Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOoY, June 1982, p. 1266-12730021-9193/82/061266-08$02.00/0
Vol. 150, No. 3
Occurrence and Properties of Composite Transposon Tn2672:Evolution of Multiple Drug Resistance Transposons
CHRISTINE HANNI, JURG MEYER, SHIGERU IIDA, AND WERNER ARBER*
Department ofMicrobiology, Biozentrum, University ofBasel, CH40S6 Basel, Switzerland
Received 28 September 1981/Accepted 8 January 1982
We found Tn2671 (the 23-kb long ISl-flanked r-determinant of NR1-Basel)inserted into the ampicillin resistance gene bla of the Tn3-related transposonTn9O2. The resulting 28-kilobase-long composite transposon Tn2672 (= Tn9O2bla::Tn2671) is stable, and it translocates as a unit into various loci including IS)of the resistance transfer factor of R100-1. These results are discussed withrespect to the evolution of R plasmids providing multiple drug resistance.
Transposons carried by resistance plasmidsoften contain several drug resistance determi-nants. For example, the ISI-flanked r-deter-minant (r-det) of RI determines resistance tochloramphenicol (Cm), fusidic acid (Far), strep-tomycin (Sm'), sulfonamide (Su'), ampicillin(Ap'), and kanamycin (Kmi) and contains theApr transposon Tn3, the Apr Smr Sur transposonTn4, and the Kmr transposon Tn2350 (Fig. 1; 6,23, 24). Since terminal IS] elements can poten-tially mediate the transposition of any DNAsequence (19, 20, 28, 30) the entire r-det of Rlshould also be a transposon. Similarly, the IS)-flanked r-det of NR1 (or R100) specifying Cmr,Far, Smr, Sur, and resistance to mercury (Hg') iscalled transposon Tn2670 and includes the SmrSur Hgr transposon Tn2l (Fig. 1; 14, 19a, 23, 24).By comparing the structure of these and otherrelated R plasmids, it has been speculated thatsequential acquisition of individual transposonsby a plasmid results in the formation of suchmultiple resistance transposons (8, 24).
In studies of transposition of Tn2671 (= r-detof NR1-Basel) which is a Tn2670 derivativecontaining a 1.25-kilobase (kb) IS element calledIS30 (1, 17, 19, 19a), we have isolated a bacterio-phage P7 derivative carrying Tn2671 (19a). Thesite of insertion of Tn2671 in the P7 genome wasin the bla gene for Apr, i.e., within transposonTn902 (18, 19a, 33). Here we present evidencethat Tn902 bla::Tn2671 is stable and transposesas a unit. This composite transposon is calledTn2672 (Fig. 1). Furthermore, we have observedthe transposition of Tn2672 into other transpos-able DNA elements. These results provide ex-amples for the evolution of composite transpo-sons with multiple drug resistance determinants.
MATERILS AND METHODSBacterial strains, phages, and plasmids. Table 1 lists
the bacterial strains, phages, and plasmids used. The
resistance transfer factor (RTF) carried by strainLC524 is a derivative of R100-1 carrying one copy ofIS) and TnlO (3). Both phages P7 and P7bla::r-det204carry the thermoinducible cI ts48 mutation (31). Plas-mids pHC230, 238, 240, and 241 are Apr derivatives ofpHC219, 220, 221, and 222, respectively.Media. Tris-glucose minimal (T) medium, LBMg
medium, LB agar, and Davis minimal agar containingsulfathiazol were described before (17, 25). Antibioticswere added to LB agar without glucose and CaC12 atthe following concentrations: ampicillin (150 gg/ml),chloramphenicol (25 4g/ml), HgCk2 (25 ag/ml), nalidix-ic acid (80 ,ug/ml), rifampin (80 pg/ml), streptomycin(20 ig/ml), and tetracycline (25 pg/ml).Isolatn and chra tin of RTF::Tn2672. Bac-
teria lysogenic for P7::r-det2O4 express P7 immunityand P1-specific restriction and modification. Since theP7bla::r-det204 genome is not packageable intact intoP7 phage particles, defective killer particles and fewplaque-forming phages (less than about 1% of thephage particles) are produced upon heat induction(19a).LC524 cells grown in TG medium with appropriate
supplement were treated with CaCl2 and infected withP7bla::r-det2O4 particles at a multiplicity of about 10-4plaque-forming phages per cell as described previously(17). Cmr transductants were collected and mated withstrain CH102 as described before (15, 19a), and CmrRifr transconjugants were isolated. From these, CmrSur Hgr Tcr colonies were screened for absence of P7immunity, Pl-specific restriction and modification,and production of killer particles (17). These Cmr SurHgr Tcr RifT transcorjugants were further mated withstrain CH101, and Cmr Smr Sur Hgl Tcr Nal' colonieswere isolated. From the resulting cells, plasmids wereisolated and subjected to restriction cleavage (2) andelectron microscopic analysis.
Electron mcroscopy. Plasmid DNA was alkali dena-tured and after neutralization was immediately mount-ed for electron microscopy by the formamide-cyto-chrome c technique of Davis et al. (10). PM2 DNA andfd DNA were added as length standards for double-and single-stranded DNA, respectively. However, thesnap-back structure of TnlO (11, 22) within RTFprovided another, intramolecular length reference.
1266
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

EVOLUTION OF COMPOSITE TRANSPOSONS 1267
Slarcm Sm Su Hg ISlb_- hT21
Tn 2670-
IShCm Sm Su
Ap _,Cm SrSum
Si5Ap n
ISibKm n
-Tn3- -Tn2350OTn#
ISIb IHg rLAP'
-.------- Tn21 :: 1S304n902-
-Th 2671 (Tn2670::1530)-- th272 (Tn902::Tn2671)
FIG. 1. Schematic representation oftransposons found in r-det of R100, RI,det2O4. The r-det of R100 and RI carry twedirectly repeated ISIs, respectively (7, 14)mediated transposons Tn2350, Tn2670,were described by Clerget et al. (6) and(19a). Tn2671 is the r-det of NR1-BaselIS30 (1, 19a). Tn3, Tn4, and Tn2l in thedescribed by Kopecko and co-workers (23,is a relative to Tn3 and was found in phage I
Tn9O2 bla::Tn2671 found in P7::r-det204scribed by lida et al. (19a). Ap' and Ap"indicate the split parts of the Apr gene.
Electron micrographs were taken with a Fi301 microscope at 40 kV and a magnificatiorDNA molecules were measured from 10:negatives by using a Numonics digitizer.
RI1ORESULTS
Transposition of Tu9O2 bla::Tn2671 from P7 toRTF. Recently, we described the integration ofTn2671 into the bla gene of Tn9O2 carried in the
RI phage P7 genome (Fig. 1; 19a). Here we presentevidence that the composite structure Tn9O2bla::Tn2671 is able to translocate from the P7derivative P7::r-det2O4 to plasmid RTF.
P7::r-d.t2Og Since Tn9O2 bla::Tn2671 is Ap5, we first usedthe markers carried by Tn2671 to detect thetransposition of Tn9O2 bla::Tn2671. BecauseP7::r-det2O4 is an oversized specialized trans-
composite ducing phage for Cmr, Smr, Sur, and Hgr, theand P7::r- majority ofCmr transductants obtained by infec-and three tion with P7::r-det2O4 at low multiplicity of
I. The ISI- infection was expected to carry the transposed r-nd Tn2671 det segment (17; 19a). Cmr transductants ap-
lida et al. peared with a frequency of about 2 per plaque-containing forming phage when the LC524 recipient cells24). Tr9e2 carrying RTF were infected with P7::r-det2O4 at4(18T33) a multiplicity of 10-4 PFU per cell. To distin-
4 was de- guish a Cmr marker on the RTF plasmid fromin Tn2672 one transposed to the host chromosome or still
in the P7 prophage, Cmr transductants werecollected and mated with the Rifr strain CH102.About half of the Cmr RifT transconjugants
?hilips EM showed no P7 immunity, no Pl-specific restric-Iof 10,000. tion and modification, no production of killerx-enaged particles upon induction and had a Cmr Sur Hgr
Tcr phenotype. Furthermore, these drug resist-
TABLE 1. Bacterial strains, phages, and plasmidsa
D~esignation Relevant characters Reference orGenotype Phenotype source
Escherichia coli K-12LC524 leu thy A(lac-proB) (RTF) L. CaroCH101 thr met leu hsdK nal Derived from
WA921 (32)CH102 rpo str P1I Derived from
W3110Phages
P7clts48 Tn9O2 Apr (31)P7bla::r-det2O4c1ts48 Tn9O2bla::Tn2671 = Tn2672 Ap5 Cmr Smr Sur Hgr (19a)
PlasmidspHC219 RTF::Tn2672 Tcr Ap8 Cmr Smr Sur Hgr This studypHC220 RTF::Tn2672 Tcr Ap' Cmr Smr Sur He This studypHC221 RTF::Tn2672 (ISl::Tn2672) Tcr Ap8 Cmr Smr Sur Hgr This studypHC222 RTF::Tn2672 Tcr Ap5 Cmr Smr Sur Hgr This studypHC230 RTF::Tn9O2 Tcr Apr, derived from pHC219 This studypHC238 RTF::Tn9O2 Tcr Apr, derived from pHC220 This studypHC240 RTF::Tn9O2 Tcr Apr, derived from pHC221 This studypHC241 RTF::Tn9O2 Tcr Apr, derived from pHC222 This studypHC257 RTFtetA::cat' aad+ sul+ mer+ Tc' Ap5 Cmr Smr Sur HgT This studypHC258 RTF::Tn2672 tet Tc' Ap5 Cmr Smr Sur HgT This studya Phage P7 carries the Apr transposon Tn9O2 (18, 33). P7bla::r-det2O4 is P7bla::Tn2671, which carries the
Tn2671 inserted within the bla gene of Tn9O2.
VOL. 150, 1982
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

1268 HANNI ET AL.
ance determinants could be transferred togetherto the Nalr strain CH101. The resulting Cmr SmrSur Hgr Tcr Nair transconjugants yielded AprCms Sms Su' Hg5 Tcr Nagr colonies with afrequency of 2 x 10-10 to 9 x 10-10. This couldbe explained by precise excision of the r-det(Tn2671) from the Tn9O2 bla::Tn2671. This char-acterization suggested that the Cmr Smr Sur HgrTcr Nair transconjugants were derivatives ofstrain CH101 carrying the plasmid RTF::Tn9O2bla::Tn2671. This was the first evidence thatTn9O2 bla::Tn2671 is a composite transposon,and we call it Tn2672.
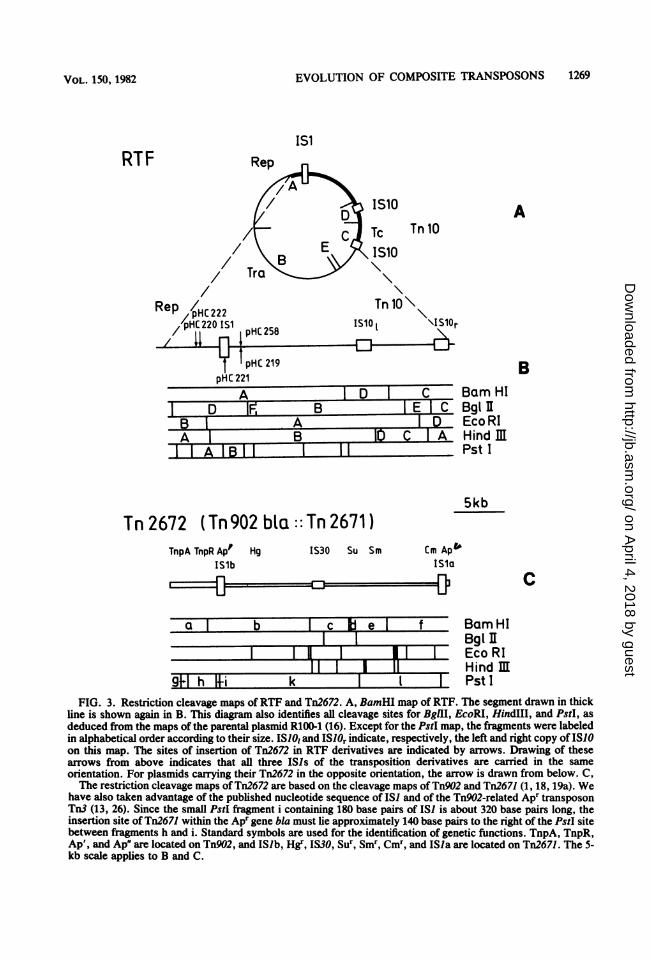
Localizion of Tn2672 on four RTF plasumidderivatives. To physically verify the transposi-tion of Tn2672 as a unit from the P7 genome toRTF, four independent RTF::Tn2672 deriva-tives, plasmids pHC219, 220, 221, and 222, weresubjected to restriction cleavage analysis. AsFig. 2 shows, the four internal BamHI fragmentsb, c, d and e, derived from Tn2672 (Fig. 3C),appeared in all four plasmids. This, and inparticular the presence ofthe BamHI fragment b(unction between Tn9O2 and Tn2671) confirmedthat Tn2672 transposed into RTF as a unit. Thedata in Fig. 2 indicate that the Tn2672 in all fourplasmids examined is inserted within BamHIfiagmentA ofRTF. The sites and the orientationof Tn2672 were more precisely mapped by re-striction cleavage analysis with BglII, EcoPU,and HindIll (data not shown). The results of thisanalysis were confirmed by PstI cleavage of theRTF::Tn9O2 plasmids pHC230, 238, 240, and241 (Fig. 2) which had been derived fromRTF::Tn2672 by precise excision of Tn2671.These restriction cleavage analyses revealedthat all four plasmids examined contain Tn2672in a relatively narrow region of RTF and thatpHC221 carries the insertion in its IS) (Fig. 3B).Furthermore, the two IS) elements of the r-det(Tn2671) carried by pHC219 and pHC221 areopposite in orientation to that of RTF, whereaspHC220 and pHC222 carry all three IS) ele-ments in the same orientation.To independently determine the site of inser-
tion of Tn2672 within the IS) of pHC221, puri-fied pHC221 DNA was denatured, rapidly rena-tured, and observed in the electron microscopefor intramolecular renatured regions (snapbacks). Taking the orientation ofTn2672 relativeto the resident IS) into account, it was expectedthat besides TnlO two other snap backs shouldappear; the size of the two stems should add upto the length of IS) (0.77 kb; 26), and the addedsize of the two loops should correspond to thatof Tn9O2 plus one IS) element (5.7 kb; (19a).The distance between these two structuresshould be equivalent to the r-det minus two ISls(approximately 22 kb; 19a), and the distance of
1 2 3 4 5 6 7 8 9
-b
-C-e
A-h-
B-
d~~~~FIG. 2. Electrophoresis patterns of plasmids after
treatment with restriction endonucleases. Slots 1through 4 show firgments produced by BamHI cleav-age of various RTF::Tn2672 plasmids. Electrophoresiswas carried out in 0.8% aarose. Capital and smallletters identify restriction cleavage fragments derivedfrom RTF and Tn2672, respectively, as illustrated inFig. 3B and C. Slots: 1, pHC220; 2, pHC219; 3,pHC222; 4, pHC221. The largest fragment A fromRTF disappeared due to the Tn2672 insertion. The twojunction fragments containing part of the RTF andTn2672 segments were not labeled, since their sizevaries from one plasmid to the other. In slot 1 bothjunction fragments migrated roughly to the same posi-tion below B. In slot 2 one junction fragment migratedwith B, and the other migrated between B and C. Slots5 through 9 show fiagments produced by PstI cleavageofvarious RTF::Tn9O2 derivatives from RTF::Tn2672.Electrophoresis was carried out in 1.3% agarose.Slots: 5, RTF; 6, pHC238 derived from pHC220; 7,pHC241 derived from pHC222; 8, pHC240 derivedfrom pHC221; 9, pHC230 derived from pHC219. Capi-tal letters A and B identify the two restriction cleavagefiagments ofRTF containing IS) sequences (Fig. 3B).Fragment h was the largest PstI fiagment derived fromthe internal part of Tn9O2. The same fiagment alsoappeared from Tn2672 (Fig. 3C). The arrows indicatethe larer junction fragments containing part of RTFand Tn2672 segments. The other junction fiagmentsare either too small to be seen or overlap with anotherfragnent.
one of the snap backs from TnlO should amountto 14.4 kb (14). In fact, such a structure waseasily detected, and the measurements corre-spond to the lengths predicted (Fig. 4A and B).The insertion site of Tn2672 within IS) wasmapped 0.57 kb from its left-hand end. This is ingood agreement with the result from the restric-tion cleavage analysis. Similarly, the site ofinsertion and orientation of Tn2672 in pHC219
J. BACTERIOL.
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
EVOLUTION OF COMPOSITE TRANSPOSONS 1269
IS1RTF Rep
/Z A 1I10 A
TC TnlO
/ Tra
Rep 'HC222 Tn 10\
/p 20S1pHC258 1510o \IslOr
-C *~~~~~~~-pHC 219 B
pHC 221
A IDl C Bam HID F. I E C Bgl I
B I -EcoRI
A B C A Hind mAIB Pst I
5kb
Tn 2672 (Tn 902 bla:: Tn 2671)TnpA TnpR App Hg IS30 Su Sm Cm Ap6*
ISlb IS1Q
,* O 9 ~~~~C
a b c IJ e I f BamHIBglII
III I Pt EcoRI_I I 4V~ Hind m
9 h i k Pst IFIG. 3. Restriction cleavage maps ofRTF and Tn2672. A, BamHI map of RTF. The segment drawn in thick
line is shown again in B. This diagram also identifies all cleavage sites for BglIH, EcoRI, HindIll, and PstI, asdeduced from the maps of the parental plasmid R100-1 (16). Except for the PstI map, the fiagments were labeledin alphabetical order according to their size. ISIO, and IS1Or indicate, respectively, the left and right copy of ISI0on this map. The sites of insertion of Tn2672 in RTF derivatives are indicated by arrows. Drawing of thesearrows from above indicates that all three ISIs of the transposition derivatives are carried in the sameorientation. For plasmids carrying their Tn2672 in the opposite orientation, the arrow is drawn from below. C,The restriction cleavage maps of Tn2672 are based on the cleavage maps of Tn902 and Tn2671 (1, 18, 19a). We
have also taken advantage of the published nucleotide sequence of IS) and of the Tn902-related Apr transposonTn3 (13, 26). Since the small PstI fragment i containing 180 base pairs of IS] is about 320 base pairs long, theinsertion site of Tn2671 within the Apr gene bla must lie approximately 140 base pairs to the right of the PstI sitebetween fragments h and i. Standard symbols are used for the identification of genetic functions. TnpA, TnpR,Ap', and Ap' are located on Tn902, and ISlb, Hgr, IS30, Sur, Smr, Cmr, and ISla are located on Tn2671. The 5-kb scale applies to B and C.
VOL. 150, 1982
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

1270 HANNI ET AL.
was confirmed by electron microscopic analysisof snap-back structures formed by annealing ofthe inverted copies of IS] (data not shown).
Insertion of drug resistance markers into andthe restructuring of TnlO on RTF. We have alsolooked for an RTF derivative carrying the trans-poson inserted within the Tcr gene of TnlO. ApsCmr Smr Sur Hgr Tc' transconjugants were iso-lated in the same way as described for Ap5 CmrSmr Sur Hgr Tcr transconjugants. Among those,two independently isolated clones which carriedpHC257 and pHC258 were analyzed in detail.Strain CHIOI(pHC258) gave Cmr Smr Sur HgtTcr revertants with a frequency of 10-9, suggest-ing that the inactivation of the Tcr gene was notdue to the insertion of Tn2672. Indeed, restric-tion cleavage analysis and electron microscopyrevealed that Tn2672 was inserted near the IS]ofRTF (Fig. 3) and that no significant structuralalteration was observed in the TnlO region ofpHC258. Thus, pHC258 appears to carry a spon-taneous point mutation in the Tcr gene.
Neither Tcr nor Apr revertants (<1011) werefound from strain CH1O1(pHC257). Restrictioncleavage analysis ofpHC257 indicated that dele-tion of part of TnlO had occurred after insertionof either Tn2671 or Tn2672 (in the latter case,the tnpA and tnpR part of Tn902 was alsodeleted) into the region common to fragmentsBgllI-E and EcoRI-D (Fig. 3B). The presence ofa deletion was also indicated by the snap-back-structure observed in single-stranded pHC257DNA (Fig. 4C). Although the ISIO stem ap-peared intact, the size of the loop measured 23.7kb (Fig. 4D), which is less than expected forTnlO::Tn2671 or TnlO::Tn2672, but which corre-sponds to the value expected from the restric-tion analysis. We therefore interpret the geno-type of pHC257 to be RTFtetA::cat' aad+ sul+mer+ (Table 1). The deletion formation mighthave been mediated either by ISlb or by IS10, asdefined in Fig. 3.
DISCUSSIONWe have demonstrated that the composite
transposon Tn2672, which consists of an ISI-flanked r-det (Tn2671) inserted into the bla geneof the Tn3-related transposon Tn902, translo-cates as a unit from the P7 genome to a transfer-able R plasmid. Tn3 is known to contain the
three genes tnpA, tnpR, and bla of which onlytnpA and tnpR are involved in the transpositionprocess (5, 12, 13). Since the structure of Tn902resembles that of Tn3, (18, 33), an insertion intothe bla gene is not expected to inactivate theTn902 transposition functions. We thereforeconclude that Tn902 transposition functions arepreserved in Tn2672 and promote also the trans-position of the composite transposon Tn2672. Inour selection of transposition products only theCmr marker was involved. Therefore, not onlythe transposition of Tn2672 but also that ofTn2671 (r-det) could have been detected. How-ever, five of six RTF derivatives examinedwhich had acquired the Cmr Smr Sur He mark-ers carried an intact Tn2672. This indicates thattransposition of Tn2672 occurs more frequentlythan that of Tn2671.Tn2672 does not confer Apr, but it contains a
split bla gene which can be reconstituted byprecise excision ofTn2671. Therefore, via trans-position of Tn2672 a silent bla gene can spreadto other replicons and later become reactivated.We observed that the composite transposon
Tn2672 (or perhaps Tn2671) inserted into twoother transposable elements, TnlO and IS). Wedo not know yet whether these composite struc-tures are able to transpose. The transposition ofTnlO is mediated by the flanking ISIO (4, 11).Since the two copies of ISIO appear to haveremained intact, the TnlO tetA::cat' aad+ sul+mer+ probably still behaves as a transposon.This deletion derivative could illustrate yet an-other aspect of r-det evolution. Provided thatTn2672 had transposed, the deletion of Tn902sequences abolishes its transposition functionsand thus stabilizes the insertion within TnlO(A).
Similarly, IS) derivatives having inserted atthe unique PstI site either a Kmr fragment orpBR322 were shown to transpose as a unit (21,29). It would be interesting to know whether theISl::Tn2672 also behaves as a transposon.Translocation of this composite structure mightalso be brought about by recA-mediated recipro-cal recombination between either part of thesplit IS) and an IS) resident on the targetreplicon, by a translocation mechanism dis-cussed by lida et al. (19, 21).
Previously we have demonstrated that a DNAsequence could become an ISI-flanked transpo-
FIG. 4. Snap-back structures observd in RTF::Tn2672 plasmids. A, Electron micrograph ofa single-strandedpHC221 molecule showing thme snap-back structures: one representing TnlO and two formed by the IS)elements flanking the r-det annealing to the split resident IS) ofRTF (arrows). B, Interpretation of the structureshown in A. Length measurements (in kb) of the various regions are based on 16 molecules. Tn9O2 sequences areindicated by a wavy line. C, Electron micrograph of single-stranded pHC257 DNA documenting the snap-backstructure formed by ISIO and the increase in loop size of TnlO. D, Length measurements (in kb) ofthe snap-backstructure shown in C, based on 13 molecules. They are interpreted to indicate integration of Tn2671 or Tn2672within the unique part of TnlO and deletion of a DNA segment.
J. BACTERIOL.
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 150, 1982 EVOLUTION OF COMPOSITE TRANSPOSONS 1271
) -
iK-jK
tu.--s Zlm sV. -
4.N
± 0.0
48.31.6 0. 40~~0.2300 Il ..2±01
21.8t0.8 RTF ~ ~ 237 ±0.B D~~~~r-e
pHC221~~~~~~ K~~~~.23t00
$'j,:~~~p-~*~-~ ,.44..,4... 21. 03Cs 2 01
TnlO 0.5A*"'
1.35 044.123LL7.0'7
4-., k. 7Z *1 iml*&4j. IIA4~L,
MN4,,4.4.44- '4. 44" .
!R.L.'--.''4.4io
'4.,.~~~~.4
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

1272 HANNI ET AL.
son (20). ISl-flanked transposons might evolveby duplication of IS] in transpositional cointe-gration or inversion and subsequent ISl-mediat-ed deletion (9, 19, 20, 27). We speculated thatnot only IS) but also other IS elements couldproduce transposons in a similar manner, andthat these newly generated transposons couldeither transpose into another unrelated transpo-son or serve as a target for different transposons(20). Either of these processes can result in anew composite transposon. Here we describeexamples for such processes and for the furtherevolution of composite transposons: stable trans-location of a composite transposon, its integra-tion into yet other transposable elements, pre-cise excision of one component, and deletionformation within the composite transposon. Fur-ther restructuring might occur which is mediatedby transposable elements within or outside thecomposite transposon. The stability of newlyformed composite transposons depends on therelative activities of the participating transpo-sons and IS elements as well as on the efficiencyof reciprocal recombination acting on homolo-gous sequences. These mechanisms must cer-tainly be involved in the genesis and subsequentdecay of multiple drug resistance r-det on Rplasmids.
ACKNOWLEDGMENSThis work was supported by the Swiss National Science
Foundation grant 3.479-0.79.
LIMRATURE CITED1. Arber, W., S. ida, H. Jane, P. Casper, J. Meyer, and C.
HIAni. 1978. Rearangements of genetic material in Esch-erichia col as observed on the bacteriophage P1 plasmid.Cold Spring Harbor Symp. Quant. Biol. 43:1197-1208.
2. Bikhl, B., ad W. Arer. 1977. Physical mapping of Bgmll,BamHI, EcoRI, HindII and PstI restriction fragments ofbacteriophage P1 DNA. Mol. Gen. Genet. 153:311-324.
3. Chandler, M., B. Alet, E. Gaflay, E. Boy de la Tour, andL. Caro. 1977. Involvement of ISI in the dissociation ofthe r-determinant and RTF components of the plasmidR100-1. Mol. Gen. Genet. 153:289-295.
4. Chander, M.,E. IRlet, L. Siver, E. Boy de Ia Tour, andL. Caro. 1979. TnIlO mediated integration of the plasmidR100-1 in the bacterial chromosome: inverse transposi-tion. Mol. Gen. Genet. 173:23-30.
5. Chon, J., P. G. Leaum, M. J. C db, ad S. H.Cohen. 1979. Transposition protein of Tn3: identificationand characterization of an essential repressor-controlledgene product. Nature (London) 282:801-806.
6. Clrget, M., M. Chandler, and L. Caro. 1980. Isolton ofan ISI flanked kanamycin resistance transposon fromRldrdl9. Mol. Gen. Genet. 13:123-127.
7. Clerget, M., M. Chandler, ad L. Caro. 1981. The struc-ture of Rldrdl9: A revised physical map of the plasmid.Mol. Gen. Genet. 181:183-191.
8. Cohb, S. N., J. Brevet, F. Cabello, A. C. Y. Cbag, J.Chon, D. J. Kopecko, P. J. KreCc_m, P. NI, andK.Tlunns. 1978. Macro- and microevolution of bacterialplasmids, p. 217-220. In D. Schlessinger (ed.), Microbiol-ogy-1978. American Society for Microbiology,Washing-ton, D.C.
9. Cornels, G., and H. Saedler. 1980. Deletions and an
inversion induced by a resident IS) of the lactose transpo-son Tn9SI. Mol. Gen. Genet. 178:367-374.
10. Davis, R. W., M. Shion, and N. Davido. 1971. Electronmicroscope heteroduplex methods for mapping regions ofbase sequence homology in nucleic acids. Methods Enzy-mol. 21:413-428.
11. Foster, T. J., M. A. Davis, D. E. Roberts, K. Takesh_ts,and N. Kleckner. 1981. Genetic organizaton of transpo-son TnJO. Cell 23:201-213.
12. Gill, R. E., F. Hdhn, and S. Falkow. 1979. Identificationof the protein encoded by the transposable element Tn3which is required for its transposition. Nature (London)282:797-01.
13. Hdebo, F., B. J. McCarthy, H. Obtabo, and E. Obtabo.1979. DNA sequence analysis of the tasposon Tn3:three genes and three sites involved in transposition ofTn3. Cell 18:1153-1163.
14. Ru, S., E. Obtabo, N. Davidson, ad H. Saedler. 1975.Electron microscope heteroduplex studies of sequencereltions among bacterial plasmids: identification andmapping of the insertion sequence IS1 and IS2 in F and Rplasmids. J. Bacteriol. 122:764-775.
15. lIda, S. 1977. Directed integration of an F' plasmid byintegative suppression. Isolation of plaque forming lamb-da transducing phage for dnaC gene. Mol. Gen. Genet.15S:153-162.
16. lida, S. 1980. A cointegrate of the bacteriophage P1genome and the conjugative R plasmid R100. Plasmid3:278-290.
17. lId., S., and W. Arber. 1977. Plaque forming specializedtransducing phage P1: isolation of PlCmSmSu, a precur-sor of PlCm. Mol. Gen. Genet. 153:259-269.
18. Iida, S., and W. Arber. 1979. Multiple physical differencesin the genome structure of functionally related bacterio-phages P1 and P7. Mol Gen. Genet. 173:249-261.
19. Ilda, S., and W. Amber. 1980. On the role of ISI in theformation of hybrids between the bacteriophage P1 andthe R plasmid NR1. Mol. Gen. Genet. 177:261-270.
19a.Ida, S., C. Hani, C. Echartl, and W. Arber. 1981. Is theISI flanked r-determinant of the R plasmid NR1 atranspo-son? J. Gen. Microbiol. 126:413-425.
20. ISd, S., J. Meyer, nd W. Arber. 1980. Genesis andnatural history of IS-mediated transposons. Cold SpringHarbor Symp. Quant. Biol. 45:27-37.
21. lida, S., J. Meyer, and W. Arber. 1981. Cointegratesbetween bacteriophage P1 DNA and plasmid pBR322derivatives suggest molecular mechanisms for P1-mediat-ed transduction of small plasmid. Mol. Gen. Genet. 184:1-10.
22. K1ckn, N., R. K. Cha, B. K. Tye, and D. Botedn.1975. Mutagenesis by insertion of a drug-resistance ele-ment carrying an inverted repetition. J. Mol. Biol. 97:561-575.
23. Kopecko, D. J. 1980. Involvement of specialized recombi-nation in the evolution and expression of bacterial ge-nomes, p. 165-205. In C. Stuttard and K. R. Rozee (ed.),Plasmids and transposons, enviroamental effects andmaintenance mechanisms. Academic Press, Inc., NewYork.
24. Kopecko, D. J., J. Brevet, and S. N. Cdon. 1976. Involve-ment of multiple translocating DNA segments and recom-binational hotspots in the structural evolution of bacterialplasmids. J. Mol. Biol. 18:333-360.
25. MIse, K., andW. Arber. 1976. Plaque-forming transducingbacteriophage P1 derivatives and their behavior in lyso-genic conditions. Virology 69:191-205.
26. Obtsubo, H., and E. Ohtsubo. 1978. Nucleotide sequenceof an insertion element IS]. Proc. Natl. Acad. Sci. U.S.A.75:615-619.
27. Obeubo, E., M. Zen7-a, and H. Obbubo. 1980. Plas-mids containing insertion elements are potential transpo-sons. Proc. Natl. Acad. Sci. U.S.A. 77:750-754.
28. ReIf, H.J.1980. Genetic evidence for absence of transpo-sition functions from the internal part of Tn981, a relativeof Tn9. Mol. Gen. Genet. 177:667-674.
J. BACTrERIOL.
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

EVOLUTION OF COMPOSITE TRANSPOSONS 1273
29. Reif, H. J., and W. Arber. 1980. Analysis of transpositionof ISI-Kan and its relatives. Cold Spring Harbor Symp.Quant. Biol. 45:40-43.
30. Roaer, J. L., and M. S. Guyer. 1980. Transposition ofIS1-XBIO-ISJ from a bacteriophage A derivative carryingthe ISI-cat-ISl transposon (Tn9). Mol. Gen. Genet.178:111-120.
31. Wandem_an, C., and M. Yarmoinsky. 1977. Bipartitecontrol of immunity conferred by the related heteroim-
mune plasmid prophages, P1 and P7 (formerly + Amp).Virology 77:386400.
32. Wood, W. B. 1966. Host specificity ofDNA produced byEscherichia coli: bacterial mutations affecting the restric-tion and modification of DNA. J. Mol. Biol. 16:118-133.
33. Yon, T., and D. Vapuek. 1977. Electron microscopicanalysis of bacteriophages P1, PlCm and P7. Determina-tion ofgenome sizes, sequence homology, and location ofantibiotic-resistance determinants. Virology 77:376-385.
VOL. 150, 1982
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from





























