Embed Size (px)
Citation preview

The multifunctional choroid
Debora L. Nickla a,*, Josh Wallman b
aDepartment of Biosciences, New England College of Optometry, 424 Beacon St., Boston, MA 02115, USAbDepartment of Biology, City College of New York, CUNY 160 Convent Ave., New York, N.Y. 10031, USA
Keywords:MyopiaDefocusEmmetropizationNitric oxideGrowth factorsIntrinsic choroidal neurons
a b s t r a c t
The choroid of the eye is primarily a vascular structure supplying the outer retina. It has several unusualfeatures: It contains large membrane-lined lacunae, which, at least in birds, function as part of thelymphatic drainage of the eye and which can change their volume dramatically, thereby changing thethickness of the choroid as much as four-fold over a few days (much less in primates). It contains non-vascular smooth muscle cells, especially behind the fovea, the contraction of which may thin the choroid,thereby opposing the thickening caused by expansion of the lacunae. It has intrinsic choroidal neurons,also mostly behind the central retina, which may control these muscles and may modulate choroidalblood flow as well. These neurons receive sympathetic, parasympathetic and nitrergic innervation.
The choroid has several functions: Its vasculature is the major supply for the outer retina; impairmentof the flow of oxygen from choroid to retina may cause Age-Related Macular Degeneration. The choroidalblood flow, which is as great as in any other organ, may also cool and warm the retina. In addition to itsvascular functions, the choroid contains secretory cells, probably involved in modulation of vasculari-zation and in growth of the sclera. Finally, the dramatic changes in choroidal thickness move the retinaforward and back, bringing the photoreceptors into the plane of focus, a function demonstrated by thethinning of the choroid that occurs when the focal plane is moved back by the wearing of negative lenses,and, conversely, by the thickening that occurs when positive lenses are worn.
In addition to focusing the eye, more slowly than accommodation and more quickly than emmetrop-ization, we argue that the choroidal thickness changes also are correlated with changes in the growth ofthe sclera, and hence of the eye. Because transient increases in choroidal thickness are followed bya prolonged decrease in synthesis of extracellular matrix molecules and a slowing of ocular elongation,and attempts to decouple the choroidal and scleral changes have largely failed, it seems that the thick-ening of the choroid may be mechanistically linked to the scleral synthesis of macromolecules, and thusmay play an important role in the homeostatic control of eye growth, and, consequently, in the etiology ofmyopia and hyperopia.
! 2009 Elsevier Ltd. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .1452. Structure and “classical” functions of the choroid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .145
2.1. Histology of the choroid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1462.1.1. Choriocapillaris . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1462.1.2. Choroidal vascular layers and suprachoroid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 146
2.2. Innervation of the choroid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1472.2.1. Parasympathetic innervation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1472.2.2. Sympathetic innervation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1492.2.3. Sensory innervation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 149
2.3. Special characteristic features of the choroid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1492.3.1. Intrinsic choroidal neurons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 149
* Corresponding author. Tel.: þ1 617 266 2030x5314.E-mail address: [email protected] (D.L. Nickla).
Contents lists available at ScienceDirect
Progress in Retinal and Eye Research
journal homepage: www.elsevier .com/locate/prer
1350-9462/$ e see front matter ! 2009 Elsevier Ltd. All rights reserved.doi:10.1016/j.preteyeres.2009.12.002
Progress in Retinal and Eye Research 29 (2010) 144e168

2.3.2. Non-vascular smooth muscle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1502.3.2.1. Are non-vascular smooth muscle cells myofibroblasts? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 150
2.3.3. Fluid-filled lacunae: lymphatics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1512.4. Choroidal blood flow: nourishment of the retina . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1512.5. Does the choroidal blood flow exhibit autoregulation? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1522.6. Choroidal blood flow: thermoregulation of the retina? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1522.7. Choroid and pathology: age-related macular degeneration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 153
3. Modulation of choroidal thickness . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .1543.1. Possible mechanisms underlying choroidal thickness changes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154
3.1.1. Changes in the synthesis of osmotically active molecules . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1553.1.2. Changes in vascular permeability . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1553.1.3. Increases in fluid flux from the anterior chamber to the choroid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1553.1.4. Movement of fluid across the RPE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1553.1.5. Changes in the tonus of non-vascular smooth muscle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 156
4. The choroid and emmetropization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .1564.1. Choroidal roles in controlling ocular elongation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157
4.1.1. Does the size of the choroid determine the size of the eye or vice versa? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1574.1.2. Are the choroidal and scleral responses independent? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158
4.1.2.1. Evidence for independent responses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1584.1.2.2. Evidence for choroidal thickness modulating ocular elongation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158
4.1.2.2.1. The enduring effects of brief episodes of myopic defocus and transient choroidal thickening . . . . . . . . . . . . . . . 1584.1.2.2.2. Pharmacological treatments affecting both choroidal thickening and ocular elongation . . . . . . . . . . . . . . . . . . . 159
4.1.3. The choroid as a secretory tissue: evidence for a direct role in ocular growth . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1594.2. Pharmacological effects on choroidal thickness and ocular elongation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 160
4.2.1. Dopamine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1604.2.1.1. Evidence for a role in emmetropization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1604.2.1.2. Effects on choroidal thickness and ocular elongation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161
4.2.2. Acetylcholine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1614.2.2.1. Evidence for a role in emmetropization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1614.2.2.2. Effects on choroidal thickness and ocular elongation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161
4.2.3. Nitric oxide . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1624.2.3.1. Evidence for a role in emmetropization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1624.2.3.2. Effects on choroidal thickness and ocular elongation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162
4.3. Implications of these pharmacological effects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1635. The diurnal rhythm in choroidal thickness and ocular growth . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .1636. Significance to human refractive disorders . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .1647. Future directions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .164
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165
1. Introduction
The choroid does not appear to be a mysterious tissue. It consistsmostly of blood vessels, it supplies the outer retina, and choroidaldefects cause degenerative changes and neovascularization.However, it is becoming increasingly evident that the choroid has atleast three other functions: thermoregulation, adjustment of theposition of the retina by changes in choroidal thickness, and secre-tion of growth factors. The last of these is likely to play an importantrole in emmetropizationethe adjustment of eye shape duringgrowth to correctmyopia or hyperopia.What remainsmysterious, atpresent, are the mechanisms behind the changes in choroidalthickness, the nature of its secretory functions and the relationshipbetween these two processes.
In this review, we will summarize the anatomy, histology,innervation and functions of the choroid, discuss the control ofchoroidal thickness by visual signals, show evidence for a secretoryrole for the choroid and speculate on the relationship betweenchanges in choroidal thickness and ocular elongation, and therefore,emmetropization.
2. Structure and “classical” functions of the choroid
The choroid is the posterior part of the uvea, the middle tunic ofthe eye (Fig. 1). The uvea develops from the mesenchyme
surrounding the two vesicles that bud off the embryonic forebrain atthe end of the first month in humans, eventually becoming the eyes.At about that time, melanocyte precursors migrate into the uveafrom the neural crest; these do not differentiate into pigmentedmelanocytes until 7e8 months of gestation. The mesenchyme thatforms the choriocapillaris at about 2monthsmust be in contact withthe developing retinal pigment epithelium (RPE) in order to differ-entiate. Therefore, the choroid derives from different cell lines thando the retina and RPE, which both derive from the neural ectoderm.The choroid is comprised of blood vessels, melanocytes, fibroblasts,
Fig. 1. Photomicrograph of the three tunics at the back of the primate eye. From:Remington, LA; Clinical Anatomy of the Visual System; 2nd Edition; 2005. Reproducedwith permission ! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 145

resident immunocompetent cells and supporting collagenous andelastic connective tissue. As one of the most highly vascularizedtissues of the body, its main function has been traditionally viewedas supplying oxygen and nutrients to the outer retina, and, in specieswith avascular retinas, to the inner retina as well. Other likelyfunctions include light absorption (in species with pigmentedchoroids), thermoregulation via heat dissipation, and modulation ofintraocular pressure (IOP) via vasomotor control of blood flow. Thechoroid also plays an important role in the drainage of the aqueoushumor from the anterior chamber, via the uveoscleral pathway.This pathway is responsible for approximately 35% of the drainagein humans, a higher percentage, between 40 and 60%, in non-humanprimates, and a much lower percentage in the cat (about 3%) andrabbit (3e8%) (Alm and Nilsson, 2009).
2.1. Histology of the choroid
The choroid extends from the margins of the optic nerve to thepars plana, where it continues anteriorly, becoming the ciliary body.Its innermost layer is the complex 5-laminar structure of Bruch'smembrane, and its outermost one is the suprachoroid outside ofwhich is the suprachoroidal space between choroid and sclera.
Histologically, the choroid has variously been divided into 4e6layers, depending onwhether the vascular region is considered as 1or 2 layers (Sattler's and Haller's), and whether the lamina fusca isconsidered to be of scleral or choroidal origin. It is most commonlydescribed as five layers: starting from the retinal (inner) side, theseare Bruch's membrane, the choroiocapillaris, the two vascularlayers (Haller's and Sattler's), and the suprachoroidea (Fig. 2Aand B) (Hogan et al., 1971).
In humans, the choroid is approximately 200 mm thick at birthand decreases to about 80 mm by age 90 (Ramrattan et al., 1994).In non-human primates, the choroid is thinner, approximately95 mm at the fovea, thinning to 55 mm at the periphery (Krebs andKrebs, 1991). In chickens, it is about 250 mm thick in the centerunder normal visual conditions (Wallman et al., 1995; Nickla et al.,1998), thinning to about 100 mm in the periphery (Wallman et al.,1995). Large melanocytes are abundant in the primate choroid,imparting a dark pigmentation (Krebs and Krebs, 1991). In otherspecies, such as birds, only sparse melanocytes are present in theouter choroid, resulting in lighter pigmentation (De Stefano and
Mugnaini, 1997) (although there is variability in pigmentationamong avian groups).
2.1.1. ChoriocapillarisThe choriocapillaris is a highly anastomosed network of capil-
laries, forming a thin sheet apposed to Bruch's membrane (Fig. 3A).The fibrous basement membrane of the capillary endothelialcells forms the outermost layer of Bruch's membrane in humans.The choriocapillaris is about 10 mm thick at the fovea, where there isthe greatest density of capillaries, thinning to about 7 mm in theperiphery. The capillaries arise from the arterioles in Sattler's layer,each of which gives rise to a hexagonal (or lobular)-shaped domainof a single layer of capillaries, giving a patch-like structure to thechoriocapillaris (Fig. 3B). The capillaries are fenestrated and, inhumans, are relatively large in diameter (20e40 mm), althoughpossessing passages narrow enough to hinder the flow of 9 mmspheres (Bill et al., 1983). Because of the large area of the chorio-capillaris, the velocity of red blood cells here is only about 77%that of the velocity of red blood cells in the retinal capillaries(Wajer et al., 2000). In birds, the capillaries are polarized, with thefenestrations almost exclusively facing the retina (De Stefano andMugnaini, 1997); in primates, this is mainly true, although there area few fenestrations on the outer walls (McLeod et al., 2009). Thefenestrated capillaries are highly permeable to proteins, contrib-uting to the high oncotic pressure in the extravascular stroma,which fosters themovement of fluids from the retina to the choroid.
Adjacent to Sattler's layer at the outer choriocapillaris there isa fibrous layer which is attached to the outer fibrous layer of Bruch'smembrane by columns of collagen fibers running between thecapillaries; these columns (or “pillars”) may function to keep thecapillary diameters constant (at least in primates) (Krebs and Krebs,1991). Finally, the innermost choroidal layer is Bruch's membrane,a 5-layered structure consisting of (from outer to inner): basementmembrane of the choroiocapillaris, outer collagenous zone, elasticlayer, inner collagenous zone, and basement membrane of .theretinal pigment epithelium.
2.1.2. Choroidal vascular layers and suprachoroidThe vascular region of the choroid consists of the outer Haller's
layer of large blood vessels and the inner Sattler's layer of mediumand small arteries and arterioles that feed the capillary network, andveins (Fig. 2). The stroma (extravascular tissue) contains collagen
Fig. 2. Histology of the choroid. A. Schematic of the layers of the choroid. Reproduced with permission from Remington, LA (2005) Clinical Anatomy of the Visual System ! Elsevier.B. Semithin resin section of the outer retina and choroid in the primate eye. RPE: retinal pigment epithelium; CC: choriocapillaris; SL: Sattler’s layer; HL: Haller’s layer. Reproducedwith permission from Forrester et al., 2002 The Eye: Basic Sciences in Practice ! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168146

and elastic fibers, fibroblasts, non-vascular smooth muscle cellsand numerous very large melanocytes that are closely-apposed tothe blood vessels. As in other types of connective tissue, there arenumerous mast cells, macrophages, and lymphocytes.
The suprachoroid is a transitional zone between choroid andsclera containing elements of bothe collagen fibers, fibroblasts andmelanocytes. In primates (Krebs and Krebs, 1988, 1991), birds(De Stefano and Mugnaini, 1997; Junghans et al., 1998; Liang et al.,2004), and rabbits (Gomez et al., 1988), this layer contains large,endothelium-lined spaces (referred to as “lacunae” in the avianliterature), which empty into veins (Fine and Yanoff, 1979; Krebsand Krebs,1988; De Stefano andMugnaini,1997). These presumablylymphatic structureswill be discussed inmore detail under “Specialcharacteristic features of the choroid.”
The lamina fusca is the outermost layer of the suprachoroid(Fig. 4A), approximately 30 mm thick, consisting of several layersof closely-apposed, flattened fusiform melanocytes and fibroblast-like cells (Fig. 4B), with interspersed bundles of myelinated axons(De Stefano and Mugnaini, 1997).1 The fibroblast-like cells formsheaths, or plates, that are connected to one another by adheringand occluding junctions (Krebs and Krebs, 1988). These cellssynthesize and secrete the usual extracellular matrix componentselastin, collagens and proteoglycans. The function of the melano-cytes, apart from providing pigmentation, is unknown. However,
their membranes contain endothelin-B receptors, which maymediate the endothelin 1 e induced influx of calcium ions (Hiroseet al., 2007), the functional implications of which are also notknown.
2.2. Innervation of the choroid
The smoothmuscle of the vesselwalls of the choroid, like those ofskeletal and cardiac muscle blood vessels, are innervated byboth divisions of the autonomic nervous system, which form denseplexuses of fibers around the vessels (“perivascular plexus”). Axonterminals are also found throughout the stroma, terminating onnon-vascular smoothmuscle, intrinsic choroidal neurons (ICNs), andpossibly other cell types. There are also primary afferent sensoryfibers that project to the trigeminal ganglion via the ophthalmicnerve; some of these give rise to peptide-positive collaterals (Mayet al., 2004) that terminate on and around the vessels and intrinsicchoroidal neurons (Schrödl et al., 2001a; Schrödl et al., 2003).
2.2.1. Parasympathetic innervationIn mammals, the main parasympathetic input to the choroid
originates from the pterygopalatine ganglion (Ruskell,1971; Uddmanet al., 1980; Bill, 1985, 1991; Stone, 1986; Stone et al., 1987), locatedwithin the pterygopalatine fossa or its equivalent. These fibers arepredominantly cholinergic and are rich in the vasodilators vasoactiveintestinal polypeptide (VIP) and nitric oxide (NO) (Uddman et al.,1980; Stone et al., 1987; Yamamoto et al., 1993a,b; Alm et al., 1995)(Fig. 5A and B). Although there is some physiological evidence for
Fig. 3. Histology of the choriocapillaris. A. The choriocapillaris is located adjacent to Bruch's membrane. Reproduced with permission from Remington, LA (2005) Clinical Anatomyof the Visual System ! Elsevier. B. Each feeder arteriole in Sattler's layer supplies a hexagonally-arrayed area of capillaries. From: Forrester et al., 2002 “The Eye: Basic sciences inpractice”. Reproduced with permission ! Elsevier.
1 Some anatomists identify the lamina fusca as the innermost layer of the sclera(Forrester et al., 2002; Watson and Young, 2004).
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 147
innervation from the ciliary ganglion (Bill et al., 1976; Stjernschantzet al., 1976; Gherezghiher et al., 1990; Nakanome et al.,1995), this hasnot been documented anatomically.
In birds, on the other hand, the main parasympathetic inner-vation originates from a well-defined region of the ciliary ganglion,which receives input from the medial aspect of the Edinger-West-phal nucleus (Reiner et al., 1983, 1991; Fitzgerald et al., 1990;
Fitzgerald et al., 1996). These ciliary “choroid” neurons releasesomatostatin (Gray et al., 1989; Epstein et al., 1988; De Stefano et al.,1993; Schrödl et al., 2006) in addition to acetylcholine (Merineyand Pilar, 1987; Reiner et al., 1991; Cuthbertson et al., 1996) andnitric oxide (Sun et al., 1994). The other source of parasympatheticinnervation is from the pterygopalatine ganglia, which in birds arean interconnected series of microganglia located adjacent to
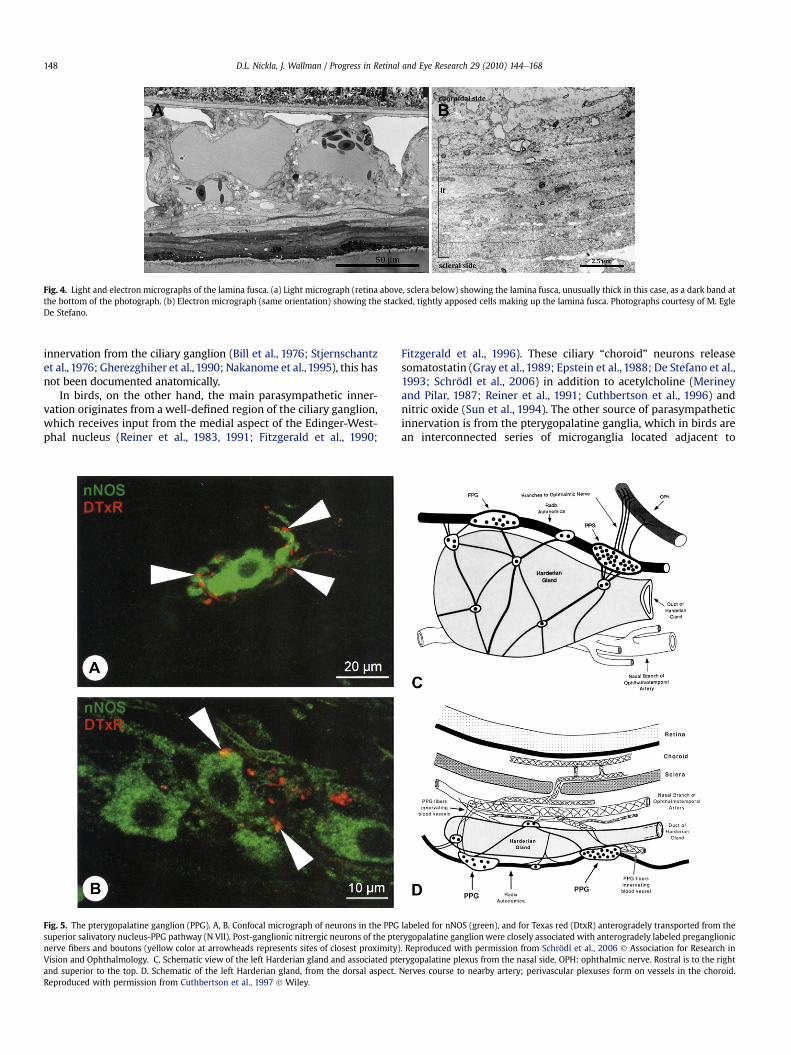
Fig. 5. The pterygopalatine ganglion (PPG). A, B. Confocal micrograph of neurons in the PPG labeled for nNOS (green), and for Texas red (DtxR) anterogradely transported from thesuperior salivatory nucleus-PPG pathway (N VII). Post-ganglionic nitrergic neurons of the pterygopalatine ganglion were closely associated with anterogradely labeled preganglionicnerve fibers and boutons (yellow color at arrowheads represents sites of closest proximity). Reproduced with permission from Schrödl et al., 2006 ! Association for Research inVision and Ophthalmology. C. Schematic view of the left Harderian gland and associated pterygopalatine plexus from the nasal side. OPH: ophthalmic nerve. Rostral is to the rightand superior to the top. D. Schematic of the left Harderian gland, from the dorsal aspect. Nerves course to nearby artery; perivascular plexuses form on vessels in the choroid.Reproduced with permission from Cuthbertson et al., 1997 ! Wiley.
Fig. 4. Light and electron micrographs of the lamina fusca. (a) Light micrograph (retina above, sclera below) showing the lamina fusca, unusually thick in this case, as a dark band atthe bottom of the photograph. (b) Electron micrograph (same orientation) showing the stacked, tightly apposed cells making up the lamina fusca. Photographs courtesy of M. EgleDe Stefano.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168148

the Harderian gland on the nasal side of the orbit (Baumel, 1975;Nickel et al., 1977; Gienc and Kuder, 1985) (Fig. 5C). Two majormicroganglia are located along the superior aspect of the Harderiangland, the larger of these more rostrally (Cuthbertson et al., 1997).There are also numerous small microganglia, distributed along theupper, medial and lateral aspects of the gland. The preganglionicinput to the pterygopalatine is called the radix autonomica (N VII).The pterygopalatine neurons are positive for VIP (Walcott et al.,1989; Cuthbertson et al., 1997), nitric oxide and choline acetyl-transferase (ChAT) (Cuthbertson et al., 1997; Schrödl et al., 2006). Inall species studied, parasympathetic fibers terminate on vessels inperivascular plexuses (Fig. 5D), andmediate increases in blood flowby vasodilation. These fibers also terminate on non-vascularsmooth muscle cells and intrinsic choroidal neurons.
2.2.2. Sympathetic innervationThe sympathetic innervation of the choroid comes from the
superior cervical ganglion (Kirby et al., 1978; Guglielmone and Can-tino,1982; Bill, 1985). These noradrenergic neurons terminate on theblood vessels and mediate vasoconstriction. In birds, the sympa-thetics also innervate non-vascular smoothmuscle (Guglielmone andCantino, 1982; Poukens et al., 1998) and intrinsic choroidal neurons(Schrödl et al., 2001b), the same targets as the parasympatheticsystem. Schrödl et al. (2001a,b) have speculated that the intrinsicchoroidal neurons may act as intermediaries in the sympatheticsystem between the post-ganglionic neurons and the muscle, andincrease smooth muscle tone, although the effects of noradrenalineon the ICNs and non-vascular smooth muscle are unknown.
2.2.3. Sensory innervationMany organs, including the eye, have been shown to use
peptides such as substance-P and calcitonin-gene-related peptidein a pre-central reflex arc, or axon reflex, a non-synaptic response inwhich a local stimulus (chemical or mechanical) depolarizesa sensory terminal which travels to the nearest collateral (branch),
releasing the peptide onto the effector tissue (reviews: Holzer,1988; Bill, 1991). Evidence for this reflex has been found in theprimary sensory afferents from the trigeminal ganglion in the uveaand choroid, which use both peptides; the reflex may mediatechanges in blood flow or a variety of other functions. For instance,in both mammals and birds, sensory fibers projecting to thetrigeminal ganglion from the choroid via the ophthalmic branch ofthe trigeminal nerve elicit vasodilation (Shih et al., 1999b). Theseterminals are positive for substance-P and calcitonin-gene-relatedpeptide (Stone, 1985; Reiner, 1987; Stone andMcGlinn,1988); othervasoactive peptides such as cholecystokinin are also found,differing between species (Bill, 1991). Recent work has shown thatthese primary afferents also contact the intrinsic choroidal neurons,which are believed to be analogous to the intrinsic neurons in theenteric nervous system which function as local signal integratorsresponding to local mechanical, thermal or chemical stimuli(Schrödl et al., 2001a).
2.3. Special characteristic features of the choroid
2.3.1. Intrinsic choroidal neuronsFirst described from choroids of higher primates including man
(Mueller, 1859), intrinsic choroidal neurons are present in primates(humans and monkeys: Flugel et al., 1994; May et al., 1997; Berguaet al., 2003; Schrödl et al., 2003; May et al., 2004), tree shrews:(Flugel et al., 1994) and birds (Bergua et al., 1996, 1998; Schrödlet al., 2004; review: Schrödl, 2009), but are scarce or absent in rats,rabbits and cats (Flugel et al., 1994). In humans, ICNs are especiallynumerous: choroids have approximately 2000 per eye, largelyconcentrated in the temporal and central regions, decreasingtowards the periphery. In other primates, the numbers are lower,about 500 per eye, and are more evenly distributed than those ofhumans. In birds, the numbers vary, from fewer than 1000 in quailand ducks, to about 6000 in geese (Schrödl et al., 2004). Theseneurons are usually small (between 20 and 40 mm) and multipolar.
Fig. 6. The intrinsic neurons of the human choroid immunolabeled for nNOS (left) and vasoactive intestinal polypeptide (VIP) (right). Left: Nitric oxide synthase labeling in thecytoplasm of ICNs (arrows) and weak staining in the axons of these neurons (arrowheads). Right: Staining for antibodies against VIP in cytoplasm (arrow) and axon (arrowhead).Approximately 200X magnification. Reproduced with permission from Flugel et al., 1994 ! Association for Research in Vision and Ophthalmology.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 149

In all species, most ICNs are positive for NADPH-diaphorase and/or neuronal nitric oxide synthase (nNOS) (Flugel et al., 1994; Berguaet al., 1996, 1998; Cuthbertson et al., 1997), indicating that they usenitric oxide as a transmitter (Fig. 6A). Many are also positive for VIP(Miller et al., 1983; Flugel et al., 1994; Cuthbertson et al., 1997)(Fig. 6B). In humans, about half the population are positive forcalretinin (May et al., 2004). There is evidence that these neuronsreceive both sympathetic and parasympathetic innervation:They are contacted by boutons that are positive for both tyrosinehydroxylase and dopamine-B-hydroxylase, which in combinationare a marker for post-ganglionic sympathetic neurons or collaterals;in addition they are contacted by boutons that are positive for nNOSand VIP, indicating inter-ICN connectivity and/or input from thepterygopalatine ganglia.
Now, 150 years after their discovery, the functions of the ICNsremain unknown. They probably play a role in blood flow regulationbecause they terminate on themuscle walls of arteries (Meriney andPilar, 1987; Flugel-Koch et al., 1996) and because they release NO, apotent vasodilator. Furthermore, because they are found adjacentto the non-vascular smooth muscle that span the stroma andsuprachoroid around the large lymphatic lacunae andmay innervatethese muscles, it is possible that these cells are involved in changingthe choroidal thickness in response to retinal defocus (Poukens et al.,1998), a topic which will be further discussed.
2.3.2. Non-vascular smooth muscleNon-vascular smooth muscle cells were first described in
human choroids by Mueller (1859). Subsequent work has foundthem in the choroids of birds (Kajikawa, 1923; Walls, 1942;Guglielmone and Cantino, 1982; Meriney and Pilar, 1987; Wallmanet al., 1995), in which they are found in the stroma. These havebeen hypothesized to play a role in the changes in choroidalthickness in response to retinal defocus (Wallman et al., 1995).
Inmammals, non-vascular smoothmuscle cells have been found inprimates (Krebs and Krebs, 1991; Flugel-Koch et al., 1996; Poukenset al.,1998) and rabbits (Haddad et al., 2001), but are absent indog, cat,rat, tree shrew, pig and cow (May, 2003). Among primates, thenumber of smooth muscle cells are higher in species with well-defined foveas (May, 2003). These non-vascular smooth muscle cellsare sometimes referred to as myofibroblasts, and are positive fora-actin. In primates, they are found in two populations: in
the suprachoroid, forming a reticulum of flattened lamellae, and ina single layer immediately beneath and parallel to Bruch'smembrane.In the suprachoroid, the lamellae run obliquely to the choroidalsurface between “spaces”, presumably lacunae (Fig. 7A) (Poukenset al., 1998). Non-vascular smooth muscle cells are concentratedbeneath the fovea and are sparcer anteriorly (although in a few casesconnections with ciliary muscle fibers were seen). In the rabbit, theyare arranged in bundles of up to 20 cell layers, running along thelongitudinal axis of blood vessels (Haddad et al., 2001), possiblyindicating a role in the choroidal blood flow autoregulation shown inthis species (Kiel and Shepherd, 1992). Neighboring non-vascularsmooth muscle cells are connected by zonula adherens, forminga syncytium,with their organelles clustered around thenucleus. Thesecells were identified as regular smooth muscle, as opposed to myofi-broblasts (Haddadet al., 2001),which typically appear as isolated cells,do not have junctional complexes, nor are their organelles clusterednear the nucleus. In birds, non-vascular smoothmuscle cells are foundin both the suprachoroid and the vascular layer (Wallman et al., 1995;Fig. 7B). The arrangement of the non-vascular smoothmuscle obliqueor tangential to the surface, between the lacunae, makes it plausiblethat they might play a role in the changes in choroidal thickness inresponse to retinal defocus, to be discussed later.
The predominant sub-foveal localization has suggested to Flu-gel-Koch et al. (1996) that these cells may stabilize the positionof the fovea against movement caused by the contracting ciliarymuscle during accommodation. It is not certain whether thesecontractile cells are controlled by the autonomic nervous system,but the presence of nerve terminals positive for NADPH-diaphoraseor tyrosine hydoxylase in close proximity suggests inputs from bothparasympathetic nitrergic and sympathetic adrenergic systems(Poukens et al., 1998).2
2.3.2.1. Are non-vascular smooth muscle cells myofibroblasts? Non-vascular smooth muscle is prominent in the human uvea, includingthe iris constrictor and the ciliary muscles. The iris dilator, however,
Fig. 7. Non-vascular smooth muscle of the primate (A) and chick (B) eyes. A. Suprachoroid and inner sclera contain cells positive for smooth muscle a-actin (blue chromogencounterstained with nuclear fast red). Reproduced with permission from Poukens et al., 1998 ! Association for Research in Vision and Ophthalmology. B. Electron micrograph ofchoroidal cells labeled with antibodies to smooth muscle actin (black dots), showing long fibers not associated with blood vessels. M, muscle cell; F, fibroblast; C, collagen.Reproduced with permission from Wallman et al., 1995 ! Association for Research in Vision and Ophthalmology.
2 In fact, these authors noted that the arrangement of the reticulum of the non-vascular smooth muscle and the adjacent open spaces is reminiscent of the lacunaein the suprachoroid of the bird, and hence might mediate similar functions, notably,that of regulating fluid flux between the vasculature and the extracellular matrix.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168150

is not typical smooth muscle, but is classified as myofibroblast,a cell type intermediate between smooth muscle and fibroblasts,and not widely present in the body except around the lumena ofglands. One distinguishing characteristic is the absence of theintermediate filament desmin found in “regular” smooth muscle(Flugel-Koch et al., 1996).
In the Poukens et al. (1998) study of primate choroids, the non-vascular smooth muscle cells were identified as predominantlymyofibroblasts. These were arranged in flattened lamellae in thesuprachoroid, oblique to the choroidal surface, between whichwere fluid-filled lacunae. In the inner part of the choroid they ranparallel to Bruch's membrane in the sub-foveal inner chorioca-pillaris. Similarly, Flugel-Koch et al. (1996) describe a network ofa-actin-positive but desmin-negative, spindle-to star-shapedmyofibroblasts in the suprachoroid, which were connected to anelastic fiber net of the stroma and to the adventitia of the vessels.Again, the arrangement was densest in the submacular region.
In contrast, an immunohistochemical study of 42 human eyes ofa wide range of ages described all of the non-vascular smoothmuscle cells as typical smooth muscle (May, 2005), based on theexpression in these of the protein smoothelin, which is notexpressed in myofibroblasts (van der Loop et al., 1996; Bar et al.,2002). The cells also stain for a-myosin, a-actin and caldesmon,markers for contractibility. The smooth muscles were located inthree distinct subgroups: (1) in the suprachoroid and sclera form-ing a semi-circle around the entering posterior ciliary arteries, (2)in the outer vascular choroid (Haller's) between large blood vesselsin the posterior segment, and (3) in the suprachoroid of the fovealregion of the temporal quadrant, where they form dense plaques.Interestingly, this third group showed the greatest inter-individualvariation, being extremely numerous in some people but absent inothers. It was speculated that the location and variability of thispopulation may indicate a visual function, as opposed to a vascularone, although without corroborating evidence (May, 2005).
2.3.3. Fluid-filled lacunae: lymphaticsLarge, fluid-filled lacunae in the choroid were first observed
over 50 years ago by Walls (1942). In 1987, Meriney & Pilardescribed fluid-filled lacunae in the stroma of chicken choroids thatwere connected to arterioles, and suggested that they functioned asa drainage reservoir involved in regulating IOP (Meriney and Pilar,1987). Although the authors did not distinguish between thestroma and the suprachoroid, the photomicrographs show them inthe suprachoroid as well. In 1997, a careful and thorough EMinvestigation of the chicken choroid showed that very large,endothelium-lined lacunae are present in the suprachoroid,continuing as smaller ones in the stroma of the vascular layers andending in the choriocapillaris as blind-end structures (Fig. 8;De Stefano andMugnaini,1997). These structures were identified astrue lymphatics based on (1) the endothelial lining being extremelythin, with elaborate junctional complexes, pinocytotic vesicles, andlarge fenestrations, all ultrastructural characteristics of lym-phoendothelium (De Stefano and Mugnaini, 1997) (Krebs andKrebs, 1988; Junghans et al., 1998), and (2) paracentesis (drainingthe anterior chamber) resulting in the backflow of blood cells intothese spaces, indicating a connection with the venous system (DeStefano and Mugnaini, 1997). These lymphatic spaces are sur-rounded by lamellae of elastin fibers and nerves, and very largemelanocytes.
It was presumed at the time of their discovery that theselacunae were a peculiarity of the bird eye. Indeed, the traditionalview is that the primate eye does not contain a lymphatic drainagesystem (Bill, 1975), although the conjunctiva and eyelids drain intothe submandibular and parotid lymph nodes (Wobig, 1981).However, some more recent work has described lymphatics in the
baboon and macaque choroids (Krebs and Krebs, 1988; Sugita andInokuchi, 1992; Poukens et al., 1998), which allegedly had beendismissed by earlier anatomists as artifacts. In these primates, thelymphatics are located both in the stroma, between arteries andveins, and in the suprachoroid (in the Japanese monkey (Macacafuscata) they were found in the “outer choroid” (Sugita and Ino-kuchi, 1992)). In man, one study describes these lymphatics in thesuprachoroid (Poukens et al., 1998). It should be emphasized,however, that the identification of these spaces as lymphatics inprimate choroids, especially humans, is still controversial. Forinstance, Schrödl et al. (2008) found that LYVE-1 (lymphatic vesselendothelial hyaluronan receptor-1) and podoplanin, two markersfor lymphatic endothelium, were absent in the vessel walls of thehuman choroid, although the same antibody labeled them inchoroidal macrophages. On the other hand, positive labeling forboth of these markers on vessel endothelium in the human ciliarybody supports the existence of lymphatics in the primate uvea,perhaps affecting fluid dynamics via the uveoscleral outflow (Yücelet al., 2009). Regardless of their anatomical identity, these fluidreservoirs are the subject of interest because of evidence that theymay contribute to the changes in choroidal thickness found inresponse to retinal defocus (Wallman et al., 1995; Junghans et al.,1998, 1999; Pendrak et al., 1998; Liang et al., 2004), a topic to bediscussed later.
2.4. Choroidal blood flow: nourishment of the retina
Despite the conspicuousness of the retinal blood vessels, themajor blood supply to the retina is the choroid. Because the photo-receptors are extremely metabolically active, especially in darkness,when the light-gated ion channels are open, and active transport ofions is required tomaintain ion homeostasis, over 90% of the oxygendelivered to the retina is consumed by the photoreceptors. Indarkness, ninety percent of the oxygen comes from the choroidalcirculation (Linsenmeier et al., 1981; Bill et al., 1983; Linsenmeierand Braun, 1992). To obtain this high transport of oxygen from thechoroid, despite the barriers of Bruch's membrane and the retinalpigment epithelium, requires a steep gradient of oxygen tension,which is maintained by the high blood flow in the choroid, probablythe highest of any tissue in the body per unit tissue weight, ten-foldhigher than the brain (Alm and Bill, 1973; Alm, 1992). Consequently,the oxygen tension in the choroid stays high, with an arterial/venousdifference of only 3% versus 38% for the retinal circulation.
Fig. 8. Diagram of the lymphatic system of the avian choroid. Veins (V) and arteries (A)traverse the eye wall from the sclera (S) through the suprachoroid and stroma, wherethey branch into arterioles (a) and venules (v), to the choriocapillaris where they formcapillaries (c). Large lymphatic vessels (L) are in the suprachoroid and branch intolymphatic capillaries (l) that reach the choriocapillaris. Reproduced with permissionfrom DeStefano & Mugnaini, 1997 ! Association for Research in Vision andOphthalmology.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 151
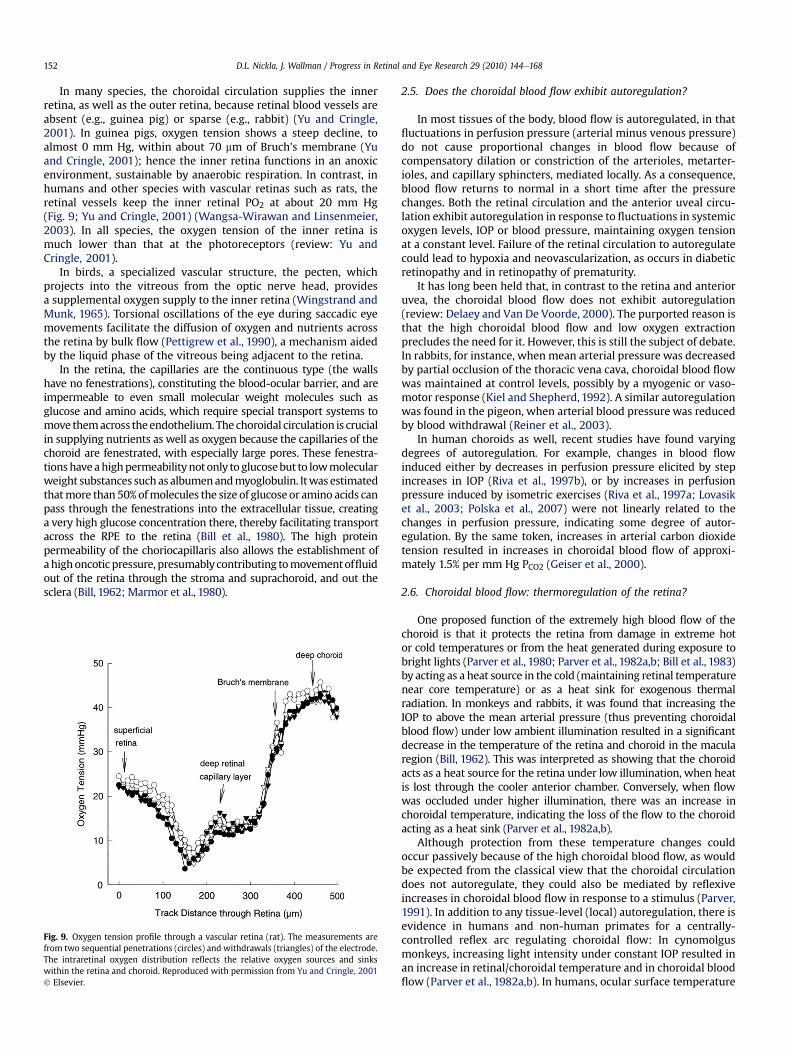
In many species, the choroidal circulation supplies the innerretina, as well as the outer retina, because retinal blood vessels areabsent (e.g., guinea pig) or sparse (e.g., rabbit) (Yu and Cringle,2001). In guinea pigs, oxygen tension shows a steep decline, toalmost 0 mm Hg, within about 70 mm of Bruch's membrane (Yuand Cringle, 2001); hence the inner retina functions in an anoxicenvironment, sustainable by anaerobic respiration. In contrast, inhumans and other species with vascular retinas such as rats, theretinal vessels keep the inner retinal PO2 at about 20 mm Hg(Fig. 9; Yu and Cringle, 2001) (Wangsa-Wirawan and Linsenmeier,2003). In all species, the oxygen tension of the inner retina ismuch lower than that at the photoreceptors (review: Yu andCringle, 2001).
In birds, a specialized vascular structure, the pecten, whichprojects into the vitreous from the optic nerve head, providesa supplemental oxygen supply to the inner retina (Wingstrand andMunk, 1965). Torsional oscillations of the eye during saccadic eyemovements facilitate the diffusion of oxygen and nutrients acrossthe retina by bulk flow (Pettigrew et al., 1990), a mechanism aidedby the liquid phase of the vitreous being adjacent to the retina.
In the retina, the capillaries are the continuous type (the wallshave no fenestrations), constituting the blood-ocular barrier, and areimpermeable to even small molecular weight molecules such asglucose and amino acids, which require special transport systems tomove themacross theendothelium. The choroidal circulation is crucialin supplying nutrients as well as oxygen because the capillaries of thechoroid are fenestrated, with especially large pores. These fenestra-tionshaveahighpermeabilitynotonly to glucosebut to lowmolecularweight substances suchas albumenandmyoglobulin. Itwas estimatedthatmore than 50%ofmolecules the size of glucoseor amino acids canpass through the fenestrations into the extracellular tissue, creatinga very high glucose concentration there, thereby facilitating transportacross the RPE to the retina (Bill et al., 1980). The high proteinpermeability of the choriocapillaris also allows the establishment ofahighoncotic pressure, presumablycontributing tomovementoffluidout of the retina through the stroma and suprachoroid, and out thesclera (Bill, 1962; Marmor et al., 1980).
2.5. Does the choroidal blood flow exhibit autoregulation?
In most tissues of the body, blood flow is autoregulated, in thatfluctuations in perfusion pressure (arterial minus venous pressure)do not cause proportional changes in blood flow because ofcompensatory dilation or constriction of the arterioles, metarter-ioles, and capillary sphincters, mediated locally. As a consequence,blood flow returns to normal in a short time after the pressurechanges. Both the retinal circulation and the anterior uveal circu-lation exhibit autoregulation in response to fluctuations in systemicoxygen levels, IOP or blood pressure, maintaining oxygen tensionat a constant level. Failure of the retinal circulation to autoregulatecould lead to hypoxia and neovascularization, as occurs in diabeticretinopathy and in retinopathy of prematurity.
It has long been held that, in contrast to the retina and anterioruvea, the choroidal blood flow does not exhibit autoregulation(review: Delaey and Van De Voorde, 2000). The purported reason isthat the high choroidal blood flow and low oxygen extractionprecludes the need for it. However, this is still the subject of debate.In rabbits, for instance, when mean arterial pressure was decreasedby partial occlusion of the thoracic vena cava, choroidal blood flowwas maintained at control levels, possibly by a myogenic or vaso-motor response (Kiel and Shepherd, 1992). A similar autoregulationwas found in the pigeon, when arterial blood pressure was reducedby blood withdrawal (Reiner et al., 2003).
In human choroids as well, recent studies have found varyingdegrees of autoregulation. For example, changes in blood flowinduced either by decreases in perfusion pressure elicited by stepincreases in IOP (Riva et al., 1997b), or by increases in perfusionpressure induced by isometric exercises (Riva et al., 1997a; Lovasiket al., 2003; Polska et al., 2007) were not linearly related to thechanges in perfusion pressure, indicating some degree of autor-egulation. By the same token, increases in arterial carbon dioxidetension resulted in increases in choroidal blood flow of approxi-mately 1.5% per mm Hg PCO2 (Geiser et al., 2000).
2.6. Choroidal blood flow: thermoregulation of the retina?
One proposed function of the extremely high blood flow of thechoroid is that it protects the retina from damage in extreme hotor cold temperatures or from the heat generated during exposure tobright lights (Parver et al., 1980; Parver et al., 1982a,b; Bill et al., 1983)by acting as a heat source in the cold (maintaining retinal temperaturenear core temperature) or as a heat sink for exogenous thermalradiation. In monkeys and rabbits, it was found that increasing theIOP to above the mean arterial pressure (thus preventing choroidalblood flow) under low ambient illumination resulted in a significantdecrease in the temperature of the retina and choroid in the macularegion (Bill, 1962). This was interpreted as showing that the choroidacts as a heat source for the retina under low illumination, when heatis lost through the cooler anterior chamber. Conversely, when flowwas occluded under higher illumination, there was an increase inchoroidal temperature, indicating the loss of the flow to the choroidacting as a heat sink (Parver et al., 1982a,b).
Although protection from these temperature changes couldoccur passively because of the high choroidal blood flow, as wouldbe expected from the classical view that the choroidal circulationdoes not autoregulate, they could also be mediated by reflexiveincreases in choroidal blood flow in response to a stimulus (Parver,1991). In addition to any tissue-level (local) autoregulation, there isevidence in humans and non-human primates for a centrally-controlled reflex arc regulating choroidal flow: In cynomolgusmonkeys, increasing light intensity under constant IOP resulted inan increase in retinal/choroidal temperature and in choroidal bloodflow (Parver et al., 1982a,b). In humans, ocular surface temperature
Fig. 9. Oxygen tension profile through a vascular retina (rat). The measurements arefrom two sequential penetrations (circles) and withdrawals (triangles) of the electrode.The intraretinal oxygen distribution reflects the relative oxygen sources and sinkswithin the retina and choroid. Reproduced with permission from Yu and Cringle, 2001! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168152

increases in response to increased light intensity, which mayimply that choroidal blood flow is increased as well under theseconditions (Parver et al., 1983). In birds, changes in light intensitymediate reflexive changes in choroidal blood flow, which may ormay not be thermoregulatory (Fitzgerald et al., 1996). The nature ofthe thermal sensory receptors and the pathways mediating thereflex arc are both as yet unknown.
This view of temperature regulation by the choroid is not,however, universally accepted. An argument can be made that thelight-evoked increase in choroidal/retinal temperature of 0.4 "CwithIOP held constant is evidence for a lack of thermoregulation andnot the opposite. Furthermore, core body temperature shows fluc-tuations normally as large as those retinal ones measured underconditions of raised IOP and increased light intensity, hence it would
be unlikely that core temperature would maintain a stable retinaltemperature.
2.7. Choroid and pathology: age-related macular degeneration
Because water and ions, as well as nutrients and plasma-borneprotein molecules, move in both directions across Bruch's membrane,impairment of this movement in some disease states and in thenormal aging process can have serious consequences for visual func-tion. In the normal process of aging, a thickeningof Bruch'smembraneand buildup of materials in the inner collagenous layer resultsin a decrease in water permeability; this is also seen in age-relatedmacular degeneration (AMD), the cause of 70% of blindness (Zarbin,1998, 2004). Impaired diffusion across Bruch's membrane may result
Fig. 10. Choroidal modulation of refractive state. A. Unfixed hemisected chick eyes. Arrowheads delimit choroidal boundaries. B. Plastic-embedded sections of the back of the eyes.L, lacuna; P, pigment cells; PE, retinal pigment epithelium; arrowhead, choriocapillaris. C, One-mm-thick section of the posterior eye wall. Ch, choroid, delimited by arrows.Reproduced with permission fromWallman et al., 1995 ! Elsevier. D. Thickness of the retinaþ choroid as a function of retinal defocus induced by spectacle lenses. Reproduced withpermission from Wildsoet and Wallman, 1995 ! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 153
in impaired diffusion of waste products from the RPE and impaireddelivery of hormones and oxygen to the RPE, eventually leading toatrophy of the RPE and retina. The thickness of the choriocapillarisand the capillary lumen diameters also decrease with age and AMD(Ramrattan et al., 1994; however, see Spraul et al., 1999). If a decreasein choroidal blood flow results in decreased clearance of debris fromthe RPE cells, this might contribute to the pathological changes inBruch's membrane that accompany AMD. It is also possible, however,that the RPE degeneration is the primary factor in the underlyingdeterioration of the choroid.
In the atrophic (dry) form of AMD the submacular chorioca-pillaris degenerates; it is unknown if this is a cause or consequenceof the inflammatory response that causes the pathologic changes inthe choroidal/RPE extracellular matrix (Zarbin, 2004). However,recent evidence showing a close association between degenerationof the RPE and that of the underlying choriocapillaris suggeststhat atrophy of the RPE occurs first (McLeod et al., 2009). In theexudative (wet) form of AMD there is choroidal neovascularization,which often leads to hemorrhage and retinal detachment. Thisneovascularization has been hypothesized to be the result of theRPE responding to the oxidative stress by synthesizing vasculargrowth factors such as vascular endothelial growth factor (VEGF).In this form of AMD, choriocapillaris degeneration initiallyoccurs in the presence of a viable RPE, suggesting that the neo-vascularization associated with it is a response to the ischemiainduced by the primary capillary degeneration, with subsequenteffects on the RPE (McLeod et al., 2009).
3. Modulation of choroidal thickness
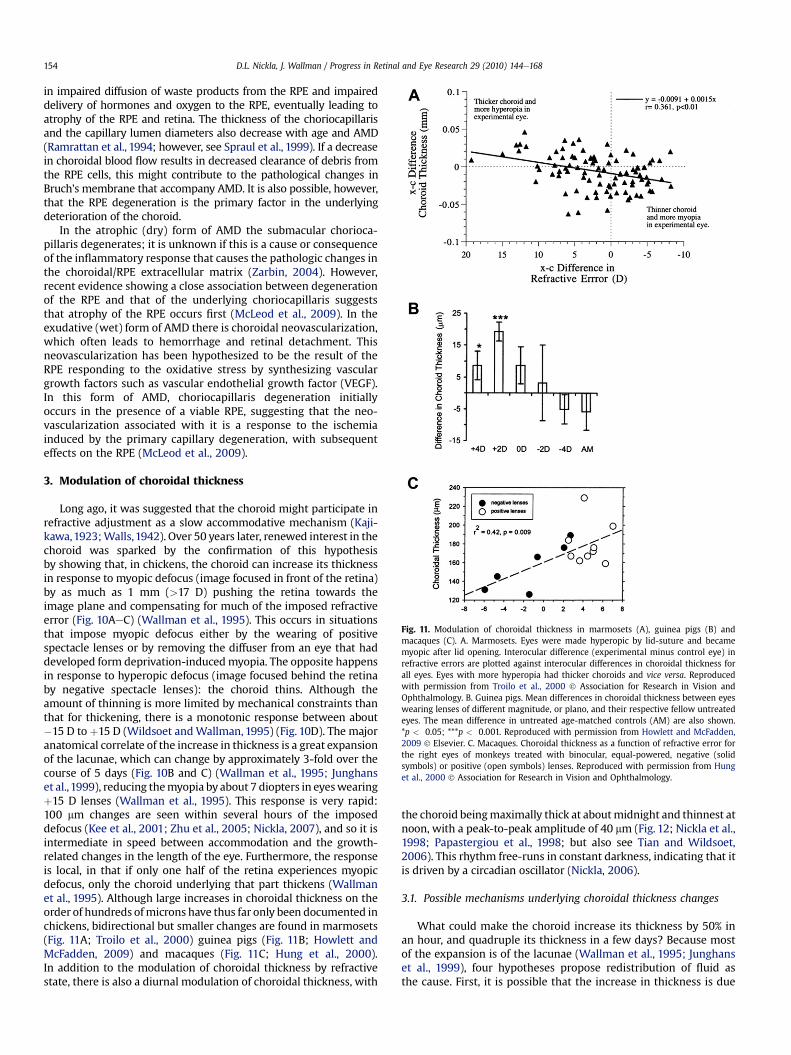
Long ago, it was suggested that the choroid might participate inrefractive adjustment as a slow accommodative mechanism (Kaji-kawa,1923;Walls,1942). Over 50 years later, renewed interest in thechoroid was sparked by the confirmation of this hypothesisby showing that, in chickens, the choroid can increase its thicknessin response to myopic defocus (image focused in front of the retina)by as much as 1 mm (>17 D) pushing the retina towards theimage plane and compensating for much of the imposed refractiveerror (Fig. 10AeC) (Wallman et al., 1995). This occurs in situationsthat impose myopic defocus either by the wearing of positivespectacle lenses or by removing the diffuser from an eye that haddeveloped form deprivation-inducedmyopia. The opposite happensin response to hyperopic defocus (image focused behind the retinaby negative spectacle lenses): the choroid thins. Although theamount of thinning is more limited by mechanical constraints thanthat for thickening, there is a monotonic response between about#15 D to þ15 D (Wildsoet andWallman, 1995) (Fig. 10D). The majoranatomical correlate of the increase in thickness is a great expansionof the lacunae, which can change by approximately 3-fold over thecourse of 5 days (Fig. 10B and C) (Wallman et al., 1995; Junghanset al.,1999), reducing themyopia by about 7 diopters in eyeswearingþ15 D lenses (Wallman et al., 1995). This response is very rapid:100 mm changes are seen within several hours of the imposeddefocus (Kee et al., 2001; Zhu et al., 2005; Nickla, 2007), and so it isintermediate in speed between accommodation and the growth-related changes in the length of the eye. Furthermore, the responseis local, in that if only one half of the retina experiences myopicdefocus, only the choroid underlying that part thickens (Wallmanet al., 1995). Although large increases in choroidal thickness on theorder of hundreds ofmicrons have thus far only been documented inchickens, bidirectional but smaller changes are found in marmosets(Fig. 11A; Troilo et al., 2000) guinea pigs (Fig. 11B; Howlett andMcFadden, 2009) and macaques (Fig. 11C; Hung et al., 2000).In addition to the modulation of choroidal thickness by refractivestate, there is also a diurnal modulation of choroidal thickness, with
the choroid beingmaximally thick at aboutmidnight and thinnest atnoon, with a peak-to-peak amplitude of 40 mm (Fig. 12; Nickla et al.,1998; Papastergiou et al., 1998; but also see Tian and Wildsoet,2006). This rhythm free-runs in constant darkness, indicating that itis driven by a circadian oscillator (Nickla, 2006).
3.1. Possible mechanisms underlying choroidal thickness changes
What could make the choroid increase its thickness by 50% inan hour, and quadruple its thickness in a few days? Because mostof the expansion is of the lacunae (Wallman et al., 1995; Junghanset al., 1999), four hypotheses propose redistribution of fluid asthe cause. First, it is possible that the increase in thickness is due
Fig. 11. Modulation of choroidal thickness in marmosets (A), guinea pigs (B) andmacaques (C). A. Marmosets. Eyes were made hyperopic by lid-suture and becamemyopic after lid opening. Interocular difference (experimental minus control eye) inrefractive errors are plotted against interocular differences in choroidal thickness forall eyes. Eyes with more hyperopia had thicker choroids and vice versa. Reproducedwith permission from Troilo et al., 2000 ! Association for Research in Vision andOphthalmology. B. Guinea pigs. Mean differences in choroidal thickness between eyeswearing lenses of different magnitude, or plano, and their respective fellow untreatedeyes. The mean difference in untreated age-matched controls (AM) are also shown.*p < 0.05; ***p < 0.001. Reproduced with permission from Howlett and McFadden,2009 ! Elsevier. C. Macaques. Choroidal thickness as a function of refractive error forthe right eyes of monkeys treated with binocular, equal-powered, negative (solidsymbols) or positive (open symbols) lenses. Reproduced with permission from Hunget al., 2000 ! Association for Research in Vision and Ophthalmology.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168154
to an increased synthesis of large, osmotically active proteogly-cans, which would pull water into the choroid. Second, it could bethe result of an increase in the size or number of the fenestrationsin the choriocapillaris, which could similarly increase the amountof osmotically active molecules in the choroidal matrix. Third,fluid could enter the choroid as part of the drainage from theanterior chamber. Fourth, the fluid could be a result of alteredtransport of fluid from the retina across the RPE. Finally, in addi-tion to other processes, changes in the tonus of the non-vascularsmooth muscle that spans the width of the choroid in both birdsand primates might play a role. It is likely that more than one ofthese is involved.
3.1.1. Changes in the synthesis of osmotically active moleculesBecause water is most commonly moved by moving osmotically
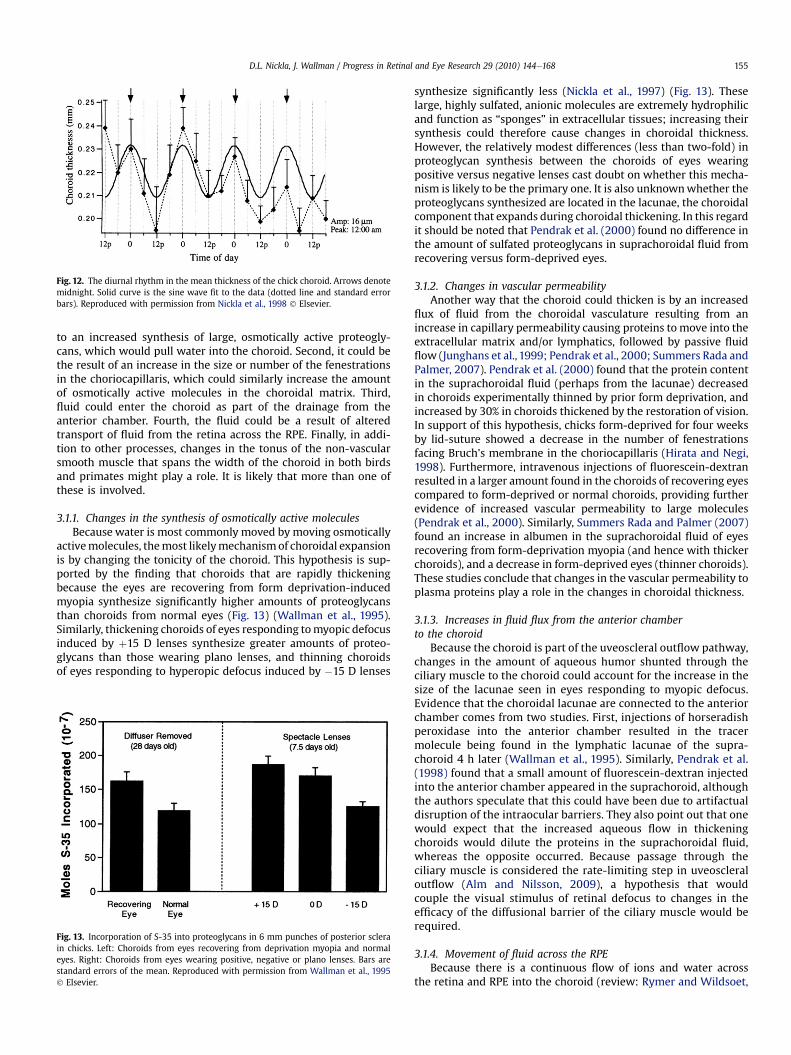
activemolecules, themost likelymechanismof choroidal expansionis by changing the tonicity of the choroid. This hypothesis is sup-ported by the finding that choroids that are rapidly thickeningbecause the eyes are recovering from form deprivation-inducedmyopia synthesize significantly higher amounts of proteoglycansthan choroids from normal eyes (Fig. 13) (Wallman et al., 1995).Similarly, thickening choroids of eyes responding tomyopic defocusinduced by þ15 D lenses synthesize greater amounts of proteo-glycans than those wearing plano lenses, and thinning choroidsof eyes responding to hyperopic defocus induced by #15 D lenses
synthesize significantly less (Nickla et al., 1997) (Fig. 13). Theselarge, highly sulfated, anionic molecules are extremely hydrophilicand function as “sponges” in extracellular tissues; increasing theirsynthesis could therefore cause changes in choroidal thickness.However, the relatively modest differences (less than two-fold) inproteoglycan synthesis between the choroids of eyes wearingpositive versus negative lenses cast doubt on whether this mecha-nism is likely to be the primary one. It is also unknownwhether theproteoglycans synthesized are located in the lacunae, the choroidalcomponent that expands during choroidal thickening. In this regardit should be noted that Pendrak et al. (2000) found no difference inthe amount of sulfated proteoglycans in suprachoroidal fluid fromrecovering versus form-deprived eyes.
3.1.2. Changes in vascular permeabilityAnother way that the choroid could thicken is by an increased
flux of fluid from the choroidal vasculature resulting from anincrease in capillary permeability causing proteins to move into theextracellular matrix and/or lymphatics, followed by passive fluidflow (Junghans et al., 1999; Pendrak et al., 2000; Summers Rada andPalmer, 2007). Pendrak et al. (2000) found that the protein contentin the suprachoroidal fluid (perhaps from the lacunae) decreasedin choroids experimentally thinned by prior form deprivation, andincreased by 30% in choroids thickened by the restoration of vision.In support of this hypothesis, chicks form-deprived for four weeksby lid-suture showed a decrease in the number of fenestrationsfacing Bruch's membrane in the choriocapillaris (Hirata and Negi,1998). Furthermore, intravenous injections of fluorescein-dextranresulted in a larger amount found in the choroids of recovering eyescompared to form-deprived or normal choroids, providing furtherevidence of increased vascular permeability to large molecules(Pendrak et al., 2000). Similarly, Summers Rada and Palmer (2007)found an increase in albumen in the suprachoroidal fluid of eyesrecovering from form-deprivation myopia (and hence with thickerchoroids), and a decrease in form-deprived eyes (thinner choroids).These studies conclude that changes in the vascular permeability toplasma proteins play a role in the changes in choroidal thickness.
3.1.3. Increases in fluid flux from the anterior chamberto the choroid
Because the choroid is part of the uveoscleral outflow pathway,changes in the amount of aqueous humor shunted through theciliary muscle to the choroid could account for the increase in thesize of the lacunae seen in eyes responding to myopic defocus.Evidence that the choroidal lacunae are connected to the anteriorchamber comes from two studies. First, injections of horseradishperoxidase into the anterior chamber resulted in the tracermolecule being found in the lymphatic lacunae of the supra-choroid 4 h later (Wallman et al., 1995). Similarly, Pendrak et al.(1998) found that a small amount of fluorescein-dextran injectedinto the anterior chamber appeared in the suprachoroid, althoughthe authors speculate that this could have been due to artifactualdisruption of the intraocular barriers. They also point out that onewould expect that the increased aqueous flow in thickeningchoroids would dilute the proteins in the suprachoroidal fluid,whereas the opposite occurred. Because passage through theciliary muscle is considered the rate-limiting step in uveoscleraloutflow (Alm and Nilsson, 2009), a hypothesis that wouldcouple the visual stimulus of retinal defocus to changes in theefficacy of the diffusional barrier of the ciliary muscle would berequired.
3.1.4. Movement of fluid across the RPEBecause there is a continuous flow of ions and water across
the retina and RPE into the choroid (review: Rymer and Wildsoet,
Fig. 12. The diurnal rhythm in the mean thickness of the chick choroid. Arrows denotemidnight. Solid curve is the sine wave fit to the data (dotted line and standard errorbars). Reproduced with permission from Nickla et al., 1998 ! Elsevier.
Fig. 13. Incorporation of S-35 into proteoglycans in 6 mm punches of posterior sclerain chicks. Left: Choroids from eyes recovering from deprivation myopia and normaleyes. Right: Choroids from eyes wearing positive, negative or plano lenses. Bars arestandard errors of the mean. Reproduced with permission from Wallman et al., 1995! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 155

2005), modulation of this flow could modulate choroidal thickness,if the outflow from the choroid did not match the inflow acrossthe RPE. Because there is also a flow of fluid and solutes from theblood vessels of the choriocapillaris into the tissues and back intothe blood vessels, it is unclear whether the flow across the RPEwould be large enough to upset this equilibrium.
One group of researchers has advanced the view that, at leastin the case of the choroidal expansion that follows restoration ofvision after form deprivation, there is a region of hyperosmolaritythat moves from the outer retina during form deprivation to theouter choroid after vision is restored (Junghans et al., 1999; Lianget al., 2004). Electron microscopy shows a sequential thickeningof the retina, RPE and lastly the choroid, concurrent with changesin the concentrations of sodium and chloride in these tissues.The fact that these concentrations decrease over the first 72 h ofrecovery, while choroidal thickness increases is presented asevidence that the choroidal thickening results from edema, andnot from a visually driven refractive compensation.
Correlated with these changes in thickness, the basal membraneof the RPE cells has fewer infoldings in the deprived eye, whichincrease over the time of the restored vision, as do the numberof membrane-bound fluid vesicles (Liang et al., 2004). The authorsinterpreted these changes as evidence of edema. Furthermore, thefenestrations of the endothelial cells of the lacunae increase insize in the thickened choroids of recovering eyes, consistent withchanges in fluid flux (Junghans et al., 1999). Their interpretationis that the return to emmetropia from form-deprivation myopiarepresents the re-establishment of normal physiology and ultra-structure, as fluid moves from the vitreous to the choroidallymphatics, and finally out of the eye, rather than being part ofa homeostatic regulation of refractive state.
These findings, however, do not preclude the possibility thatthe choroidal changes are visually regulated; that is, that they aredriven by changes in refractive error, rather than by passive edema.Furthermore, the fact that the choroidal thickness is closely relatedto lens-induced defocus in both directions, and that there is nearlyprecise refractive compensation for induced defocus, seriouslyweakens this hypothesis.
3.1.5. Changes in the tonus of non-vascular smooth muscleBecause the choroid can thin very rapidly, by about 100 mm in
3e4 h in young chicks (Kee et al., 2001), one is inclined to look formuscular, rather than osmotic, mechanisms. Given that the choroidcontains abundant non-vascular smoothmuscle, probably controlledby both sympathetic and parasympathetic inputs, the contraction ofthese muscles might squeeze fluid out of the choroid, thereby thin-ning it. Thus, we can consider the possibility that the lacunae ofthe choroid are always somewhat hypertonic and tend to acquirefluid, and this tendency is opposed by the tonus of the non-vascularsmoothmuscle, so that if they contract, the choroid becomes thinner,whereas if they relax, the choroid becomes thicker. This hypothesis issupported by the finding that drastically lowering the IOP causeschoroidal expansion (Abelsdoff and Wessely, 1909). However,because the non-vascular smooth muscle is not aligned perpendic-ular to the plane of the choroid, it is also possible that contraction ofthese muscles facilitate the filling of the lacunae.
While the innervation of the choroidal vasculature has beenextensively studied, that of the non-vascular smooth muscle isstill poorly understood. However, most evidence indicates thatthe non-vascular smooth muscle is under dual sympathetic andparasympathetic control. First, in human choroids, non-vascularsmooth muscle cells are contacted by axon terminals that arepositive for NADPH-diaphorase (a co-factor for NOS), as well as fortyrosine hydroxylase, the catecholamine rate-limiting enzyme(Schrödl et al., 2001b), suggesting innervation by the
pterygopalatine or possibly the ciliary ganglion (although only 1%of these neurons are nitrergic), and the superior cervical ganglion(Poukens et al., 1998). Second, in birds, tyrosine hydroxylase is co-localized with dopamine-B-hydroxylase in terminals on the non-vascular smooth muscle, the double-localization of which isa marker for post-ganglionic sympathetic input (Schrödl et al.,2001b). Furthermore, lesions of the superior cervical ganglionresult in loss of adrenergic fibers in the choroidal stroma, possiblyreflecting an input onto non-vascular smooth muscle (Guglielmoneand Cantino, 1982). In both humans and birds, the presence ofterminals labeled for NADPH-diaphorase (Poukens et al., 1998) andnNOS and VIP (May et al., 2004), indicates innervation from theparasympathetic system, specifically the pterygopalatine ganglion,or the ICNs (Schrödl et al., 2003; May et al., 2004). The direction ofaction of the parasympathetic innervation is shown by the fact thatelectrical stimulation of the post-ganglionic axons from the chickciliary ganglion cause contraction of explant choroids, and thiscontraction is blocked by atropine but not by curare, showing thatacetylcholine causes contraction of at least some choroidal smoothmuscle (Meriney and Pilar, 1987).
4. The choroid and emmetropization
As the eye develops from birth to maturity it undergoesadjustments of its optical components so that most eyes eventuallybecome emmetropic (focused for objects at distance). It is generallyaccepted that this “emmetropization” is determined by a combi-nation of environmental (i.e., visual) and genetic influences. Whenthis process goes awry, the eyes develop refractive errors (hyper-opia or myopia). In the United States approximately 25% of thepopulation is myopic (Sperduto et al., 1983) while in other societiesthe vast majority of the educated population is myopic (Lin, 1996).Understanding the mechanisms underlying emmetropization hasdirect clinical relevance for the prevention of myopia.
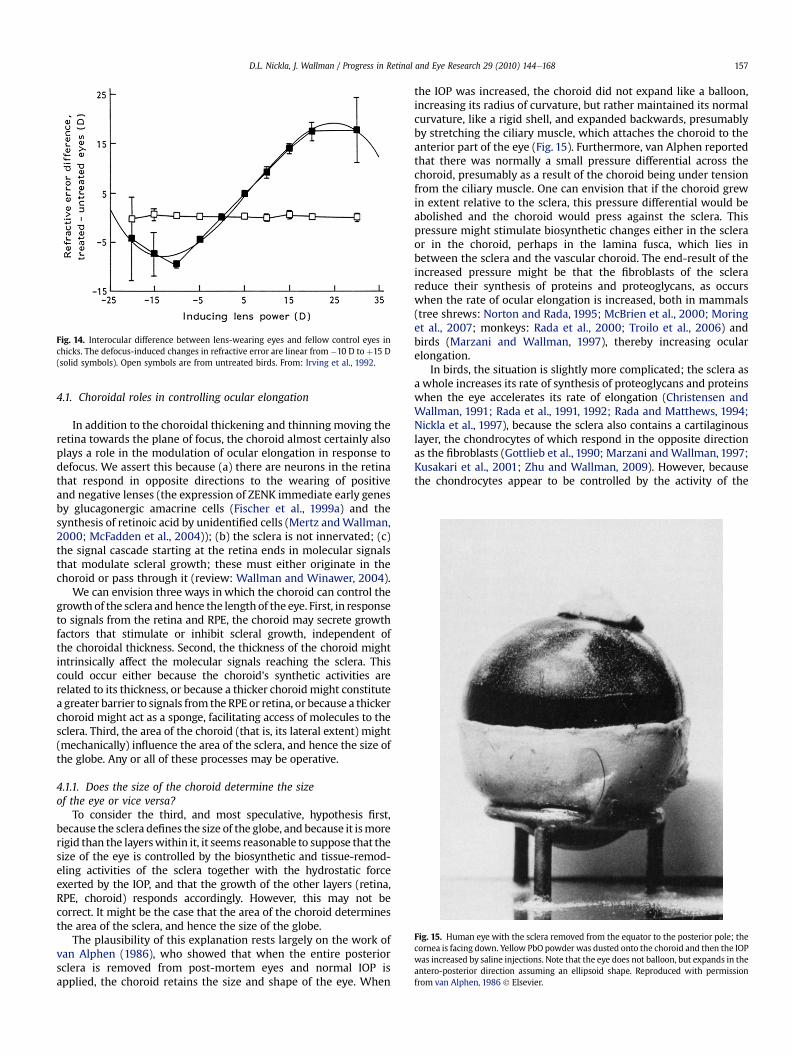
Work with animal models has been crucial in establishingthe importance of the visual environment in the regulation of oculargrowth (reviews: Wallman, 1993; Norton, 1999; Wallman andWinawer, 2004). The original findings showed that depriving theeye of form vision by lid-suture or plastic diffusers resulted inexcessive ocular elongation and consequent myopia in all speciesstudied (Sherman et al., 1977; Wiesel and Raviola, 1977; Wallmanet al., 1978). The strongest evidence for the visual regulation of eyegrowth however comes from studies using spectacle lenses toimpose defocus on the retina. Chick eyes show nearly completecompensation to both the magnitude and the sign of the defocuswhile the lenses are worn (Schaeffel et al., 1990; Irving et al., 1992;Wildsoet andWallman, 1995): Eyes made functionally myopic withpositive lenses (image focused in front of the retina) compensate forthe myopia by becoming hyperopic during the period of lens-wearwhile those made functionally hyperopic with negative lenses(image focused behind the retina) becamemyopic (Fig.14). After thelenses were removed, the eyes compensated in the oppositedirection, returning to emmetropia (Wildsoet and Wallman, 1995).In chick eyes, the compensatory response has two components:(1) changes in the size of the globe, accompanied by changes in thesynthesis of extracellular matrix macromolecules in the sclera(Rada et al., 1991, 1992), and (2) changes in the thickness of thechoroid,moving the retina forward in the case ofmyopic defocus, orbackward, in the case of hyperopic defocus (Wallman et al., 1995;Wildsoet and Wallman, 1995) (Fig. 10). We ask, first, in what waysdoes the choroid participate in the modulation of ocular growth;and second, what is the relationship between choroidal thickeningand inhibition of ocular elongation? Are they independenthomeostatic responses to defocus, or does the choroid influenceocular elongation?
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168156
4.1. Choroidal roles in controlling ocular elongation
In addition to the choroidal thickening and thinning moving theretina towards the plane of focus, the choroid almost certainly alsoplays a role in the modulation of ocular elongation in response todefocus. We assert this because (a) there are neurons in the retinathat respond in opposite directions to the wearing of positiveand negative lenses (the expression of ZENK immediate early genesby glucagonergic amacrine cells (Fischer et al., 1999a) and thesynthesis of retinoic acid by unidentified cells (Mertz andWallman,2000; McFadden et al., 2004)); (b) the sclera is not innervated; (c)the signal cascade starting at the retina ends in molecular signalsthat modulate scleral growth; these must either originate in thechoroid or pass through it (review: Wallman and Winawer, 2004).
We can envision three ways in which the choroid can control thegrowth of the sclera andhence the length of the eye. First, in responseto signals from the retina and RPE, the choroid may secrete growthfactors that stimulate or inhibit scleral growth, independent ofthe choroidal thickness. Second, the thickness of the choroid mightintrinsically affect the molecular signals reaching the sclera. Thiscould occur either because the choroid's synthetic activities arerelated to its thickness, or because a thicker choroidmight constitutea greater barrier to signals from the RPE or retina, or because a thickerchoroid might act as a sponge, facilitating access of molecules to thesclera. Third, the area of the choroid (that is, its lateral extent) might(mechanically) influence the area of the sclera, and hence the size ofthe globe. Any or all of these processes may be operative.
4.1.1. Does the size of the choroid determine the sizeof the eye or vice versa?
To consider the third, and most speculative, hypothesis first,because the sclera defines the size of the globe, and because it ismorerigid than the layerswithin it, it seems reasonable to suppose that thesize of the eye is controlled by the biosynthetic and tissue-remod-eling activities of the sclera together with the hydrostatic forceexerted by the IOP, and that the growth of the other layers (retina,RPE, choroid) responds accordingly. However, this may not becorrect. It might be the case that the area of the choroid determinesthe area of the sclera, and hence the size of the globe.
The plausibility of this explanation rests largely on the work ofvan Alphen (1986), who showed that when the entire posteriorsclera is removed from post-mortem eyes and normal IOP isapplied, the choroid retains the size and shape of the eye. When
the IOP was increased, the choroid did not expand like a balloon,increasing its radius of curvature, but rather maintained its normalcurvature, like a rigid shell, and expanded backwards, presumablyby stretching the ciliary muscle, which attaches the choroid to theanterior part of the eye (Fig. 15). Furthermore, van Alphen reportedthat there was normally a small pressure differential across thechoroid, presumably as a result of the choroid being under tensionfrom the ciliary muscle. One can envision that if the choroid grewin extent relative to the sclera, this pressure differential would beabolished and the choroid would press against the sclera. Thispressure might stimulate biosynthetic changes either in the scleraor in the choroid, perhaps in the lamina fusca, which lies inbetween the sclera and the vascular choroid. The end-result of theincreased pressure might be that the fibroblasts of the sclerareduce their synthesis of proteins and proteoglycans, as occurswhen the rate of ocular elongation is increased, both in mammals(tree shrews: Norton and Rada, 1995; McBrien et al., 2000; Moringet al., 2007; monkeys: Rada et al., 2000; Troilo et al., 2006) andbirds (Marzani and Wallman, 1997), thereby increasing ocularelongation.
In birds, the situation is slightly more complicated; the sclera asa whole increases its rate of synthesis of proteoglycans and proteinswhen the eye accelerates its rate of elongation (Christensen andWallman, 1991; Rada et al., 1991, 1992; Rada and Matthews, 1994;Nickla et al., 1997), because the sclera also contains a cartilaginouslayer, the chondrocytes of which respond in the opposite directionas the fibroblasts (Gottlieb et al., 1990; Marzani andWallman, 1997;Kusakari et al., 2001; Zhu and Wallman, 2009). However, becausethe chondrocytes appear to be controlled by the activity of the
Fig. 14. Interocular difference between lens-wearing eyes and fellow control eyes inchicks. The defocus-induced changes in refractive error are linear from #10 D to þ15 D(solid symbols). Open symbols are from untreated birds. From: Irving et al., 1992.
Fig. 15. Human eye with the sclera removed from the equator to the posterior pole; thecornea is facing down. Yellow PbO powder was dusted onto the choroid and then the IOPwas increased by saline injections. Note that the eye does not balloon, but expands in theantero-posterior direction assuming an ellipsoid shape. Reproduced with permissionfrom van Alphen, 1986 ! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 157

fibroblasts (Marzani and Wallman, 1997), the perichondrial fibro-blasts adjacent to the choroid might be the cells that respond to thepressure. A difficulty raised by this conjecture is that it would seemthat a thickened choroid would also exert more force on the sclera,and yet choroidal thickening is associated with decreased ocularelongation.
One could also envision the opposite relationship: The choroidcould grow to fit the sclera. In this scenario, the same mechano-sensory stimulation just mentioned of the choroid pushing upagainst the sclera could operate to inhibit choroidal expansion,rather than to stimulate scleral expansion. In either case, it seemsplausible that the coordination of the growth of the tissues at theback of the eyemay be guided bymechanical forces,whichmodulatethe release of growth factors. Other examples of such mechano-sensory homeostasis are changes in fibroblast gene expressionresulting from mechanical forces on the extracellular matrix (Chi-quet et al., 2009) and the expression of growth factor receptors beingmodulated by mechanical factors during angiogenesis (Mammotoet al., 2009).
4.1.2. Are the choroidal and scleral responses independent?The general association of choroidal thinning with acceleration
of ocular elongation and of choroidal thickeningwith slowed ocularelongation raises the question of whether these two processes arecausally linked. That is, does the choroid have a single phenotypeor physiological state that determines both whether the choroidthickens or thins and whether ocular elongation (and therefore,scleral growth) is stimulated or inhibited, or are the two responsesindependent?
4.1.2.1. Evidence for independent responses. In support of the inde-pendence of the choroidal thickening and scleral modulatingresponses there are several studies that show differential effects ofparticular visual conditions on the two responses. For example,when the eye receives myopic defocus from positive lenses for briefbut infrequent periods (darkness for the rest of the time), thereis inhibition of axial growth, without choroidal thickening.Conversely, when the eye receives hyperopic defocus from negativelenses for brief but infrequent periods, there is choroidal thinningwithout axial elongation (Winawer and Wallman, 2002). Further-more, if a light diffuser is put over a positive lens, this eliminatesthe choroidal thickening that the positive lens would normallyprovoke, but with no effect on the lens-induced inhibition ofocular elongation (Park et al., 2003). Most provocatively, if chickswear lenses in dim blue or red light, such that only the short-wavelength-sensitive cones or long-wavelength-sensitive conesare stimulated, the eyes compensate in both cases in terms ofrefractive status, but only the choroid thickness changes in the caseof the red light, and only the ocular length changes in the case ofthe blue light (Rucker and Wallman, 2008).
Although these examples are certainly compatible with separatevisual signals guiding the choroidal and scleral responses, theycould also indicate that the same visual signals are integrateddifferently, so that visual signal are more effective in thinning thanthickening the choroid, and in decelerating than accelerating ocularelongation. When tested explicitly by giving episodes of lens-wearat a range of different durations and intervals (with darkness inbetween), the major result was that episodes of positive lens-wearhad a four times more enduring effect on inhibiting ocular elon-gation than on thickening the choroid, whereas episodes ofnegative lens-wear had an eight times more enduring effect onthinning the choroid than on accelerating ocular elongation (Zhuand Wallman, 2009).
While the integration of these episodes of lens-wear could beoccurring in either the retina, the RPE or the choroid, the implication
of the differential action on choroid and sclera might be that thesame signals act differently on different tissues, depending on theirintensity and time course. The implication of different temporalrequirements for the effects of negative versus positive lenses mightbe that the signals controlling the thickening versus thinning of thechoroid are different from those controlling the acceleration versusdeceleration of the ocular elongation, or it might simply imply thatsome signals are easier to turn up than to turn down, or vice versa.For example, one can imagine a brief episode of lens-wear causingan increase in transcription of a gene, resulting in a long-lastingincrease in a peptide, whereas a decrease in the same peptide mightrequire a more continuous expression of a molecule that degradesthe peptide or its mRNA.
4.1.2.2. Evidence for choroidal thickness modulating ocularelongation. The preceding section has shown that the usual linkingof changes in choroidal thickness and rate of ocular elongation canbe disrupted by experimental manipulations. However, more recentwork has shown that brief exposure to myopic defocus causestransient thickening of the choroid, lasting only hours (Nickla, 2007),which therefore would not be detected in the usual experiments,such as those in the preceding section in which eyes are measuredat the start and end of the experiment. Because brief exposureto myopic defocus has an effect on the rate of ocular elongation thatis over 10 times more enduring than the effect of hyperopic defocus(Zhu and Wallman, 2009), it may be that the transient episodes ofchoroidal thickening are responsible for the enduring effect of briefperiods of myopic defocus.
4.1.2.2.1. The enduring effects of brief episodes of myopic defocusand transient choroidal thickening. It is known that long periods ofform deprivation can be largely overridden by brief daily periodsof either unrestricted vision (Napper et al., 1995), or myopicdefocus (Winawer and Wallman, 2002; Nickla, 2007), suggestinga long time-constant for myopiagenic stimuli. Similarly, negativelenses need to be worn nearly continuously to produce myopia inchicks (Schmid and Wildsoet, 1996; Winawer and Wallman,2002), tree shrews (Shaikh et al., 1999) and macaques (Kee et al.,2007). On the other hand, if a chick living in a normal visualenvironment wears a positive lens for only 2 min four times a day,hyperopia and a slowing of ocular elongation results (Zhu et al.,2005). Consistent with this asymmetry between signs of defocus,when negative and positive lenses are alternated, brief periods ofpositive lens-wear balance out much longer periods of negativelens-wear (Winawer and Wallman, 2002; Zhu and Wallman,2009). The developmental utility of it being easier to halt ocularelongation than to accelerate it might be to prevent the develop-ment of myopia from transient visual disturbances (such as periodsof near vision with hyperopic defocus because of accommodativelag), given that the eye can correct its refractive error more easilyby growing than by shrinking.
Because even 10 min of myopic defocus induces transientchoroidal thickening within an hour (Zhu et al., 2005), perhaps thistransient thickening causes the ensuing longer-lasting oculargrowth inhibition. This hypothesis is supported by the finding thatseveral daily visual manipulations that block myopia also causetransient choroidal thickening. Specifically, whether the myopia-genic stimulus was form deprivation or negative lens-wear, myopiawas inhibited by 2 daily hours of unrestricted vision or by 1 dailyhour of stroboscopic illumination, both of which produced tran-sient choroidal thickening within 3 h (Fig. 16) (Nickla, 2007) thattapered off over the ensuing 24 h. Similarly, if the myopia caused byconstant darkness was inhibited by either a half hour per day ofmyopic defocus, 2 min per hour per day of myopic defocus, or onedaily hour of unrestricted vision, again, all three stimuli resulted intransient choroidal thickening (Nickla, 2007). Interestingly, in the
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168158
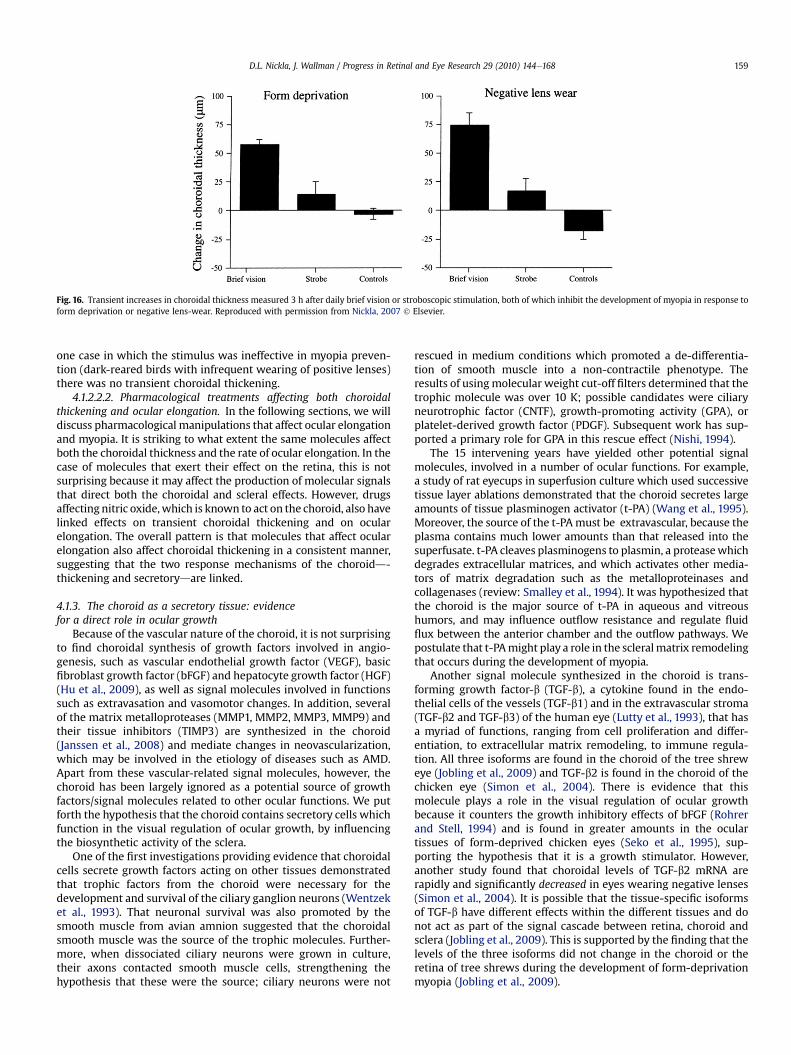
one case in which the stimulus was ineffective in myopia preven-tion (dark-reared birds with infrequent wearing of positive lenses)there was no transient choroidal thickening.
4.1.2.2.2. Pharmacological treatments affecting both choroidalthickening and ocular elongation. In the following sections, we willdiscuss pharmacological manipulations that affect ocular elongationand myopia. It is striking to what extent the same molecules affectboth the choroidal thickness and the rate of ocular elongation. In thecase of molecules that exert their effect on the retina, this is notsurprising because it may affect the production of molecular signalsthat direct both the choroidal and scleral effects. However, drugsaffecting nitric oxide,which is known to act on the choroid, also havelinked effects on transient choroidal thickening and on ocularelongation. The overall pattern is that molecules that affect ocularelongation also affect choroidal thickening in a consistent manner,suggesting that the two response mechanisms of the choroidd-thickening and secretorydare linked.
4.1.3. The choroid as a secretory tissue: evidencefor a direct role in ocular growth
Because of the vascular nature of the choroid, it is not surprisingto find choroidal synthesis of growth factors involved in angio-genesis, such as vascular endothelial growth factor (VEGF), basicfibroblast growth factor (bFGF) and hepatocyte growth factor (HGF)(Hu et al., 2009), as well as signal molecules involved in functionssuch as extravasation and vasomotor changes. In addition, severalof the matrix metalloproteases (MMP1, MMP2, MMP3, MMP9) andtheir tissue inhibitors (TIMP3) are synthesized in the choroid(Janssen et al., 2008) and mediate changes in neovascularization,which may be involved in the etiology of diseases such as AMD.Apart from these vascular-related signal molecules, however, thechoroid has been largely ignored as a potential source of growthfactors/signal molecules related to other ocular functions. We putforth the hypothesis that the choroid contains secretory cells whichfunction in the visual regulation of ocular growth, by influencingthe biosynthetic activity of the sclera.
One of the first investigations providing evidence that choroidalcells secrete growth factors acting on other tissues demonstratedthat trophic factors from the choroid were necessary for thedevelopment and survival of the ciliary ganglion neurons (Wentzeket al., 1993). That neuronal survival was also promoted by thesmooth muscle from avian amnion suggested that the choroidalsmooth muscle was the source of the trophic molecules. Further-more, when dissociated ciliary neurons were grown in culture,their axons contacted smooth muscle cells, strengthening thehypothesis that these were the source; ciliary neurons were not
rescued in medium conditions which promoted a de-differentia-tion of smooth muscle into a non-contractile phenotype. Theresults of usingmolecular weight cut-off filters determined that thetrophic molecule was over 10 K; possible candidates were ciliaryneurotrophic factor (CNTF), growth-promoting activity (GPA), orplatelet-derived growth factor (PDGF). Subsequent work has sup-ported a primary role for GPA in this rescue effect (Nishi, 1994).
The 15 intervening years have yielded other potential signalmolecules, involved in a number of ocular functions. For example,a study of rat eyecups in superfusion culture which used successivetissue layer ablations demonstrated that the choroid secretes largeamounts of tissue plasminogen activator (t-PA) (Wang et al., 1995).Moreover, the source of the t-PA must be extravascular, because theplasma contains much lower amounts than that released into thesuperfusate. t-PA cleaves plasminogens to plasmin, a proteasewhichdegrades extracellular matrices, and which activates other media-tors of matrix degradation such as the metalloproteinases andcollagenases (review: Smalley et al., 1994). It was hypothesized thatthe choroid is the major source of t-PA in aqueous and vitreoushumors, and may influence outflow resistance and regulate fluidflux between the anterior chamber and the outflow pathways. Wepostulate that t-PAmight play a role in the scleralmatrix remodelingthat occurs during the development of myopia.
Another signal molecule synthesized in the choroid is trans-forming growth factor-b (TGF-b), a cytokine found in the endo-thelial cells of the vessels (TGF-b1) and in the extravascular stroma(TGF-b2 and TGF-b3) of the human eye (Lutty et al., 1993), that hasa myriad of functions, ranging from cell proliferation and differ-entiation, to extracellular matrix remodeling, to immune regula-tion. All three isoforms are found in the choroid of the tree shreweye (Jobling et al., 2009) and TGF-b2 is found in the choroid of thechicken eye (Simon et al., 2004). There is evidence that thismolecule plays a role in the visual regulation of ocular growthbecause it counters the growth inhibitory effects of bFGF (Rohrerand Stell, 1994) and is found in greater amounts in the oculartissues of form-deprived chicken eyes (Seko et al., 1995), sup-porting the hypothesis that it is a growth stimulator. However,another study found that choroidal levels of TGF-b2 mRNA arerapidly and significantly decreased in eyes wearing negative lenses(Simon et al., 2004). It is possible that the tissue-specific isoformsof TGF-b have different effects within the different tissues and donot act as part of the signal cascade between retina, choroid andsclera (Jobling et al., 2009). This is supported by the finding that thelevels of the three isoforms did not change in the choroid or theretina of tree shrews during the development of form-deprivationmyopia (Jobling et al., 2009).
Fig. 16. Transient increases in choroidal thickness measured 3 h after daily brief vision or stroboscopic stimulation, both of which inhibit the development of myopia in response toform deprivation or negative lens-wear. Reproduced with permission from Nickla, 2007 ! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 159
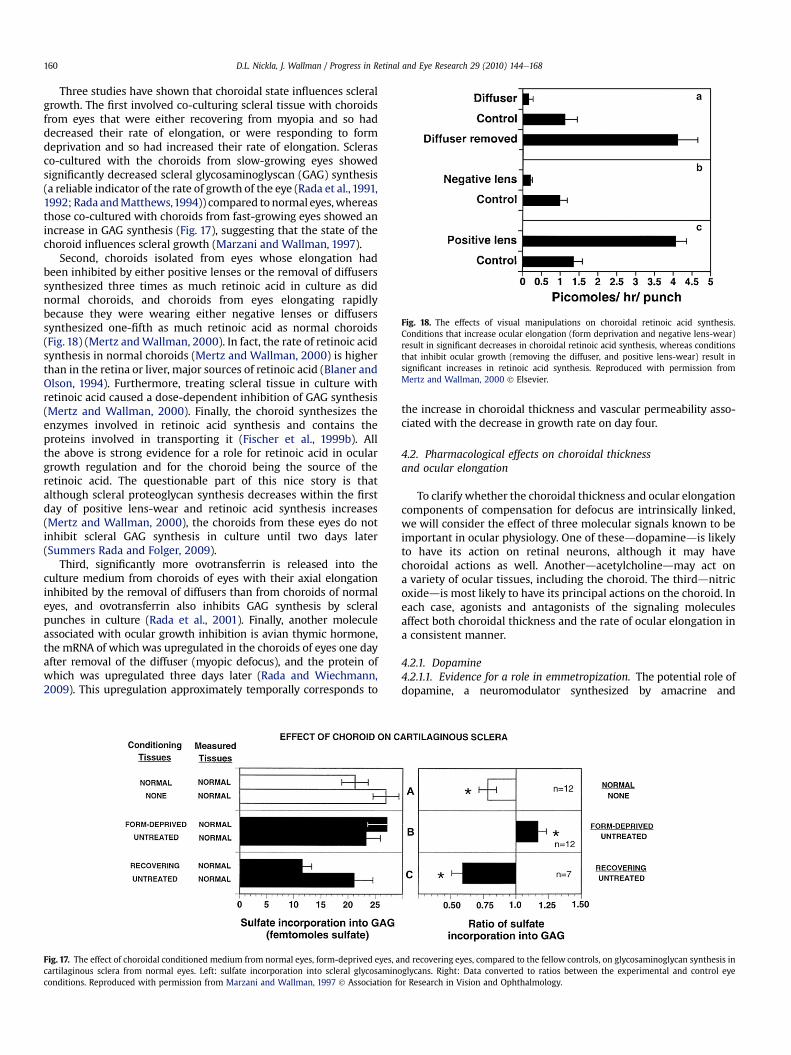
Three studies have shown that choroidal state influences scleralgrowth. The first involved co-culturing scleral tissue with choroidsfrom eyes that were either recovering from myopia and so haddecreased their rate of elongation, or were responding to formdeprivation and so had increased their rate of elongation. Sclerasco-cultured with the choroids from slow-growing eyes showedsignificantly decreased scleral glycosaminoglyscan (GAG) synthesis(a reliable indicator of the rate of growth of the eye (Rada et al., 1991,1992; Rada andMatthews,1994)) compared to normal eyes,whereasthose co-cultured with choroids from fast-growing eyes showed anincrease in GAG synthesis (Fig. 17), suggesting that the state of thechoroid influences scleral growth (Marzani and Wallman, 1997).
Second, choroids isolated from eyes whose elongation hadbeen inhibited by either positive lenses or the removal of diffuserssynthesized three times as much retinoic acid in culture as didnormal choroids, and choroids from eyes elongating rapidlybecause they were wearing either negative lenses or diffuserssynthesized one-fifth as much retinoic acid as normal choroids(Fig. 18) (Mertz andWallman, 2000). In fact, the rate of retinoic acidsynthesis in normal choroids (Mertz and Wallman, 2000) is higherthan in the retina or liver, major sources of retinoic acid (Blaner andOlson, 1994). Furthermore, treating scleral tissue in culture withretinoic acid caused a dose-dependent inhibition of GAG synthesis(Mertz and Wallman, 2000). Finally, the choroid synthesizes theenzymes involved in retinoic acid synthesis and contains theproteins involved in transporting it (Fischer et al., 1999b). Allthe above is strong evidence for a role for retinoic acid in oculargrowth regulation and for the choroid being the source of theretinoic acid. The questionable part of this nice story is thatalthough scleral proteoglycan synthesis decreases within the firstday of positive lens-wear and retinoic acid synthesis increases(Mertz and Wallman, 2000), the choroids from these eyes do notinhibit scleral GAG synthesis in culture until two days later(Summers Rada and Folger, 2009).
Third, significantly more ovotransferrin is released into theculture medium from choroids of eyes with their axial elongationinhibited by the removal of diffusers than from choroids of normaleyes, and ovotransferrin also inhibits GAG synthesis by scleralpunches in culture (Rada et al., 2001). Finally, another moleculeassociated with ocular growth inhibition is avian thymic hormone,the mRNA of which was upregulated in the choroids of eyes one dayafter removal of the diffuser (myopic defocus), and the protein ofwhich was upregulated three days later (Rada and Wiechmann,2009). This upregulation approximately temporally corresponds to
the increase in choroidal thickness and vascular permeability asso-ciated with the decrease in growth rate on day four.
4.2. Pharmacological effects on choroidal thicknessand ocular elongation
To clarify whether the choroidal thickness and ocular elongationcomponents of compensation for defocus are intrinsically linked,we will consider the effect of three molecular signals known to beimportant in ocular physiology. One of theseddopaminedis likelyto have its action on retinal neurons, although it may havechoroidal actions as well. Anotherdacetylcholinedmay act ona variety of ocular tissues, including the choroid. The thirddnitricoxidedis most likely to have its principal actions on the choroid. Ineach case, agonists and antagonists of the signaling moleculesaffect both choroidal thickness and the rate of ocular elongation ina consistent manner.
4.2.1. Dopamine4.2.1.1. Evidence for a role in emmetropization. The potential role ofdopamine, a neuromodulator synthesized by amacrine and
Fig. 17. The effect of choroidal conditioned medium from normal eyes, form-deprived eyes, and recovering eyes, compared to the fellow controls, on glycosaminoglycan synthesis incartilaginous sclera from normal eyes. Left: sulfate incorporation into scleral glycosaminoglycans. Right: Data converted to ratios between the experimental and control eyeconditions. Reproduced with permission from Marzani and Wallman, 1997 ! Association for Research in Vision and Ophthalmology.
Fig. 18. The effects of visual manipulations on choroidal retinoic acid synthesis.Conditions that increase ocular elongation (form deprivation and negative lens-wear)result in significant decreases in choroidal retinoic acid synthesis, whereas conditionsthat inhibit ocular growth (removing the diffuser, and positive lens-wear) result insignificant increases in retinoic acid synthesis. Reproduced with permission fromMertz and Wallman, 2000 ! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168160

interplexiform cells, in the signal cascade mediating emmetropiza-tion has been the subject of interest and debate for about 20 years(Stone et al., 1989; Iuvone et al., 1991; Rohrer et al., 1993). The initialinterest was sparked by the finding that dopamine was decreased inthe retinas of form-deprived eyes (Stone et al., 1989) and negativelens-wearing eyes (Guo et al., 1995) and increased in retinas of eyesrecovering from form-deprivation myopia (Pendrak et al., 1997).Interestingly, only daytime levels of dopamine were affected, sug-gesting the involvement of a circadian component. Furthermore, thedevelopment of both form-deprivation myopia (Stone et al., 1989;Iuvone et al., 1991; Rohrer et al., 1993) and lens-induced myopia(Schmid and Wildsoet, 2004) was inhibited by the administration ofthe non-specific dopamine agonist apomorphine, further supportinga role for dopamine in ocular growth inhibition. The D2 receptoragonist quinpirole, but not the D1 receptor agonist SKF-38393,inhibited the development of myopia in eyes wearing diffusers ornegative lenses, suggesting that the site of action was on the D2receptors in the retina rather than the D1 receptors on RPE cells(Schaeffel et al., 1995). Accordingly, the D2 receptor antagonist sul-piride enhanced deprivation myopia (Schaeffel et al., 1995) andprevented brief periods of daily vision from blocking the develop-ment of deprivation myopia (McCarthy et al., 2007), consistent withthe idea that dopamine is required for this inhibition of excessivegrowth.
However, other studies using dopaminergic agents have yieldedconflicting results thatmake this interpretationproblematic (Schaeffelet al., 1995). For instance, intravitreal injections of haloperidol,a dopamine antagonist, suppressed the development of deprivationmyopia (Stone et al., 1989). Similarly, 6-hydroxy-dopamine, a neuro-toxin that depletes catecholamine levels, reduced the developmentof deprivation myopia (Li et al., 1992) and did not alter the responseto lens-wear (Schaeffel et al., 1994). The same was true of reserpine,a depletor of both dopamine and serotonin (Schaeffel et al., 1995).
Despite these problematic issues, dopamine is still a potentialcandidate in ocular growth regulation (McCarthy et al., 2007).Furthermore, it has been shown that dopamine stimulates theproduction of nitric oxide (Melis et al., 1996), another potentialgrowth candidate molecule synthesized in the choroid, making itpossible that dopamine is part of the signal cascade upstream ofthe choroidal changes. It is also possible, however, that there is anas yet unknown source of dopamine in the chicken choroid: the co-localization of dopamine-b-hydroxylase and tyrosine hydroxylasein nerve terminals in the duck choroid indicates that dopamineis synthesized as an intermediate product in the biosynthesis ofnoradrenaline, but in the chicken choroidal terminals, dopamine-beta-hydroxylase appears to be absent, suggesting the possibilitythat dopamine rather than noradrenaline might be the transmitterfor these neurons (Falk Schrödl, personal communication).
4.2.1.2. Effects on choroidal thickness and ocular elongation. Ifdopamine were part of the signal cascade mediating ocular growthinhibition, and if it acted upstream of the choroid, then injections ofthe effective agonists should cause transient increases in choroidalthickness, similar to the effects induced by 2 h of myopic defocus orremoving the diffuser or negative lens. Intravitreal injections of bothapomorphine (a non-specific agonist) and quinpirole (a D2 agonist),both effective inhibitors of myopia, into eyes wearing negativelenses produced significant transient increases in choroidal thick-ness (Fig. 19) that were associated both with the inhibition of axialelongation and the resultant myopia (Dhillon and Nickla, 2008). TheD1 agonist SKF-38393, which did not prevent the development ofmyopia, also did not elicit choroidal thickening (Dhillon and Nickla,2008). Together these results support a role for dopaminergic D2receptor-mediated mechanisms, and choroidal thickening, in oculargrowth inhibition.
4.2.2. Acetylcholine4.2.2.1. Evidence for a role in emmetropization. Atropine, a non-specific muscarinic acetylcholine antagonist, has long been knownto inhibit the development of myopia in humans (Bedrossian, 1971;Shih et al., 1999a; Kennedy, 1995; Kennedy et al., 2000), and toprevent form-deprivation myopia in several species (chicks: Stoneet al., 1991; McBrien et al., 1993b; Schmid and Wildsoet, 2004;Schwahn et al., 2000; Luft et al., 2003; tree shrews: McKanna andCasagrande,1981; monkeys: Young,1965; Raviola andWiesel, 1985;Tigges et al., 1999). The effect is believed to be mediated by M1 orM4 receptors, because pirenzepine (M1 and M4 antagonist) is alsoeffective (Stone et al., 1991; McBrien et al., 1993a,b; Leech et al.,1995; Cottriall and McBrien, 1996; Schwahn et al., 2000; Luft et al.,2003). Of a large number of muscarinic antagonists examined, onlypirenzepine and oxyphenonium (non-selective), but not scopol-amine (M1) were effective in inhibiting axial elongation in responseto form deprivation in chicks (Luft et al., 2003). Because M1 recep-tors are reportedly absent in chicks, it is likely that pirenzepine actson M4, rather than M1 receptors (McBrien et al., 2009).
It is unclear on what tissue of the eye atropine acts to exert itsanti-myopiagenic effects. Contrary to the early views, the suppres-sion ofmyopia by atropine does not depend on its cycloplegic action,as atropine is also effective in preventing myopia in birds (Stoneet al., 1991; McBrien et al., 1993b), inwhich the ciliary muscle, beingstriated, is nicotinic, and in a non-accommodating mammal(McBrien et al., 1993a). The view that it acts at the retinal level is notsupported, as lesions of cholinergic retinal neurons do not alterthe effects of form deprivation, nor do they affect the inhibitoryinfluence of atropine (Fischer et al., 1998). It is possible that atropinehas an indirect retinal action, by causing the release of dopamine orother neurotransmitters, or by inducing spreading depression inthe retina (Schwahn et al., 2000). Finally, evidence in tree shrews(Cottriall et al., 2000), guinea pigs (Liu et al., 2007) and chicks(Lind et al., 1998) indicate that the site of actionmay be scleral: formdeprivation causes an increase in the expression of M1 and M4receptor mRNA in the guinea pig sclera (Liu et al., 2007).
4.2.2.2. Effects on choroidal thickness and ocular elongation. Theconnection between muscarinic receptors and both transientchoroidal thickening and inhibition of the excessive ocular elon-gation is shown by the finding that, when injected into the vitreousof birds wearing #10D lenses, atropine, pirenzepine and oxy-phenonium, drugs that are effective in eliciting ocular growthinhibition (Luft et al., 2003) also caused a significant and transient
-20
0
20
40
60
Cha
nge
in c
horo
id th
ickn
ess
(µm
)
Apomorph Quinpirole Saline Fellow
Fig. 19. Effects of dopamine agonists on choroidal thickness over 3 h from the injectioninto eyes wearing #10 D lenses. Both effective growth inhibitors result in transientincreases in choroidal thickness. Unpublished data.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 161
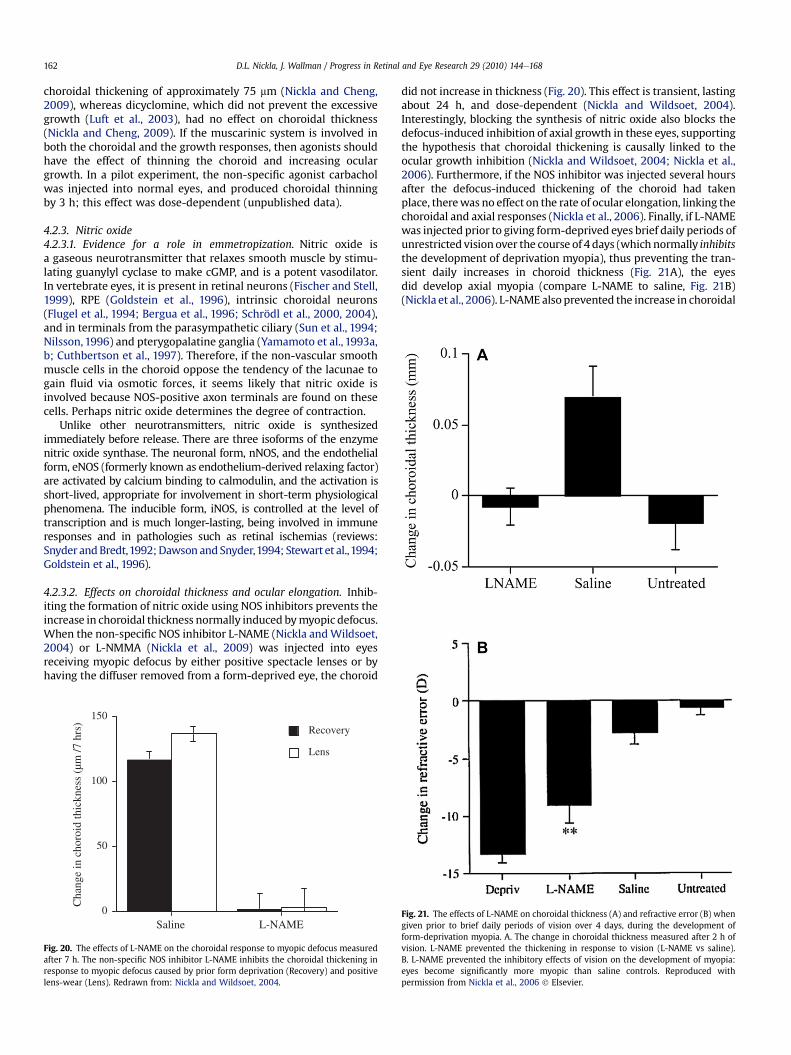
choroidal thickening of approximately 75 mm (Nickla and Cheng,2009), whereas dicyclomine, which did not prevent the excessivegrowth (Luft et al., 2003), had no effect on choroidal thickness(Nickla and Cheng, 2009). If the muscarinic system is involved inboth the choroidal and the growth responses, then agonists shouldhave the effect of thinning the choroid and increasing oculargrowth. In a pilot experiment, the non-specific agonist carbacholwas injected into normal eyes, and produced choroidal thinningby 3 h; this effect was dose-dependent (unpublished data).
4.2.3. Nitric oxide4.2.3.1. Evidence for a role in emmetropization. Nitric oxide isa gaseous neurotransmitter that relaxes smooth muscle by stimu-lating guanylyl cyclase to make cGMP, and is a potent vasodilator.In vertebrate eyes, it is present in retinal neurons (Fischer and Stell,1999), RPE (Goldstein et al., 1996), intrinsic choroidal neurons(Flugel et al., 1994; Bergua et al., 1996; Schrödl et al., 2000, 2004),and in terminals from the parasympathetic ciliary (Sun et al., 1994;Nilsson, 1996) and pterygopalatine ganglia (Yamamoto et al., 1993a,b; Cuthbertson et al., 1997). Therefore, if the non-vascular smoothmuscle cells in the choroid oppose the tendency of the lacunae togain fluid via osmotic forces, it seems likely that nitric oxide isinvolved because NOS-positive axon terminals are found on thesecells. Perhaps nitric oxide determines the degree of contraction.
Unlike other neurotransmitters, nitric oxide is synthesizedimmediately before release. There are three isoforms of the enzymenitric oxide synthase. The neuronal form, nNOS, and the endothelialform, eNOS (formerly known as endothelium-derived relaxing factor)are activated by calcium binding to calmodulin, and the activation isshort-lived, appropriate for involvement in short-term physiologicalphenomena. The inducible form, iNOS, is controlled at the level oftranscription and is much longer-lasting, being involved in immuneresponses and in pathologies such as retinal ischemias (reviews:Snyder andBredt,1992;Dawson and Snyder,1994; Stewart et al.,1994;Goldstein et al., 1996).
4.2.3.2. Effects on choroidal thickness and ocular elongation. Inhib-iting the formation of nitric oxide using NOS inhibitors prevents theincrease in choroidal thickness normally induced bymyopic defocus.When the non-specific NOS inhibitor L-NAME (Nickla andWildsoet,2004) or L-NMMA (Nickla et al., 2009) was injected into eyesreceiving myopic defocus by either positive spectacle lenses or byhaving the diffuser removed from a form-deprived eye, the choroid
did not increase in thickness (Fig. 20). This effect is transient, lastingabout 24 h, and dose-dependent (Nickla and Wildsoet, 2004).Interestingly, blocking the synthesis of nitric oxide also blocks thedefocus-induced inhibition of axial growth in these eyes, supportingthe hypothesis that choroidal thickening is causally linked to theocular growth inhibition (Nickla and Wildsoet, 2004; Nickla et al.,2006). Furthermore, if the NOS inhibitor was injected several hoursafter the defocus-induced thickening of the choroid had takenplace, therewas no effect on the rate of ocular elongation, linking thechoroidal and axial responses (Nickla et al., 2006). Finally, if L-NAMEwas injected prior to giving form-deprived eyes brief daily periods ofunrestricted visionover the course of 4 days (which normally inhibitsthe development of deprivation myopia), thus preventing the tran-sient daily increases in choroid thickness (Fig. 21A), the eyesdid develop axial myopia (compare L-NAME to saline, Fig. 21B)(Nickla et al., 2006). L-NAME also prevented the increase in choroidal
0
50
100
150
Cha
nge
in c
horo
id th
ickn
ess
(µm
/7 h
rs)
Saline L-NAME
Lens
Recovery
Fig. 20. The effects of L-NAME on the choroidal response to myopic defocus measuredafter 7 h. The non-specific NOS inhibitor L-NAME inhibits the choroidal thickening inresponse to myopic defocus caused by prior form deprivation (Recovery) and positivelens-wear (Lens). Redrawn from: Nickla and Wildsoet, 2004.
Fig. 21. The effects of L-NAME on choroidal thickness (A) and refractive error (B) whengiven prior to brief daily periods of vision over 4 days, during the development ofform-deprivation myopia. A. The change in choroidal thickness measured after 2 h ofvision. L-NAME prevented the thickening in response to vision (L-NAME vs saline).B. L-NAME prevented the inhibitory effects of vision on the development of myopia:eyes become significantly more myopic than saline controls. Reproduced withpermission from Nickla et al., 2006 ! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168162
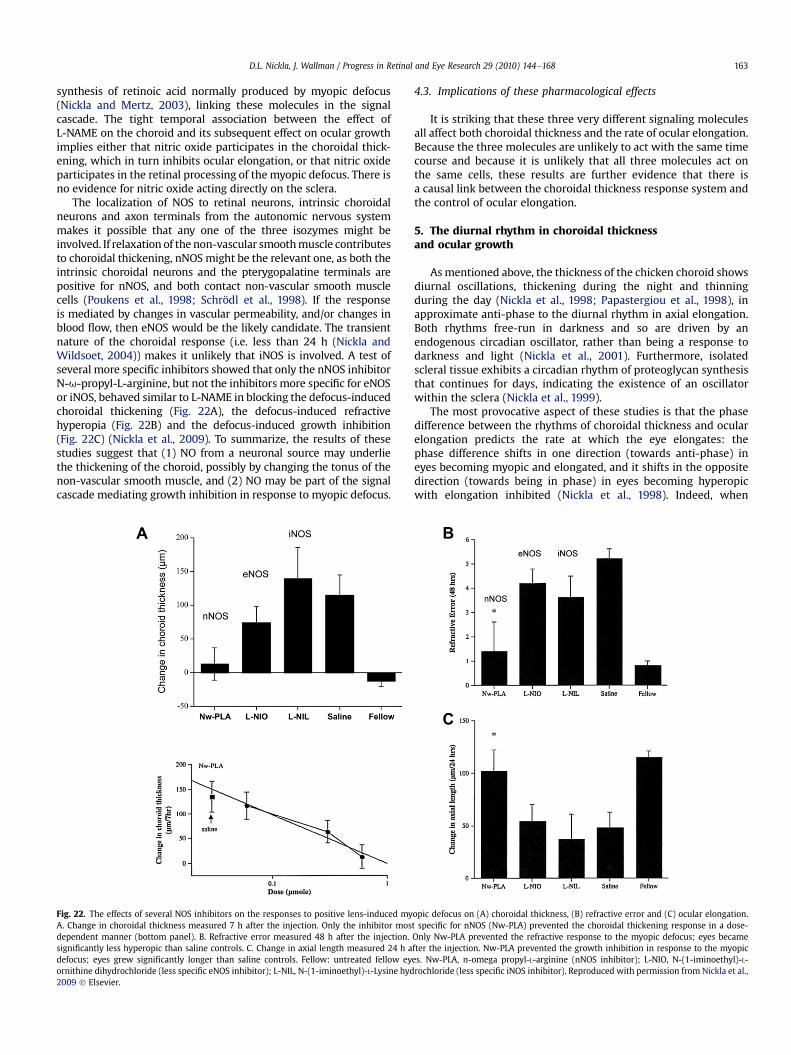
synthesis of retinoic acid normally produced by myopic defocus(Nickla and Mertz, 2003), linking these molecules in the signalcascade. The tight temporal association between the effect ofL-NAME on the choroid and its subsequent effect on ocular growthimplies either that nitric oxide participates in the choroidal thick-ening, which in turn inhibits ocular elongation, or that nitric oxideparticipates in the retinal processing of the myopic defocus. There isno evidence for nitric oxide acting directly on the sclera.
The localization of NOS to retinal neurons, intrinsic choroidalneurons and axon terminals from the autonomic nervous systemmakes it possible that any one of the three isozymes might beinvolved. If relaxation of the non-vascular smoothmuscle contributesto choroidal thickening, nNOS might be the relevant one, as both theintrinsic choroidal neurons and the pterygopalatine terminals arepositive for nNOS, and both contact non-vascular smooth musclecells (Poukens et al., 1998; Schrödl et al., 1998). If the responseis mediated by changes in vascular permeability, and/or changes inblood flow, then eNOS would be the likely candidate. The transientnature of the choroidal response (i.e. less than 24 h (Nickla andWildsoet, 2004)) makes it unlikely that iNOS is involved. A test ofseveral more specific inhibitors showed that only the nNOS inhibitorN-u-propyl-L-arginine, but not the inhibitors more specific for eNOSor iNOS, behaved similar to L-NAME in blocking the defocus-inducedchoroidal thickening (Fig. 22A), the defocus-induced refractivehyperopia (Fig. 22B) and the defocus-induced growth inhibition(Fig. 22C) (Nickla et al., 2009). To summarize, the results of thesestudies suggest that (1) NO from a neuronal source may underliethe thickening of the choroid, possibly by changing the tonus of thenon-vascular smooth muscle, and (2) NO may be part of the signalcascade mediating growth inhibition in response to myopic defocus.
4.3. Implications of these pharmacological effects
It is striking that these three very different signaling moleculesall affect both choroidal thickness and the rate of ocular elongation.Because the three molecules are unlikely to act with the same timecourse and because it is unlikely that all three molecules act onthe same cells, these results are further evidence that there isa causal link between the choroidal thickness response system andthe control of ocular elongation.
5. The diurnal rhythm in choroidal thicknessand ocular growth
Asmentioned above, the thickness of the chicken choroid showsdiurnal oscillations, thickening during the night and thinningduring the day (Nickla et al., 1998; Papastergiou et al., 1998), inapproximate anti-phase to the diurnal rhythm in axial elongation.Both rhythms free-run in darkness and so are driven by anendogenous circadian oscillator, rather than being a response todarkness and light (Nickla et al., 2001). Furthermore, isolatedscleral tissue exhibits a circadian rhythm of proteoglycan synthesisthat continues for days, indicating the existence of an oscillatorwithin the sclera (Nickla et al., 1999).
The most provocative aspect of these studies is that the phasedifference between the rhythms of choroidal thickness and ocularelongation predicts the rate at which the eye elongates: thephase difference shifts in one direction (towards anti-phase) ineyes becoming myopic and elongated, and it shifts in the oppositedirection (towards being in phase) in eyes becoming hyperopicwith elongation inhibited (Nickla et al., 1998). Indeed, when
Fig. 22. The effects of several NOS inhibitors on the responses to positive lens-induced myopic defocus on (A) choroidal thickness, (B) refractive error and (C) ocular elongation.A. Change in choroidal thickness measured 7 h after the injection. Only the inhibitor most specific for nNOS (Nw-PLA) prevented the choroidal thickening response in a dose-dependent manner (bottom panel). B. Refractive error measured 48 h after the injection. Only Nw-PLA prevented the refractive response to the myopic defocus; eyes becamesignificantly less hyperopic than saline controls. C. Change in axial length measured 24 h after the injection. Nw-PLA prevented the growth inhibition in response to the myopicdefocus; eyes grew significantly longer than saline controls. Fellow: untreated fellow eyes. Nw-PLA, n-omega propyl-L-arginine (nNOS inhibitor); L-NIO, N-(1-iminoethyl)-L-ornithine dihydrochloride (less specific eNOS inhibitor); L-NIL, N-(1-iminoethyl)-L-Lysine hydrochloride (less specific iNOS inhibitor). Reproduced with permission from Nickla et al.,2009 ! Elsevier.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 163
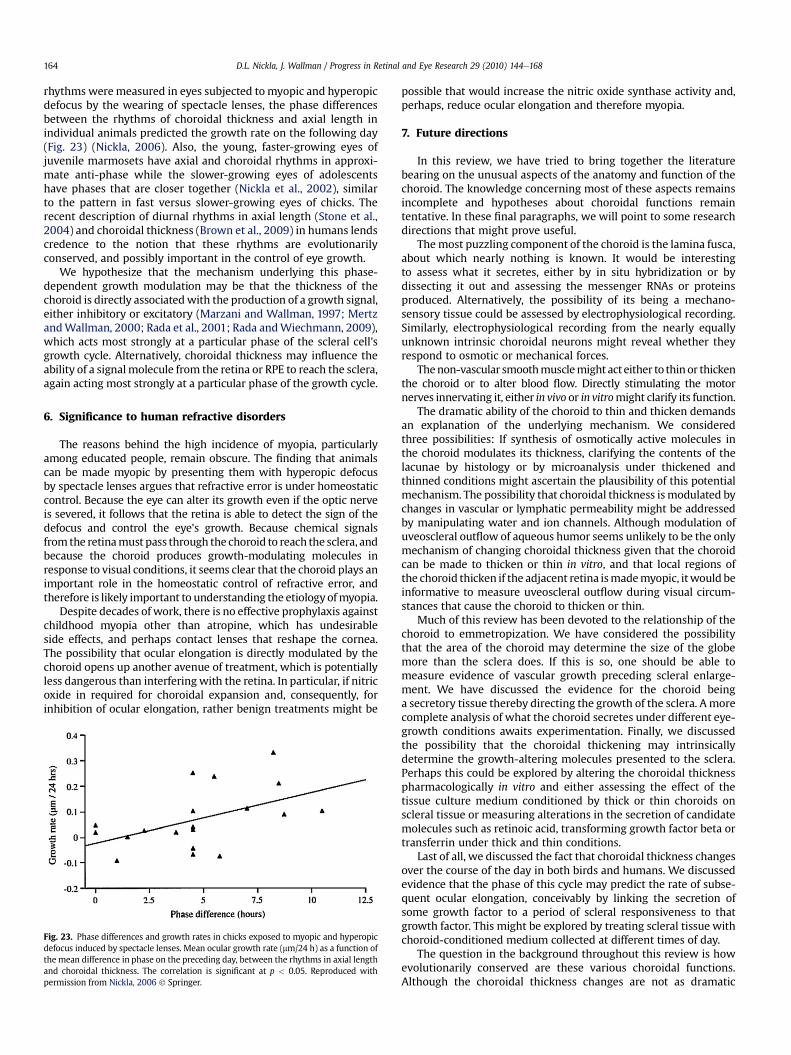
rhythms weremeasured in eyes subjected tomyopic and hyperopicdefocus by the wearing of spectacle lenses, the phase differencesbetween the rhythms of choroidal thickness and axial length inindividual animals predicted the growth rate on the following day(Fig. 23) (Nickla, 2006). Also, the young, faster-growing eyes ofjuvenile marmosets have axial and choroidal rhythms in approxi-mate anti-phase while the slower-growing eyes of adolescentshave phases that are closer together (Nickla et al., 2002), similarto the pattern in fast versus slower-growing eyes of chicks. Therecent description of diurnal rhythms in axial length (Stone et al.,2004) and choroidal thickness (Brown et al., 2009) in humans lendscredence to the notion that these rhythms are evolutionarilyconserved, and possibly important in the control of eye growth.
We hypothesize that the mechanism underlying this phase-dependent growth modulation may be that the thickness of thechoroid is directly associatedwith the production of a growth signal,either inhibitory or excitatory (Marzani and Wallman, 1997; MertzandWallman, 2000; Rada et al., 2001; Rada andWiechmann, 2009),which acts most strongly at a particular phase of the scleral cell'sgrowth cycle. Alternatively, choroidal thickness may influence theability of a signalmolecule from the retina or RPE to reach the sclera,again acting most strongly at a particular phase of the growth cycle.
6. Significance to human refractive disorders
The reasons behind the high incidence of myopia, particularlyamong educated people, remain obscure. The finding that animalscan be made myopic by presenting them with hyperopic defocusby spectacle lenses argues that refractive error is under homeostaticcontrol. Because the eye can alter its growth even if the optic nerveis severed, it follows that the retina is able to detect the sign of thedefocus and control the eye's growth. Because chemical signalsfrom the retinamust pass through the choroid to reach the sclera, andbecause the choroid produces growth-modulating molecules inresponse to visual conditions, it seems clear that the choroid plays animportant role in the homeostatic control of refractive error, andtherefore is likely important to understanding the etiology ofmyopia.
Despite decades of work, there is no effective prophylaxis againstchildhood myopia other than atropine, which has undesirableside effects, and perhaps contact lenses that reshape the cornea.The possibility that ocular elongation is directly modulated by thechoroid opens up another avenue of treatment, which is potentiallyless dangerous than interfering with the retina. In particular, if nitricoxide in required for choroidal expansion and, consequently, forinhibition of ocular elongation, rather benign treatments might be
possible that would increase the nitric oxide synthase activity and,perhaps, reduce ocular elongation and therefore myopia.
7. Future directions
In this review, we have tried to bring together the literaturebearing on the unusual aspects of the anatomy and function of thechoroid. The knowledge concerning most of these aspects remainsincomplete and hypotheses about choroidal functions remaintentative. In these final paragraphs, we will point to some researchdirections that might prove useful.
Themost puzzling component of the choroid is the lamina fusca,about which nearly nothing is known. It would be interestingto assess what it secretes, either by in situ hybridization or bydissecting it out and assessing the messenger RNAs or proteinsproduced. Alternatively, the possibility of its being a mechano-sensory tissue could be assessed by electrophysiological recording.Similarly, electrophysiological recording from the nearly equallyunknown intrinsic choroidal neurons might reveal whether theyrespond to osmotic or mechanical forces.
Thenon-vascular smoothmusclemight act either to thinor thickenthe choroid or to alter blood flow. Directly stimulating the motornerves innervating it, either in vivo or in vitromight clarify its function.
The dramatic ability of the choroid to thin and thicken demandsan explanation of the underlying mechanism. We consideredthree possibilities: If synthesis of osmotically active molecules inthe choroid modulates its thickness, clarifying the contents of thelacunae by histology or by microanalysis under thickened andthinned conditions might ascertain the plausibility of this potentialmechanism. The possibility that choroidal thickness ismodulated bychanges in vascular or lymphatic permeability might be addressedby manipulating water and ion channels. Although modulation ofuveoscleral outflow of aqueous humor seems unlikely to be the onlymechanism of changing choroidal thickness given that the choroidcan be made to thicken or thin in vitro, and that local regions ofthe choroid thicken if the adjacent retina ismademyopic, itwould beinformative to measure uveoscleral outflow during visual circum-stances that cause the choroid to thicken or thin.
Much of this review has been devoted to the relationship of thechoroid to emmetropization. We have considered the possibilitythat the area of the choroid may determine the size of the globemore than the sclera does. If this is so, one should be able tomeasure evidence of vascular growth preceding scleral enlarge-ment. We have discussed the evidence for the choroid beinga secretory tissue thereby directing the growth of the sclera. Amorecomplete analysis of what the choroid secretes under different eye-growth conditions awaits experimentation. Finally, we discussedthe possibility that the choroidal thickening may intrinsicallydetermine the growth-altering molecules presented to the sclera.Perhaps this could be explored by altering the choroidal thicknesspharmacologically in vitro and either assessing the effect of thetissue culture medium conditioned by thick or thin choroids onscleral tissue or measuring alterations in the secretion of candidatemolecules such as retinoic acid, transforming growth factor beta ortransferrin under thick and thin conditions.
Last of all, we discussed the fact that choroidal thickness changesover the course of the day in both birds and humans. We discussedevidence that the phase of this cycle may predict the rate of subse-quent ocular elongation, conceivably by linking the secretion ofsome growth factor to a period of scleral responsiveness to thatgrowth factor. This might be explored by treating scleral tissue withchoroid-conditioned medium collected at different times of day.
The question in the background throughout this review is howevolutionarily conserved are these various choroidal functions.Although the choroidal thickness changes are not as dramatic
Fig. 23. Phase differences and growth rates in chicks exposed to myopic and hyperopicdefocus induced by spectacle lenses. Mean ocular growth rate (mm/24 h) as a function ofthe mean difference in phase on the preceding day, between the rhythms in axial lengthand choroidal thickness. The correlation is significant at p < 0.05. Reproduced withpermission from Nickla, 2006 ! Springer.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168164

in mammals as in birds, they seem to occur in the same circum-stances, and three of the unusual features of choroid dintrinsicchoroidal neurons, endothelium-lined lacunae, and non-vascularsmooth muscle cells, occur in both birds and humans. How deepthese similarities extend remains a question for the future.
Acknowledgements
Supported by: NIH-EY-013636 (DLN) and EY-02727 andRR03060 (JW).
The authors are grateful to the following people for their importantcontributions: Jeffrey Kiel for his insightful discussions on the choroidand thermoregulation, and Falk Schrödl, Maria Egle De Stefano andGerald Lutty for their excellent critical readings of the manuscript.
References
Abelsdoff, G., Wessely, K., 1909. Vergleichendphysiologische Untersuchungen fiberden Flussigkeit-swechsel des auges in der Wirbeltierreihe. Arch. Augenheilk 64,65e124.
Alm, A., 1992. Ocular circulation. In: Hart, W.M. (Ed.), Adler's Physiology of the Eye.Mosby-Year Book Inc., St. Louis, MO, pp. 198e227.
Alm, A., Bill, A., 1973. Ocular and optic nerve blood flow at normal and increasedintraocular pressure in monkeys. Exp. Eye Res. 15, 15e29.
Alm, P., Uvelius, B., Ekstrom, J., Holmqvist, B., Larsson, B., Andersson, K.E., 1995.Nitric oxide synthase-containing neurons in rat parasympathetic, sympatheticand sensory ganglia: a comparative study. Histochem. J. 27, 819e831.
Alm, A., Nilsson, F.E., 2009. Uveoscleral outflow: a review. Exp. Eye Res. 88,760e768.
van Alphen, G.W.H.M., 1986. Choroidal stress and emmetropization. Vis. Res. 26,723e734.
Bar, H., Wende, P., Watson, L., 2002. Smoothelin is an indicator or reversiblephenotype modulation of smooth muscle cells in baloon-injured rat carotidarteries. Basic Res. Cardio 97, 9e16.
Baumel, J.J., 1975. Aves nervous system. In: Sisson and Grossman's the Anatomy ofDomestic Animals. W.B. Saunders, Philadelphia, pp. 2019e2066.
Bedrossian, R.H., 1971. The effect of atropine on myopia. Ann. Ophthalmol. 3,891e897.
Bergua, A., Mayer, B., Neuhuber, W., 1996. Nitrergic and VIP-ergic neurons in thechoroid and ciliary ganglion of the duck Anis carina. Anat. Embryol. 193,239e248.
Bergua, A., Schrödl, F., Brehmer, A., Neuhuber, W.L., 1998. Ultrastructural charac-teristic of different neuron types in the choroid of the duck. Exp. Eye Res. 67, 78.
Bergua, A., Junnemann, A., De Laet, A., Timmermans, J.P., Brehmer, A., Neuhuber, W.,Schrödl, F., 2003. Topography of nitrergic intrinsic choroidal neurons (ICN) inthe human eye. Invest. Ophthalmol. Vis. Sci. 44.
Bill, A., 1962. Intraocular pressure and blood flow through the uvea. Arch. Ophthal67, 336e348.
Bill, A., 1975. Blood circulation and fluid dynamics in the eye. Physiol. Rev. 55,383e417.
Bill, A., 1985. Some aspects of the ocular circulation (Friedenwald Lecture). Invest.Ophthalmol. Vis. Sci. 26, 410e424.
Bill, A., 1991. Effects of some neuropeptides on the uvea. Exp. Eye Res. 53, 3e11.Bill, A., Stjernschantz, J., Alm, A., 1976. Effects of hexamethonium, biperiden and
phentolamine on the vasoconstrictive effects of oculomotor nerve stimulationin rabbits. Exp. Eye Res. 23, 615e622.
Bill, A., Tornqvist, P., Alm, A., 1980. The permeability of the intraocular blood vessels.Trans. Ophthal. Soc. UK 100, 332e336.
Bill, A., Sperber, G., Ujiie, K., 1983. Physiology of the choroidal vascular bed. Int.Ophthalmol. 6, 101e107.
Blaner, W.S., Olson, J.A., 1994. Retinol and retinoic acid metabolism. In: Sporn, M.B.,Roberts, A.B., Goodman, D.S. (Eds.), The Retinoids: Biology, Chemistry andMedicine. Raven Press, pp. 229e255.
Brown, J.S., Flitcroft, D.I., Ying, G., Francis, E.L., Schmid, G.F., Quinn, G.E., Stone, R.A.,2009. In vivo human choroidal thickness measurements: evidence for diurnalfluctuations. Invest. Ophthalmol. Vis. Sci. 50, 5e12.
Chiquet, M., Gelman, L., Lutz, R., Maier, S., 2009. From mechanotransduction toextracellular matrix gene expression in fibroblasts. Biochim. Biophys. Acta 1793,911e920.
Christensen, A.M., Wallman, J., 1991. Evidence that increased scleral growthunderlies visual deprivation myopia in chicks. Invest. Ophthalmol. Vis. Sci. 32,2143e2150.
Cottriall, C.L., Truong, H.T., Gentle, A., McBrien, N.A., 2000. Changes in scleralmetabolism following in vivo application of pirenzepine to prevent axialmyopia. Invest. Ophthalmol. Vis. Sci. 41 (Suppl), S133.
Cottriall, C.L., McBrien, N., 1996. The m1 muscarinic antagonist pirenzepine reducesmyopia and eye enlargment in the tree shrew. Invest. Ophthalmol. Vis. Sci. 37,1368e1379.
Cuthbertson, S.L., White, J., Fitzgerald, M., Shih, Y.F., Reiner, A., 1996. Distributionwithin the choroid of cholinergic nerve fibers from the ciliary ganglion inpigeons. Vis. Res. 36, 775e786.
Cuthbertson, S., Jackson, B., Toledo, C., Fitzgerald, M.E.C., Shih, Y.F., Zagvazdin, Y.,Reiner, A., 1997. Innervation of orbital and choroidal blood vessels by thepterygopalatine ganglion in pigeons. J. Comp. Neurol. 386, 422e442.
Dawson, T.M., Snyder, S.H., 1994. Gases as biological messengers: nitric oxide andcarbon monoxide in the brain. J. Neurosci. 14, 5147e5159.
De Stefano, M., Ciofi Luzzatto, A., Mugnaini, E., 1993. Neuronal ultrastructure andsomatostatin immunolocalization in the ciliary ganglion of chicken and quail.J. Neurocytol 22, 868e892.
De Stefano, M., Mugnaini, E., 1997. Fine structure of the choroidal coat of the avianeye: lymphatic vessels. Invest. Ophthalmol. Vis. Sci. 38, 1241e1260.
Delaey, C., Van De Voorde, J., 2000. Regulatory mechanisms in the retinal andchoroidal circulation. Ophthalmic Res. 32, 249e256.
Dhillon, B., Nickla, D., 2008. The ocular growth inhibition effected by dopamineagonists and atropine is associated with transient increases in choroidalthickness in chicks. Invest. Ophthalmol. Vis. Sci. E-Abstract 1732.
Epstein, M.L., Davis, J.P., Gellman, L.E., Lamb, J.R., Dahl, J.L., 1988. Cholingergicneurons of the chicken ciliary ganglion contain somatostatis. Neuroscience 25,1053e1060.
Fine, B.S., Yanoff, M., 1979. Ocular Histology. Harper & Row, Hagerstown, MD,pp. 195.
Fischer, A., Miethke, P., Morgan, I., Stell, W., 1998. Cholinergic amacrine cells arenot required for the progression and atropine-mediated suppression of formdeprivation myopia. Brain Res. 794, 48e60.
Fischer, A., McGuire, J., Schaeffel, F., Stell, W., 1999a. Light and defocus-dependentexpression of the transcription factor ZENK in the chick retina. Nat. Neurosci. 2,706e712.
Fischer, A., Wallman, J., Mertz, J., Stell, W., 1999b. Localization of retinoid bindingproteins, retinoid receptors, and retinaldehyde dehydrogenase in the chick eye.J. Neurocytol 28, 597e609.
Fischer, A., Stell, W., 1999. Nitric oxide synthase-containing cells in the retina,pigmented epithelium, choroid, and sclera of the chick eye. J. Comp. Neurol.405, 1e14.
Fitzgerald, M.E.C., Vana, B.A., Reiner, A., 1990. Control of choroidal blood flow bythe nucleus of Edinger-Westphal in pigeons: a laser doppler study. Invest.Ophthalmol. Vis. Sci. 31, 2483e2492.
Fitzgerald, M., Gamlin, P., Zagvazdin, Y., Reiner, A., 1996. Central neural circuits forthe light-mediated reflexive control of choroidal blood flow in the pigeon eye.Vis. Neurosci. 13, 655e669.
Flugel, C., Tamm, E.R., Mayer, B., Lutjen-Drecoll, E., 1994. Species differences inchoroid vasodilative innervation: evidence for specific intrinsic nitrergic andVIP-positive neurons in the human eye. Invest. Ophthalmol. Vis. Sci. 35,592e599.
Flugel-Koch, C., May, A., Lutjen-Drecoll, E., 1996. Presence of a contractile cellnetwork in the human choroid. Ophthalmologica 210, 296e302.
Forrester, J.V., Dick, A.D., McMenamin, P.G., Lee, W.R., 2002. The Eye. Basic Sciencesin Practice. Saunders, Edinburgh.
Geiser, M.H., Riva, C., Dorner, G.T., Diermann, U., Luksch, A., Schmetterer, L., 2000.Response of choroidal blood flow in the foveal region to hyperoxia andhyperoxia-hypercapnia. Curr. Eye Res. 21, 669e676.
Gherezghiher, T., Hey, J., Koss, M., 1990. Parasympathetic nervous control of intra-ocular pressure. Exp. Eye Res. 50, 457e462.
Gienc, J., Kuder, T., 1985. Morphology and topology of the pterygopalatine ganglionin pigeons. Zool. Pol 32, 229e235.
Goldstein, I.M., Ostwald, P., Roth, S., 1996. Nitric oxide: a review of its role in retinalfunction and disease. Vis. Res. 36, 2979e2994.
Gomez, D.G., Manzo, R.P., Fenstermacher, J.D., Potts, D.G., 1988. Cerebrospinal fluidabsorption in the rabbit. Graefe's Arch. Clin. Exp. Ophthalmol. 266, 17.
Gottlieb, M.D., Joshi, H.B., Nickla, D.L., 1990. Scleral changes in chicks with formdeprivation myopia. Curr. Eye Res. 9, 1157e1165.
Gray, D.B., Pilar, G.R., Ford, M.J., 1989. Opiate and peptide inhibition of transmitterrelease in p- arasy.m- pathetic nerve terminals. J. Neurosci. 9, 1683e1692.
Guglielmone, R., Cantino, D., 1982. Autonomic innervation of the ocular choroidmembrane in the chicken. Cell Tissue Res. 222, 417e431.
Guo, S.S., Sivak, J.G., Callender, M.G., Diehljones, B., 1995. Retinal dopamine andlens-induced refractive errors in chicks. Curr. Eye Res. 14, 385e389.
Haddad, A., Laicine, E.M., Tripathi, B.J., Tripathi, R.C., 2001. An extensive system ofextravascular smooth muscle cells exists in the choroid of the rabbit eye.Exp. Eye Res. 73, 345e353.
Hirata, A., Negi, A., 1998. Morphological changes of choriocapillaris in experi-mentally induced chick myopia. Graefe's Arch. Clin. Exp. Ophthalmol. 236,132e137.
Hirose, A., Azuma, H., Tokoro, T., Hirai, K., 2007. Endothelin-B receptors on supra-choroidal melanocytes mediate and endothelin-1-induced increase in theintracellular calcium concentration of rabbit ocular suprachoroidal tissue. Curr.Eye Res. 32, 585e591.
Hogan, M.J., Alvarado, J.A., Weddell, J.E., 1971. Histology of the Human Eye. SaundersCompany, Philadelphia.
Holzer, P., 1988. Local effector functions of capsaicin-sensitive sensory nerveendings: involvement of tachykinins, calcitonin gene-related peptide and otherneuropeptides. Neuroscience 24, 739e768.
Howlett, M., McFadden, S., 2009. Spectacle lens compensation in the pigmentedguinea pig. Vis. Res. 49, 219e227.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 165

Hu,W., Criswell, M.H., Fong, S.L., Temm, C.J., Rajashekhar, G., Cornell, T.L., Clauss, M.A.,2009. Differences in the temporal expression of regulatory growth factors duringchoroidal neovascular development. Exp. Eye Res. 88, 79e91.
Hung, L.-F., Wallman, J., Smith, E., 2000. Vision-dependent changes in the choroidalthickness of Macaque monkeys. Invest. Ophthalmol. Vis. Sci. 41, 1259e1269.
Irving, E.L., Sivak, J.G., Callender, M.G., 1992. Refractive plasticity of the developingchick eye. Ophthal. Physiol. Opt. 12, 448e456.
Iuvone, P.M., Tigges, M., Stone, R.A., Lambert, S., Laties, A.M., 1991. Effects ofapomorphine, a dopamine receptor agonist, on ocular refraction and axial elon-gation in a primate model of myopia. Invest. Ophthalmol. Vis. Sci. 32, 1674e1677.
Janssen, A., Hoellenriegel, J., Fogarasi, M., Schrewe, H., Seeliger, M., Tamm, E.,Ohlmann, A., May, C.A., Weber, B.H., Stohr, H., 2008. Abnormal vessel formationin the choroid of mice lacking tissue inhibitor of metalloprotease-3. Invest.Ophthalmol. Vis. Sci. 49, 2812e2822.
Jobling, A.I., Wan, R., Gentle, A., Bui, B.V., McBrien, N., 2009. Retinal and choroidalTGF-B in the tree shrew model of myopia: isoform expression, activation andeffects on function. Exp. Eye Res. 88, 458e466.
Junghans, B.M., Crewther, S.G., Crewther, D.P., 1998. Choroidal lympthatics: anactive storage reservoir? Invest. Ophthalmol. Vis. Sci. 39, S504.
Junghans, B., Crewther, S., Liang, H., Crewther, D., 1999. A role for choroidallymphatics during recovery from form deprivation myopia? Optom. Vis. Sci. 76,796e803.
Kajikawa, J., 1923. Beiträge zur Anatomie und Physiologie des Vogelauges. Albrechtvon Graefes Arch. Ophthalmol. 112, 260e346.
Kee, C.S., Marzani, D., Wallman, J., 2001. Differences in time course and visualrequirements of ocular responses to lenses and diffusers. Invest. Ophthalmol.Vis. Sci. 42, 575e583.
Kee, C.S., Hung, L.F., Qiao-Grider, Y., Ramamirtham, R., Winawer, J., Wallman, J.,Smith, E.L., 2007. Temporal constraints on experimental emmetropization ininfant monkeys. Invest. Ophthalmol. Vis. Sci. 48, 957e962.
Kennedy, R.H., 1995. Progression of myopia. Trans. Am. Ophthalmol. Soc. 93,755e800.
Kennedy, R.H., Dyer, J.A., Kennedy, M.A., Parulkar, S., Kurland, L.T., Herman, D.C.,McIntire, D., Jacobs, D., Luepker, R.V., 2000. Reducing the progression of myopiawith atropine: a long term cohort study of Olmsted County students. Binocul.Vis. Strabismus Q. 15, 281e304.
Kiel, J.W., Shepherd, A.P., 1992. Autoregulation of choroidal blood flow in the rabbit.Invest. Ophthalmol. Vis. Sci. 33, 2399e2410.
Kirby, M.L., Diab, I.M., Mattio, T.G., 1978. Development of adrenergic innervation ofthe iris and fluorescent ganglion cells in the choroid of the chick eye. Anat. Rec.191, 311e320.
Krebs, W., Krebs, I.P., 1988. Ultrastructural evidence for lymphatic capillaries in theprimate retina. Arch. Ophthal 106, 1615e1616.
Krebs, W., Krebs, I., 1991. Primate Retina and Choroid: Atlas of Fine Structure in Manand Monkey. Springer-Verlag, New York.
Kusakari, T., Sato, T., Tokoro, T., 2001. Visual deprivation stimulates the exchange ofthe fibrous sclera into the cartilaginous sclera in chicks. Exp. Eye Res. 73,533e546.
Leech, E.M., Cottriall, C.L., McBrien, N., 1995. Pirenzapine prevents formdeprivation myopia in a dose dependent manner. Ophthalmic Physiol. Opt.15, 351e356.
Li, X.X., Schaeffel, F., Kohler, K., Zrenner, E., 1992. Dose-dependent effects of6-hydroxy dopamine on deprivation myopia, electroretinograms, and dopa-minergic amacrine cells in chickens. Vis. Neurosci. 9, 483e492.
Liang, H., Crewther, S., Crewther, D., Junghans, B., 2004. Structural and elementalevidence for edema in the retina, retinal pigment epithelium, and choroidduring recovery from experimentally induced myopia. Invest. Ophthalmol. Vis.Sci. 45, 2463e2474.
Lin, L.L., 1996. Epidemiological study of ocular refractions among school children(aged 6 through 18) in Taiwan. Invest. Ophthalmol. Vis. Sci. 37, S1002.
Lind, G.J., Chew, S.J., Marzani, D., Wallman, J., 1998. Muscarinic acetylcholinereceptor antagonists inhibit chick scleral chondrocytes. Invest. Ophthalmol.Vis. Sci. 39, 2217e2231.
Linsenmeier, R.A., Braun, R.D., 1992. Oxygen distribution and consumption in the catretina during normoxia and hypoxia. J. Gen. Physiol. 99, 177e197.
Linsenmeier, R.A., Goldstick, T.K., Blum, R.S., Enroth-Cugell, C., 1981. Estimation ofretinal oxygen transients from measurments made in the vitreous humor. Exp.Eye Res. 32, 369e379.
Liu, O.,Wu, J.,Wang, X., Zeng, J., 2007. Changes in themuscarinic acetylcholine receptorexpression in form deprivation myopia in guinea pigs. Mol. Vis. 13, 1234e1244.
van der Loop, F.T., Schaart, G., Timmer, E.D., Ramaekers, F.C., van Eys, G.J., 1996.Smoothelin, a novel cytoskeletal protein specific for smooth muscle cells. J. CellBiol. 134, 401e411.
Lovasik, J.V., Kergoat, H., Riva, C., Petrig, B.L., Geiser, M., 2003. Choroidal blood flowduring exercise-induced changes in the ocular perfusion pressure. Invest.Ophthalmol. Vis. Sci. 44, 2126e2132.
Luft, W., Ming, Y., Stell, W., 2003. Variable effects of previously untested muscarinicreceptor antagonists on experimental myopia. Invest. Ophthalmol. Vis. Sci. 44,1330e1338.
Lutty, G.A., Merges, C., Threlkeld, A.B., Crone, S., McLeod, D.S., 1993. Heterogeneity inlocalization of isoforms of TGF-beta in human retina, vitreous, and choroid.Invest. Ophthalmol. Vis. Sci. 34, 477e487.
Mammoto, A., Connor, K.M., Mammoto, T., Yung, C.W., Huh, D., Aderman, C.M.,Mostoslavsky, G., Smith, L.E., Ingber, D.E., 2009. A mechanosensitive tran-scriptional mechanism that controls angiogenesis. Nature 457, 1103e1108.
Marmor, M.F., Abdul-Rahim, A.S., Cohen, S.D., 1980. The effect of metabolic inhibi-tors on retinal adhesion and subretinal fluid reabsorbtion. Invest. Ophthalmol.Vis. Sci. 19, 893e903.
Marzani, D., Wallman, J., 1997. Growth of the two layers of the chick sclera ismodulated reciprocally by visual conditions. Invest. Ophthalmol. Vis. Sci. 38,1726e1739.
May, C.A., 2003. Nonvascular smooth muscle alpha-actin positive cells in thechoroid of higher primates. Curr. Eye Res. 1, 1e6.
May, C.A., 2005. Non-vascular smooth muscle cells in the human choroid: distri-bution, development and further characterization. J. Anat. 207, 381e390.
May, C.A., Hayreh, S., Furuyoshi, N., Ossoinig, K., Kaufman, P.L., Lutjen-Drecoll, E.,1997. Choroidal ganglion cell plexus and retinal vasculature in monkeys withlaser-induced glaucoma. Ophthalmologica 211, 161e171.
May, C.A., Neuhuber, W., Lutjen-Drecoll, E., 2004. Immunohistochemical classifi-cation and functional morphology of human choroidal ganglion cells. Invest.Ophthalmol. Vis. Sci. 45, 361e367.
McBrien, N.A., Moghaddam, H.O., New, R., Williams, L.R., 1993a. Experimentalmyopia in a diurnal mammal (Sciurus carolinensis) with no accommodativeability. J. Physiol. 469, 427e441.
McBrien, N.A., Moghaddam, H.O., Reeder, A.P., 1993b. Atropine reduces experi-mental myopia and eye enlargement via a nonaccommodative mechanism.Invest. Ophthal. Vis. Sci. 34, 205e215.
McBrien, N., Lawlor, P., Gentle, A., 2000. Scleral remodeling druing the developmentof and recovery from axial myopia in the tree shrew. Invest. Ophthalmol. Vis.Sci. 41, 3713e3719.
McBrien, N., Sahebjada, S., Arumugam, B., Chow, A., Jobling, A.I., Gentle, A., 2009.Role of M4 receptor antagonist in control of myopic eye growth. Invest.Ophthalmol. Vis. Sci. E-Abstract #3850.
McCarthy, C.S., Megaw, P., Devadas, M., Morgan, I., 2007. Dopaminergic agents affectthe ability of brief periods of normal vision to prevent form-deprivationmyopia. Exp. Eye Res. 84, 100e107.
McFadden, S., Howlett, M., Mertz, J., 2004. Retinoic acid signals the direction ofocular elongation in the guinea pig eye. Vis. Res. 44, 643e653.
McKanna, J.A., Casagrande, V.A., 1981. Atropine affects lid-suture myopia develop-ment: experimental studies of chronic atropinization in tree shrews. Doc.Ophthalmol. 28, 187e192.
McLeod, D.S., Grebe, R., Bhutto, I., Merges, C., Baba, T., Lutty, G.A., 2009. Relationshipbetween RPE and choriocapillaris in age-related macular degeneration. Invest.Ophthalmol. Vis. Sci. 50, 4982e4991.
Melis, M.R., Succu, S., Argiolas, A., 1996. Dopamine agonists increase nitric oxideproduction in the paraventricular nucleus of the hypothalamus: correlationwith penile erection and yawning. Eur. J. Neurosci. 8, 2056e2063.
Meriney, S., Pilar, G., 1987. Cholinergic innervation of the smooth muscle cells in thechoroid coat of the chick eye and its development. J. Neurosci. 7, 3827e3839.
Mertz, J.R., Wallman, J., 2000. Choroidal retinoic acid synthesis: a possible mediatorbetween refractive error and compensatory eye growth. Exp. Eye Res. 70,519e527.
Miller, A.S., Coster, D.J., Costa,M., Furness, J.B.,1983. Vasoactive intestinal polypeptideimmunoreactive nerve fibers in the human eye. Aust. J. Ophthalmol. 11,185e193.
Moring, A.G., Baker, J.R., Norton, T., 2007. Modulation of glycosaminoglycan levels inthe tree shrew sclera during lens-induced myopia development and recovery.Invest. Ophthalmol. Vis. Sci. 48, 2947e2956.
Mueller, H., 1859. Ueber glatte Muskeln und Nervengeflechte der chorioidea immenschlichen Auge. Vehr Physik-med Ges Wurzburg 10, 179e192.
Nakanome, Y., Karita, K., Izumi, H., Tamai, M., 1995. Two types of vasodilation in catchoroid elicited by electrical stimulation of the short ciliary nerve. Exp. Eye Res.60, 37e42.
Napper, G.A., Brennan, N.A., Barrington, M., Squires, M.A., Vessey, G.A., Vingrys, A.J.,1995. The duration of normal visual exposure necessary to prevent formdeprivation myopia in chicks. Vis. Res. 35, 1337e1344.
Nickel, R., Schummer, A., Seiferle, E., 1977. Anatomy of the Domestic Birds. Springer-Verlag, New York.
Nickla, D., 2006. The phase relationships between the diurnal rhythm in axiallength and choroidal thickness and the association with ocular growth rate inchicks. J. Comp. Physiol. A 192, 399e407.
Nickla, D., 2007. Transient increases in choroidal thickness are consistently asso-ciated with brief daily visual stimuli that inhibit ocular growth in chicks.Exp. Eye Res. 84, 951e959.
Nickla, D.L., Cheng, A.T., 2009. Atropine and apomorphine in emmetropization:evidence for inhibitory interactions on a common growth regulatory pathway.Invest. Ophthalmol. Vis. Sci. E-Abstract 3848.
Nickla, D.L., Wildsoet, C., Wallman, J., 1997. Compensation for spectacle lensesinvolves changes in proteoglycan synthesis in both the sclera and choroid. Curr.Eye Res. 16, 320e326.
Nickla, D.L., Wildsoet, C., Wallman, J., 1998. Visual influences on diurnal rhythms inocular length and choroidal thickness in chick eyes. Exp. Eye Res. 66, 163e181.
Nickla, D.L., Rada, J.A., Wallman, J., 1999. Isolated chick sclera shows a circadianrhythm in proteoglycan synthesis perhaps associated with the rhythm in ocularelongation. J. Comp. Physiol. [A] 185, 81e90.
Nickla, D., Wildsoet, C., Troilo, D., 2001. Endogenous rhythms in axial length andchoroidal thickness in chicks: implications for ocular growth regulation. Invest.Ophthalmol. Vis. Sci. 42, 584e588.
Nickla, D., Wildsoet, C., Troilo, D., 2002. Diurnal rhythms in intraocular pressure,axial length, and choroidal thickness in a primate model of eye growth, thecommon marmoset. Invest. Ophthalmol. Vis. Sci. 43, 2519e2528.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168166

Nickla, D., Wilken, E., Lytle, G., Yom, S., Mertz, J., 2006. Inhibiting the transientchoroidal thickening response using the nitric oxide synthase inhibitor L-NAMEprevents the ameliorative effects of visual experience on ocular growth in twodifferent visual paradigms. Exp. Eye Res. 83, 456e464.
Nickla, D., Damyanova, P., Lytle, G., 2009. Inhibiting the neuronal form of nitricoxide synthase has similar effects on the compensatory choroidal and axialresponses to myopic defocus in chicks as does the non-specific inhibitor L-NAME. Exp. Eye Res. 88, 1092e1099.
Nickla, D., Mertz, J., 2003. Inhibiting the recovery from form deprivation myopiapharmacologically results in decreased choroidal all-trans-retinoic acidsynthesis. Invest. Ophthalmol. Vis. Sci. E-Abstract, 2806.
Nickla, D., Wildsoet, C., 2004. The effect of the nonspecific nitric oxide synthaseinhibitor NG-nitro-L-arginine methyl ester on the choroidal compensatoryresponse to myopic defocus in chickens. Optom. Vis. Sci. 81, 111e118.
Nilsson, S.F., 1996. Nitric oxide as a mediator of parasympathetic vasodilation inocular and extraocular tissues in the rabbit. Invest. Ophthalmol. Vis. Sci. 37,2110e2119.
Nishi, R., 1994. Target-derived molecules that influence the development of neuronsin the avian ciliary ganglion. J. Neurobiol. 25, 612e619.
Norton, T., 1999. Animal models of myopia: learning how vision controls the size ofthe eye. Instit. Lab. Anim. Res. 40, 59e77.
Norton, T., Rada, J., 1995. Reduced extracellular matrix in mammalian sclera withinduced myopia. Vis. Res., 1271e1281.
Papastergiou, G.I., Schmid, G.F., Riva, C.E., Mendel, M.J., Stone, R.A., Laties, A.M.,1998. Ocular axial length and choroidal thickness in newly hatched chicks andone-year-old chickens fluctuate in a diurnal pattern that is influenced by visualexperience and intraocular pressure changes. Exp. Eye Res. 66, 195e205.
Park, T., Winawer, J., Wallman, J., 2003. Further evidence that chicks use the sign ofblur in spectacle lens compensation. Vis. Res. 43, 1519e1531.
Parver, L.M., 1991. Temperature modulating action of choroidal blood flow. Eye 5,181e185.
Parver, L.M., Auker, C., Carpenter, D.O., 1980. Choroidal blood flow as a heat dissi-pating mechanism in the macula. Am. J. Ophthalmol. 89, 641e646.
Parver, L.M., Auker, C., Carpenter, D.O., 1982a. The stabilizing effect of the choroidalcirculation on the temperature environment of the macula. Retina 2, 117e120.
Parver, L.M., Auker, C., Carpenter, D.O., Doyle, T., 1982b. Choroidal blood flow II.Reflexive control in the monkey. Arch. Ophthal 100, 1327e1330.
Parver, L.M., Auker, C., Carpenter, D.O., 1983. Choroidal blood flow III. Reflexivecontrol in human eyes. Arch. Ophthal 101, 1604e1606.
Pendrak, K., Nguyen, T., Lin, T., Capehart, C., Zhu, X.S., Stone, R.A., 1997. Retinaldopamine in the recovery from experimental myopia. Curr. Eye Res. 16, 152e157.
Pendrak, K., Papastergiou, G.I., Lin, T., Laties, A.M., Grimes, P.A., Stone, R.A., 1998.Origin of fluid in chick choroidal lacunae. Invest. Ophthalmol. Vis. Sci. 39, S504.
Pendrak, K., Papastergiou, G.I., Lin, T., Laties, A.M., Stone, R.A., 2000. Choroidalvascular permeability in visually regulated eye growth. Exp. Eye Res. 70,629e637.
Pettigrew, J.D., Wallman, J., Wildsoet, C., 1990. Saccadic oscillations facilitate ocularperfusion from the avian pecten. Nature 343, 362e363.
Polska, E., Simader, C., Weigert, G., Doelemeyer, A., Kolodiaschna, J., Scharmann, O.,Schmetterer, L., 2007. Regulation of choroidal blood flow during combinedchanges in intraocular pressure and arterial blood pressure. Invest. Ophthalmol.Vis. Sci. 48, 3768e3774.
Poukens, V., Glasgow, B.J., Demer, J.L., 1998. Nonvascular contractile cells in scleraand choroid of humans and monkeys. Invest. Ophthalmol. Vis. Sci. 39,1765e1774.
Rada, J.A., Thoft, R.A., Hassell, J.R., 1991. Increased aggrecan (cartilage proteoglycan)production in the sclera of myopic chicks. Dev. Biol. 147, 303e312.
Rada, J.A., McFarland, A.L., Cornuet, P.K., Hassell, J.R., 1992. Proteoglycan synthesis byscleral chondrocytes is modulated by a vision dependent mechanism. Curr. EyeRes. 11, 767e782.
Rada, J.A., Nickla, D.L., Troilo, D., 2000. Decreased proteoglycan synthesis associatedwith form deprivation myopia in mature primate eyes. Invest. Ophthalmol. Vis.Sci. 41, 2050e2058.
Rada, J., Huang, Y., Rada, K., 2001. Identification of choroidal ovotransferrin asa potential ocular growth regulator. Curr. Eye Res. 22, 121e132.
Rada, J.A., Matthews, A.L., 1994. Visual deprivation upregulates extracellular matrixsynthesis by chick scleral chondrocytes. Invest. Ophthalmol. Vis. Sci. 35,2436e2447.
Rada, J., Wiechmann, A., 2009. Ocular expression of avian thymic hormone: changesduring the recovery from induced myopia. Mol. Vis. 15, 778e792.
Ramrattan, R.S., van der Schaft, T.L., Mooy, C.M., de Bruijn, W.C., Mulder, P.G.H.,de Jong, P., 1994. Morphometric analysis of Bruch's membrane, the chorio-capillaris and the choroid in aging. Invest. Ophthalmol. Vis. Sci. 35,2857e2864.
Raviola, E., Wiesel, T.N., 1985. An animal model of myopia. N. Engl. J. Med. 312,1609e1615.
Reiner, A., 1987. The presence of substance P/CGRP-containing nerve fibers, VIP-containing fibers and numerous cholinergic fibers on blood vessels of the avianchoroid. Invest. Ophthalmol. Vis. Sci. 28, S81.
Reiner, A., Karten, H.J., Gamlin, P.D.R., Erichsen, J.T., 1983. Parasympathetic ocularcontrol: functional subdivisions and circuitry of the avian nucleus of Edinger-Westphal. Trends Neurosci. 6, 140e145.
Reiner, A., Erichsen, J., Cabot, J.B., Evinger, C., Fitzgerald, M., Karten, H.J., 1991.Neurotransmitter organization of Edinger-Westphal and its projection to theavian ciliary ganglion. Vis. Neurosci. 6, 451e472.
Reiner, A., Zagvazdin, Y., Fitzgerald, M., 2003. Choroidal blood flow in pigeonscompensates for decreases in arterial blood pressure. Exp. Eye Res. 76,273e282.
Remington, L.A., 2005. Clinical Anatomy of the Visual System, second ed. Elsevier.Riva, C., Titze, T., Hero, M., Movaffaghy, A., Petrig, B.L., 1997a. Choroidal blood flow
during isometric exercises. Invest. Ophthalmol. Vis. Sci. 38, 2338e2343.Riva, C., Titze, T., Hero, M., Petrig, B.L., 1997b. Effect of acute decreases of perfusion
pressure on choroidal blood flow in humans. Invest. Ophthalmol. Vis. Sci. 38,1752e1760.
Rohrer, B., Spira, A.W., Stell, W.K., 1993. Apomorphine blocks form-deprivationmyopia in chickens by a dopamine D2-receptor mechanism acting in retina orpigmented epithelium. Vis. Neurosci. 10, 447e453.
Rohrer, B., Stell, W.K., 1994. Basic fibroblast growth-factor (Bfgf) and TransformingGrowth-Factor-Beta (TGF-Beta) act as stop and go signals to modulate postnatalocular growth in the chick. Exp. Eye Res. 58, 553e561.
Rucker, F.J., Wallman, J., 2008. Cone signals for spectacle lens compensation:differential responses to short and long wavelengths. Vis. Res. 48, 1980e1991.
Ruskell, G.L., 1971. Facial parasympathetic innervation of the choroidal blood vesselsin monkeys. Exp. Eye Res. 12, 166e172.
Rymer, J., WIldsoet, C.F., 2005. The role of the retinal pigment epithelium in eyegrowth regulation and myopia: a review. Vis. Neurosci. 22, 251e261.
Schaeffel, F., Troilo, D., Wallman, J., Howland, H.C., 1990. Developing eyes that lackaccommodation grow to compensate for imposed defocus. Vis. Neurosci. 4,177e183.
Schaeffel, F., Hagel, G., Bartmann, M., Kohler, K., 1994. 6-hydoxy dopamine does notaffect lens-induced refractive errors but suppresses deprivation myopia. Vis.Res. 34, 143e149.
Schaeffel, F., Bartmann, M., Hagel, G., Zrenner, E., 1995. Studies on the role of theretinal dopamine/melatonin system in experimental refractive errors inchickens. Vis. Res. 35, 1247e1264.
Schmid, K., Wildsoet, C.F., 1996. Effects of the compensatory responses to positiveand negative lenses of intermittent lens wear and ciliary nerve section inchicks. Vis. Res. 36, 1023e1036.
Schmid, K.L., Wildsoet, C.F., 2004. Inhibitory effects of apomorphine and atropineand their combination on myopia in chicks. Invest. Ophthalmol. Vis. Sci. 81,137e147.
Schrödl, F., 2009. Intrinsic choroidal neurons. In: Troger, J., Kieselbach, G., Bechrakis, N.(Eds.), Neuropeptides in the Eye. Research Signpost, Kerala, pp. 169e197.
Schrödl, F., De Laet, A., Tassignon, M.J., Van Bogaert, P., Brehmer, A., Neuhuber, W.,Timmermans, J.P., 2003. Intrinsic choroidal neurons in the human eye:projections, targets, and basic electrophysiological data. Invest. Ophthalmol.Vis. Sci. 44, 3705e3712.
Schrödl, F., Brehmer, A., Neuhuber, W., Kruse, F.E., May, C.A., Cursiefen, C., 2008. Thenormal human choroid is endowed with a significant number of lymphaticvessel endoethelial hyaluronate receptor 1(LYVE-1)-positive macrophages.Invest. Ophthalmol. Vis. Sci. 49, 5222e5229.
Schrödl, F., Bergua, A., Brehmer, A., Neuhuber, W.L., 1998. Nonvascular smoothmuscleas a main target for intrinsic choroid neurons in duck. Exp. Eye Res. 67, 78.
Schrödl, F., Brehmer, A., Neuhuber, W., 2000. Intrinsic choroidal neurons in the duckeye express galanin. J. Comp. Neurol. 425, 24e33.
Schrödl, F., Schweigert, M., Brehmer, A., Neuhuber, W.L., 2001a. Intrinsic neurons inthe duck choroid are contacted by CGRP-immunoreactive nerve fibers:evidence for a local pre-central reflex arc in the eye. Exp. Eye Res. 72, 137e146.
Schrödl, F., Tines, R., Brehmer, A., Neuhuber, W., 2001b. Intrinsic choroidal neuronsin the duck eye receive sympathetic input: anatomical evidence for adrenergicmodulation of nitrergic functions in the choroid. Cell Tissue Res. 304, 175e184.
Schrödl, F., De Stefano, M., Reese, S., Brehmer, A., Neuhuber, W., 2004. Comparativeanatomy of nitrergic intrinsic choroidal neurons (ICN) in various avian species.Exp. Eye Res. 78, 187e196.
Schrödl, F., Brehmer, A., Neuhuber, W., Nickla, D., 2006. The autonomic facial nervepathway in birds: a tracing study. Invest. Ophthalmol. Vis. Sci. 47, 3225e3233.
Schwahn, H.N., Kaymak, H., Schaeffel, F., 2000. Effects of atropine on refractivedevelopment, dopamine release, and slow retinal potentials in the chick. Vis.Neurosci. 17, 165e176.
Seko, Y., Shimokawa, H., Tokoro, T., 1995. Expression of bFGF and TGF-B2 inexperimental myopia in chicks. Invest. Ophthalmol. Vis. Sci. 36, 1183e1187.
Shaikh, A.W., Siegwart, J.T., Norton, T., 1999. Effect of interrupted lens wear oncompensation for a minus lens in tree shrews. Optom. Vis. Sci. 76, 308e315.
Sherman, S.M., Norton, T.T., Casagrande, V.A., 1977. Myopia in the lid-sutured treeshrew (Tupaia glis). Brain Res. 124, 154e157.
Shih, Y.F., Chen, C.H., Chou, A.C., Ho, T.C., Lin, L.L., Hung, P.T., 1999a. Effects ofdifferent concentrations of atropine on controlling myopia in myopic children.J. Ocul. Pharm. Therapeut 15, 85e90.
Shih, Y.F., Fitzgerald, M., Cuthbertson, S.L., Reiner, A., 1999b. Influence of opthalmicnerve fibers on choroidal blood flow and myopic eye growth in chicks. Exp. EyeRes. 69, 9e20.
Simon, P., Feldkaemper, M., Bitzer, M., Ohngemach, S., Schaeffel, F., 2004. Earlytranscriptional changes of retinal and choroidal TGFB-2, RALD-2, and ZENKfollowing imposed positive and negative defocus in chickens. Mol. Vis. 10,588e597.
Smalley, D.M., Fitzgerald, J.E., Taylor, D.M., O'Rourke, J., 1994. Tissue plasminogenactivator in human aqueous humor. Invest. Ophthalmol. Vis. Sci. 35, 48e53.
Snyder, S.H., Bredt, D., 1992. Biological roles of nitric oxide. Scient. Amer. 266, 68e77.Sperduto, R.D., Seigel, D., Roberts, J., Rowland, M., 1983. Prevalence of myopia in the.
United States. Arch. Ophthalmol. 101, 405e407.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168 167

Spraul, C.W., Lang, G.E., Grossniklaus, H.E., Lang, G.K., 1999. Histologic andmorphometric analysis of the choroid, Bruch's membrane, and retinal pigmentepithelium in postmortem eyes with age-related macular degeneration andhistologic examination of surgically excised choroidal neovascular membranes.Surv. Ophthalmol. 44, 10e32.
Stewart, A.G., Phan, L., Grigoriadis, G., 1994. Physiological and pathophysiologicalroles of nitric oxide. Microsurgery 15, 693e702.
Stjernschantz, J., Alm, A., Bill, A., 1976. Effects of intracranial oculomotor nervestimulation on ocular blood flow in rabbits: modification by indomethacin.Invest. Ophthalmol. Vis. Sci. 18, 99e103.
Stone, R.A., 1985. Substance P-like immunoreactive nerves in the human eye. Arch.Ophthalmol. 103, 1207e1211.
Stone, R.A., 1986. Vasoactive intestinal polypeptide and the ocular innervation.Invest. Ophthalmol. Vis. Sci. 27, 951e957.
Stone, R.A., Kuwayama, Y., Laties, A.M., 1987. Regulatory peptides in the eye.Experentia 43, 791e800.
Stone, R.A., Lin, T., Laties, A.M., Iuvone, P.M., 1989. Retinal dopamine and form-deprivation myopia. Proc. Nat. Acad. Sci. 86, 704e706.
Stone, R.A., Lin, T., Laties, A.M., 1991. Muscarinic antagonist effects on experimentalchick myopia. Exp. Eye Res. 52, 755e7588.
Stone, R.A., Quinn, G.E., Francis, E.L., Ying, G., Flitcroft, D.I., Parekh, P., Brown, J.,Orlow, J., Schmid, G., 2004. Diurnal axial length fluctuations in human eyes.Invest. Ophthalmol. Vis. Sci. 45, 63e70.
Stone, R.A.,McGlinn, A.M.,1988. Calcitonin gene-relatedpeptide immunoreactive nervesin human and rhesus monkey eyes. Invest. Ophthalmol. Vis. Sci. 29, 305e310.
Sugita, A., Inokuchi, T., 1992. Lymphatic sinus-like structures in choroid. Jpn.J. Ophthalmol. 36, 436e442.
Summers Rada, J., Folger, R.H., 2009. Biphasic scleral response during the recoveryform induced myopia: is the choroid involved? Invest. Ophthalmol. Vis. Sci. E-Abstact 3930.
Summers Rada, J., Palmer, L., 2007. Choroidal regulation of scleral glycosamino-glycan synthesis during recovery from induced myopia. Invest. Ophthalmol. Vis.Sci. 48, 2957e2966.
Sun, W., Erichsen, J.T., May, P., 1994. NADPH-diaphorase reactivity in ciliary ganglionneurons: a comparison of distributions in the pigeon, cat and monkey. Vis.Neurosci. 11, 1e5.
Tian, Y., Wildsoet, C.F., 2006. Diurnal fluctuations and developmental changes inocular dimensions and optical aberrations in young chicks. Invest. Ophthalmol.Vis. Sci. 47, 4168e4178.
Tigges, M., Iuvone, P.M., Fernandes, A., Sugrue, M., Mallorga, P., Laties, A.M.,Stone, R., 1999. Effects of muscarinic cholinergic receptor antagonists on post-natal eye growth of rhesus monkeys. Optom. Vis. Sci. 76, 397e407.
Troilo, D., Nickla, D., Wildsoet, C., 2000. Choroidal thickness changes during alteredeye growth and refractive state in a primate. Invest. Ophthalmol. Vis. Sci. 41,1249e1258.
Troilo, D., Nickla, D.L., Mertz, J., Summers-Rada, J., 2006. Change in the synthesis ratesof ocular retinoic acid and scleral glycosaminoglycan during experimentallyaltered eye growth in marmosets. Invest. Ophthalmol. VIs. Sci. 47, 1767e1777.
Uddman, R.J., Alumets, B., Ehinger, R., Hakanson, I., Loren, I., Sundler, F., 1980.Vasoactive intestinal peptide nerves in ocular and orbital structures of the cat.Invest. Ophthalmol. Vis. Sci. 19, 878e885.
Wajer, S.D., Taomoto, M., McLeod, D.S., McCally, R.L., Nishiwaki, H., Fabry, R.L.,Lutty, G.A., 2000. Velocity measurements of normal and sickle cell red bloodcells in the rat retinal and choroidal vasculatures. Microvasc. Res. 60, 281e293.
Walcott, B., Sibony, P.A., Keyser, K.T., 1989. Neuropeptides and the innervation of theavian lacrimal gland. Invest. Ophthalmol. Vis. Sci. 30, 1666e1674.
Wallman, J., 1993. Retinal control of eye growth and refraction. In: Osbourne, N.,Chader, G. (Eds.), Progress in Retinal Research, vol. 12. Pergamon Press, Oxford,pp. 133e153.
Wallman, J., Turkel, J., Trachtman, J., 1978. Extreme myopia produced by modestchanges in early visual experience. Science 201, 1249e1251.
Wallman, J., Wildsoet, C., Xu, A., Gottlieb, M., Nickla, D., Marran, L., Krebs, W.,Christensen, A., 1995. Moving the retina: choroidal modulation of refractivestate. Vis. Res. 35, 37e50.
Wallman, J., Winawer, J., 2004. Homeostasis of eye growth and the question ofmyopia. Neuron 43, 447e468.
Walls, G.L., 1942. The Vertebrate Eye and its Adaptive Radiations. Bloomfield Hills, MI.Wang, Y., Gilliles, C., Cone, R.E., O'Rourke, J., 1995. Extravascular secretion of t-PA by
the intact superfused choroid. Invest. Ophthalmol. Vis. Sci. 36, 1625e1632.Wangsa-Wirawan, N.D., Linsenmeier, R.A., 2003. Retinal oxygen: fundamental and
clinical aspects. Arch. Ophthalmol. 121, 547e557.Watson, P.G., Young, R.D., 2004. Scleral structure, organization and disease. A
review. Exp. Eye Res. 78, 609e623.Wentzek, L.A.F., Bowers, C.W., Khairallah, L., Pilar, G., 1993. Choroid tissue supports
the survival of ciliary ganglion neurons in vitro. J. Neurosci. 13, 3143e3154.Wiesel, T.N., Raviola, E., 1977. Myopia and eye enlargement after neonatal lid fusion
in monkeys. Nature 266, 66e68.Wildsoet, C., Wallman, J., 1995. Choroidal and scleral mechanisms of compensation
for spectacle lenses in chicks. Vis. Res. 35, 1175e1194.Winawer, J., Wallman, J., 2002. Temporal constraints on lens compensation in
chicks. Vis. Res. 42, 2651e2668.Wingstrand, K., Munk, O., 1965. The pecten oculi of the pigeon with particular
regard to its function. Biol. Skr Dan Vid Selsk 14, 1e89.Wobig, J.L., 1981. The blood vessels and lymphatics of the orbit and lid. In:Wobig, J.L.,
Reeh, M.J., Wirtschafter, J.D. (Eds.), Ophthalmic Anatomy. American Academy ofOphthalmology, San Francisco, p. 77.
Yamamoto, R., Bredt, D.S., Dawson, T.M., Snyder, S.H., Stone, R.A., 1993a. Enhancedexpression of nitric oxide synthase by rat retina following pterygopalatineparasympathetic denervation. Brain Res. 631, 83e88.
Yamamoto, R., Bredt, D.S., Snyder, S.H., Stone, R.A.,1993b. The localization of nitric oxidesynthase in the rat eye and related cranial ganglia. Neuroscience 54, 189e200.
Young, F.A., 1965. The effect of atropine on the developement of myopia inmonkeys. Am. J. Optom. Arch. Am. Acad. Optom. 42, 439e449.
Yu, D.Y., Cringle, S., 2001. Oxygen distribution and consumption within the retina invascularized and avascular retinas and in animal models of retinal disease. Prog.Retin. Res. 20, 175e208.
Yücel, Y.H., Johnston, M.G., Ly, T., Patel, M., Drake, B., Gumus, E., Fraenkl, S.A.,Moore, S., Tobbia, D., Armstrong, D., Horvath, E., Gupta, N., 2009. Identificationof lymphatics in the ciliary body of the human eye: a novel “uveolymphatic”outflow pathway. Exp. Eye Res. 89, 810e819.
Zarbin, M.A., 1998. Age-related macular degeneration: review of pathogenesis.Eur. J. Ophthalmol. 8, 199e206.
Zarbin, M.A., 2004. Current concepts in the pathogenesis of age-related maculardegeneration. Arch. Ophthalmol. 122, 598e614.
Zhu, X., Park, T., Winawer, J., Wallman, J., 2005. In a matter of minutes, the eye canknow which way to grow. Invest. Ophthalmol. Vis. Sci. 46, 2238e2241.
Zhu, X., Wallman, J., 2009. Temporal properties of compensation for positive andnegative spectacle lenses in chicks. Invest. Ophthalmol. Vis. Sci. 50, 37e46.
D.L. Nickla, J. Wallman / Progress in Retinal and Eye Research 29 (2010) 144e168168





























