Embed Size (px)
Citation preview

Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004), pp. 1912–1921 ( C© 2004)
Malignant Transformation of the MouseAnorectal Epithelium Induced by an Inoculated
Human Cancer Cell Line
HIROYUKI KUWANO, MD,* TATSUYA MIYAZAKI, MD,* SOICHI TSUTSUMI, MD,*MINORU FUKUCHI, MD,* KEN-ICHI NOMOTO, MD,* TATSUO SHIMURA, MD,* ERITO MOCHIKI, MD,*
HIROYUKI KATO, MD,* TAKAYUKI ASAO, MD,* and YASUSHI TOH, MD†
Four kinds of human cancer cell lines and one mouse caner cell line were inoculated into thesubepithelial area of the anorectum of female nude mice. Among the cell lines, two cell lines(KATO III and Lu 135) showed the potential enforcement of atypical changes in the adjacent mouseanorectal epithelium. Moreover, the submucosal invasion of the malignant transformed cells ofthe mouse epithelium was demonstrated in specimens obtained from three KATO III-inoculatedmice. This exciting and novel phenomenon clearly demonstrates the need to change the presentgeneral concept of a single-cell origin of cancer tissue. This valuable and novel discovery maychange the basis of oncology research while also providing new ideas for projects to investigatethe mechanisms of carcinogenesis from several aspects such as molecular biology, cell biology, andpathology. Moreover, the novel experimental design itself is also extremely useful as a simple modelfor investigating the mechanisms of oncogenesis.
KEY WORDS: malignant transformation; paracrine; paratransformation; KATO III; Lu 135.
It is generally considered that malignant tumors growdue to multiplication of preexisting cancer cells and thenspread by invasion and destruction of the surrounding tis-sues (1). With tumors of B cell lineage, monoclonality ismost easily established by the fact that all cells synthesizethe same immunoglobulin. To prove this concept, therehave been some demonstrations of this phenomenon inthe literature.
One demonstration of this comes from an analysis of thecells’ DNA. In almost all patients with “chronic myeloge-
Manuscript received June 1, 2004; accepted July 7, 2004.From the *Department of General Surgical Science (Surgery I),
Gunma University Graduate School of Medicine, Maebashi, and†Department of Gastroenterologic Surgery, National Kyushu CancerCenter, Fukuoka, Japan.
Address for reprint requests: Hiroyuki Kuwano, MD, Departmentof General Surgical Science (Surgery I), Gunma University GraduateSchool of Medicine, 3-39-22 Showa-machi, Maebashi, Gunma, 371-8511 Japan; [email protected].
nous leukemia,” for example, leukemic white blood cellsare distinguished from the normal cells by a specific chro-mosomal abnormality (the so-called Philadelphia chro-mosome). When the DNA at the site of translocation iscloned and sequenced, the site of breakage and rejoiningof the translocated fragments is found to be identical inall the leukemic cells in any given patient, but this differsslightly from one patient to another, as expected if eachcase of leukemia arises from a unique accident occurringin a single cell (2, 3).
Another way to show that a cancer has a monoclonalorigin is by exploiting the phenomenon of X chromosomeinactivation. A normal woman is a random mixture, or mo-saic, of two classes of cells—those in which the paternalX chromosome is inactivated and those in which the ma-ternal X chromosome is inactivated. The inactivation ofone or the other of the X chromosomes occurs at randomin each cell early in embryonic development, but once the
1912 Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004)0163-2116/04/1200-1912/0 C© 2004 Springer Science+Business Media, Inc.

MALIGNANT TRANSFORMATION OF MOUSE ANORECTAL EPITHELIUM
choice has been made it is irreversible, so that when acell divides it passes on its own state of X inactivation toits daughters. Consequently, the state of X-chromosomeinactivation—maternal or paternal—can be used as a her-itable marker to trace the lineage of cells in the body. Inthe great majority of tumors that have been analyzed—both benign and malignant—all the tumor cells have beenfound to have the same inactivated X chromosome, thusstrongly suggesting that they are derived from a single de-ranged cell (4, 5). Recently, Tatematsu et al. demonstratedthe primary monoclonal and secondary polyclonal growthof colon neoplasms in C3H/HeN ↔ BALB/C chimericmice treated with 1,2-dimethylhydrazine by the immuno-histochemical detection of C3H strain-specific antigen andsimple sequence length polymorphism analysis of DNA(6). They concluded that in murine colon carcinogenesis,when multiple tumors are induced, the incidence of ap-parently polyclonal tumors resulting from the coalescenceof adjacent monoclonal carcinogenesis may be relativelyhigh. To extend this argument, malignant cells require theacceptance of autocrine stimulation as suggested by Spornand Todaro (7). The premise is that for cancer cells, asfor normal cells, proliferation is not the norm but occursin response to a sequence of polypeptide growth factorswhich bind to high-affinity membrane receptors. While inhealthy tissue in general one cell synthesizes the growthfactors required by another (some of the growth factorsonly act locally at the site of production), a characteristicof cancer cells is that they constitutively synthesize growthfactors (referred to as transforming growth factor; TGF)for which they have membrane receptors and to whichthey themselves respond.
In order to elucidate the histogenesis of esophageal can-cer, we previously demonstrated a high incidence of eitherglandular or mucus-secreting components in esophagealsquamous cell carcinoma (8), a frequent coexistence of in-traepithelial carcinoma contiguous to the main squamouscell carcinoma in early esophageal cancer (9), a close re-lationship between multiplicity and the existence of intra-epithelial carcinoma with squamous cell carcinoma (10),and a high incidence of the coexistence of intra-epithelialcarcinoma and glandular differentiation, particularly inearly cancers (11). The above findings support the conceptof either multicentric or field carcinogenesis in esophagealcancer.
However, although a few reports have suggested aparacrine mechanism of some genes in the transforma-tion of epithelial cells in vitro (12), there have so far beenno reports presenting any direct evidence of the malignanttransformation of the cells induced by another type of can-cer cell in vivo. Recently, we established a model of ex-perimental lymph node metastasis by intrarectal implan-
tation of human cancer cells in nude mice, and this modelis considered to enable us to investigate the process oflymph node metastasis and the effect of anticancer agentssimulating human rectal cancers (13). Among the rectalspecimens obtained in this experiment, either rectal or analepithelium adjacent to the inoculated tumor cell lines of afew specimens showed mild atypia. We therefore startedto perform systematic experiments to investigate the ma-lignant transformation of the mouse anorectal epitheliumadjacent to the inoculated tumor cell lines. This reportdemonstrates novel evidence regarding the phenomenonof “paratransformation.”
MATERIALS AND METHODS
Animals. Female nude mice (BALB/cAnNCrj-nu) aged8 weeks were used. They were purchased from Charles RiverJapan Inc., Kanagawa. All animals were housed at the Laboratoryfor Animal Experiments, Research Institute, Gunma UniversityFaculty of Medicine, under laminar airflow conditions. Housingwas temperature controlled with a 12-hr light and dark cycle.The procedures used in this study were approved by the Re-view Committee on Animal Use at Gunma University, Maebashi,Japan.
Cell Lines. Four human cancer cell lines and one micecell line were used: MKN45, HT-29, KATO III, Lu 135, andColon-26. MKN45 cells were derived from undifferentiated gas-tric adenocarcinoma (14). HT-29 cells were derived from well-differentiated colon adenocarcinoma (15). KATO III cells werederived from signet-ring cell types of gastric adenocarcinoma.The spanning coding sequence of the p53 gene is deleted inKATO III cells, and so KATO III cells do not have mRNA andprotein of p53 (16). Lu 135 cells derive from a small cell type oflung carcinoma. This line has a point mutation in exon 7 of p53(17). Colon 26 was derived from BALB/c mouse undifferentiatedcolon adenocarcinoma induced by N -nitroso-N -methylurethane(18). The cells were cultured in RPMI-1640 medium (NissuiPharmaceutical Co., Ltd.) supplemented with 10% fetal bovineserum. The cells were passaged when they became confluent.
Technique. Log-phase cancer cells were harvested with1 mM EDTA in phosphate-buffered saline (PBS), washed threetimes with PBS, and then resuspended at a cell density of1 × 107 ml in PBS containing 500 µg/ml of Matrigel basementmembrane matrix (Becton Dickinson Labware, Two Oak Park,Bedford, MA). Tumor cells (1 × 106) were suspended in 0.1 mlof PBS containing 50 µg of Matrigel to prevent the suspen-sion from leaking out of the rectum, and the cells were slowlyinjected submucosally with a 29-gauge needle as described pre-viously (13). As a negative control, only PBS containing 50 µgof Matrigel was injected into five mice. Furthermore, to examinewhether or not the tumor cell content affects the mouse rectalmembrane, cell lysate was injected into five mice. Tumor cells(1 × 106) were suspended in PBS. Subsequently, the cells werebroken up by sonication using a probe sonicator W-220F (HEATSYSTEMS-Ultrasonic Inc., Plainview, NY) and injected by thesame method.
Measurement. The tumor volume was calculated by the fol-lowing formula: tumor volume (mm3) = 1/2 × (long diameter) ×(short diameter2).
Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004) 1913

KUWANO ET AL.
Selection of Cell Line. Regarding screening, all five celllines were injected into the mice. Five mice were sacrificed at4–6 weeks when the tumors had grown sufficiently. The anus andrectum with the tumors, were excised fixed in 10% phosphate-buffered formalin for 24 hr at room temperature, and then em-bedded in paraffin.
Evaluation of Tumor Growth. Six mice were then sacri-ficed, at 1, 2, 4, 6, 8, and 10 weeks after tumor implantation,using KATO III. Groups of five mice each were sacrificed at dif-ferent times, 1, 2, 4, 6, 8, and 10 weeks after tumor implantation,using Lu 135. The extracted specimens were all treated in thesame manner.
Immunohistochemistry. Formalin-fixed, paraffin-embeddedtissues were cut into 3-µm sections and then stainedwith hematoxylin–eosin (HE). In an immunohistochemi-cal study, sections were stained by the avidin–biotin–peroxidase complex (ABC) method (Vectastain Laboratories,Inc., Burlingame, CA) using antibodies against p53 antibody(CM5; Novocastra, Newcastle, UK) as described previously(19).
Evaluation for p53. Immunostaining to detect p53 in mouserectal epithelium surrounding the tumor was graded as positivewhen >1% of the cells were stained or as negative when <1%were stained. The anorectal epithelium was classified as either asquamous epithelium or a glandular epithelium.
RESULTS
No instances of operative mortality were observed, andthere was also no postoperative mortality or complica-tions. In several experiments, implanted tumor cells de-veloped at the site of direct injection and formed a nodulein all mice.
Selection of Cell LinesRegarding screening, all five cell lines were injected
into the mice. Five mice injected with colon 26 were sac-rificed at 4 weeks. In addition, the group of five miceinjected with the other cells were killed at 6 weeks. Allmice had rectal tumors. Histopathologically, in the low-power field, implanted cancer cells mainly developed inthe submucosal layer, with involvement of the muscularispropria and submucosa. Anorectal epithelium of controlmice showed no change and no atypism. Mice injectedwith the broken cells also did not show a rectal tumoror any atypical membrane. Anorectal epithelium injectedwith MKN45, HT-29, and Colon 26 showed no atypicalchanges either. The specimens in two cases of five miceimplanted with KATO III revealed atypical change of themouse glandular epithelium. In one case of five mice im-planted with Lu 135, the same changes appeared (Table 1).As a result, further serial experiments were performed us-ing the cell lines KATO III and Lu 135. The volume of theinoculated tumors increased in size with time in these celllines as well as in others (Figure 1).
TABLE 1. ATYPICAL CHANGE IN THE MOUSE RECTAL MEMBRANE
Cell line
KATOIII Lu 135 MKN45 HT-29 Colon-26Site (n = 5) (n = 5) (n = 5) (n = 5) (n = 5)
Squamous 0 0 0 0 0epithelium
Glandular 2 1 0 0 0epithelium
Evaluation of Transformation in Tumor GrowthHistopathological Study. As the glandular epithelium
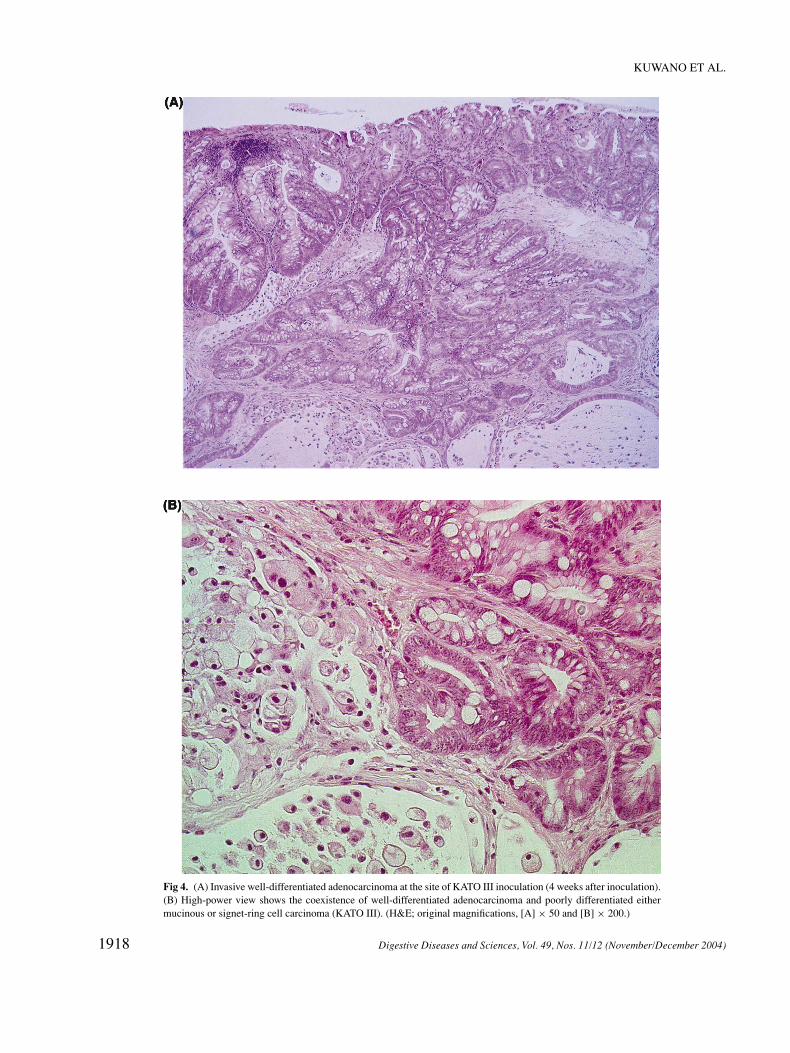
implanted with KATO III or Lu 135 demonstrated atypicalchanges, we investigated the changes with time in the miceimplanted with KATO III and Lu 135 (Table 2). Atypicalchanges in the mice implanted with KATO III appearedat 2W (weeks) and the incidence of atypical epitheliumwas 33% at this time. The incidence of an atypical ep-ithelium tended to increase with the passage of time. Thetotal incidence was 47.2% and the incidence after 4W was62.5%. However, there were some cases without any atyp-ical epithelial changes even in the late phase after KATOIII inoculation (Figure 2). Atypical changes in the glan-dular epithelium of mice implanted with Lu 135 appearedat 2W and the incidence of atypical epithelium was 20%at this moment. The total incidence was 26.7% and the in-cidence after 4W was 35%. Moreover, in two specimensobtained from mice sacrificed 2 and 4 weeks after theinoculation of KATO III, there were foci of invasive ade-nocarcinoma (Figures 3 and 4 and Table 2). In the formerspecimen, there was an area of invasive squamous cell car-cinoma with squamous epithelial atypia concomitant withinvasive adenocarcinoma, although there were no foci ofdefinite atypia in the squamous epithelium of other mousespecimens.
Fig 1. Growth curves of KATO III and Lu 135 inoculated in the mouseanorectal region.
1914 Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004)

MALIGNANT TRANSFORMATION OF MOUSE ANORECTAL EPITHELIUM
TABLE 2. ATYPICAL CHANGE IN TUMOR GROWTH
(A) KATO III
Site 1W (n = 6) 2W (n = 6) 4W (n = 6) 6W (n = 6) 8W (n = 6) 10W (n = 6)
Squamous epithelium 0 0 0 0 0 0(Advanced cancer) (1)*
Glandular epithelium 0 2 3 4 5 3(Advanced cancer) (1)* (1)
(B) Lu 135
Site 1W (n = 5) 2W (n = 5) 4W (n = 5) 6W (n = 5) 8W (n = 5) 10W (n = 5)
Squamous epithelium 0 0 0 0 0 0Glandular epithelium 0 1 2 2 2 1
*Coexistence of advanced adenocarcinoma and squamous cell carcinoma.
Immunohistochemical Study. Atypical glandular ep-ithelium in most cases showed expression of p53 protein(Figure 3C). In the cases implanted with KATO III, expres-sion of p53 was found in the glandular epithelium fromthe 2W time point and the incidence of p53 positivity was33% (Table 3). The total incidence of p53 positivity inglandular epithelium was 36.1% and the incidence after4W was 45.8%. Expression of p53 in the squamous ep-
Fig 2. A specimen of a case without atypical changes in the anorectal epithelium, 6 weeks after KATO IIIinoculation. There was no atypical change in either the glandular or the squamous epithelium. (H&E; originalmagnification, × 50.)
ithelium was found after 2W. The total incidence of p53positivity in squamous epithelium was 19.4% and the in-cidence after 4W was 29.2%. However, no p53-positiveKATO III cells were observed in any specimens in this ex-periment. Interestingly, sporadic p53-positive cells werefound in the areas of invasive foci of the two adenocarci-nomas and the one squamous cell carcinoma mentionedabove.
Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004) 1915

KUWANO ET AL.
Fig 3. (A) Coexistence of invasive adenocarcinoma and squamous cell carcinoma of the mouse anorectal mucosa2 weeks after inoculation with KATO III. (B) High-power view of invasive adenocarcinoma and squamous cellcarcinomas intermingling among a poorly differentiated adenocarcinoma (KATO III.) (C) Immunostaining for p53protein. There are some p53-positive cells in nests of invasive well-differentiated adenocarcinoma and squamouscell carcinima. On the other hand, there are no p53-positive KATO III cells. (H&E; original magnifications, [A] ×50, [B] × 100, and [C] × 200.)
1916 Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004)

MALIGNANT TRANSFORMATION OF MOUSE ANORECTAL EPITHELIUM
Fig 3. continued
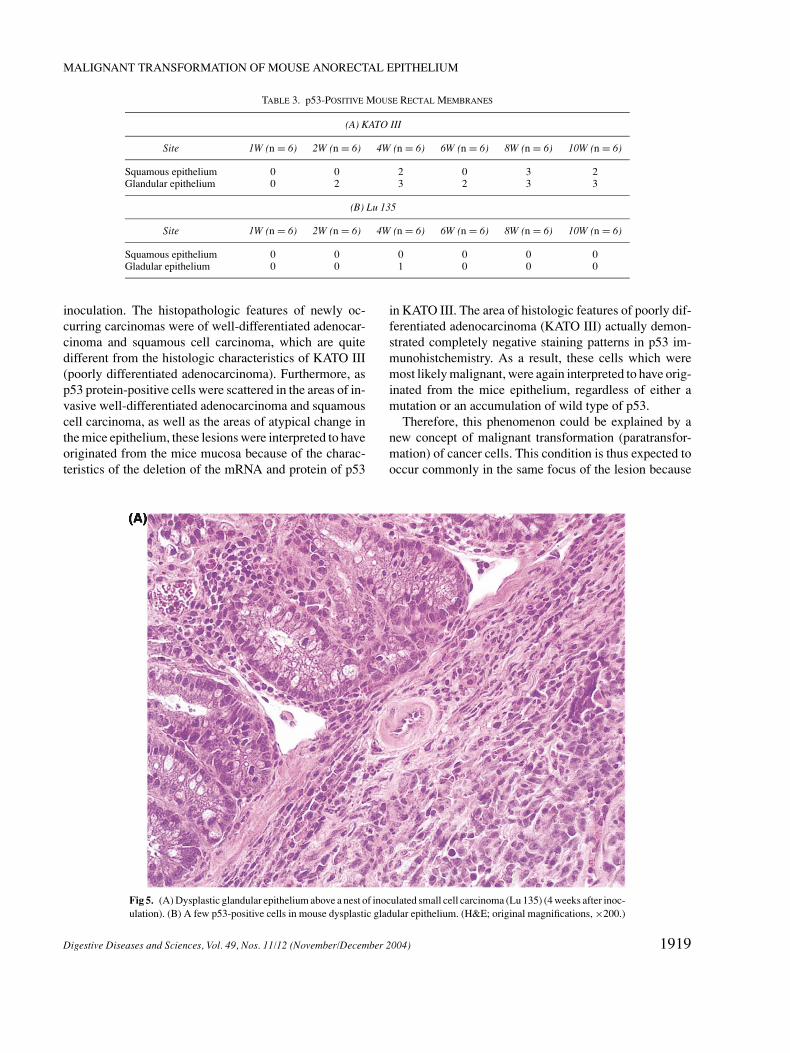
On the other hand in the cases implanted with Lu135,expression of p53 was found in the glandular epitheliumof only one case at 4W (Table 3 and Figure 5).
DISCUSSION
This study demonstrated the novel phenomenon thatcancer cells do have the ability to transform neighboringnormal cells into malignant cells. Although the factor re-sponsible for the inoculated malignant cells is still notclear at present, either certain effects or continuous stim-uli from the viable cancer cells to the adjacent host cellsare the likely cause of this phenomenon because inocula-tion of the homogenized malignant cell lines did not causethis phenomenon. As this cell-to-cell interaction occurredeven between the cells derived from different species, sucha phenomenon is thus expected to take place frequently be-tween the malignant and the adjacent cells in the same pa-tients or between some organs. From a clinicopathologicviewpoint some cases with malignant lesions composed ofmixed histologic types are frequently encountered (8), andtherefore it is possible that one malignant type of cancerstimulates the other types of cells, resulting in the coexis-tence of the different types of cancer cells. Moreover, theoccurrence of multiple primary cancers appears to be theresult of such cell-to-cell interaction. In such malignantlesions composed of mixed histologic types or those with
multiple foci of cancerous lesions, we are able to recognizethem as different phenotypes of the lesion. Even in lesionswith monotonous histologic phenotypes they would alsoresult from the cell-to-cell interaction of malignant trans-formation, such as the intraepithelial spread of esophagealsquamous cell carcinoma (20).
In this experiment, among the five malignant cell lines,KATO III and Lu 135 tumors, which were derived fromthe signet-ring cell type of human gastric adenocarcinomaand from the small cell type of lung carcinoma, showed thepotential for atypical or malignant transformation in theadjacent anorectal epithelium of tumor-inoculated mice.There have been reports of multifocal carcinogenesis ofsignet-ring cell carcinoma in the human stomach (21, 22)and of the coexistence of adenocarcinoma adjacent to thenest of small cell carcinoma of the stomach (23). Thisphenomenon observed in human histopathologic investi-gations is probably due to the prominent potential for para-transformation of signet-ring cell carcinoma and small cellcarcinoma.
The current study demonstrated atypical changes ofthe anorectal epithelium of female nude mice inducedby inoculation of KATO III and Lu 135. Moreover, def-inite invasive carcinomas originating from the glandularand squamous epithelium of the specimens were obtainedfrom at least three mice in the groups with KATO III
Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004) 1917
KUWANO ET AL.
Fig 4. (A) Invasive well-differentiated adenocarcinoma at the site of KATO III inoculation (4 weeks after inoculation).(B) High-power view shows the coexistence of well-differentiated adenocarcinoma and poorly differentiated eithermucinous or signet-ring cell carcinoma (KATO III). (H&E; original magnifications, [A] × 50 and [B] × 200.)
1918 Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004)
MALIGNANT TRANSFORMATION OF MOUSE ANORECTAL EPITHELIUM
TABLE 3. p53-POSITIVE MOUSE RECTAL MEMBRANES
(A) KATO III
Site 1W (n = 6) 2W (n = 6) 4W (n = 6) 6W (n = 6) 8W (n = 6) 10W (n = 6)
Squamous epithelium 0 0 2 0 3 2Glandular epithelium 0 2 3 2 3 3
(B) Lu 135
Site 1W (n = 6) 2W (n = 6) 4W (n = 6) 6W (n = 6) 8W (n = 6) 10W (n = 6)
Squamous epithelium 0 0 0 0 0 0Gladular epithelium 0 0 1 0 0 0
inoculation. The histopathologic features of newly oc-curring carcinomas were of well-differentiated adenocar-cinoma and squamous cell carcinoma, which are quitedifferent from the histologic characteristics of KATO III(poorly differentiated adenocarcinoma). Furthermore, asp53 protein-positive cells were scattered in the areas of in-vasive well-differentiated adenocarcinoma and squamouscell carcinoma, as well as the areas of atypical change inthe mice epithelium, these lesions were interpreted to haveoriginated from the mice mucosa because of the charac-teristics of the deletion of the mRNA and protein of p53
Fig 5. (A) Dysplastic glandular epithelium above a nest of inoculated small cell carcinoma (Lu 135) (4 weeks after inoc-ulation). (B) A few p53-positive cells in mouse dysplastic gladular epithelium. (H&E; original magnifications, ×200.)
in KATO III. The area of histologic features of poorly dif-ferentiated adenocarcinoma (KATO III) actually demon-strated completely negative staining patterns in p53 im-munohistchemistry. As a result, these cells which weremost likely malignant, were again interpreted to have orig-inated from the mice epithelium, regardless of either amutation or an accumulation of wild type of p53.
Therefore, this phenomenon could be explained by anew concept of malignant transformation (paratransfor-mation) of cancer cells. This condition is thus expected tooccur commonly in the same focus of the lesion because
Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004) 1919

KUWANO ET AL.
Fig 5. continued.
this phenomenon was identified even in the combinedspecimens of epithelium and cancer tissue obtained fromdifferent species.
It is thus expected that the mechanism of carcinogen-esis is continuously progressing even after the malignanttransformation of a single or a few cells and that the pro-cesses of carcinogenesis and proliferation of such malig-nant cells occur simultaneously in cancer tissue. Fromthese standpoints, research on cancer clonality, chronol-ogy, and strategies for cancer gene therapy or chemother-apy therefore needs to be thoroughly reevaluated. This pa-per demonstrates for the first time direct evidence of thephenomenon that malignant cancer cells have the poten-tial to transform themselves into other kinds of epithelialcells, and these novel findings would provide us with a newconcept for oncology research in the future. This valuableand novel discovery may change the basis of oncologyresearch while also providing new ideas for projects toinvestigate the mechanisms of carcinogenesis from sev-eral aspects such as molecular biology, cell biology, andpathology. Moreover, the novel experimental design itselfis also extremely useful as a simple model for investigatingthe mechanisms of oncogenesis.
ACKNOWLEDGMENTS
The authors thank Brian Quinn for critical comments. Thiswork was supported in part by Grants-in-Aid for Scientific Re-
search (A) 11307021 and 14207050 from the Japan Society forthe Promotion of Science.
REFERENCES
1. Alberts B, Bray D, Lewis J, Raff M, Roberts K, Watson JD: Molec-ular Biology of the Cell, 3rd ed. New York, London, Garland, 1994,pp 1275–1294
2. Groffen J, Stephenson JR, Heisterkamp N, de Klein A, Bartram CR,Grosveld G: Philadelphia chromosomal breakpoints are clusteredwithin a limited region, bcr, on chromosome 22. Cell 36(1):93–99,1984
3. Nowell PC, Hungerford DA: A minute chromosome in humanchronic granulocytic leukemia. Science 132:1497, 1960
4. Fearon ER, Hamilton SR, Vogelstein B: Clonal analysis of humancolorectal tumors. Science 238(4824):193–197, 1987
5. Fialkow PJ: Clonal origin of human tumors. Biochim Biophys Acta12;458(3):283–321, 1976
6. Tatematsu M, Masui T, Fukami H, et al.: Primary mono-clonal and secondary polyclonal growth of colon neoplastic le-sions in C3H/HeN ↔ BALB/c chimeric mice treated with 1,2-dimethylhydrazine immunohistochemical detection of C3H strain-specific antigen and simple sequence length polymorphism analysisof DNA. Int J Cancer 66(2):234–238, 1996
7. Sporn MB, Todaro GJ: Autocrine secretion and malignant transfor-mation of cells. N Engl J Med 303(15):878–880, 1980
8. Kuwano H, Ueo H, Sugimachi K, Inokuchi K, Toyoshima S,Enjoji M: Glandular or mucus-secreting components in squamouscell carcinoma of the esophagus Cancer 56:514–518, 1985
9. Kuwano H, Matsuda H, Matsuoka H, Kai H, Okudaira Y,Sugimachi K: Intra-epithelial carcinoma concomitant withesophageal squamous cell carcinoma Cancer 59:783–787, 1987
1920 Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004)

MALIGNANT TRANSFORMATION OF MOUSE ANORECTAL EPITHELIUM
10. Kuwano H, Ohno S, Matsuda H, Mori M, Sugimachi K: Serial his-tologic evaluation of multiple primary squamous cell carcinomas ofthe esophagus. Cancer 61:1635–1638, 1988
11. Kuwano H, Nagamatsu M, Ohno S, Matsuda H, Mori M, SugimachiK: Coexistence of intraepithelial carcinoma and glandular differen-tiation in esophageal squamous cell carcinoma. Cancer 62:1568–1572, 1988
12. Jue SF, Bradley RS, Rudnicki JA, Varmus HE, Brown AM: Themouse Wnt-1 gene can act via a paracrine mechanism in transfor-mation of mammary epithelial cells Mol Cell Biol 12(1):321–328,1992
13. Tsutsumi S, Kuwano H, Morinaga N, Shimura T, Asao T: Animalmodel of para-aortic lymph node metastasis. Cancer Lett 169(1):77–85, 2001
14. Motoyama T, Hojo H, Watanabe H: Comparison of seven cell linesderived from human gastric carcinomas. Acta Pathol Jpn 36(1):65–83, 1986
15. von Kleist S, Chany E, Burtin P, King M, Fogh J: Immuno-histology of the antigenic pattern of a continuous cell linefrom a human colon tumor. J Natl Cancer Inst 55(3):555–560,1975
16. Matozaki T, Sakamoto C, Matsuda K, et al.: Missense mutationsand a deletion of the p53 gene in human gastric cancer BiochemBiophys Res Commun 182(1):215–223, 1992
17. Fujita T, Kiyama M, Tomizawa Y, Kohno T, Yokota J: Compre-hensive analysis of p53 gene mutation characteristics in lung car-cinoma with special reference to histological subtypes. Int J Oncol15(5):927–934, 1999
18. Tsuruo T, Yamori T, Naganuma K, Tsukagoshi S, Sakurai Y: Char-acterization of metastatic clones derived from a metastatic variantof mouse colon adenocarcinoma 26. Cancer Res 43(11):5437–5442,1983
19. Miyazaki T, Kato H, Shitara Y, et al.: Mutation and expression ofthe metastasis suppressor gene KAII in esophageal squamous cellcarcinoma. Cancer 89(5):955–962, 2000
20. Kuwano H, Saeki H, Kawaguchi H, et al.: Proliferative activity ofcancer cells in front and center areas of carcinoma in situ and in-vasive sites of esophageal squamous-cell carcinoma. Int J Cancer78(2):149–152, 1998
21. Chun YS, Lindor NM, Smyrk TC, et al.: Germline E-cadheringene mutations: Is prophylactic total gastrectomy indicated? Cancer92(1):181–187, 2001
22. Huntsman DG, Carneiro F, Lewis FR, et al.: Early gastric cancer inyoung, asymptomatic carriers of germ-line E-cadherin mutations NEngl J Med 344(25):1904–1909, 2001
23. Matsui K, Kitagawa M, Miwa A, Kuroda Y, Tsuji M: Small cellcarcinoma of the stomach: A clinicopathologic study of 17 cases.Am J Gastroenterol 86(9):1167–1175, 1991
Digestive Diseases and Sciences, Vol. 49, Nos. 11/12 (November/December 2004) 1921





























