Embed Size (px)
Citation preview

Localization of Transitin mRNA, aNestin-Like Intermediate Filament
Family Member, in Chicken Radial GliaProcesses
JU-AHNG LEE AND GREGORY J. COLE*
Neurobiotechnology Center and Department of Neuroscience, The Ohio State University,Columbus, Ohio 43210
ABSTRACTWe have examined the gene expression of two radial glia intermediate filament proteins,
transitin and vimentin, in the developing chick CNS. Despite global similarities in theirmRNA distributions, marked regional differences are observed. Most notably, we show thattransitin mRNA is localized along radial glial processes and is localized to radial glia endfeet,whereas vimentin mRNA is not localized in radial glia. Localization of transitin mRNA is bestshown in the diencephalic radial glia, as well as cerebellar Bergmann glia. In addition, in theearly embryonic optic tectum, telencephalon, and retina, transitin mRNA is highly localizedto radial glia endfeet, which is suggestive of its transport in these cells. These in vivodemonstrations of transitin mRNA localization are confirmed by in situ hybridization anal-ysis of cultured chick brain radial glia, which demonstrates the presence of granular stainingfor transitin mRNA in glial processes. Transitin mRNA distribution in developing musclealso shows a highly regulated expression pattern, especially along the Z-lines of myofibrils. Asfurther support for the transport and localization of transitin mRNA in radial glia andmuscle, we have identified a consensus RNA transport signal in transitin mRNA that isabsent from vimentin. These data suggest that the local regulation of transitin proteinsynthesis may contribute to its function as an intermediate filament protein in radial glia. J.Comp. Neurol. 418:473–483, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: vimentin; glial endfeet; RNA transport signal; mRNA localization
Intermediate filaments (IFs) represent one of the threeclasses of cytoskeletal components found within eukary-otic cells and are the most complex cytoskeletal element interms of their gene diversity and cell type specificity ofexpression. The classes of IF proteins are defined based ontheir type of a-helical rod domain, with six IF proteinclasses currently proposed to exist (Fuchs and Weber,1994; Klymkowsky, 1995). In nervous tissue, IF proteinsare members of several distinct classes, with glial cellsexpressing vimentin and glial fibrillary acidic protein(GFAP), which are type III IF proteins (Fuchs and Weber,1994), as well as nestin, which is the defining member ofthe type VI class (Lendahl et al., 1990).
During CNS development transitions occur in IF pro-tein expression in neurons (Fliegner et al., 1994; Chien etal., 1996) and glia (Levitt and Rakic, 1980; Dahl, 1981;Pixely and de Vellis, 1984) and are likely associated withthe differentiation of these cell types. In glia, vimentin isthe predominant IF protein of immature astrocytes and
radial glia (Franke et al., 1978; Pixely and de Vellis, 1984;Voigt, 1989). As development proceeds vimentin is for themost part replaced in astrocytes by GFAP (Bignami andDahl, 1974; Levitt and Rakic, 1980; Pixely and de Vellis,1984). An additional IF protein of the nervous system,nestin, is also expressed in radial glia (Lendahl et al.,1990; Dahlstrand et al., 1995). It has also been shown thatreactive astrocytes induce nestin expression, with this IFprotein being coexpressed in these cells with vimentin andGFAP (Frisen et al., 1995). Thus, it appears that, duringthe life cycle of astrocytes, different combinations of IF
Grant sponsor: NIH; Grant number: NS33981.*Correspondence to: Dr. Gregory J. Cole, Neurobiotechnology Center,
The Ohio State University, 184 Rightmire Hall, 1060 Carmack Road,Columbus, OH 43210. E-mail: [email protected]
Received 10 August 1999; Revised 10 November 1999; Accepted 24November 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 418:473–483 (2000)
© 2000 WILEY-LISS, INC.

proteins can be expressed, and the developmental state ofastrocytes may be defined by the expression of these IFproteins.
Our laboratory has recently described the molecularcloning of chicken transitin, an IF protein with highesthomology to nestin and tanabin (Yuan et al., 1997). Basedon gene structure, transitin is a member of the nestin typeVI IF class (Napier et al., 1999). Accordingly, the a-helicalcores of transitin and nestin are 45% identical (Yuan etal., 1997), and transitin and nestin are both expressed byradial glia. Likewise, a recently characterized chickenskeletal muscle IF protein, paranemin, is a transitin iso-form that exhibits an expression pattern in muscle iden-tical to that of nestin (Hemkin et al., 1997). In developingCNS, transitin is a developmentally regulated IF proteinexpressed by radial glia, particularly midline radial glia(McCabe and Cole, 1992; McCabe et al., 1992). In radialglia, transitin and vimentin exhibit many similarities intheir expression pattern, as well as some differences (Coleand Lee, 1997). For example, floor plate cells expressvimentin protein but not transitin protein. These datatherefore suggest that, although transitin and vimentinare IF proteins of radial glia, subpopulations of glia arepresent in the developing CNS that have a unique require-ment for specific IF protein expression, such that transitinand vimentin are not always coexpressed by all radial glia.
In the present study, we have compared the distributionof mRNAs encoding chicken transitin and vimentin. Onerationale for undertaking these experiments was our ob-servation that transitin mRNA appeared to be localized inradial glial processes. Therefore, we were interested indetermining whether other glial IF mRNAs would also belocalized. Insofar as a hallmark of radial glia is their beinghighly polarized cells, it seemed reasonable to speculatethat the transport and localization of transitin mRNA intoradial glial processes could serve as one mechanism toregulate the morphology of radial glia. Consistently withthis hypothesis, it has become increasingly clear that onemechanism cells can utilize to establish polarity is torestrict the synthesis of specific proteins to selected cyto-plasmic compartments. This is often accomplished bytransporting and localizing the mRNAs to the specificcellular compartment. An example of this localization in-volves b-actin protein and mRNA localization to the la-mellipodia of muscle cells and fibroblasts (Lawrence andSinger, 1986; Hoock et al., 1991; Herman, 1993). In addi-tion, in the nervous system, it has been well documentedthat the transport of MAP2 and tau proteins into thedendritic and axonal compartments, respectively, canserve to localize these proteins to specific neuronal com-partments (Bernhardt and Matus, 1984; Kosik and Finch,1987). In the case of these neuronal proteins, MAP2mRNA is localized into dendrites (Kleiman et al., 1994),and tau mRNA is localized into the proximal segment ofthe axon (Litman et al., 1993). Other important examplesof mRNA localization in neurons include the localizationof specific mRNAs to the distal dendrites of the neuron(Bian et al., 1996; Paradies et al., 1997). Of particularinterest is that synaptic activity regulates dendritic local-ization of specific mRNAs to activated postsynaptic sites(Steward et al., 1998), suggesting that mRNA localizationmay have functional significance in synaptic plasticity. Incells of the glial lineage, GFAP has also been shown to belocalized to Muller cell radial glial processes (Sarthy et al.,1989), and myelin basic protein (MBP) mRNA is localized
to oligodendrocyte processes (Ainger et al., 1993). In thepresent study, we show that transitin mRNA is indeedlocalized in radial glial processes, whereas vimentinmRNA does not appear to be localized. Based on thislocalization of transitin mRNA in radial glia, we proposethat one function of transitin protein may be to provide anIF protein in radial glia that can undergo dynamicchanges to regulate morphological changes in these cellsduring CNS development.
MATERIALS AND METHODS
In situ hybridization of embryonic chickensections with
35S-labeled probes
The expression of transitin mRNA was analyzed duringchick brain development by in situ hybridization accordingto published protocols (Yuan et al., 1997). Chick brains wereremoved at various ages ranging from E3 to E13, fixed over-night in 4% paraformaldehyde in phosphate-buffered saline(PBS; pH 7.4), cryoprotected overnight in 20% sucrose, andfrozen at 285°C. Frozen cryostat sections (12 mm) werecollected onto Superfrost Plus slides and were heated at42°C for 30–45 minutes to adhere sections to the slides. Foranalysis of transitin mRNA expression, the pEAP3 cDNA(nucleotides 2,713–4,638 of transitin cDNA) was used tosynthesize [35S]CTP-labeled riboprobe using an RNA tran-scription kit (Promega, Madison, WI). For analysis of vimen-tin mRNA expression, a vimentin cDNA was isolated byRT-PCR of chick glia cDNA using the primer pair 59-ACGATGACTTGGAGTAAAGCTG (forward primer) and 59-GAGTTGTCAACCGTATAGTAATTCC (reverse primer)that spanned nucleotides 1,454–1,875 of the chicken vimen-tin cDNA. This cDNA was then used to synthesize vimentin[35S]CTP-labeled riboprobe. A second vimentin cDNA wasalso prepared by RT-PCR and spanned nucleotides 441–874in the coding regions. This vimentin cDNA, when used toprepare riboprobe, gave identical results to the vimentin1,454–1,875 probe and thus data are only shown for vimen-tin 1,454–1,875. For riboprobe analysis, radiolabeled ribo-probe in hybridization cocktail (Amresco, Solon, OH) wasapplied to slides, which were incubated at 55°C for 24 hours.The slides were washed 2 3 15 minutes with 23 SSC at45°C, treated for 30 minutes at 37°C with 20 mg/ml RNase A,and then washed in succession with 23 SSC (5 3 15 min-utes) at 37°C, 0.53 SSC (2 3 15 minutes) at 45°C, and 0.13SSC (3 3 15 minutes) at 65°C. Slides were dried by washingwith ethanol and exposed to Reflections X-ray film (Dupont,Boston, MA) for 3–7 days. For emulsion analysis, slides weredipped in NTB-2 liquid emulsion (Kodak, Rochester, NY)and incubated at 4°C for 1–2 weeks. Slides were then coun-terstained with cresyl violet prior to viewing.
In situ hybridization of embryonic chickensections with digoxigenin-labeled probes
Cryostat sections were prepared as described above.Digoxigenin-labeled riboprobes were synthesized usingDIG RNA labeling mix (Boehringer Mannheim, Indianap-olis, IN). Probes were hybridized in hybridization cocktail(Amresco) at 55°C for 24 hours. The slides were washed asdescribed above. After washing in 0.13 SSC, the slideswere incubated in TBST (Tris-buffered saline, pH 7.5,with Tween 20 and KCl) with 1% normal sheep serum for1 hour at room temperature. The slides were rinsed withTBST twice and then incubated with antidigoxigenin–AP
474 J.-A. LEE AND G.J. COLE

Fab fragments diluted 1:500 in chicken brain powder-equilibrated TBST for 1 hour. The slides were washedwith TBST (5 3 10 minutes) and Tris-buffered saline withTween 20 and MgCl2 (3 3 10 minutes) at room tempera-ture. Chromogenic reaction was performed with NBT(6.75 ml/ml) and BCIP (3.5 ml/ml) until the antisense/sensesignal ratio became maximized. The reaction was stoppedby washing with Tris-buffered saline with Tween 20 andMgCl2 (2 3 5 minutes) at room temperature. The slideswere serially dehydrated with ethanol and treated withxylene for 10 minutes before being coverslipped with per-mount.
In situ hybridization analysis of chick radialglial cultures
Primary glial cultures were prepared according to pub-lished protocols (Cole and Lee, 1997). Embryonic brains ofproper stages were dissected and triturated with trypsinunder sterile conditions. Centrifuged pellets were dissoci-ated with pipetting and aliquoted to 35 mm culture dishes.After about 1 week, cells were replated following dissoci-ation with trypsin and incubated in chamber slides (Lab-Tek, Naperville, IL) for about 2 days, until the glial cellsformed a highly radial morphology. Cells were then fixedin 220°C MeOH for 20 minutes. Digoxigenin labeled ribo-probes were hybridized to the slides at 55°C for 3 hoursand washed in succession with 23 SSC (5 minutes) atroom temperature, 0.53 SSC (15 minutes) at 45°C, and0.13 SSC (3 3 15 minutes) at 65°C. The slides wereincubated with normal donkey serum in PBS (1:500, roomtemperature, 30 minutes) and antidigoxigenin mousemonoclonal antibody in PBS 1 2% BSA (1:200, room tem-perature, 1 hour). The slides were washed in PBS (threetimes) and incubated with anti-mouse-Cy3 secondary an-tibody in PBS 1 2%BSA (room temperature, 1 hour in thedark). After washing in PBS, the slides were coverslippedwith 20% glycerol in PBS and observed with a scanninglaser confocal microscope (Bio-Rad). Figures were pro-duced using Adobe Photoshop 4.0.
RESULTS
Transitin mRNA distribution indiencephalon and cerebellum is associated
with radial glial processes
When the cellular distribution of transitin mRNA wasexamined by in situ hybridization in E11 diencephalonand E13 cerebellum, it was clear that transitin mRNAexpression was restricted in these developing brain re-gions. As is shown in Figure 1A, transitin mRNA expres-sion was localized in proximity to the third ventricle indiencephalon, with cellular processes containing transitinmRNA extending away from the third ventricle (arrow-heads). When E13 cerebellum was analyzed for transitinmRNA expression, a similar phenomenon was observed inthat transitin mRNA was localized in a fibrous patternthat appeared to be associated with radial glial cell pro-cesses (Fig. 1B). Interestingly, the transitin mRNA distri-bution in cerebellum appeared to exist as punctate gran-ules in a radial pattern (Fig. 1B, arrowheads), which hasbeen shown previously for other transported mRNAs toresult from RNA granules undergoing transport (Aingeret al., 1993). These data were therefore highly suggestiveof transitin mRNA being transported in radial glial cellprocesses.
Transitin mRNA is transported andlocalized into glial processes in vitro
To extend our analyses of whether transitin might be atransported mRNA in radial glia, we examined whethertransitin mRNA was present in glial cell processes ofcultured cells. At low density, chick brain astrocytes arefor the most part bipolar glial cells, based on their mor-phology. When these cells were analyzed by in situ hybrid-ization, we always observed the localization of transitinmRNA in glial cell processes (Fig. 2). An interesting ob-servation we made in these experiments was that in manycells transitin mRNA was enriched in the endfeet/growthcones of the bipolar glial cell (Fig. 2A,B) and that when anastrocyte was spreading via a lamellipodium we could
Fig. 1. In situ localization of transitin mRNA in embryonic chickdiencephalon and cerebellum using digoxigenin-labeled riboprobe.Transitin mRNA is localized to radial glial processes in E11 chickdiencephalon (A) and E13 chick cerebellum (B). Arrowheads denote
radial glial process labeling with digoxigenin-labeled riboprobe, and itcan be seen in cerebellum that transitin mRNA is distributed in apunctate pattern that may suggest transport of transitin mRNA ingranules in these cellular processes.
475TRANSITIN mRNA IS TRANSPORTED IN DEVELOPING RADIAL GLIA

observe transitin mRNA along the leading edge of the cell(Fig. 2B). Thus, these data are consistent with transitinmRNA being transported to specific compartments of ra-dial glial cells. In addition, at higher magnification gran-ules that were positive for transitin mRNA could be ob-served in bipolar glial processes (Fig. 2D), againsuggesting that transitin mRNA localization in glial pro-cesses arises via transport of transitin mRNA.
Is transitin mRNA localization in radial gliaunique to this IF protein?
Vimentin and transitin are both IF proteins, of differentIF classes, that are expressed by radial glia during CNS
development. Thus, it is of interest to determine whethertransitin mRNA transport and localization in radial gliaare unique to this IF protein. We therefore examined themRNA expression patterns for vimentin and transitinmRNAs during chick CNS development. As is shown inFigure 3, when transitin and vimentin mRNA expressionwas examined in E11 diencepahlon, it was apparent thatboth mRNAs were localized to cells in proximity to thethird ventricle, where radial glia cell bodies reside. How-ever, it was also apparent that the transitin mRNA ex-pression pattern was more diffuse and extended awayfrom the third ventricle, whereas vimentin mRNA wasexpressed only in proximity to the third ventricle (Fig.3A,B). Furthermore, radial glial cell processes could bedetected that contained transitin mRNA (Fig. 3A, arrow-heads), whereas vimentin mRNA was not localized to ra-dial glia cell processes.
The differences in expression of vimentin and transitinmRNAs were even more apparent when Muller cells wereanalyzed in early chick retina (E4). Transitin mRNA isclearly localized to Muller cell endfeet (Fig. 3C), which wehave shown previously to be enriched in transitin protein(McCabe and Cole, 1992). Conversely, vimentin mRNAwas not detected in Muller cell endfeet (Fig. 3D). When weextended these analyses to E4 telencephalon, we againobserved transitin mRNA in radial glia cell endfeet at thepial surface (Fig. 3E), whereas vimentin mRNA was notlocalized to telencephalic radial glia endfeet (Fig. 3F).
Comparison of transitin and vimentin mRNAexpression in embryonic chick spinal cord
Transitin and vimentin are also expressed in developingchicken spinal cord, so we examined the expression ofthese intermediate filament protein mRNAs in this CNSregion. We focused on early spinal cord development, be-cause both proteins are almost completely down-regulatedin the later stages of embryonic spinal cord development.As is shown in Figure 4A,B, during early spinal corddevelopment (E3) vimentin and transitin exhibited somesimilarity in mRNA expression pattern, with both mRNAsexpressed in the ventricular zone, which is suggestive ofexpression by glial progenitors. However, vimentin mRNAwas clearly expressed in ventral spinal cord, whereastransitin mRNA was absent from this region (Fig. 4A,B).
At E4 and E5 of spinal cord development, vimentinmRNA expression is particularly robust in the ventricularzone, but transitin mRNA expression exhibits an apparentdown-regulation in this region of the spinal cord (Fig.4C–H). In addition, vimentin mRNA expression is readilyapparent in the floor plate, with transitin not being ex-pressed in floor plate. These data are in agreement withour recent studies on vimentin and transitin protein ex-pression, which showed that only vimentin protein waspresent in spinal cord floor plate (Cole and Lee, 1997).Interestingly, vimentin mRNA expression also appears inthe ventral horn of E4 and E5 spinal cord (Fig. 4D,F,J),which could be suggestive of expression by early motorneurons. We also considered the possibility that the vi-mentin probe we used contained homology to a motorneuron mRNA, so we used a second vimentin riboprobe,from nucleotides 441–874 in the coding region, to conductin situ hybridization. These results were identical in thathybridization was observed in the ventral horn, indicatingthat vimentin mRNA is expressed in these cells (data notshown). In situ hybridization analysis of the E5 spinal
Fig. 2. Demonstration of transitin mRNA transport in primarychick radial glial cell cultures. E9 chick brains were dissociated andplated on tissue culture plastic. Cells were then split and replated onplastic slide chambers and permeabilized with methanol. Cells werehybridized with sense or antisense digoxigenin-labeled EAP3 ribo-probe, washed, and then incubated with mouse antidigoxigenin mono-clonal antibody and anti-mouse Cy3-conjugated secondary antibody.Cells were then viewed by scanning laser confocal microscopy. Cellshybridized with sense riboprobe exhibited no fluorescence (data notshown). A single astrocyte is shown in each panel and clearly showsthat transitin mRNA is transported by astrocytes in vitro. A,B: Thearrow indicates the presence of abundant transitin mRNA in theendfoot region of the glial cell process. Arrowheads in B illustrate thepresence of transitin mRNA in lamellipodia of a flattened region of theglial cell body. Interestingly, we have previously shown that transitinprotein is distributed in endfeet and lamellipodia of astrocytes withthis morphology (McCabe et al., 1992). C: A punctate staining patternis apparent in a radial glial cell process. The boxed region is shown athigher magnification in D, and the arrows indicate granules in radialglia processes that express transitin mRNA, which is indicative oftransitin RNA transport in granules in glial processes. A, 3200; B,C,3400; D, 31,200.
476 J.-A. LEE AND G.J. COLE

cord using 35S-labeled riboprobe reveals another interest-ing distinction between transitin and vimentin mRNAdistributions: They are almost completely complementaryto each other at this stage (Fig. 4K,L). Where transitinmRNA is abundant, vimentin mRNA is absent, and viceversa.
Transitin and vimentin mRNAs havedistinctive expression patterns in
embryonic chick brain
When embryonic chick brain is analyzed globally fortransitin and vimentin mRNA expression, it becomes ap-parent that key differences in the expression of theseradial glial IF mRNAs can be detected (Fig. 5). For exam-ple, in the early neural tissue of E4 brain, transitin mRNAis shown to be transported along the radial glial processesand localized at the endfeet of the cortical plate (Fig. 5A,arrowheads). In addition, transitin mRNA is highly local-ized at the endfeet of Muller cells in E4 retinas (Fig. 5A,arrows). These features were reminiscent of the resultsshown in Figure 3. By contrast, such transport and local-ization are not detected with vimentin mRNA (Fig. 5B).However, vimentin mRNA is clearly detected in the non-neural mesenchyme (Fig. 5B, arrows).
In E7 brain, further marked differences were detected(Fig. 5C–J). First, at the floor plate of developing hind-
brain, vimentin mRNA is highly expressed, whereas tran-sitin mRNA is absent (Fig. 5C,D,G,H, curved arrows).Second, the mRNA expression pattern of transitin is morediffuse and extended away from the fourth ventricle,again suggesting that transitin mRNA is transported andlocalized along the radial glial processes. This is reminis-cent of the third ventricle expression pattern shown inFigure 3. Conversely, vimentin mRNA is expressed inproximity to the fourth ventricle [compare in Fig. 5C andD area at line of arrowheads in C (fourth ventricle)].Third, localization of transitin mRNA along the radialglial processes can be shown on the cellular level in thediencephalic ventricular layer (Fig. 5E). In the adjacentsection, vimentin mRNA is distributed along the ventric-ular layer only (Fig. 5F). Fourth, 35S-labeled riboprobe insitu hybridization analysis of the E7 brain reveals addi-tional differences between the two IF mRNAs. Thus, non-neural mesenchymal tissue in the head shows a high levelof vimentin mRNA, whereas transitin mRNA is absent(Fig. 5H, solid arrows). Furthermore, connective tissuethat encloses the marginal layer of the telencephalic neu-ral tissue contains a high level of vimentin mRNA, but nottransitin mRNA (Fig. 5H, arrowheads). In addition, me-dial rectal eye muscle shows a marked discrepancy be-tween the two IF mRNAs, i.e., high transitin mRNA andvery low vimentin mRNA expression (Fig. 5G,H, open
Fig. 3. In situ hybridization analysis of transitin and vimentinmRNA in embryonic chick diencephalon, retina, and telencephalon.Digoxigenin-labeled transitin or vimentin riboprobes were hybridizedwith E12 chick brain sagittal sections (A,B), E4 retina (C,D), or E4telencephalon (E,F). A: Transitin mRNA expression is observed incells located in proximity to the third ventricle (3V), and radial glialprocesses containing transitin mRNA can be detected emanating fromthe third ventricle (arrowheads). Note that the width of stainingadjacent to the third ventricle is wider than that observed for vimen-
tin in B. B: Vimentin mRNA expression is detected in cells near thethird ventricle, but radial glial processes containing vimentin mRNAare not detected. C: Transitin mRNA expression is observed in Mullercell endfeet in E4 retina (arrow), whereas vimentin expression isabsent from endfeet (D). E: In E4 telencephalon transitin mRNA isagain detected in endfeet along the pial surface, whereas vimentin (F)mRNA expression is absent from pial endfeet. GCL, ganglion celllayer; ONL, outer nuclear layer; V, ventricle; P, pial surface.
477TRANSITIN mRNA IS TRANSPORTED IN DEVELOPING RADIAL GLIA

arrows). Marked differences are also present in neuraltissue. For example, two layers of high-level expressionare easily detected with transitin mRNA outside the earlylateral ventricles (Fig. 5C,G,I, arrowheads and solid ar-rows). Such mRNA expression pattern is not found in thevimentin mRNA analysis (Fig. 5H,J). Compared to bright-field images (Fig. 5I,J, right panels), the inner layer of
transitin mRNA expression (Fig. 5I, arrowheads) is foundto correspond to the subplate of the developing cortex, andthe outer layer of transitin mRNA expression (Fig. 5I,arrows) corresponds to the pial surface. Of particular in-terest is that these are the two regions of the early cortexwhere different populations of radial glial endfeet arepositioned (Marı́n-Padilla, 1995); these data suggest thattransitin mRNA is also being localized to radial glial end-feet in the chicken telencephalon.
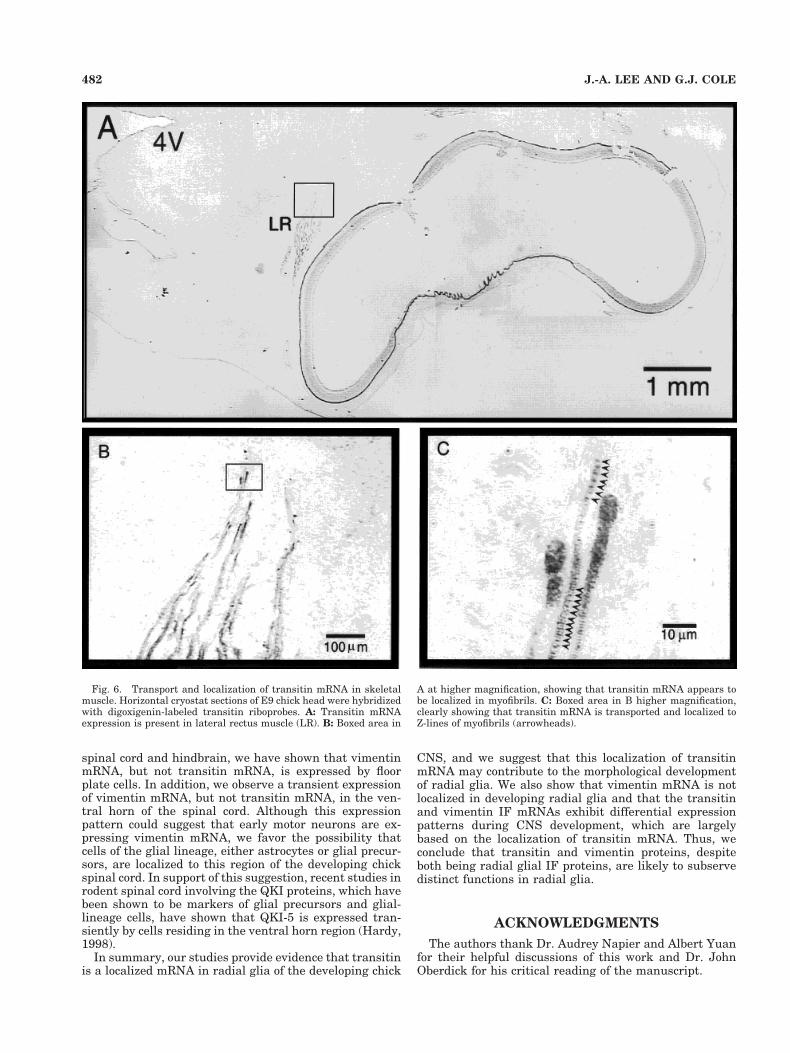
Transitin mRNA is localized inskeletal muscle
Because previous studies have shown that transitin pro-tein is abundantly expressed in embryonic skeletal muscle(McCabe et al., 1992), transitin mRNA expression wasexamined in skeletal muscle. Previous in vitro studiesinvolving vimentin mRNA have indicated that myoblastsdo not transport and localize vimentin mRNA (Lawrenceand Singer, 1986; Cripe et al., 1993) and that developingmyotubes do not localize vimentin mRNA (Cripe et al.,1993). However, mature myotubes have been shown tolocalize vimentin mRNA and protein to Z-lines and cos-tameres (Cripe et al., 1993; Morris and Fulton, 1994).Thus, these studies suggest that, although vimentinmRNA is not transported and localized in radial glia, itsmRNA may be able to be localized in specific cell types.Based on these data concerning vimentin, we postulatedthat transitin would be localized to Z-lines in developingskeletal muscle. As is shown in Figure 6A, when analyzingthe expression of transitin mRNA in developing chickCNS, we could detect transitin mRNA in the lateral rectusmuscle, located in proximity to the developing eye. Whenviewed at higher magnification, individual myofibrils canbe shown to express transitin mRNA, and the mRNA isclearly localized to Z-lines (Fig. 6B,C, arrowheads). Wecould not detect such localized expression along theZ-lines with vimentin mRNA at these stages of embryonicskeletal muscle development (data not shown; see Discus-sion).
DISCUSSION
Our present studies demonstrate that chicken transitinmRNA is localized in radial glial cells into glial processes,with the mRNA often being enriched in the glial endfeet.This observation is important, insofar as it adds transitinto the growing list of mRNAs that are transported andlocalized in somatic cells. In particular, for a morphologi-cally polarized cell such as a radial glial cell, it is tempting
Fig. 4. Comparison of mRNA expression patterns of transitin andvimentin in early embryonic spinal cord. Cryostat sections of E3(A,B), E4 (C,D,G,H), and E5 (E,F,I–L) spinal cord were hybridizedwith antisense digoxigenin-labeled riboprobe to transitin (left column)or vimentin (right column). Localization of transitin mRNA to glialendfeet is prominent only at E3 (A). Note that transitin is not ex-pressed in floor plate (C,E,G), whereas vimentin is expressed in floorplate (D,F,H). E and F are higher magnification images of the floorplate region (boxed areas in C and D). Vimentin mRNA is also abun-dantly expressed by cells in the ventral horn of the E4–E5 spinal cord(D,H,J), whereas transitin is never detected in this spinal cord region.I and J are high magnification images of the ventral horn area (boxedareas in G and H). Note also the striking complementary patterns oftransitin and vimentin mRNA distribution in K and L.
478 J.-A. LEE AND G.J. COLE

to speculate that transitin mRNA transport and localiza-tion to cellular processes and endfeet may serve to regu-late the morphological differentiation of these cells duringnervous system development. In this context, the trans-port of transitin mRNA can permit local regulation oftransitin protein synthesis in radial glial cells, allowing aspecific type of IF protein to be localized to specific cellcompartments. Consistently with this hypothesis, usingmonoclonal antibodies to transitin an isoform of the pro-tein has been shown to localize to the cytoplasmic face ofthe membrane (Kelly et al., 1995), and a similar localiza-tion of this transitin isoform occurs in cardiac muscle (Maet al., 1999). This raises the interesting possibility thatRNA localization of transitin can contribute to this distri-bution of the transitin protein.
In support of our postulate that transitin mRNA local-ization may be crucial to the morphological differentiationof radial glia, it has become increasingly clear that onepossible mechanism cells can utilize to establish polarityis to restrict the synthesis of specific proteins to selectedcytoplasmic compartments by transporting and localizingtheir mRNAs. A classic example of this localization in-volves b-actin protein localization to the lamellipodia ofmuscle cells and fibroblasts (Hoock et al., 1991; Herman,1993). Tau and MAP2 protein have also been shown toplay an integral role in the establishment of polarity inneurons (Caceres and Kosik, 1990), and the localization oftheir mRNAs to neuronal processes (Litman et al., 1993;Kleiman et al., 1994) is consistent with the hypothesisthat mRNA localization is one mechanism cells utilize tomaintain polarity. Our demonstration that transitinmRNA is transported to the leading edge of bipolar glialcells in vitro, and throughout radial glial processes andinto endfeet in vivo, suggests that transitin is anotherexample of a cytoskeletal component that may have itsmRNA transported and localized in cells in order to reg-ulate the acquisition or maintenance of the polarized cellstate.
Another intriguing hypothesis can be raised with regardto the functional significance of transitin mRNA trans-port, based on transitin mRNA’s localized expression pat-tern in radial glial endfeet: Transitin may play a role asone of the components that link the cellular cytoskeletonto the extracellular matrix, a notable example being thehemidesmosome. Supporting evidence for this hypothesis,although indirect, is that glial endfeet make a direct con-tact with the basement membrane of the pial surfaceusing hemidesmosomes (Nakano et al., 1996; Green andJones, 1996) and cytokeratin, the de facto IF protein as-sociated with hemidesmosomes, is absent in chicken ra-dial glia (Tapscott et al., 1981; Page, 1989). Furthermore,IF proteins other than cytokeratin can be involved inhemidesmosomal attachment (Kartenbeck et al., 1984;Schroder et al., 1997). Because transitin protein andmRNA are highly enriched at radial glial endfeet duringperiods when the integrity of pial–endfeet adhesion maybe most critical (McCabe and Cole, 1992; present study)and an isoform of transitin has been shown to be highlylocalized to the cytoplasmic face of the plasma membraneof the cultured astrocytes (Kelly et al., 1995), it is tempt-ing to speculate that transitin’s localization to radial glialendfeet either serves to regulate hemidesmosome functionor occurs in response to hemidesmosomes with thesestructures, possibly regulating transitin function.
A possible explanation regarding the two layers of tran-sitin mRNA localization in E7 chick telencephalon (seeFig. 5I) is related to our hypothesis that transitin proteinmight be involved in radial glia hemidesmosome function.Thus, the outer band of transitin mRNA localization cor-responds to the pial surface, whereas the inner band cor-responds to the subplate. Two different types of radial gliahave been proposed to exist during early telencephalicdevelopment (Marı́n-Padilla, 1995). Type I radial glia, re-taining their ependymal and pial contacts, are utilized inneuronal migration, and later lose their pial contacts anddifferentiate into astrocytes. Type II radial glia prema-turely lose their pial attachment, and the large, roundterminals of type II radial glia become aligned along thesubplate. Thus, the two bands of transitin mRNA local-ization appear to correspond to the endfeet of type I andtype II radial glia. Whereas the type II radial glia losetheir pial contacts, the number of remaining type I radialglial endfeet is progressively increased by undergoing se-quential branching of their pial segments. In addition, astype II radial glia mature, they establish new endfeet-mediated contacts with developing capillaries within thesubplate (Marı́n-Padilla, 1995). Under such active mor-phological changes, the cytoskeletal components of theradial glial endfeet would be expected to undergo signifi-cant changes, and transitin protein may contribute to thisprocess. In this regard, timely changes in cytoskeletalstructures at a long distances from the glial cell bodywould be best achieved if a reservoir of mRNA was local-ized at the same site, allowing proteins to be synthesizedlocally rather than requiring synthesis and transport ofthe protein to the endfeet.
Based on our in situ hybridization analyses, whichclearly show that transitin mRNA is localized to radialglial processes, it is of interest to ask whether cis-actingsequences will be contained with transitin that regulateits localization. Several classes of cis-acting elements havebeen described that may regulate RNA transport, includ-ing the zip code element found in the 39-UTR of severalRNAs (Singer, 1993) and a consensus RNA transport sig-nal (RTS) that was originally identified in the 39-UTR ofMBP mRNA (Ainger et al., 1997). Interestingly, the RTScomprises two overlapping 10mer sequences, with at leastone of the 10mer sequences having been shown to reside inthe 39-UTR or coding region of many transported mRNAs(Ainger et al., 1997). This observation therefore suggeststhat this cis-acting element may be an important regula-tor of RNA transport (Ainger et al., 1997). Interestingly,one of the RTS 10mer sequences (GCCAGAGAGC) is con-tained within the open-reading frame of transitin, provid-ing support for our initial observation that transitin is atransported mRNA in radial glia. Interestingly, vimentinlacks an RTS in its mRNA sequence, which may contrib-ute to its apparent lack of transport in radial glia. It willtherefore be important in future studies to examine theprecise role of transitin’s RTS in its localization.
The studies reported here have also focused on an anal-ysis and comparison of the distribution of transitin andvimentin mRNAs in the developing CNS. Part of the ra-tionale for conducting these studies was that, althoughboth of these IF proteins are expressed by radial glia, onlytransitin contains a consensus RTS. Our data showmarked differences in the expression pattern of transitinand vimentin mRNAs. Particularly intriguing is the ap-parent lack of vimentin mRNA transport in radial glia
479TRANSITIN mRNA IS TRANSPORTED IN DEVELOPING RADIAL GLIA

expressing and transporting transitin mRNA. For exam-ple, we demonstrate that radial glia express both IF mol-ecules in retina, diencephalon, and telencephalon, withonly transitin exhibiting RNA transport. These datatherefore provide additional evidence that transitin hasunique functions as an IF protein compared to other radialglial IF proteins.
We have also shown in the present study that tran-sitin mRNA is selectively localized to Z-lines in skeletal
muscle. These data are of particular interest, in thatprevious in vitro studies on cultured muscle cells haveshown that vimentin mRNA becomes localized to Z-linesas muscle cells differentiate (Cripe et al., 1993; Morrisand Fulton, 1994). Our failure to detect vimentin mRNAlocalization to Z-lines in vivo may suggest that vimentinmRNA localization in muscle is delayed relative to tran-sitin mRNA localization. These data also raise the in-teresting possibility that different cell types (i.e., radial
Figure 5
480 J.-A. LEE AND G.J. COLE

glia and muscle) may employ distinct mechanisms totransport and/or localize mRNAs, because vimentinmRNA localized in muscle cells in vitro but is not local-ized in radial glia.
Our comparison of transitin and vimentin mRNA trans-port has also provided important new information relatingto differences in the expression pattern of these two IFproteins during CNS development. For early developing
Fig. 5. Comparison of mRNA expression patterns of transitin (leftcolumn) and vimentin (right column) in early embryonic chick brain.A: Transitin mRNA expression in E4 head. Arrowheads denote thetransitin mRNA expression pattern that spans the telencephalic neu-ral tube and is localized in the pial layer where radial glial endfeet arepresent. Such localized transitin mRNA pattern is also denoted alongthe vitreal layer of the developing retinas where Muller cell endfeetare present (arrows). B: Adjacent sections of vimentin in situ hybrid-ization show no such localized mRNA expression patterns. However,in the developing mesenchyme, there is a high level of vimentinmRNA. C–J: In situ hybridization of transitin and vimentin in E7chick brain. Note the broader mRNA distribution pattern of transitinalong the fourth ventricle (4V) and the narrow pattern of vimentinmRNA (doubly arrowed lines at the fourth ventricle in C and D).Arrowheads in C denote the telencephalic region where radial gliaendfeet are present, which contain transitin mRNA. Transitin mRNAis also expressed in radial glia of diencephalon (boxed area), which isshown at higher magnification in E. These radial glia also transporttransitin mRNA. The hindbrain region of E7 brain also expressestransitin mRNA, but transitin is noticeably absent from floor plate(curved arrow). It can also be seen that at E7 the retinal Muller cellscontinue to localize transitin mRNA to endfeet. D: Vimentin mRNAexpression in E7 brain. Note that vimentin mRNA is for the most part
restricted to the ventricular zone of telencephalon, diencephalon, andhindbrain. However, clear differences in vimentin expression are ev-ident compared to transitin. Vimentin is not transported into radialglial processes, or localized to endfeet, in any brain region. VimentinmRNA is also expressed at high levels in the hindbrain floor plate,where transitin is absent at this stage of development. F: Highermagnification view of vimentin mRNA expression in diencephalon,showing that vimentin is restricted to the ventricular zone and is nottransported into radial glial cell processes (compare to E for tran-sitin). G–J: 35S-labeled in situ hybridization images of E7 brainslightly dorsal to C and D. In comparison to vimentin (H), two bandsof high level mRNA expression are clearly shown with transitin probe(solid, straight arrows and arrowheads in G). Note also the broadermRNA distribution around the fourth ventricle and diencephalic neu-ral tissue. The curved arrow denotes the lack of transitin mRNA atthe floor plate. Open arrows (G and H) compare the mRNA level oftransitin and vimentin in the medial rectus muscle. The two telence-phalic signal bands of transitin mRNA are better shown in the mag-nified images in I. The outer band (arrows) corresponds to the pialsurface and the inner band (arrowheads) to the subplate. The dashedline in the J indicates the pial surface of the neural tissue to distin-guish it from the outer connective tissue that contains high level ofvimentin mRNA.
481TRANSITIN mRNA IS TRANSPORTED IN DEVELOPING RADIAL GLIA
spinal cord and hindbrain, we have shown that vimentinmRNA, but not transitin mRNA, is expressed by floorplate cells. In addition, we observe a transient expressionof vimentin mRNA, but not transitin mRNA, in the ven-tral horn of the spinal cord. Although this expressionpattern could suggest that early motor neurons are ex-pressing vimentin mRNA, we favor the possibility thatcells of the glial lineage, either astrocytes or glial precur-sors, are localized to this region of the developing chickspinal cord. In support of this suggestion, recent studies inrodent spinal cord involving the QKI proteins, which havebeen shown to be markers of glial precursors and glial-lineage cells, have shown that QKI-5 is expressed tran-siently by cells residing in the ventral horn region (Hardy,1998).
In summary, our studies provide evidence that transitinis a localized mRNA in radial glia of the developing chick
CNS, and we suggest that this localization of transitinmRNA may contribute to the morphological developmentof radial glia. We also show that vimentin mRNA is notlocalized in developing radial glia and that the transitinand vimentin IF mRNAs exhibit differential expressionpatterns during CNS development, which are largelybased on the localization of transitin mRNA. Thus, weconclude that transitin and vimentin proteins, despiteboth being radial glial IF proteins, are likely to subservedistinct functions in radial glia.
ACKNOWLEDGMENTS
The authors thank Dr. Audrey Napier and Albert Yuanfor their helpful discussions of this work and Dr. JohnOberdick for his critical reading of the manuscript.
Fig. 6. Transport and localization of transitin mRNA in skeletalmuscle. Horizontal cryostat sections of E9 chick head were hybridizedwith digoxigenin-labeled transitin riboprobes. A: Transitin mRNAexpression is present in lateral rectus muscle (LR). B: Boxed area in
A at higher magnification, showing that transitin mRNA appears tobe localized in myofibrils. C: Boxed area in B higher magnification,clearly showing that transitin mRNA is transported and localized toZ-lines of myofibrils (arrowheads).
482 J.-A. LEE AND G.J. COLE

LITERATURE CITED
Ainger K, Avossa D, Morgan F, Hill SJ, Barry C, Barbarese E, Carson JH.1993. Transport and localization of exogenous myelin basic proteinmRNA microinjected into oligodendrocytes. J Cell Biol 123:431–441.
Ainger K, Avossa D, Diana AS, Barry C, Barbarese E, Carson JH. 1997.Transport and localization elements in myelin basic protein mRNA.J Cell Biol 138:1077–1087.
Bernhardt R, Matus A. 1984. Light and electron microscopic studies of thedistribution of microtubule-associated protein 2 in rat brain: a differ-ence between dendritic and axonal cytoskeletons. J Comp Neurol 226:203–221.
Bian F, Chu T, Schilling K, Oberdick J. 1996. Differential mRNA transportand the regulation of protein synthesis: selective sensitivity of Purkinjecell dendritic mRNAs to translational inhibition. Mol Cell Neurosci7:116–133.
Bignami A, Dahl D. 1974. Astrocyte specific protein and neuroglial differ-entiation: an immunofluorescence study with antibodies to the glialfibrillary acidic protein. J Comp Neurol 153:27–38.
Caceres A, Kosik KS. 1990. Inhibition of neurite polarity by tau antisenseoligonucleotides in primary cerebellar neurons. Nature 343:461–463.
Chien C-L, Mason CA, Liem RKH. 1996. a-Internexin is the only neuronalintermediate filament expressed in developing cerebellar granule neu-rons. J Neurobiol 29:304–318.
Cole GJ, Lee J-A. 1997. Immunocytochemical localization of a novel radialglial intermediate filament protein. Brain Res Dev Brain Res101:225–238.
Cripe L, Morris E, Fulton AB. 1993. Vimentin mRNA location changesduring muscle development. Proc Natl Acad Sci USA 90:2724–2728.
Dahl D. 1981. The vimentin–GFAP protein transition in rat cytoskeletonoccurs at the time of myelination. J Neurosci Res 6:741–748.
Dahlstrand J, Lardelli M, Lendahl U. 1995. Nestin mRNA expressioncorrelates with the central nervous system progenitor cell state inmany, but not all, regions of developing central nervous system. BrainRes Dev Brain Res 84:109–129.
Fliegner KH, Kaplan MP, Wood TA, Pintar JE, Liem RKH. 1994. Expres-sion of the gene for the neuronal intermediate filament proteina-internexin coincides with the onset of neuronal differentiation in thedeveloping rat nervous system. J Comp Neurol 342:161–173.
Franke WW, Schmid E, Osborn M, Weber K. 1978. Different intermediatesized filaments distinguished by immunofluorescence microscopy. ProcNatl Acad Sci USA 75:5034–5038.
Frisen J, Johansson CB, Totok C, Risling M, Lendahl U. 1995. Rapid,widespread, and longlasting induction of nestin contributes to thegeneration of glial scar tissue after CNS injury. J Cell Biol 131:453–464.
Fuchs E, Weber K. 1994. Intermediate filaments. Structure, dynamics,function and disease. Annu Rev Biochem 63: 345–382.
Green KJ, Jones JC. 1996. Desmosomes and hemidesmosomes: structureand function of molecular components. FASEB J 18:871–881.
Hardy RJ. 1998. QKI expression is regulated during neuron–glial cell fatedecisions. J Neurosci Res 54:46–57.
Hemken PM, Bellin RM, Sernett SW, Becker B, Huiatt TW, Robson RM.1997. Molecular characteristics of the novel intermediate filament pro-tein paranemin. Sequence reveals EAP-300 and IFAPa-400 are highlyhomologous to paranemin. J Biol Chem 272:32489–32499.
Herman IM. 1993. Actin isoforms. Curr Opin Cell Biol 5:48–55.Hoock TC, Newcomb PM, Herman IM. 1991. b-Actin and its mRNA are
localized at the plasma membrane and the regions of moving cytoplasmduring cellular response to injury. J Cell Biol 112:653–664.
Kartenbeck J, Schwechheimer K, Moll R, Franke WW. 1984. Attachment ofvimentin filaments to desmosomal plaques in human meningiomalcells and arachnoidal tissue. J Cell Biol 98:1072–1081.
Kelly MM, Phanhthourath C, Brees DK, McCabe CF, Cole GJ. 1995.Molecular characterization of EAP-300: a high molecular weight, em-bryonic polypeptide containing an amino acid repeat comprised ofmultiple leucine-zipper motifs. Brain Res Dev Brain Res 85: 31–47.
Kleiman R, Banker G, Steward O. 1994. Development of subcellular mRNAcompartmentation in hippocampal neurons in culture. J Neurosci 14:1130–1140.
Klymkowsky MW. 1995. Intermediate filaments: new proteins, some an-swers, more questions. Curr Opin Cell Biol 7:46–54.
Kosik KS, Finch EA. 1987. MAP2 and tau segregate into dendritic andaxonal domains after the elaboration of morphologically distinct neu-rites: an immunocytochemical study of cultured rat cerebrum. J Neu-rosci 7:3142–3153.
Lawrence JB, Singer RH. 1986. Intracellular localization of messengerRNAs for cytoskeleton proteins. Cell 45:407–415.
Lendahl U, Zimmerman LB, McKay RDG. 1990. CNS stem cells express anew class of intermediate filament protein. Cell 60:585–595.
Levitt P, Rakic P. 1980. Immunoperoxidase localization of glial fibrillaryacidic protein in radial glial cells and astrocytes of the developingrhesus monkey brain. J Comp Neurol 193:815–840.
Litman P, Barg J, Rindzoonski L, Ginzburg I. 1993. Subcellular localiza-tion of tau mRNA in differentiating neuronal cell culture: implicationsfor neuronal polarity. Neuron 10:627–639.
Ma X, Cole GJ, Juge N, Vincent M. 1999. Different intracellular localiza-tion of transitin isoforms during in vitro differentiation of avian myo-genic cells. Submitted for publication.
Marı́n-Padilla M. 1995. Prenatal development of fibrous (white matter),protoplasmic (gray matter), and layer I astrocytes in the human cere-bral cortex: a golgi study. J Comp Neurol 357:554–572.
McCabe CF, Cole GJ. 1992. Expression of the barrier-associated proteinsEAP-300 and claustrin in the developing central nervous system. BrainRes Dev Brain Res 70:9–24.
McCabe CF, Thompson RP, Cole GJ. 1992. Distribution of the noveldevelopmentally-regulated protein EAP-300 in the embryonic chicknervous system. Brain Res Dev Brain Res 66:11–23.
Morris EJ, Fulton AB. 1994. Rearrangement of mRNAs for costamereproteins during costamere development in cultured skeletal musclefrom chicken. J Cell Sci 107:377–386.
Nakano I, Iwatsubo T, Otsuka N, Kamei M, Matsumura K, Mannen T.1996. Some unusual responses of astrocytes to ghost tangles in a longduration case of juvenile Alzheimer’s disease: an electron microscopicstudy. J Neurol Sci 136:41–46.
Napier A, Yuan Y, Cole GJ. 1999. Characterization of the chicken transitingene reveals a strong relationship to the nestin intermediate filamentclass. J Mol Neurosci 12:11–22.
Page M. 1989. Changing patterns of cytokeratins and vimentin in the earlychick embryo. Development 105:97–107.
Paradies MA, Steward O. 1997. Multiple subcellular mRNA distributionpatterns in neurons: a nonisotopic in situ hybridization analysis.J Neurobiol 33:473–493.
Pixely SKR, de Vellis J. 1984. Transition between immature radial glia andmature astrocytes studies with a monoclonal antibody to vimentin.Brain Res Dev Brain Res 15:201–209.
Sarthy PV, Fu M, Huang J. 1989. Subcellular localization of an interme-diate filament protein and its mRNA in glial cells. Mol Cell Biol9:4556–4559.
Schroder R, Mundegar RR, Treusch M, Schlegel U, Blumcke I, Owaribe K,Magin TM. 1997. Altered distribution of plectin/HD1 in dystrophinopa-thies. Eur J Cell Biol 74:165–171.
Singer RH. 1993. RNA zipcodes for cytoplasmic addresses. Curr Biol3:719–721.
Steward O, Wallace CS, Lyford GL, Worley PF. 1998. Synaptic activationcauses the mRNA for the IEG Arc to localize selectively near activatedpostsynaptic sites on dendrites. Neuron 21:741–751.
Tapscott SJ, Bennett GS, Toyama Y, Kleinbart F, Holtzer H. 1981. Inter-mediate filament proteins in the developing chick spinal cord. Dev Biol86:40–54.
Voigt T. 1989. Development of glial cells in the cerebral wall of ferrets:direct tracing of their transformation from radial glia into astrocytes.J Comp Neurol 289:74–88.
Yuan Y, Lee J-A, Napier A, Cole GJ. 1997. A midline radial glial proteindefines a new class of intermediate filament protein with a novelcentral heptad repeat domain and alternative splice variants lackingthe repeat region. Mol Cell Neurosci 10:71–86.
483TRANSITIN mRNA IS TRANSPORTED IN DEVELOPING RADIAL GLIA





























