wei-lems, R. L. Hdnrison, and R. Hen. 1981. Purifica-tion, physical characterization, and NHrterminal se-quence of glutamine synthetase from the cyanobacteriumAnabaena 7120. J. Biol. Chem. 256:13091-13098.
22. Pan, F. L., and J. G. Coote. 1979. Glutamine synthetaseand glutamate synthetase activities during growth andsporulation in Bacillus subtilis. J. Gen. Microbiol.112:373-377.
23. Pete, G., and B. Mayne. 1974. The Azolla-Anabaenaazollae relationship. I. Initial characterization of the asso-
ciation. Plant Physiol. 53:813-819.24. Ray, T. B., G. A. Peters, R. E. Toia, Jr., and
B. C. Mayne. 1978. Azolla-Anabaena relationship. VII.Distribution of ammonia assimilating enzymes, protein,and chlorophyll between host and symbiont. Plant Phy-siol. 62:463-467.
25. Rebello, J. L., and N. Strauss. 1969. Regulation of synthe-sis of glutamine synthetase in Bacillus subtilis. J. Bacter-iol. 98:683-688.
26. Rowel, P., S. Entlcott, and W. D. P. Stewart. 1977. Gluta-mine synthetase and nitrogenase activity in the blue-greenalga Anabaena cylindrica. New Phytol. 79:41-54.
27. Scoeilhorn, R., ad R. Burrs. 1967. Reduction of azideby the N2-fixing enzyme system. Proc. Natl. Acad. Sci.U.S.A. 57:1317-1323.
28. Stacey, G., C. Van Baalen, and F. R. Tabita. 1979. Nitro-gen and ammonia assimilation in the cyanobacteria: regu-lation of glutamine synthetase. Arch. Biochem. Biophys.194:457-467.
29. Stadtnan, E. R., and A. Ginsburg. 1974. The glutaminesynthetase of Escherichia coli: structure and control, p.755-807. In P. Boyer (ed.), The enzymes, vol. 10, 3rd. ed.Academic Press, Inc., New York.
30. Stewart, W. D. P., and P. Rowell. 1975. Effects of L-methionine-D,L-sulphoximine on the assimilation of newlyfixed NH3, acetylene reduction and heterocyst productionin Anabaena cylindrica. Biochem. Biophys. Res. Com-mun. 65:846-856.
31. Tull, R., and J. Thomas. 1981. In vivo regulation ofglutamine synthetase by ammonium in the cyanobacter-ium Anabaena L-31. Arch. Biochem. Biophys. 206:181-189.
32. Van Gorkom, H. J., and M. Donze. 1971. Localization ofnitrogen fixation in Anabaena. Nature (London) 234:231-232.
33. Weeke, B. 1973. General remarks on principles, equip-ment, reagents and procedures, p. 15-35. In N. Axelsen,J. Kroll, and B. Weeke (ed.), A manual of quantitativeimmunoelectrophoresis. Universitetsforlaget, Oslo.
34. Wood, N. B., and R. Haselkorn. 1979. Proteinase activityduring heterocyst differentiation in nitrogen-fixing cyano-bacteria, p. 159-166. In G. N. Cohen and H. Holzer (ed.),Limited proteolysis in microorganisms. U. S. Departmentof Health, Education and Welfare Publication (NIH) 79-1591. U. S. Government Printing Office, Washington,D.C.
35. Woolfolk, C. A., B. Shapiro, and E. R. Stadtman. 1966.Regulation of glutamine synthetase. I. Purification andproperties of glutamine synthetase from Escherichia coli.Arch. Biochem. Biophys. 116:177-192.
635VOL. 152, 1982
on July 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























