Embed Size (px)
Citation preview

Thesis for the degree of Doctor of Medicine
Muscarinic Receptors in the Urinary Bladder
The role of the urothelium regarding cholinergic
and nitrergic effects in inflammation
Michael Andersson
2010
Department of Pharmacology
Institute of Neuroscience and Physiology
The Sahlgrenska Academy at the University of Gothenburg
Sweden

Printed by Intellecta Infolog AB, Gothenburg, Sweden, 2010
Previously published papers were reproduced with permission
from the publishers
ISBN 978-91-628-8198-6

Till min familj

4
Abstract
Inflammation alters the functional properties of the urinary bladder.
Interstitial cystitis (IC) is a chronic inflammatory syndrome in man
that is characterized by urgency, frequency and visceral pain. The
overall aim of this study was to investigate how the rat urinary
bladder is affected by inflammation, and what specific part the
urothelium plays in this.
Methods: Cystitis was induced in rats by a single injection of
cyclophosphamide (CYP; 100 mg/kg). This treatment causes a
disease state which is highly comparable to IC. Data comparing the
properties of the healthy and inflamed bladder were gathered from
(1) contraction experiments in vitro in an organ bath setup, (2)
cystometrical studies in vivo in anaesthetized rats and (3) wake,
freely moving rats in a metabolism cage. Cell cultures were also
cultivated in order to investigate if proliferation of urothelial cells is
influenced by receptor activation.
Key findings: Induction of cystitis by CYP altered the cholinergic
response of the urinary bladder. In vitro studies showed a
significantly lower response to carbachol in the inflamed bladder.
Both in vitro and in vivo, the altered cholinergic response could be
normalized by either removal of the urothelium, blockade of nitric
oxide (NO) synthase or blockade of muscarinic M1/M3/M5
receptors. These findings indicate that during CYP-induced cystitis,
NO is released from the urothelium upon muscarinic receptor
activation. Further characterization in vitro revealed the M5 receptor
as the most likely candidate for mediating this release.
In vivo experiments in the metabolism cage showed that micturition
parameters are affected by CYP-induced cystitis. Increasing doses of
a muscarinic antagonist eliminated these differences, and a
connection between the effects of antimuscarinic and antinitrergic

5
drugs was indicated. These findings underline the importance of
muscarinic receptors and NO in the alterations seen during cystitis.
Proliferation experiments indicated that adrenergic, but not
muscarinic, nicotinic or EGF receptors, are involved in the regulation
of urothelial cell proliferation.
Conclusions: In CYP-induced cystitis in the rat, the urothelium
exerts an inhibitory influence on the cholinergic response of the
urinary bladder. We conclude that this is caused by the release of NO
upon activation of urothelial muscarinic M5 receptors.
Keywords: urinary bladder, inflammation, cyclophosphamide-induced
cystitis, muscarinic receptor, urothelium, nitric oxide, M5 receptor, rat,
proliferation, micturition

6
Populärvetenskaplig sammanfattning
Urinblåsan utgör en central del av urinvägarna. Dess uppgifter består
av lagring och tömning av urin. Så länge urinblåsan fungerar som den
ska är man omedveten om dess existens, men när den krånglar kan
man inte låta bli att tänka på den. I Sverige lider runt 400.000
personer av inkontinens, och 4,5% av all diagnostiserad cancer har
sitt ursprung i urinblåsan. Flera av de behandlingar som finns idag är
otillräckliga, vilket visar på behovet av mer kunskap om urinblåsan
och dess sjukdomar. I denna avhandling presenteras ny kunskap om
de förändringar som sker i urinblåsan vid inflammation.
Att urinblåsan blir inflammerad är mycket vanligt och kan bero på
flera orsaker. Till exempel kan inflammation i urinblåsan, s.k. cystit,
orsakas av en bakterieinfektion eller närvaro av kemiska ämnen i
urinen som irriterar urinblåsans insida till den grad att en
inflammation uppstår. Urinblåsans insida täcks av en typ av celler
som kallas urotelceller. Dessa trodde man ända fram till för 10-15 år
sedan var passiva celler med ett enda syfte: att skydda urinblåsan
och resten av kroppen från toxiska ämnen i urinen. Senare forskning
har dock visat att dessa celler har flera viktiga uppgifter, inte minst
vid diverse sjukdomstillstånd. Interstitiell cystit (IC) är ett av dessa
sjukdomstillstånd, och kännetecknas av kronisk (långvarig)
inflammation, smärta, överaktivitet samt förtjockning av urinblåsan.
Kunskaper om vad som orsakar IC är begränsade, men man vet att IC
inte orsakas av en infektion. Flera studier har visat att urinblåsans
funktionalitet förändras vid cystit. Det övergripande målet med
denna avhandling var att förstå vilka skillnader som uppstår vid
inflammation i urinblåsan, och hur dessa påverkar urinblåsans
funktion. Fokus lades till stor del på urotelceller och hur dessa
förändras vid cystit. Känt sedan tidigare var att urotelceller har
förmågan att frisätta kvävemonoxid (NO). Avhandlingsarbetet
syftade till att klargöra hur denna frisättning påverkas vid cystit, och
vad NO-frisättningen har för påverkan på urinblåsan. Det var också

7
känt sedan tidigare att en speciell sorts receptor, den muskarina M5
receptorn, uttrycks mer i urotelcellerna vid cystit. Vi ville ta reda på
om detta ökade receptoruttryck påverkar urinblåsans funktion.
För att framkalla ett tillstånd som efterliknar IC användes en
etablerad metod i vilken råttor injicerades med substansen
cyklofosfamid (CYP). Denna substans används inom sjukvården för
att behandla vissa former av cancer och svåra varianter av multipel
skleros (MS). En känd biverkning av denna behandling är
uppkomsten av cystit, och två dygn efter injektion med CYP
uppvisade 100% av försöksdjuren inflammation i urinblåsan. Denna
cystit är på flera sätt mycket lik den som finns hos patienter med IC.
Under studierna noterades konsekvent att lagret av urotelceller var
tjockare i inflammerade urinblåsor. Detta ledde till ett intresse att
undersöka hur celldelningen av urotelceller är reglerad. Specifikt
ville vi undersöka om det fanns någon receptor som var extra viktig
för denna celldelning. Dessa undersökningar gjordes på både friska
urotelceller och cancerurotelceller från människa.
Våra djurförsök visade att inflammation (cystit) orsakad av CYP
påverkar urinblåsans förmåga att kontrahera. Eftersom blockad av
frisättningen av NO från urotelceller vid cystit konsekvent
förbättrade urinblåsans förmåga att kontrahera drar vi slutsatsen att
NO utövar en dämpande effekt på funktionen i den inflammerade
urinblåsan. En minskad frisättning av NO var möjlig att åstadkomma
genom att antingen blockera bildningen av NO, blockera muskarina
receptorer eller helt enkelt ta bort urotelcellerna. Resultaten från
dessa studier bekräftade därmed att NO frisätts från urotelceller och
visade att denna frisättning kunde minskas genom att blockera
muskarina receptorer. Utifrån experimenten konstaterade vi att
denna muskarina receptor med ganska hög sannolikhet är av typen
M5. Om våra resultat är giltiga även för människa kan det innebära
en möjlighet att behandla den överaktivitet som uppstår hos den
inflammerade urinblåsan med ett läkemedel som selektivt påverkar

8
den muskarina M5 receptorn, och därigenom påverkar frisättningen
av NO.
Våra försök på urotelceller indikerade att s.k. adrenoceptorer är
viktiga för regleringen av celldelning. Vi kunde se att aktivering av en
typ av adrenoceptor (β) ledde till ökad celldelning, medan aktivering
av en annan typ (α) ledde till minskad celldelning. Kolinerg
stimulering, t.ex. via muskarina receptorer, tycktes inte påverka
celldelningen.
En intressant fråga som ännu inte besvarats är exakt vilken roll (eller
roller) NO spelar under utvecklingen av cystit. Det kan vara så att NO
enbart är en produkt av inflammationen, eller så är NO en av de
molekyler som driver på utvecklingen av inflammationen. Det kan till
och med vara så att det är en kombination av båda dessa. Om NO är
med och driver på utvecklingen av inflammationen skulle det
teoretiskt sett vara möjligt att förhindra uppkomsten av cystit genom
att hämma bildningen av NO. Detta skulle visserligen kräva att man
utvecklar ett test för att upptäcka inflammationen när den är i sin
linda, men skulle potentiellt sett kunna minska lidandet för
miljontals människor världen över.
Sammanfattningsvis visar denna avhandling att råttans urinblåsa
förändras vid inflammation. Betydelsen av NO frisatt från
urotelceller ökar, och detta NO minskar urinblåsans förmåga att
kontrahera. Frisättningen sker, åtminstone till viss del, som svar på
aktivering av muskarina receptorer i urotelcellerna. Dessa
muskarina receptorer är med stor sannolikhet av typen M5. Framtida
studier bör göras för att avgöra om våra fynd är tillämpbara även på
människa. Framtida experiment bör också göras för att undersöka
betydelsen av NO vid utvecklingen av cystit.

9
List of Publications
This thesis is based on the following papers, which are referred to by
their Roman numerals in the text:
I. Andersson MC., G. Tobin & D. Giglio, 2007
Cholinergic nitric oxide release from the urinary bladder
mucosa in cyclophosphamide-induced cystitis of the
anaesthetized rat
Br J Pharm 2008 Apr;153(7):1438-44. Epub 2008 Feb 4
II. Andersson M., P. Aronsson, D. Doufish, A. Lampert &
G. Tobin
The muscarinic M5 receptor is the primary mediator of
urothelium-derived nitric oxide effects in the rat urinary
bladder
Manuscript
III. Andersson M., P. Aronsson, D. Giglio, A. Wilhelmson, P.
Jeřábek & G. Tobin, 2010
Pharmacological modulation of the micturition pattern in
normal and cyclophosphamide-pretreated conscious rats
Auton Neurosci 2010. Epub 2010 Sep 17
IV. Andersson M., P. Aronsson, D. Eskandari, D. Giglio & G. Tobin
Characterization of receptor-mediated proliferation in the
human bladder urothelial UROtsa and T24 cell line
Manuscript

10
Table of contents
ABSTRACT .............................................................................................................. 4
POPULÄRVETENSKAPLIG SAMMANFATTNING ........................................................ 6
LIST OF PUBLICATIONS ........................................................................................... 9
TABLE OF CONTENTS ............................................................................................ 10
LIST OF ABBREVIATIONS ....................................................................................... 12
INTRODUCTION .................................................................................................... 14
ANATOMY OF THE URINARY BLADDER ............................................................................. 14
Urothelial morphology ...................................................................................... 16
NORMAL BLADDER FUNCTION ....................................................................................... 17
The efferent nervous system ............................................................................. 20
The afferent nervous system ............................................................................. 25
The urothelium .................................................................................................. 26
Nitric oxide ........................................................................................................ 28
URINARY TRACT DISORDERS – DISEASES AND TREATMENTS ................................................. 29
OAB - Overactive bladder.................................................................................. 29
Acute and chronic incontinence ........................................................................ 30
Interstitial cystitis .............................................................................................. 31
Urinary bladder cancer ..................................................................................... 32
AIMS .................................................................................................................... 34
SPECIFIC AIMS ........................................................................................................... 34
METHODS AND MATERIALS .................................................................................. 35
CYCLOPHOSPHAMIDE-INDUCED CYSTITIS ......................................................................... 35
REMOVAL OF UROTHELIUM .......................................................................................... 35
CYSTOMETRICAL STUDIES ............................................................................................. 36
IN VITRO FUNCTIONAL STUDIES – THE ORGAN BATH ........................................................... 37
IN VIVO FUNCTIONAL STUDIES – THE METABOLISM CAGE .................................................... 38
CELL CULTIVATION ...................................................................................................... 39
MEASUREMENT OF PROLIFERATION ............................................................................... 39
MATERIALS ............................................................................................................... 40
Drugs ................................................................................................................. 40
Solutions and reagents ..................................................................................... 41

11
STATISTICAL CALCULATIONS .......................................................................................... 41
Paper I ............................................................................................................... 41
Paper II .............................................................................................................. 42
Paper III ............................................................................................................. 42
Paper IV............................................................................................................. 42
RESULTS AND DISCUSSION ................................................................................... 44
PAPER I .................................................................................................................... 44
PAPER II ................................................................................................................... 48
PAPER III .................................................................................................................. 53
PAPER IV .................................................................................................................. 58
CONCLUDING REMARKS ....................................................................................... 63
ACKNOWLEDGEMENTS ......................................................................................... 67
REFERENCES ......................................................................................................... 69

12
List of Abbreviations
4-DAMP 4-diphenylacetoxy-N-methylpiperidine
methobromide
AC adenylate cyclase
ACh acetylcholine
ANOVA analysis of variance
APF anti-proliferative factor
ATP adenosine 5’-triphosphate
BPH benign prostatic hyperplasia
BPS bladder pain syndrome
Ca2+ calcium
cAMP cyclic adenosine monophosphate
CYP cyclophosphamide
DAG diacylglycerol
DMEM Dulbecco´s modified Eagle medium
DO detrusor overactivity
EGF epidermal growth factor
eNOS endothelial nitric oxide synthase
HB-EGF heparin-binding epidermal growth factor-like
growth factor
IC interstitial cystitis
IL-6 interleukin-6
IP3 inositol triphosphate
L-748,337 N-[[3-[(2S)-2-Hydroxy-3-[[2-[4-
[(phenylsulfonyl)amino]phenyl]ethyl]amino]
propoxy]phenyl]methyl]-acetamide
L-NAME Nω-nitro-L-arginine methyl ester hydrochloride
L-NNA Nω-nitro-L-arginine
LUTD lower urinary tract dysfunctions
MeCh methacholine
MS multiple schlerosis
NA noradrenaline
NANC non-adrenergic, non-cholinergic

13
NO nitric oxide
NOS nitric oxide synthase
OAB overactive bladder
PBS phosphate buffered saline
PEST penicillin/streptomycin
pFHHSiD p-fluoro-hexahydro-sila-diphenidol
hydrochloride
PIP2 phosphatidylinositol 4,5-bisphosphate
PLC phospholipase C
PPADS pyridoxalphosphate-6-azophenyl-2’,4’-disulfonic
acid
SR59230A 3-(2-Ethylphenoxy)-1-[[(1S)-1,2,3,4-
tetrahydronaphth-1-yl]amino]-(2S)-2-propanol
oxalate salt
UTP uridine 5’-triphosphate

14
Introduction
Imagine life without the urinary bladder. Day in and day out it
enables us to store and dispose of numerous waste products filtered
from the blood. Without it, the urinary system would be
dysfunctional. For most people, the actions of the urinary bladder
passes unnoticed, until it malfunctions. In Sweden more than
400 000 people have daily symptoms of urinary incontinence, and
annually approximately 4.5% of all diagnosed cancer has its origin in
the urinary bladder. Many of the treatments for diseases affecting the
urinary bladder are suboptimal, rendering the need for greater
knowledge, in order to be able to generate improved treatments.
Anatomy of the urinary bladder
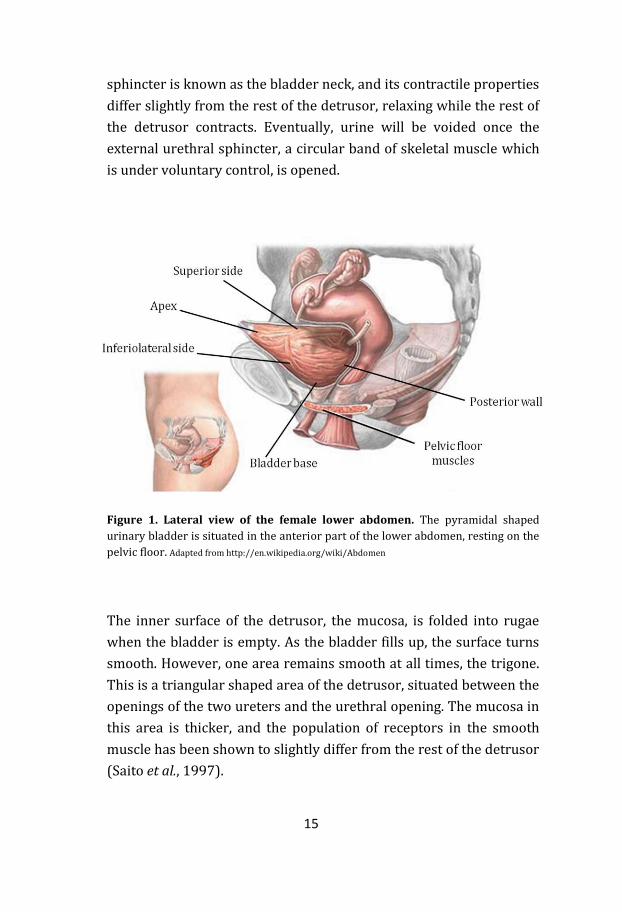
The urinary bladder is a hollow muscular organ situated in the
anterior part of the lower abdomen, resting on the pelvic floor
(Figure 1). Its basic functions are to store and void urine. The
maximum capacity of the urinary bladder varies interindividually,
but various sources say somewhere between 0.5 – 1 L in the healthy
bladder. Urine produced in the kidneys enters the bladder via the
two ureters. The entrances for these are situated on each side of the
posterior wall of the bladder, also known as the bladder base. The
empty bladder has a pyramidal shape, giving it, apart from the
posterior base, a superior side, two inferiolateral sides and an apex
(Figure 1).
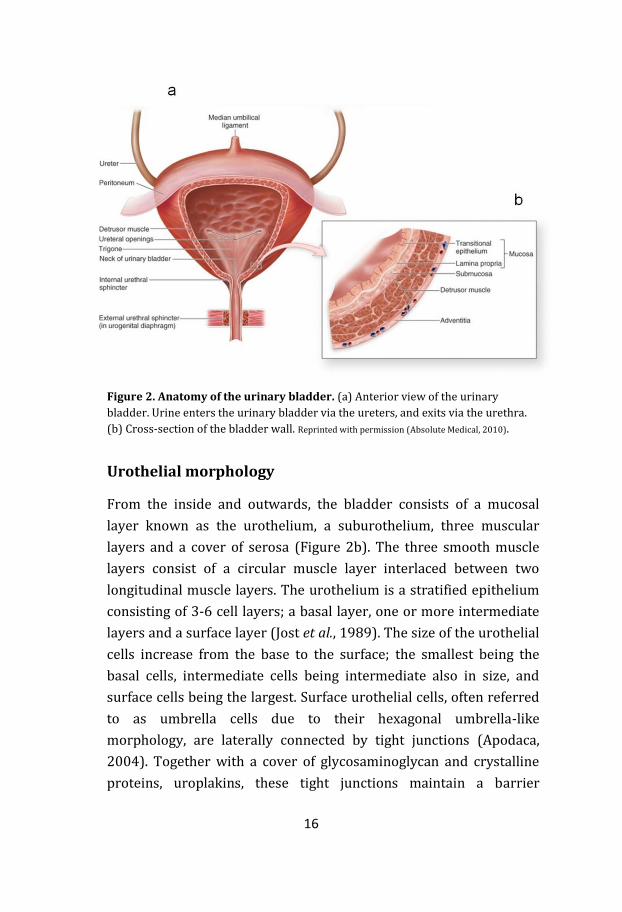
Voiding is performed by contraction of the large smooth muscular
layer known as the detrusor. Urine thereby exits the bladder via the
urethra, which is situated in the most inferior part of the bladder
(Figure 2a). The involuntarily controlled internal sphincter keeps the
urine from leaking into the urethra and remains closed until the
detrusor contracts and the pressure inside the bladder, the
intravesical pressure, rises. The area surrounding the internal
15
sphincter is known as the bladder neck, and its contractile properties
differ slightly from the rest of the detrusor, relaxing while the rest of
the detrusor contracts. Eventually, urine will be voided once the
external urethral sphincter, a circular band of skeletal muscle which
is under voluntary control, is opened.
Figure 1. Lateral view of the female lower abdomen. The pyramidal shaped
urinary bladder is situated in the anterior part of the lower abdomen, resting on the
pelvic floor. Adapted from http://en.wikipedia.org/wiki/Abdomen
The inner surface of the detrusor, the mucosa, is folded into rugae
when the bladder is empty. As the bladder fills up, the surface turns
smooth. However, one area remains smooth at all times, the trigone.
This is a triangular shaped area of the detrusor, situated between the
openings of the two ureters and the urethral opening. The mucosa in
this area is thicker, and the population of receptors in the smooth
muscle has been shown to slightly differ from the rest of the detrusor
(Saito et al., 1997).
16
Figure 2. Anatomy of the urinary bladder. (a) Anterior view of the urinary
bladder. Urine enters the urinary bladder via the ureters, and exits via the urethra.
(b) Cross-section of the bladder wall. Reprinted with permission (Absolute Medical, 2010).
Urothelial morphology
From the inside and outwards, the bladder consists of a mucosal
layer known as the urothelium, a suburothelium, three muscular
layers and a cover of serosa (Figure 2b). The three smooth muscle
layers consist of a circular muscle layer interlaced between two
longitudinal muscle layers. The urothelium is a stratified epithelium
consisting of 3-6 cell layers; a basal layer, one or more intermediate
layers and a surface layer (Jost et al., 1989). The size of the urothelial
cells increase from the base to the surface; the smallest being the
basal cells, intermediate cells being intermediate also in size, and
surface cells being the largest. Surface urothelial cells, often referred
to as umbrella cells due to their hexagonal umbrella-like
morphology, are laterally connected by tight junctions (Apodaca,
2004). Together with a cover of glycosaminoglycan and crystalline
proteins, uroplakins, these tight junctions maintain a barrier

17
between the urine and the bloodstream (Parsons et al., 1990; Hu et
al., 2002; Born et al., 2003). Immediately beneath the urothelium one
finds a thin basal lamina, separating the urothelium from the lamina
propria. The lamina propria is today often referred to as part of the
suburothelium. The suburothelium consists of connective tissue
(bundles of collagen and elastic fibers), muscular bundles, afferent
and efferent nerve fibers and a numerous amount of capillaries.
Thus, the urothelium is not vascularized nor directly innervated by
the nervous system, but is still in close vicinity to capillaries and
nerve endings. Within the suburothelium one also finds
myofibroblasts. While the function of these cells has not been
entirely established, it has been suggested that they function much in
the same way as cells of Cajal do in the gut, modulating
neurotransmission and bladder function (Wiseman et al., 2003;
Tobin et al., 2009).
Normal bladder function
The urinary bladder has two basic functions; storage of urine and
emptying of urine. Since the production of urine is constantly active,
at all given times one of these functions are active. One often refers to
the urinary bladder being in either the filling or the emptying phase.
The sensations during the filling phase can be divided into three
distinct happenings; initial sensation of filling, first desire to void and
strong desire to void. During the filling phase, there is a concomitant
increasing contraction of the bladder outlet region (bladder neck and
urethra), and increasing relaxation of the detrusor. At the same time,
muscles and connective fibers in the pelvic floor help maintain
continence (Sampselle & DeLancey, 1998). The increasing relaxation
of the detrusor as the bladder fills up yields a situation where despite
bladder filling, the intravesical pressure remains rather constant.
However, the bladder cannot fill up forever and eventually the

18
intravesical pressure will increase. As the bladder continues to fill up,
the balance is shifted towards decreased contraction of the bladder
outlet region, and increased contraction of the detrusor. Afferent
pressure sensing systems (stretch receptors) are activated,
rendering awareness to us that the bladder needs to be emptied.
Before we can empty the bladder the outer sphincter has to be
opened. This sphincter is via the pudendal nerve under our voluntary
control, at least in healthy individuals. Our voluntary opening of the
outer sphincter initiates the micturition reflex, causing the bladder
outlet region to relax and the detrusor to contract.
Like most organs in the human body, the urinary bladder is
innervated by both sympathetic and parasympathetic nerve fibers.
Activation of these nerve fibers leads to activation of muscarinic and
adrenergic receptors in the detrusor, respectively. The muscarinic
receptor population in the urinary bladder consists of five subtypes,
denoted M1-M5 (Caulfield & Birdsall, 1998; Giglio & Tobin, 2009).
Several studies have shown that M2 is the most abundantly
expressed subtype in the detrusor, with roughly a 3:1 ratio over the
functionally most important muscarinic receptor, the M3 receptor
(Eglen et al., 1994; Wang et al., 1995; Yamaguchi et al., 1996; Hegde
& Eglen, 1999; Sigala et al., 2002). The overall distribution of
muscarinic receptors varies throughout the urinary bladder, with a
generally lower concentration in the bladder neck (Saito et al., 1997).
Three of the muscarinic receptor subtypes, namely the M1, M3 and
M5 receptor, are Gq/11 coupled receptors whose activation induces
the hydrolysis of phosphatidylinositol 4,5-bisphosphate (PIP2) to
diacylglycerol (DAG) and inositol triphosphate (IP3) by
phospholipase C (PLC; Figure 3). Both products (DAG and IP3)
increase the amount of intracellular calcium (Ca2+). In the detrusor
this results in an increased tonus. The other two subtypes are Gi
coupled, and even if several pathways have been suggested (Bolton &
Zholos, 1997; Togashi et al., 1998), in man they predominantly
inhibit adenylate cyclase (AC) upon activation (Figure 3).

19
Figure 3. Main intracellular pathways of muscarinic and β-adrenergic
receptors in the urinary bladder. An increase of intracellular calcium causes an
increase in detrusor tonus, while an increase of intracellular cAMP causes a decrease
in detrusor tonus. The increase of intracellular calcium upon muscarinic M1/M3/M5
receptor activation is achieved through two separate mechanisms; (1) diacylglycerol
(DAG) in the cell membrane causes an influx via L-type Ca2+ channels and (2)
calcium is released from the sarcoplasmatic reticulum (SR) upon inositol
triphosphate (IP3) stimulation.
The adrenergic receptors, or adrenoceptors, which are widespread
throughout the lower urinary tract, are divided into the α- and β-
adrenoceptors. The α-adrenoceptors are further subdivided into the
α1- (α1A, α1B and α1D) and α2-(α2A, α2B and α2C)adrenoceptors. The β-
adrenoceptors are divided in to three subtypes; the β1-, β2- and β3-
adrenoceptor. All subtypes are found in the urinary bladder of both
man and rat, predominantly in the bladder body (Levin et al., 1988;
Seguchi et al., 1998). One study determined the β3 subtype to be the
most abundant in the human bladder in regard to mRNA expression
(Nomiya & Yamaguchi, 2003). Activation of β-adrenoceptors mainly
20
leads to the formation of cyclic adenosine phosphate (cAMP), and the
subsequent lowering of detrusor tonus (Figure 3).
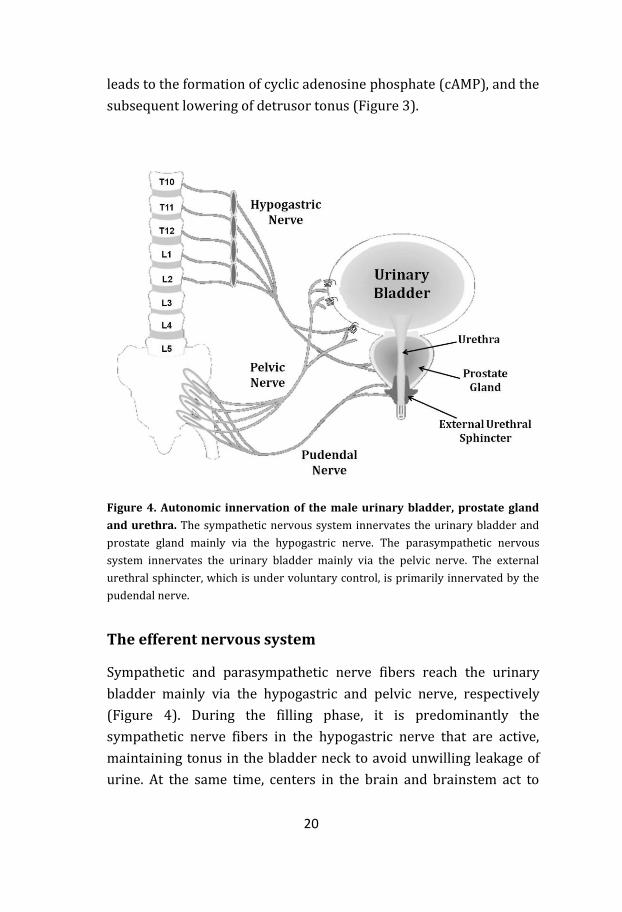
Figure 4. Autonomic innervation of the male urinary bladder, prostate gland
and urethra. The sympathetic nervous system innervates the urinary bladder and
prostate gland mainly via the hypogastric nerve. The parasympathetic nervous
system innervates the urinary bladder mainly via the pelvic nerve. The external
urethral sphincter, which is under voluntary control, is primarily innervated by the
pudendal nerve.
The efferent nervous system
Sympathetic and parasympathetic nerve fibers reach the urinary
bladder mainly via the hypogastric and pelvic nerve, respectively
(Figure 4). During the filling phase, it is predominantly the
sympathetic nerve fibers in the hypogastric nerve that are active,
maintaining tonus in the bladder neck to avoid unwilling leakage of
urine. At the same time, centers in the brain and brainstem act to

21
suppress the parasympathetic activity. When the threshold is
reached, and we have willingly opened the outer sphincter, the
balance between sympathetic and parasympathetic activation is
reversed. Then the pelvic nerve signals dominate, causing a
contraction of the detrusor and a relaxation of the bladder neck and
urethra.
During the filling phase, several receptor subtypes are involved in
the maintaining of constant intravesical pressure. Adrenergic α1-
receptors, particularly of the α1A-subtype, are dominant in the
bladder neck region, causing contraction of the smooth muscle
(Caine et al., 1975; Andersson et al., 1984; Walden et al., 1997; Chang
et al., 2000). Partly therefore, α1 antagonists are used to relieve outlet
obstruction symptoms in elderly men with benign prostatic
hyperplasia (BPH). However, α1-adrenoceptors are poorly expressed
in the rest of the detrusor (Goepel et al., 1997; Malloy et al., 1998;
Sigala et al., 2004), and their physiological relevance there seems to
be of low importance (Ueda et al., 1984; Lluel et al., 2003). Instead, β-
adrenoceptors are responsible for relaxation of the detrusor during
the filling phase (Figure 5; Elmer, 1974; Nergardh et al., 1977; Abdel-
Rahman et al., 1983). In man, it is predominantly the β3-
adrenoceptor that mediates relaxation, while in rat it is a
combination of β2- and β3-adrenoceptor activation (Oshita et al.,
1997; Igawa et al., 1998; Yamazaki et al., 1998; Igawa et al., 2001).
Studies have shown that β-adrenergic tone increases with increasing
intravesical pressure (Lecci et al., 1998) and that the nerve fibers
that mediate this relaxation are sensitive to both prostaglandins,
neurokinin A and capsaicin (Tucci et al., 2002).
Noradrenaline (NA) is the key signal substance of efferent
sympathetic innervation. Upon activation, the release of NA can be
modulated by several mechanisms, including negative feedback on
pre-junctional α2-adrenoceptors (Mattiasson et al., 1987; Somogyi &
de Groat, 1990). However, in some species such as pig, cat and
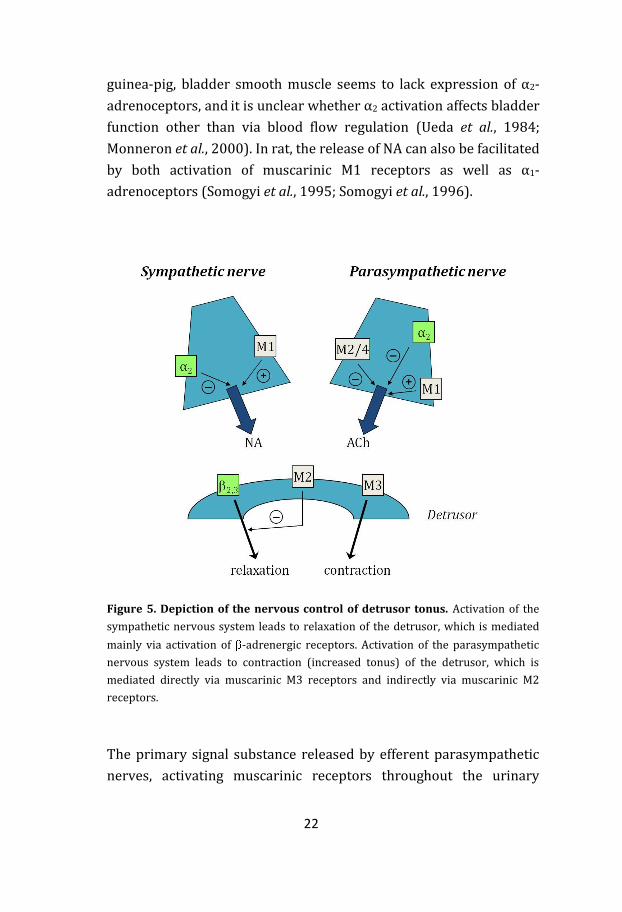
22
guinea-pig, bladder smooth muscle seems to lack expression of α2-
adrenoceptors, and it is unclear whether α2 activation affects bladder
function other than via blood flow regulation (Ueda et al., 1984;
Monneron et al., 2000). In rat, the release of NA can also be facilitated
by both activation of muscarinic M1 receptors as well as α1-
adrenoceptors (Somogyi et al., 1995; Somogyi et al., 1996).
Figure 5. Depiction of the nervous control of detrusor tonus. Activation of the
sympathetic nervous system leads to relaxation of the detrusor, which is mediated
mainly via activation of -adrenergic receptors. Activation of the parasympathetic
nervous system leads to contraction (increased tonus) of the detrusor, which is
mediated directly via muscarinic M3 receptors and indirectly via muscarinic M2
receptors.
The primary signal substance released by efferent parasympathetic
nerves, activating muscarinic receptors throughout the urinary

23
bladder, is acetylcholine (ACh). Binding of ACh to muscarinic
receptors in the detrusor causes it to contract, rendering the bladder
to empty. Even though it is the M3 receptor that is considered the
functionally most important subtype for detrusor contraction, it is
thought that there is a dual effect between the activation of M3 and
M2 receptors (Figure 5; Hegde & Eglen, 1999). In the healthy human
bladder, the M3 subtype has been shown in functional studies to be
responsible for direct contraction (Chess-Williams et al., 2001;
Fetscher et al., 2002). Second messenger studies have confirmed
these results, showing an increased formation of IP3 upon M3
activation (Noronha-Blob et al., 1989; Andersson et al., 1991). Also,
studies on M3 knock-out mice revealed a 95% reduction in bladder
contractile response to carbachol (Matsui et al., 2000).
Activation of M2 receptors has been suggested to yield an indirect
contraction by inhibiting β-adrenoceptor and purinergic receptor
mediated relaxation (Hegde et al., 1997; Giglio et al., 2001; Chess-
Williams, 2002; Giglio et al., 2005a). These findings are supported by
second messenger studies that show a decrease of cAMP upon
carbachol stimulation (Noronha-Blob et al., 1989; Harriss et al.,
1995). Even though it has never been shown in man that M2 plays a
significant role in the direct contraction of the healthy urinary
bladder, some studies indicate an increased role of M2-mediated
detrusor contraction during disease (Eglen et al., 1994; Braverman et
al., 1999).
The release of ACh from parasympathetic nerve endings can be both
inhibited and facilitated via activation of different muscarinic
receptors. Prejunctional M1 receptors have been shown to facilitate
the release of ACh in several species (Somogyi et al., 1994; Tobin &
Sjogren, 1995; Inadome et al., 1998). It has also been shown that this
facilitatory effect can increase during hyperreactive disease states
(Somogyi et al., 1998). Which prejunctional muscarinic receptor that
inhibits ACh release via negative feedback seems to vary among

24
species (for review see Somogyi et al. (1999)). Prejunctional M2/M4
receptors have been found to inhibit the release of ACh in rat and
rabbit (Tobin, 1995; Tobin & Sjogren, 1995; Braverman et al., 1998),
and later studies indicated that M4 receptors do the same in rat and
man (D'Agostino et al., 1997; D'Agostino et al., 2000). Apart from
muscarinic receptors, also the activation of α1- and α2-adrenoceptors
has been shown to facilitate and inhibit the release of ACh,
respectively (Nakamura et al., 1984; Somogyi et al., 1995; Tobin &
Sjogren, 1998; Szell et al., 2000).
In several species it has been shown that there can be other signal
substances than ACh that are involved in bladder emptying. In cat
and rat for instance, adenosine 5’-triphosphate (ATP) is co-released
together with ACh from parasympathetic nerve fibers (MacKenzie et
al., 1982; Chancellor et al., 1992; Palea et al., 1993; Theobald, 1995).
In these species ATP acts upon P2X1 receptors (Aronsson et al.,
2010), causing a fast transient contraction which possibly is the
initiator of the detrusor contraction (for review see Burnstock,
2009). The co-release of ATP makes up the most important part of
the so called atropine-resistant component of parasympathetically
induced bladder contraction, a part that hence cannot be blocked by
the non-selective muscarinic antagonist atropine. In man, even
though it has been hard to show that the atropine-resistant
component is of any great importance in the healthy bladder, ATP
can play a role in the activation of the detrusor during certain disease
states and in the aged bladder (Sjogren et al., 1982; Bayliss et al.,
1999; Yoshida et al., 2001). On the other hand, the atropine-resistant
component in the healthy bladder has been shown to be of greater
importance in both rabbit (Longhurst et al., 1984), guinea-pig
(Chesher & Thorp, 1965), cat (Theobald, 1996) and rat (Giglio et al.,
2007).

25
The afferent nervous system
Several afferent systems in the urinary bladder work together in
order to maintain optimal bladder function. Afferent, or sensory,
nerve fibers are widespread throughout the detrusor, some which
extend into the basal layer of the urothelium (Andersson, 2002a).
These nerve fibers are mainly conveyed via the pelvic nerves, but
also via the hypogastric and pudendal nerve (Andersson, 2002a). The
sensory nerves involved are both of the myelinated Aδ-fiber and
unmyelinated C-fiber type. Roughly, the Aδ-fibers convey sensation
of filling while the C-fibers mediate thermal stimulus and relay
information of chemical irritation (Janig & Morrison, 1986; Fall et al.,
1990; Habler et al., 1990). Stretch receptors in the bladder wall are
stimulated by increased intravesical pressure. These receptors
activate the Aδ afferent nerve fibers in the pelvic nerve that carry the
signal to the sacral spinal cord. The afferent signaling can be
modulated by several transmitters, including ATP (Andersson,
2002b).
Adrenergic α1A-receptors found in the urothelium have been shown
to modulate bladder afferent activity in rats, shortening the
intercontraction interval (Yanase et al., 2008). The physiological
relevance of this finding is debatable, but perhaps valid during
certain patophysiological conditions that can give increased
concentrations of NA in the urine. Several other substances have also
been shown to be able to modulate afferent activity including
neutrophins, prostaglandins and tachykinins (Yoshimura & de Groat,
1997). While prostaglandins mainly seem to act as sensitizers of
afferents, intravesically administered tachykinins are able to cause
direct contraction of the detrusor (Ishizuka et al., 1995a, b).

26
The urothelium
For a long time the urothelium was considered solely a protective
barrier, shielding the urinary bladder, as well as the rest of the body,
from the toxic and corrosive urine. But during the last decade several
studies have showed that the urothelium does much more than so.
For instance, the urothelium has been shown to respond to its
environment in regard to luminal pressure and urine composition by
releasing diffusible agents such as acetylcholine (ACh), ATP and
nitric oxide (NO) (Ferguson et al., 1997; Klapproth et al., 1997; de
Groat, 2004). To exemplify, the mechanism of stretch-induced ATP
release from the basolateral side of urothelial cells is well established
(Ferguson et al., 1997). It has also been shown that the release of
ATP can increase upon infection (Save & Persson, 2010). This
release, and that of other diffusible agents, can in turn affect afferent
nerves and smooth muscle bundles in the suburothelium, thereby
assigning the urothelium modulatory properties. Within the
suburothelium one finds nerve fibers reactive to cholinergic,
adrenergic, purinergic (P2X) and peptidergic stimuli (Jen et al.,
1995). Since receptor stimulation on some of these nerves has been
shown to evoke exocytosis, they could be both of an afferent and
paracrine/modulatory nature at the same time. Interestingly, the
expression and function of the diffusible agents from the urothelium
that can affect these nerves can be altered by urinary bladder
diseases such as cystitis (Sun et al., 2001; Giglio et al., 2005b; Smith
et al., 2005). Further, the urothelium is capable of directly
responding to various stresses, such as ischemia and urinary tract
infections (Mysorekar et al., 2002).

27
Receptor Activator Method of detection
References
Muscarinic (M1-M5)
ACh
IHC, IB RT-PCR
Giglio et al., 2005 Tyagi et al., 2006
Nicotinic (subunits alpha 3, 5, 7, 9, 10 and beta 3, 4)
ACh RT-PCR Beckel et al., 2006 Bschleipfer et al.,
2007
Purinergic (P1A1, P1A2A, P1A2B, P1A3)
adenosine IB Yu et al., 2006 Säve et al., 2009
Purinergic (P2X2,3 and P2Y1,2,4)
ATP ICC, IB, PCR RT-PCR
Chopra et al., 2008 Tempest et al., 2004
Adrenergic (α1A, α1D and β1-3)
A/NA IB, IHC RT-PCR
ISH
Ishihama et al., 2006 Otsuka et al., 2008 Walden et al., 1997
PAC1, VPAC1, VPAC2 VIP/PACAP ICC RT-PCR
Braas et al., 2006 Girard et al., 2008
EGFR EGF IHC Røtterud et al., 2005
Tachykinin (NK1) substance P RT-PCR, IB IHC
Sanchez Freire et al., 2010
Vanilloid (TRPV; VR1) pH, heat, capsaicin RT-PCR, ICC
Birder et al., 2001
Bradykinin (B2) bradykinin RT-PCR, ICC
Chopra et al., 2005
Table 1. Receptor expression in urothelial cells. ACh - acetylcholine; ATP -
adenosine 5’-triphosphate; A - adrenaline; NA - noradrenaline; VIP - vasointestinal
peptide; PACAP - pituitary adenylate cyclase activating polypeptide; EGF - epidermal
growth factor; RT-PCR - reverse transcriptase polymerase chain reaction; IHC -
immunohistochemistry; IB - immunoblotting; ICC - immunocytochemistry; ISH - in
situ hybridization.

28
All five muscarinic receptor subtypes have been shown to be present
in both human and rat urothelium (Giglio et al., 2005b; Tyagi et al.,
2006). Activation of these has been shown to lead to a number of
events, showing that they possess functional properties. For instance,
in both man and pig an unidentified relaxant factor has been shown
to be released from the urothelium upon muscarinic receptor
activation (Hawthorn et al., 2000; Chaiyaprasithi et al., 2003). Others
have shown that activation of urothelial muscarinic receptors can
lead to the release of ATP, which possibly modifies the bladders
afferent response (Kullmann et al., 2008a; Kullmann et al., 2008b;
Munoz et al., 2010). In turn, ATP and uridine 5’-triphosphate (UTP)
have been shown to yield larger contractions in intact than
urothelium-denuded strips of guinea-pig detrusor (Sui et al., 2008),
further demonstrating the presence of functional urothelial
receptors. The most likely candidates for this functional response are
of the P2X subtype, but also the presence of P1 and P2Y receptors
has been shown in the urothelium (Tempest et al., 2004; Yu et al.,
2006; Chopra et al., 2008). Apart from muscarinic and purinergic
receptors, urothelial cells express a vast array of receptors (see
Table 1).
Nitric oxide
Nitric oxide (NO) is generally accepted as one of the non-adrenergic,
non-cholinergic (NANC)-transmittors affecting the bladder (Bult et
al., 1990; Andersson & Persson, 1994). Studies using a NO scavenger
have shown that NO can be involved in normal bladder activity, and
that the removal of NO can cause a condition similar to overactive
bladder (Pandita et al., 2000). During conditions such as
cyclophosphamide (CYP)-induced cystitis, nitric oxide synthase
(NOS) has been shown to be up-regulated (Alfieri & Cubeddu, 2000;
Giglio et al., 2005b), and NO has been suggested to not only affect the
detrusor, but also act directly on afferent pathways (Ozawa et al.,
1999; Yoshimura et al., 2001).

29
NO can be released in the bladder in several ways. Studies have
shown the presence of nitric oxide containing nerve fibers in the
suburothelium, but their exact function is still unclear (Smet et al.,
1996). Others have seen release of NO from cultured urothelial cells
in response to muscarinic agonists (Kullmann et al., 2008b), and
from urinary bladder strips in response to adrenergic agonists
(Birder et al., 1998). A recent study by Munoz et al. (2010) could
show the release of NO from the urothelium upon muscarinic
receptor stimulation, but also other parts of the rat urinary bladder
could release NO, most likely the suburothelium.
Urinary tract disorders – diseases and treatments
OAB - Overactive bladder
One of many lower urinary tract dysfunctions (LUTD) is overactive
bladder (OAB), defined as urgency, with or without incontinence,
usually accompanied by increased frequency and nocturia (Abrams et
al., 2002). For a patient to be diagnosed with OAB, a concomitant
infection has to be ruled out. The prevalence of OAB is about 17%
(Milsom et al., 2001), but few prospective studies have been
performed. The cause of OAB is not well understood, but the general
risk of developing OAB increases with age. One possible cause of this
has been shown in a study by Li et al. (2003) where they found the
amount of binding sites for β-adrenoceptor agonists to decrease with
age. Since it has been shown that the most important function of β-
adrenoceptors is mediating relaxation of the detrusor during bladder
filling, it would be logical that a decrease of sympathetic activity
yields an increase in detrusor sensitivity. However, the most
widespread theory is increased afferent activity, either depending on
increased sensitivity or an increased number of afferents (Drake,
2008).

30
β3-agonists, due to their hampering of bladder contraction, have been
suggested to be a possible treatment for OAB, but to date no such
drug has been successfully developed. During the testing of
experimental β3-agonists on rats, induced cystitis and renal tubular
necrosis was reported (Waghe et al., 1999). Since the beneficial site
of action very well may be at the spinal level (Durant et al., 1988),
intrathecal administration could be a possible route of
administration, but that is something that most patients would
probably wish to avoid. However, the adverse effects seem to vary
considerably between different substances, and some promising oral
β3 agonist candidates are currently in phase II or phase III trials (for
review see Drake, 2008).
Despite the promising future, the drug treatments available today are
few. Patients are advised to manage their fluid intake and avoid
certain drinks, for instance those containing caffeine. The by far most
common drug treatment is antimuscarinic drugs, which give good
clinical effect initially, but regular long-term loss of function and
compliance problems (Andersson, 2009). Apart from the
antimuscarinic drugs and β3-adrenoceptor agonists, Ca2+-antagonists
and K+-channel openers have been suggested as possible inhibitors
of detrusor activity (Guarneri et al., 1991; Pandita et al., 2006). Also,
drugs that reduce afferent activity and/or affect NK-1 receptors at
the spinal level are under development and are currently in clinical
phase II or III trials (for review see Drake, 2008).
Acute and chronic incontinence
Urinary incontinence, or enuresis, is defined as involuntary leakage of
urine (Abrams et al., 2002). This involuntary leakage can either be a
constant dribbling or large emptying of the bladder. The large
emptying can be caused by stresses such as sneezing or coughing, or
be so called urge incontinence, a condition where the patient feels
the urge to micturate, but does not have time to reach the toilet

31
before the expel of urine. While stress incontinence is by far the most
common form in women (80%), in men urge incontinence is the
most widespread form (Hampel et al., 2004). Incontinence in any
form is a very common condition, and prevalence increases with
increasing age. It has been estimated that 250 million people
worldwide suffer from incontinence, and in Sweden the prevalence
among 80-year olds is as high as 25% (Fall, 1999; Milsom, 2009).
Treatment of acute and chronic incontinence has, apart from lifestyle
changes, focused on antimuscarinic drugs. Older drugs, such as
oxybutynin, are basically unselective, whereas the development of
new treatments has focused on finding M3-selective compounds.
However, no drug has yet been launched that does not cause the
most common adverse effects; dry mouth and gastrointestinal
disturbances. The main reason for that is surely due to the fact that
these adverse effects per se are M3 receptor-mediated (Tobin, 1995;
Barras et al., 1999). The mechanism of action of antimuscarinic drugs
was first thought to be solely by blocking efferent muscarinic
receptors in the detrusor. But further studies have shown that both
the urothelium and afferent nerve fibers are possible sites of action
(Kim et al., 2005; Abrams et al., 2006; Yamaguchi, 2010).
Other possible treatments include β agonists. One study found that in
bladder strips taken from patients with urge incontinence,
clenbuterol, a fairly β2-selective agonist, gave a significantly greater
relaxation than in bladder strips taken from patients without any
history of incontinence (Hudman et al., 2001). This finding in itself is
interesting, but as previously discussed, it is mainly the β3-
adrenoceptor that mediates relaxation in man.
Interstitial cystitis
Interstitial cystitis (IC), which is also known as bladder pain
syndrome (BPS), is a non-infectious chronic pain syndrome that is

32
characterized by painful urination, urgency, frequency, inflammation,
stiffening of the bladder wall and visceral pain. In its classic form one
finds ulcers upon cytoscopic examination, but in most cases (~80%)
the disease is ulcer-free (Koziol et al., 1996). As the symptoms are
similar to other LUTD, and the objective diagnostic criteria are few,
the epidemiology of IC is difficult to determine (Hanno, 2002).
Estimations vary between 10 per 100 000 to 70 per 100 000
(Oravisto, 1975; Curhan et al., 1999). The etiology of IC is mostly
unknown, and treatment regimens are mainly symptomatic.
However, sensitization of afferent pathways, as well as altered
release of ATP and NO, has been linked to the condition (Sun et al.,
2001; Logadottir et al., 2004). Further, the expression of muscarinic
receptors has been shown to be up-regulated, and studies have
linked the increase of certain inflammatory markers (mainly IL-6)
and changes in the levels of proliferative factors such as anti-
proliferative factor (APF), epidermal growth factor (EGF) and
heparin-binding epidermal growth factor-like growth factor
(HB-EGF) in urine to the condition (Keay et al., 1997; Erickson et al.,
2002; Sanchez Freire et al., 2010).
Urinary bladder cancer
Urinary bladder cancer accounts for 4-5% of all diagnosed cancer in
Sweden, making it the 6th most common form of cancer (Cancer
Incidence in Sweden 2007, Socialstyrelsen, 2008; IARC Globocan,
2008). Worldwide it is the 11th most common form of cancer,
accounting for about 3% of all reported cancers (IARC Globocan,
2008). However, the worldwide estimation is probably somewhat on
the small side as several developing countries, with a lower
probability of reporting, have high incidences of Bilharzia (also
known as Schistosomiasis) infections, a known cause of bladder
cancer (Elsebai, 1977; Schwartz, 1981).
The major causative agents for bladder cancer are various chemical

33
carcinogens, with different metabolites from cigarette smoking
accounting for about half of the cases (Shirai, 1993). More
particularly, these carcinogenic metabolites are thought to
accumulate in the urine, irritate certain urothelial cells to the degree
that they develop point mutations, and eventually cause them to
transform (for review see Brandt, 2009). As a result of this, most
urinary bladder cancer (~90%) has its origin in the urothelium
(Cordon-Cardo, 2008). Chemical carcinogens aside, also chronic
inflammation, caused by infections, indwelling catheters or
pharmacological treatment, has been shown to increase the risk of
developing bladder cancer (for review see Michaud, 2007). Studies
on Bilharzia, both clinical and pre-clinical, have shown a chain of
events starting with infection, continuing with chronic inflammation
and eventually leading to bladder cancer (Samaras et al., 2010). The
coupling between inflammation and cancer is well-established and
has been shown in several other tissues (Coussens & Werb, 2002;
Ohshima et al., 2003). Apart from Bilharzia infection, it has also been
shown that long-term treatment (>10 years) with CYP, generating a
state similar to chronic inflammation, increases the risk of
developing bladder cancer (Travis et al., 1995; Talar-Williams et al.,
1996).
Presently, the exact mechanisms of transformation of urothelial cells
from healthy to malignant are incompletely understood, and
therefore we today only have treatment options for already arisen
tumors. In the future we hopefully will have greater knowledge of the
transformational mechanisms, giving us the opportunity to develop
treatment strategies that prevent the origination of bladder cancer.

34
Aims
The main aim of this thesis was to investigate the effects of CYP pre-
treatment in rats, with special interest in the release of NO from the
urothelium.
Specific aims
In vivo in the anaesthetized rat (Paper I), we wondered how CYP pre-
treatment affected the muscarinic bladder contractile responses and
if NO might be released from the urothelium by cholinergic stimuli.
Our studies confirmed that the muscarinic bladder contractile
responses were altered by CYP pre-treatment, and our data indicated
that one component of this alteration was regulated by the
cholinergic release of NO from the urothelium. This led us to a study
in vitro on bladder strips (Paper II) where we wanted to characterize
the muscarinic changes involved in the alterations during CYP-
induced cystitis. This study confirmed the results found in vivo and
raised the question whether or not CYP pre-treatment altered the
micturition parameters in the conscious rat and, further, if
muscarinic, purinergic or NOS blockade could alter these changes
(Paper III). In all studies on CYP pre-treated rats, we could see
morphological changes, most prominently an increase in the
thickness of the bladder wall. Several studies have shown a coupling
between chronic CYP-induced cystitis, including bladder wall
thickening, and the development of urothelial bladder cancer. This
led us to wonder how the proliferation of urothelial cells is regulated
(Paper IV); more specifically, if the proliferation rate of healthy and
malignant urothelial cells is receptor mediated and if there are any
changes in this regulation after transformation.

35
Methods and Materials
All experiments conducted during this thesis were approved by the
local ethics committee at the University of Gothenburg. Consistently,
male rats of the Sprague-Dawley (300 – 400 g) strain were used.
Cyclophosphamide-induced cystitis
Cyclophosphamide (CYP)-induced cystitis is a common and well
documented method for inducing experimental cystitis (deVries &
Freiha, 1990). CYP is a DNA alkylating agent used in man to treat
neoplastic diseases and multiple schlerosis. Its metabolite acrolein
accumulates in the bladder lumen, where it causes severe irritation
of the urothelium, subsequently inducing haemorrhagic cystitis (Cox,
1979). To induce cystitis we have given a single intraperitoneal dose
of CYP (100 mg/kg) in combination with the analgesic
buprenophinum (Temgesic®; 10 g/kg). As previously seen in our
lab, the peak inflammatory state arises after 60 h (Giglio et al.,
2005b). Depending on which type of experiment was to be carried
out, we waited for an appropriate time before we conducted the
experiments. I.e., for cystometrical studies and in vitro functional
studies in the organ bath, experiments were carried out 60 h after
induction of cystitis. For in vivo functional studies in the metabolism
cage, in which the experiments were conducted during a 24 h period,
the experiments were started 48 h after induction of cystitis and thus
continued until 72 h post induction, comprising the 60 h peak
inflammatory time point.
Removal of urothelium
To study the direct impact of the urothelium on bladder function we
conducted both in vivo and in vitro experiments where the
urothelium was removed. In order to achieve this, we injected

36
collagenase (0.1% in Hank’s balanced salt solution) directly into the
bladder of an anaesthetized rat, either via a secured catheter or
needle. The volume injected varied between 0.2 - 0.5 mL, depending
on the size of the bladder. Careful notice was taken to first empty the
bladder of urine, and then properly fill it without distending it
extensively. Once injected, the collagenase was left in the bladder for
30 min, and then removed via the catheter/needle. Thereafter, the
serosal side of the bladder was gently rubbed, and loose urothelial
cells were removed by rinsing the bladder with saline. Regularly
after the conduction of the experiments, the bladder was
microscopically examined in order to ensure that the urothelium was
properly removed.
Cystometrical studies
Rats (300 - 350 g) were anaesthetized with an intraperitoneal
injection of pentobarbitone (45 mg/kg). During the remainder of the
experiment supplementary doses were given intravenously when
needed. The rats were placed on a thermostatically controlled
blanket, keeping their body temperature at 38°C. Airways were kept
free by tracheotomy and placement of a cannula in their trachea.
Catheters were placed in both the femoral artery and femoral vein, in
order to monitor blood pressure and administer drugs, respectively.
Two cannulas were inserted through a small incision at the apex of
the bladder, where they also were fixed to the bladder with a
ligature. One of the bladder cannulas was used to maintain bladder
pressure at 10 - 15 mmHg at all times by injection of small volumes
of saline (0.05 - 0.20 mL), and the other was used to monitor
intravesical pressure. When needed, the same cannula that was used
to maintain intravesical pressure was also used to instill collagenase.
Both the catheter in the femoral artery and the pressure sensing
cannula in the urinary bladder were connected to pressure
transducers, as part of the MP100WSW data acquisition system. All
data was recorded using the Acquire software (Biopac, Goleta, USA).

37
Adrenergic effects were blocked in all experiments by administration
of the phentolamine (1 mg/kg i.v.) and propranolol (1 mg/kg i.v.). At
the beginning of each experiment, regardless of series, the
unselective muscarinic agonist methacholine (MeCh) was given in
increasing doses (0.5 – 5 µg/kg) in order to establish a dose-
response curve. Dose-response curves were therefore generated for
untreated control rats and CYP pre-treated rats, with and without
urothelium. In one experimental series, dose-response curves were
also generated for CYP pre-treated rats, before and after removal of
the urothelium, in the presence of the NOS inhibitor
Nω-nitro-L-arginine methyl ester hydrochloride (L-NAME; 30 mg/kg
i.v.). In another experimental series, L-NAME (30 mg/kg i.v.) or saline
(0.9% i.v.) was given to normal rats with an intact urothelium and
CYP pre-treated rats with or without urothelium. Thereafter,
repeated doses of MeCh (2 µg/kg i.v.) were given after increasing
doses of the muscarinic M1/M3/M5-selective antagonist
4-diphenylacetoxy-N-methyl-piperidine methobromide (4-DAMP;
0.1 – 1000 µg/kg i.v.). After each experiment, rats were killed with an
overdose pentobarbitone and their urinary bladders were examined
macroscopically and microscopically.
In vitro functional studies – the organ bath
Experiments on detrusor contractility were conducted in an organ
bath setup. Rats (300 - 350 g; n=52), either CYP(100 mg/kg i.p.) or
saline (0.9%; 1 mL/kg i.p.; serving as controls) pre-treated, were
anaesthetized with an intraperitoneal injection of medetomidine
(Domitor®; 1 mg/kg). Thereafter, they were killed with an overdose
of carbon dioxide and the urinary bladder, with an intact urothelium
or after removal of the urothelium, was removed and placed in Krebs
bicarbonate solution. Via a cut along the posterior surface from the
urethral opening to the apex the bladder was opened, and two or
three full thickness strips (6 x 2 mm) were excised from the detrusor

38
medially to the orifices of the two ureters. The detrusor strips were
mounted between two steel rods, of which one was fixed and the
other was adjustable and connected to an isometric force transducer
(Linton). Thereafter, they were immersed in 25 mL organ baths
containing Krebs bicarbonate solution of the following composition
(mM): NaCl 118, KCl 4.6, CaCl2 1.25, KH2PO4 1.15, MgSO4 1.15,
NaHCO3 25, and glucose 5.5. The baths were kept at 37°C and gassed
with 5% CO2 in O2. The strips were repeatedly stretched for 45 min,
in order to achieve a stable tension of about 5 mN. At the beginning
of each experiment, a high K+ Krebs solution (containing 124 mM K+;
obtained by exchanging Na+ for equimolar amounts of K+) was
administered in order to assess the viability of each strip
preparation. All antagonists, as well as the NOS inhibitor
Nω-nitro-L-arginine (L-NNA), were administered 20 min before each
series of the unselective muscarinic agonist carbachol. All drugs were
administered to the organ baths at a volume of 125 µl. Data were
recorded using a MP100WSW data acquisition system and the
Acquire software (Biopac, Goleta, USA).
In vivo functional studies – the metabolism cage
For these experiments, rats (310 - 400 g; n=100) were pre-treated
with either saline (0.9%; 1 mL/kg i.p.; serving as controls) or CYP
(100 mg/kg i.p.), both in combination with the analgesic
buprenorphinum (10 µg/kg i.p.). Forty-eight hours later, they were
injected intraperitoneally with either saline (0.9%), 4-DAMP
(1 mg/kg), the purinergic P2 receptor antagonist
pyridoxalphosphate-6-azophenyl-2’,4’-disulfonic acid (PPADS;
10 mg/kg) or L-NAME (30 mg/kg) at a volume of 1 mL/kg.
Thereafter, they were placed in a metabolic cage with free supply of
water. During the following 24 h observation period, the rats were
kept in a light-dark-light cycle of 6, 12 and 6 h. Expelled urine was
gathered in a graded bottle below the cage and a laser Doppler
registered each drop of urine continuously. This allowed the urine

39
output, number of micturitions per hour and voided volume per
micturition to be calculated. Total water consumption was also
measured after each experiment. All data were recorded using a
MP100WSW data acquisition system and the Acquire software
(BioPac, Goleta, USA).
Cell cultivation
Two human urothelial cell lines, the UROtsa and T24 cell lines, were
cultivated, both growing as a monolayer. The UROtsa cell line is an
immortalized urothelial cell line derived from the ureter urothelial
lining (Petzoldt et al., 1995). It has been shown to have several
characteristics that closely resemble the intermediate layers of
normal human urothelium (Rossi et al., 2001). The T24 cell line is a
transitional carcinoma cell line that has been shown to be non-
tumorigenic (Flatow et al., 1987), and is frequently used as a cell
culture model to study urothelial cancer cell proliferation (Chen et
al., 2008; Teng et al., 2008; Nakanishi et al., 2009). Both cell lines
were grown at 37°C in a humidified atmosphere containing 5% CO2.
The UROtsa cell line was cultured in Dulbecco´s Modified Eagle
Medium (DMEM; Sigma-Aldrich, St.Louis, USA) while the T24 cell line
was cultured in McCoy’s 5a medium (LGC Promochem, Boras,
Sweden). Both mediums were supplemented with 10% fetal bovine
serum (Sigma-Aldrich, St.Louis, USA) and 1% penicillin/
streptomycin (PEST; PAA Laboratories, Pasching, Austria). Medium
was renewed every other day, and when the cells reached 90%
confluency they were split to a 1:3 ratio.
Measurement of proliferation
In order to measure the proliferation rate of the UROtsa and T24 cell
lines, the MTT Cell Proliferation Assay was utilized. In this assay,
confluent (90-95%) cells were trypsinized and resuspended in
growth medium at a concentration of 400 000 cells per mL. This

40
concentration was determined, by a standard procedure, to yield a
linear relationship between the number of cells and absorbance for
both cell lines. The cells were subsequently pipetted (100 µL) into
separate wells. Two hours later, drugs were added to the wells at a
volume of 10 µL, and the plate was incubated overnight at 37°C in a
5% CO2-atmosphere. Untreated cells and medium not containing
cells were used as control and blank, respectively. The following day,
MTT Reagent (10 µL), which enabled a spectrophotometrical
quantitative measurement of the proliferation rate, was added to
each well. The plate was then returned to the incubator for
approximately three hours. In order for the cells to lysate, a
detergent reagent (100 µL) was added to each well and the plate was
put in dark, at room temperature, overnight. The proliferation rate
(i.e. number of cells), equivalent to the amount of absorbance at
570 nm, was measured the following day.
Materials
Drugs
The drugs employed in this thesis were buprenorphine
hydrochloride (Temgesic®, Apoteket AB, Stockholm, Sweden),
cyclophosphamide monohydrate (CYP), collagenase type I
(Invitrogen, Paisley, UK), acetyl- -methylcholine chloride
(methacholine; MeCh), carbamylcholine chloride (carbachol),
nicotine hemisulfate salt (nicotine), 4-diphenylacetoxy-N-
methylpiperidine methobromide (4-DAMP), p-fluoro-hexahydro-sila-
diphenidol hydrochloride (pFHHSiD), pirenzepine dihydrochloride
(pirenzepine), pyridoxalphosphate-6-azophenyl-2’,4’-disulfonic acid
(PPADS), Nω-nitro-L-arginine (L-NNA), Nω-nitro-L-arginine methyl
ester hydrochloride (L-NAME), chlorbenzoyl-5-methoxy-2-
methylindole-3-acetic acid (indomethacin), phenylephrine
hydrochloride (phenylephrine), isoprenaline hydrochloride

41
(isoprenaline), phentolamine methansulphate (phentolamine),
doxazosin mesylate (doxazosin), propranolol hydrochloride
(propranolol; ICI Pharmaceuticals, London, UK), N-[[3-[(2S)-2-
Hydroxy-3-[[2-[4-[(phenylsulfonyl)amino]phenyl]ethyl]amino]-
propoxy]phenyl]methyl]-acetamide (L-748,337; Tocris, Ellisville,
USA), 3-(2-Ethylphenoxy)-1-[[(1S)-1,2,3,4-tetrahydronaphth-1-yl]-
amino]-(2S)-2-propanol oxalate salt (SR59230A), epidermal growth
factor (EGF) and heparin-binding EGF-like growth factor (HB-EGF).
Unless otherwise stated, all drugs were purchased from Sigma-
Aldrich (St Louis, USA).
Solutions and reagents
The solutions and reagents used in this thesis for cell cultivation and
cell proliferation assays were phosphate buffered saline (PBS; Sigma-
Aldrich, St Louis, USA), Dulbecco´s Modified Eagle Medium (DMEM;
Sigma-Aldrich, St.Louis, USA), McCoy’s 5a medium (LGC Promochem,
Boras, Sweden), fetal bovine serum (Sigma-Aldrich, St.Louis, USA),
penicillin/streptomycin (PEST; PAA Laboratories, Pasching, Austria),
MTT reagent (ATCC, Manassas, VA, USA), MTT detergent (ATCC,
Manassas, VA, USA) and trypsin (PAA laboratories, Pasching,
Austria).
Statistical calculations
Paper I
Comparisons were made between raw data or between relative data.
Raw data comparisons were made when increases in bladder
pressure (Δ bladder pressure) are compared between two groups.
Relative data comparisons were made when comparing the effect
that 4-DAMP had (% decrease) on basal contraction to MeCh.

42
Statistical significance was determined by Student’s t-test for paired
or unpaired data.
Paper II
Statistical comparisons using one-way analysis of variance (ANOVA),
followed by the Bonferroni correction test, was made between raw
data of mean contractile responses to carbachol in repetitive rounds.
When comparing raw data (of mean contractile responses to
carbachol) between control and CYP pre-treated bladder
preparations, Student’s t-test for unpaired data was used to
determine statistical significance. Schild plots and pA2-values were
estimated by obtaining dose ratios of full carbachol response curves
from three antagonist concentrations (Arunlakshana & Schild, 1959).
The estimated Schild slopes were compared to unity utilizing a built-
in test in the GraphPad software (GraphPad Software Inc., San Diego,
USA). Likewise, pharmacodynamic modeling was performed by
applying built-in models for nonlinear regression.
Paper III
One- or two-way ANOVA, followed by the Bonferroni correction test,
was used to determine statistical significance when comparing either
(1) data measured at several time points in the saline pre-treated
(control) group to data measured at several time points in the CYP
pre-treated group, (2) data from saline treated (control) rats to data
from rats treated with an antagonist or L-NAME, (3) data from one
treatment group to combined data from two treatment groups or (4)
data from saline pre-treated rats to data from CYP pre-treated rats
where both groups also received equivalent treatment with 4-DAMP.
Paper IV
In this paper three types of comparisons were made. Either (1) the

43
proliferation rate of UROtsa cells treated with an agonist or
antagonist was compared to the proliferation rate of T24 cells
treated with the same agonist/antagonist, (2) the proliferation rate
of cells treated with an agonist or antagonist was compared to the
proliferation rate of untreated (control) cells or (3) the proliferation
rate of cells treated with both an antagonist and an agonist was
compared to the proliferation rate of cells treated with solely the
agonist. Statistical significance was determined by Student’s t-test for
paired or unpaired data.

44
Results and Discussion
Paper I
Cholinergic nitric oxide release from the urinary bladder
mucosa in cyclophosphamide-induced cystitis of the
anaesthetized rat
In anaesthetized rats, MeCh (0.5 – 5 µg/kg i.v.) generated almost
similar bladder contractions in saline and CYP pre-treated rats with
an intact urothelium. This finding was in contrast to previous
findings in vitro, in which significant differences have been reported
(Giglio et al., 2005b; Giglio et al., 2007). However, this discrepancy
might be explained by a combination of several factors. Firstly, in
vivo the intravesical pressure can only reach a certain level before
the micturition reflex is initiated. This may have prevented the
expected differences in maximal detrusor contractility. Secondly, the
largest dose of MeCh led to a dramatic drop in blood pressure which
could have affected bladder pressure (i.e. lowered the maximal
response). Thirdly, in vivo MeCh reaches primarily the detrusor, as it
is introduced to the bladder via the blood stream, while in vitro
carbachol has equal access to detrusor and urothelium. The
inhibitory CYP-induced effects seen in vitro might therefore be
imperceptible in vivo.
When the response to MeCh (0.5 – 5 µg/kg i.v.) was tested in rats in
the presence and absence of the urothelium, larger contractions
were observed in CYP pre-treated bladders (p < 0.05; n = 5) in the
absence of the urothelium. Meanwhile, the contractions were
unaffected by removal of the urothelium in normal bladders. When
examining the dose dependent effects of MeCh on CYP pre-treated
bladders in the presence of L-NAME, no contractile alterations arose
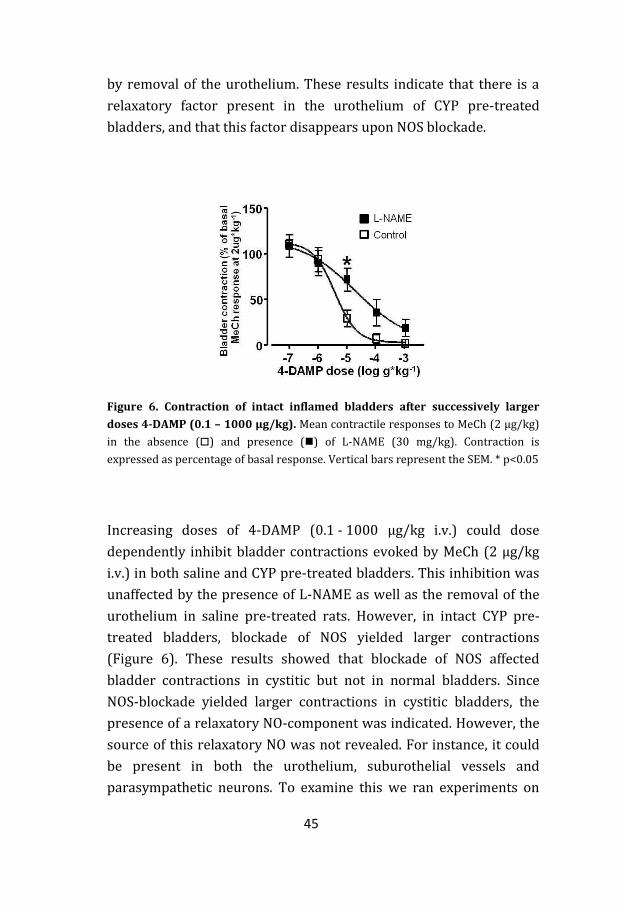
45
by removal of the urothelium. These results indicate that there is a
relaxatory factor present in the urothelium of CYP pre-treated
bladders, and that this factor disappears upon NOS blockade.
Figure 6. Contraction of intact inflamed bladders after successively larger
doses 4-DAMP (0.1 – 1000 μg/kg). Mean contractile responses to MeCh (2 μg/kg)
in the absence () and presence () of L-NAME (30 mg/kg). Contraction is
expressed as percentage of basal response. Vertical bars represent the SEM. * p<0.05
Increasing doses of 4-DAMP (0.1 - 1000 µg/kg i.v.) could dose
dependently inhibit bladder contractions evoked by MeCh (2 µg/kg
i.v.) in both saline and CYP pre-treated bladders. This inhibition was
unaffected by the presence of L-NAME as well as the removal of the
urothelium in saline pre-treated rats. However, in intact CYP pre-
treated bladders, blockade of NOS yielded larger contractions
(Figure 6). These results showed that blockade of NOS affected
bladder contractions in cystitic but not in normal bladders. Since
NOS-blockade yielded larger contractions in cystitic bladders, the
presence of a relaxatory NO-component was indicated. However, the
source of this relaxatory NO was not revealed. For instance, it could
be present in both the urothelium, suburothelial vessels and
parasympathetic neurons. To examine this we ran experiments on
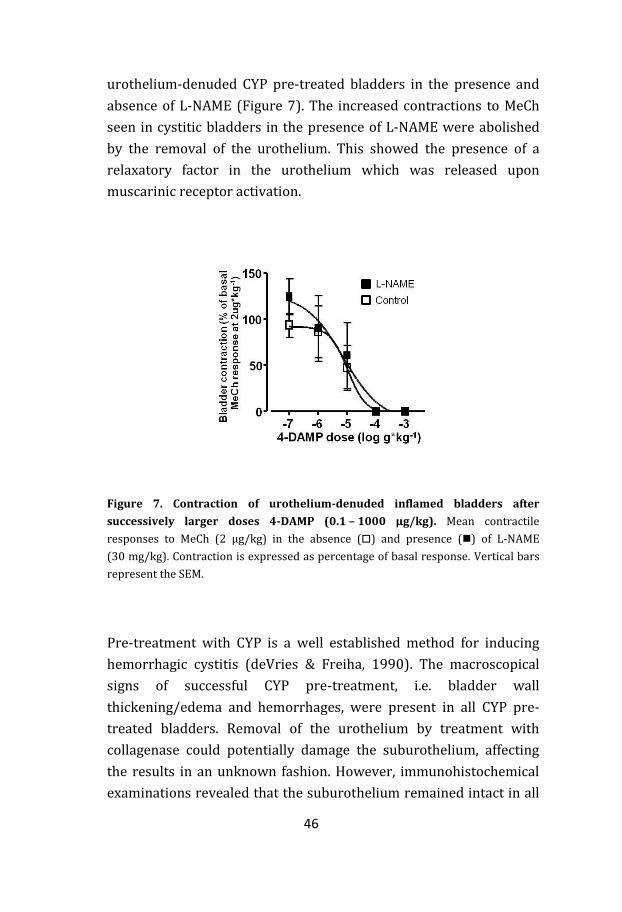
46
urothelium-denuded CYP pre-treated bladders in the presence and
absence of L-NAME (Figure 7). The increased contractions to MeCh
seen in cystitic bladders in the presence of L-NAME were abolished
by the removal of the urothelium. This showed the presence of a
relaxatory factor in the urothelium which was released upon
muscarinic receptor activation.
Figure 7. Contraction of urothelium-denuded inflamed bladders after
successively larger doses 4-DAMP (0.1 – 1000 μg/kg). Mean contractile
responses to MeCh (2 μg/kg) in the absence () and presence () of L-NAME
(30 mg/kg). Contraction is expressed as percentage of basal response. Vertical bars
represent the SEM.
Pre-treatment with CYP is a well established method for inducing
hemorrhagic cystitis (deVries & Freiha, 1990). The macroscopical
signs of successful CYP pre-treatment, i.e. bladder wall
thickening/edema and hemorrhages, were present in all CYP pre-
treated bladders. Removal of the urothelium by treatment with
collagenase could potentially damage the suburothelium, affecting
the results in an unknown fashion. However, immunohistochemical
examinations revealed that the suburothelium remained intact in all

47
bladders and that 80-90% of the urothelium was removed by the
procedure. Further, damages caused by urothelial denudation should
render a lower contractile response, which was not the case.
Detrusor overactivity (DO) and increased frequency are two of the
most common features of CYP-induced cystitis in both humans and
animals. During our experiments, we observed small spontaneous
contractions that were more frequent in cystitic bladders, especially
after treatment with L-NAME. In the state of anaesthesia, these
spontaneous contractions were too small to elicit a micturition.
However, it cannot be excluded that these contractions could yield
micturition events in the conscious rat, thereby linking the DO to
increased frequency. Many patients with DO do not respond to
antimuscarinic treatment. Minaglia et al. (2005) showed a link
between patients with DO that are refractory to antimuscarinic drugs
and concomitant IC. This non-responsiveness to antimuscarinic
treatment could perhaps in part be explained by our findings.
The current investigation shows that CYP pre-treatment alters the
cholinergic response of the urinary bladder. We show that NO is
present in the urothelium during CYP-induced cystitis and that this
NO exerts an inhibitory effect on detrusor contractility. Further, we
suggest that NO is released upon cholinergic stimulation, possibly
through activation of M5 and/or M3 receptors.

48
Paper II
The muscarinic M5 receptor is the primary mediator of
urothelium-derived nitric oxide effects in the rat urinary
bladder
In bladder strips from CYP pre-treated rats, the maximal contractile
response to carbachol (0.01 - 100 µM) was significantly reduced in
comparison to normal bladder strips (Figure 8; 19.3 ± 1.0 mN and
14.3 ± 1.2 mN in saline and CYP pre-treated strips, respectively;
p < 0.01).
Figure 8. Contraction of intact urinary bladders. Mean contractile responses to
carbachol (0.01 - 100 µM) of isolated detrusor strip preparations from saline (●) and
CYP (■) pre-treated rats. Vertical bars represent the SEM. * p<0.05; ** p<0.01
A comparison of the EC50-values also showed a significant right-shift
of the concentration-response curve in CYP pre-treated bladder
strips (logEC50 = -6.1 and -5.4 in saline and CYP pre-treated rats,
respectively; p < 0.001). The right shift, as well as the difference in
maximal contractile response, was abolished by removal of the
urothelium (Figure 9). Administration of the NOS inhibitor L-NNA
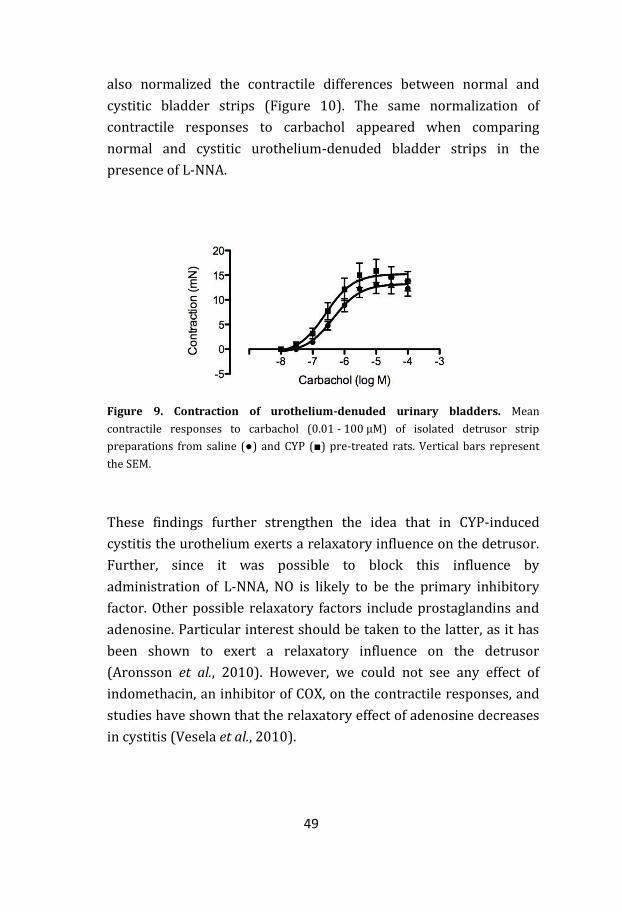
49
also normalized the contractile differences between normal and
cystitic bladder strips (Figure 10). The same normalization of
contractile responses to carbachol appeared when comparing
normal and cystitic urothelium-denuded bladder strips in the
presence of L-NNA.
Figure 9. Contraction of urothelium-denuded urinary bladders. Mean
contractile responses to carbachol (0.01 - 100 µM) of isolated detrusor strip
preparations from saline (●) and CYP (■) pre-treated rats. Vertical bars represent
the SEM.
These findings further strengthen the idea that in CYP-induced
cystitis the urothelium exerts a relaxatory influence on the detrusor.
Further, since it was possible to block this influence by
administration of L-NNA, NO is likely to be the primary inhibitory
factor. Other possible relaxatory factors include prostaglandins and
adenosine. Particular interest should be taken to the latter, as it has
been shown to exert a relaxatory influence on the detrusor
(Aronsson et al., 2010). However, we could not see any effect of
indomethacin, an inhibitor of COX, on the contractile responses, and
studies have shown that the relaxatory effect of adenosine decreases
in cystitis (Vesela et al., 2010).

50
Figure 10. Contraction of intact urinary bladders in the presence of L-NNA
(100 µM). Mean contractile responses to carbachol (0.01 - 100 µM) of isolated
detrusor strip preparations from saline (●) and CYP (■) pre-treated rats. Vertical
bars represent the SEM.
The carbachol induced contractions were concentration-dependently
inhibited by both 4-DAMP, the muscarinic M3 antagonist pFHHSiD
and the muscarinic M1 antagonist pirenzepine. While pFHHSiD and
pirenzepine inhibited the contractile responses of saline and CYP
pre-treated rats in a similar fashion, 4-DAMP inhibited the
contractile responses of CYP pre-treated bladder strips less potently
than saline pre-treated (pA2 value = 9.9 and 9.5 in saline and CYP
pre-treated bladders, respectively; p < 0.01). In the CYP pre-treated
bladders, the lowest concentration of 4-DAMP (10-10 M) even tended
to generate a left-shift of the contraction curve. By either removing
the urothelium or running the experiments in the presence of L-NNA,
the difference in inhibitory potency of 4-DAMP in CYP pre-treated
bladders was abolished. Analysis of the Schild slopes for all three
antagonists revealed that they were all significantly different from
unity in CYP pre-treated bladder strips, but not in the normal
preparations. This is in favor of the notion that a change in
contractility occurs during cystitis. Removal of the urothelium
eliminated this difference, indicating that the contractility is altered
due to changes mediated via the urothelium. Similarly, the difference

51
from unity of the Schild slopes was abolished by the presence of
L-NNA.
The antagonistic profile is different for the three examined
antagonists. While 4-DAMP inhibits M1, M3 and M5 in a similar
fashion, pFHHSiD and pirenzepine are considered mainly a M3- and
M1-antagonist, respectively (Eglen & Nahorski, 2000). The
antagonistic potency of 4-DAMP is reduced in cystitic bladder strips,
indicating that either the M1, M3 or M5 receptor (or more than one
of them) is involved in the altered contractile responses seen during
cystitis. Since neither the potency of pFHHSiD nor pirenzepine is
changed during cystitis, M5 seems to be the primary receptor
involved in this alteration. Taken together, our findings indicate that
changes occur during CYP-induced cystitis that alter NO-mediated
effects via urothelial M5 receptors.
Correlation plots for the pA2-values obtained in the study versus
–Ki values reported in the literature for the five muscarinic receptors
support the generally accepted idea that detrusor contraction is
mediated mainly by stimulation of muscarinic M3 receptors, both in
normal and inflamed bladders (r2 for M3 = 0.98 and 0.93 in normal
and inflamed bladder strips, respectively). Thus, as expected, these
plots do not support a notion that M5 receptors are directly involved
in the contractile response of the detrusor. However, the correlation
plots show that during CYP-induced cystitis, the M3-mediated
contraction seems to be less dominant, indicating the influence of
some other receptor or factor on contraction.
It is important to keep in mind that there may be differences
between CYP-induced and naturally occurring cystitis, as well as
inter-species disparities. Studies on bladder strips from cats have
shown that during naturally occurring cystitis, activation of
urothelial muscarinic receptors may actually lead to increased
contractions at low agonist doses (Ikeda et al., 2009). Therefore,

52
extrapolation of the interpretation of the data in this study to man
must be performed with caution. Further, one also has to keep in
mind that these are studies performed in an organ bath setup,
thereby excluding the possibility for compensatory effects via the
nervous system or changes in blood pressure.
To summarize, this study shows that the contractile changes seen in
CYP-induced cystitis are mediated via stimulation of muscarinic M5
receptors and the following release of NO. Further, this study agrees
with several previous studies that conclude that detrusor contraction
in the healthy rat bladder is mediated almost entirely via muscarinic
M3 receptors.
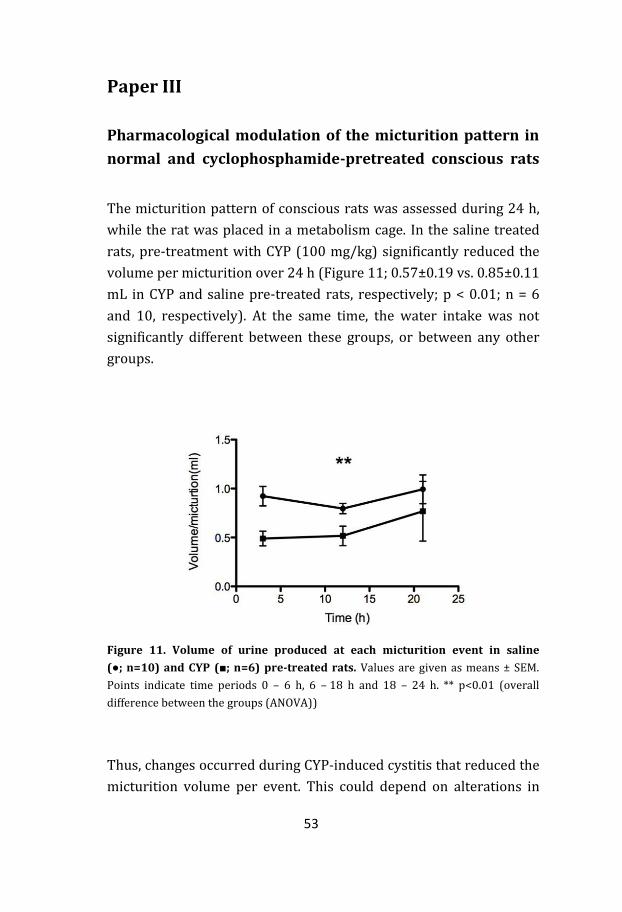
53
Paper III
Pharmacological modulation of the micturition pattern in
normal and cyclophosphamide-pretreated conscious rats
The micturition pattern of conscious rats was assessed during 24 h,
while the rat was placed in a metabolism cage. In the saline treated
rats, pre-treatment with CYP (100 mg/kg) significantly reduced the
volume per micturition over 24 h (Figure 11; 0.57±0.19 vs. 0.85±0.11
mL in CYP and saline pre-treated rats, respectively; p < 0.01; n = 6
and 10, respectively). At the same time, the water intake was not
significantly different between these groups, or between any other
groups.
Figure 11. Volume of urine produced at each micturition event in saline
(●; n=10) and CYP (■; n=6) pre-treated rats. Values are given as means ± SEM.
Points indicate time periods 0 – 6 h, 6 – 18 h and 18 – 24 h. ** p<0.01 (overall
difference between the groups (ANOVA))
Thus, changes occurred during CYP-induced cystitis that reduced the
micturition volume per event. This could depend on alterations in

54
the contractile threshold of the detrusor, or solely be a result of a
decreased detrusor contractile ability. The latter would increase the
amount of residual urine, which has been shown to occur during
CYP-induced cystitis (Ito et al., 2007). We could also see a slight, but
not significant, increase in micturition frequency in CYP pre-treated
rats. Since the volume per micturition is lower in CYP pre-treated
rats, at the same time as the water intake is unchanged, and other
studies have shown an increase in frequency during CYP-induced
cystitis (Smaldone et al., 2009), we would have expected a significant
increase in micturition frequency. However, in many other studies
observations have been made over shorter time periods (< 12 h). In
our study, measurements were made over a 24 h period, which may
conceal temporary changes occurring during shorter periods.
Others have shown that CYP-induced cystitis can cause bladder
overactivity (Borvendeg et al., 2003). During CYP-induced cystitis, an
increase in both the release of urothelial ATP and of the expression
of eNOS and muscarinic receptors has been shown to occur (Giglio et
al., 2005b; Smith et al., 2005). In the current study, in CYP pre-
treated rats, treatment with L-NAME (30 mg/kg) led to an increase in
micturition frequency (Figure 12a; 2.53 ± 0.36 vs. 1.25 ± 0.17
events/h in L-NAME and saline treated rats, respectively; p < 0.05;
n = 6 in both groups). In saline pre-treated rats, L-NAME did not
affect any micturition parameter. This is in line with findings in other
studies that show that intravesically administered NO-donors can
abolish the increased frequency seen in CYP pre-treated rats (Ozawa
et al., 1999). Our results, and those of others, show that in CYP-
induced cystitis, NO exerts an inhibitory (relaxatory) effect on the
detrusor.

55
Figure 12. Effects of treatments on micturition parameters in cystitis.
Frequency of micturition events (a) and volume of urine produced at each
micturition event (b) of CYP pre-treated rats during the 6 – 18 h dark period. Forty-
eight hours after CYP pre-treatment, saline (1 mL/kg i.p.; n=10), 4-DAMP (1 mg/kg
i.p.; n=6), L-NAME (30 mg/kg i.p.; n=6) or PPADS (10 mg/kg i.p.; n=6) was
administered. Values are given as means ± SEM. * p<0.05
The muscarinic antagonist 4-DAMP affected micturition parameters
of CYP but not saline pre-treated rats. The volume per micturition of
CYP pre-treated rats during the 6 – 18 h dark period was
significantly increased by treatment with 4-DAMP (Figure 12b;
1 mg/kg; 0.75 ± 0.05 and 0.51 ± 0.09 mL in 4-DAMP and saline
treated rats, respectively; p < 0.05; n = 6 and 10, respectively).
Similarly, in paired comparisons, treatment with different doses of
4-DAMP (10 - 1000 µg/kg) eliminated the significant difference in
volume per micturition between saline and CYP pre-treated rats.
Even though non-significant, the frequency of CYP pre-treated rats
was increased, rather than decreased, by treatment with 4-DAMP
(Figure 12a). This finding is in line with studies in man that report
that patients with DO are non-responsive to antimuscarinic
treatment (Minaglia et al., 2005). It is evident that treatment with

56
antimuscarinic drugs has limited effects in the healthy bladder of
rats, which is in line with previous studies (Modiri et al., 2002).
However, in the cystitic bladder, 4-DAMP increases the volume per
micturition, perhaps via an increase of detrusor contractility. The
absence of effect of an antimuscarinic drug in the normal rat bladder
is likely, at least partly, due to compensatory effects from the
purinergic system.
Treatment with a combination of 4-DAMP (1 mg/kg) and L-NAME
(30 mg/kg) did not affect any of the micturition parameters
differently than separate administration of the two drugs, in neither
saline nor CYP pre-treated rats. However, when combining 4-DAMP
(1 mg/kg) with PPADS (10mg/kg), a synergistic effect was seen on
the voiding frequency in CYP pre-treated rats during the 6 – 18 h
dark period. This could indicate that at least part of the effects of
muscarinic antagonists and those of NOS blockers are mediated via
the same pathway, while the effects of purinergic antagonists are
mediated via a separate pathway. The reason for why a synergistic
effect is only seen during the dark period is likely due to the fact that
it is during this period that the rats are awake, and thus the
micturition activity is the highest and most likely to reveal
differences in micturition parameters.
When given separately to CYP pre-treated rats, treatment with
4-DAMP leads to increased volumes per micturition, while treatment
with L-NAME leads to increased micturition frequency. Even though
these two parameters are not totally interlinked, they indicate an
opposite effect of 4-DAMP and L-NAME in CYP pre-treated rats. To
understand the discrepancies between a combinatory treatment and
that of 4-DAMP and L-NAME alone, one has to remember that
4-DAMP may exert its effects via two separate pathways; (a) via
blockade of muscarinic receptors in the efferent and/or afferent
pathways, leading to decreased detrusor activity, and (b) via
blockade of urothelial muscarinic receptors, which according to our

57
studies in vivo and in vitro leads to a decreased relaxatory influence
of NO on the detrusor. This kind of dual effect of 4-DAMP would
explain our findings.
Experiments on wake, freely moving rats present certain problems
when examining drugs that have a relatively small impact on
micturition parameters. More specifically, the interindividual
variations among rats are sometimes larger than the effects of the
studied drugs, making it impossible to statistically detect minute
changes. However, this observable fact strengthens the correctness
in those significant differences one does find.
The current study indicates that changes occur during CYP-induced
cystitis that not only affects the micturition parameters, but also the
effects of antimuscarinic, antipurinergic and antinitrergic drugs. It is
possible to reverse the altered micturition parameters in the
inflamed rat bladder by treatment with an antimuscarinic drug,
indicating the strong involvement of these receptors in the
alterations occurring during cystitis. Further, this study shows that
there is a connection between the effect of antimuscarinic and
antinitrergic drugs. Thus, this study strengthens the previously
proposed idea of an increased importance of NO, which is released
upon muscarinic receptor stimulation, during CYP-induced cystitis.

58
Paper IV
Characterization of receptor mediated proliferation in the
human bladder urothelial UROtsa and T24 cell lines
Studies on various types of gastrointestinal inflammation have
shown that chronic inflammation of the bowel together with
increased proliferation of the mucosa is strongly associated with the
development of cancer (Ament, 1975; MacDonald, 1992). Treatment
with CYP causes similar proliferative changes in the urothelium
(Zupancic et al., 2009). Further, more than 90% of all urinary bladder
cancer originates from the urothelium. When studying this disease,
others have reported a link between chronic inflammation and
urothelial transformation (Talar-Williams et al., 1996; Michaud,
2007). It has also been shown that the expression and/or sensitivity
of certain receptors in the urothelium can be altered in the state of
inflammation and transformation (Giglio et al., 2005b; MacLaine et
al., 2008).
The proliferative effects of adrenergic, muscarinic, nicotinic and EGF
receptor agonists were examined in an immortalized normal human
bladder urothelial cell line, designated UROtsa, and a malignant
human bladder urothelial cell line, designated T24. Regarding the cell
culturing, all experiments were performed during passage 4 – 10.
This should be well within the time frame where one would expect
cell properties to stay consistent.
The non-selective β-adrenoceptor agonist isoprenaline
concentration-dependently increased the proliferation rate of both
the UROtsa and T24 cell line (Figure 13a; pEC50 = 3.0 and 3.1,
respectively). This proliferative effect could not be antagonized by
the β1/β2-selective antagonist propranolol (2 x 10-5 M) in neither the
UROtsa nor T24 cell line. However, the selective β3-adrenoceptor
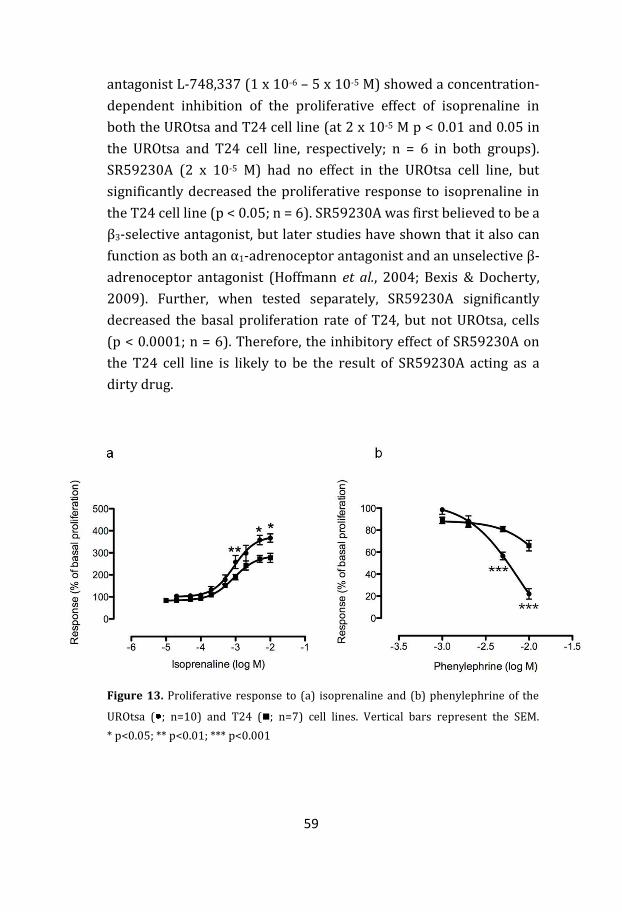
59
antagonist L-748,337 (1 x 10-6 – 5 x 10-5 M) showed a concentration-
dependent inhibition of the proliferative effect of isoprenaline in
both the UROtsa and T24 cell line (at 2 x 10-5 M p < 0.01 and 0.05 in
the UROtsa and T24 cell line, respectively; n = 6 in both groups).
SR59230A (2 x 10-5 M) had no effect in the UROtsa cell line, but
significantly decreased the proliferative response to isoprenaline in
the T24 cell line (p < 0.05; n = 6). SR59230A was first believed to be a
β3-selective antagonist, but later studies have shown that it also can
function as both an α1-adrenoceptor antagonist and an unselective β-
adrenoceptor antagonist (Hoffmann et al., 2004; Bexis & Docherty,
2009). Further, when tested separately, SR59230A significantly
decreased the basal proliferation rate of T24, but not UROtsa, cells
(p < 0.0001; n = 6). Therefore, the inhibitory effect of SR59230A on
the T24 cell line is likely to be the result of SR59230A acting as a
dirty drug.
Figure 13. Proliferative response to (a) isoprenaline and (b) phenylephrine of the
UROtsa ( ; n=10) and T24 (; n=7) cell lines. Vertical bars represent the SEM.
* p<0.05; ** p<0.01; *** p<0.001

60
These results indicate that the increased proliferative response to
isoprenaline in the UROtsa and T24 cell lines is mediated mainly via
the β3-adrenoceptor. Interestingly, β3-agonists, which have been
suggested as treatment for OAB, have been shown to induce cystitis
in rats (Waghe et al., 1999). Based on our finding in urothelial cells,
we would expect such treatment to yield thickening of the bladder
mucosa, one of the characteristic symptoms of cystitis. The reason
for why it was not possible to block the entire proliferative effect of
isoprenaline with L-748,337 could depend on one of two things (or
both of these). Either the effect is partly mediated via other pathways
than activation of the β3-adrenoceptor, or the concentrations of
L-748,337 chosen for the experiments (1 x 10-6 – 5 x 10-5 M) were not
high enough to block the entire proliferative effect. The range of
concentrations of L-748,337, and that of all other agonists and
antagonists, was based on pilot experiments, and doses showing
signs of toxic effects were excluded. Unfortunately, it was therefore
not possible to test higher concentrations of L-748,337.
In the presence of the α1-adrenoceptor agonist phenylephrine, a
decrease of the proliferation rate was seen in both the UROtsa and
T24 cell line (Figure 13b; pIC50 = 2.2 and 1.7, respectively; n = 6). At
the highest concentration (1 x 10-2 M), there was a significantly
greater anti-proliferative effect in the UROtsa cell line (Figure 13b;
p < 0.0001; n = 6). When it comes to anti-proliferative effects, one
cannot exclude that the response is not simply a toxic effect. This is
further highlighted by the fact that neither the unselective α-
adrenergic antagonist phentolamine (10-9 – 10-4 M) nor the α1-
selective adrenoceptor antagonist doxazosin (10-9 – 10-4 M) could
block the effect of phenylephrine (at 5 x 10-3 M). However, if the anti-
proliferative effect of phenylephrine was not mediated via receptor
activation, the impact should be equal in the two cell lines. Further,
the concentrations of α-adrenergic antagonists used in this study
could have been too low to inhibit the effect of phenylephrine at

61
5 x 10-3 M. Unfortunately, it was not possible to use higher
concentrations of antagonists due to toxic effects.
The expression of muscarinic receptors seems to be different in the
two studied cell lines. While all subtypes (M1 - M5) have been shown
to be present in the UROtsa cell line (Tyagi et al., 2006), another
study could only detect mRNA for the M2 and M5 receptors in the
T24 cell line (Tully et al., 2009). As we have shown in a previous
study on rats, the expression of muscarinic receptors, in particular
the M5 subtype, was altered in the state of CYP-induced cystitis
(Giglio et al., 2005b). Thus, one of the possible links between
inflammation and bladder cancer could be alterations in the
expression of muscarinic receptors. We could, however, not see any
functional effect of this altered expression, as the non-selective
muscarinic agonist methacholine did not affect the proliferation rate
of neither the UROtsa nor T24 cell line. One plausible explanation for
the lack of proliferative effects upon activation of muscarinic
receptors could be due to two or more effects cancelling each other
out. I.e., by employing a non-selective agonist, activation of one
subtype might increase proliferation, while activation of another
subtype might decrease the rate of cell division. Future experiments
should be designed to elucidate this possibility.
Nicotine, and the activation of the nicotinic receptor, has been
associated with the development of bladder cancer and modulation
of the proliferation of malignant urothelial cells (Chen et al., 2008).
However, we could not see any effect of nicotine (1 x 10-9 –
1 x 10-7 M) in neither the UROtsa nor T24 cell line. This could be due
to toxicity problems, which only rendered the possibility to use very
low concentrations.
Activation of the EGF receptor by EGF or HB-EGF is closely linked to
increased cell proliferation, and their involvement in the
transformation of urothelial cells has been suggested (Adam et al.,

62
2003). Several other studies have shown an increased proliferation
rate upon EGF receptor activation, but despite using wide ranges of
concentrations for both EGF (5 – 500 ng/mL) and HB-EGF
(1 – 200 ng/mL), we could not see this effect in neither the UROtsa
nor T24 cell line. This could possibly be explained by studies that
have shown that only the autocrine release, and not the exocrine
addition, of EGF or HB-EGF can affect the proliferation rate of
urothelial cells (Southgate et al., 1994; Rebel et al., 1995; Freeman et
al., 1997; Sterle et al., 1997). Further, other studies have suggested
that activation of the EGF receptor only increases the proliferation
rate of urothelial cells during challenges such as wound repair
(Varley et al., 2005).
Studies on cell cultures are beneficial in the sense that they offer an
experimental setup that allows the study of one specific target in one
specific cell type. However, one must always draw conclusions from
cell culture studies with caution. The proliferation of urothelial cells
is most likely an interplay between not only the urothelial cells
themselves, but also other cell types. Findings in cell cultures should
therefore, when possible, be verified in vivo.
The results in this report show the importance of adrenergic
receptors in the regulation of urothelial cell proliferation, both in a
normal and a malignant cell line. Further, since the UROtsa cell line is
affected to a greater extent than the T24 cell line by both α- and β-
adrenoceptor stimulation, we show that the impact of adrenergic
receptor stimulation on cell proliferation is decreased in the
malignant state. This report also examines the impact of muscarinic,
nicotinic and EGF receptor stimulation, but the current results do not
support the involvement of any of these in urothelial cell
proliferation during unchallenged conditions.
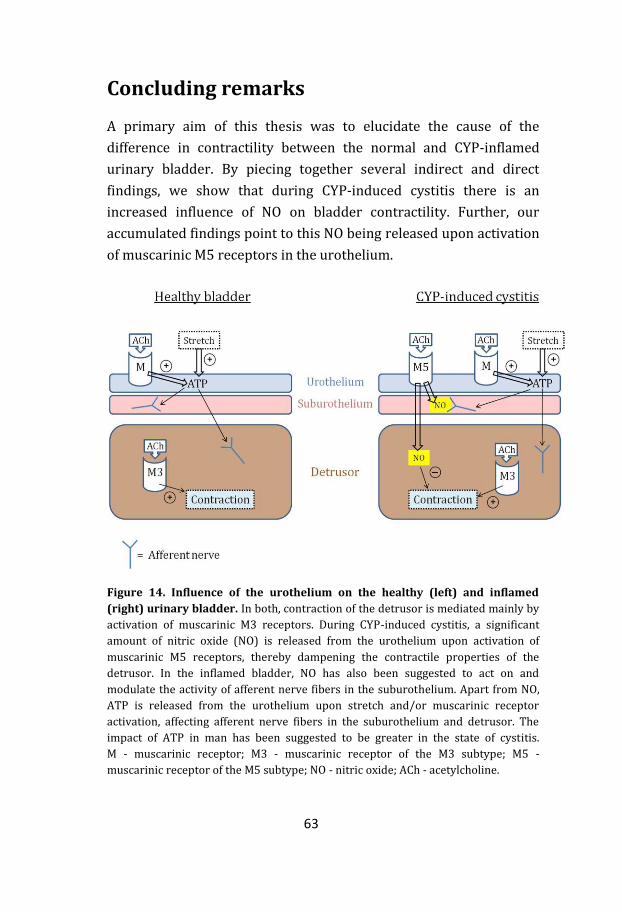
63
Concluding remarks
A primary aim of this thesis was to elucidate the cause of the
difference in contractility between the normal and CYP-inflamed
urinary bladder. By piecing together several indirect and direct
findings, we show that during CYP-induced cystitis there is an
increased influence of NO on bladder contractility. Further, our
accumulated findings point to this NO being released upon activation
of muscarinic M5 receptors in the urothelium.
Figure 14. Influence of the urothelium on the healthy (left) and inflamed
(right) urinary bladder. In both, contraction of the detrusor is mediated mainly by
activation of muscarinic M3 receptors. During CYP-induced cystitis, a significant
amount of nitric oxide (NO) is released from the urothelium upon activation of
muscarinic M5 receptors, thereby dampening the contractile properties of the
detrusor. In the inflamed bladder, NO has also been suggested to act on and
modulate the activity of afferent nerve fibers in the suburothelium. Apart from NO,
ATP is released from the urothelium upon stretch and/or muscarinic receptor
activation, affecting afferent nerve fibers in the suburothelium and detrusor. The
impact of ATP in man has been suggested to be greater in the state of cystitis.
M - muscarinic receptor; M3 - muscarinic receptor of the M3 subtype; M5 -
muscarinic receptor of the M5 subtype; NO - nitric oxide; ACh - acetylcholine.

64
Much of the research on the urothelium has focused on its role as an
integrator of afferent signaling. This led to that early studies on the
involvement of the urothelium during cystitis mainly examined the
effects that factors released from the urothelium had on afferent
nerves. However, the results presented in this thesis show that
urothelial factors have impact on not only afferent, but also efferent
mechanisms. It is also currently shown that the significance of this
impact can be altered during disease states such as cystitis. This
knowledge might highlight numerous new targets for future
pharmacological treatments. For instance, there is a debate whether
up-regulation of NO during cystitis is solely a result of the
inflammation, or if it is actually part of the cause of the inflammation
(Souza-Fiho et al., 1997). This is a matter of great interest,
particularly regarding the development of new pharmacological
therapies for IC, which are needed. A major problem today is the
uncertainty in the diagnosis of IC. Several factors have been
suggested as early markers for IC, but so far no diagnostic tests have
been developed. If an early marker for IC was found, and NO plays an
important part in the development of inflammation, it might be
possible to prevent IC by inhibiting the increased impact of NO
during cystitis. Future experiments should be conducted in order to
elucidate the development of inflammation, and if it is possible to
influence this development.
A key concern for our studies was the proper induction of cystitis. In
order for our data to be translatable, it was relevant for us that the
cystitis we studied was similar to IC as it appears in man. Several
animal models have been established to study experimental cystitis,
of which induction with CYP is the most common (deVries & Freiha,
1990). There are many similarities between CYP-induced cystitis and
IC. For one, CYP-induced cystitis gives symptoms that are very
similar to IC. Secondly, none of them give rise to inflammation due to
infection. Thirdly, CYP-induced cystitis, just as IC, has been reported
to sensitize afferent pathways (Yu & de Groat, 2008). Further, IC is

65
associated with an altered release of NO and ATP. During CYP-
induced cystitis, an up-regulation of both endothelial nitric oxide
synthase (eNOS) and muscarinic receptors, mainly of the M5
subtype, has been shown (Giglio et al., 2005b). Also, both altered
release of ATP and increased frequency have been shown to arise
during CYP-induced cystitis (Wood et al., 2001; Smith et al., 2005).
However, there are also certain differences between IC and CYP-
induced cystitis. For instance, ultrastructural studies of bladder
specimens from IC patients has shown a disrupted barrier function of
the urothelium (Elbadawi & Light, 1996). This is not equally
apparent in CYP-induced cystitis, where the common observation is a
thickening of the bladder mucosa. There may also be differences in
the initiating causes of inflammation. In CYP-induced cystitis the
inflammatory response is evoked by the presence of chemical
irritants in the urine, while the exact cause for IC remains unclear.
The results presented in this thesis may also be applicable to other
bladder disturbances than IC. In man, the classic symptom of DO is
frequency. Treatment for patients suffering from frequency is today
focused on antimuscarinic drugs. However, several studies have
shown limited long-term clinical effects of these drugs (Andersson,
2009). A possible goal for the future treatment of symptoms of
frequency could be via regulation of NO release. If our data are
translatable to man, a compound selective for the muscarinic M5
receptor might improve quality of life for millions of patients
suffering from inflammation-induced DO.
As mentioned previously, a recurrent observation during our studies
was an increased proliferation of the urothelium upon CYP-
treatment. Concomitantly, an increase of the expression of
muscarinic M5 receptors occurs in the urothelium (Giglio et al.,
2005b). This same link, between increased proliferation and
expression of M5 receptors, is observed in other diseases such as
Sjögrens syndrome (Ryberg et al., 2008). Despite these studies

66
showing a potential link between proliferation and increased
expression of M5 receptors we could not show any effect of a
muscarinic agonist on urothelial cell proliferation. The expression of
all five muscarinic receptors has been shown in normal human
urothelium (Table 1). However, in order to validate these findings,
future studies should be performed to elucidate receptor expression
in the UROtsa and T24 urothelial cell lines. Furthermore, future
proliferation experiments should be performed in order to
investigate if other results could have been obtained by the use of
selective agonists and antagonists for these receptors.

67
Acknowledgements
My sincere gratitude goes out to all of you whom have helped me
complete this thesis. You know who you are. However, some of you I
would like to thank in particular.
First and foremost I would like to thank my supervisor, Gunnar
Tobin. Without your help and encouragement I would have never
even begun my PhD-studies, let alone completed them. I owe you
one.
My co-supervisor, Daniel Giglio. Thank you for the many pointers and
good scientific discussions.
My bladder friend, Patrik Aronsson. Thank you for your company,
friendship and all our great discussions. And thank you for putting up
with me during our trips.
My fellow PhD-student in our group, Anders Ryberg. Thank you for
teaching me immunological methods in the lab and that comic strips
can be considered fine art.
Group members Hanna Asklund, Martin Johnsson and Christine
Ahlström. Thank you for fun lunches and help with my papers.
Co-authors Anna Wilhelmson, Davoud Eskandari, Deya Doufish and
Anette Lampert. Thank you for your help in gathering data for my
papers and for you good company.
My pharmafriends from the Czech Republic, Renata Vesela, Ondrej
Soukup and Pavel Jeřábek. Thank you for helping me with my papers.
And thank you for teaching me that dumplings are an essential part
of life.
Professor Scott Garrett, Univeristy of South Dakota, for kindly
providing me with the UROtsa cell line without any request for
compensation.

68
To Hans Nissbrandt and Christer Nilsson at the department of
pharmacology. Thank you for discussing sports, my favorite interest,
with me. I need that sometimes.
To all other co-workers at the department of pharmacology. Thank
you for proving such a stimulating and positive surrounding.
To my friends at other parts of the University, Johan Bourghardt,
Louise Strandberg and Marie Kalm. Thank you for numerous fun
lunches
To former and present members of the PhD committee, in particular
Malin von Otter and Karin Hultman. Thank you for your
commitment.
To my parents, for guiding me to where I am today. Thank you for
your endless support.
To my son, William. You bring a smile to my face each and every day.
Thank you for teaching me that one should not take the voluntary
control of the bladder for granted.
Finally, but mostly, to my beloved Elin. Thank you for always being
there, for always listening to me (even when I ramble) and for
showing me what is most important in life. I love you with all my
heart.
This thesis was supported by grants from Anders Otto Swärd
Foundation/Ulrika Eklund Foundation, Ferrings och Svenska Enures
Akademin, Lars Hierta Foundation, Wilhelm och Martina Lundgrens
Vetenskapsfond, Karl och Annie Leons Minnesfond and the Magn.
Bergvall Foundation

69
References
Abdel-Rahman M, Galeano C & Elhilali M. (1983). New approach to study of voiding cycle in cat. Preliminary report on pharmacologic studies. Urology 22, 91-97.
Abrams P, Andersson KE, Buccafusco JJ, Chapple C, de Groat WC, Fryer AD, Kay G, Laties
A, Nathanson NM, Pasricha PJ & Wein AJ. (2006). Muscarinic receptors: their distribution and function in body systems, and the implications for treating overactive bladder. Br J Pharmacol.
Abrams P, Cardozo L, Fall M, Griffiths D, Rosier P, Ulmsten U, van Kerrebroeck P, Victor A
& Wein A. (2002). The standardisation of terminology of lower urinary tract function: report from the Standardisation Sub-committee of the International Continence Society. Neurourol Urodyn 21, 167-178.
Adam RM, Danciu T, McLellan DL, Borer JG, Lin J, Zurakowski D, Weinstein MH,
Rajjayabun PH, Mellon JK & Freeman MR. (2003). A nuclear form of the heparin-binding epidermal growth factor-like growth factor precursor is a feature of aggressive transitional cell carcinoma. Cancer Res 63, 484-490.
Alfieri AB & Cubeddu LX. (2000). Nitric oxide and NK(1)-tachykinin receptors in
cyclophosphamide-induced cystitis, in rats. J Pharmacol Exp Ther 295, 824-829. Ament ME. (1975). Inflammatory disease of the colon: ulcerative colitis and Crohn's
colitis. J Pediatr 86, 322-334. Andersson KE. (2002a). Bladder activation: afferent mechanisms. Urology 59, 43-50. Andersson KE. (2002b). Overactive bladder--pharmacological aspects. Scand J Urol
Nephrol Suppl, 72-81. Andersson KE. (2009). Pharmacotherapy of the overactive bladder. Discov Med 8, 118-
124. Andersson KE, Holmquist F, Fovaeus M, Hedlund H & Sundler R. (1991). Muscarinic
receptor stimulation of phosphoinositide hydrolysis in the human isolated urinary bladder. J Urol 146, 1156-1159.
Andersson KE, Larsson B & Sjogren C. (1984). Characterization of the alpha-
adrenoceptors in the female rabbit urethra. Br J Pharmacol 81, 293-300. Andersson KE & Persson K. (1994). Nitric oxide synthase and nitric oxide-mediated
effects in lower urinary tract smooth muscles. World J Urol 12, 274-280. Apodaca G. (2004). The uroepithelium: not just a passive barrier. Traffic 5, 117-128. Aronsson P, Andersson M, Ericsson T & Giglio D. (2010). Assessment and
characterization of purinergic contractions and relaxations in the rat urinary bladder. Basic Clin Pharmacol Toxicol 107, 603-613.

70
Arunlakshana O & Schild HO. (1959). Some quantitative uses of drug antagonists. Br J Pharmacol Chemother 14, 48-58.
Barras M, Coste A, Eon MT & Guillot E. (1999). Pharmacological characterization of
muscarinic receptors implicated in rabbit detrusor muscle contraction and activation of inositol phospholipid hydrolysis in rabbit detrusor and parotid gland. Fundam Clin Pharmacol 13, 562-570.
Bayliss M, Wu C, Newgreen D, Mundy AR & Fry CH. (1999). A quantitative study of
atropine-resistant contractile responses in human detrusor smooth muscle, from stable, unstable and obstructed bladders. J Urol 162, 1833-1839.
Beckel JM, Kanai A, Lee SJ, de Groat WC & Birder LA. (2006). Expression of functional
nicotinic acetylcholine receptors in rat urinary bladder epithelial cells. Am J Physiol Renal Physiol 290, F103-110.
Bexis S & Docherty JR. (2009). Role of alpha 1- and beta 3-adrenoceptors in the
modulation by SR59230A of the effects of MDMA on body temperature in the mouse. Br J Pharmacol 158, 259-266.
Birder LA, Apodaca G, De Groat WC & Kanai AJ. (1998). Adrenergic- and capsaicin-evoked
nitric oxide release from urothelium and afferent nerves in urinary bladder. Am J Physiol 275, F226-229.
Birder LA, Kanai AJ, de Groat WC, Kiss S, Nealen ML, Burke NE, Dineley KE, Watkins S,
Reynolds IJ & Caterina MJ. (2001). Vanilloid receptor expression suggests a sensory role for urinary bladder epithelial cells. Proc Natl Acad Sci U S A 98, 13396-13401.
Bolton TB & Zholos AV. (1997). Activation of M2 muscarinic receptors in guinea-pig
ileum opens cationic channels modulated by M3 muscarinic receptors. Life Sci 60, 1121-1128.
Born M, Pahner I, Ahnert-Hilger G & Jons T. (2003). The maintenance of the permeability
barrier of bladder facet cells requires a continuous fusion of discoid vesicles with the apical plasma membrane. Eur J Cell Biol 82, 343-350.
Borvendeg SJ, Al-Khrasani M, Rubini P, Fischer W, Allgaier C, Wirkner K, Himmel HM,
Gillen C & Illes P. (2003). Subsensitivity of P2X but not vanilloid 1 receptors in dorsal root ganglia of rats caused by cyclophosphamide cystitis. Eur J Pharmacol 474, 71-75.
Brandt WD, Matsui W, Rosenberg JE, He X, Ling S, Schaeffer EM & Berman DM. (2009).
Urothelial carcinoma: stem cells on the edge. Cancer Metastasis Rev 28, 291-304.
Braverman A, Legos J, Young W, Luthin G & Ruggieri M. (1999). M2 receptors in genito-
urinary smooth muscle pathology. Life Sci 64, 429-436. Braverman AS, Kohn IJ, Luthin GR & Ruggieri MR. (1998). Prejunctional M1 facilitory and
M2 inhibitory muscarinic receptors mediate rat bladder contractility. Am J Physiol 274, R517-523.

71
Bschleipfer T, Schukowski K, Weidner W, Grando SA, Schwantes U, Kummer W & Lips KS.
(2007). Expression and distribution of cholinergic receptors in the human urothelium. Life Sci 80, 2303-2307.
Bult H, Boeckxstaens GE, Pelckmans PA, Jordaens FH, Van Maercke YM & Herman AG.
(1990). Nitric oxide as an inhibitory non-adrenergic non-cholinergic neurotransmitter. Nature 345, 346-347.
Burnstock G. (2009). Purinergic signalling: past, present and future. Braz J Med Biol Res
42, 3-8. Caine M, Raz S & Zeigler M. (1975). Adrenergic and cholinergic receptors in the human
prostate, prostatic capsule and bladder neck. Br J Urol 47, 193-202. Caulfield MP & Birdsall NJ. (1998). International Union of Pharmacology. XVII.
Classification of muscarinic acetylcholine receptors. Pharmacol Rev 50, 279-290.
Chaiyaprasithi B, Mang CF, Kilbinger H & Hohenfellner M. (2003). Inhibition of human
detrusor contraction by a urothelium derived factor. J Urol 170, 1897-1900. Chancellor MB, Kaplan SA & Blaivas JG. (1992). The cholinergic and purinergic
components of detrusor contractility in a whole rabbit bladder model. J Urol 148, 906-909.
Chang RS, Chen TB, O'Malley SS, Pettibone DJ, DiSalvo J, Francis B, Bock MG, Freidinger R,
Nagarathnam D, Miao SW, Shen Q, Lagu B, Murali Dhar TG, Tyagarajan S, Marzabadi MR, Wong WC, Gluchowski C & Forray C. (2000). In vitro studies on L-771,688 (SNAP 6383), a new potent and selective alpha1A-adrenoceptor antagonist. Eur J Pharmacol 409, 301-312.
Chen RJ, Ho YS, Guo HR & Wang YJ. (2008). Rapid activation of Stat3 and ERK1/2 by
nicotine modulates cell proliferation in human bladder cancer cells. Toxicol Sci 104, 283-293.
Chesher GB & Thorp RH. (1965). The atropine-resistance of the response to intrinsic
nerve stimulation of the guinea-pig bladder. Br J Pharmacol Chemother 25, 288-294.
Chess-Williams R. (2002). Muscarinic receptors of the urinary bladder: detrusor,
urothelial and prejunctional. Auton Autacoid Pharmacol 22, 133-145. Chess-Williams R, Chapple CR, Yamanishi T, Yasuda K & Sellers DJ. (2001). The minor
population of M3-receptors mediate contraction of human detrusor muscle in vitro. J Auton Pharmacol 21, 243-248.
Chopra B, Barrick SR, Meyers S, Beckel JM, Zeidel ML, Ford AP, de Groat WC & Birder LA.
(2005). Expression and function of bradykinin B1 and B2 receptors in normal and inflamed rat urinary bladder urothelium. J Physiol 562, 859-871.

72
Chopra B, Gever J, Barrick SR, Hanna-Mitchell AT, Beckel JM, Ford AP & Birder LA. (2008). Expression and function of rat urothelial P2Y receptors. Am J Physiol Renal Physiol 294, F821-829.
Cordon-Cardo C. (2008). Molecular alterations associated with bladder cancer initiation
and progression. Scand J Urol Nephrol Suppl, 154-165. Coussens LM & Werb Z. (2002). Inflammation and cancer. Nature 420, 860-867. Cox PJ. (1979). Cyclophosphamide cystitis--identification of acrolein as the causative
agent. Biochem Pharmacol 28, 2045-2049. Curhan GC, Speizer FE, Hunter DJ, Curhan SG & Stampfer MJ. (1999). Epidemiology of
interstitial cystitis: a population based study. J Urol 161, 549-552. D'Agostino G, Barbieri A, Chiossa E & Tonini M. (1997). M4 muscarinic autoreceptor-
mediated inhibition of -3H-acetylcholine release in the rat isolated urinary bladder. J Pharmacol Exp Ther 283, 750-756.
D'Agostino G, Bolognesi ML, Lucchelli A, Vicini D, Balestra B, Spelta V, Melchiorre C &
Tonini M. (2000). Prejunctional muscarinic inhibitory control of acetylcholine release in the human isolated detrusor: involvement of the M4 receptor subtype. Br J Pharmacol 129, 493-500.
de Groat WC. (2004). The urothelium in overactive bladder: passive bystander or active
participant? Urology 64, 7-11. deVries CR & Freiha FS. (1990). Hemorrhagic cystitis: a review. J Urol 143, 1-9. Drake MJ. (2008). Emerging drugs for treatment of overactive bladder and detrusor
overactivity. Expert Opin Emerg Drugs 13, 431-446. Durant PA, Lucas PC & Yaksh TL. (1988). Micturition in the unanesthetized rat: spinal vs.
peripheral pharmacology of the adrenergic system. J Pharmacol Exp Ther 245, 426-435.
Eglen RM & Nahorski SR. (2000). The muscarinic M(5) receptor: a silent or emerging
subtype? Br J Pharmacol 130, 13-21. Eglen RM, Reddy H, Watson N & Challiss RA. (1994). Muscarinic acetylcholine receptor
subtypes in smooth muscle. Trends Pharmacol Sci 15, 114-119. Elbadawi AE & Light JK. (1996). Distinctive ultrastructural pathology of nonulcerative
interstitial cystitis: new observations and their potential significance in pathogenesis. Urol Int 56, 137-162.
Elmer M. (1974). Inhibitory beta-adrenoceptors in the urinary bladder of the rat. Life Sci
15, 273-280. Elsebai I. (1977). Parasites in the etiology of cancer--bilharziasis and bladder cancer. CA
Cancer J Clin 27, 100-106.

73
Erickson DR, Xie SX, Bhavanandan VP, Wheeler MA, Hurst RE, Demers LM, Kushner L & Keay SK. (2002). A comparison of multiple urine markers for interstitial cystitis. J Urol 167, 2461-2469.
Fall M. (1999). [Urinary incontinence in men--a neglected problem? A quarter of all 80-
year old men suffer of urinary leakage]. Lakartidningen 96, 2227-2231. Fall M, Lindstrom S & Mazieres L. (1990). A bladder-to-bladder cooling reflex in the cat. J
Physiol 427, 281-300. Ferguson DR, Kennedy I & Burton TJ. (1997). ATP is released from rabbit urinary bladder
epithelial cells by hydrostatic pressure changes--a possible sensory mechanism? J Physiol 505 ( Pt 2), 503-511.
Fetscher C, Fleichman M, Schmidt M, Krege S & Michel MC. (2002). M(3) muscarinic
receptors mediate contraction of human urinary bladder. Br J Pharmacol 136, 641-643.
Flatow U, Rabson AB & Rabson AS. (1987). Tumorigenicity of T24 urinary bladder
carcinoma cell sublines. Int J Cancer 40, 240-245. Freeman MR, Yoo JJ, Raab G, Soker S, Adam RM, Schneck FX, Renshaw AA, Klagsbrun M &
Atala A. (1997). Heparin-binding EGF-like growth factor is an autocrine growth factor for human urothelial cells and is synthesized by epithelial and smooth muscle cells in the human bladder. J Clin Invest 99, 1028-1036.
Giglio D, Aronsson P, Eriksson L & Tobin G. (2007). In vitro characterization of
parasympathetic and sympathetic responses in cyclophosphamide-induced cystitis in the rat. Basic Clin Pharmacol Toxicol 100, 96-108.
Giglio D, Delbro DS & Tobin G. (2001). On the functional role of muscarinic M2 receptors
in cholinergic and purinergic responses in the rat urinary bladder. Eur J Pharmacol 428, 357-364.
Giglio D, Delbro DS & Tobin G. (2005a). Postjunctional modulation by muscarinic M2
receptors of responses to electrical field stimulation of rat detrusor muscle preparations. Auton Autacoid Pharmacol 25, 113-120.
Giglio D, Ryberg AT, To K, Delbro DS & Tobin G. (2005b). Altered muscarinic receptor
subtype expression and functional responses in cyclophosphamide induced cystitis in rats. Auton Neurosci 122, 9-20.
Giglio D & Tobin G. (2009). Muscarinic receptor subtypes in the lower urinary tract.
Pharmacology 83, 259-269. Girard BM, Wolf-Johnston A, Braas KM, Birder LA, May V & Vizzard MA. (2008). PACAP-
mediated ATP release from rat urothelium and regulation of PACAP/VIP and receptor mRNA in micturition pathways after cyclophosphamide (CYP)-induced cystitis. J Mol Neurosci 36, 310-320.

74
Goepel M, Wittmann A, Rubben H & Michel MC. (1997). Comparison of adrenoceptor subtype expression in porcine and human bladder and prostate. Urol Res 25, 199-206.
Guarneri L, Ibba M, Angelico P & Testa R. (1991). Effects of oxybutynin, terodiline, and
nifedipine on the cystometrogram in conscious rats with infravesical outflow obstruction. Pharmacol Res 24, 263-272.
Habler HJ, Janig W & Koltzenburg M. (1990). Activation of unmyelinated afferent fibres
by mechanical stimuli and inflammation of the urinary bladder in the cat. J Physiol 425, 545-562.
Hampel C, Artibani W, Espuna Pons M, Haab F, Jackson S, Romero J, Gavart S &
Papanicolaou S. (2004). Understanding the burden of stress urinary incontinence in Europe: a qualitative review of the literature. Eur Urol 46, 15-27.
Hanno PM. (2002). Interstitial cystitis-epidemiology, diagnostic criteria, clinical markers.
Rev Urol 4 Suppl 1, S3-8. Harriss DR, Marsh KA, Birmingham AT & Hill SJ. (1995). Expression of muscarinic M3-
receptors coupled to inositol phospholipid hydrolysis in human detrusor cultured smooth muscle cells. J Urol 154, 1241-1245.
Hawthorn MH, Chapple CR, Cock M & Chess-Williams R. (2000). Urothelium-derived
inhibitory factor(s) influences on detrusor muscle contractility in vitro. Br J Pharmacol 129, 416-419.
Hegde SS, Choppin A, Bonhaus D, Briaud S, Loeb M, Moy TM, Loury D & Eglen RM. (1997).
Functional role of M2 and M3 muscarinic receptors in the urinary bladder of rats in vitro and in vivo. Br J Pharmacol 120, 1409-1418.
Hegde SS & Eglen RM. (1999). Muscarinic receptor subtypes modulating smooth muscle
contractility in the urinary bladder. Life Sci 64, 419-428. Hoffmann C, Leitz MR, Oberdorf-Maass S, Lohse MJ & Klotz KN. (2004). Comparative
pharmacology of human beta-adrenergic receptor subtypes--characterization of stably transfected receptors in CHO cells. Naunyn Schmiedebergs Arch Pharmacol 369, 151-159.
Hu P, Meyers S, Liang FX, Deng FM, Kachar B, Zeidel ML & Sun TT. (2002). Role of
membrane proteins in permeability barrier function: uroplakin ablation elevates urothelial permeability. Am J Physiol Renal Physiol 283, F1200-1207.
Hudman D, Elliott RA, Whitaker P, Terry TR, Sandhu DP & Norman RI. (2001). Inhibition
of the contractile responses of isolated human and rat bladders by clenbuterol. J Urol 166, 1969-1973.
Igawa Y, Yamazaki Y, Takeda H, Akahane M, Ajisawa Y, Yoneyama T & Nishizawa O.
(1998). Possible beta 3-adrenoceptor-mediated relaxation of the human detrusor. Acta Physiol Scand 164, 117-118.

75
Igawa Y, Yamazaki Y, Takeda H, Kaidoh K, Akahane M, Ajisawa Y, Yoneyama T, Nishizawa O & Andersson KE. (2001). Relaxant effects of isoproterenol and selective beta3-adrenoceptor agonists on normal, low compliant and hyperreflexic human bladders. J Urol 165, 240-244.
Ikeda Y, Birder L, Buffington C, Roppolo J & Kanai A. (2009). Mucosal muscarinic
receptors enhance bladder activity in cats with feline interstitial cystitis. J Urol 181, 1415-1422.
Inadome A, Yoshida M, Takahashi W, Yono M, Seshita H, Miyamoto Y, Kawano T & Ueda S.
(1998). Prejunctional muscarinic receptors modulating acetylcholine release in rabbit detrusor smooth muscles. Urol Int 61, 135-141.
Ishihama H, Momota Y, Yanase H, Wang X, de Groat WC & Kawatani M. (2006). Activation
of alpha1D adrenergic receptors in the rat urothelium facilitates the micturition reflex. J Urol 175, 358-364.
Ishizuka O, Mattiasson A & Andersson KE. (1995a). Prostaglandin E2-induced bladder
hyperactivity in normal, conscious rats: involvement of tachykinins? J Urol 153, 2034-2038.
Ishizuka O, Mattiasson A & Andersson KE. (1995b). Tachykinin effects on bladder activity
in conscious normal rats. J Urol 154, 257-261. Ito K, Iwami A, Katsura H & Ikeda M. (2007). Therapeutic effects of the putative
P2X(3)/P2X (2/3) antagonist A-317491 on cyclophosphamide-induced cystitis in rats. Naunyn Schmiedebergs Arch Pharmacol.
Janig W & Morrison JF. (1986). Functional properties of spinal visceral afferents
supplying abdominal and pelvic organs, with special emphasis on visceral nociception. Prog Brain Res 67, 87-114.
Jen PY, Dixon JS & Gosling JA. (1995). Immunohistochemical localization of neuromarkers
and neuropeptides in human fetal and neonatal urinary bladder. Br J Urol 75, 230-235.
Jost SP, Gosling JA & Dixon JS. (1989). The morphology of normal human bladder
urothelium. J Anat 167, 103-115. Keay S, Zhang CO, Kagen DI, Hise MK, Jacobs SC, Hebel JR, Gordon D, Whitmore K,
Bodison S & Warren JW. (1997). Concentrations of specific epithelial growth factors in the urine of interstitial cystitis patients and controls. J Urol 158, 1983-1988.
Kim Y, Yoshimura N, Masuda H, de Miguel F & Chancellor MB. (2005). Antimuscarinic
agents exhibit local inhibitory effects on muscarinic receptors in bladder-afferent pathways. Urology 65, 238-242.
Klapproth H, Reinheimer T, Metzen J, Munch M, Bittinger F, Kirkpatrick CJ, Hohle KD,
Schemann M, Racke K & Wessler I. (1997). Non-neuronal acetylcholine, a signalling molecule synthezised by surface cells of rat and man. Naunyn Schmiedebergs Arch Pharmacol 355, 515-523.

76
Koziol JA, Adams HP & Frutos A. (1996). Discrimination between the ulcerous and the
nonulcerous forms of interstitial cystitis by noninvasive findings. J Urol 155, 87-90.
Kullmann FA, Artim D, Beckel J, Barrick S, de Groat WC & Birder LA. (2008a).
Heterogeneity of muscarinic receptor-mediated Ca2+ responses in cultured urothelial cells from rat. Am J Physiol Renal Physiol 294, F971-981.
Kullmann FA, Artim DE, Birder LA & de Groat WC. (2008b). Activation of muscarinic
receptors in rat bladder sensory pathways alters reflex bladder activity. J Neurosci 28, 1977-1987.
Lecci A, Giuliani S, Tramontana M, Santicioli P, Criscuoli M, Dion S & Maggi CA. (1998).
Bladder distension and activation of the efferent function of sensory fibres: similarities with the effect of capsaicin. Br J Pharmacol 124, 259-266.
Levin RM, Ruggieri MR & Wein AJ. (1988). Identification of receptor subtypes in the
rabbit and human urinary bladder by selective radio-ligand binding. J Urol 139, 844-848.
Li G, Li K, Li Z & Wang P. (2003). Age-dependent changes in beta-adrenoceptor function
in human detrusors and possible mechanisms. Chin Med J (Engl) 116, 1511-1514.
Lluel P, Deplanne V, Heudes D, Bruneval P & Palea S. (2003). Age-related changes in
urethrovesical coordination in male rats: relationship with bladder instability? Am J Physiol Regul Integr Comp Physiol 284, R1287-1295.
Logadottir YR, Ehren I, Fall M, Wiklund NP, Peeker R & Hanno PM. (2004). Intravesical
nitric oxide production discriminates between classic and nonulcer interstitial cystitis. J Urol 171, 1148-1150; discussion 1150-1141.
Longhurst PA, Belis JA, O'Donnell JP, Galie JR & Westfall DP. (1984). A study of the
atropine-resistant component of the neurogenic response of the rabbit urinary bladder. Eur J Pharmacol 99, 295-302.
MacDonald TT. (1992). Epithelial proliferation in response to gastrointestinal
inflammation. Ann N Y Acad Sci 664, 202-209. MacKenzie I, Burnstock G & Dolly JO. (1982). The effects of purified botulinum
neurotoxin type A on cholinergic, adrenergic and non-adrenergic, atropine-resistant autonomic neuromuscular transmission. Neuroscience 7, 997-1006.
MacLaine NJ, Wood MD, Holder JC, Rees RW & Southgate J. (2008). Sensitivity of normal,
paramalignant, and malignant human urothelial cells to inhibitors of the epidermal growth factor receptor signaling pathway. Mol Cancer Res 6, 53-63.
Malloy BJ, Price DT, Price RR, Bienstock AM, Dole MK, Funk BL, Rudner XL, Richardson
CD, Donatucci CF & Schwinn DA. (1998). Alpha1-adrenergic receptor subtypes in human detrusor. J Urol 160, 937-943.

77
Matsui M, Motomura D, Karasawa H, Fujikawa T, Jiang J, Komiya Y, Takahashi S & Taketo MM. (2000). Multiple functional defects in peripheral autonomic organs in mice lacking muscarinic acetylcholine receptor gene for the M3 subtype. Proc Natl Acad Sci U S A 97, 9579-9584.
Mattiasson A, Andersson KE, Elbadawi A, Morgan E & Sjogren C. (1987). Interaction
between adrenergic and cholinergic nerve terminals in the urinary bladder of rabbit, cat and man. J Urol 137, 1017-1019.
Michaud DS. (2007). Chronic inflammation and bladder cancer. Urol Oncol 25, 260-268. Milsom I. (2009). Lower urinary tract symptoms in women. Curr Opin Urol 19, 337-341. Milsom I, Abrams P, Cardozo L, Roberts RG, Thuroff J & Wein AJ. (2001). How widespread
are the symptoms of an overactive bladder and how are they managed? A population-based prevalence study. BJU Int 87, 760-766.
Minaglia S, Ozel B, Bizhang R & Mishell DR, Jr. (2005). Increased prevalence of interstitial
cystitis in women with detrusor overactivity refractory to anticholinergic therapy. Urology 66, 702-706.
Modiri AR, Alberts P & Gillberg PG. (2002). Effect of muscarinic antagonists on
micturition pressure measured by cystometry in normal, conscious rats. Urology 59, 963-968.
Monneron MC, Gillberg PG, Ohman B & Alberts P. (2000). In vitro alpha-adrenoceptor
autoradiography of the urethra and urinary bladder of the female pig, cat, guinea-pig and rat. Scand J Urol Nephrol 34, 233-238.
Munoz A, Gangitano DA, Smith CP, Boone TB & Somogyi GT. (2010). Removal of
urothelium affects bladder contractility and release of ATP but not release of NO in rat urinary bladder. BMC Urol 10, 10.
Mysorekar IU, Mulvey MA, Hultgren SJ & Gordon JI. (2002). Molecular regulation of
urothelial renewal and host defenses during infection with uropathogenic Escherichia coli. J Biol Chem 277, 7412-7419.
Nakamura T, Yoshimura M, Shinnick-Gallagher P, Gallagher JP & Akasu T. (1984). alpha 2
and alpha 1-Adrenoceptors mediate opposing actions on parasympathetic neurons. Brain Res 323, 349-353.
Nakanishi R, Oka N, Nakatsuji H, Koizumi T, Sakaki M, Takahashi M, Fukumori T &
Kanayama HO. (2009). Effect of vascular endothelial growth factor and its receptor inhibitor on proliferation and invasion in bladder cancer. Urol Int 83, 98-106.
Nergardh A, Boreus LO & Naglo AS. (1977). Characterization of the adrenergic beta-
receptor in the urinary bladder of man and cat. Acta Pharmacol Toxicol (Copenh) 40, 14-21.

78
Nomiya M & Yamaguchi O. (2003). A quantitative analysis of mRNA expression of alpha 1 and beta-adrenoceptor subtypes and their functional roles in human normal and obstructed bladders. J Urol 170, 649-653.
Noronha-Blob L, Lowe V, Patton A, Canning B, Costello D & Kinnier WJ. (1989).
Muscarinic receptors: relationships among phosphoinositide breakdown, adenylate cyclase inhibition, in vitro detrusor muscle contractions and in vivo cystometrogram studies in guinea pig bladder. J Pharmacol Exp Ther 249, 843-851.
Ohshima H, Tatemichi M & Sawa T. (2003). Chemical basis of inflammation-induced
carcinogenesis. Arch Biochem Biophys 417, 3-11. Oravisto KJ. (1975). Epidemiology of interstitial cystitis. Ann Chir Gynaecol Fenn 64, 75-
77. Oshita M, Hiraoka Y & Watanabe Y. (1997). Characterization of beta-adrenoceptors in
urinary bladder: comparison between rat and rabbit. Br J Pharmacol 122, 1720-1724.
Otsuka A, Shinbo H, Matsumoto R, Kurita Y & Ozono S. (2008). Expression and functional
role of beta-adrenoceptors in the human urinary bladder urothelium. Naunyn Schmiedebergs Arch Pharmacol 377, 473-481.
Ozawa H, Chancellor MB, Jung SY, Yokoyama T, Fraser MO, Yu Y, de Groat WC &
Yoshimura N. (1999). Effect of intravesical nitric oxide therapy on cyclophosphamide-induced cystitis. J Urol 162, 2211-2216.
Palea S, Artibani W, Ostardo E, Trist DG & Pietra C. (1993). Evidence for purinergic
neurotransmission in human urinary bladder affected by interstitial cystitis. J Urol 150, 2007-2012.
Pandita RK, Mizusawa H & Andersson KE. (2000). Intravesical oxyhemoglobin initiates
bladder overactivity in conscious, normal rats. J Urol 164, 545-550. Pandita RK, Ronn LC, Jensen BS & Andersson KE. (2006). Urodynamic effects of
intravesical administration of the new small/intermediate conductance calcium activated potassium channel activator NS309 in freely moving, conscious rats. J Urol 176, 1220-1224.
Parsons CL, Boychuk D, Jones S, Hurst R & Callahan H. (1990). Bladder surface
glycosaminoglycans: an epithelial permeability barrier. J Urol 143, 139-142. Petzoldt JL, Leigh IM, Duffy PG, Sexton C & Masters JR. (1995). Immortalisation of human
urothelial cells. Urol Res 23, 377-380. Rebel JM, Thijssen CD, Vermey M, Zwarthoff EC & Van der Kwast TH. (1995). Modulation
of intra-epithelial expansion of human T24 bladder-carcinoma cells in murine urothelium by growth factors and extracellular-matrix components. Int J Cancer 60, 707-711.

79
Rossi MR, Masters JR, Park S, Todd JH, Garrett SH, Sens MA, Somji S, Nath J & Sens DA. (2001). The immortalized UROtsa cell line as a potential cell culture model of human urothelium. Environ Health Perspect 109, 801-808.
Rotterud R, Nesland JM, Berner A & Fossa SD. (2005). Expression of the epidermal
growth factor receptor family in normal and malignant urothelium. BJU Int 95, 1344-1350.
Ryberg AT, Warfvinge G, Axelsson L, Soukup O, Gotrick B & Tobin G. (2008). Expression
of muscarinic receptor subtypes in salivary glands of rats, sheep and man. Arch Oral Biol 53, 66-74.
Saito M, Nakamura I & Miyagawa I. (1997). [Autoradiographic localization of muscarinic
receptors in diabetic rat bladder]. Nippon Hinyokika Gakkai Zasshi 88, 858-867. Samaras V, Rafailidis PI, Mourtzoukou EG, Peppas G & Falagas ME. (2010). Chronic
bacterial and parasitic infections and cancer: a review. J Infect Dev Ctries 4, 267-281.
Sampselle CM & DeLancey JO. (1998). Anatomy of female continence. J Wound Ostomy
Continence Nurs 25, 63-70, 72-64. Sanchez Freire V, Burkhard FC, Kessler TM, Kuhn A, Draeger A & Monastyrskaya K.
(2010). MicroRNAs may mediate the down-regulation of neurokinin-1 receptor in chronic bladder pain syndrome. Am J Pathol 176, 288-303.
Save S, Mjosberg J, Poljakovic M, Mohlin C & Persson K. (2009). Adenosine receptor
expression in Escherichia coli-infected and cytokine-stimulated human urinary tract epithelial cells. BJU Int 104, 1758-1765.
Save S & Persson K. (2010). Extracellular ATP and P2Y receptor activation induce a
proinflammatory host response in the human urinary tract. Infect Immun 78, 3609-3615.
Schwartz DA. (1981). Helminths in the induction of cancer II. Schistosoma haematobium
and bladder cancer. Trop Geogr Med 33, 1-7. Seguchi H, Nishimura J, Zhou Y, Niiro N, Kumazawa J & Kanaide H. (1998). Expression of
beta3-adrenoceptors in rat detrusor smooth muscle. J Urol 159, 2197-2201. Shirai T. (1993). Etiology of bladder cancer. Semin Urol 11, 113-126. Sigala S, Mirabella G, Peroni A, Pezzotti G, Simeone C, Spano P & Cunico SC. (2002).
Differential gene expression of cholinergic muscarinic receptor subtypes in male and female normal human urinary bladder. Urology 60, 719-725.
Sigala S, Peroni A, Mirabella G, Fornari S, Palazzolo F, Pezzotti G, Simeone C, Cunico SC &
Spano P. (2004). Alpha1 adrenoceptor subtypes in human urinary bladder: sex and regional comparison. Life Sci 76, 417-427.

80
Sjogren C, Andersson KE, Husted S, Mattiasson A & Moller-Madsen B. (1982). Atropine resistance of transmurally stimulated isolated human bladder muscle. J Urol 128, 1368-1371.
Smaldone MC, Vodovotz Y, Tyagi V, Barclay D, Philips BJ, Yoshimura N, Chancellor MB &
Tyagi P. (2009). Multiplex analysis of urinary cytokine levels in rat model of cyclophosphamide-induced cystitis. Urology 73, 421-426.
Smet PJ, Jonavicius J, Marshall VR & de Vente J. (1996). Distribution of nitric oxide
synthase-immunoreactive nerves and identification of the cellular targets of nitric oxide in guinea-pig and human urinary bladder by cGMP immunohistochemistry. Neuroscience 71, 337-348.
Smith CP, Vemulakonda VM, Kiss S, Boone TB & Somogyi GT. (2005). Enhanced ATP
release from rat bladder urothelium during chronic bladder inflammation: effect of botulinum toxin A. Neurochem Int 47, 291-297.
Somogyi GT & de Groat WC. (1990). Modulation of the release of [3H]norepinephrine
from the base and body of the rat urinary bladder by endogenous adrenergic and cholinergic mechanisms. J Pharmacol Exp Ther 255, 204-210.
Somogyi GT & de Groat WC. (1999). Function, signal transduction mechanisms and
plasticity of presynaptic muscarinic receptors in the urinary bladder. Life Sci 64, 411-418.
Somogyi GT, Tanowitz M & de Groat WC. (1994). M1 muscarinic receptor-mediated
facilitation of acetylcholine release in the rat urinary bladder. J Physiol 480 ( Pt 1), 81-89.
Somogyi GT, Tanowitz M & de Groat WC. (1995). Prejunctional facilitatory alpha 1-
adrenoceptors in the rat urinary bladder. Br J Pharmacol 114, 1710-1716. Somogyi GT, Tanowitz M, Zernova G & de Groat WC. (1996). M1 muscarinic receptor-
induced facilitation of ACh and noradrenaline release in the rat bladder is mediated by protein kinase C. J Physiol 496 ( Pt 1), 245-254.
Somogyi GT, Zernova GV, Yoshiyama M, Yamamoto T & de Groat WC. (1998). Frequency
dependence of muscarinic facilitation of transmitter release in urinary bladder strips from neurally intact or chronic spinal cord transected rats. Br J Pharmacol 125, 241-246.
Southgate J, Hutton KA, Thomas DF & Trejdosiewicz LK. (1994). Normal human
urothelial cells in vitro: proliferation and induction of stratification. Lab Invest 71, 583-594.
Souza-Fiho MV, Lima MV, Pompeu MM, Ballejo G, Cunha FQ & Ribeiro Rde A. (1997).
Involvement of nitric oxide in the pathogenesis of cyclophosphamide-induced hemorrhagic cystitis. Am J Pathol 150, 247-256.
Sterle M, Kreft ME & Batista U. (1997). The effect of epidermal growth factor and
transforming growth factor beta 1 on proliferation and differentiation of urothelial cells in urinary bladder explant culture. Biol Cell 89, 263-271.

81
Sui GP, Wu C, Roosen A, Ikeda Y, Kanai AJ & Fry CH. (2008). Modulation of bladder
myofibroblast activity: implications for bladder function. Am J Physiol Renal Physiol 295, F688-697.
Sun Y, Keay S, De Deyne PG & Chai TC. (2001). Augmented stretch activated adenosine
triphosphate release from bladder uroepithelial cells in patients with interstitial cystitis. J Urol 166, 1951-1956.
Szell EA, Yamamoto T, de Groat WC & Somogyi GT. (2000). Smooth muscle and
parasympathetic nerve terminals in the rat urinary bladder have different subtypes of alpha(1) adrenoceptors. Br J Pharmacol 130, 1685-1691.
Talar-Williams C, Hijazi YM, Walther MM, Linehan WM, Hallahan CW, Lubensky I, Kerr
GS, Hoffman GS, Fauci AS & Sneller MC. (1996). Cyclophosphamide-induced cystitis and bladder cancer in patients with Wegener granulomatosis. Ann Intern Med 124, 477-484.
Tempest HV, Dixon AK, Turner WH, Elneil S, Sellers LA & Ferguson DR. (2004). P2X and
P2X receptor expression in human bladder urothelium and changes in interstitial cystitis. BJU Int 93, 1344-1348.
Teng J, Wang ZY, Jarrard DF & Bjorling DE. (2008). Roles of estrogen receptor alpha and
beta in modulating urothelial cell proliferation. Endocr Relat Cancer 15, 351-364.
Theobald RJ. (1996). Pharmacological studies to examine the source of ATP released by
pelvic nerve stimulation in the feline lower urinary tract. J Auton Pharmacol 16, 111-115.
Theobald RJ, Jr. (1995). Purinergic and cholinergic components of bladder contractility
and flow. Life Sci 56, 445-454. Tobin G. (1995). Muscarinic receptor subtypes in the submandibular gland and the
urinary bladder of the rabbit: in vivo and in vitro functional comparisons of receptor antagonists. J Auton Pharmacol 15, 451-463.
Tobin G, Giglio D & Lundgren O. (2009). Muscarinic receptor subtypes in the alimentary
tract. J Physiol Pharmacol 60, 3-21. Tobin G & Sjogren C. (1995). In vivo and in vitro effects of muscarinic receptor
antagonists on contractions and release of [3H]acetylcholine in the rabbit urinary bladder. Eur J Pharmacol 281, 1-8.
Tobin G & Sjogren C. (1998). Prejunctional facilitatory and inhibitory modulation of
parasympathetic nerve transmission in the rabbit urinary bladder. J Auton Nerv Syst 68, 153-156.
Togashi H, Emala CW, Hall IP & Hirshman CA. (1998). Carbachol-induced actin
reorganization involves Gi activation of Rho in human airway smooth muscle cells. Am J Physiol 274, L803-809.

82
Travis LB, Curtis RE, Glimelius B, Holowaty EJ, Van Leeuwen FE, Lynch CF, Hagenbeek A, Stovall M, Banks PM, Adami J & et al. (1995). Bladder and kidney cancer following cyclophosphamide therapy for non-Hodgkin's lymphoma. J Natl Cancer Inst 87, 524-530.
Tucci P, Bartocci C & Bolle P. (2002). Cyclo-oxygenase- and capsaicin-sensitive afferent
fibres affect Beta-adrenoceptor-evoked response in the rat urinary bladder. Pharmacology 64, 57-62.
Tully BT, Li M, Sun Y, Berkowitz J & Chai TC. (2009). Defects in muscarinic receptor cell
signaling in bladder urothelial cancer cell lines. Urology 74, 467-473. Tyagi S, Tyagi P, Van-le S, Yoshimura N, Chancellor MB & de Miguel F. (2006). Qualitative
and quantitative expression profile of muscarinic receptors in human urothelium and detrusor. J Urol 176, 1673-1678.
Ueda S, Satake N & Shibata S. (1984). Alpha 1- and alpha 2-adrenoceptors in the smooth
muscle of isolated rabbit urinary bladder and urethra. Eur J Pharmacol 103, 249-254.
Waghe M, Westwood R, Nunn G, Kalinowski A & Aldridge A. (1999). Urinary tract toxicity
in rats following administration of beta 3-adrenoceptor agonists. Toxicol Pathol 27, 165-170.
Walden PD, Durkin MM, Lepor H, Wetzel JM, Gluchowski C & Gustafson EL. (1997).
Localization of mRNA and receptor binding sites for the alpha 1a-adrenoceptor subtype in the rat, monkey and human urinary bladder and prostate. J Urol 157, 1032-1038.
Wang P, Luthin GR & Ruggieri MR. (1995). Muscarinic acetylcholine receptor subtypes
mediating urinary bladder contractility and coupling to GTP binding proteins. J Pharmacol Exp Ther 273, 959-966.
Varley C, Hill G, Pellegrin S, Shaw NJ, Selby PJ, Trejdosiewicz LK & Southgate J. (2005).
Autocrine regulation of human urothelial cell proliferation and migration during regenerative responses in vitro. Exp Cell Res 306, 216-229.
Vesela R, Aronsson P & Tobin G. (2010). Functional and morphological examinations of
P1A(1) purinoceptors in the normal and inflamed urinary bladder of the rat. Auton Neurosci.
Wiseman OJ, Fowler CJ & Landon DN. (2003). The role of the human bladder lamina
propria myofibroblast. BJU Int 91, 89-93. Wood R, Eichel L, Messing EM & Schwarz E. (2001). Automated noninvasive
measurement of cyclophosphamide-induced changes in murine voiding frequency and volume. J Urol 165, 653-659.
Yamaguchi O. (2010). Antimuscarinics and overactive bladder: other mechanism of
action. Neurourol Urodyn 29, 112-115.

83
Yamaguchi O, Shishido K, Tamura K, Ogawa T, Fujimura T & Ohtsuka M. (1996). Evaluation of mRNAs encoding muscarinic receptor subtypes in human detrusor muscle. J Urol 156, 1208-1213.
Yamazaki Y, Takeda H, Akahane M, Igawa Y, Nishizawa O & Ajisawa Y. (1998). Species
differences in the distribution of beta-adrenoceptor subtypes in bladder smooth muscle. Br J Pharmacol 124, 593-599.
Yanase H, Wang X, Momota Y, Nimura T & Kawatani M. (2008). The involvement of
urothelial alpha1A adrenergic receptor in controlling the micturition reflex. Biomed Res 29, 239-244.
Yoshida M, Homma Y, Inadome A, Yono M, Seshita H, Miyamoto Y, Murakami S, Kawabe K
& Ueda S. (2001). Age-related changes in cholinergic and purinergic neurotransmission in human isolated bladder smooth muscles. Exp Gerontol 36, 99-109.
Yoshimura N & de Groat WC. (1997). Neural control of the lower urinary tract. Int J Urol
4, 111-125. Yoshimura N, Seki S & de Groat WC. (2001). Nitric oxide modulates Ca(2+) channels in
dorsal root ganglion neurons innervating rat urinary bladder. J Neurophysiol 86, 304-311.
Yoshiyama M & de Groat WC. (2008). The role of vasoactive intestinal polypeptide and
pituitary adenylate cyclase-activating polypeptide in the neural pathways controlling the lower urinary tract. J Mol Neurosci 36, 227-240.
Yu W, Zacharia LC, Jackson EK & Apodaca G. (2006). Adenosine receptor expression and
function in bladder uroepithelium. Am J Physiol Cell Physiol 291, C254-265. Yu Y & de Groat WC. (2008). Sensitization of pelvic afferent nerves in the in vitro rat
urinary bladder-pelvic nerve preparation by purinergic agonists and cyclophosphamide pretreatment. Am J Physiol Renal Physiol 294, F1146-1156.
Zupancic D, Vidmar G & Jezernik K. (2009). Melatonin prevents the development of
hyperplastic urothelium induced by repeated doses of cyclophosphamide. Virchows Arch 454, 657-666.





























