Embed Size (px)
Citation preview

Biological Conservation 31 ( i 985) 21-40
Phytosociology of Sinharaja A Contribution to Rain Forest Conservation in Sri Lanka
C. V. S. Gunatilleket & I. A. U. N. Gunatilleket
Arnold Arboretum of the Harvard University, 22 Divinity ~venue, Cambridge, Massachusetts 02138, USA
ABSTRACT
The structure and floristic composition of the vegetation over 30 cm girth at breast height (gbh) in five relatively undisturbed forest sites, each 5 ha in extent, in the Sinharaja International Man and Biosphere rain forest reserve investigated reveal a similarity in their (a) mean density of individuals (594-769) per ha, (b) basal area (36.0-41.6m 2) per ha and vertical distribution of individuals. The 17 427 individuals enumerated were identified into 211 species, 119 genera and 43families. Contribution of endemic species to density, basal area and floristic richness in terms of total species complement ranged between 75-93 %, 87-93 % and 64-75 % respectively.
Clusiaceae [Importance Value Index (IVI) = 27.3-78.1] and Diptero- carpaceae (IVI= 39.9-50.3) were the two most dominant families in this forest. Based on height attained by mature individuals of a species, 59 canopy, 49 subcanopy and 103 understorey tree species were discerned. In all except sample site 2, Garcinia hermonii ( Clusiaceae) followed by Xylopia championii (Annonaceae) dominated the understorey tree stratum, but in the subcanopy and canopy the dominants were more diverse.
Of the 211 species recorded, 40% had population densities of 10 individuals or less per 25 ha and 43 ~ were restricted either to one or two sites sampled. These species,particularly the endemics among them, may be considered vulnerable species and further encroachments into the reserve will jeopardise their survival. Even for those endemics once
t Permanent address: Department of Botany, University of Peradeniya, Peradeniya, Sri Lanka.
21
Biol. Conserv. 0006-3207/85/$03.30 © Elsevier Applied Science Publishers Ltd, England, 1985. Printed in Great Britain

22 C. V. S. Gunatilleke, I. A. U. N. Gunatilleke
widespread in the lowland wet zone of the &land, Sinharaja has now become the last refugium as most of their habitats have been modified, leaving a mere 9 % of the wet zone land area under natural forest cover. This too, is fragmented and inadequately protected. Conserving Sinharaja in its entirety is therefore all the more important and is highlighted in this phytosociological survey.
INTRODUCTION
The primaeval rain forests of the aseasonal perhumid southwest lowlands of Sri Lanka (Fig. 1) have an ancestry that dates back, possibly to the ancient flora of the Deccan plate (Abeywickrama, 1956). These forests are not only the most complex, but floristically the richest of the island's present day natural vegetation types and contain as much as 60% of the country's endemic flora (Abeywickrama, 1956; Peeris, 1975). Consequently they are of manifold interest to conservationists and to students of biogeography, ecology and evolutionary biology of South and Southeast Asia.
Fig. l .
S I N H A R A J A FOREST 0 4 2 km
Location of Sinharaja forest in relation to the climatic zones of Sri Lanka. 1, Arid zone; 2, Dry zone; 3, Intermediate zone; 4, Wet zone.
Rain forest conservation in Sri Lanka 23
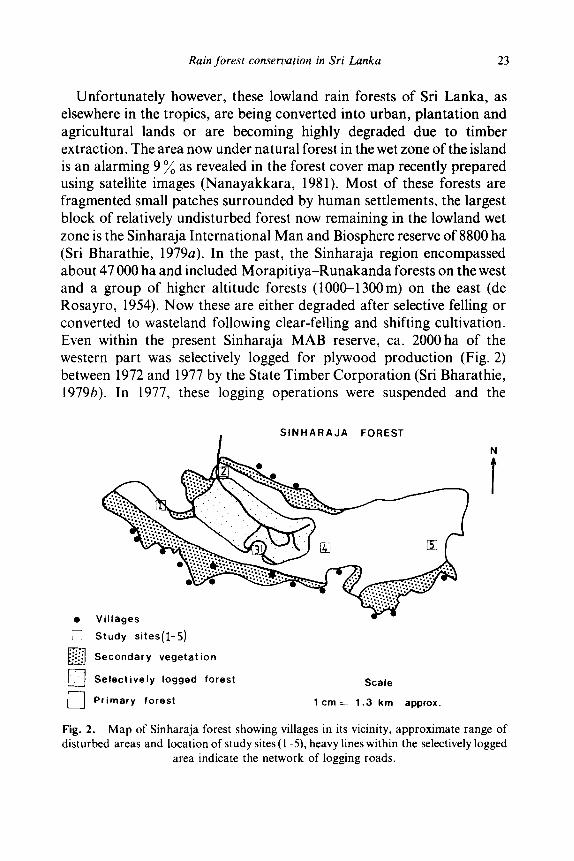
Unfortunately however, these lowland rain forests of Sri Lanka, as elsewhere in the tropics, are being converted into urban, plantation and agricultural lands or are becoming highly degraded due to timber extraction. The area now under natural forest in the wet zone of the island is an alarming 9 ~o as revealed in the forest cover map recently prepared using satellite images (Nanayakkara, 1981). Most of these forests are fragmented small patches surrounded by human settlements, the largest block of relatively undisturbed forest now remaining in the lowland wet zone is the Sinharaja International Man and Biosphere reserve of 8800 ha (Sri Bharathie, 1979a). In the past, the Sinharaja region encompassed about 47 000 ha and included Morapi t iya-Runakanda forests on the west and a group of higher altitude forests (1000-1300 m) on the east (de Rosayro, 1954). Now these are either degraded after selective felling or converted to wasteland following clear-felling and shifting cultivation. Even within the present Sinharaja MAB reserve, ca. 2000ha of the western part was selectively logged for plywood production (Fig. 2) between 1972 and 1977 by the State Timber Corporation (Sri Bharathie, 1979b). In 1977, these logging operations were suspended and the
SINHARAJA FOREST
• Villages Study sites(l- 5)
[ ] Secondary vegetation
Selectively logged forest
~ ] Primary forest
Scale
lcm ~ 1.3 km approx.
Fig. 2. Map of Sinharaja forest showing villages in its vicinity, approximate range of disturbed areas and location of study sites (I 5), heavy lines within the selectively logged
area indicate the network of logging roads.

24 C. V. S. Gunatilleke, 1. A. U. N. Gunatilleke
selectively logged part was also included in the MAB reserve. Although this status itself affords a certain amount of protection to the forest, the human impact of a large number of villages dotted round its perimeter (Fig. 2) cannot be effectively controlled by a handful of forest rangers entrusted to police the reserve. The extent of this human impact over the past several decades is quite obvious along the southern boundary of Sinharaja, where a secondary scrub vegetation prevails in place of the primary forest and the retreating tree line is clearly indicated on the recent forest cover map referred to above.
Among the earliest records on Sinharaja are those of Baker (1937, 1938), who made a reconnaissance visit and studied its climate, plants, animals and the sociology of inhabitants in villages around it. More recent publications include those of de Rosayro (1954, 1959), Andrews (1961) and Merritt & Ranatunga (1959), who studied aerial photographic maps and carried out ground surveys to determine its timber potential for selective logging on a sustained yield basis. The latter forestry-oriented studies were limited to investigations of the spatial distribution and regeneration of merchantable hardwood and firewood species and the accuracy of species identifications in these studies was not emphasised.
In the present study, the phytosociology (structure and floristic composition) of the woody vegetation over 30 cm girth at breast height (gbh) in five different sites of the forest has been examined using plot sampling (detailed below) to assess their conservation value. The results of three of these sites (site 2--Waturawa, site 3--Warukandeniya and site 4 Sinhagala) given in Gunatilleke & Gunatilleke (1981) have been incorporated in this paper so as to present a more complete description of the phytosociology of Sinharaja, now the most valuable lowland rain forest left in Sri Lanka. Phytosociological studies such as this in different parts of biosphere reserves provide base-line information, particularly on rare species, and have been strongly recommended as a prerequisite for conservation of these plant species in tropical rain forests (Ashton, 1981).
SITE DESCRIPTION AND FIELD SAMPLING METHODS
The geography, geology, climate and soils of the Sinharaja region have been described by de Rosayro (1959), Moorman & Panabokke (1961), Cooray (1967) and Gunatilleke & Gunatilleke (1983). Topographically,

Rain forest conservation in Sri Lanka 25
the area is dissected with steep slopes and valleys. An intricate network of streamlets drain the area into two streams on the northern and southern boundaries of the forest. The sampling sites in this study were located in different areas of the forest, as far apart from one another as possible, so as to obtain a reasonable representation of the entire range. These sites, numbered 1-5 from west to east respectively, have an altitudinal variation between 335 m and 915 m above mean sea level (Fig. 2 and Table 1).
At each sampling site twenty 0.25 ha plots (mostly 100 m × 25 m and occasionally 50 m x 50 m), subjectively located to maintain topographic heterogeneity within a plot at a minimum, were demarcated by using alternating bands of red and white paint on trees immediately outside the plot boundary. Thus in all, 100 plots, or 25 ha of what appeared to be undisturbed primary forest, were sampled. Usually two plots were located contiguously so that plot size could be doubled, if required during analysis. When sampling, each plot was subdivided into 25 m x 25 m subplots, and within each the girth of all individuals over 30 cm gbh were recorded. A herbarium voucher specimen from each individual was also collected for identification of the species and an aluminium number tag nailed to each so that they could be relocated when required. Although local names of species were recorded, species identifications were done entirely by comparing specimens collected with those in the National Herbarium at Peradeniya (PDA) as well as by referring to the standard floras by Trimen ( 1893-1900), Alston (1931) and Dassanayake & Fosberg (1980-1982).
RESULTS
Structure
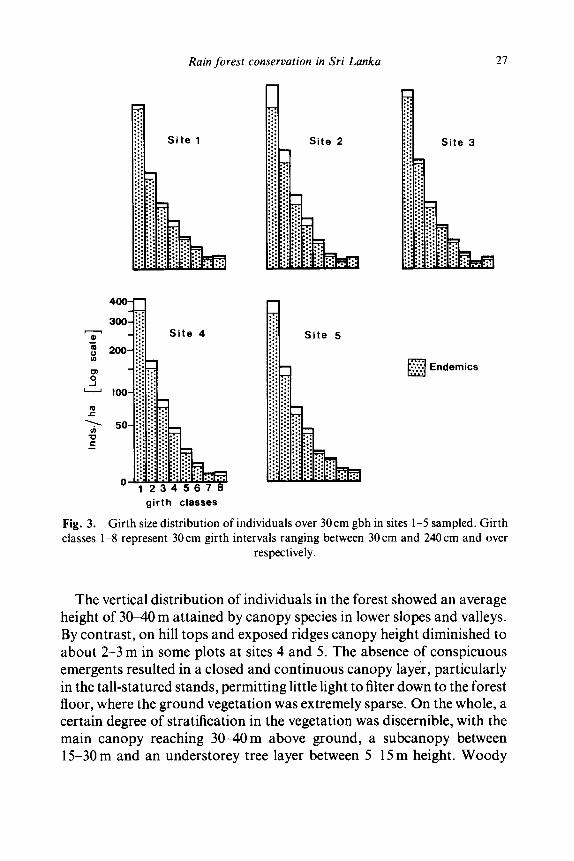
A total of 17 427 individuals over 30 cm gbh were enumerated in the 25 ha sampled in this study. While mean density and basal area in the total sample were found to be 696 individuals ha -1 and 38.7m2ha -1 respectively, the corresponding data for individual sites are given in Table 1. The girth size distribution of individuals for each site (Fig. 3) presents typical reverse 'J' curves with relatively high densities of individuals in the lower girth classes, a feature characteristic of mature tropical rain forests (Whitmore, 1975). The mean number of individuals over 150 cm gbh (the merchantable timber class) in the total sample was 50 individuals ha-1 and ranged between 45 and 56 individuals ha-1 in the different sites.

26 C. V. S. Gunatilleke, I. A. U. N. Gunatilleke
r ~ , . o
0
L)
oo
~ tg5 r',.
I'~ oo oO
tg)
tt~
" ~ ~ 0
~ e ~ e a
oO
oO
¢¢5
oo
Rain forest conservation in Sri Lanka 27
Site 1 : r :
:!: Site 2
o : , ° : ,
m
Site 3
i:i
40O-
300-
200-
O~ 0
._1
I 00
50- " 0 c
D
Fig. 3.
Site 4
i:i l : ' : I
2 3 4 5 6 7 8 girth c lasses
o:o o:o
i~i Site 5 :~ :
:~:
iii- :~ : r . !
H irlL i:~:
[ ] Endemics
Girth size distribution of individuals over 30 cm gbh in sites 1-5 sampled. Girth classes 1-8 represent 30cm girth intervals ranging between 30 cm and 240cm and over
respectively.
The vertical distribution of individuals in the forest showed an average height of 30-40 m attained by canopy species in lower slopes and valleys. By contrast, on hill tops and exposed ridges canopy height diminished to about 2-3 m in some plots at sites 4 and 5. The absence of conspicuous emergents resulted in a closed and continuous canopy layer, particularly in the tall-statured stands, permitting little light to filter down to the forest floor, where the ground vegetation was extremely sparse. On the whole, a certain degree o f stratification in the vegetation was discernible, with the main canopy reaching 30-40 m above ground, a subcanopy between 15-30 m and an understorey tree layer between 5-15 m height. Woody

28 C. V. S. Gunatilleke, I. A. U. N. Gunatilleke
lianas, when present, formed an intricate network in the crowns of canopy trees. On assigning each species to a particular stratum based on the position its individuals attain at maturity, it was possible to recognise 59 canopy, 49 subcanopy and 103 understorey tree species.
The contribution of endemic species to density of individuals in the forest stand is as high as 86 ~ for the total sample. The corresponding data for each of the sites are given in Table 1 and Fig. 3. Since publication of the preliminary report of this study (Gunatilleke & Gunatilleke, 1981) the nomenclature and endemicity of some species have been revised; most notable among them is the dominant canopy species, Mesuaferrea Auct. (non L.) revised as Mesua nagassarium (Burm. f.) Kosterm. Its variety pulchella found at Sinharaja is reported endemic to the island (Kostermans, 1980). Updated figures of the proportion of endemics at each site are given in Table 1. In each girth-size class the proportion of individuals of endemic species is consistently high and over 72 ~o (Fig. 3). Similarly, their contribution to total basal area is appreciably high and over 87 ~ (Table 1).
Floristic composition
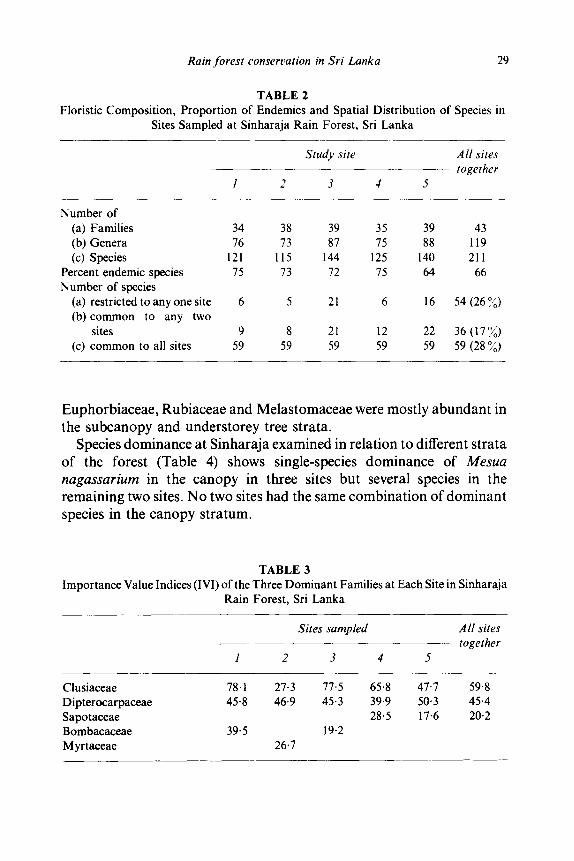
The floristic data and proportion of endemics at individual sites and all sites together, as well as the distribution of species between sites, are given in Table 2.
At the sites sampled, both family and species dominance have been examined based on IVI as proposed by Curtis & McIntosh (1950). Either Clusiaceae or Dipterocarpaceae is the leading family in sites 1 4 as indicated by their high IVI, which are almost double those of the second leading family. At site 5, however, the close similarity of IVI between these two families suggest their co-dominance in this stand (Table 3). These two families together comprise 37 ~ of the 17 427 individuals and 45 ~ of the total basal area (966.7 m 2) sampled. At all sites, Bombacaceae, Myrtaceae and Sapotaceae were among the third, fourth and fifth leading families.
When the number of species in each family was examined, Euphorbiaceae with 22 species ranked first, followed by Myrtaceae with 17, Dipterocarpaceae and Rubiaceae each with 16, Clusiaceae, Ebenaceae and Sapotaceae each with 12 and Melastomaceae with 10. Thus over 50 ~o of the tree species sampled at Sinharaja belonged to these eight families. Further, while species of Myrtaceae, Dipterocarpaceae, Clusiaceae and Sapotaceae were present in all three strata of the forest, those of
Rain forest conservation in Sri Lanka 29
TABLE 2 Floristic Composition, Proportion of Endemics and Spatial Distribution of Species in
Sites Sampled at Sinharaja Rain Forest, Sri Lanka
Study site All sites together
1 2 3 4 5
Number of (a) Families 34 38 39 35 39 43 (b) Genera 76 73 87 75 88 119 (c) Species 121 115 144 125 140 211
Percent endemic species 75 73 72 75 64 66 Number of species
(a) restricted to any one site 6 5 21 6 16 54 (26 ~/o) (b) common to any two
sites 9 8 21 12 22 36 (17~o) (c) common to all sites 59 59 59 59 59 59 (28 ~)
Euphorbiaceae, Rubiaceae and Melastomaceae were mostly abundant in the subcanopy and understorey tree strata.
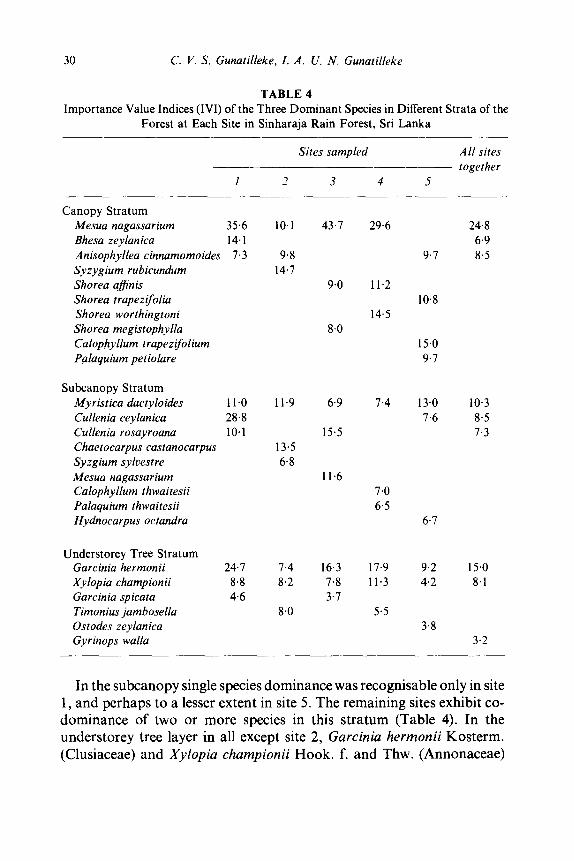
Species dominance at Sinharaja examined in relation to different strata of the forest (Table 4) shows single-species dominance of Mesua nagassarium in the canopy in three sites but several species in the remaining two sites. No two sites had the same combination of dominant species in the canopy stratum.
TABLE 3 Importance Value Indices (IVI) of the Three Dominant Families at Each Site in Sinharaja
Rain Forest, Sri Lanka
Sites sampled All sites together
I 2 3 4 5
Clusiaceae 78.1 27-3 77.5 65.8 47.7 59.8 Dipterocarpaceae 45-8 46.9 45-3 39.9 50.3 45.4 Sapotaceae 28.5 17.6 20.2 Bombacaceae 39-5 19.2 Myrtaceae 26.7
30 C. V. S. Gunatilleke, I. A. U. N. Gunatilleke
TABLE 4 Importance Value Indices (IVI) of the Three Dominant Species in Different Strata of the
Forest at Each Site in Sinharaja Rain Forest, Sri Lanka
Sites sampled All sites together
1 2 3 4 5
Canopy Stratum Mesua nagassarium Bhesa zeylanica Anisophyllea cinnamomoides Syzygium rubicundum Shorea affinis Shorea trapezifolia Shorea worthingtoni Shorea megistophylla Calophyllum trapezifolium Palaquium petiolare
Subcanopy Stratum Myristica dactyloides Cullenia ceylanica Cullenia rosayroana Chaetocarpus castanocarpus Syzgium sylvestre Mesua nagassarium Calophyllum thwaitesii Palaquium thwaitesii Hydnocarpus octandra
Understorey Tree Stratum Garcinia hermonii Xylopia championii Garcinia spicata Timonius jambosella Ostodes zeylanica Gyrinops walla
35.6 10.1 43.7 29.6 24.8 14.1 6.9 7.3 9.8 9.7 8.5
14.7 9.0
8.0
11.2
14.5 10.8
15.0 9.7
11-0 11.9 6.9 7.4 13.0 10.3 28.8 7.6 8.5 10-1 15.5 7-3
7.0 6.5
11.6
13.5 6-8
6.7
24.7 7.4 16.3 17.9 9.2 15.0 8.8 8.2 7.8 11.3 4.2 8.1 4.6 3.7
8.0 5.5 3.8
3-2
In the subc a nopy single species d o m i n a n c e was recognisable on ly in site 1, and perhaps to a lesser extent in site 5. The remain ing sites exhibi t co- d o m i n a n c e o f two or more species in this s t r a tum (Table 4). In the unde r s to r ey tree layer in all except site 2, Garcinia hermonii Kos te rm. (Clusiaceae) and Xylopia championii H o o k . f. and Thw. (Annonaceae )

Rain forest conservation in Sri Lanka 31
were the two leading species with a tendency towards single-species dominance by the former. While X. championii ranked first in site 2, Timonius jambosella (Gaertn.) Thw. (Rubiaceae) and G. hermonii, the second and third leading species, whose IVI were very closely similar to the first, together co-dominated this stratum.
The three dominant canopy species at different sites recorded a greater diversity as compared with those of the subcanopy and understorey tree layers, which showed progressively fewer species among their dominants (Table 4). On the other hand, distribution of species in different strata showed a reverse trend; viz. there were 59 canopy, 49 subcanopy and 103 understorey tree species. The latter strata also have juvenile individuals of canopy species and had they been included, diversity of species in the lower strata would be more pronounced and as high as 211 species in the understorey and 108 (i.e. 59 + 49) in the subcanopy.
The leading species in different strata in all sites were endemics, except at site 2 where two non-endemics, Syzygium rubicundum and Chaetocarpus castanocarpus Thw. (Euphorbiaceae), ranked first in the canopy and subcanopy respectively.
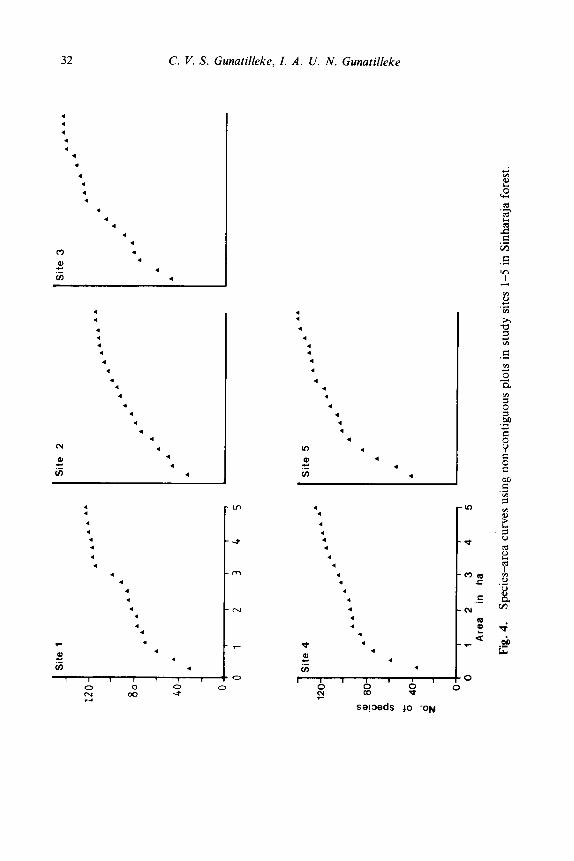
The species-area curves drawn using non-contiguous plots for each site (Fig. 4) in this study reveal the following: (i) that adequate areas have been sampled to give a fair representation of the vegetation in each site. This is indicated by a levelling-off of the respective curves characterising each site; (ii) with the exception of that in site 2, the step-wise rise in the graphs
TABLE 5 Proportion of Species in Different Population Size Classes at Each Site given as a
Percentage of the Total Number of Species Recorded Therein
Sites samph, d All sites together
2 3 4 5
Area sampled (ha) 5 5 Percentage of species in each site with: "/ /o 'YO
(i) 1 individual/species 16 11 (ii) 2-10 individuals/species 48 41
(iii) 11-100 individuals/species 29 37 (iv) 101-200 individuals/species 05 10 (v) 201 individuals and over/
species 02 01
5 5 5 25
% % % % 25 19 13 10 38 37 43 30 33 37 38 40 02 04 05 10
02 03 01 10
32 C. 1I. S. Gunatilleke, I. A. U. N. Gunatilleke
v-
cr~
4
4
4
4
4
4
4
4
i i T r I i o o o
N o o "4"
Lr~
",4"
o
4
4
4
Q) •
4 4
4
4 4
4
u~
I ~ ,
4
& ' 6 ' CO '~"
s e ! o e d s ) o "o N
.E -¢'~1
- m , -
0
0
tg~ I
0
0 ?
0
&
,4

Rain forest conservation in Sri Lanka 33
suggests a higher localisation of species, particularly at sites 1, 3 and 5; and (iii) the floristic richness of sites 3 and 5 and floristic paucity of site 2.
Population densities of the species computed for the total sample (Table 5) reveal that 10 ~o of the species are extremely poorly represented with only one individual per 25 ha. A further 30 ~o had 2-10 individuals per 25 ha. While these species may be considered rare, over half of them are endemic to the island, and confined to these lowland wet zone forests. The proportion of species with different population densities at each site is given in Table 5. Sixty-two percent of the species with small population densities are restricted to the understorey tree stratum, 18~o ~ to the subcanopy and 20 ~ to the canopy.
DISCUSSION
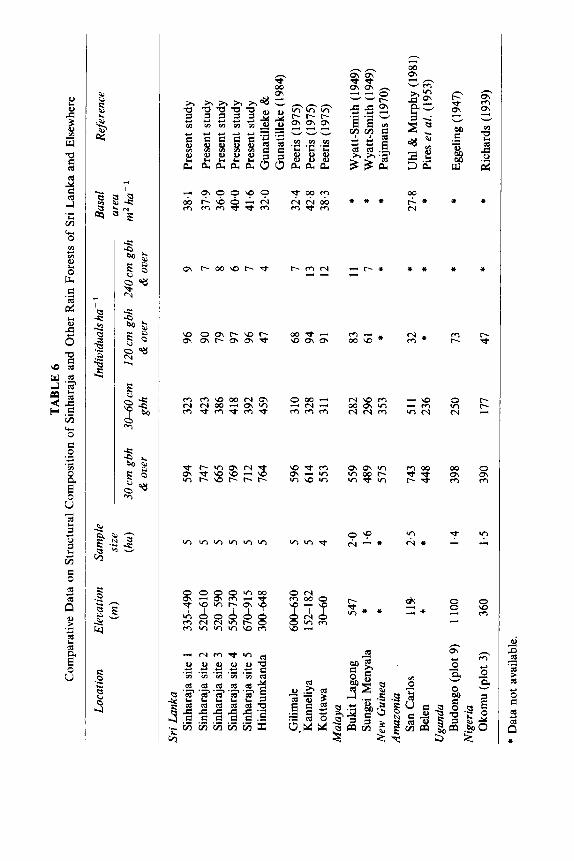
Despite the altitudinal range (335-915 m) of the five forest stands sampled in Sinharaja, a similarity was observed in their (a) mean density of individuals over 30cm gbh; (b) basal area per ha; and (c) vertical distribution of individuals. These data, on the whole, are in agreement with corresponding values for other lowland wet zone forests of the country (Table 6) (Peeris, 1975; Gunatilleke & Gunatilleke, 1984). A noteworthy difference, however, is the somewhat higher proportion of trees over 240cm gbh recorded at Kanneliya and Kottawa forests, possibly accounted for by their lower altitude, viz. 152m-182m and 30 m-60 m respectively. The average density of stems over 30 cm gbh in all stands sampled at Sinharaja are higher than those recorded for forests of Malaya (Cousens, 1951 ; Wyatt-Smith, 1949; Ashton, 1971), New Guinea (Paijmans, 1970) and Africa (Eggeling, 1947; Richards, 1939) but in between those of Amazonia (Uhl & Murphy, 1981; Pires et aL, 1953) (Table 6).
The floristic richness and localisation of species varied between sites sampled and was highest at sites 3 and 5, both located on the southern side of the reserve where human impact was also greatest. Unlike the western and central parts of the reserve which have a network of roads that facilitate effective policing, the southern part, by its relative inaccessibility, receives much less protection. The species richness of the Sinharaja stands sampled ranges between 115-144 and is similar to those of the other wet lowland forests of Sri Lanka. However, these figures are much lower than
" 0
c~
0
E ' ~ . ~
0
0
0
~S
\
L~
I
oo
c~ c~

Rain forest conservation in Sri Lanka 35
those reported for the Malayan region (Wyatt-Smith, 1949; Cousens, 1951; Ashton, 1964).
Species area curves of the Sinharaja forest stands depict a minimal area of 3.75 ha (Fig. 4) and again conform with the value arrived at for other wet lowland forests of the Island (Peeris, 1975).
Based on Importance Value Indices, Clusiaceae followed by Diptero- carpaceae are the two most dominant families in the Sinharaja sites sampled (Table 3). In this respect too, they differ from the lower altitude Kanneliya and Kottawa forests where Dipterocarpaceae was the leading dominant family (Peeris, 1975).
Although Mesua nagassarium depicts single species dominance in the canopy stratum in all Sinharaja sites except site 2, the combination of the three most dominant species differed at each site, resulting in a number of species among the leading dominants in this stratum. Contrastingly, few species dominated the subcanopy and even fewer the understorey tree stratum, where Garcinia hermonii and Xylopia champion ii always ranked among the dominants. If dominance of a species reflects its adaptability to a particular area at a given time, then Mesua nagassarium, Garcinia hermonii and Xylopia championii are the most successful irrespective of topographic, altitudinal and microclimatic variations that prevail in this 8800 ha forest. The greater diversity of species in the understorey tree layer as compared to the canopy, 49 ~ and 28~o respectively, also suggests that the understorey has a much more diverse environment and more habitat niches than the overstorey strata. In spite of this habitat diversity in the understorey tree layer, it is remarkable that the same few species dominate this layer in different parts of the forest.
The altitudinal range (450 m-900 m) and dominant species (Mesua and Shoreaspp.) of Sinharaja sites 2, 3 and 4 are similar to those of de Rosayro's Mesua-Doona (Shorea) community (de Rosayro, 1942). Site 1 also has the same dominant species and hence could be included in this community, but the lower limit of its altitudinal range (335m) falls outside that described by de Rosayro, again confirming our contention that the Mesua-Doona (Shorea) community occurs over a wider altitudinal range than that proposed by him (Peeris, 1975). Site 5 on the other hand, by its paucity of Mesua spp., does not belong to any other of de Rosayro's high forest community type.
As in all wet lowland forest sampled for the island, the endemic tree species at Sinharaja not only make a significant contribution to the structural and floristic composition of this primaeval forest ecosystem,

36 C. V. S. Gunatilleke, I. A. U. N. Gunatilleke
but they also dominate it. In the wet lowlands of Sri Lanka, there are 217 endemic tree species and woody lianas (Peeris, 1975). Of these 139 have been recorded in this phytosociological survey. Of the 27 genera reported endemic to Sri Lanka (Abeywickrama, 1956), nine--Schumacheria Vahl. (Dilleniaceae), Stemonoporus Thw. (Dipterocarpaceae), Gleniea Hk.f. (Sapindaceae), Nargedia Bedd., Leucocodon Gardn., Schizostigma Arn. (Rubiaceae), Podadenia Thw. (Euphorbiaceae), Hortonia Wight (Monimiaceae) and Loxococcus Wendl. and Drud. (Palmae)--have been recorded at Sinharaja. With the exception of Schumacheria, Stemonoporus and Hortonia these genera are also monotypic (Willis, 1973).
Since a high localisation of species has been recorded even within the small area of 25 ha sampled at Sinharaja, it is not unlikely that more endemic woody species may be present in areas not sampled by us. However, there is still a large number of endemic species restricted to other forests in the wet lowlands of the country (Table 7 and references therein). Will these species be able to survive in small patches of degraded forest or could Sinharaja be made a home for them too? Mason-Hale (1981) after an exhaustive study of the lichen family Thelotremataceae in Sri Lanka in which 35 ~ of the species are endemic to the wet zone of the island, says 'most of them are abundant in the undisturbed lowland rain forest and logging is particularly damaging for lichens and many other organisms, since exposure to direct sunshine and desiccation can wipe
TABLE 7 Comparat ive Data on Floristic Composi t ion of Sinharaja and Other Lowland Rain
Forests of Sri Lanka
Location Area Number oJ ~0 ReJerence sampled endemic
(ha) Families Genera Species species
Sinharaja site 1 5 34 76 121 75 Present study Sinharaja site 2 5 38 73 115 73 Present study Sinharaja site 3 5 39 87 144 72 Present study Sinharaja site 4 5 35 75 125 75 Present study Sinharaja site 5 5 39 88 140 64 Present study Hinidumkanda 5 39 92 140 75 Gunatilleke & Gunatilleke
(1984) Kanneliya 5 34 77 125 65 Peeris (1975) Kottawa 4 31 65 101 67 Peeris (1975) Gilimale 5 41 88 136 59 Peeris (1975)

Rain forest conservation in Sri Lanka 37
them out in a matter of months after the canopy is destroyed'. The same could perhaps be said of the low-density (less than 10 individuals 25 ha- 1) species which make up 40 ~o of the Sinharaja tree flora sampled. Among them, those restricted to the understorey (62 ~o) and those which inhabit a single site would be the most adversely affected. The herbaceous, shrub and epiphytic species, whose very survival certainly also depends on the performance of the overhead strata, would be susceptible to any form of disturbance. So far, the ecology of these life forms in the lowland wet zone forests has received little attention (Mitchell & Tilakeratne, 1980). Consequently, much remains to be learnt about them before habitats are further disturbed.
The differences observed at site 2 as compared to the other sites--(i) lowest floristic richness; (ii) greater homogeneity of the vegetation as observed in the species-area curve; (iii) prevalence of non-endemics among the leading species in the canopy and subcanopy and (iv) fewer species restricted to this site-- make one wonder whether this site has in fact been disturbed in the past. This could well be true because the site is more or less surrounded by patches of secondary vegetation in different stages of succession. However, its structural composition gave no such indication at the time of its selection for sampling. This leads us to speculate that the disturbance would have been of a thinning nature rather than one that opened up large gaps in the canopy, allowing secondary forest species to invade.
Sinharaja, surrounded by wastelands of Dicranopteris linearis (Burm. f.) Underw. now being planted with exotic Pinus caribaea, plantations of tea and rubber and human settlements, has virtually become the last refugium for most of Sri Lanka's lowland wet zone plant species. It would not be incorrect to say that most plant species at Sinharaja belong at least to the category of vulnerable species, and some even to that of endangered species as classified by the International Union for Conservation of Nature and Natural Resources (Lucas & Synge, 1981).
This phytosociological survey has given baseline information on the tree species of Sinharaja and laid a foundation for on-going and future research. This survey shows that different areas of the forest, represented by five sampling sites, have their own unique floristic features and a single area is not representative of the entire forest. Therefore, every endeavour must be taken to conserve this largest relatively undisturbed patch of lowland rain forest in its entirety. In the unwelcome event that part of this

38 C. V. S. Gunatilleke, I. A. U. N. Gunatilleke
forest reserve has to be released for timber exploitation, the data collected should be examined in the light of scientific criteria laid down for selection of conservation areas (Jacobs, 1979; Shaffer, 1981) before deciding upon the best and most appropriate area that should be conserved. At the same time we have information as to which species would be excluded in the process, so that an attempt could at least be made to conserve them in botanical gardens or arboreta.
A C K N O W L E D G E M E N T S
The authors extend their thanks and grateful appreciation to the following: International Foundation for Science, Sweden for their generous financial support, prompt attention and amicable solutions to the many administrative problems during various phases of the project; Natural Resources, Energy and Science Authority of Sri Lanka for local administration of the project and provision of transport; Forest Department and the State Timber Corporation of Sri Lanka for facilities during field work; Department of Botany, University of Peradeniya, Sri Lanka and the Arnold Arboretum of Harvard University, Cambridge, USA for laboratory and library facilities and all who helped with field work and data analysis, without whose assistance this project could never have been successfully completed.
REFERENCES
Abeywickrama, B. A. (1956). The origin and affinities of the flora of Ceylon. Proc. Ann. Sess. Ceylon A.A.S., 1 lth, Part 2, 99-121.
Alston, A. H. G. (1931). A handbook to theflora of Ceylon, 6. London, Dulav and Co.
Andrews, J. R. T. (1961). Forest inventory of Ceylon (A Canadian-Ceylon Colombo-Plan Project). Colombo, Sri Lanka, Ceylon Government Press.
Ashton, P. S. (1964). Ecological studies in the mixed dipterocarp forests of Brunei State. Oxf. For. Mem., 25, 1-75.
Ashton, P. S. (1971). Pasoh Forest Reserve vegetation survey. Second report, IBP Malayan Project. Kepong, Malaysia, Forest Research Institute. (Mimeographed.)
Ashton, P. S. (1981). Techniques for the identification and conservation of threatened species in tropical forests. In The biological aspects of rare plant conservation, ed. by Hugh Synge, 155-64. Chichester, John Wiley.
Baker, J. R. (1937). The Sinharaja Rain Forest, Ceylon. Geogr. Jl, 89, 539-51. Baker, J. R. (1938). Rain forest in Ceylon. Kew Bull., 1, 9-16.

Rain.[brest conservation in Sri Lanka 39
Cooray, P. G. (1967). An introduction to the geology of Ceylon. Spolia Zeylanica, 31, 1 314.
Cousens, J. E. (1951). Some notes on the composition of lowland tropical rain forest in Rengam Forest Reserve, Johore. Malay. Forester, 14, 131-9.
Curtis, J. T. & Mclntosh, R. P. (1950). The interrelations of certain analytic and synthetic phytosociological characters. Ecology, 31, 434-54.
Dassanayake, M. D. & Fosberg, F. R. (1980-1982). A revised handbook to the flora of Ceylon, 1, 2, and 3. New Delhi, Amarind Publishing Co.
de Rosayro, R. A. (1942). The soils and ecology of the wet evergreen forests of Ceylon. Trop. Agric. (Ceylon), 98, 70-80; 153-75.
de Rosayro, R. A. (1954). A reconnaissance of Sinharaja rain forest. Ceylon Forester, 1(3), N.S., 68-74.
de Rosayro, R. A. (1959). The application of aerial photography to stock- mapping and inventories on an ecological basis in rain forests in Ceylon. Empire Forestry Review, 38, 141-74.
Eggeling, W. J. (1947). Observations on the ecology of Budongo rain forest, Uganda. J. Ecol., 34, 20-87.
Gunatilleke, C. V. S. & Gunatilleke, I. A. U. N. (1981). The floristic composition of Sinharaja--A rain forest in Sri Lanka with special reference to endemics and Dipterocarps. Malay. Forester, 44, 386 96.
Gunatilleke, C. V. S. & Gunatilleke, I. A. U. N. (1983). A forestry case study of the Sinharaja rain forest in Sri Lanka. In Forest and watershed development and conservation in Asia and the Pacific, ed. by L. S. Hamilton, 289-357. Colorado, Westview Press.
Gunatilleke, I. A. U. N. & Gunatilleke, C. V. S. (1984). Distribution of endemics in the tree flora of a lowland hill forest in Sri Lanka. Biol. Conserv., 28, 275 85.
Jacobs, M. (1979). Botanical philosophy on the selection of rain forest reserves in Malesia. Flora Malesiana Bull., 32, 3247-50.
Kostermans, A. J. G. H. (1980). Clusiaceae (Guttiferae). In A revised handbook to the flora of Ceylon, I, ed. by M. D. Dassanayake and F. R. Fosberg, 72-110. New Delhi, Amarind Publishing Co.
Lucas, G. & Synge, H. (1981). The assessment and conservation of threatened plants around the world. In The biological aspects oJ rare plant conservation, ed. by H. Synge, 3-18. Chichester, Wiley.
Mason-Hale, E. Jr (1981). A revision of the lichen family Thelotremataceae in Sri Lanka. Bull. Br. Mus. Nat. Hist. (Bot.), 8(3), 227-332.
Merritt, V. G. & Ranatunga, M. S. (1959). Aerial photographic survey of Sinharaja Forest. Ceylon Forester, N.S., 4(2), 103 56.
Mitchell, R. J. & Tilakeratne, L. K. D. (1980). Understorey vegetation structure of the Sinharaja tropical wet evergreen forest type. Sri Lanka Forester, N.S., 14(3 and 4), 181-6.
Moorman, F. R. & Panabokke, C. R. (1961). Soils of Ceylon. Trop. Agric. (Ceylon), 117, 4-65.
Nanayakkara, S. D. F. C. (1981). Sri Lanka Forest cover map. Colombo, Center for Remote Sensing, Survey Department.

40 C. V. S. Gunatilleke, I. A. U. N. Gunatilleke
Paijmans, K. (1970). An analysis of four tropical rain forest sites in New Guinea. J. Ecol., 58, 77 101.
Peeris, C. V. S. (1975). The ecology of endemic tree species of Sri Lanka in relation to their conservation. PhD thesis, University of Aberdeen.
Pires, J. M., Dobzhansky, T. & Black, G. A. (1953). An estimate of the number of species of trees in an Amazonian Forest Community. Bot. Gaz., 114, 467-77.
Richards, P. W. (1939). Ecological studies on the rain forest of southern Nigeria, 1. The structure and floristic composition of the primary forest. J. Ecol., 27, 1-16.
Shaffer, M. L. (1981). Minimum population sizes for species conservation. Bioscience, 31, 131~4.
Sri Bharathie, K. P. (1979a). Man and Biosphere reserves in Sri Lanka. Sri Lanka Forester, N.S., 14, 37-40.
Sri Bharathie, K. P. (1979b). Natural regeneration in the exploited section of the Sinharaja Reserve. Sri Lanka Forester, N.S., 14, 41-2.
Trimen, J. (1893-1900). A handbook to the flora of Ceylon. London, Dulav Co. Uhl, C. & Murphy, P. G. (1981). Composition, structure and regeneration of a
tierra firme forest in the Amazon Basin of Venezuela. Trop. Ecol., 22, 219-37.
Whitmore, T. C. (1975). Tropical rainjorests of the Far East. Oxford, Clarendon Press.
Willis, J. C. (1973). A dictionary of the flowering plants andjerns, 8th edn, revised by H. K. Airy Shaw, Cambridge, Cambridge University Press.
Wyatt-Smith, J. (1949). A note on tropical lowland evergreen rain forest in Malaya. Malay. Forester, 12, 58-64.





























