Embed Size (px)
Citation preview

WAGENINGEN UNIVERSITY
LABORATORY OF ENTOMOLOGY
Overwintering strategy of Harmonia axyridis Pallas
(Coleoptera: Coccinellidae) may contribute to high
invasiveness
No.: 010.14 ..................................................................................
Name: Jeltje Stam .......................................................................
Study programme: MBI ..............................................................
Period: August 2010 ...................................................................
Thesis ENT-80439 ......................................................................
1st Examiner: Joop van Lenteren...............................................
1st Supervisor / 2nd Examiner: Lidwien Raak – Van den Berg
2nd Supervisor: Peter de Jong....................................................

2
Table of Contents
Preface 3
Abstract 4 Dutch Summary (Nederlandse Samenvatting) 5
Introduction 6 Material and Methods 8
Experiment 1. - Winter survival at central site 8 Experiment 2. - Winter survival at original sites 9
Survival at constant temperature 9
Mortality caused by handling 10 Experiment 3. - Ovary development during hibernation 10
Experiment 4. - Natural enemies 10 Experiment 5. - Colour variation 11
Sex ratio 11
Experiment 6. - Alighting in autumn and leaving the hibernation site in spring 11 Statistical Analysis 12
Results 13 Experiment 1. - Winter survival at central site 13
Experiment 2. - Winter survival at original sites 13 Survival at constant temperature 14
Mortality caused by handling 15
Experiment 3. - Ovary development during hibernation 15 Experiment 4. - Natural enemies 15
Experiment 5. - Colour variation 16 Sex ratio 16
Experiment 6. - Alighting in autumn and leaving the hibernation site in spring 16
Discussion 17 Winter survival 17
Influence of temperature on survival 17 Survival at constant temperature 18
Influence of place of overwintering on survival 18
Colour morphology 19 Sex ratio 20
Ovary development 20 Landing and dispersal 20
Natural enemies 21 Conclusion 22
Acknowledgements 23
References 24 Appendixes 28
Appendix 1. - Figures and tables 28 Appendix 2. - Detailed information on monitoring survival 53
Appendix 3. - Detailed information on the end of overwintering 54

3
Preface During the winter of 2009-2010 I have spent most of my time counting beetles. I counted the number of dead ones, the number of alive ones, the number of red beetles and the number of beetles with
two or four spots. I even counted the number of beetles with fungi or other parasites. Most of this
counting was done outside in the field or in a cold chamber, to not disturb the beetles in their winter hibernation. Although this was the most 'comfortable' for the beetles, for me this meant getting cold
fingers.
Al this counting and, after that, analyzing the data and writing down my conclusions, led to this thesis
report. It took me little more time longer than expected because the gathered data was better than initially hoped for. This gave me the opportunity however, to reconsider all that I have done,
experienced and learned while doing this thesis. Apart from knowing a great deal more about ladybirds now, I also learned more about designing and organizing experiments, asking others for
help, keeping a logbook and making friends during coffee breaks.
At least one thing that I have learned is that although it seems to be the easiest part of research,
counting the beetles actually appeared to be a difficult task!

4
Abstract Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) is a ladybird beetle, native in Asia. It has been released in the USA, after which it has spread and established in most of the country. After
introduction in Europe, it has established here in large parts of the continent as well. Some of its
characteristics may classify it as a useful species for biological control. These same characteristics also mean it has a higher risk as an invasive species.
Life history characteristics influence the degree of invasiveness of exotic species. Winter survival during unfavourable conditions is one of these characteristics. Low mortality during overwintering
results in large populations at the start of the new breeding season, in spring. The occurrence of
few natural enemies may also contribute to low mortality. To assess winter survival of Harmonia axyridis in the Netherlands, a semi-field experiment was
carried out. Overwintering H. axyridis adults were collected in late autumn at several sites in the Netherlands and subsequently introduced to a semi-field situation. Some collected beetles were
kept at a central site, resembling a general Dutch overwintering situation, and others at the original sites where the beetles were found. At these original sites beetles were kept at two orientations,
facing southwest and northeast. During the winter season the number of dead and surviving
beetles were counted eight and four times at the central and original sites respectively. Live beetles frozen at regular intervals were monitored for ovary development and checked for the prevalence
of natural enemies. Beetles that died during the winter were checked for fungus infection. Results indicate a high winter survival of H. axyridis in the Netherlands during the winter of 2009-
2010. Survival was on average 69.2% (ranging from 15.0% - 100.0%) at the central site and
89.1% at the original sites (ranging from 35.0% - 100.0%). The orientation at which H. axyridis overwinters influences winter survival: overwintering at a location oriented southwest resulted in
higher survival when compared to a north-eastern location. The orientation with the highest survival is not always the orientation were most beetles were found to overwinter. H. axyridis overwintering in leaf litter, a place where it normally is hardly found, showed high mortality. The results suggest that the overwintering strategy of H. axyridis is generally a good one. Colour and
gender did not influence the winter survival. Some parasites (mites Coccipopipus hippodamiae,
nematodes, fungi and the parasitoid Dinocompus cocinellae) were found in overwintering beetles, but their numbers were generally lower than found in other studies.
The high survival of H. axyridis during overwintering results in large beetle populations in spring, which might contribute to the invasiveness of this species.
Keywords: Coccinellidae, exotic species, hibernation, life history characteristics, natural enemies, winter survival

5
Dutch Summary (Nederlandse Samenvatting)
DE OVERWINTERINGSSTRATEGIE VAN HARMONIA AXYRIDIS PALLAS (COLEOPTERA: COCCINELLIDAE) KAN BIJDRAGEN AAN ZIJN HOGE INVASIVITEIT Het lieveheersbeestje Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) is afkomstig uit Azië. Het
is geïntroduceerd in de VS, waarna het zich heeft verspreid en gevestigd in het grootste deel van
Noord Amerika. Na introductie in Europa, heeft H. axyridis zich in grote delen van het continent gevestigd. Door bepaalde eigenschappen is het een bruikbare soort voor de biologische bestrijding, .
Door dezelfde eigenschappen vormt het lieveheersbeestje echter ook een groot risico als een invasieve soort.
De eigenschappen van een soort bepalen hoe invasief een soort is. Overleving tijdens ongunstige perioden, zoals de winter, is één van die eigenschappen. Een lage mortaliteit tijdens de overwintering
resulteert in grote populaties aan het begin van een nieuw paarseizoen in de lente. Het ontbreken
natuurlijke vijanden kan ook bijdragen aan lage sterfte. Om de overleving tijdens de winter te bepalen van H. axyridis in Nederland, werd een semi-veld
experiment opgezet. Overwinterende volwassen lieveheersbeestjes werden in de late herfst op verschillende plekken in Nederland verzameld en geplaatst in overwinteringskooitjes. Een deel van de
populatie werd naar een centrale plaats gebracht, die de overwinteringssituatie in Nederland
nabootste. Het ander deel van de populatie continueerde de overwintering op de locatie waar ze werden gevonden. Daar werden de kevers in twee verschillende windrichtingen in of op de gebouwen
geplaatst, op het Zuidwesten en het Noordoosten. Gedurende de winter werd op zowel de centrale als de oorspronkelijke locaties het aantal dode en levende kevers geteld, elke twee of elke vier week
respectievelijk. Op de tijdstippen waarop de kevers geteld werden, werd ook een monster levende kevers ingevroren. Van dit monster werd de ontwikkeling van de ovariën gecontroleerd, en de kevers
werden gecontroleerd op het voorkomen van natuurlijke vijanden. Kevers die dood gingen tijdens de
winter werden op schimmelinfecties gecontroleerd. Tijdens de winter van 2009-2010 in Nederland laat H. axyridis een grote overleving zien. De
overleving was gemiddeld 69,2% (15,0% tot 100,0%) op de centrale locatie en 89,1% op de oorspronkelijke locatie (35,0% tot 100,0%). De windrichting waarin H. axyridis overwinterde,
beïnvloedde de mate van overleving: de kevers aan de Zuidwest zijde van het gebouw hadden een
hogere overleving dan die aan de Noordoostelijke zijde. De windrichting waar de overleving het hoogst was, is niet altijd de windrichting waar de meeste overwinterende kevers werden gevonden
tijdens het verzamelen. H. axyridis die in bladafval overwinterde, een plek waar de kever normaal nauwelijks wordt aangetroffen, had een lage overleving. De resultaten suggereren dat de
overwinteringsstrategie van H. axyridis meestal een goede is. De kleur en geslacht beïnvloedden de
mate van overleving niet. In de overwinterende kevers werden enkele parasieten (mijten van de soort Coccipopipus hippodamiae, nematoden, schimmels en de sluipwesp Dinocompus coccinellae)
gevonden, maar in lagere aantallen dan in andere studies werd gevonden. De hoge overleving van H. axyridis tijdens de overwintering resulteert in grote populaties kevers in het
voorjaar, wat kan bijdragen aan de invasiviteit van deze soort.
Trefwoorden: Coccinellidae, exotische soort, levenseigenschappen, natuurlijke vijanden, overleving,
overwintering

6
Introduction The Multicoloured Asian ladybird beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae), is indigenous in Asia. After numerous releases in the USA since 1916, the first established population
was recorded in 1988. It has spread through most of the USA, as well as through Canada (Koch et al.,
2004). After introduction in Europe in 1982, it took until 2000-2001 until it established in Europe (Lombaert et al., 2007). Since then, it has spread rapidly over the continent. Currently, it is abundantly
present in a large part of Europe, from southern France up to Denmark in the North, and reaches as far as the United Kingdom in the West and the Czech Republic in the East of Europe. In autumn 2002
it was first recorded in the field in the Netherlands, and from 2004 their numbers increased greatly
(Brown et al., 2008).
Until recently, H. axyridis has been used in classical and augmentative biological control, because of its ability to consume large amounts of numerous aphid species (Koch, 2003; Berkvens et al., 2009). H. axyridis has several characteristics that make it useful in biological control, but at the same time these characteristics qualify it as a risk to become an invasive species (Berkvens et al., 2009; Koch and
Galvan, 2008; Van Lenteren et al., 2008). It is multivoltine, has a high dispersal capacity, and a high
cold tolerance. It is polyphagous, consuming aphids as well as pollen, nectar and larvae of several coccinellid species, including its own. It thus has a low host specificity (Berkvens et al., 2009).
Therefore, H. axyridis scored the second highest risk among several commercially available biological control agents on the risk-index developed by Van Lenteren et al. (2003). Today, it is hardly used as a
biological control agent in parts of Europe, because of strict regulations (Brown et al., 2008; Loomans,
pers. comm.).
To be able to settle in a new environment, such as Europe, H. axyridis must be capable of surviving unfavourable conditions, for example the winter season. During winter, H. axyridis hibernates in
clusters (Berkvens et al., 2009). In its natural environment, it searches a suitable hibernation place by hypsotactic behaviour; it searches for conspicuous objects on the horizon by visual cues (Obata et al.,
1986; Nalepa et al., 2005; Koch, 2003). On warm, calm days in autumn the beetles fly towards the
north-east and land on trees and rocks or, in case of a more urban environment as in the Netherlands, on buildings. They may aggregate there in large groups, possibly regulated by pheromones or faeces
of conspecifics (Koch, 2003; Tanigishi, 1976). When the ladybird beetles land and overwinter in buildings, they may become a nuisance to humans,
as they may reflex bleed or even bite on disturbance. Some cases of allergic reactions have also been
reported (Koch, 2003). Next to that, the beetles may cause problems in fruit production, as they may feed on various fruits and can for example ruin wine production when they become crushed between
the grapes (Koch, 2003).
During hibernation, ladybird beetles may go into diapause. Diapause may be defined as a
physiological state in which metabolism is lowered and development is inhibited. It is initiated by environmental cues before the onset of unfavourable conditions, and will end only after certain cues
(environmental or physiological), even when the conditions may be favourable for development again (Hodek et al., 1996; Tauber et al., 1986). In contrast, quiescence is a state in which development is
suppressed in reaction to unfavourable conditions, and development immediately resumes when conditions become favourable again (Hodek et al., 1996; Tauber et al., 1986). Diapause increases the
chances of survival under unfavourable conditions, such as the winter period, by inhibiting costly
processes as development (Hodek et al., 1996). Ovary regression may be viewed as an indicator of diapause (Iperti and Bèrtand et al., 2001; Hodek et
al., 1996; Sakurai et al., 1986). As diapause initiates inhibited development before the onset of unfavourable conditions, and development may start even after favourable conditions occur, it is
expected that the beetles will show ovary regression during the whole winter period if H. axyridis has
indeed a true diapause. Previous results showed however that H. axyridis lays eggs quickly after transfer to long-day conditions, even in mid-winter (Raak-van den Berg, unpublished results).

7
In the winter period, H. axyridis may encounter low temperatures during their hibernation. The cold
tolerance of overwintering H. axyridis is high, and it tolerates temperatures below 0 °C (Berkvens et al., 2009). The supercooling point is the temperature at which the beetles freeze, and is a good
indication of its cold hardiness. This point was found to be at least -13.2 °C for H. axyridis (Berkvens et al., 2009; Koch et al., 2004; Watanabe, 2002). The supercooling point decreases during the winter
season, which may increase the survival of hibernating H. axyridis adults (Koch et al., 2004;
Watanabe, 2002). Ladybird beetles may die however, at a temperature above the supercooling point, due to chill injury
(Koch et al., 2004; Watanabe, 2002). H. axyridis was found to survive better at lower temperatures than higher temperatures in a controlled environment (Watanabe, 2002; Labrie et al., 2008). A
depletion of energy reserves, or desiccation, rather than chill injury could explain this (Watanabe, 2002).
To what extend the supercooling point determines the overwintering success, and hence fitness, of H. axyridis, remains unclear. In the North of the United States the supercooling point did not adequately explain the distribution of H. axyridis (Koch et al., 2004), suggesting that in this area, cold tolerance is
not constraining the distribution of this beetle. H. axyridis had a high winter survival when overwintering outside in South East France (Iperti and
Bèrtand, 2001), and northern Italy (Bazzocchi et al., 2004), while only slightly more than half the
population survived in Quebec, Canada, when they overwintered inside houses (Labrie et al., 2008). The native species Coleomegilla maculata failed to survive inside, but did outside in leaf litter. In leaf
litter outside, H. axyridis failed to survive at all (Labrie et al., 2008). Native coccinellidae in Europe, such as Propylea quatuordecimpunctata, Adalia bipunctata and Coccinella septempunctata showed a
relative low survival (Honek, 1997; Brakefield, 1985; Zhou et al., 1995; Bazzochi et al., 2004) when compared to the survival of H. axyridis in Europe. Apparently, H. axyridis is well able to survive in its
relative new environment, compared to native species.
Next to cold tolerance, biotic mortality factors such as parasitic infections may influence the mortality
during winter hibernation as well. Potential parasites are among others fungi, mites, nematodes, and/or parasitoids (Riddick et al., 2009).
Previously, several species of fungi (Lecanicillium, Isarina fariosa and Beauveria bassia) were found on
H. axyridis which caused some mortality of overwintering beetles (Steenberg and Harding, 2009), as well as the ectoparasitic fungus Hesperomyces virescens did (Riddick and Cottrell, 2010; Riddick,
2010). Also the mite Coccipopipus hippodamiae is found on H. axyridis and it is shown that it can control H. axyridis populations (Rhule et al., 2010). Shapiro-Ilan and Cottrell (2005) found that
although exotic species were less susceptible to nematodes than native species were, nematodes may
influence survival of H. axyridis. The Hymenopteran parasitoid Dinocampus coccinellae is a specialist in Coccinellids, including H. axyridis, though its impact on natural field populations is not yet known
(Riddick et al., 2009; Hodek, 1973).
In this study, the survival of H. axyridis was assessed in two semi-field experiments during the winter of 2009-2010 to elucidate the overwintering success in the Netherlands and factors that influence this.
This includes the site and orientation of overwintering and the prevalence of parasitic infections (fungi,
mites, nematodes and parasitoids) in overwintering Dutch H. axyridis populations. Next to that, the ovary development was studied as an indication of the degree of diapause. The influence of the
overwintering characteristics and prevalence of natural enemies which occur during winter on the
invasiveness of H. axyridis is discussed.

8
Materials and Methods Experiment 1. - Winter survival at central site Collecting beetles Adult H. axyridis have been collected from 13 November – 27 November 2009 at their site of
hibernation at 5 different sites where large aggregations had been formed. Aggregations were defined as a cluster of beetles ranging from 5 – 1000 individuals. The sites were distributed over the
Netherlands and consisted of either private houses situated in the woods (Kootwijk, Loenen and Zundert), or church towers (Alkmaar and Houten). The beetles were found mostly at the exterior of
the buildings: behind wooden shutters and hinges, in window or door frames, in crevices between
brick walls or inside in the corners between walls and ceiling. During transport to Wageningen, where the central site was located, beetles were kept in plastic boxes containing wrinkled filter paper, and
covered with a gauze lid (ambient temperature). Until use, the beetles were kept in an unheated outdoor shed. Temperature and relative humidity in the shed were similar to outside conditions, but
sheltered from rain.
Experimental set-up To monitor overwintering, collected beetles were transferred to outdoor cages on 4 December 2009. The outdoor cages consisted of wooden shutters (60x105cm, 3cm deep; figure 1), resembling the
situation in which many beetles were found to overwinter naturally in the Netherlands. Overwintering was monitored at the experimental farm 'Droevendaal' for organic farming of Wageningen University
(central Netherlands; the 'central site').
Six identical shutters were located at a south-facing wall: shutters 1-3 at the left, and 4-6 at the right of a central door. At the inside of the building, a heating was located next to the wall with shutters 1-
3, and a cold chamber was located behind shutters 4-6 (this was not arranged intentionally for the experiment, but happened to be the situation at the location of the cages). Shutters at the left were a
little more shaded in the late afternoon than shutters at the right, but were similar in all other aspects. The beetles were placed in gauze bags (10x50cm, gauze diameter 1x1mm) hanging vertically,
because previous studies had shown that the beetles have the tendency to stay in top corners during
winter hibernation (Hodek, 1996; Raak-van den Berg, personal observation; Tanigishi, 1976). A metal spiralling frame (3x6x40cm) allowed free movement within the bags. Beetles collected at the 5
different sites where placed in separate bags. Bags were placed at one level in random order concerning the sample sites behind each shutter (figure 1).
Each bag contained 100 beetles (50 in case of location Loenen) which is equivalent to 5 cm2 available
for each beetle, according to the methods of Labrie et al. (2005). A series of bags to monitor ovary development and parasitic infections (see below) were placed in between these bags.
Monitoring survival A time-series was made, with shutters 1-6 representing 6 replicates. Every 2 weeks, from 19
December 2009 until 18 March 2010, the number of alive and dead beetles was counted at 4 °C (See appendix 2 for more details on the method used). Live beetles, defined as walking around, moving
body parts or reflex bleeding, were put back into the bags to continue hibernation, and dead ones were collected in Eppendorf tubes to determine colour morph, gender, and fungus infections.
Two dataloggers, behind shutter 2 and 5, recorded temperature every 30 minutes during the whole experimental period.
End of overwintering Towards the end of winter, experiments were stopped to avoid mortality caused by increased post-
hibernation activity during higher temperatures. This moment of experiment termination was determined by observing the movement of beetles in
opened bags, which were placed in half and full shade. Also, beetle activity in the field was monitored,
and the observation of a large number of individuals moving around during a warm period was an indication that they had left their overwintering sites. (See further details on the end of overwintering
in appendix 3).

9
Thus, on March 18, 2010 all remaining beetles in the bags were counted and colour morph was
determined. Afterwards beetles were sexed. Beetles were scored as either 'alive' (walking around
normally), or 'potentially dead' (not or hardly moving). The potentially dead beetles were kept at room temperature overnight, after which normally walking beetles were scored as 'alive'. Those that barely
moved or did not move at all, would not make it to the breeding area in a field situation and were thus considered dead in further analysis (Labrie et al., 2008). Dead beetles were scored as such, but
were no longer used to check for fungus infections due to a lack of time, as was done with all other
beetles dying during the overwintering experiments (see below).
Experiment 2. - Winter survival at original sites
Experimental set-up Winter survival was also monitored at the 5 sites where the beetles were initially collected for
experiment 1.
At each site, the collected beetles were placed in gauze bags with a spiralling frame at two orientations: the southwest-facing orientation where the majority of beetles was found aggregating,
and the northeast-facing orientation where few or no beetles were found. The bags were either placed at the inside of the outer stone walls of the tower (Alkmaar, Houten), or they were placed behind
wooden shutters at the outer stone wall of the building (Kootwijk, Loenen, Zundert). At each site, four
bags with each 100 beetles (50 in case of Loenen) were placed at the each of the orientations. The sites Kootwijk, Loenen and Zundert were located in the woods. The site Kootwijk was directly
surrounded by trees, except for a small open space at the northeast and southwest. At Loenen there was only an open space in the trees to the northwest, while the trees at the site Zundert were not so
dense (figure 2). At the site Kootwijk overwintering at four additional orientations could be monitored: First, at the
north of the building at the inside of a wooden door of an unheated adjacent attached barn; second
and third, beetles from this northern and the south-western orientation were brought to an unheated attic; and finally, beetles were placed in leaf litter under a hedge near the house. The latter was
repeated with beetles originating from the same location in leaf litter under a hedge at the experimental farm, Wageningen.
A datalogger recording temperature was placed among the bags at the orientation were most beetles
were found at every site.
Monitoring survival As in experiment 1, a time-series was made, with the four bags representing 4 replicates. All beetles
were counted four times during the winter season (early January – end of March, 2010), as described
above (experiment 1). Dead beetles were collected in Eppendorf tubes to determine colour and gender and checked for fungal infections (see below). At the end of the overwintering season all bags
containing the beetles were transported to Wageningen, and the beetles were scored as 'alive' or 'dead' after 24 hours, as described above.
The survival was monitored at the following sites: (figure 3, table 1) – Alkmaar (north-western Netherlands)
– Houten (central Netherlands)
– Kootwijk (central-eastern Netherlands) – Loenen (central-eastern Netherlands)
– Zundert (southern Netherlands)
Survival at constant temperature Hatherly et al. (2005) argued that an LTime50 (the time required to kill 50% of the population) at 5°C
corresponded best of several factors with survival in the field. Thus, an experiment was set up in a climate chamber kept at 5 °C, (0:24 L:D), representing another shutter as at the experimental farm.
The number, contents and handling of bags was similar as at the experimental farm. Bags with

10
beetles from each of the 5 sites were placed randomly and at one level next to each other. Survival
was monitored identical to the winter survival experiment at the experimental farm.
Because of the constant conditions in the climate room, and thus the absence of spring conditions, the survival could be determined over a prolonged period. Therefore the LTime50 could be determined,
which was not always possible for field survival when less than 50% of the population died before the end of the winter. Survival was continuously monitored every two weeks until August 29, 2010.
Mortality caused by handling
Because the site Loenen did not provide sufficient beetles to fill all bags for the ovary development experiment, surplus bags with beetles from another site (Deelen, not far from the site Loenen)
replaced these gaps behind 3 random shutters (2, 6 and constant temperature). These bags containing beetles were taken to 4 °C, as was done with the other bags at the experimental farm to
count the beetles of experiment 1. The surplus bags were however not opened, and the beetles thus
not disturbed by counting (removing beetles from the bags and replacing them into the bags after counting). At the end of the winter season, the mortality of these beetles was determined to assess
whether extra mortality, in comparison the regularly counted beetles of experiment 1, was caused by disturbance during counting and handling the beetles.
Experiment 3. – Ovary development during hibernation
Beetles were collected from the five original sites as well to monitor ovary development during winter. About 250 beetles per bag from every site were hung randomly behind the shutters, in between those
for monitoring survival (experiment 1). Similarly, 250 beetles from every location (except the site Loenen due to a shortage of beetles) were placed in between the bags in the climate chamber with
constant temperature (5 °C). Every two weeks, about 25 beetles from each of these bags were
harvested in Eppendorf tubes per 5 individuals. Subsequently, they were frozen at -25 °C until further use. The frozen beetles were later dissected under a dissecting microscope. Beetles were harvested
from 4 December 2009 (at the start of the overwintering experiments) until 12 April 2010 (the last harvest from the experiment at constant temperature).
The ovary states were found to be defined in several ways in literature (table 2). Therefore, a new
definition of the ovary states were made, combining definitions found in literature. In state 1, no eggs or follicles are present in the ovarioles, as no or hardly any constrictions of the ovarioles and no lighter
coloured spots (indicating follicles) are visible. At state 2, the first follicles are developing, and become clear by constrictions of the ovariole around the still transparent (whitish) follicles. From state 3
onwards, the follicles become vitellinized, colouring yellow, and the number of follicles per ovariole
increases from 1-2 in state 3, 2 in state 4, and 3 mature eggs in state 5. The follicles increase in size with increasing development state.
The size of a few ovarioles of some beetles was measured as a reference, using digital pictures taken with a digital camera (Euromox CMEX DC.1300) mounted on the microscope.
Experiment 4. – Natural enemies
4.1. Fungi Dead beetles collected during the winter survival experiments at the central and original sites were
checked for fungal infection. Dead beetles were first dipped in 70% ethanol to avoid colonization by saprophytic fungi. Next,
beetles were placed in Petri dishes lined with moist filter paper. These were then placed at 25 °C to
allow fungal growth and beetles were checked for fungi after 10 days. Beetles with fungi were stored at 5 ºC in sealed petridishes until further identification.
The fungi were identified at the Fungal Biodiversity Centre (CBS, Utrecht, the Netherlands). They were first identified using morphological characters, after which the potential entomopathogenic fungi were
inoculated on malt extract agar and oatmeal agar to obtain pure cultures. Detailed identification of these pure cultures under a light microscope was conducted two weeks later. Fungi that could not be

11
identified morphologically (mainly the Aspergillus ochaceus-group) were molecularly identified by
sequencing.
The prevalence of the ectoparasitic fungus Hesperomyces virescens on dead beetles collected from the winter survival experiments was scored as well.
4.2. Mites, nematodes and parasitic wasp Dinocampus coccinellae
During dissection of beetles to monitor ovary development, the occurrence of parasites was scored as
well. The beetles were frozen alive, so any parasites present on or in the live beetles were still present at dissection.
Individuals of the parasitic mite Coccipolipus hippodamiae can be found under the elytra of living Coccinellidae (Webberly et al., 2004, Rhule et al., 2010). The mites are sessile as adults, and attach to
the elytra of the beetle. Therefore the undersides of the elytra were checked under a dissecting microscope. The number of mites under each elytrum and the life stage (larvae or adults) was scored.
Any nematodes present were visible after opening the abdomen of the beetles. The nematodes drifted
out into the dissection buffer [PBS] of nematode infected beetles and were thus clearly visible under the dissecting microscope.
The prevalence of larval stages of the parasitic wasp Dinocampus coccinellae occurring inside the abdomen or thoracic region was scored during or after the removal of the ovarioles.
Experiment 5. - Colour variation
In addition to the beetles collected at the five sites mentioned above, beetles were collected at 15 other sites in the Netherlands, from which the number and colour morph was determined. The
following colour morphs were recognized: f. succinea (red with zero – nineteen dark spots), f. spectabilis and f. conspicua, (black with four or two red spots respectively), as well as the more rare
morphologies f. equicolor (red with black lower part of the elytra) and f. axyridis (red with twelve black
spots).
Sex ratio
All dead and life beetles from the 5 original sites that overwintered at the central site were sexed
under a dissecting microscope.
Experiment 6. – Alighting in autumn and leaving the hibernation site in spring
Alighting of beetles Beetles were observed on a warm, sunny day in autumn (23rd of October 2009) when landing on the coloured sunscreens of the building where the laboratory of Entomology is housed (Radix building).
The screens were coloured in three shades of green and one shade of brown. The dark background in gaps between the screens formed a contrast with the bright coloured screens (figure 4).
Every three minutes, the number of beetles on the screens was scored during half an hour in the afternoon. The screen colour, the number of beetles landing on each screen and the position on the
screen (within 10 cm of the edge or in the middle) was scored.
The number of beetles landing on each colour per surface unit was calculated.
Beetles flying away At the end of the overwintering season (April 14th, 2010) beetles were observed leaving their
hibernation site. Two gauze bags with a metal spiralling frame were filled with about 200 beetles each
and placed behind two shutters at the experimental farm. The door of one of the shutters remained open, while the other remained closed during observation (only to be opened quickly when checking
the beetles). After opening the bags the number of beetles escaping was scored every 3 minutes, as well as the
weather condition (sun or overcast). The experiment was carried out from 15:27 until 16:37, at which time the sun no longer reached the shutters.

12
Statistical Analysis
Overwintering survival was analysed using the Cox proportional hazards model (Cox, 1972). This
model analyses effects of covariates on a hazard rate, which is the conditional probability of an event to occur per unit of time. The condition is that the event has not occurred yet. In this study, the event
is the death of a beetle. Hence, at every point in time, there is a certain chance for each beetle still alive until that moment, to die. The model assumes a certain baseline mortality rate (the baseline
hazard), multiplied by a factor (the regression coefficient) that accounts for the effect of the
covariates; in this case, for example the site or shutter at which the beetles overwintered are different covariates.
The following model is used:
q
i ii zthZth100 )exp()(),(
where h0 (t, Z) and h0(t) are the hazard rate and baseline hazard respectively, at time t, with covariate values Z=(z1, …, zq)
T. βi (i = 1,…, q) are the regression coefficients that give the relative contribution
of the covariates zi (Cox, 1972). The hazard ratio, given by the expression exp (Σ βi zi) gives the effect of a covariate; when it is lower
than 1, the survival is reduced, while with a hazard ratio higher than one, the survival is increased. The model also takes censored data into account, the cases in which the dying of a beetle could not
be recorded. Censored data occurs when a beetle is either lost from the study (without dying) before
the end of the study, or by dying due to another cause than winter mortality (e.g. it has become crushed accidentally), or it has not died yet upon the end of the study. A „censored‟ beetle at a certain
point in time generates valuable information, because although the exact time of dying is unknown, it is known that the beetle has lived until that point in time, and could die after that moment.
For the winter survival analysis on sites and orientations, the number of covariates is q=6, namely βi
(i=1,…,4) codes for sites (Alkmaar, Houten, Kootwijk, Loenen) compared to the baseline site Zundert. β5 and β6 are left and right shutters compared to the baseline for the experiment at constant
temperature. If the lines of the covariates in the survival curve cross, it means that the survival curves of the
covariates could not be obtained by multiplying the baseline hazard with a certain factor βi. In other
words, the covariates are not proportional to each other. In that case, the Cox model could not be used, and the Log-rank test was used instead to distinguish between significantly different mortality
patterns. When only one of several covariates (e.g. orientation and site of overwintering) appeared to be non-
proportional, the model can be stratified on this covariate, which means that although the covariate is not proportional, the model does take the effect of the covariate on the survival into account.
It should be noted that figures based on the model do not display the actual survival curve but an
estimate of the survival. The log-rank test does give the actual survival, which was therefore used to create the figures in this report for a better comprehension.
The gender and colour ratios were analysed with an r x k contingency table (Pearsons Chi-Square),
the alighting behaviour was analysed using the Kruskal-Wallis test for the different screen colours and
the Mann-Whitney U-test (non-parametric rank tests) for the position on the screens. The software package SPSS was used for analysis (version 15.0.1.1, July 2007).

13
Results Experiment 1. - Winter survival at central site In total, 2690 beetles have overwintered at the central site (Table 3a). The average survival was
69.2% (ranging from 15% - 100%). Survival remained above 90% until 1st March, i.e. 88 days after
the beginning of the experiment (4 December 2009), after which survival decreased quickly until a minimum of 15% at March 18, 90 days after the first day of counting (figure 5a, b). Differences in
colour morph did not significantly influence winter survival (Log–rank test, all pairwise comparisons have Χ2
1 < 2.3 and P >0.14). Likewise, the gender did not influence winter survival (Log–rank test, Χ21
<< 0.001, P =0.988).
As there were no statistical differences with regard to morph colour and sex concerning winter survival, these factors could be ignored in further analysis.
Thereafter, the data were analysed by means of Cox regression in which the sites and locations were categorical covariates. As stated before, the winter survival analysis was performed with six covariates
for sites and locations, namely covariates 1 to 4 codes for sites compared to the baseline site Zundert and covariates five and six code for left and right shutters respectively, compared to the baseline in
the experiment at constant temperature.
Survival differed significantly between sites (Cox regression Wald test = 664.53, df=6, P < 0.001). The beetles originating from the sites in the woods (Loenen, Zundert and Kootwijk) had a longer
mean survival time than the beetles originating from the church towers in Alkmaar and Houten (table 4a). In addition, the shutters behind which the beetles overwintered influenced survival, with the
shutters at the left having a shorter mean survival time than beetles overwintering behind shutters at
the right (table 4b). The effect exp(β) for the left hand shutter compared to the baseline (the experiment at constant 5 °C) was 2.03, while the effect of the right hand shutter compared to the
baseline was 0.10. This means that the baseline hazard is multiplied with a larger number (a larger difference) to obtain the survival curve for the left hand shutter, than for the survival curve for the
right hand shutter. A difference in temperature was found between the shutters. Figure 6 shows the daily average
temperatures at the central site. Temperature data indicate that the shutters at the left were generally
warmer than the shutters at the right. When temperatures were summed over the winter season (figure 7) the shutters at the left-hand side had a higher cumulative temperature at the end of the
winter season than the right-hand shutters. The experiment in the laboratory at constant 5 °C had an intermediate cumulative temperature.
Experiment 2. - Winter survival at original sites
In total, 5304 beetles were monitored in an experimental setting at the Dutch sites where aggregations of hibernating beetles were found (Table 3b). The average survival over all sites and
orientations, excluding leaf litter, was 89.1% (ranging from 35.0% - 100%). Because survivor curves
of the different sites crossed (the curves were not proportional), a stratified analysis for site is performed to reveal the effect of orientation.
The beetles overwintering in leaf litter at Kootwijk had low average survival (7.13%, ranging from 1.1% - 11.5%), with relatively high mortality from the start of the winter onwards. This was
significantly lower than survival at the other places at Kootwijk where overwintering was monitored (Log–rank test, Χ2
5 > 464.59, P < 0.001). The beetles overwintering in leaf litter at the central site had
an even lower median survival time (Log–rank test, Χ 21 = 87.74, N = 780, P < 0.001).
The survival at the different sites was compared including only the south- and north-facing
orientations of all sites. The survival differed significantly between the sites (survival at south- and north were combined; Log–rank test, overall Χ2
4 > 5.48, P < 0.019), except for the sites Alkmaar –
Loenen (Log–rank test, pairwise Χ 21= 3.528, P = 0.06), and Kootwijk - Loenen (Log–rank test,
pairwise Χ 21= 2.094, P = 0.148).
The sites were not proportional (the survival curves crossed), so a stratified Cox was necessary to
take into account the non proportional effect of the different sites on the survival, when analyzing the

14
effect of orientation on survival. While the orientation (southwest or northeast) was entered as
covariate, the sites at which the beetles had overwintered were entered as strata, to account for the
differences between the sites. The survival differed significantly between the orientations facing the southwest or northeast when all
sites were included, with the beetles overwintering at the southwest having the highest survival (Cox regression Wald test = 48.27, df = 1, P < 0.001; table 4d). The effect exp(β) was -0.68 for southwest
with respect to baseline northeast, which means that the survival curve of beetles overwintering at the
northeast is multiplied with this number to obtain the survival curve of beetles overwintering at the northeast.
Beetles overwintering at the sites located in the church towers (Alkmaar, Houten) had a higher mean survival time than the sites located in the woods (Kootwijk, Loenen, Zundert; Cox regression model,
stratified on site, Wald test = 48.27, df = 1, P < 0.001; table 4c).
Winter survival: zooming in on Kootwijk Although at Kootwijk the overwintering at the southwest did not differ significantly from the northeast, some of the additionally tested orientations (table 4d) at this site did differ.
When collecting the beetles, many were found at the north-facing side unexpectedly. The beetles overwintering here survived significantly better than those at the northeast side. At the northeast side
no beetles were found originally, but they were placed here experimentally (Log-rank test, pairwise
Χ21 = 8.312, P = 0.004). Both sides where beetles were found (southwest and north) did not differ
significantly in survival from each other (P=0.217), although the survival did differ when beetles from
both sides were brought to an unheated attic. Beetles from the south-facing side survived better at the attic than those from the north-facing side did at the attic (Log-rank test, Χ 2
1=6.189, P = 0,013).
At the three sides where beetles overwintered inside (at the inner wall of an attached barn and twice at the unheated attic), the survival was significantly higher than the sides where beetles overwintered
behind shutters (south-west and north-east) (Cox regression Wald test = 1713.82, df = 2, P < 0.001).
Church towers versus woods When overwintering was compared between the site types ('woods' or 'church tower'), the beetles overwintering at the church towers had a significantly higher mean survival time than the beetles
overwintering at the woods (Cox regression Wald test = 93.23, df = 1, P < 0.001).
Figure 9 shows the daily average temperature at the 5 original sites. All sites had comparable temperatures, although Kootwijk had slightly lower, and Alkmaar had slightly higher temperatures.
The cumulative temperatures during the winter season showed that Alkmaar has the highest cumulative temperature at the end of the winter season, followed by Loenen, Kootwijk, Zundert and
Houten respectively (figure 10).
Original sites versus central site The beetles had a significantly higher survival time when overwintering at the original sites than when overwintering at the central site (Log–rank test, stratified, Χ2
1 = 753.55, P < 0.001).
Survival at constant temperature
Survival under controlled circumstances (constant 5 °C, no light) after 15 weeks was high (mean survival 90.4%, ranging from 85.0% – 96.1%). Survival did not significantly differ from the beetles
overwintering behind the shutters at the right hand side of the experimental farm (figure 5c), but it was significantly higher than the survival of the beetles overwintering behind the shutters at the left
(P <0.001).
The monitoring of the survival under controlled circumstances was continued until 17 August 2010 (242 days after the first time counting). Figure 8 shows that the survival declined quickly after the
moment that the beetles in field started to disperse (90 days after first day of counting), until a survival of 2.2% (ranging from 0.0 – 4.0%). The LTime50, the median number of days when a survival
of 50% was reached, was 171 days.

15
Mortality caused by handling
The survival of beetles in the surplus bags that were not counted had a slightly higher mean survival (75.3%) than the average survival in the bags that were counted (71.8%), although the variation of
the survival in the surplus bags is larger. This suggests that handling did not systemically influence survival.
Experiment 3. – Ovary development during hibernation
In total 448 ladybirds were dissected, of which 276 were female. Only one beetle, harvested at 29 March 2010, showed a clear ovary state 2 (table 2, table 5), with first follicles in the ovarioles partly
developed (constrictions around the follicles are visible), although the follicles have not became vitellinezed yet (no yellow colouring; see figure 11). A few other beetles from this date showed some
constriction of the ovarioles, but no follicle development (lighter coloured spots at the base of the
ovariole) could be seen. Most other ovarioles showed no follicle development at all (table 5). As hardly any follicle development could be seen, only the sizes of the whole ovariole could be
measured, which was on average 790.12 ± 160.81 μm.
Experiment 4. – Natural enemies 4.1 Fungi Of the 1818 beetles that were checked for fungal growth, 494 had some sort of fungus infection. After the first rough selection at the CBS, the fungal mycelium from 28 of these beetles was isolated to
obtain a pure culture for further morphological determination. After sequencing of the fungi that could not be identified morphologically, four species of fungi from
the Aspergillus ochraceus-group (A. ochraceous, A. persii, A. sclerotiorum and A. westerdijkiae) were
found in 11 individual beetles, and 3 cases of fungi in this group could not be further identified. Additionally, one case of Myriodontium spp., one case of Chromelosporium fulvum, one case of
Paeciliomyces lilianus and two cases of Lecanicillium spp. were found. Of the 19 cases (1.1%) of beetles infected with a fungus that were thought to be pathogenic (table
6), none of them appeared to be truly entomopathogenic after discussion.
Among the dead beetles at least 5 (0.28%) beetles were found with an infection of Hesperomyces virescens, an ectoparasitic fungus. The fungi originated from beetles overwintering at the sites Alkmaar (7 cases), Houten (5 cases),
Kootwijk (4 cases), and Loenen (3 cases). None of the beetles originating from the location Zundert or
from the leaf litter at Kootwijk of the experimental farm carried (entomopathogenic) fungi. They were about evenly distributed over the place of overwintering (the central or the original sites), and they
were found about equally frequent at the south or north-facing orientations at the original sites. 4.2 Mites, nematodes and parasitic wasp Dinocampus coccinellae In total, 448 beetles were dissected and checked for ovary development and parasites (Table 7).
Three beetles (0.7%, 2 females and 1 male) carried the mite C. hippodamiae (Figure 12). In two
cases both larvae and adults were found under both elytra, in the other case only one single adult was found under one of the elytra.
In total 14 (3.13%) beetles with nematodes inside the beetles were found (8 beetles were female, 6 male). In most cases, the abdomen was filled with nematodes that drifted out as soon as the beetle
was opened (figure 13). Unfortunately, the nematode species could not be identified in the scope of
this study. Only one larvae of the parasitic wasp Dinocampus coccinellae was found in the abdomen of one
female beetle (figure 14).

16
Experiment 5. - Colour variation
In total 21545 beetles were collected at all sites. From these, 76.88% were non-melanic (f. succinea),
18.41% f. spectabilis and 4.55% f. conspicua. Only 2 individuals of f. equicolour were found (0.01%), and no f. axyridis (table 8).
Colour ratio differed significantly between all the 20 different sites (Χ238 = 766.17, N = 21510, P <
0,001), as well as between the 5 sites used for the overwintering experiments (Χ210 = 614.59, N =
17895, P < 0,001). Even when the two melanic colour morphs, f. spectabilis and f. conspicua, were
pooled, the sites differed significantly from each other (Χ25 = 312.10, N = 17895, P < 0,001).
However, there was no clear correlation between colour ratio (red versus melanic) and the location
(coordinates) in the Netherlands, the R2 of linear regression was below 0.2 in all cases (data not shown).
Moreover, no clear trend in geographical distribution in the Netherlands could be found when grouping sites into 7 regions (figure 15); the non-melanic versus melanic ratio was very similar in for
example the northern region (25 km from the coast) and the southern region (about 150 km from the
coast), although they differed significantly from the region in central Netherlands. At some sites, H. axyridis was collected in autumn 2008 as well. Only at the site Loenen the number
of beetles was about twice as low as previous year, at the other sites (Kootwijk, Deelen) the numbers of beetles was equal to the numbers found the previous year. The ratio between non-melanic and
melanic beetles was however almost equal in the two years at the sites Loenen (85.2:17.5) and
Deelen (around 84:16). At Kootwijk however, slightly more melanics were found in 2009 than in 2008 (78.0:22 and 84.1:15.9 respectively).
Sex ratio Of the 2658 beetles tested at the central site, 1163 (43.75%) were male and 1495 (56.25%) were
female. A significant difference in sex ratio was found between the different sites (Χ24 = 21.2, P <
0.001). More females than males were found at the sites located in the woods (41.78 : 58.22 M:F), while in the church towers the sex ratio was close to equal (48.35 : 51.65 M:F). This difference
between sites in the woods and church towers was significant (Χ21= 11.21, P < 0.001).
Experiment 6. - Alighting in autumn and leaving the hibernation site in spring Alighting
No significant difference in the number of beetles per unit of surface landing on the different coloured screens was found (Kruskal-Wallis test; H(3) = 3.39, p = 0.335). In contrast, significantly more
beetles landed on the edge of the screens than on the middle (Mann-Whitney test, U = 605, z = -
5.83, N = 110, P < 0.001).
Flying away Almost all beetles flew out of the opened bags in half an hour. During observations sunshine
alternated quickly with overcast conditions. A significant difference existed between the number of beetles flying away with sunshine or shade (Χ2 = 4.91, N = 527, P < 0.028). About 70% of the
beetles flew when the sun was shining, the remaining 28.1% when there was shade.
There was a difference in activity of the beetles between the opened and closed shutter. The beetles were more active at the opened shutter (they flew away immediately at the moment they had left the
bag) than at the closed one, where they stayed longer on the bag or wall (even when the shutter was quickly opened to count the number of beetles). They also walked upwards more quickly when in the
opened shutter than in the closed one. The beetles were generally more active in reaction to direct
sunshine, compared with those in the shade, due to either the closed shutter or overcast.

17
Discussion Winter survival The survival experiments indicate a general high survival of Harmonia axyridis in the Netherlands. In
Canada, no H. axyridis beetles survived outside, although 24.81% did survive inside houses (Labrie et
al., 2008). Furthermore, less than 10% of the beetles that overwintered outside in Connecticut, USA survived the winter. In contrast, in France more than 91% of the beetles overwintering in hills and
mountains survived (Iperti and Bertand, 2001). In Italy, a survival of 68.1% and 62.3% was recorded for beetles hibernating in an outdoor shelter (Bazzochi et al., 2004). Survival in the Netherlands lies in
the same range as in these European countries. Apparently, H. axyridis is well able to survive
European winters outside, although it could only survive in Canada (and probably northern USA), where winters are generally much colder than in Europe, when it searched 'cold free spaces' inside
(Labrie et al., 2008). The survival of two native species, Propylea quatuordecimpunctata and Adalia bipunctata in Italy was
much lower than that of H. axyridis (Bazzochi et al., 2004). A native Canadian species, Coleomegilla maculata, could survive outside but did not survive inside, and their (outside) winter survival ranged
from very low to 100%. C. septempunctata in Europe shows a similar range of survival (Labrie et al.,
2008). A little less than half the population of C. septempunctata survived in the Czech Republic (Honek, 1997) and only 17% survived in England and the Czech Republic (Zhou et al., 1995). The
survival of the native species A. bipunctata in the Netherlands was also low (16.9%, Brakefield, 1985) compared to the survival of H. axyridis found in this study. Apparently, H. axyridis is equally well, or
even better capable of surviving the European winters than native species are. Further research is
needed to elucidate whether differences in survival between native and invasive ladybird species indeed exist.
Influence of temperature on survival At the end of the winter, the survival at the central site was not uniform. Survival was lower at the left-hand side, than at either the right-hand side or the experiment at constant temperature. The left-
hand side showed the highest cumulative temperature, which indicates that the beetles endured
higher temperatures for a longer time. This phenomenon of lower survival at higher temperatures was found by other researchers as well,
especially for temperatures above 0 °C. Labrie et al. (2008) found 45% survival of H. axyridis at -5 °C but no survival at 10°C. Likewise, Watanabe (2002) found that H. axyridis had a lower survival at 5 °C
than at 0 °C, although at -5 °C the survival was again lower. A similar trend in survival at 15 °C, 10 °C
and 5 °C respectively was recorded by Lombaert et al. (2007), with higher survival at 5 °C than at 10 °C, but again a lower survival at 5 °C. No or hardly any aggregations were found at the two higher
temperatures however, and possibly therefore the beetles staying at these temperatures could not be considered as being in hibernation.
In contrast, at temperatures below 0 °C, survival declines with temperature, as was shown by
Berthiaume et al. (2003). Survival at 5 °C and 0 °C were both high, but the survival declined over decreasing temperatures (-5 °C, -10 °C, -20 °C). Berkvens et al. (2010) showed a similar pattern, with
a lower survival at -5 °C than at 0 °C, as was also clear from the results of Watanabe (2002) at -5 °C and 0 °C.
The difference in mortality rates between higher and lower temperatures may be due to different causes for this mortality. A difference between freezing and chilling injury exists. Chilling injury is the
mortality caused by exposure to low temperatures on a long term. It can be represented by the
Ltime50 (Berkvens et al., 2010). On the other hand, freezing is represented by the supercooling point, the temperature at which the beetles die of direct freezing. The supercooling point and thus the cold
tolerance vary throughout the winter, the lowest supercooling points occur during November (Berkvens et al., 2010) or December-February (Watanabe, 2002; Koch et al., 2004). The lethal
temperatures measured by Berkvens et al. (2010), Koch et al. (2004) and Watanabe (2002) in mid
winter ranged from -20 °C to -13.2°C. These are temperatures well below the normal temperatures H. axyridis will encounter during winter in temperate regions as the Netherlands. The lowest minimal
temperatures measured behind the shutters during the experiments in the winter of 2009-2010 were -

18
5.8 °C and -10.9 °C for the central site and original sites respectively. Temperatures during this winter
season were normal, compared with data („Langjarig gemiddelde van Nederland') of 1971-2000
(KNMI). The Canadian winters, during which a high winter mortality of H. axyridis was found, are usually much colder than winters in Europe (Labrie et al., 2008). Thus, the winter mortality in Europe
is more likely caused by an accumulation of chilling injury, rather than direct freezing (Watanabe, 2002; Berkvens et al., 2010).
The higher mortality at higher temperatures found in controlled environments as well as in the field,
could be explained by a depletion of energy reserves, rather than chilling injury (Watanabe, 2002). When temperature rises, the beetles become more active, causing a depletion of energy reserves.
Iperti and Bèrtand (2001) found a decreasing fat body during the winter period. Correspondingly, more lipids were found in H. axyridis at 5 °C than at 10 °C, indicating more activity at warmer
temperatures (Labrie et al., 2008). The fresh weight of adults kept under natural climatic conditions decreased from December to early March, as did the fat body from February onwards (Sakurai et al.,
1992).
Survival at constant temperature Hatherly et al. (2005) found that survival at 5 °C represented field survival the best for several species. The experiment at constant 5 °C showed a high survival until the end of the winter season,
which was comparable with the right-hand shutters at the experimental farm and with the survival at
the original sites. The total survival time (about 36 weeks) at 5 °C is comparable with the survival from November onwards found by Watanabe (2002). The Ltime50 (171 days) was slightly lower than
found by Watanabe (2002) at 5 °C, but slightly higher than found by Berkvens et al. (2010) at 0 °C when comparing populations overwintering from November onwards.
The high survival at the controlled temperature indicates that during the winter season, the experiment at constant 5 °C was representative for field survival in the Netherlands. Only when the
winter season was ended (from 18 March onwards), survival dropped quickly to 2.2% at August 17,
2010. From half March onwards, beetles in the field would disperse from their hibernation site to feeding and breeding areas, and no longer hibernate anymore. Only the strongest beetles were
capable to survive the constant low temperatures for a total of 9 months.
Influence of place of overwintering on survival The survival at the central site was lower for the beetles that originated from the sites in church towers (Alkmaar and Houten), than for those originating from the sites located in the woods
(Kootwijk, Loenen, Zundert). As these beetles all overwintered at the same location (the experimental farm), this difference in survival can be attributed to a difference between populations, rather than
the site of overwintering in the Netherlands. One of the possibilities is that less food can be found in
the urban area in autumn, so the beetles overwintering in the church towers had a poorer physical condition, resulting in a higher mortality. It is however not known how far from the feeding areas in
autumn the beetles fly towards their hibernations site, and to what extend the food availability here influences the survival chances during winter. Other factors so far unknown to us can be the cause of
this difference between sites as well.
The results of the overwintering at the original sites in the Netherlands (52.0 °N), resemble those of
Watanabe (2002) in Japan (36.0 °N), the native range of H. axyridis. Watanabe found a survival of 90% until the end of February, when first mating was observed, after which survival declined.
Survival at the original sites was highest for beetles overwintering at the south-west orientations, when compared to the north-east orientations. The largest aggregations of beetles were usually found
at the south-west facing side. At the site Loenen however, the largest aggregations were found at the
north-western side. Hardly any beetles aggregated at the south-facing orientations at this site. Large numbers of beetles were found at the north-facing side of Kootwijk as well.
The location of the largest beetle aggregations could be explained by the way the beetles fly towards the hibernation place in late autumn. Beetles fly in the afternoon towards surfaces illuminated by the
sun, so they alight on south-west facing surfaces (Obata et al., 1986; Nalepa et al., 2005; Koch, 2003). At the two sites where beetles were found on the north-facing side as well, however, the

19
buildings were closely surrounded by trees, except for the north-facing side (Loenen) or north- and
south-facing side (Kootwijk, where beetles were found on both sides). The trees at site Zundert were
less dense, and beetles were found here at the south-facing side only, similar to the situation at the church towers, which were high enough to be approached from all sides. Thus, when the south-facing
side, where the beetles would normally alight, is blocked, beetles are able to approach the building from another side and find a hibernation place there. The buildings in the denser woods were white,
which stands out against the darker background of the trees surrounding them. Therefore the beetles
should be able to find the building even without direct sunshine on the north-facing side, as they search for contrasting objects (Nalepa et al., 2005). Survival was however lower at this northern side,
though still high enough to survive the winter in large numbers. The strategy to fly to south-west facing surfaces is apparently a good strategy in most cases.
Comparing survival at the original sites, the sites located in the church towers showed a higher
survival than the sites located in the woods. This is in contrast to what was found at the central site.
Survival at the different orientations at Kootwijk confirms the hypothesis that the beetles survived better inside than outside. At the unheated attic or inside the attached barn, beetles survived better
than behind the shutters. Labrie et al. (2008) found that H. axyridis survived inside houses, which provided a stable environment and 'cold-free space' during winter. Apparently the inside of the church
towers or houses provided better shelter against the winter conditions (extreme temperatures and/or
humidity) than the shutters at the outside of the building, although both kept dry during snow and rainfall. Temperature data show that the site Alkmaar was slightly warmer and had less extremes
during the winter period than the other sites, although Houten did not show this trend. Beetles collected at the south-facing side in Kootwijk transferred to the unheated attic, survived better
than beetles collected at the north-facing side transferred to the attic. In contrast, the survival of beetles overwintering at the south-facing and north-facing side did not differ. This suggests that a
difference in survival between the two orientations is expressed only at the attic, and should thus be
attributed to a population, rather than to the specific place of overwintering. What may cause this difference is however yet unknown.
At the original sites the survival was higher than at the central site. This is possibly caused by more
handling at the central site (beetles were counted every two weeks at the central site, but only every
4 weeks at the original sites), although the influence of handling was minimal as found from the surplus bags. Another explanation might be that the artificial shutters at the central site are less
robust than those at the three sites in the woods, thus providing less protection against extremes in for example temperature and humidity.
Colour morphology The frequency of the colour morphs varies with the season and the geographical distribution for H. axyridis as well as Adalia bipunctata (Koch, 2003; de Jong and Brakefield, 1998; Brakefield, 1984). The significant variation in colour morphology showed no clear geographical trend in the Netherlands,
which was in contrast to earlier results with A. bipunctata (Brakefield and de Jong, in prep.; de Jong and Brakefield, 1998; Brakefield, 1984). From 1980 until 2004, a larger proportion of melanic A. bipunctata individuals was found inlands than near the Dutch coast, confirming the hypothesis that
the melanic individuals have an advantage in colder regions. Nowadays however this trend has disappeared for A. Bipunctata (Brakefield and de Jong, in prep.). The variation has largely disappeared
as well, the percentage melanics being about 20% everywhere in the Netherlands now (Brakefield and de Jong, in prep.). Possibly the same trend has disappeared for H. axyridis as well.
In its native range, the frequencies of melanic forms appeared to be linked to differences in climate,
although this was not the case in Japan (Michie et al., 2010). The non-melanic morph of H. axyridis, f. succinea, showed an adaptation of spot size to different temperatures, indicating a fast adaptation to
temperature differences. Also the frequency of melanic forms declined after introduction in Britain, suggesting a genetic adaptation to the new environment of the invasive H. axyridis (Michie et al.,
2010). This refutes the argument that H. axyridis has not been long enough in Europe to have adapted to differences in climate by variable colour morph frequencies. Further research is needed to

20
determine whether or not a geographical trend of colour morphs in the Netherlands exists, and what
may cause this.
Of the rarer colour morphs in Europe, only 2 f. equicolour were found and no f. axyridis, although the latter has been found at least 3 times before in the Netherlands (Raak – van den berg, unpublished
results). The prominent colour morph in parts of Asia is f axyridis, while f. succinea is dominant in Europe (Brown et al., 2008). It is suggested that the higher adaptability in spot variation of this colour
morph, compared to that of the melanic ones, explains the dominance of f. succinea in areas where it
is invasive (Michie et al., 2010).
Sex ratio The sex ratio differed between the sites; relatively fewer females were found in the church towers,
compared to the sites in the woods. Apparently, there is a difference in sex ratio between the different populations in the Netherlands. It is possible that Dutch populations are not homogeneously
distributed over the country, as was also clear from the variation in colour morphs. A restricted
migration between populations may create this non-homogenous pattern in sex ratio (and in colour morphology). However, large differences in sex ratio may also be caused by unequal sampling of the
population or may even be caused by micro-organisms as Wolbachia and other male-killing bacteria that influence the sex ratio (Koch, 2003).
Nevertheless, both colour and gender did not influence the survival, which is in accordance with
earlier results (Koch et al., 2004; Labrie et al., 2008; Berkvens et al., 2010).
Ovary development Hardly any ovary development was seen during the whole winter period, and the development that
was found (one beetle with ovaries in state 2), was found only very late in the overwintering season. At that time the overwintering experiments were stopped because beetles became active and would
normally disperse from their hibernation site. Sakurai et al. (1992) and Ceryngier et al. (2004) found
similar results, with hardly developed ovaries during the winter period, until March or April. Sakurai et al. (1988, 1992) concluded that H. axyridis showed true diapause, regulated by the corpus allatum.
The undeveloped ovaries during winter indicate H. axyridis do is in diapause (Iperti and Bèrtand, 2001; Sakurai et al., 1988; 1992; Hodek et al., 1996), and this is confirmed by the ovary regression
seen in this study. However, H. axyridis was shown to be capable of oviposition after only 1 to 2
weeks after transfer to 25 °C and long-day conditions, even in mid-winter (Raak et al., in prep.). This suggests a fast ovary development when the beetles were brought to suitable conditions, which is in
contrast to the results indicating true diapause. Therefore the question whether H. axyridis shows a true diapause or a mere quiescence, remains debatable.
Landing and dispersal During the observation in autumn, many beetles alighted on the coloured sunscreens. The colour of
the screens, varying from light green to brown, did not influence the landing behaviour of the beetles. However, more beetles were found landing near the edge of the screens, where they form a sharp
contrast to the darker background, than in the middle of the screens. This is in accordance with Nalepa et al. (2005), Obata et al. (1986) and Tanigishi (1976). Although Obata et al. (1986) argued
that H. axyridis is attracted to lighter coloured objects, Nalepa et al. (2005) found this was particularly
due to the contrast of these lighter objects to a darker background. Nalepa et al. (2005) observed that beetles preferred vertical contrasting lines, which are assumed to resemble vertical, and thus dry,
cracks and crevices in rocks; a typical place where H. axyridis overwinters in its native range (Nalepa et al., 2005; Koch, 2003; Obata et al., 1986).
Some research is done on the landing of beetles at their hibernations spot (Nalepa et al., 2005; Obata et al., 1986; Koch, 2003; Tanigishi, 1976), but little research on the leaving behaviour of the
hibernation spot is carried out (Sakurai et al., 1988; Iperti and Bèrtand, 2001; Tanigishi, 1976). Probably this is partly caused by contrast between the „massiveness‟ with which the beetles alight in
autumn, and the less pronounced spring dispersal.

21
The beetles left their hibernation spot with large numbers in a small amount of time. The radiation of
the sun greatly influenced the flying behaviour of the beetles when leaving their hibernation place.
Direct sunshine stimulated walking upwards and flying away, whereas the beetles moved much slower when in shade, caused either by the closed shutter door or by clouds. Obata et al. (1986) found that
alighting of beetles in autumn was sometimes interrupted by overcast. Apparently, H. axyridis reacts to the same cues when to fly in autumn, as in early spring upon leaving the hibernation site.
Natural enemies Next to abiotic factors influencing overwintering, biotic factors may play a role in winter survival as
well. Several fungus species were found on the dead beetles, though none of them seemed to be truly entomopathogenic. Steenberg and Harding (2009) found on average 4.5% of a H. axyridis population
infected with fungi. In accordance with Steenberg and Harding (2009), some Lecanicillium were found, but surprisingly not the species Isarina fariosa and Beauveria bassiana. The latter two are
known to be entomopathogenic to H. axyridis (Roy et al., 2008; Steenberg and Harding, 2009; Riddick
et al., 2009). Lecanicillium and Aspergillus ocharceous species may cause diseases in beetles, but there is not
enough known about these fungi on H. axyridis to conclude with certainty that the beetles were indeed infected by entomopathogenic fungi. The number of fungi that were thought to be possibly be
pathogenic were too low to make certain statements about fungal infections in this population of H. axyridis. The fungi that were thought possibly entomopathogenic at first (Myriodontium spp., Chromelosporium fulvum and Paecilomyces lilaciunus) were therefore considered to have not been
truly pathogenic in this population of H. axyridis (R. Samson, pers. comm.). The relative large amount of Aspergillus fumigatus found on the beetles indicates that the used
incubation temperature (25 °C) might have been too high for true entomopathogenic species to grow (R. Samson, pers. comm.).
In general, H. axyridis as an exotic species is known to be less susceptible to fungal infections (Roy et
al., 2008; Riddick et al., 2009), although the numbers found by Steenberg and Harding (2009) were similar to those found on other coccinellids.
Hesperomyces virescens is an ectoparasitic fungus that occurs on coccinellids (Riddick and Cottrell,
2010, Riddick, 2010; Riddick et al., 2009). It is horizontally transmitted during contact of beetles, for
example during winter aggregations. Riddick (2010) found 3.6% – 43.5% of an overwintering H. axyridis population infected with H. virescens, which is much more than found in this study. Although
survival was lower for beetles infected with the ectoparasite, especially in combination with the mite Coccipolipus hippodamiae (Riddick, 2010), H. virescens is not believed to be able to control coccinellid
field populations (Riddick et al., 2009).
The ectoparasitic mite Coccipolipus hippodamiae lives as sessile adult under the elytra of ladybird
beetles, where it consumes the heamolymph of the beetles. The motile larvae migrate between the beetles upon social contact (Rhule et al., 2010; Hurst et al., 1995; Webberly et al., 2004). Riddick
(2010) found that between 1.8% and 17.4% of overwintering H. axyridis in the USA carried a mite infection. The numbers found in this study are much lower. In 2009, C. hippodamiae was found to
have established in Polish populations of H. axyridis, although not in Britain (Rhule et al., 2010). This
study indicates that the mite also occurs in Dutch populations, although not in such high numbers as found in the USA or Poland. Whether this indicates a spread throughout Europe remains to be seen.
Rhule et al. (2010) found evidence that C. hippodamiae is well adapted to H. axyridis, and may cause sterility, although the current numbers in natural populations as found by Rhule et al. (2010) are not
high enough to influence the population size (Rhule et al., 2010).
Although the nematodes in this study could not be identified, they might belong to one (or both) of
the genera Steinernema or Hetherorhabditis, as these are known to be highly virulent in coccinellids (Riddick et al., 2009; Shapiro-Ilan and Cottrell, 2005). Shapiro-Ilan and Cottrell (2005) found H. axyridis susceptible to the nematodes S. carpocasae and H. bacteriophora, although they were less virulent on exotic ladybird species, than on native ones.

22
The parasitoid D. coccinellae parasitizes different species of coccinellids in varying levels (Riddick et
al., 2009, Hodek, 1973). Hoogendoorn and Heimpel (2002) found in a field survey that between 8.9% and 23.8% of H. axyridis was parasitized by D. coccinellae in the USA in summer. A much lower
number was found in this study, but possibly this is due to the fact that here only alive overwintering adults were examined. The percentage of parasitation of H. axyridis was similar to that of a native
species in the USA, Coleomegilla maculata, according to Hoogendoorn and Heimpel (2002). The
impact of the parasitoid on exotic species in comparison to native species in the Netherlands remains question for further research.
In this study, only low numbers of natural enemies were found in the field population of H. axyridis in
comparison with other studies. Although not all endosymbionts found may have caused direct mortality, they may have influenced mortality throughout the winter, and accounted for a small
percentage of the mortality found. Thus, the mortality caused by chill injury or starvation due to
hibernation may be even lower than initially found. It should be noted, however, that only beetles that were alive upon freezing were checked for infections. Beetles that died during the overwintering
period were only checked for fungus growth and not for mites, nematodes or parasitoids. The number of natural enemies in this population of H. axyridis may thus be higher than found in this study.
Future research is needed to determine the effects of natural enemies on the survival of overwintering
H. axyridis.
Conclusion The winter survival of H. axyridis in the Netherlands is high. This is due to several factors; H. axyridis is found to hibernate at suitable places to overwinter, in urban and rural environment. Moreover the ladybird can tolerate temperatures well below temperatures it will usually encounter during Dutch
winters. Higher temperatures however will cause higher mortality at the end of the overwintering season due to a depletion of energy reserves. Furthermore, the numbers of parasites that could
influence winter survival, such as fungi, mites, nematodes or parasitoids, found in H. axyridis in the Netherlands are generally low.
In conclusion, H. axyridis has an overwintering strategy that allows it to survive the Dutch winters in
large numbers. This will lead to large populations of H. axyridis in the breeding season in early spring, giving them a head start over other Coccinellidae. The overwintering strategy of H. axyridis may thus
contribute to its high invasiveness.

23
Acknowledgements First of all I would like to thank Lidwien Raak – van den Berg, my supervisor, for all her help with the experiment set-up, the counting of beetles, the thinking about and creating solutions to problems and
sending me home when I was working too hard. I would also like to thank Peter de Jong and Joop
van Lenteren, second supervisor and examinator, for their critical remarks and advises on the set-up of the experiments and the writing of the report. Thank you to Lia Hemerik for her advice on the
statistics and the design of the experiments as well. All other staff at Entomology who helped me one way or another with my research or created a good
atmosphere at the coffee breaks, as well as the people from the experimental farm, Andries Siepel,
Gerard Mekking, and Henk van Roekel, and of course Antoon Loomans for helping to collect the ladybeetles; thanks!
Of course I am grateful as well to all people who provided their home or other place to collect and count the beetles during winter: Mary Huijsman (Kootwijk), Koen Helling (Houten), Ebbo Voorhout
(Alkmaar), Roelof Brandsma (Loenen) and Hans Schep (Zundert). The following people helped as well to collect the beetles: Peter van Wieringen, Jan Doevedans, Dr. John Slogget, Martin Hermans, Theo
Meeks, Anneke van Zandvoort, Anje and Hendrik Buining, Tikie de Haan, Ria van Dijk and Margriet
and Hendrik-Jan Boneschaker. Many thanks to Prof. Rob A. Samson and Jos Houbraken (CBS Fungal Biodiversity Centre) for
identifying the fungi, as well as Emma Rhule (University of Cambridge; UK Ladybird Survey) for identification and advise on the mites, Hein Overmars and Sven van den Elsen (Wageningen
University, Laboratory of Nematology) for help with identifying nematodes and Dr. Piotr Ceryngier
(Center for Ecological Research, Dziekanów Leśny) for advise on dissection and parasitic wasp identification.

24
References Bazzocchi, G.G.; Lanzoni, A.; Accinelli, G.; Burgio, G. (2004). Overwintering, phenology and fecundity
of Harmonia axyridis in comparison with native coccinellid species in Italy. BioControl 49:
245–260
Berkvens, N; Baverstock, J; De Clercq, P; Majerus, M.E.N.; Pell, J.K.; Roy, H.E; Tirry, L; Wells, P.M.
(2008). Good and bad traits of Harmonia axyridis – from biological control to invasion. Procceedings of the Third International Symposium on Biological Control of Arthropods, Christchurch, New Zealand. Peter G. Mason, David R. Gillespie & Charles Vincent (2008)
Berkvens, N.; Bale, J. S.; Berkvens, D.; Tirry, L.; De Clerq, P. (2010). Cold tolerance of the harlequin
ladybird Harmonia axyridis in Europe. Journal of Insect Physiology 56: 438-444
Berthiaume, R.; Hébert, C.; Bauce, É. (2003). Impact de la température et de la durée d‟exposition au froid sur la survie des adultes de la coccinelle asiatique, Harmonia axyridis (Pallas)
Phytoprotection 84(2): 85-91
Brakefield, P.M.(1984). Ecological Studies on the Polymorphic Ladybird Adalia bipunctata in the
Netherlands. I. Population Biology and Geographical Variation of Melanism. Journal of Animal Ecology 53(3): 761-774
Brakefield, P.M. (1985). Differential winter mortality and seasonal selection in the polymorphic
ladybird Adalia bipunctata (L) in the Netherlands. Biological Journal of the Linnean Society 24:
189-206
Brakefield, P.M.; De Jong, P.W. (in preperation). A steep cline in ladybird melanism has decayed over
25 years: an effect of climate change?
Brown, P.M.J.; Adriaens, T.; Bathon, H.; Cuppen, J.; Goldarazena, A.; Hägg, T.; Kenis, M.; Klausnitzer, B.E.M.; Kovář, I.; Loomans, A.J.M.; Majerus, M.E.N.; Nedved, O.; Pedersen, J.; Rabitsch, W.;
Roy, H.E.; Ternois, V.; Zakharov, I.A.; Roy, D.B. (2008). Harmonia axyridis in Europe: spread
and distribution of a non-native coccinellid. BioControl 53: 5-21
Ceryngier, P.; Havelka, J.; Hodek, I. (2004). Mating and activity of gonads in pre-dormant and dormant ladybirds (Coleoptera: Coccinellidae). Invertebrate Reproduction and Development 42(2): 127-135
Cox, D.R. (1972). Regression models and Life-Tables. Journal of the Royal Statistical Society, Series B
(Methodological). 34(2): 187-220
Hatherly, I.S.; Hart, A.J.; Tullett, A.G.; Bale, J.S. (2005). Use of thermal data as a screen for the establishment potential of non-native biological control agents in the UK. Biocontrol 50(5):
687-698
Hodek, I. (1973). Biology of coccinellidae. Academia, Praha, 260 p.
Hodek, I. (1996). Diapause development, diapause termination and the end of diapause. European
Journal of Entomology 93(3): 475-487
Hodek, I.; Honek, A.; Ceryngier, P.; Kovar, I. (1996). Ecology of Coccinellidae. Series entomologica,
Volume 54. Kluwer, Dordrecht [etc], 464 p.

25
Honek, A. (1997). Factors determining winter survival in Coccinella septempunctata (Col.:
Coccinellidae). Entomophaga 42(1/2):119-124
Hoogendoorn, M.; Heimpel, G.E. (2002). Indirect interactions between an introduced and a native
ladybird beetle species mediated by a shared parasitoid. Biological Control 25(3): 224-230
Hurst, G.D.D.; Sharpe, R.G.; Broomfield, A.H.; Walker, L.E; Majerus, T.M.O.; Zakharov, I.A.; Majerus,
M.N. (1995). Sexually transmitted disease in a promiscuous insect, Adalia bipunctata; Ecological Entomology 20:230-236
Iperti, G., Bèrtand, E. (2001). Hibernation of Harmonia axyridis (Coleoptera: Coccinellidae) in South-
Eastern France. Acta Soc. Zool. Bohem. 65: 207-210
Jong, P.W., de; Brakefield, P.M. (1998). Climate and change in clines for melanism in the two-spot
ladybird, Adalia bipunctata (Coleoptera: Coccinellidae). Proc. R. Soc. Lond. B. 265:39-43
Koch, R.L.; Galvan, T.L. (2008). Bad side of a good beetle: the North American experience with Harmonia axyridis. BioControl 53:23-35
Koch. R.L; Carrillo, M.A.; Venette, R.C.; Cannon, C.A.; Hutchison, W.D. (2004). Cold Hardiness of the Multicolored Asian Lady Beetle (Coleoptera: Coccinellidae). Environ. Entomol 33(4): 815-822
Koch, R.L. (2003). The multicolored Asian lady beetle, Harmonia axyridis: A review of its biology, uses
in biological control, and non-target impacts. Journal of Insect Science 3: pp 32
Labrie, G.; Coderre, D.; Lucas, E. (2008). Overwintering Strategy of Multicolored Asian Lady Beetle
(Coleoptera: Coccinellidae): Cold-Free Space As a Factor of Invasive Success. Ann. Entomol. Soc. Am. 101(5): 860-866
Lefebvre, D.; Pierre, J.; Outreman, Y.; Pierre, J.S. (2007). Patch departure rules in Bumblebees:
evidence of a decremental motivational mechanism. Behavioral Ecology and Sociobiology
61(11): 1707-1715
Lenteren, J.C., van; Babendreier, D.; Bigler, F.; Burgio, G.; Hokkanen, H.M.T.; Kuske, S.; Loomans, A.J.M.; Menzler-Hokkanen, I.; Van Rijn, P.C.J.; Thomas, M.B.; Tommasini, M.G.; Zeng, Q.Q.
(2003). Environmental risk assessment of exotic natural enemies used in inundative biological
control. Biocontrol 48(1):3-38
Lenteren, J.C., van; Loomans, A.J.M.; Babendreier, D.; Bigler, F. (2008). Harmonia axyridis: an environmental risk assessment for Northwest Europe. BioControl 53:37-54
Lombaert, E.; Malausa, T.; Devred, R.; Estoup, A. (2008). Phenotypic variation in invasive and
biocontrol populations of the harlequin ladybird, Harmonia axyridis. BioControl 53(1): 89-102
Michie, L.J.; Mallard, F.; Majerus. M.E.N.; Jiggins, F.M. (2010). Melanic through nature or nurture:
genetic polymorphism and phenotypic plasticity in Harmonia axyridis. Journal of Evolutionary Biology 23: 1699-1707
Nalepa, C.A.; Kennedy, G.G.; Brownie, C. (2005). Role of Visual Contrast in the Alighting Behavior of Harmonia axyridis (Coleoptera: Coccinellidae) at Overwintering Sites. Envoron. Entomol. 34(2): 425-431

26
Obata, S. (1986). Determination of the hibernation site in the Ladybird Beetle, Harmonia axyridis (Pallas) (Coleoptera: coccinellidae) The Entomological Society of Japan; Kontyû, Japan 54(2):
218-223.
Obata, S.; Johki, Y.; Hidaka, T. (1986). Location of hibernation sites in the ladybird beetle, Harmonia axyridis. Hodek I. (ed.) Ecology of Aphidophaga. Academia, Prague 193-198.
Rhule. E.L.; Majerus, M.E.N.; Jiggins, F.M.; Ware, R.L. (2010); Potential role of the sexually transmitted mite Coccipolipus hippodamiae in controlling populations of the invasive ladybird
Harmonia axyridis. Biological Control, 53(2): 243-247
Riddick, E.W.; Cottrell, T.E.; Kidd, K.A. (2009). Natural Enemies of the Coccinellidae: Parasites, pathogens, and parasitoids; Biological Control 51(2): 306-312
Riddick, E.W. (2010). Ectoparasitic mite and fungus on an invasive lady beelte: parasite coexistence and influence on host survival. Bulletin of Insectology 63(1):13-20
Riddick, E.W.; Cottrell, T.E. (2010). Is the prevalence and intensity of the ectoparasitic fungus
Hesperomyces virescens related to the abundance of entomophagous coccinellids? Bulletin of Insectology 63(1):71-78
Roy, H.E.; Brown, P.M.J.; Rothery, P.; Ware, R.L.; Majerus, M.E.N. (2008). Interactions between the fungal pathogen Beauveria bassiana and three species of coccinellid: Harmonia axyridis,
Coccinella septempunctata and Adalia bipunctata. BioControl 53(1): 265-276
Sakurai, H.; Kawai, T.; Takeda, S. (1992). Physiological changes in relation to diapause of the lady
beetle, Harmonia axyridis (Coleoptera: Coccinellidae). Appl. Entomol. Zool. 27(4):479-487
Sakurai, H.; Kumada, Y.; Takeda, S.. (1993). Seasonal prevalence and Hibernating-Diapause Behavior in the Lady Beetle, Harmonia axyridis. Research Bulletin of the Faculty of Agricultrue, Gifu University 58: 51-55
Sakurai, H.; Hirano, T.; Takeda, S. (1986). Physiological Distinction between Aestivation and
Hibernation in the Lady Beetle, Coccinella septempunctata bruckii (Coleoptera: Coccinellidae). Appl. Ent. Zool. 21(3): 424-429
Sakurai, H.; Takeda, S.; Kawai, T. (1988). Diapause regulations in the lady beetle, Harmonia axyridis. Ecology and Effectiveness of Aphidophaga. pp. 67-70.
Shapiro-Ilan, D.I.; Cottrell, T.E. (2005). Susceptibility of lady beetles (Coleoptera: Coccinellidae) to
entomopathogeneic nematodes. Journal of Invertebrate Pathology 89(2): 150-156
Steenberg, T.; Harding, S. (2009). Entomophagic fungi recorded from the harlequin ladybird,
Harmonia axyridis. Journal of Invertebrate Pathology 102: 88-89
Tanigishi, K. (1976); Hibernation of the lady beetle, Harmonia axyridis. Insectarium 13:294-298
Tauber, M.J.; Tauber, C. A.; Masaki, S. (1986). Seasonal adaptations of insects. Oxford University
Press, New York, 411 p.
Webberley, K.M.; Hurst, G.D.D.; Husband, R.W.; Hinrich, J.; Schulenburg, G.V.D.; Sloggett, J.J.; Isham, V.; Buszko, J.; Majerus, M.E.N. (2004). Host reproduction and a sexually transmitted
disease: causes and consequences of Coccipolipus hippodmiae distribution on coccinellid beetles. Journal of Animal Ecology 73: 1-10.

27
Watanabe, M. (2002). Cold tolerance and myo-inositol accumulation in overwintering adults of a lady
beetle, Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol 99: 5-9
Zhou, X.; Honek, A.; Powell, W.; Carter, N.(1995). Variations in body length, weight, fat content and survival in Coccinella septempunctata at different hibernation sites. Entomologica Experimentalis et Applicata 75: 99-107

28
Appendix 1 - Figures and tables Numbered according to numbers in text
Appendix contents Figure 1 – Shutters at the experimental farm 29
Figure 2 – Schematic overview of original sites 30 Figure 3 – Sites where survival was monitored 31
Figure 4 – Coloured screens on which H. axyridis alighted in autumn 32 Figure 5 – Survival curves, central site 33
Figure 6 – Daily average temperature, central site 35
Figure 7 – Cumulative temperature, central site 36 Figure 8 – Survival curve, constant temperature 37
Figure 9 – Daily average temperature at original sites 38 Figure 10 – Cumulative temperature at original sites 39
Figure 11 – Ovaries of overwintering H. axyridis 40
Figure 12 – Mites C. hippodamiae under the elytra of H. axyridis 41 Figure 13 – Nematodes found in the abdomen of H. axyridis 42
Figure 14 – Larve of the parasitoid D. coccinellae found in the abdomen of H. axyridis 43 Figure 15 – Map of the Netherlands indicating collection sites 44
Table 1 – Geographical coordinates of sites where survival was monitored 45
Table 2 – States of ovary development 46
Table 3 – Number of beetles overwintering at the central and original sites 47 Table 4 – Mean survival times 48
Table 5 – Ovary development results 49 Table 6 – Fungi found on H. axyridis 50
Table 7 – Parasites found in overwintering H. axyridis 51
Table 8 – Total numbers of H. axyridis found, specified to colour morph 52

29
Figure 1 – Shutters at the experimental farm
Figure 1. Shutters at the experimental farm Droevendaal, Wageningen. (a) Gauze bags contain
ladybeetles, for both the overwintering experiment (yellow labels) and ovary dissection (blue labels).
Datalogger to record temperature and relative humidity. (b) Inside sizes of shutter are 60x105cm,
3cm deep. (c) The experimental farm, showing the three shutters at the left, and three at the right of
the central door.
A
C
B

30
Figure 2 – Schematic overview of original sites
Figure 2. Schematic overview of the original sites (a) Alkmaar; (b) Houten; (c) Kootwijk; (d) Loenen
and (e) Zundert. Black: building; white: open area,
light gray background (a, b): urban surroundings;
dark grey background (c, d, e): trees. Red dots
indicate places were bags were placed. At Kootwijk,
the two bags inside the building indicate bags on
unheated attic; the bag in tree-area is placed in leaf
litter. The other three are from left to right: south-
west (many beetles found), north-east (no beetles
found) and north (many beetles found). North is
pointed upwards in all figures; figures are not on
scale.
A B
E
D C
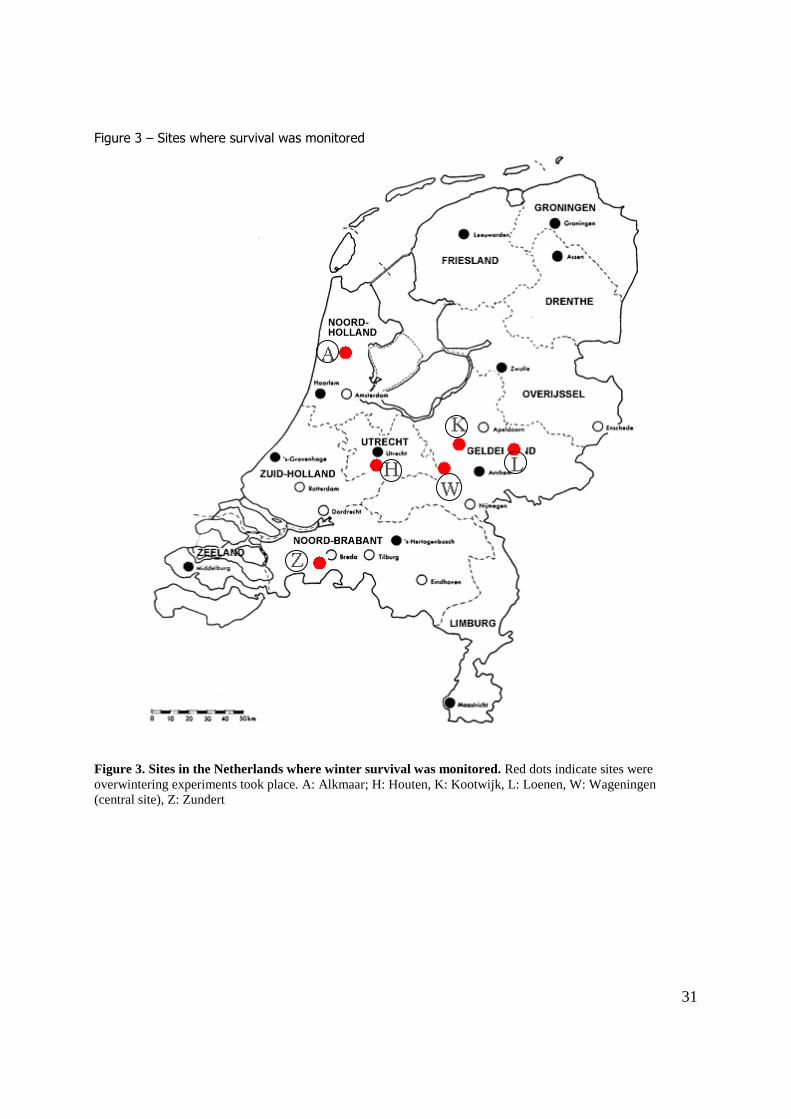
31
Figure 3 – Sites where survival was monitored
Figure 3. Sites in the Netherlands where winter survival was monitored. Red dots indicate sites were
overwintering experiments took place. A: Alkmaar; H: Houten, K: Kootwijk, L: Loenen, W: Wageningen
(central site), Z: Zundert

32
Figure 4 – Coloured screens on which H. axyridis alighted in autumn
Figure 4. Coloured sunscreens on which landing of H. axyridis in late autumn was observed.
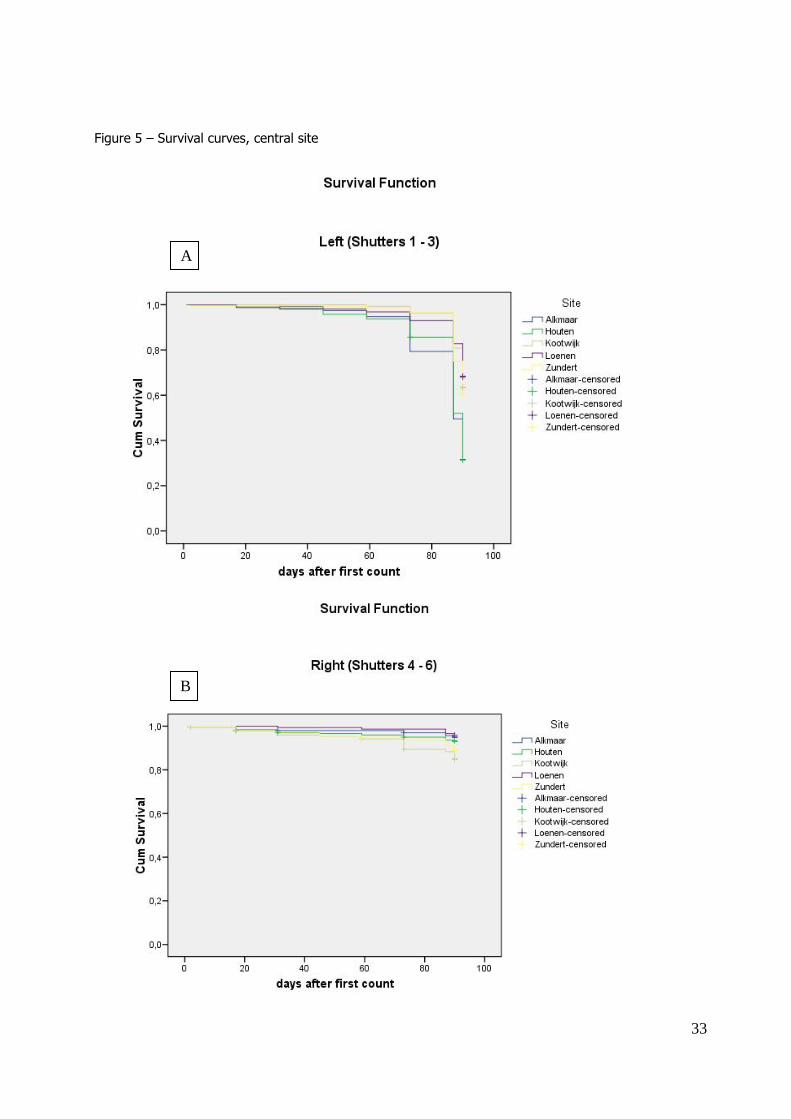
33
Figure 5 – Survival curves, central site
B
A

34
Figure 5. Survival curves at the central site. (a) Shutters 1-3 (left-hand side of central site), (b) shutters 4-6
(right-hand side of central site), (c) constant 5 °C, laboratory.
C
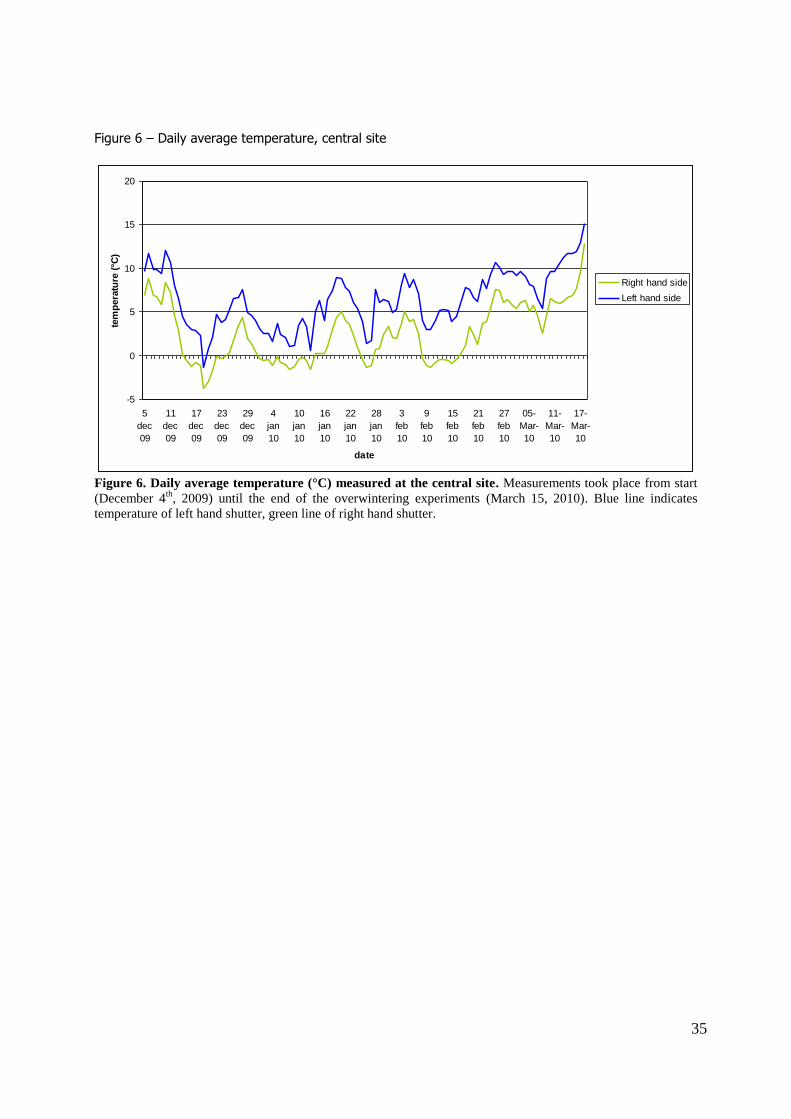
35
Figure 6 – Daily average temperature, central site
-5
0
5
10
15
20
5
dec
09
11
dec
09
17
dec
09
23
dec
09
29
dec
09
4
jan
10
10
jan
10
16
jan
10
22
jan
10
28
jan
10
3
feb
10
9
feb
10
15
feb
10
21
feb
10
27
feb
10
05-
Mar-
10
11-
Mar-
10
17-
Mar-
10
date
tem
pera
ture
(°C
)
Right hand side
Left hand side
Figure 6. Daily average temperature (°C) measured at the central site. Measurements took place from start
(December 4th
, 2009) until the end of the overwintering experiments (March 15, 2010). Blue line indicates
temperature of left hand shutter, green line of right hand shutter.
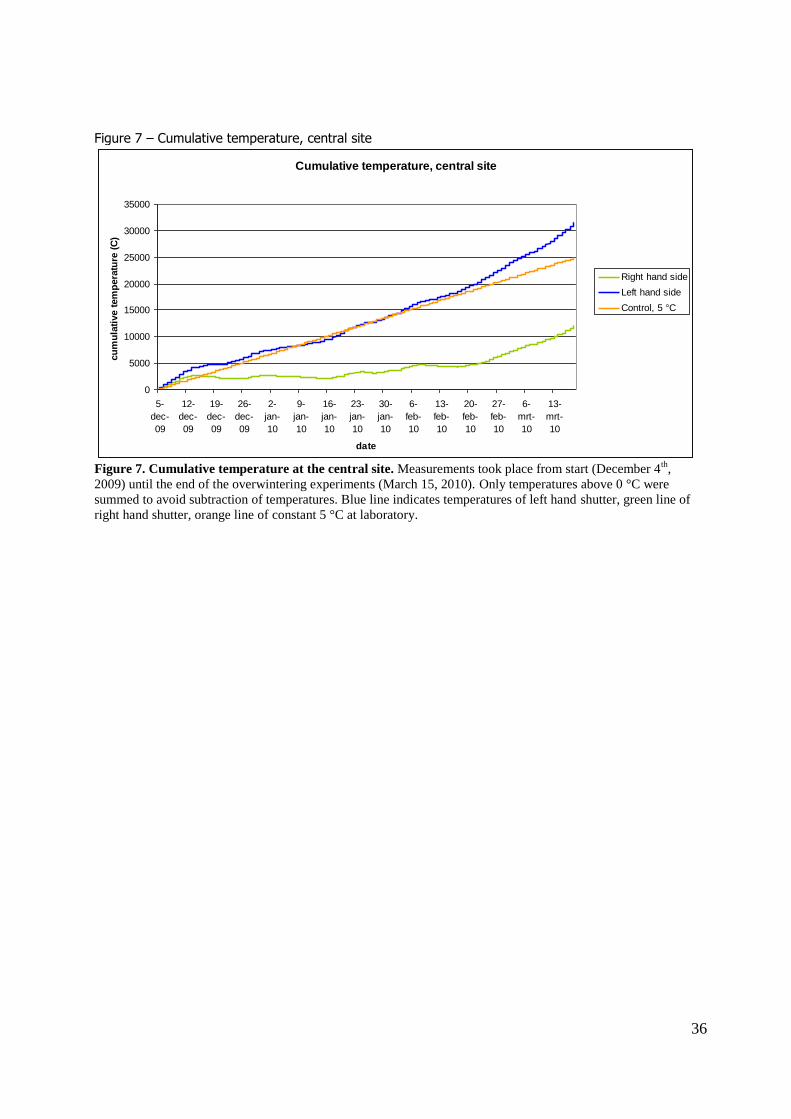
36
Figure 7 – Cumulative temperature, central site
Cumulative temperature, central site
0
5000
10000
15000
20000
25000
30000
35000
5-
dec-
09
12-
dec-
09
19-
dec-
09
26-
dec-
09
2-
jan-
10
9-
jan-
10
16-
jan-
10
23-
jan-
10
30-
jan-
10
6-
feb-
10
13-
feb-
10
20-
feb-
10
27-
feb-
10
6-
mrt-
10
13-
mrt-
10
date
cu
mu
lati
ve t
em
pera
ture
(C
)
Right hand side
Left hand side
Control, 5 °C
Figure 7. Cumulative temperature at the central site. Measurements took place from start (December 4
th,
2009) until the end of the overwintering experiments (March 15, 2010). Only temperatures above 0 °C were
summed to avoid subtraction of temperatures. Blue line indicates temperatures of left hand shutter, green line of
right hand shutter, orange line of constant 5 °C at laboratory.
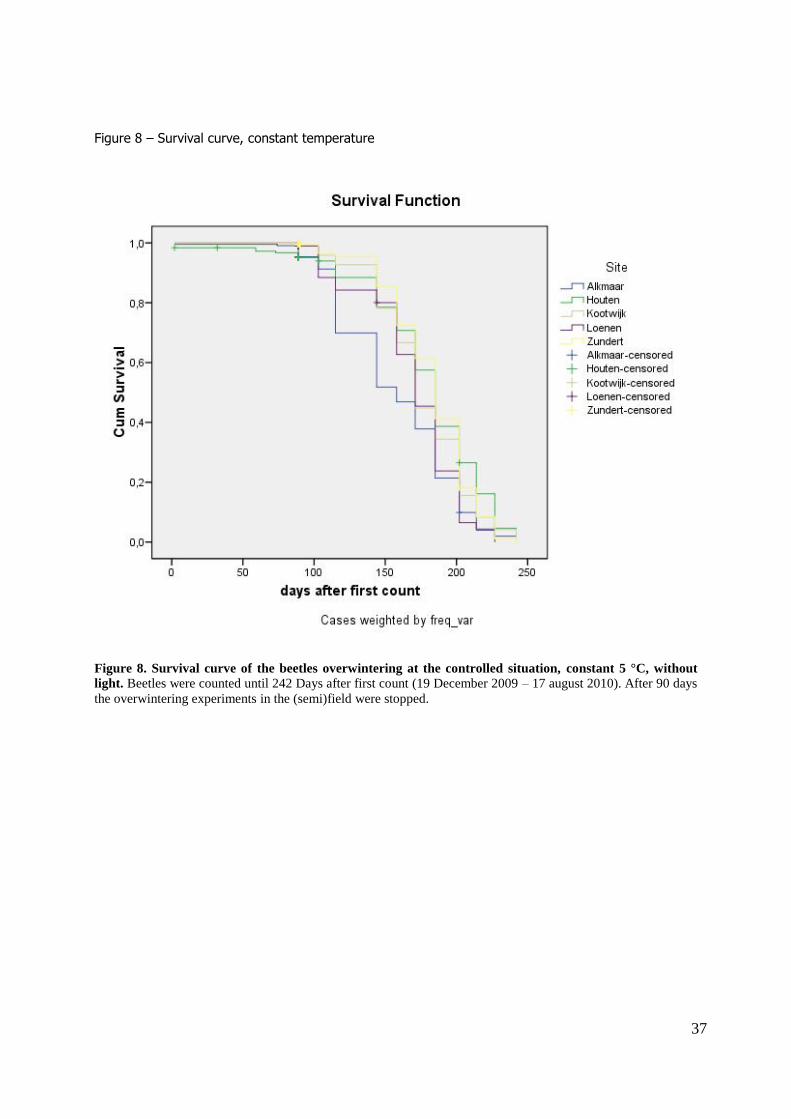
37
Figure 8 – Survival curve, constant temperature
Figure 8. Survival curve of the beetles overwintering at the controlled situation, constant 5 °C, without
light. Beetles were counted until 242 Days after first count (19 December 2009 – 17 august 2010). After 90 days
the overwintering experiments in the (semi)field were stopped.
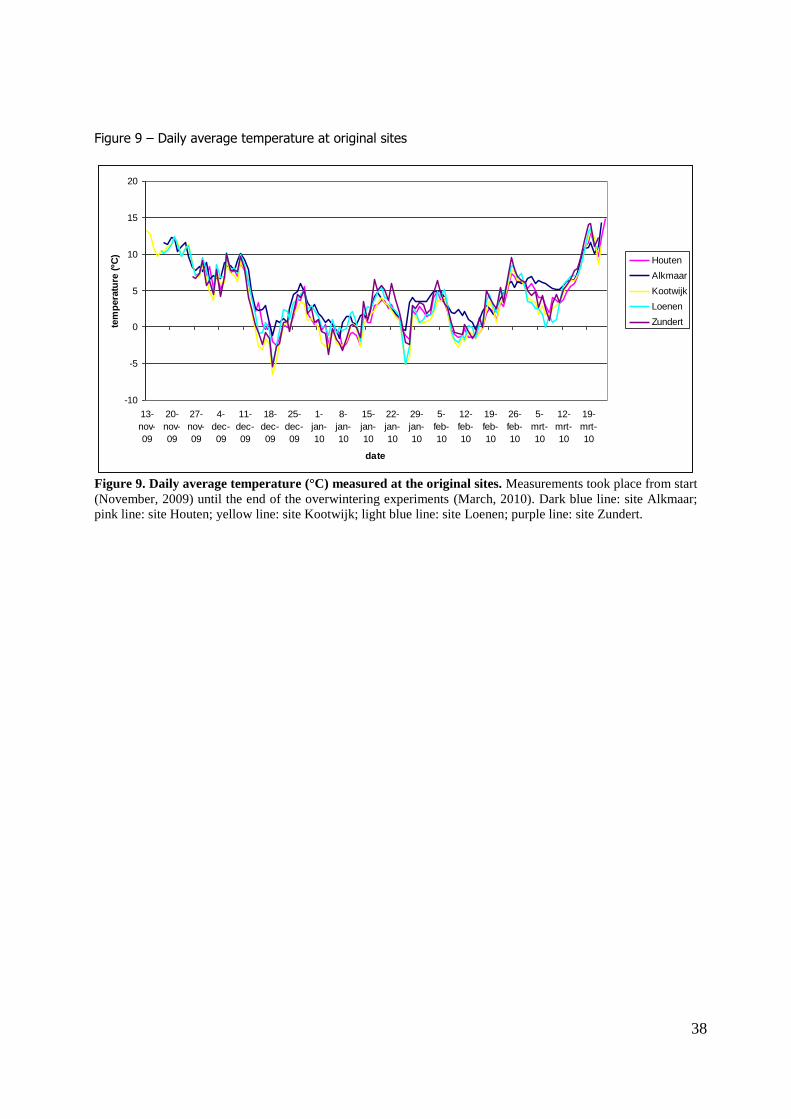
38
Figure 9 – Daily average temperature at original sites
-10
-5
0
5
10
15
20
13-
nov-
09
20-
nov-
09
27-
nov-
09
4-
dec-
09
11-
dec-
09
18-
dec-
09
25-
dec-
09
1-
jan-
10
8-
jan-
10
15-
jan-
10
22-
jan-
10
29-
jan-
10
5-
feb-
10
12-
feb-
10
19-
feb-
10
26-
feb-
10
5-
mrt-
10
12-
mrt-
10
19-
mrt-
10
date
tem
pera
ture
(ºC
)
Houten
Alkmaar
Kootwijk
Loenen
Zundert
Figure 9. Daily average temperature (°C) measured at the original sites. Measurements took place from start
(November, 2009) until the end of the overwintering experiments (March, 2010). Dark blue line: site Alkmaar;
pink line: site Houten; yellow line: site Kootwijk; light blue line: site Loenen; purple line: site Zundert.
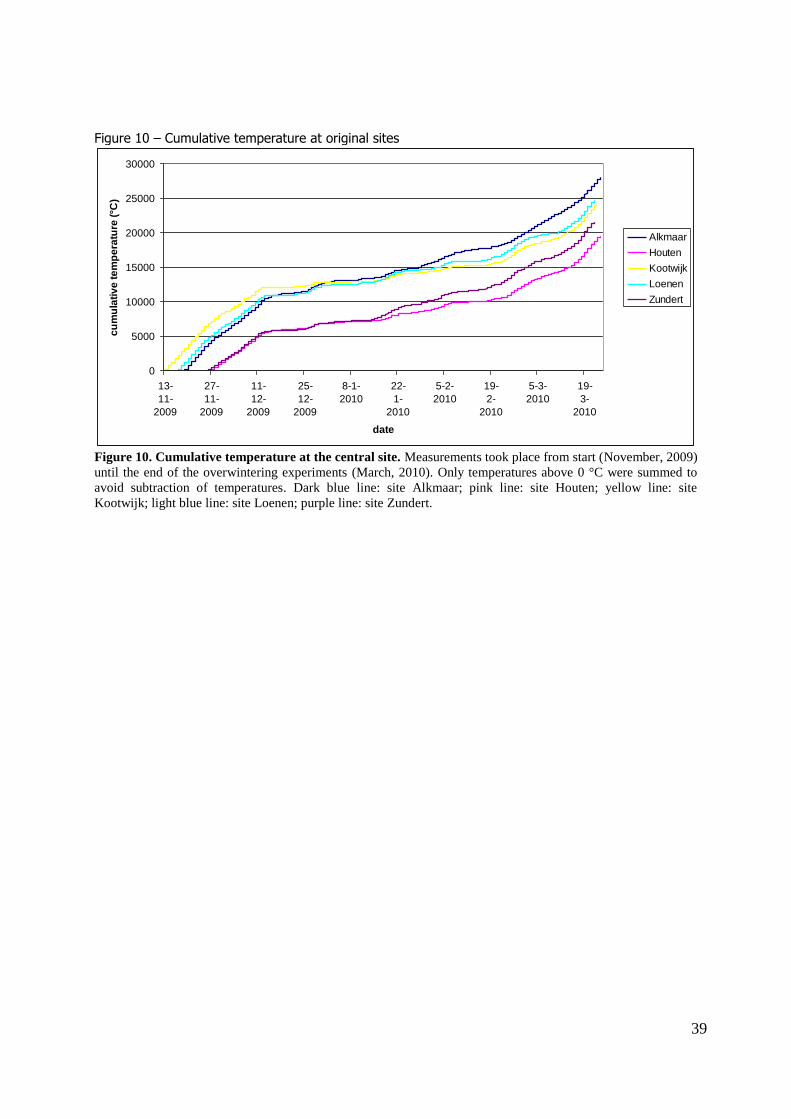
39
Figure 10 – Cumulative temperature at original sites
0
5000
10000
15000
20000
25000
30000
13-
11-
2009
27-
11-
2009
11-
12-
2009
25-
12-
2009
8-1-
2010
22-
1-
2010
5-2-
2010
19-
2-
2010
5-3-
2010
19-
3-
2010
date
cu
mu
lati
ve
te
mp
era
ture
(°C
)
Alkmaar
Houten
Kootwijk
Loenen
Zundert
Figure 10. Cumulative temperature at the central site. Measurements took place from start (November, 2009)
until the end of the overwintering experiments (March, 2010). Only temperatures above 0 °C were summed to
avoid subtraction of temperatures. Dark blue line: site Alkmaar; pink line: site Houten; yellow line: site
Kootwijk; light blue line: site Loenen; purple line: site Zundert.
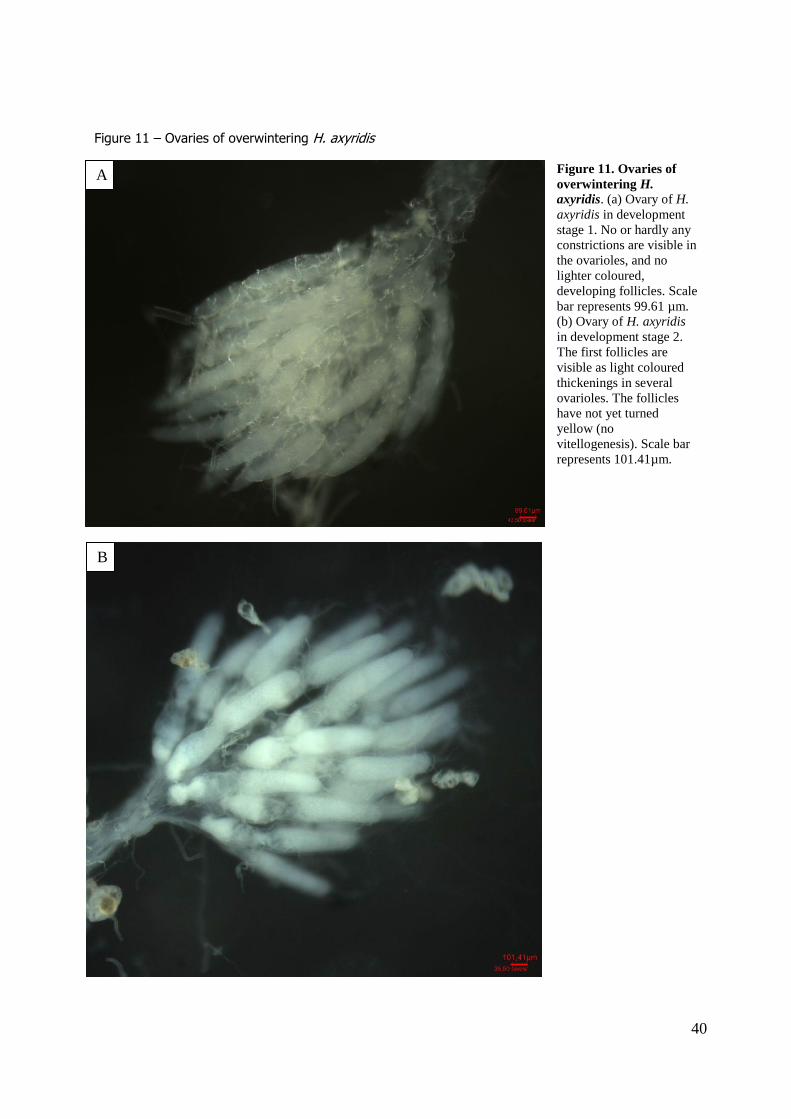
40
Figure 11 – Ovaries of overwintering H. axyridis Figure 11. Ovaries of
overwintering H.
axyridis. (a) Ovary of H.
axyridis in development
stage 1. No or hardly any
constrictions are visible in
the ovarioles, and no
lighter coloured,
developing follicles. Scale
bar represents 99.61 µm.
(b) Ovary of H. axyridis
in development stage 2.
The first follicles are
visible as light coloured
thickenings in several
ovarioles. The follicles
have not yet turned
yellow (no
vitellogenesis). Scale bar
represents 101.41µm.
A
B
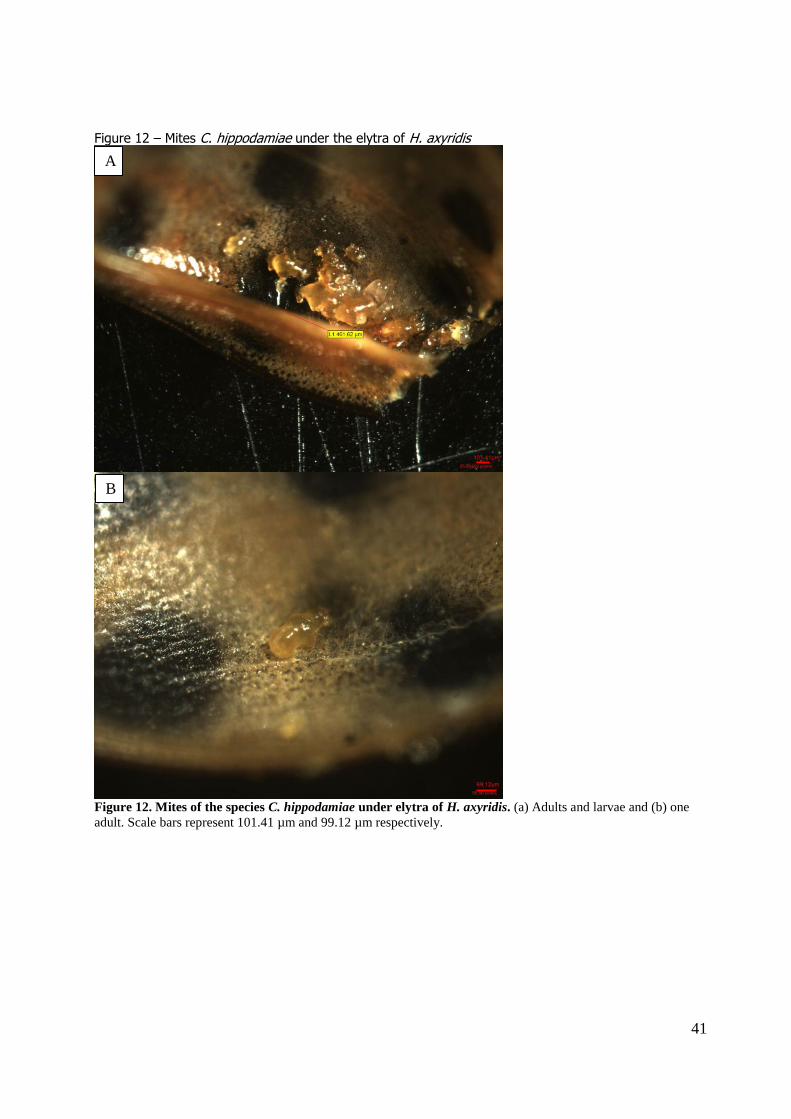
41
Figure 12 – Mites C. hippodamiae under the elytra of H. axyridis
Figure 12. Mites of the species C. hippodamiae under elytra of H. axyridis. (a) Adults and larvae and (b) one
adult. Scale bars represent 101.41 µm and 99.12 µm respectively.
B
A
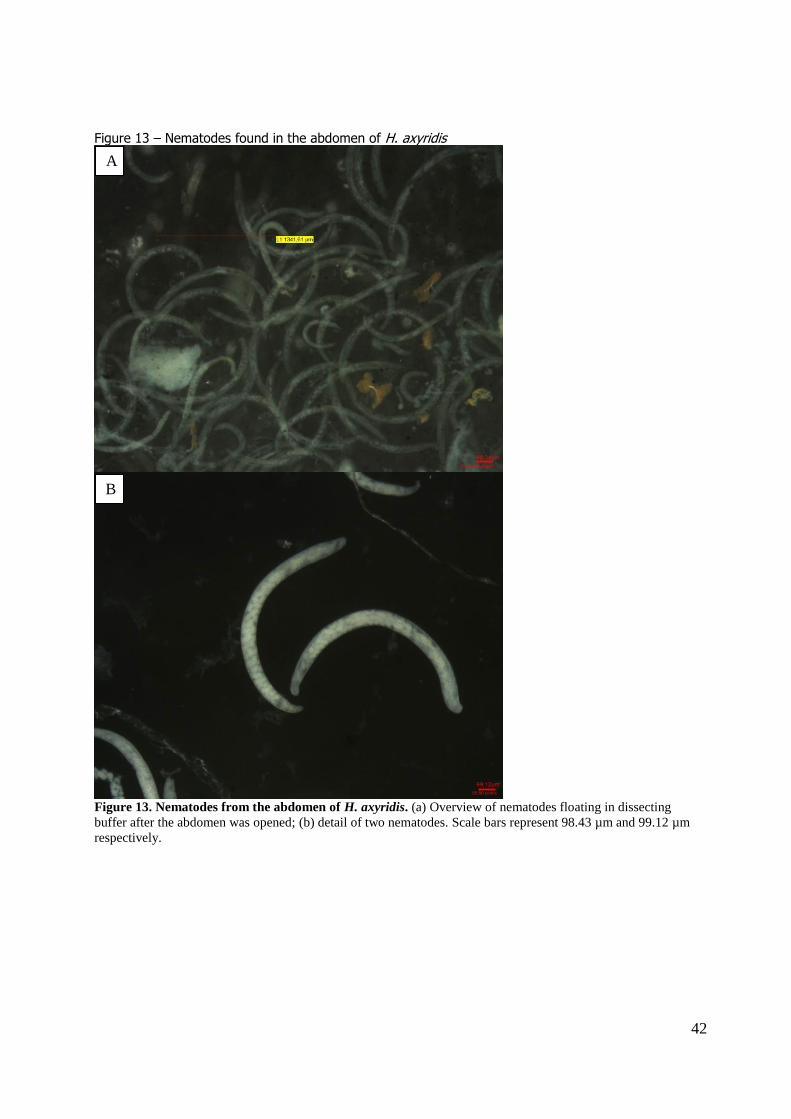
42
Figure 13 – Nematodes found in the abdomen of H. axyridis
Figure 13. Nematodes from the abdomen of H. axyridis. (a) Overview of nematodes floating in dissecting
buffer after the abdomen was opened; (b) detail of two nematodes. Scale bars represent 98.43 µm and 99.12 µm
respectively.
A
B

43
Figure 14 – Larve of the parasitoid D. coccinellae found in the abdomen of H. axyridis
Figure 14. Larve of Dinocampus coccinellae found in the abdomen of H. axyridis. (a) Overview of whole
larvae; (b) detail of head. Scale bar in (b) represents 100.41 µm.
A
B

44
Figure 15 – Map of the Netherlands indicating collection sites
Figure 15. Map of the Netherlands with 20 sites where H. axyridis beetles were collected, grouped into 7
areas. Yellow circle: Groningen; Green circle: Alkmaar; Blue circle: Ommen; Red circle: middle Netherlands
(Houten, Kootwijk, Loenen, Deelen, Wageningen, Alphen); light blue circle: Zundert and Tholen; Grey circle:
Waalre; Purple circle: Born and Landgraaf. Green points indicate five sites were overwintering experiments were
conducted, blue and yellow points are additional sites.
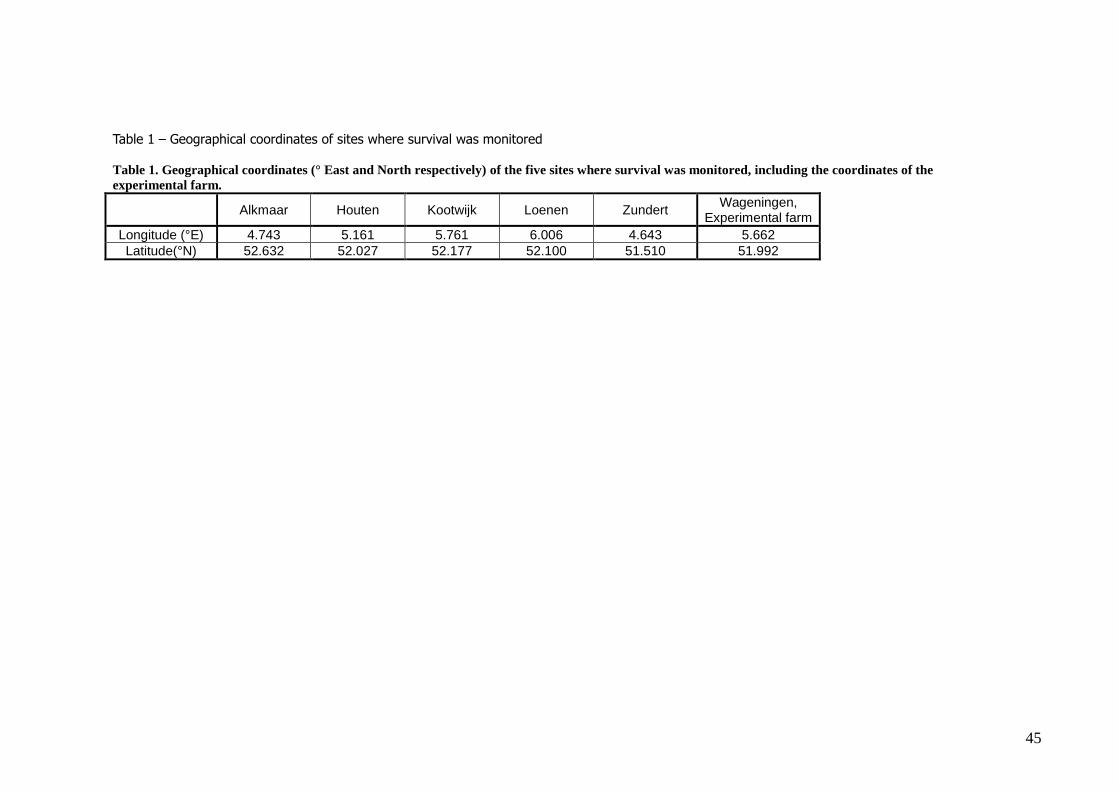
45
Table 1 – Geographical coordinates of sites where survival was monitored Table 1. Geographical coordinates (° East and North respectively) of the five sites where survival was monitored, including the coordinates of the
experimental farm.
Alkmaar Houten Kootwijk Loenen Zundert Wageningen,
Experimental farm
Longitude (°E) 4.743 5.161 5.761 6.006 4.643 5.662
Latitude(°N) 52.632 52.027 52.177 52.100 51.510 51.992
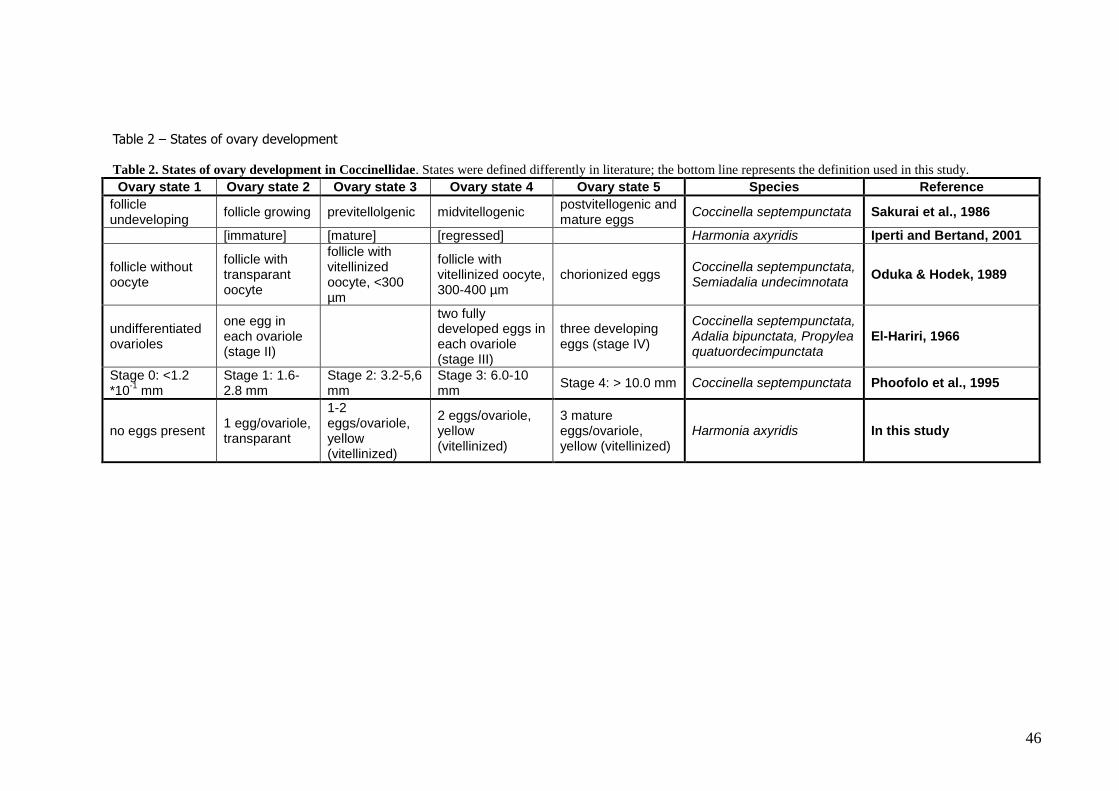
46
Table 2 – States of ovary development Table 2. States of ovary development in Coccinellidae. States were defined differently in literature; the bottom line represents the definition used in this study.
Ovary state 1 Ovary state 2 Ovary state 3 Ovary state 4 Ovary state 5 Species Reference
follicle undeveloping
follicle growing previtellolgenic midvitellogenic postvitellogenic and mature eggs
Coccinella septempunctata Sakurai et al., 1986
[immature] [mature] [regressed] Harmonia axyridis Iperti and Bertand, 2001
follicle without oocyte
follicle with transparant oocyte
follicle with vitellinized oocyte, <300 µm
follicle with vitellinized oocyte, 300-400 µm
chorionized eggs Coccinella septempunctata, Semiadalia undecimnotata
Oduka & Hodek, 1989
undifferentiated ovarioles
one egg in each ovariole (stage II)
two fully developed eggs in each ovariole (stage III)
three developing eggs (stage IV)
Coccinella septempunctata, Adalia bipunctata, Propylea quatuordecimpunctata
El-Hariri, 1966
Stage 0: <1.2 *10
-1 mm
Stage 1: 1.6-2.8 mm
Stage 2: 3.2-5,6 mm
Stage 3: 6.0-10 mm
Stage 4: > 10.0 mm Coccinella septempunctata Phoofolo et al., 1995
no eggs present 1 egg/ovariole, transparant
1-2 eggs/ovariole, yellow (vitellinized)
2 eggs/ovariole, yellow (vitellinized)
3 mature eggs/ovariole, yellow (vitellinized)
Harmonia axyridis In this study
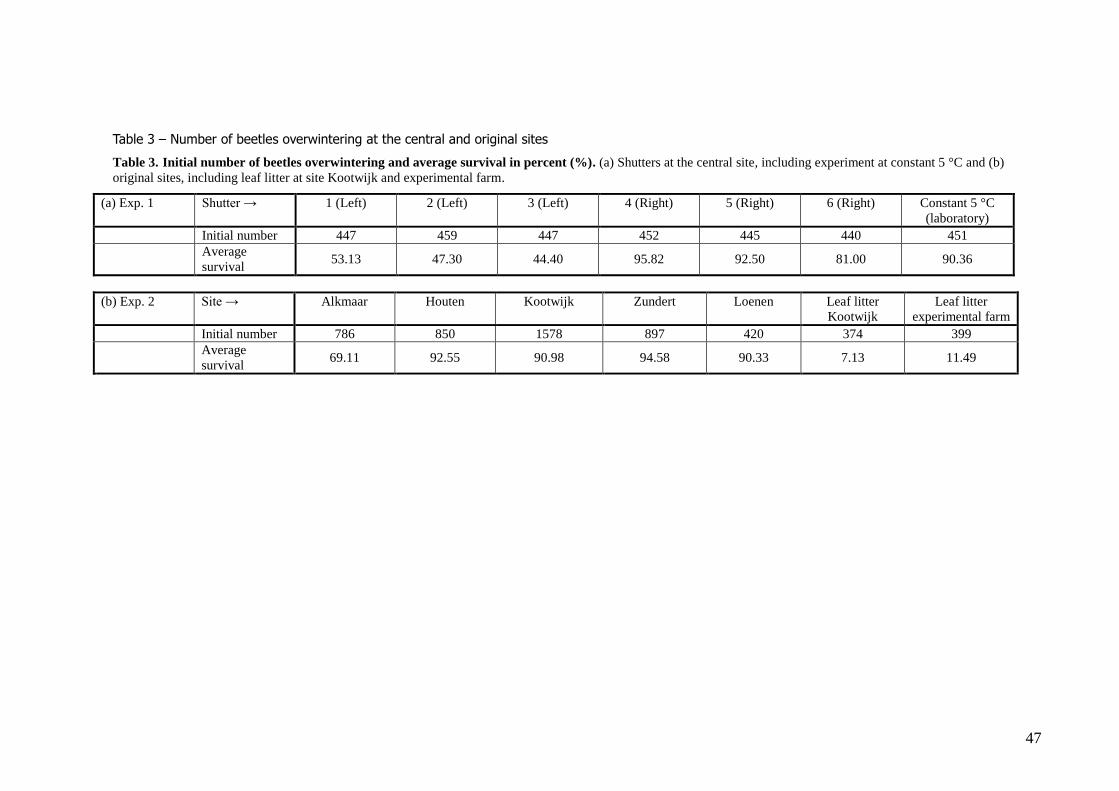
47
Table 3 – Number of beetles overwintering at the central and original sites
Table 3. Initial number of beetles overwintering and average survival in percent (%). (a) Shutters at the central site, including experiment at constant 5 °C and (b)
original sites, including leaf litter at site Kootwijk and experimental farm.
(a) Exp. 1 Shutter → 1 (Left) 2 (Left) 3 (Left) 4 (Right) 5 (Right) 6 (Right) Constant 5 °C
(laboratory)
Initial number 447 459 447 452 445 440 451
Average
survival 53.13 47.30 44.40 95.82 92.50 81.00 90.36
(b) Exp. 2 Site → Alkmaar Houten Kootwijk Zundert Loenen Leaf litter
Kootwijk
Leaf litter
experimental farm
Initial number 786 850 1578 897 420 374 399
Average
survival 69.11 92.55 90.98 94.58 90.33 7.13 11.49
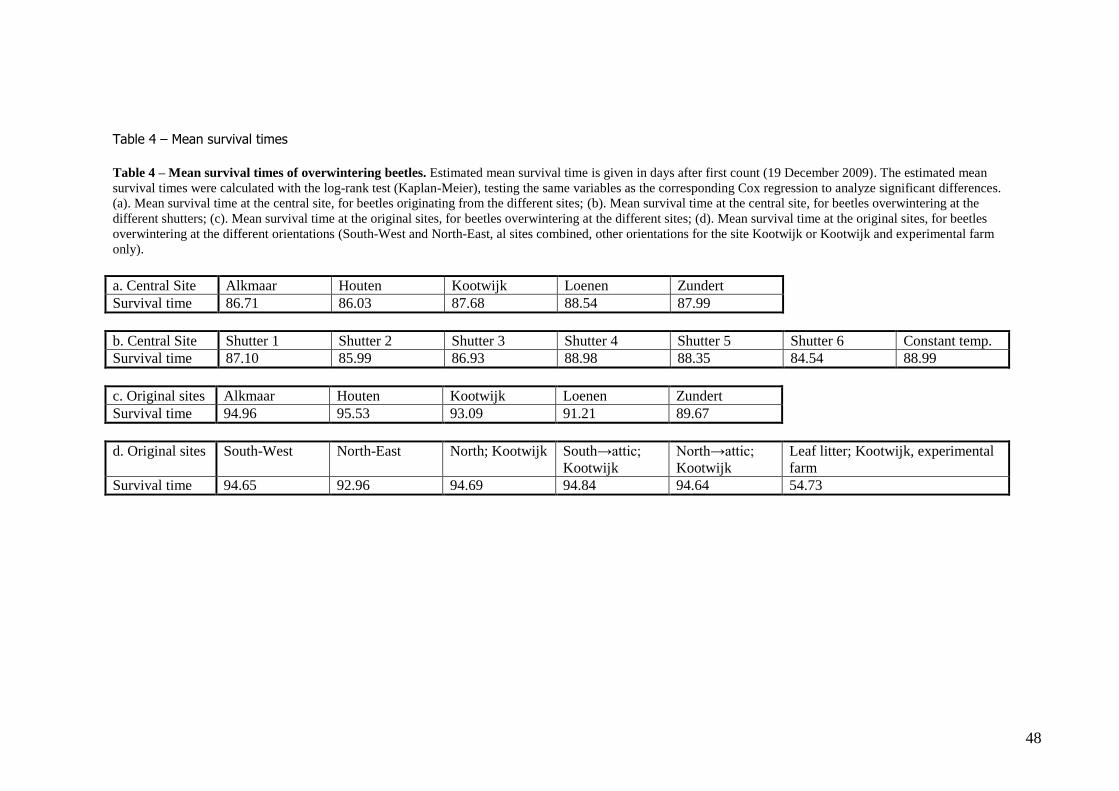
48
Table 4 – Mean survival times
Table 4 – Mean survival times of overwintering beetles. Estimated mean survival time is given in days after first count (19 December 2009). The estimated mean
survival times were calculated with the log-rank test (Kaplan-Meier), testing the same variables as the corresponding Cox regression to analyze significant differences.
(a). Mean survival time at the central site, for beetles originating from the different sites; (b). Mean survival time at the central site, for beetles overwintering at the
different shutters; (c). Mean survival time at the original sites, for beetles overwintering at the different sites; (d). Mean survival time at the original sites, for beetles
overwintering at the different orientations (South-West and North-East, al sites combined, other orientations for the site Kootwijk or Kootwijk and experimental farm
only).
a. Central Site Alkmaar Houten Kootwijk Loenen Zundert
Survival time 86.71 86.03 87.68 88.54 87.99
b. Central Site Shutter 1 Shutter 2 Shutter 3 Shutter 4 Shutter 5 Shutter 6 Constant temp.
Survival time 87.10 85.99 86.93 88.98 88.35 84.54 88.99
c. Original sites Alkmaar Houten Kootwijk Loenen Zundert
Survival time 94.96 95.53 93.09 91.21 89.67
d. Original sites South-West North-East North; Kootwijk South→attic;
Kootwijk
North→attic;
Kootwijk
Leaf litter; Kootwijk, experimental
farm
Survival time 94.65 92.96 94.69 94.84 94.64 54.73
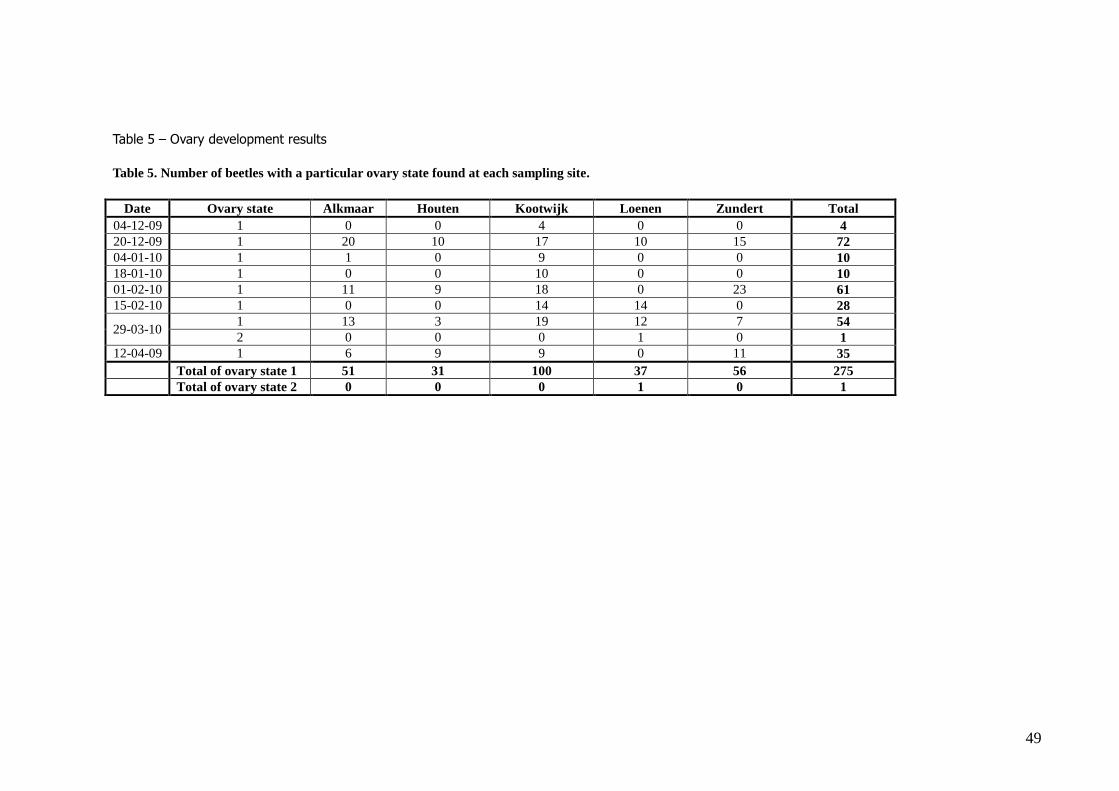
49
Table 5 – Ovary development results Table 5. Number of beetles with a particular ovary state found at each sampling site.
Date Ovary state Alkmaar Houten Kootwijk Loenen Zundert Total
04-12-09 1 0 0 4 0 0 4
20-12-09 1 20 10 17 10 15 72
04-01-10 1 1 0 9 0 0 10
18-01-10 1 0 0 10 0 0 10
01-02-10 1 11 9 18 0 23 61
15-02-10 1 0 0 14 14 0 28
29-03-10 1 13 3 19 12 7 54
2 0 0 0 1 0 1
12-04-09 1 6 9 9 0 11 35
Total of ovary state 1 51 31 100 37 56 275
Total of ovary state 2 0 0 0 1 0 1
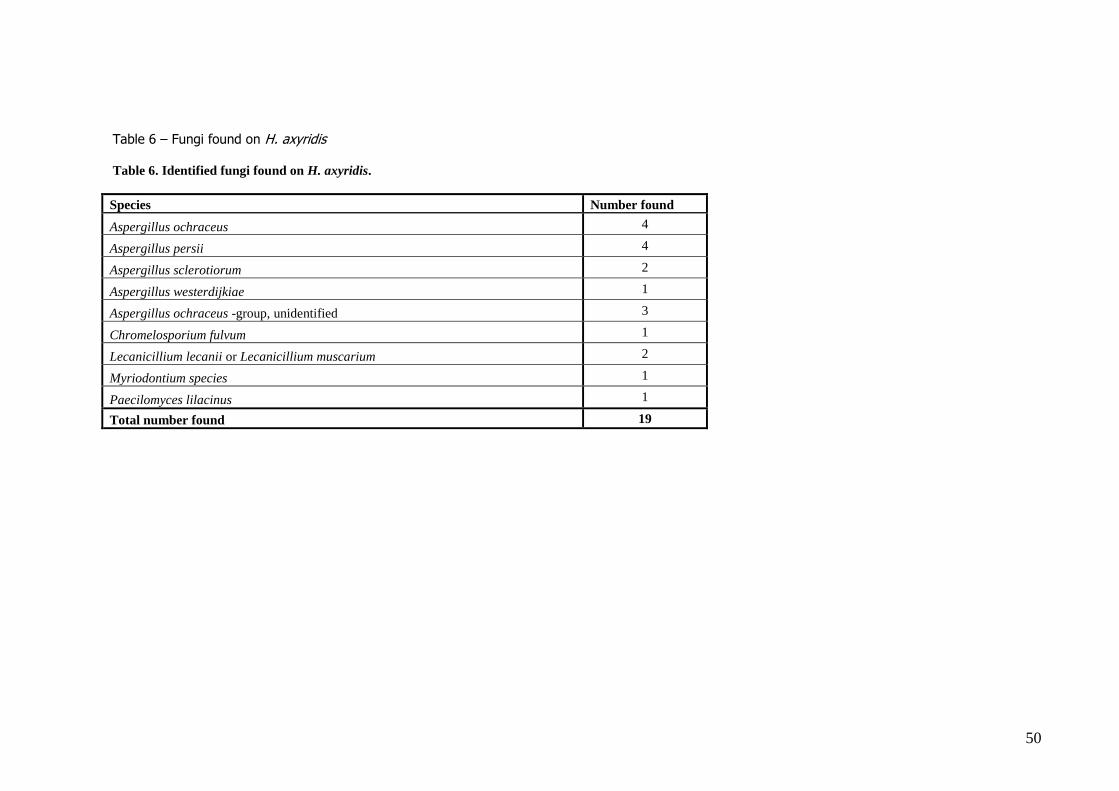
50
Table 6 – Fungi found on H. axyridis Table 6. Identified fungi found on H. axyridis.
Species Number found
Aspergillus ochraceus 4
Aspergillus persii 4
Aspergillus sclerotiorum 2
Aspergillus westerdijkiae 1
Aspergillus ochraceus -group, unidentified 3
Chromelosporium fulvum 1
Lecanicillium lecanii or Lecanicillium muscarium 2
Myriodontium species 1
Paecilomyces lilacinus 1
Total number found 19
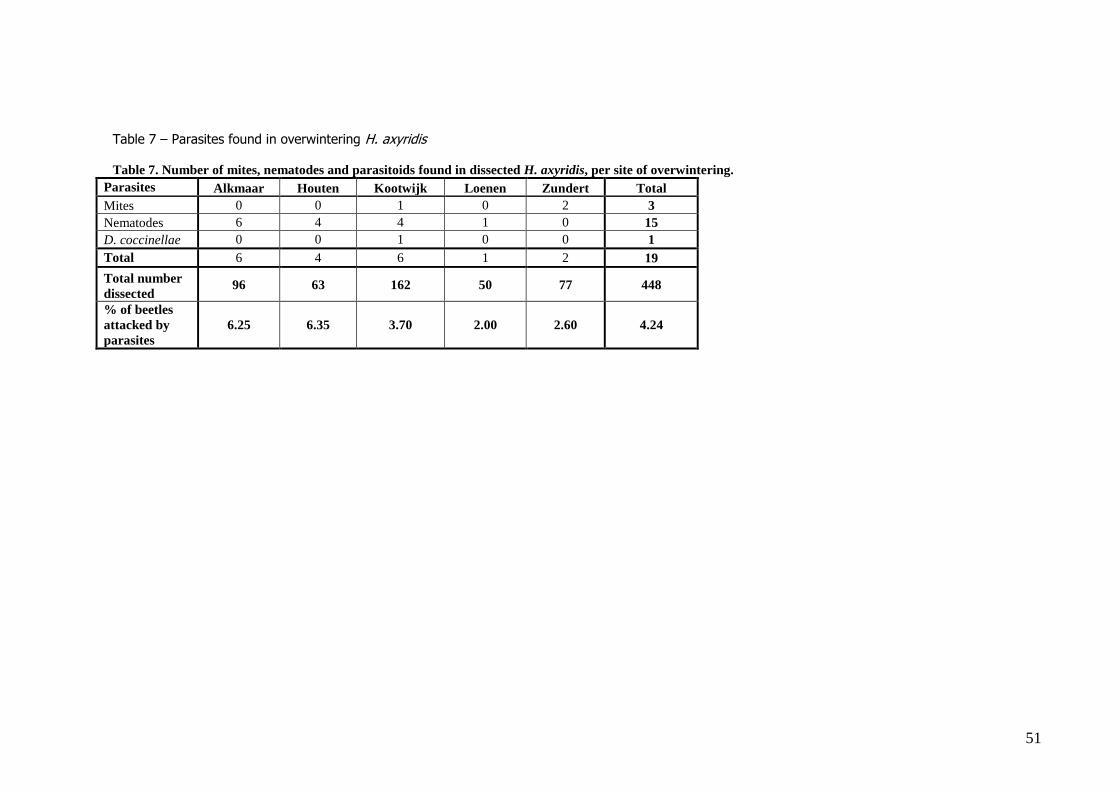
51
Table 7 – Parasites found in overwintering H. axyridis
Table 7. Number of mites, nematodes and parasitoids found in dissected H. axyridis, per site of overwintering.
Parasites Alkmaar Houten Kootwijk Loenen Zundert Total
Mites 0 0 1 0 2 3
Nematodes 6 4 4 1 0 15
D. coccinellae 0 0 1 0 0 1
Total 6 4 6 1 2 19
Total number
dissected 96 63 162 50 77 448
% of beetles
attacked by
parasites
6.25 6.35 3.70 2.00 2.60 4.24
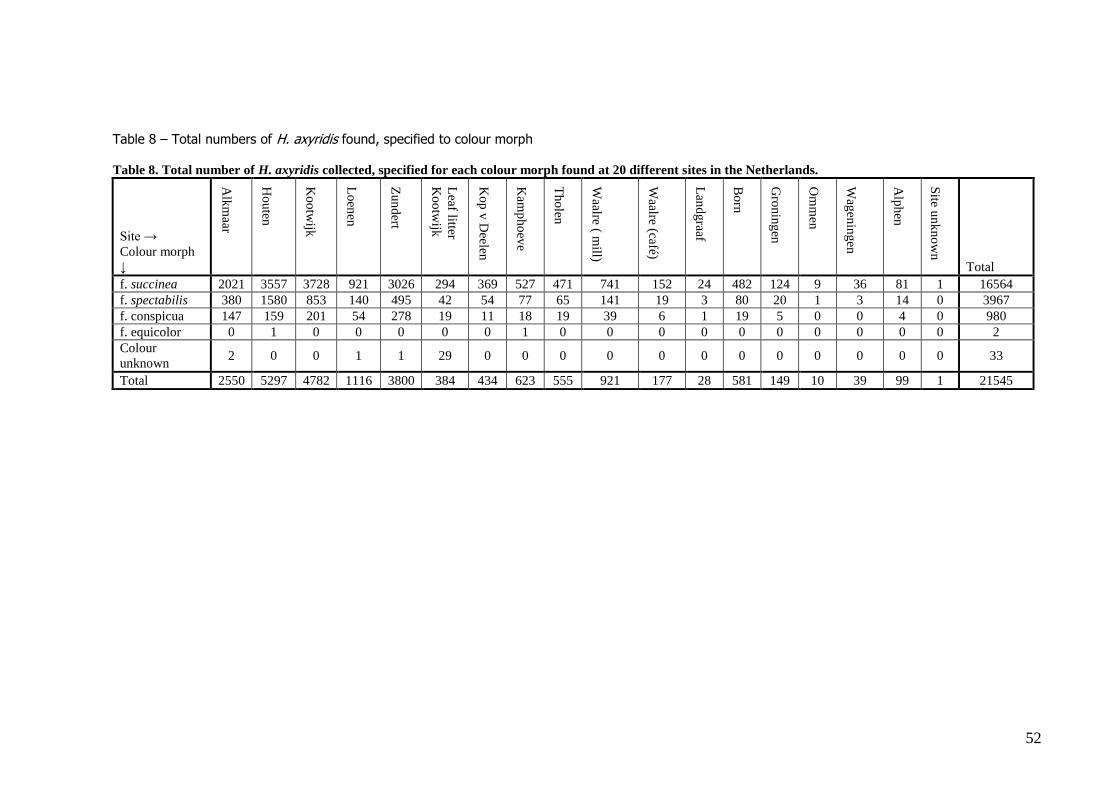
52
Table 8 – Total numbers of H. axyridis found, specified to colour morph
Table 8. Total number of H. axyridis collected, specified for each colour morph found at 20 different sites in the Netherlands.
Site →
Colour morph
↓
Alk
maar
Ho
uten
Ko
otw
ijk
Lo
enen
Zu
nd
ert
Leaf litter
Ko
otw
ijk
Ko
p v
Deelen
Kam
ph
oev
e
Th
olen
Waalre ( m
ill)
Waalre (café)
Lan
dg
raaf
Bo
rn
Gro
nin
gen
Om
men
Wag
enin
gen
Alp
hen
Site u
nk
no
wn
Total
f. succinea 2021 3557 3728 921 3026 294 369 527 471 741 152 24 482 124 9 36 81 1 16564
f. spectabilis 380 1580 853 140 495 42 54 77 65 141 19 3 80 20 1 3 14 0 3967
f. conspicua 147 159 201 54 278 19 11 18 19 39 6 1 19 5 0 0 4 0 980
f. equicolor 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 2
Colour
unknown 2 0 0 1 1 29 0 0 0 0 0 0 0 0 0 0 0 0 33
Total 2550 5297 4782 1116 3800 384 434 623 555 921 177 28 581 149 10 39 99 1 21545

53
Appendix 2 – Detailed information on monitoring survival Counting beetles For the ladybird beetles from each location, a time series was made, with 6 replicates (the 6
shutters of the experimental farm) each. Behind every shutter, one bag of each location was placed, containing about 100 beetles each. Temperature and relative humidity were measured
with a datalogger. These were placed between the bags behind two of the shutters of the
experimental farm, shutters 2 and 5. Dataloggers measured every 30 minutes during whole winter period.
Every 2 weeks, from 19 December 2009 until 18 March 2010, the beetles in the bags of each location, from each shutter were counted in the following way:
The bags were counted in random order, in a cold chamber (4 ° C) to prevent disturbance by
a higher temperature. Every bag was opened and carefully turned upside-down, to allow dead beetles to fall down. Dead beetles are supposed to be unable to hold themselves to the gauze.
When these beetles, in addition to some alive individuals who also accidentally felt down, do not walk away immediately, but cannot yet be identified as dead, they were scored as
'potentially dead'. Potentially dead beetles were temporarily placed at room temperature, and checked for mortality shortly after (within an hour). Antennae movement and position of the
legs (close to the body when alive, a condition called thanatosis (Hodek et al., 1996)) are
signs that the beetles are alive after all. These alive beetles were then immediately placed back into the bag, while dead beetles were scored and collected in Eppendorf tubes for further
experiments. The colour morph and, later, gender of these beetles was scored. All live beetles remaining in the bags (and those fallen out accidentally) were counted when
still in the bags. The bags thus counted were replaced behind the shutters afterwards, and
recounted again next time. All beetles were counted on the same day as far as possible, so no variance in time occurred.

54
Appendix 3 – Detailed information on the end of overwintering End of overwintering The last time beetles were counted, all remaining beetles (dead and alive) were counted,
sexed and colour morph was determined. In addition to this, all „potentially dead‟ beetles were kept at room temperature in small cups (approximately 10 cm high) overnight, according to
the methodology of Labrie et al. (2008). The beetles that walked out of the cups or were able
to do so, were scored as being alive. Those that were alive, but could hardly move, were scored as being “half alive”. These would not be able to reach feeding areas in natural
conditions, and thus were considered to not have survived the winter period. Dead beetles were scored as such (but not used for experiment 4, fungus infection, any more).
To prevent the beetles from dying by another cause than „normal‟ overwintering (for example
due to warming by sunshine in early spring), the overwintering experiment was stopped when beetles normally would move away from their hibernation spot, or search for another place to
hibernate. This date was determined by placing 6 additional bags with 50 ladybird beetles each, in large cages (23x25x40cm), with the difference that these bags were left open. Three
cages with each one opened bag were placed outside in a shaded place, to resemble a more exposed hibernation spot. Three additional cages were likewise placed near the entrance of an
unheated, dark shed, to resemble a less exposed hibernation spot. When beetles became
active, they walked out of the bags (but still remained inside the larger cage). The overwintering experiments were stopped when 25% or more beetles had moved out of the
bags.





























