Embed Size (px)
Citation preview

Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
Contents lists available at ScienceDirect
Journal of Molecular and Cellular Cardiology
j ourna l homepage: www.e lsev ie r.com/ locate /y jmcc
Original article
Electrical stimulation of sympathetic neurons induces autocrine/paracrine effects ofNGF mediated by TrkA
Erol Saygili a,⁎,1, Patrick Schauerte a,1, Florian Küppers a, Lutz Heck a, Joachim Weis b, Christian Weber c,Robert H.G. Schwinger d, Rainer Hoffmann a, Jörg W. Schröder a, Nikolaus Marx a, Obaida R. Rana a
a Department of Cardiology, University RWTH Aachen, Aachen, Germanyb Institute for Neuropathology, University RWTH Aachen, Aachen, Germanyc Institute for Molecular Cardiovascular Research, University RWTH Aachen, Aachen, Germanyd Medical Clinic II, Klinikum Weiden, Weiden, Germany
Abbreviations: AF, Atrial fibrillation; NRAM, NeonSuperior cervical ganglia; SNS, Sympathetic nerve sprelectrical field stimulation; NGF, Nerve growth factor;Tyrosine kinase A; GAP-43, Growth associated protein 4⁎ Corresponding author. University Hospital RWTH A
ogy,Medical Clinic I, Pauwelsstr. 30, D-52074Aachen, Gerfax: +49 241 8082482.
E-mail address: [email protected] (E. Saygili).1 Both authors contributed equally to this work.
0022-2828/$ – see front matter © 2010 Elsevier Ltd. Adoi:10.1016/j.yjmcc.2010.01.019
a b s t r a c t
a r t i c l e i n f oArticle history:Received 22 October 2009Received in revised form 26 January 2010Accepted 27 January 2010Available online 2 February 2010
Keywords:Atrial fibrillationSympathetic neuronsNerve growth factorNeurotrophin-3Tyrosine kinase A receptor
Neuronal remodeling with increased sympathetic innervation density has been implicated in the patho-genesis of atrial fibrillation (AF). Recently, increased transcardiac nerve growth factor (NGF) levels wereobserved in a canine model of AF. Whether atrial myocytes or cardiac sympathetic neurons are the source ofneurotrophins, and whether NGF is the main neurotrophic factor contributing to sympathetic nerve sprouting(SNS) in AF still remains unclear. Therefore, neonatal rat atrial myocytes were cultured under conditions ofhigh frequency electrical field stimulation (HFES) to mimic rapid atrial depolarization. Likewise, sympatheticneurons from the superior cervical ganglia of neonatal rats were exposed to HFES to simulate the physiologicaleffect of sympathetic stimulation. Real-time PCR, ELISA and Western blots were performed to analyze theexpression pattern of NGF and neurotrophin-3 (NT-3). Baseline NGF and NT-3 content was 3-fold higher insympathetic neurons than in atrial myocytes (relative NGF protein expression: 1±0.0 vs. 0.37±0.11, all n=5,pb0.05). HFES of sympathetic neurons induced a frequency dependent NGF and NT-3 gene and protein up-regulation (relative NGF protein expression: 0 Hz=1±0.0 vs. 5 Hz=1.13±0.19 vs. 50 Hz=1.77±0.08, alln=5, 0 Hz/5 Hz vs. 50 Hz pb0.05), with a subsequent increase of growth associated protein 43 (GAP-43)expression and morphological SNS. Moreover, HFES of sympathetic neurons increased the tyrosine kinase A(TrkA) receptor expression. HFES induced neurotrophic effects could be abolished by lidocaine, TrkA blockadeor NGF neutralizing antibodies, while NT-3 neutralizing antibodies had no significant effect on SNS. In neonatalrat atrial myocytes, HFES resulted inmyocyte hypertrophy accompanied by an increase in NT-3 and a decreasein NGF expression. In summary, this study provides evidence that high-rate electrical stimulation of sym-pathetic neurons mediates nerve sprouting by an increase in NGF expression that targets the TrkA receptor inan autocrine/paracrine manner.
atal rat atrial myocytes; SCG,outing; HFES, High frequencyNT-3, Neurotrophin-3; TrkA,3.achen, Department of Cardiol-many. Tel.:+49241 8036887;
ll rights reserved.
© 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Atrial fibrillation (AF) is a complex arrhythmia that requires atrigger for initiation and a substrate for maintenance. Sympatheticnerve sprouting (SNS)with increased sympathetic innervation density
and sympathetic hyperactivity has been implicated in the generationand maintenance of AF. Different groups in recent studies could con-vincingly demonstrate in animal models as well as in humans thatpersistent AF is accompanied by an increase in tyrosine hydroxylasepositive nerve fibers in the atria [1–4]. Furthermore, Tan et al. revealedin a canine model of paroxysmal AF a positive correlation betweensympathetic hyperinnervation and transcardiac nerve growth factor(NGF) levels [5], indicating a potential role of NGF in AF-associatedSNS. NGF is the best characterized member of the neurotrophin familyresponsible for survival, differentiation and neurite outgrowth of sym-pathetic neurons by targeting the tyrosine kinase A (TrkA) receptor[6]. Whether atrial myocytes or cardiac sympathetic neurons are thesource of NGF in AF, and whether NGF is the main neurotrophic factorcontributing to SNS still remains unclear. Therefore, to clarify thecellular source and the regulation of NGF, we used pure cell cultures

80 E. Saygili et al. / Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
of neonatal rat atrial myocytes and sympathetic neurons from thesuperior cervical ganglia of Sprague Dawley rats under conditionsof high frequency electrical field stimulation (HFES). While HFES inmyocytes mimics rapid atrial electrical depolarization as during AF,in neurocytes HFES simulates the physiological effect of neuronalstimulation and hyperactivity.
Since AF is not only characterized by high-rate atrial activity, aswell as by irregularity, the in-vitro model chosen herein is not trans-ferable to AF in-vivo. High-rate stimulation of neuronal or atrial cellsdoes not resemble the situation of these cells in the fibrillating atrium.
2. Methods
2.1. Cell culture
All animal experiments were approved by the local and state Ethicsin Animal Research Committee. Atrial myocytes (NRAM) and massculture of sympathetic neurons from the superior cervical ganglia(SCG) of neonatal Sprague Dawley rats (Charles River, Germany)wereisolated and cultured as described previously [7,8]. n represents thenumber of cell preparations. For each SCG cell preparation we usedin average 25–30 neonatal rats. For each NRAM cell preparation weused in average 15–20neonatal rats. Regularly the purity of cell culturewas 95% assessed by specific immunocytofluorescence staining fortroponin T-C (for NRAM) or tyrosine hydroxylase (for sympatheticneurons).
2.2. High frequency electrical field stimulation of atrial myocytes andsympathetic neurons
Three days after cell isolation serum containing medium waschanged to serum-free medium and thereafter NRAMs and sympa-thetic neurons were subjected to high frequency bipolar electricalfield stimulation (HFES) for 48 h with the C-PaceEP external pacingsystem (IonOptix, The Netherlands). The ratio of atrial activationduring sinus rhythm compared to AF in humans is nearly 1:10 [9]. Ourprimary cultures of NRAM show inhomogeneously averaged spon-taneous beating rates of up to 3–5 Hz as described previously [8].Therefore, we used a complementary frequency of 50 Hz to simulatethe ratio of 1:10 observed in humans. Cells cultured in parallel with-out HFES were used as controls. Cell cultures of atrial myocytesshowed a capture efficiency of about 90% at electrical field strengthof 1 V/cm and impulse duration (ID) of 1 ms, as confirmed by micro-scopic examination of cell shortening at the beginning and the endof stimulation. Therefore, all experiments were performed at 2-foldthreshold intensity of 2 V/cm at an ID of 1 ms. To assess the integrity ofthe cultures, the numbers of cells of identical microscopic fields werecounted before and after exposure to HFES. If the cell number per fielddeviated by more than 10% cultures were discarded.
2.3. ELISA and Western blot
For NGF and NT-3 ELISA each well was normalized to 300,000 cells(for NRAM) or 100,000 cells (for sympathetic neurons) andHFES stepsfrom 3 Hz up to 5, 20, 50 and 99 Hz were analyzed. The conditionedmedia was collected at 0–24 and 48 h after exposure to HFES. Sampleswere assayed by NGF and NT-3 ELISA kits from R&D Systems (Min-neapolis, USA) according to the manufacturer's instructions. Westernblots were performed with normalized protein amounts of 30 µg/slot[8].
2.4. Immunocytofluorescence
Immunocytofluorescence experiments for tyrosine hydroxylase(TH), growth associated protein-43 (GAP-43) and troponin T-C wereperformed as described previously [7].
2.5. Drugs and antibodies
Drugs: Lidocaine (Sigma-Aldrich) at a concentration of 1 mmol/l was chosen to completely block membrane depolarization [10].
Primary antibodies forWestern blot and immunocytofluorescence:rabbit anti-GAPDH (Cell Signaling), rabbit anti-GAP-43 (Santa Cruz),mouse anti-troponin-T-C (Santa Cruz), mouse anti-TH (Santa Cruz),rabbit anti-NGF (Santa Cruz), goat anti-NT-3 (Santa Cruz), rabbit anti-TrkA (Santa Cruz), mouse anti-beta-myosin heavy chain (beta-MHC,Santa Cruz).
Neurotrophin and TrkA receptor neutralizing antibodies: anti-ratbeta-NGF (R&D Systems). The neutralization dose50 (ND50) of anti-ratβ-NGF antibody was determined to be 0.1 μg/ml; anti-human NT-3(R&D Systems). The ND50 of anti-human NT-3 antibody was deter-mined to be 0.1 mg/ml, anti-TrkA (R&D Systems).
2.6. RNA preparation, first-strand cDNA synthesis and quantitative real-time reverse transcription-PCR
RNA extraction, first-strand cDNA synthesis and quantitativereal-time PCR experiments were performed as described previously[11]. PCR primers and fluorogenic probes for target genes and theendogenous control were purchased from Applied Biosystems (FosterCity, CA). The assay numbers were as follows: Rn00560865_m1(beta-2 microglobulin), Rn01533872_m1 (NGFb), Rn00579280(NT-3), Rn00572130_m1 (TrkA), Rn00561661_m1 (Nppa), andRn00568328_m1 (myh7, beta-myosin heavy chain).
2.7. Statistical analysis
All values are expressed as mean±SEM. Comparisons of twogroups were performed by Student's t-test and multiple groups by1-way ANOVA followed by LSD post-hoc test. Values of pb0.05 wereconsidered statistically significant.
3. Results
3.1. Baseline NGF and NT-3 content is 3-fold more abundant insympathetic neurons than in atrial myocytes
We first quantified relative and absolute values of NGF and NT-3in un-stimulated control cultures. Fig. 1A presents the amount ofNGF and NT-3 in cell-conditioned medium of NRAM and sympatheticneurons, with a range between 35 and 45 pmol/ml. Although NRAMswere cultured with 300,000 cells/well compared to sympatheticneurons with 100,000 cells/well, the absolute NGF and NT-3 con-centration was roughly the same (sympathetic neurons vs. NRAM:NGF 45.34±2.35 pg/ml vs. 44.17±3.27 pg/ml; NT-3 33.71±2.73 pg/ml vs. 35.77±3.03 pg/ml, all n=10), demonstrating analmost 3-fold higher secretion of these neurotrophins by sympatheticneurons. These data could be confirmed by Western blot analysis.Figs. 1B–C shows the relative protein amount of NGF and NT-3 innormalized protein samples (30 µg) of NRAM and sympatheticneurons (sympathetic neurons vs. NRAM: NGF 1±0.0 vs. 0.37±0.11,pb0.05; NT-3 1±0.0 vs. 0.39±0.07, pb0.05, all n=5).
3.2. The impact of HFES on NGF and NT-3 expression by NRAM andsympathetic neurons
NRAM and sympathetic neurons were exposed to HFES for 48 has described in Methods. Since it has been shown that NT-3 is asefficient as NGF in mediating neurite outgrowth in NGF-dependentsympathetic neurons [12], we also examined the expression patternof NT-3 in both cell cultures. In sympathetic neurons HFES induceda significant increase in NGF and NT-3 expression, confirmed bymRNA and protein measurements (relative NGF protein expression:

Fig. 1. Baseline NGF andNT-3 content is 3-foldmore abundant in the cell-conditionedmedium of SCGs than in NRAM. Control cells were cultured without HFES for 48 h in serum-freemedium. (A), ELISA results showing NGF and NT-3 concentrations in cell-conditioned medium of NRAM and sympathetic neurons (n=10). (B–C), Western blots showing relativeprotein amount of NGF and NT-3 in normalized protein samples (30 µg) of NRAM and sympathetic neurons (n=5). *pb0.05 vs. SCG samples.
81E. Saygili et al. / Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
0 Hz=1±0.0 vs. 5 Hz=1.13±0.19 vs. 50 Hz=1.77±0.08, pb0.05,all n=5/relative NT-3 protein expression: 0 Hz=1±0.0 vs. 5 Hz=1.07±0.13 vs. 50 Hz=1.43±0.15, pb0.05, all n=5). In the presenceof lidocaine the HFES induced up-regulation of NGF and NT-3 wascompletely blocked at a frequency of 50 Hz (NGF: 0.93±0.13/NT-3:1.03±0.17, pb0.05, all n=3) (Figs. 2A–D). By contrast, in NRAMHFESinduced a significant down-regulation of NGF, whereas NT-3 mRNAand protein expression increased significantly (relative NGF proteinexpression: 0 Hz=1±0.0 vs. 5 Hz=1.07±0.18 vs. 50 Hz=0.39±0.13, pb0.05, all n=5/relative NT-3 protein expression: 0 Hz=1±0.0 vs. 5 Hz=0.97±0.17 vs. 50 Hz=1.37±0.11, pb0.05, all n=5)(Figs. 2E–H). ELISA experiments with cell-conditioned medium ofNRAM or sympathetic neurons revealed the impact of HFES on NGFand NT-3 release in the cell culture medium. Using ELISAs we also
analyzed the relative changes in NGF and NT-3 secretion 24 and 48 hafter exposure to HFES. In line with real-time PCR or Western blotexperiments, HFES of sympathetic neurons resulted in an up-regulation of NGF and NT-3 protein secretion in the cell culturemedium in a frequency and time dependent manner (Figs. 3A–B).Furthermore, we could observe a statistically significant differencebetween the groups 24 and 48 h after exposure to HFES at frequencieswith 50 and 99 Hz as indicated in Figs. 3A–B (relative NGF secretion at50 Hz: 24 h vs. 48 h, pb0.05; and 99 Hz: 24 h vs. 48 h, pb0.05). In thepresence of lidocaine HFES inducedNGF andNT-3 release into the cell-culture medium of sympathetic neurons was completely blocked atfrequencies of 50 Hz (Figs. 3C–D). By contrast, HFES of NRAM resultedin a down-regulation of NGF release into the cell-culture medium,whereas NT-3 secretion increased significantly (Figs. 3E–F).

Fig. 2. The impact of HFES on NGF and NT-3 expression in sympathetic neurons and NRAM. Both cell cultures were exposed to HFES for 48 h. (A–D), On both the mRNA and proteinlevels, HFES of sympathetic neurons induced a significant increase in NGF and NT-3 expression (n=5). In the presence of lidocaine (1 mmol/l) the HFES induced NGF and NT-3 up-regulation was completely blunted (n=3). (E–H), By contrast in NRAM, HFES induced a down-regulation of NGF whereas NT-3 expression increased significantly (n=5). *pb0.05vs. 0 Hz. #pb0.05 vs. 5 Hz. **pb0.05 vs. 50 Hz.
82 E. Saygili et al. / Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
3.3. HFES contributes to sympathetic nerve sprouting and atrial myocytehypertrophy
In the next step we analyzed morphological changes in sympa-thetic neurons and NRAM induced by HFES. Accompanied by an in-crease in NGF and NT-3 expression, HFES of sympathetic neuronsresulted in morphological SNS with an increase in GAP-43 proteinexpression as a marker for neuronal growth (relative GAP-43 proteinexpression: 0 Hz=1±0.0 vs. 5 Hz=1.2±0.23 vs. 50 Hz=2.3±0.34,pb0.05, all n=5) (Figs. 4A–B). In the presence of lidocaine no SNScould be observed with any increase in GAP-43 protein expression ata frequency of 50 Hz (relative GAP-43 protein expression: 0 Hz=1±0.0 vs. 5 Hz=1.13±0.15 vs. 50 Hz=2.25±0.21, pb0.05/vs. 50 Hz+lidocaine=0.93±0.11, pb0.05, all n=15) (Fig. 5). Atrial myocytesincreased frequency dependent mRNA expression of atrial natriureticpeptide factor (ANP) and both mRNA and protein expression of beta-
myosin heavy chain (b-MHC) as characteristic signs for cardiomyo-cyte hypertrophy (Fig. 4C).
3.4. HFES of sympathetic neurons up-regulates the TrkA receptor
On the receptor level, HFES of sympathetic neurons resulted inan increase in TrkA mRNA and protein expression. Lidocaine com-pletely prevented HFES induced TrkA up-regulation (relative TrkAprotein expression: 0 Hz=1±0.0 vs. 5 Hz=1.07±0.15 vs. 50 Hz=1.79±0.15, pb0.05/vs. 50 Hz+lidocaine=0.97±0.17, pb0.05, alln=5) (Figs. 6A–B). To analyze whether NGF and NT-3 mediatesSNS via the TrkA receptor, TrkA blocking antibodies were appliedduring HFES. Neuronal TrkA inhibition by specific antibodiesabolished the HFES induced neurite outgrowth quantified byWesternblot for GAP-43 protein expression (relative GAP-43 protein ex-pression: 0 Hz=1±0.0 vs. 5 Hz=1.1±0.09 vs. 50 Hz=1.87±

Fig. 3. ELISA measurements revealed the degree of changes in NGF and NT-3 protein secretion in a frequency and time dependent manner. (A–B), HFES of sympathetic neuronsresulted in an increase in NGF and NT-3 protein release into the cell culture medium from 0 up to 99 Hz over a time period of 24 and 48 h (n=15). At the frequencies of 50 Hz and99 Hz we also observed a statistically significant difference between the groups 24 and 48 h. (C–D), Lidocaine was able to block the HFES induced increase in neurotrophin secretionsignificantly (n=5). (E–F), NRAM showed a decrease in NGF secretion and an increase in NT-3 secretion by HFES (n=15). *pb0.05 vs. 0 Hz–5 Hz. #pb0.05 vs. 50 Hz and 99 Hz.¥pb0.05 vs. 24 h.
83E. Saygili et al. / Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
0.13, pb0.05/vs. 50 Hz+lidocaine=0.87±0.11, pb0.05, all n=5)(Fig. 6C).
3.5. SNS via the TrkA receptor is mediated by NGF
We demonstrated that HFES of sympathetic neurons induced anup-regulation of the neurotrophins NGF and NT-3 resulting in mor-phological SNS by targeting the TrkA receptor. To investigate whetherNGF or NT-3 is the main neurotrophic factor contributing to SNS viathe TrkA receptor we performed loss of function experiments usingNGF or NT-3 neutralizing antibodies during HFES. NGF neutralizingantibodies completely prevented the HFES induced SNS confirmed byWestern blotting for GAP-43 protein expression, whereas anti-NT-3had no significant effect on HFES induced SNS (Fig. 7). These datashow that HFES contributes to SNS by an increase in neuronal NGFexpression that targets the TrkA receptor in an autocrine/paracrinemanner.
4. Discussion
The major findings of the present study are: (a) baseline NGF andNT-3 content is 3-fold more abundant in sympathetic neurons thanin atrial myocytes. (b) Electrical stimulation of sympathetic neuronscontributes to morphological SNS by an up-regulation of neuronalNGF expression that targets the TrkA receptor in an autocrine/para-crine manner. (c) NRAMs decrease their NGF expression by HFES,whereas NT-3 expression increases significantly.
4.1. NGF expression and function in target cells and sympathetic neurons
NGF is known as a target-derived protein which plays a crucial rolein sympathetic neuron survival and differentiation [6]. Several studieshave shown that various tissues control their neuronal innervationdensity under physiological or pathological conditions by expressingneurotrophins including NGF [13,14]. Recently we have demonstratedthat ventricularmyocytes for example, express and secrete NGFwhichdecreases under conditions of mechanical stretch, sufficient to sup-press sympathetic axon outgrowth [7]. Beside these observations,Hasan et al. first reported that sympathetic neurons themselves areable to synthesize and secrete NGF which acts in an autocrine/para-crine manner [15].
4.2. HFES contributes to NGF, NT-3 and TrkA up-regulation in sympatheticneurons
In the present study we applied HFES to cultured neurocytes tosimulate the physiological effect of sympathetic stimulation. Sympa-thetic neurons in cell culture respond to HFES with an increase in NGFand NT-3 expression and secretion. In the presence of lidocaine, ablocking agent of the fast voltage gated sodium channels in the cellmembrane, these effects could not be observed. Although NT-3 isreported to be as efficient as NGF in promoting neurite outgrowth anddifferentiation, in our in-vitro model, NGF appears to be the mainneurotrophic factor contributing to SNS (Fig. 8). Even though NT-3has the ability to induce sympathetic nerve outgrowth via the TrkA

Fig. 4. HFES contributes to SNS and myocardial hypertrophy. SNS was confirmed by GAP-43 protein expression measurements in sympathetic neurons. We observed a frequencydependent increase in GAP-43 protein expression as shown in figure (A–B) (all n=5). (In figure (A)—yellow staining indicates GAP-43 protein expression and red staining indicatesTH positive neurons. Atrial myocytes showed an increase in the expression of characteristic hypertrophy markers (C) (all n=7). *pb0.05 vs. 0 Hz. #pb0.05 vs. 5 Hz.
84 E. Saygili et al. / Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
receptor, its binding affinity to TrkA is lower and is also inconstantas compared to NGF [16]. Furthermore, NGF is almost 10-fold moreefficient than NT-3 in mediating acute, short-term TrkA activity asreported by Belliveau et al., too [12]. Due to the fact that our cell cul-ture experiments do not represent a chronic stimulation over severaldays or weeks (HFES was exposed for 48 h), we cannot exclude apossible significant effect of NT-3 on sympathetic nerve outgrowthin a chronic scenario.
In parallel to the increase of NGF, HFES of sympathetic neuronsalso increased the expression of the TrkA receptor, confirmed bymRNA and protein measurements. These data highlight the impor-tance of physiological positive feed-back loops, which in some casesare boostered by pathological conditions. For example, a current studyby Tsunoda et al. revealed that an up-regulation in NGF expressioncorrelated with an increase in TrkA receptor expression in oesopha-geal squamous cell carcinoma [17]. Thus, the fact that sympatheticneurons respond to HFESwith an increase in TrkA receptor expressioneven though NGF is up-regulated strengthens the hypothesis thatneuronal remodeling once occurred is a self amplifying mechanismwhich is coupled to a NGF-TrkA autocrine positive feed-back loop.
4.3. HFES contributes to NGF down-regulation and NT-3 up-regulation inatrial myocytes
During AF the atria undergoes structural remodeling processesresulting in hypertrophy and fibrosis. Goette et al. could convincinglydemonstrate that AF alters intracellular calcium homeostasis and in-duces cellular hypertrophy of atrial myocytes via the calcium-depen-dent calcineurin-NFAT pathway [18].
Although, in the present study we did not investigate a possibleinvolvement of the Cn-NFAT pathway, our data are in line with theseobservations. In the present study, we applied HFES to cultured atrialmyocytes to simulate rapid atrial depolarization as during AF. Wefound that HFES contributes to myocyte hypertrophy confirmed byANP mRNA expression and beta-MHC mRNA and protein expression.These observations were accompanied by a down-regulation of NGFand an up-regulation of NT-3 expression in a time and frequencydependent manner. NGF is not only known as a neurotrophic factorfor survival and differentiation of neuronal cell lines, it acts also asa pro-survival factor for cardiomyocytes [19]. Likewise, in a recentstudy we demonstrated that a down-regulation of myocyte NGF
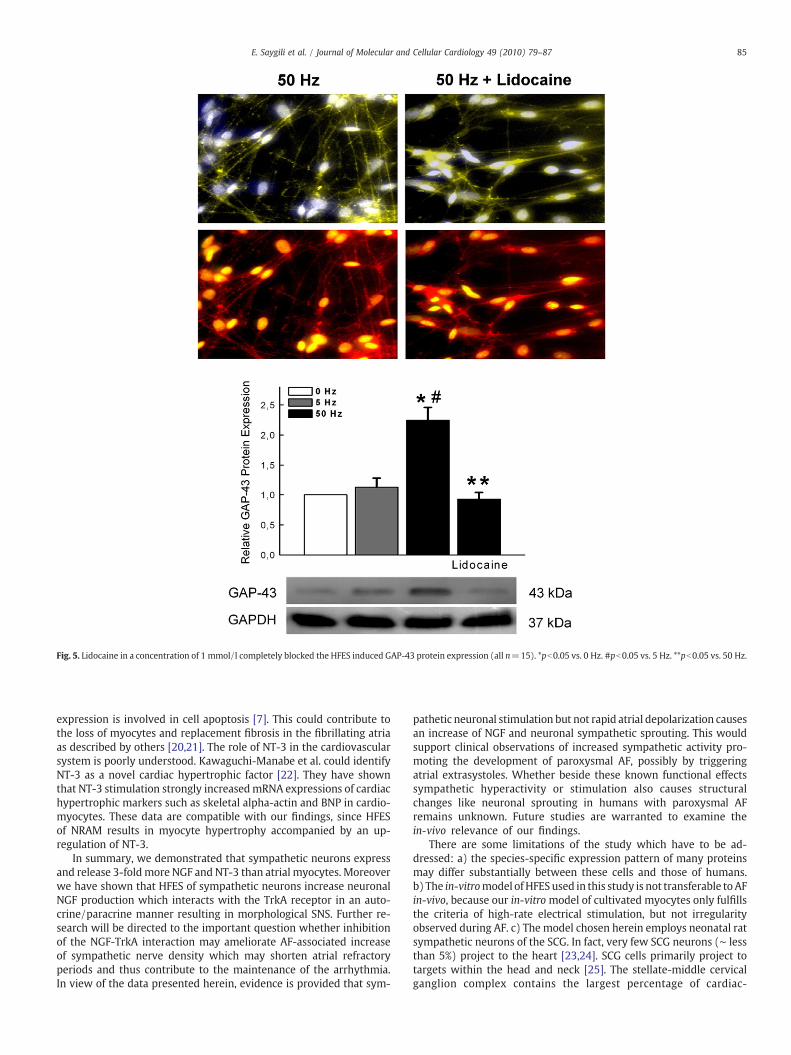
Fig. 5. Lidocaine in a concentration of 1 mmol/l completely blocked the HFES induced GAP-43 protein expression (all n=15). *pb0.05 vs. 0 Hz. #pb0.05 vs. 5 Hz. **pb0.05 vs. 50 Hz.
85E. Saygili et al. / Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
expression is involved in cell apoptosis [7]. This could contribute tothe loss of myocytes and replacement fibrosis in the fibrillating atriaas described by others [20,21]. The role of NT-3 in the cardiovascularsystem is poorly understood. Kawaguchi-Manabe et al. could identifyNT-3 as a novel cardiac hypertrophic factor [22]. They have shownthat NT-3 stimulation strongly increasedmRNA expressions of cardiachypertrophic markers such as skeletal alpha-actin and BNP in cardio-myocytes. These data are compatible with our findings, since HFESof NRAM results in myocyte hypertrophy accompanied by an up-regulation of NT-3.
In summary, we demonstrated that sympathetic neurons expressand release 3-foldmore NGF and NT-3 than atrial myocytes. Moreoverwe have shown that HFES of sympathetic neurons increase neuronalNGF production which interacts with the TrkA receptor in an auto-crine/paracrine manner resulting in morphological SNS. Further re-search will be directed to the important question whether inhibitionof the NGF-TrkA interaction may ameliorate AF-associated increaseof sympathetic nerve density which may shorten atrial refractoryperiods and thus contribute to the maintenance of the arrhythmia.In view of the data presented herein, evidence is provided that sym-
pathetic neuronal stimulation but not rapid atrial depolarization causesan increase of NGF and neuronal sympathetic sprouting. This wouldsupport clinical observations of increased sympathetic activity pro-moting the development of paroxysmal AF, possibly by triggeringatrial extrasystoles. Whether beside these known functional effectssympathetic hyperactivity or stimulation also causes structuralchanges like neuronal sprouting in humans with paroxysmal AFremains unknown. Future studies are warranted to examine thein-vivo relevance of our findings.
There are some limitations of the study which have to be ad-dressed: a) the species-specific expression pattern of many proteinsmay differ substantially between these cells and those of humans.b) The in-vitromodel ofHFESused in this study is not transferable toAFin-vivo, because our in-vitromodel of cultivated myocytes only fulfillsthe criteria of high-rate electrical stimulation, but not irregularityobserved during AF. c) The model chosen herein employs neonatal ratsympathetic neurons of the SCG. In fact, very few SCG neurons (∼ lessthan 5%) project to the heart [23,24]. SCG cells primarily project totargets within the head and neck [25]. The stellate-middle cervicalganglion complex contains the largest percentage of cardiac-
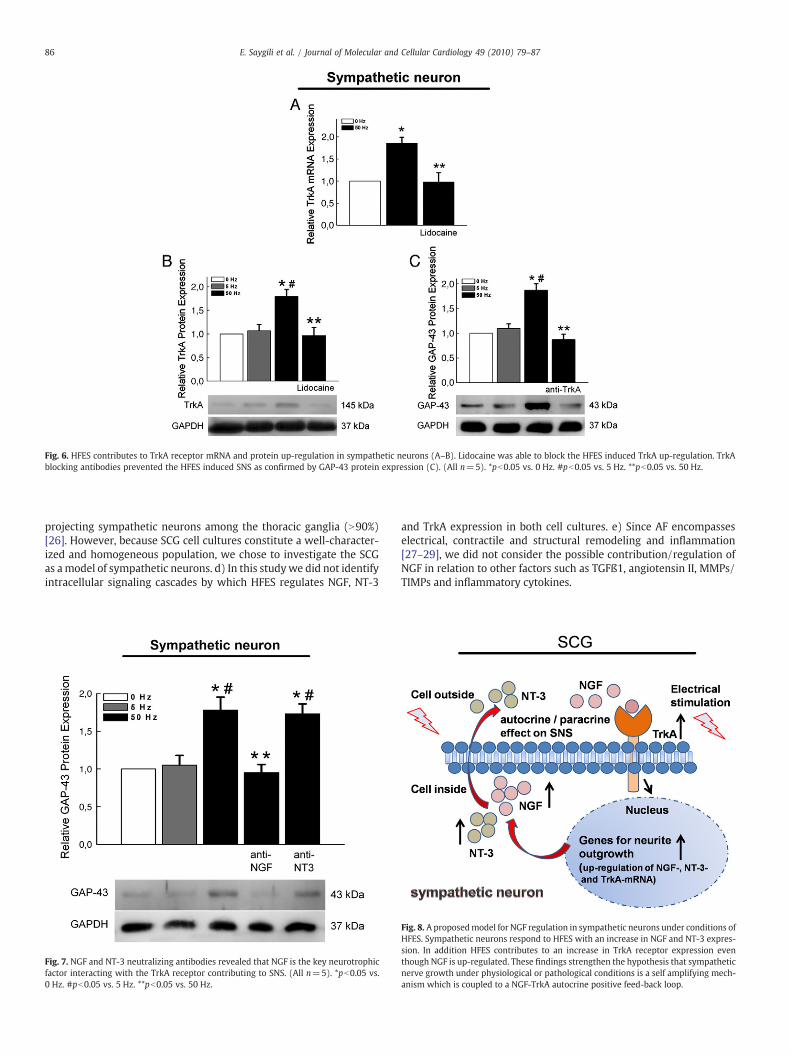
Fig. 6. HFES contributes to TrkA receptor mRNA and protein up-regulation in sympathetic neurons (A–B). Lidocaine was able to block the HFES induced TrkA up-regulation. TrkAblocking antibodies prevented the HFES induced SNS as confirmed by GAP-43 protein expression (C). (All n=5). *pb0.05 vs. 0 Hz. #pb0.05 vs. 5 Hz. **pb0.05 vs. 50 Hz.
86 E. Saygili et al. / Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
projecting sympathetic neurons among the thoracic ganglia (N90%)[26]. However, because SCG cell cultures constitute a well-character-ized and homogeneous population, we chose to investigate the SCGas amodel of sympathetic neurons. d) In this studywe did not identifyintracellular signaling cascades by which HFES regulates NGF, NT-3
Fig. 7. NGF and NT-3 neutralizing antibodies revealed that NGF is the key neurotrophicfactor interacting with the TrkA receptor contributing to SNS. (All n=5). *pb0.05 vs.0 Hz. #pb0.05 vs. 5 Hz. **pb0.05 vs. 50 Hz.
and TrkA expression in both cell cultures. e) Since AF encompasseselectrical, contractile and structural remodeling and inflammation[27–29], we did not consider the possible contribution/regulation ofNGF in relation to other factors such as TGFß1, angiotensin II, MMPs/TIMPs and inflammatory cytokines.
Fig. 8. A proposedmodel for NGF regulation in sympathetic neurons under conditions ofHFES. Sympathetic neurons respond to HFES with an increase in NGF and NT-3 expres-sion. In addition HFES contributes to an increase in TrkA receptor expression eventhough NGF is up-regulated. These findings strengthen the hypothesis that sympatheticnerve growth under physiological or pathological conditions is a self amplifying mech-anism which is coupled to a NGF-TrkA autocrine positive feed-back loop.

87E. Saygili et al. / Journal of Molecular and Cellular Cardiology 49 (2010) 79–87
Acknowledgments
This work contains data from the doctoral thesis of Florian Küppersand Lutz Heck (University RWTH Aachen, Aachen, Germany).
References
[1] Chang CM, Wu TJ, Zhou S, Doshi RN, Lee MH, Ohara T, et al. Nerve sprouting andsympathetic hyperinnervation in a canine model of atrial fibrillation produced byprolonged right atrial pacing. Circulation 2001;103:22–5.
[2] Gould PA, Yii M, McLean C, Finch S, Marshall T, Lambert GW, et al. Evidencefor increased atrial sympathetic innervation in persistent human atrial fibrillation.Pacing Clin Electrophysiol 2006;29:821–9.
[3] Hamabe A, Chang CM, Zhou S, Chou CC, Yi J, Miyauchi Y, et al. Induction of atrialfibrillation and nerve sprouting by prolonged left atrial pacing in dogs. Pacing ClinElectrophysiol 2003;26:2247–52.
[4] Nguyen BL, Fishbein MC, Chen LS, Chen PS, Masroor S. Histopathological substratefor chronic atrial fibrillation in humans. Heart Rhythm 2009;6:454–60.
[5] Tan AY, Zhou S, Ogawa M, Song J, Chu M, Li H, et al. Neural mechanisms ofparoxysmal atrial fibrillation and paroxysmal atrial tachycardia in ambulatorycanines. Circulation 2008;118:916–25.
[6] Levi-Montalcini R. The nerve growth factor: its role in growth, differentiation andfunction of the sympathetic adrenergic neuron. Prog Brain Res 1976;45:235–58.
[7] Rana OR, Saygili E, Meyer C, Gemein C, Krüttgen A, Andrzejewski MG, et al.Regulation of nerve growth factor in the heart: the role of the calcineurin–NFATpathway. J Mol Cell Cardiol 2009;46:568–78.
[8] Saygili E, Rana OR, Meyer C, Gemein C, Andrzejewski MG, Ludwig A, et al. Theangiotensin–calcineurin–NFAT pathway mediates stretch-induced up-regulationof matrix metalloproteinases-2/-9 in atrial myocytes. Basic Res Cardiol 2009;104:435–48.
[9] Lin YJ, Tai CT, Kao T, Tso HW, Higa S, Tsao HM, et al. Frequency analysis in differenttypes of paroxysmal atrial fibrillation. J Am Coll Cardiol 2006;47:1401–7.
[10] Scholz A, Kuboyama N, Hempelmann G, Vogel W. Complex blockade of TTX-resis-tant Na+ currents by lidocaine and bupivacaine reduce firing frequency in DRGneurons. J Neurophysiol 1998;79:1746–54.
[11] Saygili E, Rana OR, Saygili E, Reuter H, Frank KF, Schwinger RH, et al. Losartanprevents stretch induced electrical remodeling in cultured atrial neonatalmyocytes.Am J Physiol Heart Circ Physiol 2007;292:H2898–905.
[12] Belliveau DJ, Krivko I, Kohn J, Lachance C, Pozniak C, Rusakov D, et al. NGF andneurotrophin-3 both activate TrkA on sympathetic neurons but differentiallyregulate survival and neuritogenesis. J Cell Biol 1997;136:375–88.
[13] Snider WD. Functions of the neurotrophins during nervous system development:what the knockouts are teaching us. Cell 1994;77:627–38.
[14] Hassankhani A, Steinhelper ME, Soonpaa MH, Katz EB, Taylor DA, Andrade-Rozental A, et al. Overexpression of NGFwithin the heart of transgenic mice causeshyperinnervation, cardiac enlargement, and hyperplasia of ectopic cells. Dev Biol1995;169:309–21.
[15] Hasan W, Pedchenko T, Krizsan-Agbas D, Baum L, Smith PG. Sympathetic neuronssynthesize and secrete pro-nerve growth factor protein. J Neurobiol 2003;57:38–53.
[16] Reichardt LF. Neurotrophin-regulated signaling pathways. Philos Trans R Soc LondB Biol Sci 2006;361:1545–64.
[17] Tsunoda S, Okumura T, Ito T,Mori Y, SomaT,Watanabe G, et al. Significance of nervegrowth factor overexpression and its autocrine loop in oesophageal squamous cellcarcinoma. Br J Cancer 2006;95:322–30.
[18] Bukowska A, Lendeckel U, Hirte D, Wolke C, Striggow F, Röhnert P, et al. Activationof the calcineurin signaling pathway induces atrial hypertrophy during atrial fibril-lation. Cell Mol Life Sci 2006;63:333–42.
[19] Caporali A, Sala-Newby GB, Meloni M, Graiani G, Pani E, Cristofaro B, et al. Iden-tification of the prosurvival activity of nerve growth factor on cardiac myocytes.Cell Death Differ 2008;15:299–311.
[20] Grammer JB, Böhm J, Dufour A, Benz M, Lange R, Bauernschmitt R. Atrial fibrosis inheart surgery patients decreased collagen III/I ratio in postoperative atrial fibril-lation. Basic Res Cardiol 2005;100:288–94.
[21] Boldt A, Wetzel U, Lauschke J, Weigl J, Gummert J, Hindricks G, et al. Fibrosis in leftatrial tissue of patients with atrial fibrillation with and without underlying mitralvalve disease. Heart 2004;90:400–5.
[22] Kawaguchi-ManabeH, IedaM, KimuraK,Manabe T,Miyatake S, KanazawaH, et al. Anovel cardiac hypertrophic factor, neurotrophin-3, is paradoxically downregulatedin cardiac hypertrophy. Life Sci 2007;81:385–92.
[23] Pardini BJ, Lund DD, Schmid PG. Organization of the sympathetic postganglionicinnervation of the rat heart. J Auton Nerv Syst 1989;28:193–201.
[24] Richardson RJ, Grkovic I, Allen AM, Anderson CR. Separate neurochemical classesof sympathetic postganglionic neurons project to the left ventricle of the rat heart.Cell Tissue Res 2006;324:9–16.
[25] Flett DL, Bell C. Topography of functional subpopulations of neurons in the superiorcervical ganglion of the rat. J Anat 1991;177:55–66.
[26] Yang Q, Sumner AD, Puhl HL, Ruiz-Velasco V. M(1) and M(2) muscarinic acetyl-choline receptor subtypes mediate Ca(2+) channel current inhibition in rat sym-pathetic stellate ganglion neurons. J Neurophysiol 2006;96:2479–87.
[27] Boldt A, Scholl A, Garbade J, ResetarME, Mohr FW, Gummert JF, et al. ACE-inhibitortreatment attenuates atrial structural remodeling in patients with lone chronicatrial fibrillation. Basic Res Cardiol 2006;101:261–7.
[28] Dobrev D, Ravens U. Remodeling of cardiomyocyte ion channels in human atrialfibrillation. Basic Res Cardiol 2003;98:137–48.
[29] Rana OR, Zobel C, Saygili E, Brixius K, Gramley F, Schimpf T, et al. A simple deviceto apply equibiaxial strain to cells cultured on flexible membranes. Am J PhysiolHeart Circ Physiol 2008;294:H532–40.





























