Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Erik Willems . Luc Leyns
Patterning of mouse embryonic stem cell-derived pan-mesodermby Activin A/Nodal and Bmp4 signaling requires fibroblast growthfactor activity
Received June 2, 2007; accepted in revised form November 1, 2007
Abstract Embryonic stem (ES) cells have the poten-tial to differentiate into all cell types of the adult body,and could allow regeneration of damaged tissues. Thechallenge is to alter differentiation toward functionalcell types or tissues by directing ES cells to a specificfate. Efforts have been made to understand the molec-ular mechanisms that are required for the formation ofthe different germ layers and tissues from ES cells, andthese mechanisms appear to be very similar in themouse embryo. Differentiation toward mesoderm andmesoderm derivatives such as cardiac tissue or hem-angioblasts has been demonstrated, however, the rolesof Activin A/Nodal, bone morphogenetic protein(BMP), and fibroblast growth factor (FGF) signalingin the early patterning of ES cell-derived pan-mesodermand anterior visceral endoderm (aVE) have not beenreported yet. We therefore analyzed the roles of ActivinA/Nodal, BMP, and FGF signaling in the patterning ofES cell-derived mesoderm as well as specification of theaVE by using a dual ES cell differentiation system com-bining a loss-of-function with a gain-of-function ap-proach. We found that Activin A or Nodal directed thenascent mesoderm toward axial mesoderm and mesen-doderm, while Bmp4 was inducing posterior and extra-embryonic mesoderm at the expense of anteriorprimitive streak cells. FGF signaling appeared to havean important role in mesoderm differentiation by al-lowing an epithelial-to-mesenchymal transition of thenewly formed mesoderm cells that would lead to theirfurther patterning. Moreover, inhibition of FGF sig-
naling resulted in increased expression of axial meso-derm markers. Additionally, we revealed that theformation of aVE cells from ES cells requires FGF-dependent Activin A/Nodal signaling and the attenua-tion of Bmp4 signaling.
Key words embryonic stem cells � mesoderm �patterning � aVE � mouse
Introduction
Embryonic stem (ES) cells are derived from the innercell mass of a blastocyst embryo (Evans and Kaufman,1981; Thomson et al., 1998). Because of their unlimitedself-renewal and pluripotency, ES cells have consider-able promise as an alternative for organ transplantation(reviewed in Smith, 2001). They can differentiate into allcell types of the adult body and would thus be able togenerate organ tissues in vitro or could allow regener-ation of damaged tissues in vivo. The challenge, how-ever, is to control ES cell fate so that differentiation canbe altered to enrich a specific cell population. Effortshave already been made to understand the molecularmechanisms that determine whether an ES cell remainspluripotent or whether it differentiates toward endo-derm, mesoderm, or ectoderm cell types. Recently,mimicking of the molecular signals in mouse develop-ment to direct ES cell differentiation has demonstratedgreat promise. Using such approaches, ES cell differen-tiation could not only be exploited as a therapeutic po-tential, but also as an in vitro model to study molecularinteractions in, for example, mesoderm development(D’Amour et al., 2006; Gadue et al., 2006).
In the mouse embryo, gastrulation is initiated around6.0 dpc with the emergence of the primitive streak onthe posterior side of the embryo. At the same time, the
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
D I F 2 5 7 B Dispatch: 30.11.07 Journal: DIF CE: Swatilekha
Journal Name Manuscript No. Author Received: No. of pages: 15 PE: Ratna/prasath
Erik Willems � Luc Leyns ( .*)Laboratory for Cell GeneticsVrije Universiteit BrusselPleinlaan 2, 1050 BrusselsBelgiumFax: 13226292759E-mail: [email protected]
Differentiation (2007) DOI: 10.1111/j.1432-0436.2007.00257.xr 2007, Copyright the AuthorsJournal compilation r 2007, International Society of Differentiation
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

distal visceral endoderm (dVE) will have moved to theanterior side of the embryo (now called the anteriorvisceral endoderm, aVE), where it will allow neural in-duction by blocking the signals emanating from theprimitive streak (Tam et al., 2001, 2006). As gastrulat-ion proceeds, epiblast cells become mesenchymal andmigrate through the primitive streak, in between theendoderm and ectoderm layer, forming the mesoderm.Further patterning of the formed mesoderm leads to theformation of the mesendoderm in the anterior primitivestreak and posterior and extraembryonic mesoderm inthe posterior primitive streak. Mesendoderm will giverise to the somitic or paraxial mesoderm and the defin-itive endoderm, while posterior mesoderm will contrib-ute to tail and cardiac mesoderm. Extraembryonicmesoderm mostly forms the amnion and hemangio-blasts, the precursors of blood and blood vessels (Tamet al., 2001, 2006). Patterning of the epithelial epiblastcells positioned immediately anterior of the anteriorprimitive streak will lead to the formation of axial me-soderm, comprising the node and notochord and whichare involved in gut and neural specification (Kinderet al., 2001; Blum et al., 2007).
Several of the molecular players that control pat-terning of the embryo have been characterized by com-plementing mouse models with ES cell models (reviewedin Tam and Loebel, 2007). Wnt signaling is required forthe induction of mesoderm and is needed for the move-ment of the dVE toward the anterior side of the embryo(Haegel et al., 1995; Liu et al., 1999; Gadue et al., 2006).Later in gastrulation, Wnt proteins are needed for thespecification of the presomitic mesoderm (Takada et al.,1994; Ikeya and Takada, 1998). Nodal is important forthe formation of the anterior–posterior axis in the em-bryo, because it induces the dVE as well as the primitivestreak (Conlon et al., 1994; Brennan et al., 2001). Fur-thermore, Nodal patterns the newly formed mesoderm(Vincent et al., 2003; Kubo et al., 2004; Tada et al.,2005). Of the bone morphogenetic protein (BMP) pro-tein family, only Bmp2 and Bmp4 were found to haveseveral important roles in early development. Bmp2� /�
embryos have malformations in the amnion and chori-on and have heart malformations (Zhang and Bradley,1996). Ablation of Bmp4 expression results in reducedmesoderm formation, tail truncations, defective dVEmigration, and blood cell formation (Winnier et al.,1995; Soares et al., 2005; Kennedy et al., 2006). Fibro-blast growth factor signaling on the other hand is notnecessary for mesoderm induction in the mouse em-bryo, but it is essential for the migration of the meso-derm away from the primitive streak by allowing newlyformed mesoderm cells to undergo an epithelial to me-senchymal transition (EMT) (Deng et al., 1994; Yam-aguchi et al., 1994; Ciruna et al., 1997; Ciruna andRossant, 2001). In absence of FGF activity, patterningis thus significantly reduced because of a defective mi-gration of epiblast cells through the primitive streak
(Deng et al., 1994; Yamaguchi et al., 1994; Sun et al.,1999; Guo and Li, 2007).
Differentiation of ES cells toward primitive streakderivatives such as hemangioblasts, cardiac tissue, anddefinitive endoderm has been demonstrated, however,the roles of Activin A or Nodal, Bmp4, and FGF sig-naling in the early formation of the different mesodermsubtypes and the aVE still need to be determined. Wehave therefore studied Activin A/Nodal, Bmp4, andFGF signaling in mesoderm patterning and aVE for-mation from ES cells, by combining a loss-of-functionapproach in serum-containing media (SCM) (using spe-cific signaling inhibitors) with a gain-of-function setupin serum-free media (SFM) (using specific signalingactivators). During ES cell differentiation, Activin Aor Nodal induced mesendoderm and axial mesodermmarkers in embryoid bodies (EB), whereas Bmp4enhanced expression of posterior and extraembryonicmesoderm genes at the expense of anterior primitivestreak patterning. Moreover, we found that mesodermpatterning by Activin A/Nodal and Bmp4 signaling inES cells was dependent on an FGF-mediated EMT andthat FGF signaling was important in determining cel-lular fate of the future node cells. Additionally, we re-vealed that specification of aVE cells from ES cellsrequires FGF-dependent Activin A/Nodal signalingand the attenuation of Bmp4 signaling.
Materials and methods
ES cell culture and differentiation
Mouse E14Tg2a feeder-free ES cells were cultured on gelatin-coat-ed dishes in Knock Out DMEM (GIBCO Q1) supplemented with 10%fetal calf serum (FCS) (Hyclone Q2), 0.1mM amercaptoethanol (GI-BCO), 1% non-essential amino acids (GIBCO), 2mM L-glutamine(Sigma Q3), antibiotics (Sigma), and 1,000U/ml of LIF (Sigma).Differentiation in a SFM containing N2 and B27 supplements
without retinoic acid was performed as described elsewhere (Gadueet al., 2006). EB were allowed to form from 50,000 cells/ml andafter 2 days, EB were transferred to fresh SFM containing mouserecombinant Nodal, human recombinant Activin A, human re-combinant Fgf2, mouse recombinant Fgf4, and Fgf8b or mouserecombinant Bmp4 for 2 more days (R&D Systems Q4). FGF inhibi-tion using the FGF receptor blocker SU5402 (Sigma) was allowedfor 2 hr before the addition of Activin A or Bmp4.Differentiation in SCM was performed in ES cell medium with-
out LIF. EB were allowed to form from 50,000 cells/ml for 3 days,and were cultured for another day in the presence of the Activin A/Nodal receptor inhibitor SB-431542 (Sigma), SU5402, or the BMPantagonist Noggin (R&D Systems).
Quantitative reverse transcription PCR (qRT-PCR)
Cell samples were collected 4 days after the initiation of EB for-mation and RNA was extracted using the RNagents RNA extrac-tion kit (Promega Q5) and qRT-PCR was performed as describedpreviously (Willems et al., 2006). Primer sequences for all markersare available in the Real-Time Primer Database and are listedin Supplementary Table 1 (http://medgen.ugent.be/rtprimerdb/)(Pattyn et al., 2003). qRT-PCR data were calculated as described
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
2
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

using Actb as a reference gene (Willems et al., 2006). All treatedsamples are represented as the expression level of the gene relativeto their corresponding untreated control (control value equals toone). The results are presented as the mean of three independentexperiments with the standard error of the mean.
Statistical analysis
Statistical significance of the differential expression levels wasassessed by the calculation of a 95% confidence interval (CI) forthree independent experiments. To this purpose, normalizedexpression levels were log transformed and subsequently standard-ized so that the average expression and standard deviation of thevarious conditions in a given experiment are equalized over theexperiments.
Whole mount in situ hybridization (WISH)
WISH was performed as described elsewhere (Belo et al., 2000).Staining for alkaline phosphatase (AP) was stopped simultaneouslyin all samples. BCIP was used as a substrate for AP for the Bra-chyury riboprobe, BM purple was used for all other riboprobes(Roche).The riboprobes used for Brachyury, Shh, and Lhx1 WISH are
described (Wilkinson et al., 1990; Echelard et al., 1993; Shawlot andBehringer, 1995). Evx1 and Snai1 riboprobes were a kind gift ofP. Dolle and F. Gofflot.
Fluorescent-assisted cell sorting (FACS)
EB were dissociated to single cells by light trypsinization (3min at371C) and were washed thoroughly in PBS to remove all traces oftrypsin and serum. Cells were then incubated for 45min at 41C withAPC-labeled Flk1 antibody (eBioscienceQ6 ), after which unboundantibody was removed by washing with PBS. Labeled cells weredirectly measured with a FACSCanto II flow cytometer (BDBiosciencesQ7 ).
Results
Here, we investigated the molecular signals that are re-quired to pattern ES cell-derived pan-mesoderm in EBtoward the different mesoderm subtypes such as mes-endoderm, axial mesoderm, posterior mesoderm, andextraembryonic mesoderm. By using a dual ES cell cul-ture system based on SFM and SCM, we were able tostudy gain-of-function as well as loss-of-function ofActivin A/Nodal, Bmp4, and FGF signaling by geneexpression analysis of specific markers (SupplementaryTable 1). Differentiation in SFM results in low spon-taneous mesoderm patterning, allowing the analysis ofthe role of added growth factors on the different me-soderm subtypes (Johansson and Wiles, 1995; Gadue etal., 2006). By adding specific inhibitors for Activin A/Nodal, BMP, and FGF signaling to EB after sponta-neous mesoderm induction had occurred in SCM (atday 3 of embryoid body formation), we were able tostudy their effect on mesoderm patterning.
Concentration-dependent mesoderm patterning byActivinA/Nodal signaling
Activin A/Nodal signaling induces anterior mesodermand endoderm in ES cells (Kubo et al., 2004; Pfendler etal., 2005; Tada et al., 2005). Because these reports how-ever do not address the effect of graded Activin A/Nodal activity on ES cell differentiation toward differ-ent mesoderm derivatives, we have studied Activin A/Nodal activation and inhibition during mesoderm pat-terning. Graded effects of Activin A/Nodal signaling inES cell differentiation were obtained by generating agradient using a combination of a concentration rangeof the Activin A/Nodal signaling inhibitor SB-431542(SB, 10 and 1 mM) in SCM with a series of Activin A (5,15, 30, and 50 ng/ml) or Nodal protein concentrations(40, 80, 200, and 400 ng/ml) in SFM, both comparedwith their respective untreated controls (Fig. 1A, Sup-plementary Fig. 1A). The gradation of Activin A/Nodalsignaling activity was demonstrated by the expression ofLefty1 and Lefty2, two direct target genes of the ActivinA/Nodal signaling pathway (Fig. 1B, SupplementaryFig. 1B). The comparison of the effect of Activin A andNodal on the expression of Lefty1 and Lefty2 clearlyshows that Activin A protein is more active than Nodalprotein. Owing to the lower induction capacity of Nodalprotein, only Activin A data are presented here. Forcomparison, the effect of a Nodal concentration gradi-ent on the analyzed mesoderm markers is included inSupplementary Fig. 1.
Brachyury expression marking pan-mesoderm andSox1 expression marking neuroectoderm were studiedto demonstrate the low effect of the Activin A/Nodalsignaling gradient on the establishment of the pan-mesoderm. In SFM, only 5 ng/ml of Activin A inducedBrachyury expression, and further increasing the Act-ivin A concentration reduced Brachyury levels back tocontrol, which was confirmed by WISH (Figs. 1C,2A,2E,2I). Sox1 expression on the other hand wasstrongly down-regulated by Activin A (Fig. 1C). In theloss-of-function setup, reduction of Activin A/Nodalsignaling after pan-mesoderm induction in SCM hadoccurred, only led to a slight decrease in Brachyuryand a strong induction of Sox1 (Fig. 1C). Furthermore,WISH analysis demonstrated that SB treatment didnot affect the Brachyury expressing domain in EB(Figs. 2Q,2U). The follow up of Brachyury expressionthus indicated that pan-mesoderm formation was hard-ly affected by the Activin A/Nodal activity gradient.
When looking at markers for posterior mesoderm celltypes in SFM cultures, we found that Evx1 and Mesp1but not Flk1, responded to the addition of higher Act-ivin A concentrations (Fig. 1D). WISH analysis ofEvx1 expression in EB showed that Activin A was in-deed inducing low levels of Evx1 in the cells of the EB(Figs. 2C,2G,2K). Moreover, Activin A/Nodal signal-ing inhibition with 10 mM SB led to reduced posterior
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
3
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)
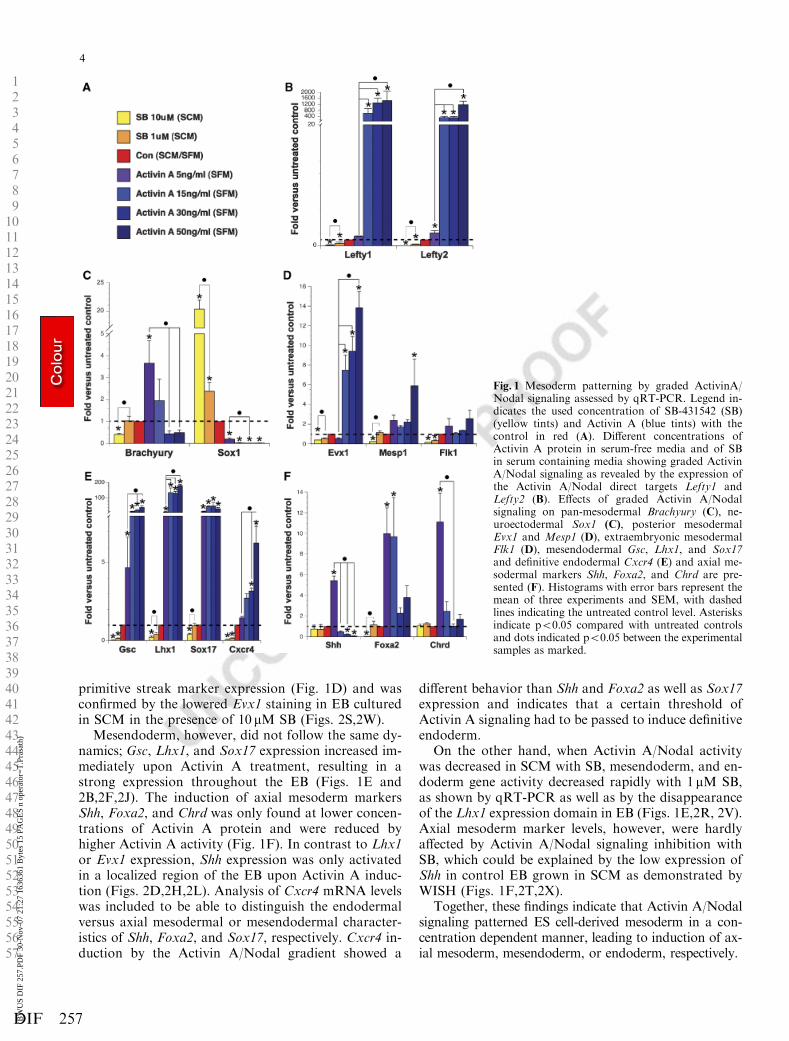
primitive streak marker expression (Fig. 1D) and wasconfirmed by the lowered Evx1 staining in EB culturedin SCM in the presence of 10 mM SB (Figs. 2S,2W).
Mesendoderm, however, did not follow the same dy-namics; Gsc, Lhx1, and Sox17 expression increased im-mediately upon Activin A treatment, resulting in astrong expression throughout the EB (Figs. 1E and2B,2F,2J). The induction of axial mesoderm markersShh, Foxa2, and Chrd was only found at lower concen-trations of Activin A protein and were reduced byhigher Activin A activity (Fig. 1F). In contrast to Lhx1or Evx1 expression, Shh expression was only activatedin a localized region of the EB upon Activin A induc-tion (Figs. 2D,2H,2L). Analysis of Cxcr4 mRNA levelswas included to be able to distinguish the endodermalversus axial mesodermal or mesendodermal character-istics of Shh, Foxa2, and Sox17, respectively. Cxcr4 in-duction by the Activin A/Nodal gradient showed a
different behavior than Shh and Foxa2 as well as Sox17expression and indicates that a certain threshold ofActivin A signaling had to be passed to induce definitiveendoderm.
On the other hand, when Activin A/Nodal activitywas decreased in SCM with SB, mesendoderm, and en-doderm gene activity decreased rapidly with 1 mM SB,as shown by qRT-PCR as well as by the disappearanceof the Lhx1 expression domain in EB (Figs. 1E,2R, 2V).Axial mesoderm marker levels, however, were hardlyaffected by Activin A/Nodal signaling inhibition withSB, which could be explained by the low expression ofShh in control EB grown in SCM as demonstrated byWISH (Figs. 1F,2T,2X).
Together, these findings indicate that Activin A/Nodalsignaling patterned ES cell-derived mesoderm in a con-centration dependent manner, leading to induction of ax-ial mesoderm, mesendoderm, or endoderm, respectively.
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
Fig. 1 Mesoderm patterning by graded ActivinA/Nodal signaling assessed by qRT-PCR. Legend in-dicates the used concentration of SB-431542 (SB)(yellow tints) and Activin A (blue tints) with thecontrol in red (A). Different concentrations ofActivin A protein in serum-free media and of SBin serum containing media showing graded ActivinA/Nodal signaling as revealed by the expression ofthe Activin A/Nodal direct targets Lefty1 andLefty2 (B). Effects of graded Activin A/Nodalsignaling on pan-mesodermal Brachyury (C), ne-uroectodermal Sox1 (C), posterior mesodermalEvx1 and Mesp1 (D), extraembryonic mesodermalFlk1 (D), mesendodermal Gsc, Lhx1, and Sox17and definitive endodermal Cxcr4 (E) and axial me-sodermal markers Shh, Foxa2, and Chrd are pre-sented (F). Histograms with error bars represent themean of three experiments and SEM, with dashedlines indicating the untreated control level. Asterisksindicate po0.05 compared with untreated controlsand dots indicated po0.05 between the experimentalsamples as marked.
4
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)
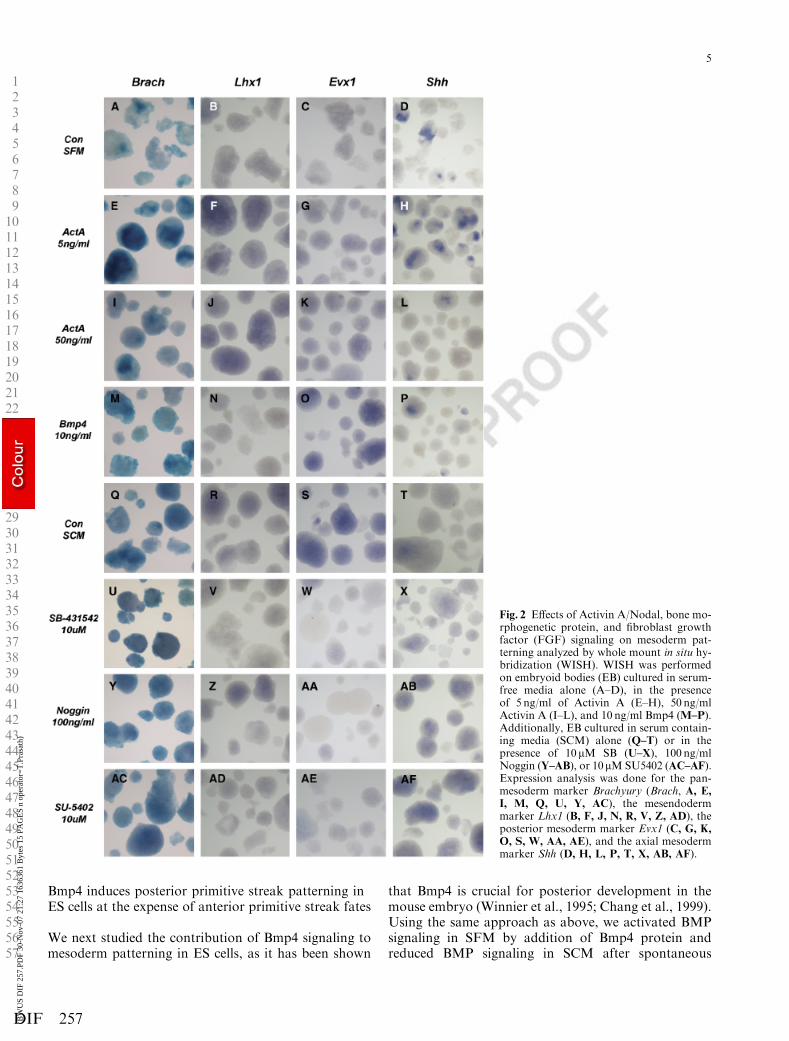
Bmp4 induces posterior primitive streak patterning inES cells at the expense of anterior primitive streak fates
We next studied the contribution of Bmp4 signaling tomesoderm patterning in ES cells, as it has been shown
that Bmp4 is crucial for posterior development in themouse embryo (Winnier et al., 1995; Chang et al., 1999).Using the same approach as above, we activated BMPsignaling in SFM by addition of Bmp4 protein andreduced BMP signaling in SCM after spontaneous
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
Fig. 2 Effects of Activin A/Nodal, bone mo-rphogenetic protein, and fibroblast growthfactor (FGF) signaling on mesoderm pat-terning analyzed by whole mount in situ hy-bridization (WISH). WISH was performedon embryoid bodies (EB) cultured in serum-free media alone (A–D), in the presenceof 5 ng/ml of Activin A (E–H), 50 ng/mlActivin A (I–L), and 10 ng/ml Bmp4 (M–P).Additionally, EB cultured in serum contain-ing media (SCM) alone (Q–T) or in thepresence of 10mM SB (U–X), 100 ng/mlNoggin (Y–AB), or 10mMSU5402 (AC–AF).Expression analysis was done for the pan-mesoderm marker Brachyury (Brach, A, E,I, M, Q, U, Y, AC), the mesendodermmarker Lhx1 (B, F, J, N, R, V, Z, AD), theposterior mesoderm marker Evx1 (C, G, K,
O, S, W, AA, AE), and the axial mesodermmarker Shh (D, H, L, P, T, X, AB, AF).
5
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)
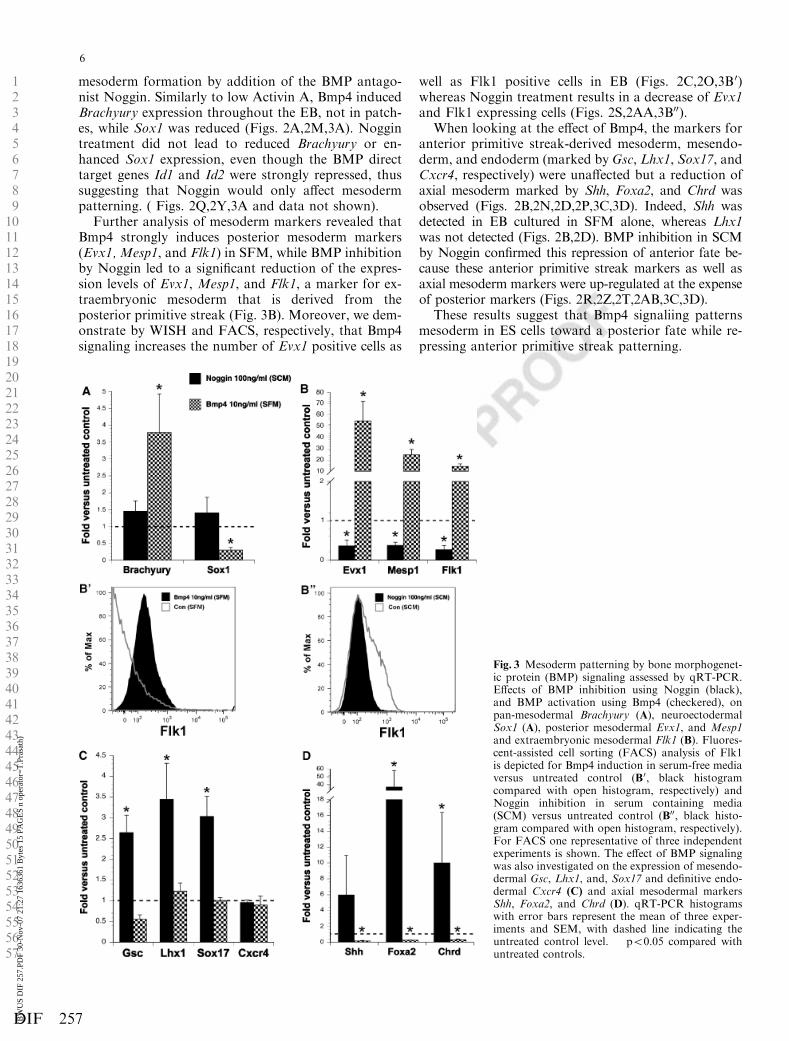
mesoderm formation by addition of the BMP antago-nist Noggin. Similarly to low Activin A, Bmp4 inducedBrachyury expression throughout the EB, not in patch-es, while Sox1 was reduced (Figs. 2A,2M,3A). Noggintreatment did not lead to reduced Brachyury or en-hanced Sox1 expression, even though the BMP directtarget genes Id1 and Id2 were strongly repressed, thussuggesting that Noggin would only affect mesodermpatterning. ( Figs. 2Q,2Y,3A and data not shown).
Further analysis of mesoderm markers revealed thatBmp4 strongly induces posterior mesoderm markers(Evx1, Mesp1, and Flk1) in SFM, while BMP inhibitionby Noggin led to a significant reduction of the expres-sion levels of Evx1, Mesp1, and Flk1, a marker for ex-traembryonic mesoderm that is derived from theposterior primitive streak (Fig. 3B). Moreover, we dem-onstrate by WISH and FACS, respectively, that Bmp4signaling increases the number of Evx1 positive cells as
well as Flk1 positive cells in EB (Figs. 2C,2O,3B 0)whereas Noggin treatment results in a decrease of Evx1and Flk1 expressing cells (Figs. 2S,2AA,3B00).
When looking at the effect of Bmp4, the markers foranterior primitive streak-derived mesoderm, mesendo-derm, and endoderm (marked by Gsc, Lhx1, Sox17, andCxcr4, respectively) were unaffected but a reduction ofaxial mesoderm marked by Shh, Foxa2, and Chrd wasobserved (Figs. 2B,2N,2D,2P,3C,3D). Indeed, Shh wasdetected in EB cultured in SFM alone, whereas Lhx1was not detected (Figs. 2B,2D). BMP inhibition in SCMby Noggin confirmed this repression of anterior fate be-cause these anterior primitive streak markers as well asaxial mesoderm markers were up-regulated at the expenseof posterior markers (Figs. 2R,2Z,2T,2AB,3C,3D).
These results suggest that Bmp4 signaliing patternsmesoderm in ES cells toward a posterior fate while re-pressing anterior primitive streak patterning.
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
Fig. 3 Mesoderm patterning by bone morphogenet-ic protein (BMP) signaling assessed by qRT-PCR.Effects of BMP inhibition using Noggin (black),and BMP activation using Bmp4 (checkered), onpan-mesodermal Brachyury (A), neuroectodermalSox1 (A), posterior mesodermal Evx1, and Mesp1and extraembryonic mesodermal Flk1 (B). Fluores-cent-assisted cell sorting (FACS) analysis of Flk1is depicted for Bmp4 induction in serum-free mediaversus untreated control (B0, black histogramcompared with open histogram, respectively) andNoggin inhibition in serum containing media(SCM) versus untreated control (B00, black histo-gram compared with open histogram, respectively).For FACS one representative of three independentexperiments is shown. The effect of BMP signalingwas also investigated on the expression of mesendo-dermal Gsc, Lhx1, and, Sox17 and definitive endo-dermal Cxcr4 (C) and axial mesodermal markersShh, Foxa2, and Chrd (D). qRT-PCR histogramswith error bars represent the mean of three exper-iments and SEM, with dashed line indicating theuntreated control level. � po0.05 compared withuntreated controls.
6
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)
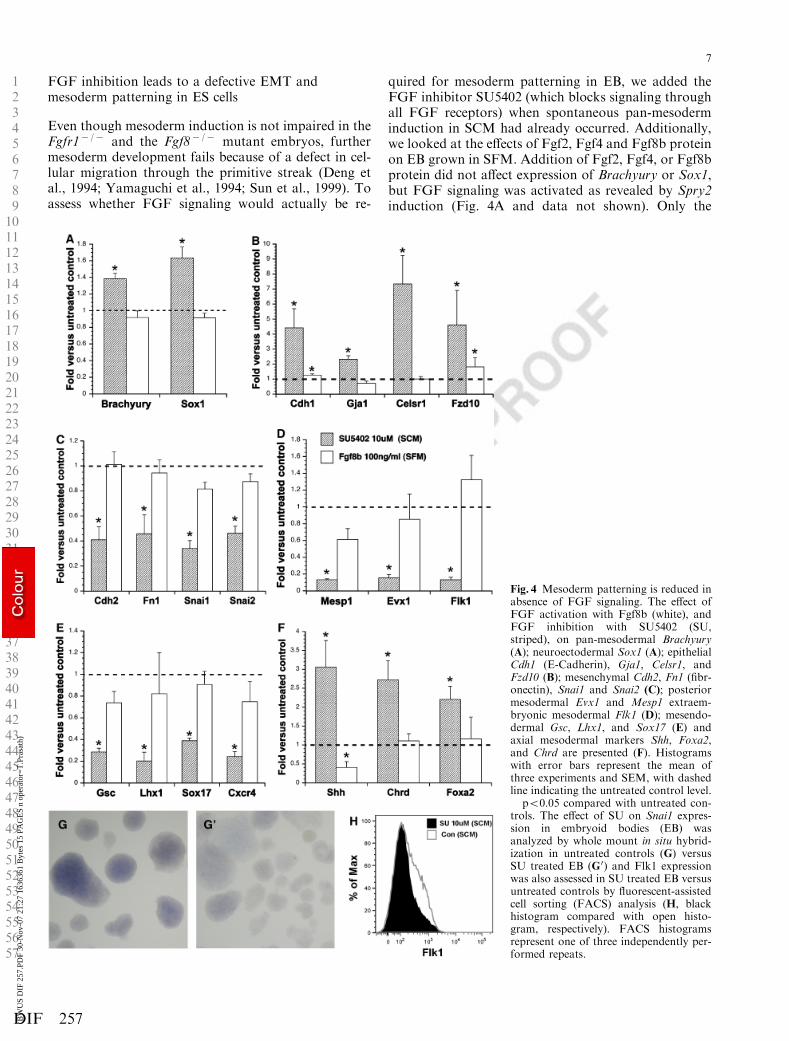
FGF inhibition leads to a defective EMT andmesoderm patterning in ES cells
Even though mesoderm induction is not impaired in theFgfr1� /� and the Fgf8� /� mutant embryos, furthermesoderm development fails because of a defect in cel-lular migration through the primitive streak (Deng etal., 1994; Yamaguchi et al., 1994; Sun et al., 1999). Toassess whether FGF signaling would actually be re-
quired for mesoderm patterning in EB, we added theFGF inhibitor SU5402 (which blocks signaling throughall FGF receptors) when spontaneous pan-mesoderminduction in SCM had already occurred. Additionally,we looked at the effects of Fgf2, Fgf4 and Fgf8b proteinon EB grown in SFM. Addition of Fgf2, Fgf4, or Fgf8bprotein did not affect expression of Brachyury or Sox1,but FGF signaling was activated as revealed by Spry2induction (Fig. 4A and data not shown). Only the
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
Fig. 4 Mesoderm patterning is reduced inabsence of FGF signaling. The effect ofFGF activation with Fgf8b (white), andFGF inhibition with SU5402 (SU,striped), on pan-mesodermal Brachyury(A); neuroectodermal Sox1 (A); epithelialCdh1 (E-Cadherin), Gja1, Celsr1, andFzd10 (B); mesenchymal Cdh2, Fn1 (fibr-onectin), Snai1 and Snai2 (C); posteriormesodermal Evx1 and Mesp1 extraem-bryonic mesodermal Flk1 (D); mesendo-dermal Gsc, Lhx1, and Sox17 (E) andaxial mesodermal markers Shh, Foxa2,and Chrd are presented (F). Histogramswith error bars represent the mean ofthree experiments and SEM, with dashedline indicating the untreated control level.� po0.05 compared with untreated con-trols. The effect of SU on Snai1 expres-sion in embryoid bodies (EB) wasanalyzed by whole mount in situ hybrid-ization in untreated controls (G) versusSU treated EB (G0) and Flk1 expressionwas also assessed in SU treated EB versusuntreated controls by fluorescent-assistedcell sorting (FACS) analysis (H, blackhistogram compared with open histo-gram, respectively). FACS histogramsrepresent one of three independently per-formed repeats.
7
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

treatment with Fgf8b is shown, because Fgf8b, the mostpotent isoform of Fgf8, is the only gene of the studiedFGF family members that has been found to be crucialin mesoderm development in the mouse (Guo and Li,2007). Similarly, mesoderm and neuroectoderm forma-tion was not affected by FGF inhibition with SU5402 inSCM (Figs. 2Q,2AC,4A).
As the primary defect in the Fgfr1� /� and Fgf8� /�
mutants is a deficient migration of mesoderm cells,caused by a failure in EMT, we first checked whetherlevels of epithelial and mesenchymal markers as well asEMT target genes were affected by FGF signaling inEB. Even though Fgf8b (or Fgf2 and Fgf4) addition toEB cultured in SFM did not reveal any effect on thesemarkers, FGF inhibition by SU5402 in EB maintainedin SCM revealed that the epithelial markers Cdh1 (thegene encoding E-cadherin), Gja1 (the gene encodingConnexin43), Celsr1, and Fzd10 were strongly enhanced(Fig. 4B). In contrast, the mesenchymal markers Cdh2(the gene encoding N-cadherin) and Fn1 (the gene en-coding fibronectin) including the EMT target genesSnai1 and Snai2 were reduced, suggesting that FGFinhibition in EB indeed resulted in a defective EMT(Fig. 4C). WISH analysis confirms that Snai1 expres-sion was lost in EB treated with SU5402 (Fig. 4G,4G 0).
We next addressed the question whether reducedFGF signaling had any effect on mesoderm patterning.Expression levels of posterior mesoderm markers Mesp1and Evx1 as well as Flk1 were strongly reduced in EBcultured in SCM in the presence of SU5402 (Fig. 4D)and the number of Evx1 and Flk1 positive cells hadclearly decreased (Figs. 2S,2AE,4H). Not only meso-derm derived from the posterior streak was affected butalso mesendoderm and endoderm markers (Gsc, Lhx1,Sox17, and Cxcr4) were reduced significantly in absenceof FGF signaling (Fig. 4E). By in situ hybridization, wefound that the Lhx1 domain was strongly decreased inintensity ( Fig. 2R,2AD). In contrast, the expression ofthe axial mesoderm markers (Shh, Foxa2 and Chrd) hadincreased throughout the EB, indicating that the forma-tion of axial mesoderm from ES cells is enhanced whenFGF activity is repressed (Figs. 2T,2F,4F).
Mesoderm patterning in ES cells by Activin A/Nodaland Bmp4 activity is FGF dependent
As we found that FGF signaling was required for theformation of posterior mesoderm, extraembryonic me-soderm, and mesendoderm but not axial mesoderm, wenext questioned whether mesoderm patterning by Act-ivin A/Nodal or Bmp4 signaling in SFM would beaffected in the absence of FGF activity.
Brachyury expression analysis revealed that FGF in-hibition did not affect mesoderm induction both byActivin A and by Bmp4, suggesting that this system
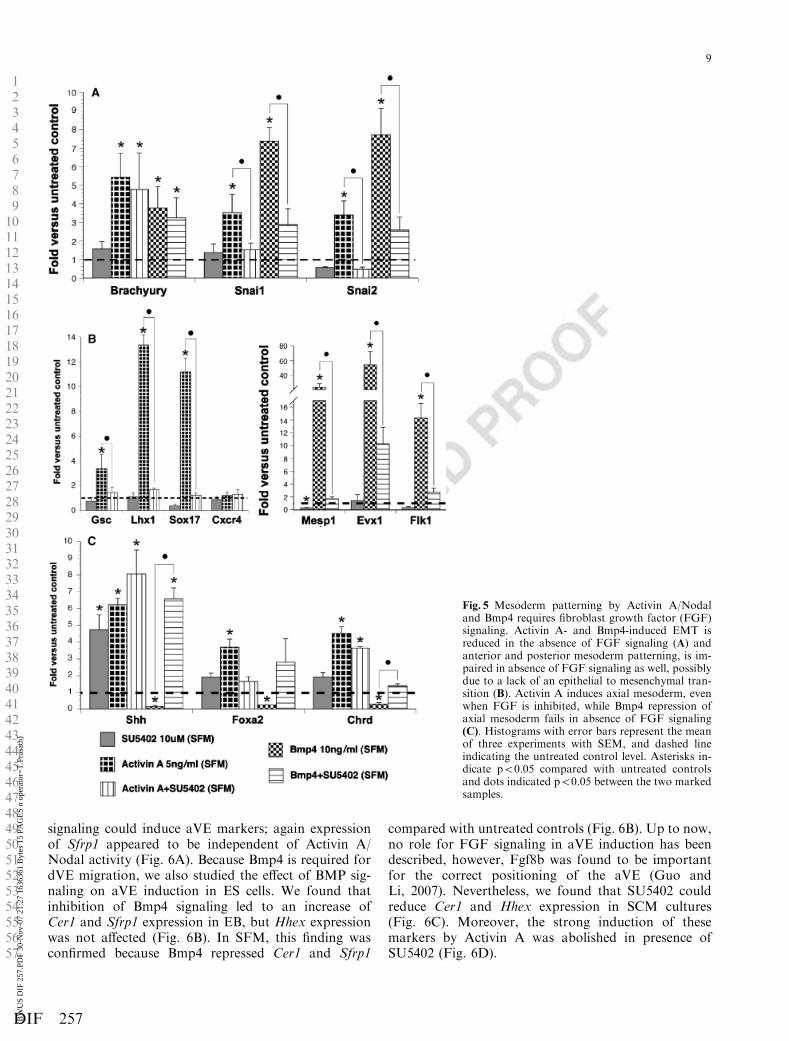
allowed an efficient interpretation of the role of FGFsignaling in mesoderm patterning (Fig. 5A).
Induction of Snai1 and Snai2 by Activin A and Bmp4suggest that both pathways are able to activate an EMT(Fig. 5A). Moreover, the increase in Snai1 and Snai2 geneexpression by Activin A or Bmp4 was blocked in thepresence of SU5402, indicating that Activin A and Bmp4induce an EMT via FGF signaling (Fig. 5A). Furtherconfirming the results shown in Fig. 4, anterior meso-derm patterning by Activin A and posterior and extra-embryonic mesoderm patterning by Bmp4 was stronglyreduced in the absence of FGF signaling (Fig. 5B).
Unlike the mesendoderm markers, axial mesodermmarkers Shh and Chrd were induced in the presence ofSU5402 alone (Fig. 5C). Activin A alone induced thesemarkers to similar levels, but could not further enhanceShh and Chrd induction by SU5402. As Foxa2 is alsodetected in the mesendoderm compartment, reducedFoxa2 levels in EB treated with Activin A and SU5042were not completely unexpected (Fig. 5C).
As shown in Fig. 5C, Bmp4 repressed axial meso-derm, but in the presence of SU5402, Bmp4 failed toblock Shh, Foxa2, and Chrd. As this phenomenon wasnot found for the mesendoderm markers (data notshown), this outcome suggested that FGF inhibitionalone is sufficient for axial mesoderm development frompan-mesoderm cells.
These results indicated that FGF signaling has animportant role in mesoderm patterning together withActivin A/Nodal and Bmp4 signaling, which probablyactivated FGF signaling in order to allow the epithelialprimitive streak cells to become mesenchymal meso-derm cells that can be patterned.
Anterior visceral endoderm differentiation in ES cellsdepends on Activin A/Nodal signaling
Up to now we have focused on the induction of pos-terior embryo tissues from ES cells, but in the embryo,the aVE has an equally important function in embryodevelopment. We therefore used our dual ES cell systemto reveal the molecular steps that were required to gen-erate aVE cells from ES cells. In the mouse embryo,Nodal and Bmp4 are required in dVE induction andmigration, respectively (Brennan et al., 2001; Soareset al., 2005). The aVE can be typically characterized byCer1 and Hhex expression and we previously demon-strated that Sfrp1 marks the aVE specifically (MartinezBarbera et al., 2000; Perea-Gomez et al., 2002; Kempet al., 2005). The expression of these three markers wastherefore further investigated when altering Activin A/Nodal, Bmp4, and FGF signaling.
When Activin A/Nodal signaling was inhibited with10 mM SB, Cer1 and Hhex were strongly down-regulat-ed, while Sfrp1 was not affected. Activation of the Act-ivin A/Nodal pathway confirmed that Activin A/Nodal
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
8
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)
signaling could induce aVE markers; again expressionof Sfrp1 appeared to be independent of Activin A/Nodal activity (Fig. 6A). Because Bmp4 is required fordVE migration, we also studied the effect of BMP sig-naling on aVE induction in ES cells. We found thatinhibition of Bmp4 signaling led to an increase ofCer1 and Sfrp1 expression in EB, but Hhex expressionwas not affected (Fig. 6B). In SFM, this finding wasconfirmed because Bmp4 repressed Cer1 and Sfrp1
compared with untreated controls (Fig. 6B). Up to now,no role for FGF signaling in aVE induction has beendescribed, however, Fgf8b was found to be importantfor the correct positioning of the aVE (Guo andLi, 2007). Nevertheless, we found that SU5402 couldreduce Cer1 and Hhex expression in SCM cultures(Fig. 6C). Moreover, the strong induction of thesemarkers by Activin A was abolished in presence ofSU5402 (Fig. 6D).
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
Fig. 5 Mesoderm patterning by Activin A/Nodaland Bmp4 requires fibroblast growth factor (FGF)signaling. Activin A- and Bmp4-induced EMT isreduced in the absence of FGF signaling (A) andanterior and posterior mesoderm patterning, is im-paired in absence of FGF signaling as well, possiblydue to a lack of an epithelial to mesenchymal tran-sition (B). Activin A induces axial mesoderm, evenwhen FGF is inhibited, while Bmp4 repression ofaxial mesoderm fails in absence of FGF signaling(C). Histograms with error bars represent the meanof three experiments with SEM, and dashed lineindicating the untreated control level. Asterisks in-dicate po0.05 compared with untreated controlsand dots indicated po0.05 between the two markedsamples.
9
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)
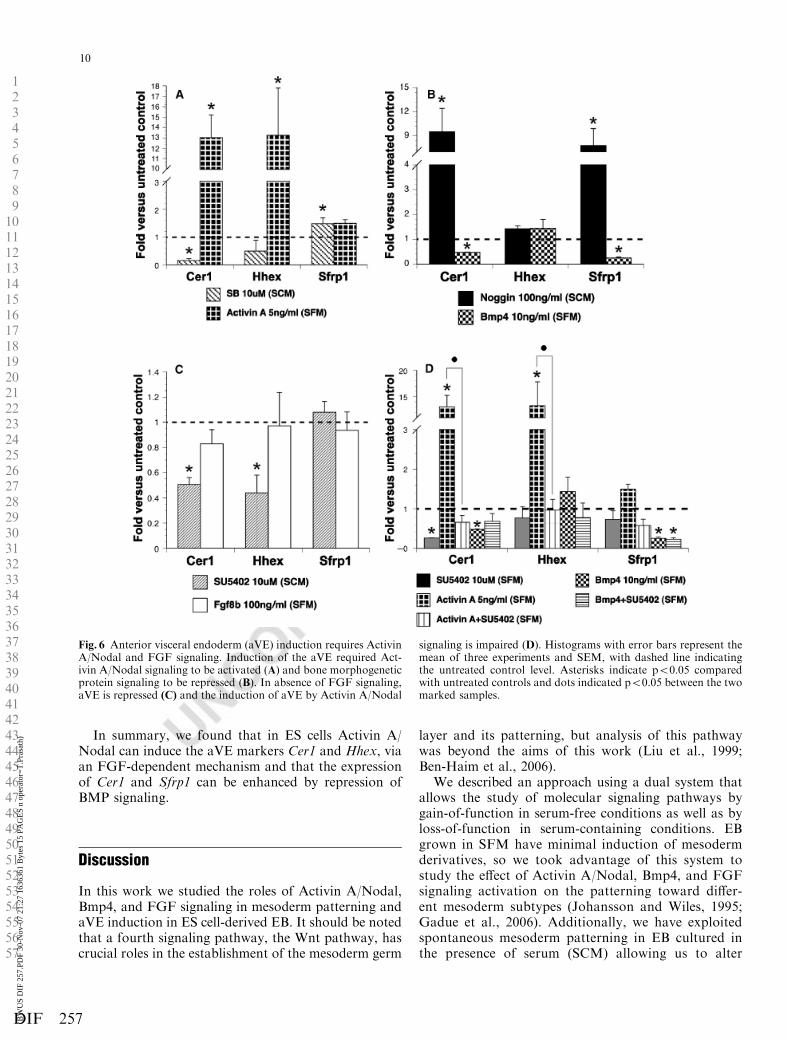
In summary, we found that in ES cells Activin A/Nodal can induce the aVE markers Cer1 and Hhex, viaan FGF-dependent mechanism and that the expressionof Cer1 and Sfrp1 can be enhanced by repression ofBMP signaling.
Discussion
In this work we studied the roles of Activin A/Nodal,Bmp4, and FGF signaling in mesoderm patterning andaVE induction in ES cell-derived EB. It should be notedthat a fourth signaling pathway, the Wnt pathway, hascrucial roles in the establishment of the mesoderm germ
layer and its patterning, but analysis of this pathwaywas beyond the aims of this work (Liu et al., 1999;Ben-Haim et al., 2006).
We described an approach using a dual system thatallows the study of molecular signaling pathways bygain-of-function in serum-free conditions as well as byloss-of-function in serum-containing conditions. EBgrown in SFM have minimal induction of mesodermderivatives, so we took advantage of this system tostudy the effect of Activin A/Nodal, Bmp4, and FGFsignaling activation on the patterning toward differ-ent mesoderm subtypes (Johansson and Wiles, 1995;Gadue et al., 2006). Additionally, we have exploitedspontaneous mesoderm patterning in EB cultured inthe presence of serum (SCM) allowing us to alter
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
Fig. 6 Anterior visceral endoderm (aVE) induction requires ActivinA/Nodal and FGF signaling. Induction of the aVE required Act-ivin A/Nodal signaling to be activated (A) and bone morphogeneticprotein signaling to be repressed (B). In absence of FGF signaling,aVE is repressed (C) and the induction of aVE by Activin A/Nodal
signaling is impaired (D). Histograms with error bars represent themean of three experiments and SEM, with dashed line indicatingthe untreated control level. Asterisks indicate po0.05 comparedwith untreated controls and dots indicated po0.05 between the twomarked samples.
10
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

patterning specifically by inhibiting Activin A/Nodal,BMP, and FGF activity .
Differentiation toward specific cell types was fol-lowed by quantifying expression levels of marker genesby qRT-PCR, as this method allows accurate monitor-ing of ES cell differentiation (Willems et al., 2006).Additionally, some markers were investigated usingWISH or FACS analysis as qRT-PCR does not allowinformation about spatial expression or the number ofmarker positive cells in the EB. The genes analyzed inthis study were selected based on their specific mRNAexpression in the different tissues formed in the gastrula(Supplementary Table 1). Moreover, the use of multiplemarkers for each tissue type allowed us to make moreaccurate conclusions.
By exploiting the combination of the two different cul-ture systems with qRT-PCR analysis and complementarycell population analysis, we could thus reveal the impor-tance of Activin A/Nodal, Bmp4, and FGF signaling inmesoderm patterning and aVE induction in ES cell-derivedEB. An overview of our most important findings in me-soderm patterning is illustrated in Supplementary Fig. 2.
Graded Activin A/Nodal signaling results in differentmesoderm subtypes
Nodal signaling has concentration dependent effects onmesoderm patterning in the mouse embryo as well as inXenopus animal caps (McDowell and Gurdon, 1999;Vincent et al., 2003). Additionally, Activin A or Nodalcan induce mesendoderm from ES cells (Kubo et al.,2004; Tada et al., 2005; Yasunaga et al., 2005). How-ever, none of these studies addressed the graded effect ofActivin A/Nodal signaling in the establishment of thedifferent mesoderm derivatives from ES cells. We there-fore generated a gradient of Activin A and Nodal ac-tivity to investigate whether different Activin A orNodal concentrations would allow the formation ofdifferent mesoderm subtypes. When comparing bothNodal and Activin A protein activity, we found thatboth proteins generally have the same effects on themesoderm markers studied, but that Activin A shows asignificant higher activity than Nodal. This could be dueto either lower activity of the Nodal protein, or becauseof Nodal induced inhibition by Leftys, whereas ActivinA cannot be inhibited by Lefty proteins. Therefore onlyActivin A is mentioned and shown as activator of Act-ivin A/Nodal signaling.
We found that Activin A would first induce axialmesoderm cells as well as mesendoderm, where furtherincrease of Activin A activity would lead to decreasedaxial mesoderm levels and enhanced mesendoderm.Secondly, a certain threshold of Activin A concentra-tion had to be passed to allow induction of endoderm asdemonstrated by Cxcr4 expression.
Reduction of Activin A/Nodal signaling further con-firmed the need of Activin A/Nodal signaling in the
specification of these mesoderm subtypes. Surprisingly,when mesoderm patterning by Activin A/Nodal signal-ing was blocked, we found a vast increase of Sox1 lev-els, even though Brachyury expression had notdecreased strongly. On the contrary, anterior meso-derm patterning by Activin A reduced Sox1 expression.In the Wnt3a� /� mutant for example, the lack of pat-terning in the primitive streak of the mouse embryoleads to enhanced neural induction, which indicates thatreduced mesoderm patterning can result in enhancedneural induction and vice versa (Takada et al., 1994).
We also found that Activin A induced markers forposterior mesoderm in cells within the EB. These ex-pression levels are however significantly lower (at least10 times) compared with mesendoderm induction andmight be caused by an indirect activation of the pat-terning of pan-mesodermal cells. It was recently dem-onstrated that the Activin A/Nodal pathway can induceBmp4, which then could induce posterior mesoderm(Ben-Haim et al., 2006).
Activin A/Nodal signaling could thus generate axialmesoderm, mesendoderm, or endoderm from ES cellsdepending on its concentration (Supplementary Fig. 2,Step 2–4). From these different subtypes one could thenfurther derive notochord, somites, cardiac mesoderm,or gut, respectively (Kinder et al., 2001; Tam et al.,2006). Closer analysis of the presented Activin A/Nodalgradient suggested that Activin A or Nodal might act asa morphogen in EB, as described in Xenopus (McDo-well and Gurdon, 1999; Gurdon and Bourillot, 2001).More experimental work would, however, be requiredto demonstrate that Nodal or Activin A could havediverse effects at different distances from a local ActivinA or Nodal source.
Bmp4 directs ES cell-derived mesoderm towardposterior mesoderm
That Bmp4 has an important role in fate decision in EScells was demonstrated by the fact that it can stronglyreduce neuroectoderm differentiation from ES cells andthat it can induce mesoderm (Johansson and Wiles,1995; Ying et al., 2003; Lindsley et al., 2006). Moreover,several recent studies have indicated a role for Bmp4 inthe differentiation of ES cells toward cardiac or he-matopoietic fates (Kennedy et al., 2006; Liu et al.,2007). We have, however, further investigated the needfor Bmp4 signaling in early mesoderm patterning.Bmp4 was necessary for the induction of Mesp1 andEvx1, two markers that are found in the posteriorprimitive streak. Mesp1 was recently described as oneof the key players that are required to extend somiticmesoderm and thus formation of the tail, suggestingthat tail truncations in the Bmp4� /� mutant mightbe due to reduced Mesp1 induction (Winnier et al.,1995; Morimoto et al., 2006). Using Noggin, our data
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
11
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

demonstrate that BMP signaling is indeed required forthe generation of posterior mesoderm from ES cells.Moreover, we confirmed that BMP signaling is neces-sary for the induction of hemangioblasts (Flk1 positivecells) (Kennedy et al., 2006).
More importantly, we show for the first time thatBMP signaling (activated by Bmp4) directs ES cell-de-rived mesoderm toward a posterior fate at the expenseof more anterior primitive streak derivatives (Supple-mentary Fig. 2, Step 1, 2, and 4). This finding, based ongain-of-function and loss-of-function, suggests that notonly Activin A/Nodal signaling is required to patternthe anterior primitive streak cells in EB, but that Bmp4has a repressive effect on mesendoderm and axial me-soderm differentiation in EB, which needs to be atten-uated for efficient anterior patterning by for exampleNoggin and Chrd expression in the node (Bachiller et al.,2000). It is unclear how Bmp4 would repress patterningin the anterior primitive streak, but a study in Xenopusindicates that Bmp4 might repress Nodal function, andthus anterior primitive streak patterning, on the level ofthe Smads by competing for commonly used Smads(Candia et al., 1997). Moreover, Smad4 inactivation inthe mouse leads to enhanced posterior patterning andreduced anterior patterning and is similar to the effectof Bmp4 we found (Chu et al., 2004).
Whereas Activin A specifically patterned anteriormesoderm subtypes from ES cells, we found that Bmp4acted as a switch factor that determines whether ES cell-derived mesoderm would form anterior or posteriorprimitive streak cells.
FGF signaling is required for mesoderm patterning inES cells
Even though we demonstrated that Activin A or Nodaland Bmp4 have crucial roles in anterior versus posteriorpatterning in ES cells, the Fgf8� /� and the Fgfr1� /�
mutants also suggest that FGF signaling has an impor-tant function in mesoderm patterning (Deng et al.,1994; Yamaguchi et al., 1994; Sun et al., 1999). As me-soderm patterning in these mutants fails because of adefective EMT, we first investigated whether inhibitionof FGF signaling would have such an effect in differ-entiating ES cells. By inhibition of FGF signaling withthe chemical inhibitor SU-5402, we were able to blockall FGF signaling through Fgfr1-3 and probably Fgfr4in EB (Mohammadi et al., 1997; Dvorak et al., 2005;Masih-Khan et al., 2006). Addition of Fgf2, Fgf4, orFgf8b allowed us to activate FGF signaling, but wecould however not demonstrate an effect of these FGFson the EMT or mesoderm patterning. Inhibition bySU5402 blocks all FGF signaling whereas we investi-gated the FGFs known to be present in the primitivestreak. These data suggest that there might be otheruncharacterized FGF proteins that could be required in
the EMT process in the primitive streak. Alternatively,the FGF proteins tested here might need the presence ofa mesoderm patterning molecule to allow their functionin the primitive streak. Indeed, Faloon et al. (2000)demonstrated that in ES cells, Fgf2 can synergize withActivin A to enhance hematopoietic development.
When looking into the EMT, we found that the cellsin the EB indeed retained a more epithelial character atthe expense of a mesenchymal one as shown by in-creased E-Cadherin levels, lost expression of the Snailgenes and reduced Fibronectin. Using Fzd10 as a markerof future mesoderm cells that still need to migrate out ofthe primitive streak, our data further confirm that inabsence of FGF signaling ES cells were converted toprimitive streak or pan-mesoderm, but that they re-tained an epithelial primitive streak character instead ofdifferentiating toward one of the mesoderm subtypes(Kemp et al., 2007). Analysis of the Snail genes dem-onstrated that Bmp4 and Activin A activate an EMT inEB, and that in the presence of SU5402, the inductionof these EMT genes is lost, indicating that Bmp4 andActivin A induce an FGF-dependent EMT before me-soderm cells are patterned in EB toward anterior orposterior mesoderm.
When looking at markers for patterned mesoderm,we found that mesoderm patterning in SCM in absenceof FGF signaling toward posterior and extraembryonicmesoderm as well as mesendoderm failed (Supplemen-tary Fig. 2, Step 1–3). Similarly, in the presence of Act-ivin A or Bmp4, FGF inhibition indeed led to impairedanterior and posterior mesoderm patterning, respec-tively. In contrast, we found that the expression of Shh,Foxa2, and Chrd, marking the cells directly anterior tothe anterior primitive streak that are destined to formthe axial mesoderm, was enhanced instead of decreasedin the absence of FGF signaling (Supplementary Fig. 2,Step 4). This finding suggests that node formation isregulated by FGF signaling in addition to Activin A/Nodal signaling. Indeed, in the presence of Activin A,FGF inhibition led to reduced mesendoderm, but axialmesoderm induction by Activin A signaling was notaffected. The fact that FGF inhibition could lead toenhanced axial mesoderm formation in ES cells can befurther explained by the fact that node cells retain anepithelial character and therefore do not undergo anEMT. In the absence of FGF signaling, more epithelial-type cells would be formed that cannot differentiate to-ward mesendoderm in the presence of Activin A orNodal, but would become node or axial mesoderm cellsinstead.
In general, our findings show similarities with thedefects observed in Fgfr1� /� and Fgf8� /� mutantembryos (Deng et al., 1994; Yamaguchi et al., 1994; Sunet al., 1999; Guo and Li, 2007). Based on these mutants,we did not expect a reduction of Bmp4 induced Flk1,because the extraembryonic tissues derived from theprimitive streak are normal in Fgfr1� /� and Fgf8� /�
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
12
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

mutant embryos (Deng et al., 1994; Yamaguchi et al.,1994; Sun et al., 1999; Guo and Li, 2007). However,SU5402 does not only block signaling through Fgfr1,but also through Fgfr2, which might explain why Flk1was reduced, because Fgfr2 is strongly expressed in theextraembryonic region and could be regulating Flk1expression (Orr-Urtreger et al., 1991; Mohammadiet al., 1997; Dvorak et al., 2005).
All together, our data demonstrated that ES cell-derived mesoderm cells need to obtain a mesenchymalcharacter via an FGF-dependent mechanism allowingtheir further patterning.
aVE induction requires Activin A/Nodal and FGFsignaling but not Bmp4 activity
In our ES cell system, we were not only able to studymesoderm patterning, but we could also study the es-tablishment of the aVE. Because day 4 EB correspondto a gastrula stage embryo, alteration of signaling path-ways could also affect the induction/maintenance of thesignaling center on the anterior side of the embryo. Byblocking and activating Activin A/Nodal signaling wecould demonstrate that aVE markers were indeed in-duced by Activin A in ES cells and that this induction isFGF dependent. On the other hand, Bmp4 was able torepress aVE markers, indicating that Bmp4 should beblocked to obtain enriched populations of aVE cells.
In the embryo, Bmp4 knockdown leads to an en-larged aVE as demonstrated by an enlarged expressiondomain of the marker Cer1, but in EB it was difficult tostudy aVE induction by the expression of the typicalaVE marker Cer1, because it is also expressed in theanterior primitive streak and is even used as a mesen-doderm marker (Soares et al., 2005; Gadue et al., 2006).We therefore included the markers Hhex and Sfrp1,which are detected more specifically in the aVE, Hhexhowever, is also detected in the most anterior definitiveendoderm at late streak stages (Thomas et al., 1998;Kemp et al., 2005). Even then, we still noted severaldiscrepancies when using Cer1, Hhex, and Sfrp1 asmarkers for the aVE. Cer1 was regulated by ActivinA/Nodal, FGF and Bmp4 signaling, whereas Sfrp1 andHhex were independent of Activin A and Bmp4 signal-ing, respectively. A different regulation of the aVEmarkers Hhex and Cer1 was also described in theb-Catenin� /� mutant, supporting our findings (Haegelet al., 1995; Huelsken et al., 2000). Therefore, we couldconclude that in EB, Bmp4, and a FGF-dependentActivin A/Nodal signaling indeed regulate the aVElike cells.
All together, we found that Activin A could inducethe aVE markers Cer1 andHhex via an FGF-dependentmechanism and that Bmp4 was able to repress theaVE markers Sfrp1 and Cer1 independently of FGFsignaling.
In conclusion, we have identified different key playersthat are required for the patterning of ES cell-derivedpan-mesoderm cells. First, we demonstrated that Act-ivin A or Nodal could induce axial mesoderm, mesen-doderm, or endoderm, depending on the level of ActivinA/Nodal signaling. Secondly, we found that Bmp4 act-ed as a switch from anterior to posterior mesoderm inES cells. Bmp4 activation led to an enhanced posteriormesoderm compartment, whereas Bmp4 attenuation al-lowed an enhanced anterior primitive streak patterning.Thirdly, we identified FGF signaling as a key partner inmesoderm patterning. FGF activity was required forboth Activin A/Nodal and Bmp4 patterning, by allow-ing an EMT of the newly formed mesoderm cells.Moreover, FGF signaling acted as an important re-pressor of axial mesoderm.
By combining loss-of-function and gain-of-functionES cell culture systems, we were thus able to determinethe early molecular steps that were required for thepatterning and EMT of mesoderm derived from ES cellsand for the induction of aVE cells from ES cells.
Acknowledgments The authors would like to thank J. Van-desompele for his help with statistical analysis, C. Kemp for herhelp with WISH and critically reading the manuscript and M. Gu-illiams for his help with FACS analysis. This research was fundedby the Horizontal Action of the Vrije Universiteit Brussel (GrantHOA1), the Flemish Fund for Scientific Research (GrantG.0485.06), and the EU (6th Framework, Grant NEST012930).
References
Bachiller, D., Klingensmith, J., Kemp, C., Belo, J.A., Anderson,R.M., May, S.R., McMahon, J.A., McMahon, A.P., Harland,R.M., Rossant, J. and De Robertis, E.M. (2000) The organizerfactors Chordin and Noggin are required for mouse forebraindevelopment. Nature 403:658–661.
Belo, J.A., Bachiller, D., Agius, E., Kemp, C., Borges, A.C., Mar-ques, S., Piccolo, S. and De Robertis, E.M. (2000) Cerberus-likeis a secreted BMP and nodal antagonist not essential for mousedevelopment. Genesis 26:265–270.
Ben-Haim, N., Lu, C., Guzman-Ayala, M., Pescatore, L., Mesnard,D., Bischofberger, M., Naef, F., Robertson, E.J. and Constam,D.B. (2006) The nodal precursor acting via activin receptors in-duces mesoderm by maintaining a source of its convertases andBMP4. Dev Cell 11:313–323.
Blum, M., Andre, P., Muders, K., Schweickert, A., Fischer, A.,Bitzer, E., Bogusch, S., Beyer, T., van Straaten, H.W. and Vie-bahn, C. (2007) Ciliation and gene expression distinguish be-tween node and posterior notochord in the mammalian embryo.Differentiation 75:133–146.
Brennan, J., Lu, C.C., Norris, D.P., Rodriguez, T.A., Beddington,R.S. and Robertson, E.J. (2001) Nodal signalling in the epiblastpatterns the early mouse embryo. Nature 411:965–969.
Candia, A.F., Watabe, T., Hawley, S.H., Onichtchouk, D., Zhang,Y., Derynck, R., Niehrs, C. and Cho, K.W. (1997) Cellular in-terpretation of multiple TGF-beta signals: intracellular antago-nism between activin/BVg1 and BMP-2/4 signaling mediated bySmads. Development 124:4467–4480.
Chang, H., Huylebroeck, D., Verschueren, K., Guo, Q., Matzuk,M.M. and Zwijsen, A. (1999) Smad5 knockout mice die at mid-gestation due to multiple embryonic and extraembryonic defects.Development 126:1631–1642.
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
13
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

Chu, G.C., Dunn, N.R., Anderson, D.C., Oxburgh, L. and Rob-ertson, E.J. (2004) Differential requirements for Smad4 in TGF-beta-dependent patterning of the early mouse embryo.Development 131:3501–3512.
Ciruna, B. and Rossant, J. (2001) FGF signaling regulates meso-derm cell fate specification and morphogenetic movement at theprimitive streak. Dev Cell 1:37–49.
Ciruna, B.G., Schwartz, L., Harpal, K., Yamaguchi, T.P. andRossant, J. (1997) Chimeric analysis of fibroblast growth factorreceptor-1 (Fgfr1) function: a role for FGFR1 in morphogeneticmovement through the primitive streak. Development 124:2829–2841.
Conlon, F.L., Lyons, K.M., Takaesu, N., Barth, K.S., Kispert, A.,Herrmann, B. and Robertson, E.J. (1994) A primary requirementfor nodal in the formation and maintenance of the primitivestreak in the mouse. Development 120:1919–1928.
D’Amour, K.A., Bang, A.G., Eliazer, S., Kelly, O.G., Agulnick,A.D., Smart, N.G., Moorman, M.A., Kroon, E., Carpenter,M.K. and Baetge, E.E. (2006) Production of pancreatic hor-mone-expressing endocrine cells from human embryonic stemcells. Nat Biotechnol 24:1392–1401.
Deng, C.X., Wynshaw-Boris, A., Shen, M.M., Daugherty, C., Or-nitz, D.M. and Leder, P. (1994) Murine FGFR-1 is required forearly postimplantation growth and axial organization. GenesDev 8:3045–3057.
Dvorak, P., Dvorakova, D., Koskova, S., Vodinska, M., Najvir-tova, M., Krekac, D. and Hampl, A. (2005) Expression and po-tential role of fibroblast growth factor 2 and its receptors inhuman embryonic stem cells. Stem Cells 23:1200–1211.
Echelard, Y., Epstein, D.J., St-Jacques, B., Shen, L., Mohler, J.,McMahon, J.A. and McMahon, A.P. (1993) Sonic hedgehog, amember of a family of putative signaling molecules, is implicatedin the regulation of CNS polarity. Cell 75:1417–1430.
Evans, M.J. and Kaufman, M.H. (1981) Establishment in culture ofpluripotential cells from mouse embryos. Nature 292:154–156.
Faloon, P., Arentson, E., Kazarov, A., Deng, C.X., Porcher, C.,Orkin, S. and Choi, K. (2000) Basic fibroblast growth factorpositively regulates hematopoietic development. Development127:1931–1941.
Gadue, P., Huber, T.L., Paddison, P.J. and Keller, G.M. (2006)Wnt and TGF-{beta} signaling are required for the induction ofan in vitro model of primitive streak formation using embryonicstem cells. Proc Natl Acad Sci USA.
Guo, Q. and Li, J.Y. (2007) Distinct functions of the major Fgf8spliceform, Fgf8b, before and during mouse gastrulation. Devel-opment 134:2251–2260.
Gurdon, J.B. and Bourillot, P.Y. (2001) Morphogen gradient in-terpretation. Nature 413:797–803.
Haegel, H., Larue, L., Ohsugi, M., Fedorov, L., Herrenknecht, K.and Kemler, R. (1995) Lack of beta-catenin affects mouse de-velopment at gastrulation. Development 121:3529–3537.
Huelsken, J., Vogel, R., Brinkmann, V., Erdmann, B., Birchmeier,C. and Birchmeier, W. (2000) Requirement for beta-catenin inanterior-posterior axis formation in mice. J Cell Biol 148:567–578.
Ikeya, M. and Takada, S. (1998) Wnt signaling from the dorsalneural tube is required for the formation of the medialdermomyotome. Development 125:4969–4976.
Johansson, B.M. and Wiles, M.V. (1995) Evidence for involvementof activin A and bone morphogenetic protein 4 in mammalianmesoderm and hematopoietic development. Mol Cell Biol15:141–151.
Kemp, C., Willems, E., Abdo, S., Lambiv, L. and Leyns, L. (2005)Expression of all Wnt genes and their secreted antagonists duringmouse blastocyst and postimplantation development. Dev Dyn233:1064–1075.
Kemp, C., Willems, E., Wawrzak, D., Hendrickx, M., AgborAgbor, T. and Leyns, L. (2007) Expression of Frizzled5, Frizz-led7, and Frizzled10 during early mouse development and inter-actions with canonical Wnt signaling. Dev Dyn, in press.
Kennedy, M., D’Souza, S.L., Lynch-Kattman, M., Schwantz, S.and Keller, G. (2006) Development of the hemangioblast definesthe onset of hematopoiesis in human ES cell differentiation cul-tures Q10. Blood.
Kinder, S.J., Tsang, T.E., Wakamiya, M., Sasaki, H., Behringer,R.R., Nagy, A. and Tam, P.P. (2001) The organizer of the mousegastrula is composed of a dynamic population of progenitor cellsfor the axial mesoderm. Development 128:3623–3634.
Kubo, A., Shinozaki, K., Shannon, J.M., Kouskoff, V., Kennedy,M., Woo, S., Fehling, H.J. and Keller, G. (2004) Development ofdefinitive endoderm from embryonic stem cells in culture. De-velopment 131:1651–1662.
Lindsley, R.C., Gill, J.G., Kyba, M., Murphy, T.L. and Murphy,K.M. (2006) Canonical Wnt signaling is required for develop-ment of embryonic stem cell-derived mesoderm. Development133:3787–3796.
Liu, P., Wakamiya, M., Shea, M.J., Albrecht, U., Behringer, R.R.and Bradley, A. (1999) Requirement for Wnt3 in vertebrate axisformation. Nat Genet 22:361–365.
Liu, Y., Asakura, M., Inoue, H., Nakamura, T., Sano, M., Niu, Z.,Chen, M., Schwartz, R.J. and Schneider, M.D. (2007) Sox17 isessential for the specification of cardiac mesoderm in embryonicstem cells. Proc Natl Acad Sci USA 104:3859–3864.
Martinez Barbera, J.P., Clements, M., Thomas, P., Rodriguez, T.,Meloy, D., Kioussis, D. and Beddington, R.S. (2000) Thehomeobox gene Hex is required in definitive endodermal tissuesfor normal forebrain, liver and thyroid formation. Development127:2433–2445.
Masih-Khan, E., Trudel, S., Heise, C., Li, Z., Paterson, J., Nadeem,V., Wei, E., Roodman, D., Claudio, J.O., Bergsagel, P.L. andStewart, A.K. (2006) MIP-1alpha (CCL3) is a downstream targetof FGFR3 and RAS-MAPK signaling in multiple myeloma.Blood 108:3465–3471.
McDowell, N. and Gurdon, J.B. (1999) Activin as a morphogenin Xenopus mesoderm induction. Semin Cell Dev Biol 10:311–317.
Mohammadi, M., McMahon, G., Sun, L., Tang, C., Hirth, P., Yeh,B.K., Hubbard, S.R. and Schlessinger, J. (1997) Structures of thetyrosine kinase domain of fibroblast growth factor receptor incomplex with inhibitors. Science 276:955–960.
Morimoto, M., Kiso, M., Sasaki, N. and Saga, Y. (2006) Coop-erative Mesp activity is required for normal somitogenesis alongthe anterior–posterior axis. Dev Biol 300:687–698.
Orr-Urtreger, A., Givol, D., Yayon, A., Yarden, Y. and Lonai, P.(1991) Developmental expression of two murine fibroblastgrowth factor receptors, flg and bek. Development 113:1419–1434.
Pattyn, F., Speleman, F., De Paepe, A. and Vandesompele, J.(2003) RTPrimerDB: the real-time PCR primer and probe da-tabase. Nucleic Acids Res 31:122–123.
Perea-Gomez, A., Vella, F.D., Shawlot, W., Oulad-Abdelghani,M., Chazaud, C., Meno, C., Pfister, V., Chen, L., Robertson, E.,Hamada, H., Behringer, R.R. and Ang, S.L. (2002) Nodal an-tagonists in the anterior visceral endoderm prevent the formationof multiple primitive streaks. Dev Cell 3:745–756.
Pfendler, K.C., Catuar, C.S., Meneses, J.J. and Pedersen, R.A.(2005) Overexpression of Nodal promotes differentiation ofmouse embryonic stem cells into mesoderm and endoderm at theexpense of neuroectoderm formation. Stem Cells Dev 14:162–172.
Shawlot, W. and Behringer, R.R. (1995) Requirement for Lim1 inhead-organizer function. Nature 374:425–430.
Smith, A.G. (2001) Embryo-derived stem cells: of mice and men.Annu Rev Cell Dev Biol 17:435–462.
Soares, M.L., Haraguchi, S., Torres-Padilla, M.E., Kalmar, T.,Carpenter, L., Bell, G., Morrison, A., Ring, C.J., Clarke, N.J.,Glover, D.M. and Zernicka-Goetz, M. (2005) Functional studiesof signaling pathways in peri-implantation development of themouse embryo by RNAi. BMC Dev Biol 5:28 Q11.
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
14
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

Sun, X., Meyers, E.N., Lewandoski, M. and Martin, G.R. (1999)Targeted disruption of Fgf8 causes failure of cell migration in thegastrulating mouse embryo. Genes Dev 13:1834–1846.
Tada, S., Era, T., Furusawa, C., Sakurai, H., Nishikawa, S., Kino-shita, M., Nakao, K. and Chiba, T. (2005) Characterization ofmesendoderm: a diverging point of the definitive endoderm andmesoderm in embryonic stem cell differentiation culture. Devel-opment 132:4363–4374.
Takada, S., Stark, K.L., Shea, M.J., Vassileva, G., McMahon, J.A.and McMahon, A.P. (1994) Wnt-3a regulates somite and tailbudformation in the mouse embryo. Genes Dev 8:174–189.
Tam, P.P., Gad, J.M., Kinder, S.J., Tsang, T.E. and Behringer,R.R. (2001) Morphogenetic tissue movement and the establish-ment of body plan during development from blastocyst to gas-trula in the mouse. Bioessays 23:508–517.
Tam, P.P. and Loebel, D.A. (2007) Gene function in mouse em-bryogenesis: get set for gastrulation. Nat Rev Genet 8:368–381.
Tam, P.P., Loebel, D.A. and Tanaka, S.S. (2006) Building themouse gastrula: signals, asymmetry and lineages. Curr OpinGenet Dev 16:419–425.
Thomas, P.Q., Brown, A. and Beddington, R.S. (1998) Hex: ahomeobox gene revealing peri-implantation asymmetry in themouse embryo and an early transient marker of endothelial cellprecursors. Development 125:85–94.
Thomson, J.A., Itskovitz-Eldor, J., Shapiro, S.S., Waknitz, M.A.,Swiergiel, J.J., Marshall, V.S. and Jones, J.M. (1998) Embryonicstem cell lines derived from human blastocysts. Science 282:1145–1147.
Vincent, S.D., Dunn, N.R., Hayashi, S., Norris, D.P. and Robert-son, E.J. (2003) Cell fate decisions within the mouse organizer aregoverned by graded Nodal signals. Genes Dev 17:1646–1662.
Wilkinson, D.G., Bhatt, S. and Herrmann, B.G. (1990) Expressionpattern of the mouse T gene and its role in mesoderm formation.Nature 343:657–659.
Willems, E., Mateizel, I., Kemp, C., Cauffman, G., Sermon, K. andLeyns, L. (2006) Selection of reference genes in mouse embryosand in differentiating human and mouse ES cells. Int J Dev Biol50:627–635.
Winnier, G., Blessing, M., Labosky, P.A. and Hogan, B.L. (1995)Bone morphogenetic protein-4 is required for mesoderm forma-tion and patterning in the mouse. Genes Dev 9:2105–2116.
Yamaguchi, T.P., Harpal, K., Henkemeyer, M. and Rossant, J.(1994) fgfr-1 is required for embryonic growth and mesodermalpatterning during mouse gastrulation. Genes Dev 8:3032–3044.
Yasunaga, M., Tada, S., Torikai-Nishikawa, S., Nakano, Y., Ok-ada, M., Jakt, L.M., Nishikawa, S., Chiba, T., Era, T. andNishikawa, S. (2005) Induction and monitoring of definitive andvisceral endoderm differentiation of mouse ES cells. NatBiotechnol 23:1542–1550.
Ying, Q.L., Nichols, J., Chambers, I. and Smith, A. (2003) BMPinduction of Id proteins suppresses differentiation and sustainsembryonic stem cell self-renewal in collaboration with STAT3.Cell 115:281–292.
Zhang, H. and Bradley, A. (1996) Mice deficient for BMP2 arenonviable and have defects in amnion/chorion and cardiac de-velopment. Development 122:2977–2986.
Supplementary Material
The following supplementary material is available forthis article online:
Fig. S1. Mesoderm patterning by graded ActivinA/Nodal signaling assessed by qRT-PCR. Legend indi-cates the used concentration of SB-431542 (SB) (yellowtints) and Nodal (blue tints) with the control in red (A).Different concentrations of nodal protein in SFM andof SB in SCM showing graded Activin A/Nodal sig-naling as revealed by the expression of the Nodal directtargets Lefty1 and Lefty2 (B). Effects of graded Nodalsignaling on pan-mesodermal Brachyury (C), ne-uroectodermal Sox1 (C), posterior mesodermal Evx1and Mesp1 (D), extraembryonic mesodermal Flk1 (D),mesendodermal Gsc, Lhx1 and Sox17 and definitiveendodermal Cxcr4 (E) and axial mesodermal markersShh, Foxa2 and Chrd are presented (F). Histogramswith error bars represent the mean of three experimentsand SEM, with dashed lines indicating the untreatedcontrol level. Asterisks indicate po0.05 compared tountreated controls and dots indicated po0.05 betweenthe experimental samples as marked.
Fig. S2. Schematic representation of the major find-ings in mesoderm patterning. Circles represent differentmesoderm subtypes that can be marked with specificgenes. Arrows indicate the direction from a pan-meso-derm cell towards a more differentiated subtype. Theeffectors that induce mesoderm subtypes are indicatedwith a plus, the ones that repress are indicated with aminus. Patterning towards posterior and extraembry-onic mesoderm (1), mesendoderm (2), endoderm (3),and axial mesoderm (4) are indicated.
Table. S1. Specific markers in the gastrulating mouseembryo used in this study.
This material is available as part of the online articlefrom: http://www.blackwell-synergy.com/doi/abs/10.1111/j.1432-0436.2007.00244.x (This link will take youto the article abstract).
Please note: Blackwell Publishing is not responsiblefor the content or functionality of any supplementarymaterials supplied by the authors. Any queries (otherthan missing material) should be directed to the corre-sponding author for the article.
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
15
DIF 257(BW
US
DIF
257
.PD
F 30
-Nov
-07
21:2
7 16
3636
1 B
ytes
15
PAG
ES
n op
erat
or=
T.P
rasa
th)

Author Query Form
_______________________________________________________
_______________________________________________________
Dear Author,
During the copy-editing of your paper, the following queries arose. Please respond to these by marking up your proofs with the necessary changes/additions. Please write your answers clearly on the query sheet if there is insufficient space on the page proofs. If returning the proof by fax do not write too close to the paper's edge. Please remember that illegible mark-ups may delay publication.
Journal DIFArticle 257
Query No. Description Author Response
Q1AQ: Please provide the manufacturer's information for GIBCO: town, state (if USA) and country.
Q2AQ: Please provide the manufacturer information for Hyclone: town, state (if USA) and country.
Q3AQ: Please provide the manufacturer information for Sigma: town and state (if USA).
Q4AQ: Please provide the manufacturer information for R&D Systems: town, state (if USA) and country.
Q5AQ: Please provide the manufacturer information for Promega: town, state (if USA) and country.
Q6AQ: Please provide the manufacturer information for eBioscience: town, state (if USA) and country.
Q7AQ: Please provide the manufacturer information for BD Biosciences: town, state (if USA) and country.
Q8AQ: Please provide the volume number for the reference Gadue et al. (2006).
Q9AQ: Please provide the volume number and page range for the reference Kemp et al. (2007).
Q10AQ: Please provide the volume number and page range for the reference Kennedy et al. (2006).
Q11AQ: Please provide the page range for the reference Soares et al. (2005).





























