Embed Size (px)
Citation preview

BIOCHEMICAL MEDICINE 30, 295-304 (1983)
Mechanism of Endotoxin-Induced Reduction in the Number of /3-Adrenergic Receptors in Dog Livers: Role
of Phospholipase A
MAW-SHUNG LIU, SANJAY GHOSH, AND GE-FEI KANG
Department of Physiology, St. Louis University School of Medicine, St. Louis, Missouri 63104
Received November 18, 1982
Previous work from this laboratory has shown that endotoxin admin- istration affects the p-adrenergic receptor and its coupling system, adenylate cyclase, in dog livers (1,2). The hepatic p-adrenergic receptor binding capacity was reduced by 39% (1) coupled with a 67% reduction in the isoproterenol (lo-’ h&stimulated adenylate cyclase activity (2) 2 hr fol- lowing endotoxin administration. Since endotoxic shock is associated with disturbances in liver glucose metabolism (3-9), the relationship of the P-adrenergic receptor-adenylate cyclase system to carbohydrate me- tabolism (10,ll) suggests that an understanding of the mechanism whereby endotoxin affects this system may be important.
The receptor is a protein molecule situated in the membrane lipid microenvironment and its proper functioning depends upon the appropriate integration with membrane lipids (12). Any change in the membrane lipid profiLe could affect P-adrenergic receptor dynamics. Limbird and Lefkowitz (13) reported that digestion of erythrocyte membranes with phospholipases A, C, or D reduced the number of p-adrenergic receptors. Since phos- pholipase A activities have been found to increase in liver (14) and heart (15) during endotoxic shock, it is possible that the endotoxin-induced reduction in the p-adrenergic receptor binding capacity is triggered by changes in phospholipase A activity. Therefore, the present study was undertaken to investigate the role of phospholipase A on the altered hepatic p-adrenergic receptor dynamics during endotoxic shock.
METHODS
Mongrel dogs of either sex weighing from 18 to 23 kg were used. They were divided into two groups: control and endotoxic. Endotoxic shock was induced by a single intravenous injection of 1 mg/kg endotoxin (lipopolysaccharide B from Escherichia co/i), and the endotoxic dogs
295 0006-2944/83 $3.00
Copyright 8 1983 by Academic &ss, Inc. All rights of reproduction in any form reserved.

296 LIU. GHOSH, AND KANG
were killed 2 hr after endotoxin administration ( 1). Control dogs received no treatment. Control animals were fasted for 4.5 hr while the endotoxic animals were fasted for 43 hr prior to and for 2 more hr after endotoxin administration. Water was permitted freely. All animals were anesthetized with 2% chloralose in 20% urethane (4 ml/kg, iv) and livers were removed and chilled immediately in ice-cold physiological saline for the isolation of plasma membranes.
Hepatic plasma membranes were prepared by the method of Neville (16) with minor modification as described by Liu and Ghosh ( 1). The purity of the liver plasma membranes was assessed by measurements of the activities of adenylate cyclase (60 and 420 pmoles/mg min in the absence and presence of 10 PM ( - )-isoproterenol, respectively) and sodium dodecyl sulfate-activated ouabain-inhibitable (Na’ + K+)-ATPase (72 nmoles/mg min).
fi-Adrenergic receptor binding studies were carried out by using ( - I- ]3H]dihyroalprenolol ([3H]DHA). a /3-adrenergic antagonist, as a labeled ligand according to the methods of Mukhetjee et al. (17) and Smith and Clark (18) with minor modification as described by Liu and Ghosh (1). The standard assay mixture in a final volume of 0.2 ml contained 10 rnbr MgCl?, 50 mM Tris-HCl (pH 7.5). and freshly prepared liver plasma membranes containing 200 pg protein. The reaction was started by the addition of 5 nM of r3H]DHA (30,000 cpm) and proceeded for 20 min at 37°C. For Scatchard analysis different concentrations of [‘HIDHA ranging from 0.5 to 10 nM were employed. At the end of each incubation, the reaction mixture was diluted with ice-cold washing buffer (10 rnhl MgCl: and 50 mM Tris-HCl, pH 7.5), filtered through a Whatman GF/C filter paper, and rewashed three times with washing buffer. The filter papers were dried and the radioactivity was then determined. The specific binding was defined as the bound radioactivity displaceable by IO ’ M of ( - I- alprenolol. It should be noted that the kinetics of ]‘H]DHA binding to dog liver plasma membranes have been characterized in this laboratory (1). Other experimental details regarding digestion of membranes with exogenous phospholipase AZ and the addition of phosphatidylcholine (PC) are stated in the legend to each individual figure.
Endogenous phospholipase A activity was measured as described by Liu and Takeda (IS) using liver homogenates as an enzyme source in the presence of 2 mM Ca”.
Total lipids were extracted from liver plasma membranes by the method of Bligh and Dyer (19). Phospholipids were separated by two-dimensional thin-layer chromatography on silica gel H plates and the phosphorus content of each individual phospholipid was then determined (20).
The protein content of liver plasma membranes was measured by the method of Lowry rt al. (21). Student’s t test (Figs. 3 and 5. Table I)

&RECEPTOR AND PHOSPHOLIPASE IN ENDOTOXIC LIVER 297
and one-way analysis of variance (Figs. 1, 2, and 4) were used for the statistical analyses of the data.
(-)-/j~opyl-2,3-~H]Dihydroalprenolol HCl (49.4 Ci/mmole) was pur- chased from New England Nuclear Corporation. Phospholipase A2 (from porcine pancreas), L-a-phosphatidylcholine (dipalmitoyl), (-)-alprenolol (tartrate salt), and silica gel H were products of Sigma Chemical Company. Endotoxin (lipopolysaccharide isolated from E. coli) was obtained from Difco. Other chemicals and reagents were of analytical grade.
RESULTS AND DISCUSSION
Figure 1 shows the effect of exogenous phospholipase A2 digestion on the specific binding of 13H]DHA in liver plasma membranes isolated from control dogs. Exogenous phospholipase A2 had a concentration-dependent inhibitory effect on the specific binding of [3H]DHA in the absence of bovine serum albumin: the specific binding was inhibited by 37.1% (Z’ < 0.01) at 0.1 unit/200 ,ug protein, and the extent of inhibition was increased to 68.3% (P < 0.01) when the concentration of exogenous phospholipase AZ was increased to 0.5 unit/200 pg protein. In the presence of 1% bovine serum albumin, the specific binding was also inhibited by increasing concentrations of exogenous phospholipase A,: the inhibitions
2 250-
\E - -BSA
JG - +BSA
$200-
b s 0” ‘0 150- ‘0
g 0 IOO- % 3 9 2 50- **p<o.o1
8 :: 8 h 0 ' 1 I I c
0 0.1 0.2 0.3 0.4 0.5 Phospholipase A2 (U/O.2 ml)
FIG. 1. The effect of exogenous phospholipase A, digestion on the specific binding of (-)-[‘HI-dihydroalprenolol in liver plasma membranes isolated from control dogs. Binding assays were carried out as described under Methods. Exogenous phospholipase A, digestion was performed by incubating 200 pg membrane protein with different amounts of exogenous phospholipase AZ, as indicated on the abscissa, for 30 min at 28°C. At the end of each digestion, the mixture was centrifuged at 30,OOOg for 15 min and the resulting pellet was suspended in washing buffer and then used for binding assays. (0) Indicates experiments carried out in the absence of, while (0) represents those in the presence of 1% bovine serum albumin (BSA) during digestion.

298 LIU, GHOSH, AND KANG
were 20.5% (P < 0.05), 37.3% (P < O.Ol), and 44.9% (P < 0.01) at 0.1, 0.2, and 0.5 units/200 pug membrane protein, respectively. Since albumin serves as an absorbent for fatty acids and lysophosphatides, the hydrolyzed products of phospholipase A?, the data presented here indicate that the inhibitory effect of phospholipase AZ digestion on 13H]DHA binding is not merely a secondary response to the increases in the concentrations of fatty acids and lysophosphatides.
Limbird and Lefkowitz (13) reported that treatment of frog erythrocyte membranes with phospholipases A, C, or D decreased /3-adrenergic receptor binding capacity. Thang et al. (12) found that solubilization of rabbit heart membranes with Lubrol PX, a lipid perturbing agent, reduced the number of /3-adrenergic receptors and the reduction in receptor number was reversible by brain phospholipid extracts. These observations together with those reported in this study demonstrate the role of phospholipids in the alteration of /3-adrenergic receptor binding.
Based on the data presented in Fig. 1, we have ruled out the possibility that the inhibition of the specific binding of [3H]DHA as a result of exogenous phospholipase A2 digestion is due to the hydrolyzed products, fatty acids. and lysophosphatides. It seems likely then that the inhibition is due to the decrease in phospholipids which act as substrates for phos- pholipase A,.
Accordingly, the composition of phosphoglycerides in liver plasma membranes of control dogs digested with exogenous phospholipase A, and that of endotoxic (2-hr postendotoxin) dogs was analyzed and the results were correlated with changes in endogenous phospholipase A activities (Table 1). As shown in Table 1, digestion of control membranes with exogenous phospholipase A? decreased PC and phosphatidylethan- olamine (PE) contents by 50.6 and 51.2%, respectively, coupled with a 1Zfold increase in lysophosphatidylcholine (LPC) and an 8.4-fold increase in lysophosphatidylethanolamine (LPE) levels. Endotoxin administration decreased PC and PE contents by 21.4 and 23.8%. respectively. coupled with a 2. l-fold increase in LPC and a 1.6fold increase in LPE levels (Table 1). These data demonstrate that changes in membrane phospholipid composition induced by endotoxin administration resemble those in control membranes digested with exogenous phospholipase A?. In addition. the endogenous phospholipase A activity was increased by 73.5% 2 hr following endotoxin administration (Table 1). Taking all these findings together, it is suggested that changes in membrane phospholipid composition induced by in viva administration of endotoxin are a result of phospholipase A
activation. It should be pointed out that although PC and PE contents were both
decreased by the in viva administration of endotoxin or by the in vitro digestion of the control membranes with exogenous phospholipase Al,
the decrease in PC content was greater than that in PE. i.e.. 713 vs I?,?
P TA
BLE
1 ii
THE
COIW
XITI
ON
OF
PHOS
PHOL
IP~D
S AN
D TH
E EN
D~CE
NOUS
PH
OSPH
OLIP
ASE
A AC
TIVI
TY
IN
DOG
LIVE
RS
FOLL
OWIN
G EI
THER
TH
E IN
VI
TRO
DI
GEST
ION
i2
WIT
H EX
OGEN
OUS
PHOS
PHOL
IPAS
E A,
OR
THE
IN
VIVO
AD
MIN
ISTR
ATIO
N OF
EN
D~TO
XIN
2
Phos
phol
ipid
co
mpo
sitio
n w
(nm
ole/
mg
prot
ein)
En
doge
nous
ph
osph
olip
ase
A 8
PC
LPC
PE
LPE
(nm
olef
mg
hr)
z
Con
trol
421
k 15
.6
13.9
”
1.8
248
k 10
12
k
1.8
3.4
2 0.
3 E
(II =
6)
2
Con
trol
+ 20
8 f
21*
183
zt 2
4*
121
+ 14
* 11
3 -c
8.8
* N
ot
dete
rmin
ed
3 ex
ogen
ous
PLA,
(-
50.6
%)
( + 1
2-fo
ld)
(-51.
2%)
(+8.
4-fo
ld)
F (n
=
6)
Endo
toxic
33
1 k
17*
43.4
k
4.8*
18
9 -c
7*
28.7
k
3.8*
5.
9 k
0.4*
%
(2 h
r po
sten
doto
xin)
(-21.
4%)
( + 2
.1 -f
old)
(-2
3.8%
) ( +
1 .C
fold
) (+
73.5
%)
5
(n
= 7)
L2
Not
e.
Valu
es
are
mea
ns
c SE
of
the
mea
n.
Exog
enou
s ph
osph
olip
ase
Ar (
PLAL
; 0.
2 un
it/20
0 pg
pr
otei
n)
dige
stio
n is
des
crib
ed
in F
ig.
1 us
ing
8
cont
rol
liver
pl
asm
a m
embr
anes
in
th
e ab
senc
e of
alb
umin
. Ex
ogen
ous
phos
phol
ipas
e A
activ
ity
is m
easu
red
as d
escr
ibed
un
der
Met
hods
us
ing
hom
ogen
ates
as
an
enzy
me
sour
ce.
The
abbr
evia
tions
ar
e PC
, ph
osph
atid
ylcho
line;
LP
C,
lyso
phos
phat
idyl
chol
ine;
PE
, ph
osph
atid
yleth
anol
amin
e;
z F;
and
LPE,
iy
soph
osph
atid
ylet
hano
lam
ine.
*
P <
0.01
. E s:
W

300 LIU. GHOSH, AND KANG
nmoles/mg decrease for PC vs PE respectively, after exogenous phos- pholipase AZ digestion; and 90 vs 59 nmoles/mg decrease for PC vs PE. respectively, following endotoxin administration (Table 1). Because of the relatively greater loss in PC content, the effect of the addition of PC on the specific binding of [3H]DHA in control liver membranes predigested with exogenous phospholipase A? was studied and the results are shown in Fig. 2. Exogenous phospholipase A? digestion caused a 33.8% inhibition on [3H]DHA binding, and the inhibition was completely reversed by the addition of PC (0.1-0.5 mM) (Fig. 2). These data suggest that the decrease in [3H]DHA binding caused by exogenous phospholipase Az digestion in control liver membranes is a result of a decrease in membrane phospholipid content, particularly in PC. A similar contention has been advanced regarding rat brain opiate receptor binding (22,23).
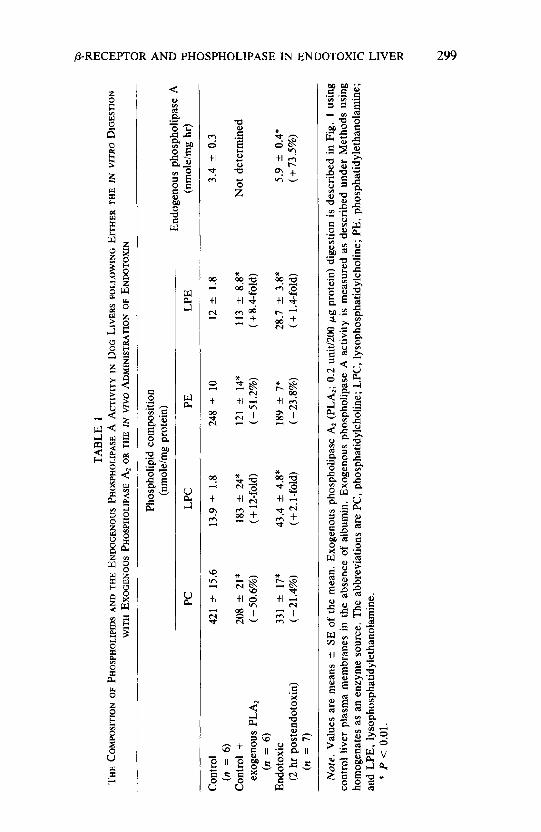
In order to clarify further the nature of phospholipase A? action and the role of PC on /I-adrenergic receptors, the Scatchard plots of [‘HIDHA binding in control liver plasma membranes under various experimental conditions were analyzed and the results are shown in Fig. 3. Control liver membranes without exogenous phospholipase AZ digestion had a K, (dissociation constant) value of 8.3 t I.2 nM and a R,,, (maximal binding capacity) of 626 -+ 51 fmoles/mg protein. Exogenous phospholipase A2 digestion decreased the B, by 31.7% (P < 0.05) to 430 k 40 fmoles/mg protein with little change in Kd value (8. I i 0.6 nM). Addition of PC to the control membranes predigested with exogenous phospholipase ‘4: restored the B,,, (610 t 52 fmoles/mg protein) to the predigested value with no significant change in the Kd (8.8 +- I. 1 nM). These data demonstrate that digestion of control membranes with exogenous phospholipase A,
PLA2W0.2 ml1 0 02 0.2 02 02
PC(mM) 0 0 01 02 05
FIG. 2. The effect of phosphatidylcholine addition on the specific binding of (- )-[‘H]dihydroalprenolol in control liver plasma membranes predigested with exogenous phospholipase A*. Phospholipase Az (PLA,) digestion and binding assays were carried out as described in Fig. I. Phosphatidylcholine (PC) was suspended in 20% ethanol and sonicated for 30 min at 4°C and the membranes were incubated with various concentrations of PC (final ethanol concentration was 1%) for 30 min at 4°C prior to binding assays. Vertical bars indicate mean t SEM. Number of experiments is shown in parentheses.
P-RECEPTOR AND PHOSPHOLIPASE IN ENDOTOXIC LIVER 301
o- ’ ’ 0 100 200 300 400 500 600
1_ 700
13Hl i-1 - Dihydroalprenolol Bound (f moles /mg 1
FIG 3. The effect of phosphatidylcholine addition on the Scatchard plots of (-)-[‘H]dihydroalprenolol binding in control liver plasma membranes predigested with exogenous phospholipase A,. The binding assays were carried out as described under Methods. Exogenous phospholipase A, (PLA,; 0.2 unit/O.2 ml) digestion and phosphati- dylcholine (PC; 0.2 mM) addition are described in Figs. 1 and 2. Triangles represent the results obtained using control liver membranes (n = 6). empty circles represent results obtained using control liver membranes predigested with PLA, (n = 7), and filled circles indicate the results obtained using control liver membranes predigested with PLA, and assayed in the presence of PC (n = 7). Vertical and horizonatal bars represent standard errors of the mean.
reduced the number but not the affinity of /3-adrenergic receptors and the reduction in receptor number was reversible by PC.
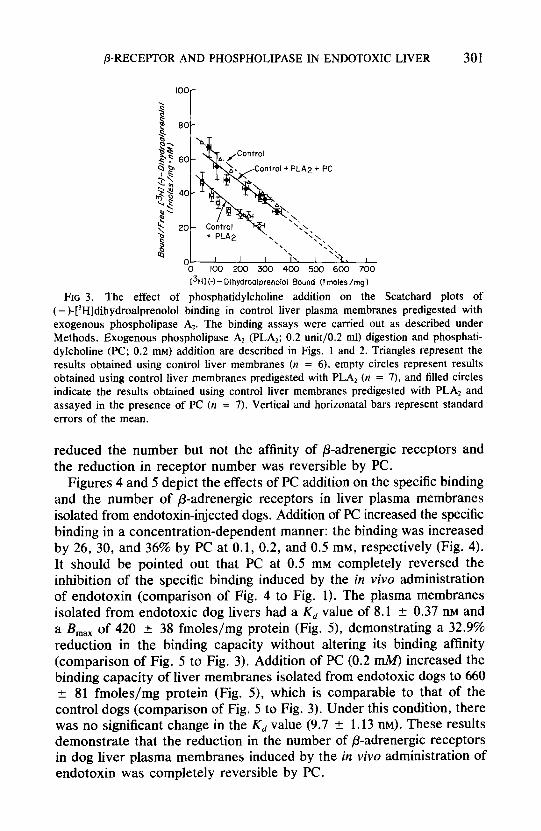
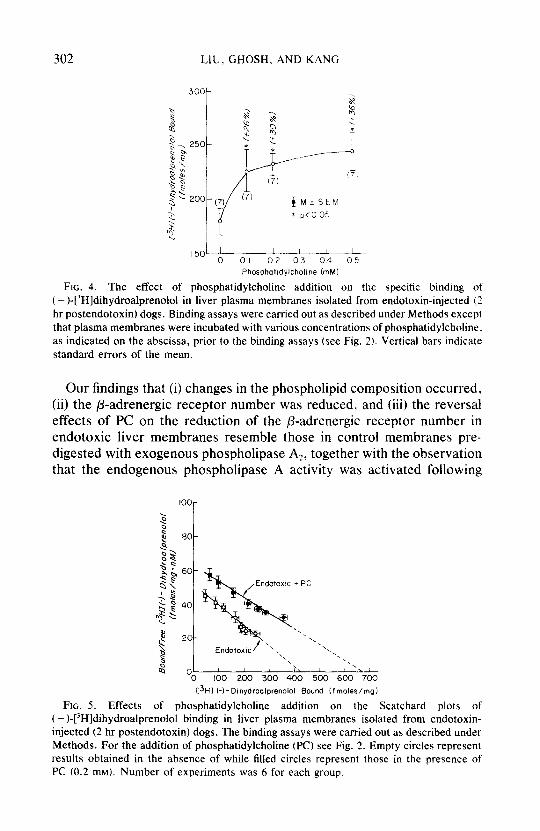
Figures 4 and 5 depict the effects of PC addition on the specific binding and the number of p-adrenergic receptors in liver plasma membranes isolated from endotoxin-injected dogs. Addition of PC increased the specific binding in a concentration-dependent manner: the binding was increased by 26, 30, and 36% by PC at 0.1, 0.2, and 0.5 mM, respectively (Fig. 4). It should be pointed out that PC at 0.5 mM completely reversed the inhibition of the specific binding induced by the in vivo administration of endotoxin (comparison of Fig. 4 to Fig. 1). The plasma membranes isolated from endotoxic dog livers had a Kd value of 8.1 + 0.37 nM and a &,, of 420 rf: 38 fmoleslmg protein (Fig. 5), demonstrating a 32.9% reduction in the binding capacity without altering its binding affinity (comparison of Fig. 5 to Fig. 3). Addition of PC (0.2 mM) increased the binding capacity of liver membranes isolated from endotoxic dogs to 660 + 81 fmoles/mg protein (Fig. 5), which is comparable to that of the control dogs (comparison of Fig. 5 to Fig. 3). Under this condition, there was no significant change in the Kd value (9.7 + 1.13 nM). These results demonstrate that the reduction in the number of p-adrenergic receptors in dog liver plasma membranes induced by the in vivo administration of endotoxin was completely reversible by PC.
302 LIU, GHOSH. AND KANG
MiSEhl
p<o 05
,50/L-- 0 01 02 03 04 05
Phosphotldylchollne (mM1
FIG. 4. The effect of phosphatidylcholine addition on the specific binding of t - )-[3H]dihydroalprenolo1 in liver plasma membranes isolated from endotoxin-injected (2 hr postendotoxin) dogs. Binding assays were carried out as described under Methods except that plasma membranes were incubated with various concentrations of phosphatidylcholine, as indicated on the abscissa, prior to the binding assays (see Fig. 2). Vertical bars indicate standard errors of the mean.
Our findings that (i) changes in the phospholipid composition occurred, (ii) the P-adrenergic receptor number was reduced, and (iii) the reversal effects of PC on the reduction of the fl-adrenergic receptor number in endotoxic liver membranes resemble those in control membranes pre- digested with exogenous phospholipase A?, together with the observation that the endogenous phospholipase A activity was activated following
80-
60-
40-
20-
OL 0
C3Hl k- DIhydroalprenolol Bound (fmoles/mgi
FIG. 5. Effects of phosphatidylcholine addition on the Scatchard plots of ( - )-[“H]dihydroalprenolol binding in liver plasma membranes isolated from endotoxin- injected (2 hr postendotoxin) dogs. The binding assays were carried out as described under Methods. For the addition of phosphatidylcholine (PC) see Fig. 2. Empty circles represent results obtained in the absence of while filled circles represent those in the presence of PC (0.2 mM). Number of experiments was 6 for each group.

P-RECEPTOR AND PHOSPHOLIPASE IN ENDOTOXIC LIVER 303
endotoxin administration, unequivocally demonstrate the role of phos- pholipase A in the reduction of the number of P-adrenergic receptors in dog livers during endotoxic shock.
SUMMARY
The role of phospholipase A on the endotoxin-induced reduction in the number of /3-adrenergic receptors in dog liver plasma membranes., was investigated. The results show that digestion of control liver plasma membranes with exogenous phospholipase A2 (0.2 unit/200 pg protein) decreased the specific binding of ( -)-[3H]dihydroalprenolol by 37.3% (P < 0.01) and reduced the number of receptor sites by 31.7% (P < 0.05). These decreases in the specific binding and the number of P-adrenergic receptors were completely reversible by the addition of phosphatidylcholine (0.2 mM). Endotoxin administration (2 hr postendotoxin) decreased the specific binding by 36% (P < 0.05) and reduced the number of @ulrenergic receptors by 33% (P < O.OS), and these decreases were completely re- versible by the addition of 0.2 mM phosphatidylcholine. Digestion of control liver membranes with exogenous phospholipase A2 decreased phosphatidylcholine and phosphatidylethanolamine levels by 50.6 and 5 1.2%, respectively, but increased lysophosphatidylcholine and lyso- phosphatidylethanolamine levels by 12- and 8.4-fold, respectively. Endo- toxin administration decreased phosphatidylcholine and phosphatidyle- thanolamine contents by 21.4 and 23.8%, respectively, but increased lysophosphatidylcholine and lysophosphatidylethanolamine contents by 2. I- and 1.4-fold, respectively. In addition, endotoxin administration in- creased endogenous phospholipase A activity by 73.5%. Based on these results, it is suggested that the decreases in the specific binding and the number of p-adrenergic receptors in dog livers during endotoxic shock are a result of phospholipase A activation.
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health Grants GM-31664 and HL- 30080.
REFERENCES 1. Liu, M. S., and Ghosh, S., Amer. J. Physiol. 244, R718 (1983). 2. Ghosh, S., and Liu, M. S., Amer. J. Physiol., in press. 3. Filkins, J. P., and Cornell, R. P., Amer. J. Physiol. 227, 778 (1974). 4. Lanoue, K. F., Mason, A. D., and Daniels, J. P., Metabolism 17, 606 (1968). 5. Williamson, J. R., Refino, C., and Lanoue, K., in “Energy Metabolism in Trauma”
(R. Porter and J. Knight, Eds.), pp. 145-154. Churchill, London, 1970. 6. Liu, M. S., and Sharma, C., Amer. J. Physiol. 240, RIO (1981). 7. Giger, O., and McCallum, R. E., Amer. J. Physiol. 231, 1285 (1976). 8. McCallum, R. E., and Berry, L. J., Infect. Immun. 7, 642 (1973). 9. Hamosh, M., and Sapiro, B., Bn’r. J. Exp. Pathol. 41, 372 (1960).

304 LIU. GHOSH, AND KANG
IO. Sutherland, E. W.. Science 177. 401 (1972). 11. Exton, J. H., Biochem. Pharmacol. 28, 2737 (1979). 12. Thang, N. X., Borsodi. A.. and Wolleman. M.. Biuchem. Phurmucol. 29, ?7Y1 (198Ob. 13. Limbird. L. E.. and Lefkowitz. R. J.. Mol. Phurmtrcwl. 12. 55Y f 1976)
14. Conde. C.. Barreno. P. G.. Municio. A. M.. and Suarer. .A.. FEB.5 Ix/r 127. I Ii
15. Liu. M. S.. and Takeda, H., Biochem. Mrd. 28, 61 t 1982). 16. Neville. D. M.. Jr.. Biochim. Bioph~~s. Acttr 154, 540 (196X).
17. Mukhejee. C., Caron, M. G.. Covet-stone. M.. and Letkowitz, R. J.. J. Bi~,l. C‘lre/~. 250, 4869 (1975).
18. Smith. P. B.. and Clark. G. F.. Biochim. Biophys. Aua 633. 274 (1980). 19. Bligh. E. G.. and Dyer. W. J.. Canad. J. Binchcwz. Ph~siol. 37. 91 I (19.59). 20. Rouser. G.. Fleischer. S., and Yamamoto. A.. Lipid 5, 494 tlY70). 21. Lowry. 0. H., Rosebrough. N. J.. Farr. A. L.. and Randall. R. J.. ./. Biol. Chrm
193, 265 (1951).
22. Lin. H. K.. and Simon, E. J.. Nature (Londo~~ 271. 383 !IY7XI.
23. Abood. L. G.. Butler. M.. and Reynolds. D.. Mol. Phormacol. 17. 190 f IY80).





























