Embed Size (px)
Citation preview

GENES, CHROMOSOMES & CANCER 3:101-107 (1991)
Interphase Cytogenetics of Brain Tumors Edo P.J. Arnoldus, lnge A. Noordermeer, A.C. Boudewijn Peters, Joan H.C. Voormolen, Gerard T.A.M. Bob, Anton K. Raap, and Mels van der Ploeg
Departments of Neurology (E.P.J.A., I.A.N., A.C.B.P.), Neurosurgery (J.H.C.V.), Neuropathology (G.T.A.M.B.), and Cytochemistry and Cytometry (E.P.J.A., I.A.N.. A.K.R., M.v.d.P.), Leiden University, Leiden, The Netherlands
The development and application of a procedure for interphase cytogenetics on brain tumor material is described. Nuclei isolated from freshly removed brain tumor tissue were investigated for chromosomal aberrations by nonradioactive in situ hybridization with a panel of chromosome-specific probes. The panel consisted of nine satellite DNA probes specific for the centromeric regions of chromosomes I, 6,7, 10, I I, 17, 18, X, and Y. For each probe, the number of hybridization signals per cell was determined in 200 nuclei. It was inferred from the hybridization results that in I I gliomas (seven astrocytomas grade 11-IV, three oligodendrogliomas, and one ependymoma) the numerical aberrations were gains of chromosomes I (once), 7 (twice), 10 (once), I I (twice), and X (twice); losses of chromosomes I (once), 10 (twice), 17 (twice), and Y (once); and complete tetraploidy (once). Among the I8 investigated meningiomas monosomy I8 and trisomy I7 were observed once and twice, respectively. An additional hybridization with a cosmid probe for the BCR gene on 22q I I indicated monosomy 22q in I I meningiomas. These results show the value of interphase cytogenetics for the analysis of solid tumors for which it is relatively difficult to obtain sufficient metaphases of good quality for conventional cytogenetics.
INTRODUCTION
During the last few years a number of reports have been published regarding the cytogenetic fea- tures of brain tumors. Metaphase spreads of glio- mas often show double minutes, a gain of chromo- some 7, and losses of chromosomes 10 and 22 and sex chromosomes (Bigner et al., 1986, 1988, 1990; Rey et al., 1987a,b; Jenkins e t al., 1989). In the majority of meningiomas, the only aberration ob- served in metaphase preparations is usually mono- somy 22 (Zang and Singer, 1967; Mark, 1973; Zang, 1982; A1 Saadi et al., 1987; Casalone et al., 1987, 1990; Maltby et al., 1988). So far, cytoge- netic changes of these solid tumors have been stud- ied mainly by karyotyping. Tissue culture of tumor material is notoriously difficult (Teyssier, 1989): Not only can cultures fail completely but they can also lead to either selective growth or spontaneous transformation of the karyotype. In addition, the yield of good quality metaphases is usually small. Therefore, approaches for the study of cytogenetic changes in solid tumors, that make culturing of the tumor material unnecessary, are urgently needed.
Taking into consideration that chromosomal ab- errations may change the normal DNA content of cells, measurement of that content by DNA flow cytometry is one of the possible choices. However, the smallest changes in nuclear DNA content de- tectable are on the order of a few percent (Corne- lisse and Tanke, 1990). A dramatic genetic change, such as the loss of a chromosome 22 (0.86% of the genome; Mendelsohn et al., 1973) in meningio- 0 1991 WILEY-LISS. INC.
mas, will actually fall below the detection thresh- old (Raap et al., 1987). Furthermore, even when an abnormal DNA content would be found, it is impossible to detect the exact structural cytoge- netic changes that have occurred.
The in situ hybridization technique with chro- mosome-specific DNA probes seems to provide a better alternative (Raap et al., 1990). An important advantage of this approach is that it can be applied to interphase nuclei as well as metaphase chromo- somes. Interphase cytogenetics (Cremer et al., 1986) not only overcomes the necessity for cultur- ing, but provides the possibility to increase the number of cells to be studied. In this way, the method can be very useful to identify tumor sub- populations with specific chromosome aberrations. In cytogenetic studies of solid tumors, interphase cytogenetics has been shown to be a reliable pro- cedure in the study of bladder carcinomas (Hop- man et al., 1988, 1989), breast tumors (Devilee et al., 1988b), testicular tumors (Walt et al., 1989), and gastric tumors (Van Dekken et al., 1990).
For the interpretation of interphase cytogenetics results, one should realize that information is gen- erated only for those chromosome segments to which the probes hybridize. Three signals in an interphase nucleus after hybridization with a probe specific for the centromeric region may reflect the presence of three complete copies of that chromo-
Received October 1 , 1990; accepted November 18, 1990. Address reprint requests to Edo P.J. Arnoldus, Department of
Cytochemistry and Cytometry, Leiden University, Wassenaarseweg 72, 2333 AL Leiden, The Netherlands.

ARNOLDUS ET AL 102
some. However, a centromeric sequence present in, e.g., a marker chromosome or a dicentric chro- mosome can also cause the third signal. Therefore, one must be careful with the interpretation of the interphase cytogenetics results. A combination of GTG-banding and in situ hybridization, as de- scribed by Smit e t al. (1990), can improve the char- acterization of complex karyotypes, although such an approach has its own limitations.
In this report, the development and application of a reliable procedure for sensitive interphase cy- togenetics on brain tumors is described. The im- plications of this technique for the study of karyo- typic changes in brain tumors are discussed.
MATERIALS AND METHODS Tumor Material
Fresh tumor tissue of seven astrocytomas (grade 11-IV, according to WHO classification; Zulch, 1979), three oligodendrogliomas, one papillary ependymoma, and 18 meningiomas were collected in culture medium during neurosurgery. The size of the specimen varied from 0.02 to 1.0 cm3. At the neuropathological examination of this material, special care was given to prevent sampling errors for the in situ hybridization studies.
T h e glioma group included eight males and three females, with a mean age of 43 years. In the meningioma group, there were seven males and 1 1 females, with a mean age of 56 years. The clinico- pathological data of the 29 patients are summarized in Tables 2 and 3.
Preparation of Nuclei Suspensions
The tumor tissue was mechanically disaggre- gated with a Potter-Elvehjem homogenizer (with a
TABLE I , Hybridization Results on Tumor A4*
Observer A Observer B (spotdnucleus) (spotdnucleus)
Chromosome I 2 3 I 2 3
I 6 7
10 I I 17 18 X Y
7 5 I
79 4 8
12 99 99
90 3 3 9 5 2 95 0 4 96 0 28 71 I 30 69 21 0 78 22 0 96 0 7 93 0 92 0 10 88 2 88 0 7 92 I
I 0 9 8 2 0 I 0 99 I 0
* For nine centromere-specific probes, we present the percentages of nuclei with one, two, or three pots as determined by two independent observers (number of nuclei counted = 200).
TABLE 2. Summary of the Chromosomal Aberrations Observed in I I Gliomas by Interphase Cytogenetics Using a
Panel of Nine Centromere-Specific Probes
Histology and Aberration
No. Sex (years) grade (% nuclei) Age
A l M 33 A2 M 42 A3 M 33 A4 M 56 A5 M 62 A6 M 36 A7 M 52 E l F 52
01 F 29 0 2 M 39 0 3 F 43
Astrocytoma II Astrocytoma II Normal Astrocytoma ll-Ill Tetraploid (34) Astrocytoma 111 Astrocytoma 111 - 10 (82) Astrocytoma IV Astrocytoma IV Papi I lary
- I7 ? (28)
- 10 (79), +7 (70)
+/+ +7 (I 2/25) -Y (loo), +X (80) +/+ + I (27/47)
ependymoma 111 +I++ 10 (30/40)
+/++X (16169) + I I (49)
Oligodendroglioma Normal Oligodendroglioma + I I (72) Oligodendroglioma - I (36), - 17 ? (26)
TABLE 3. Summary of the Chromosomal Aberrations Observed in 18 Meningiomas by lnterphase Cytogenetics Using a Panel of Nine Centromere-Specific Probes Plus a
Cosmid Probe for BCR on 22q I I
Age No. Sex (years) Type’ Aberration (% nuclei)
MI M2 M3 M4 M5 M6 M7 M8 M9 MI0 MI I T I T2 T3 T4 F I HI H2
M 30 F 41 F 54
M 60 F 67 F 67
M 68 F 68
M 69 M 76 F 83 F 43
M 43 F 51 F 57 F 41
M 35 F 49
men. men. men. men. men. men. men. men. men. men. men. tran. tran. tran. tran. fibr. hem. hem.
+ I7 (35)
Normal
Normal
Normal
+ I7 (81) Normal
- I8 (59) Normal Normal
-22 (50)
-22 (43) -22 (55)
-22 (39)
-22 (80) -22 (62) -22 (86)
-22 (58)
-22 (68) -22 (46)
-22 (49)
a men., meningothelid; tran.. transitional; fibr.. fibrous hem., hemangio- pericytic.
piston+ylindrical vessel distance of 200 pm). The suspension was fixed in 1% acid-free formaldehyde (Merck No. 3999, Darmstadt, Germany), 0.9% NaCl, 10 mM MgSO,. After centrifuging for 10 min at 400g, the pellet was dissolved in lysis buffer (0.15 M NH,Cl, 10 mM KHCO,, 10 mM EDTA, pH 7.4) to lyse the remaining erythrocytes. The

INTERPHASE CYTOGENETICS OF BRAIN TUMORS 103
cells were centrifuged again, after which the pellet was resuspended in 0.5% (w/v) pepsin (Serva No. 31855, Heidelberg, Germany) in 0.9% NaCl, 10 mM MgSO, (pH adjusted to 2.0 with 1 M HCl). The resulting suspension was filtered through a nylon filter of 80 pm to remove remaining clumps of cells. Finally, the suspension of nuclei was fixed several times in cold 70% ethanol and stored at - 20°C. Shortly before the in situ hybridization ex- periment, the nuclei were centrifuged onto glass slides with a bucket centrifuge as previously de- scribed (Arnoldus et al., 1989).
Probes and Labeling
In this study nine probes specific for the (per;)- centromeric regions of chromosomes 1 (pUC1.77; Cooke and Hindley, 1979), 6 (p308; Jabs et al., 1984), 7 (p7tl; Waye et al., 1987b), 10 (DlOZl; Devilee et al., 1988a), 11 (pLCllA, Waye et al., 1987a), 17 (p17H8; Waye and Willard 1986), 18 (L1.84; Devilee e t al., 1986), X (pBamX5; Willard et al., 1983), and Y (DYZ3; Cooke et al., 1982) were used. These probes, which recognize highly repetitive satellite DNA sequences clustered around the centromere, were selected on basis of their relevance for brain tumors and their good ap- plicability.
For the detection of chromosome 22 aberrations in meningiomas, the cosmid probe Cos-BCR-19-1 was used, because we found that the probe for the chromosome 22 centromeric region (D2223; Metz- dorf et al., 1988) is not specific enough for this purpose. This cosmid probe which contains a 34 kb fragment of the BCR gene on 22qll (Hermans et al., 1989) was combined with the cosmid clone Cos-ABL-18, which contains a 40 kb fragment of the human ABL gene in the q34 band of chromo- some 9 (Heisterkamp et al., 1983), as a control for hybridization efficiency in a double hybridization.
T h e probes were labeled by standard nick- translation with either biotin-1 1-dUTP (Sigma, St. Louis, MO) or digoxigenin-1 1-dUTP (Boehringer, Mannheim, Germany). The labeled DNA was eth- anol-precipitated with salmon sperm DNA and yeast RNA as a carrier. The precipitate was dis- solved in a mixture of 60% deionized formamide (v/v), 2x SSC ( l x SSC = 0.15 M NaCl, 0.015 M sodium citrate) to a probe concentration of 1 ng/pl of pUC1.77 and 2.5 ng/pl of the other satellite DNA probes.
In the double cosmid in situ hybridization ex- periments the hybridization mixtures contained 20 ng of each labeled cosmid probe plus a 500-fold excess of human genomic DNA as a competitor,
dissolved in 10 pl of 50% formamide, 10% dextran sulphate, and Z X SSC.
In Situ Hybridization (ISH)
Before hybridization, cells were treated with pepsin for 10 min at 37°C (0.01% pepsin [Serva 318551 in 0.01 M HCl), and postfixed for 10 min in 1% acid-free formaldehyde in PBS. For the satel- lite DNA probes 5 p1 of the hybridization mixtures were added under a coverslip of 18 X 18 mmz, and probe and target DNA were denatured simulta- neously for 10 min at 80°C. For the double cosmid hybridization the slides and the hybridization mix- ture were denatured separately. Then 10 pl of the hybridization mixture was applied and sealed un- der a 18 X 18 mm2 coverslip with a rubber solution. In situ hybridization was allowed to take place overnight at 37°C in a moist chamber with 60% formamide and 2X SSC.
lrnrnunocytochernical Detection
After hybridization, the slides that were hybrid- ized with the satellite DNA probes were washed in 60% formamide, 2X SSC at 37°C. T h e hybrids were detected by three consecutive immunocyto- chemical steps of 30 min at 37°C in 4X SSC, 5% nonfat dry milk. The first and third layer contained 5 pg avidin-D-FITC/ml (Vector Laboratories, Bur- lingame, CA). T h e second layer consisted of bio- tinylated goat antiavidin (Vector), 1: 100. Finally, the cells were dehydrated and embedded in a glyc- erol mixture with propidium iodide for counter- staining and the antifade reagent DABCO (Sigma).
For the double cosmid hybridization, the slides were washed in 50% formamide, 2x SSC at 45"C, followed by washes in 0.1 X SSC at 60°C. T h e im- munocytochemical detection consisted of four steps of 30 min at 37°C. T h e first layer contained 5 p.g avidin-D-FITC/ml in 4X SSC, 5% nonfat dry milk. The second layer consisted of biotinylated goat antiavidin, 1: 100, plus mouse antidigoxigenin (a generous gift from Boehringer, Tutzing, Ger- many), 1:1,000. T h e third layer was avidin-FITC plus rabbit antimouse-TRITC, 1: 1,000; the fourth layer was goat antirabbit-TRITC, 1: 1,000. T h e last three layers were diluted in 0.1 M Tris HCl (pH 7.5), 0.15 M NaCl, and 0.5% Boehringer blocking reagent. Finally, the cells were dehy- drated and embedded in a glycerol mixture with DAPI for counterstaining and DABCO.
Evaluation and Interpretation of In Situ Hybridization Signals
The evaluation of the in situ hybridization sig-

104 ARNOLDUS ET AL.
nals was performed using similar criteria as previ- ously described (Hopman et al., 1988; Arnoldus et al., 1989). In contrast to the method of Hopman et al., nuclei without any signals were also counted. For all probes used, the number of signals per nu- cleus after hybridization was counted in 200 nuclei. As can be seen in Table 1, the interobserver vari- ability was low.
For the sake of simplicity, we have assumed that the actual copy number of a certain chromosome was identical to the number of hybridization signals obtained with its specific centromeric probe. Thus two clear spots per nucleus represent the normal two copies of the chromosome. Nuclei with only one signal may reflect a monosomy for that specific chromosome. T o rule out the possibility that the in situ hybridization partly failed in these nuclei, dou- ble hybridizations with another probe were per- formed in those cases (Fig. lC,D). However, it is not uncommon to have 1040% nuclei with only one signal in normal tissue (Devilee et al., 1988b; Nederlof et al., 1989). In this study, therefore, one signal per nucleus was only interpreted as an indi- cation of a monosomy when it was present in more than 20% of the nuclei. More than two signals usu- ally give less interpretation problems and in this case a value of more than 7.5% of nuclei with three signals was used to indicate a subpopulation with a trisomy.
RESULTS
Results of the experiments with the panel of satellite DNA probes on glioma interphase nuclei are summarized in Table 2. Only two of the ten investigated gliomas (one astrocytoma grade I1 and one oligodendroglioma) did not show any abnor- malities with this panel of probes. The observed aberrations in the seven astrocytomas 11-IV were: - 10 (twice), - 17 (once) and - Y (once); +/+ + 7 (twice), and + X (once) and complete tetraploidy (once). T h e papillary ependymoma displayed: +/+ + 1, +/+ + 10, + 11, and +/+ +X. The ob- served aberrations in the three oligodendrogliomas were: - 1 (once), - 17 (once), and + 11 (once). For chromosome 17, control experiments on nor- mal brain tissue revealed that the centromeres are often paired in astrocytes, resulting in only one, slightly larger, spot per nucleus (Arnoldus et al., 1990a). This complicates the interpretation of the apparent loss of a chromosome 17 (indicated by a question mark in Table 2) observed in 28% and 26% of the nuclei of tumors A1 and 0 3 , respec- tively.
The results of in situ hybridization on meningi-
omas are summarized in Table 3. With the panel of probes specific for the centromeric regions only three changes were found in 18 meningiomas: - 18 (once) and +17 (twice). T h e hybridization with the cosmid probes for BCR (22qll) and ABL (9q34) revealed more aberrations. Eleven of 18 meningi- omas showed one signal for 22qll and two signals for 9q34 in 39-86% of the nuclei, implicating a loss of 22q (Fig. lE,F). The majority of the remaining nuclei showed two signals for both probes.
DISCUSSION
The first important step in a protocol for inter- phase cytogenetics is the preparation of a suspen- sion of intact nuclei from freshly collected (brain) tumor tissue that can be stored until use. Since the clinical samples are often small, it is important to obtain high nuclear yields. Mechanical disaggrega- tion with a Potter-Elvehjem homogenizer gave a much higher yield than simple cutting with razor blades; even the smallest clinical specimen (0.02 cm3) thus rendered sufficient nuclei for 16 hybrid- izations. Pepsin was then used to disaggregate the remaining clumps of cells into single nuclei. Fi- nally, the nuclei were fixed with 70% ethanol, after which the suspensions could be kept at - 20°C for months without loss of in situ hybridization effi- ciency.
Our hybridization results showed that chromo- somal abnormalities frequently occur in gliomas. Often, however, only a fraction of the nuclei showed a chromosomal aberration. Although spe- cial care was taken by the neurosurgeons to make sure that the samples used for interphase cytoge- netics consisted of tumor tissue only, it cannot be ruled out completely that they also contained some normal cells. This implicates that the percentage of nuclei with a certain aberration might be underes- timated. For instance, 79% of the nuclei of tumor A4 displayed one signal for the centromeric region of chromosome 10, whereas the remaining 21% showed the normal number of two signals. This might reflect the presence of different subpopula- tions of the tumor, but it could also be caused by normal cells in the neurosurgical sample.
Interestingly, only two of 11 gliomas (18%) showed a trisomy for chromosome 7. This is less frequent than the 50% that Rey et al. (1987b) re- ported in cultured material. Recently Heim et al. (1989) published findings of chromosomal aberra- tions in normal brain tissue after short-term cul- tures. The changes observed were - Y (in all eight males), - X (one of three females), and + 7 in three patients. In our analysis of brain tumors, we

INTERPHASE CYTOGENETICS OF BRAIN TUMORS 105
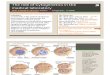
Figure I. lnterphase cytogenetics results. A Nuclei of an oligodendroglioma show two hybridization signals (FITC. yellow as a result of the filter combination) for chromosome I centromeres; red propidium iodide counterstaining. B: Nuclei of meningioma M I I display three hybridization signals (FITC) for chromosome I7 centromeres; red propidium iodide counterstaining. C: Nuclei of attrocytoma A4 show after double hybridization only one green signal for chromosome 10 centromere (FITC) but three red signals for chromosome 7 centromeres (TRITC); blue DAPl counterstaining, triple-exposure microphotoglaph. D: Nuclei of meningioma FI display after double hybridization two red signals for chromosome 7 centromeres (TRITC) but only one green signal for chromosome I8 centromere (FITC); blue DAPl counterstaining. triple-exposure microphotograph. E Hybridization with the cosmid probes for A61 and BCR on interphase nuclei of meningioma M7. Left: blue DAPl counterstaining; middle: two red ABL signals (TRITC) in each nucleus; right: the arrows indicate the single (small) green BCR signals (FITC) in each nucleus. F: Another interphase nucleus from meningioma M7 displaying two red A61 signals but only one green 6CR signal (FITC) after a double hybridization. indicating a loss of 22q; DAPl counterstaining, triple-exposure microphotograph.
found in seven gliomas from male patients only once a loss of the Y chromosome. This discrepancy might be caused by the fact that small mosaicisms, if present, would not be classified as such using our criteria for monosomy (more than 20% of nuclei
with one spot) and trisomy (more than 7.5% of nuclei with three spots). However, in a recent study with interphase cytogenetics on samples of normal brain tissue, we found - X, - Y or + 7 in an average of only 0.3%, 0.3%, and 0.5% of the

106 ARNOLDUS ET AL.
nuclei, respectively (Arnoldus et al., 1990a). These facts suggest that the numerical changes observed by Heim et al. are induced by the culture proce- dure and that they do not reflect an in vivo mosa- icism.
The majority of the meningiomas (62.5%) dis- played, as expected, only one signal per nucleus after hybridization with the cosmid probe for 22ql1, indicating a loss of 22q (Fig. lE,F). Some- times the nuclei with only one signal for 22qll (3946%) could be distinguished from the normal nuclei as a result of their slightly larger diameter. The three meningiomas with other changes (+ 17, - 18) did not show the different clinical behavior as suggested by A1 Saadi et al. (1987). There was no difference between the mean age of patients with chromosome anomalies (54.8 years) and the mean age of patients with normal findings (56.2 years).
The present study shows the feasibility of study- ing numerical chromosome aberrations in brain tu- mors by interphase cytogenetics using nonradioac- tive in situ hybridization. Recently it has been shown that interphase cytogenetics is also capable of detecting specific structural aberrations (Arnol- dus et al., 1990b). The number of meningiomas and gliomas studied is still too small to allow any generalizations about possible correlations between specific chromosome aberrations and the histopath- ological type and grade of the tumors. The results of the cosmid hybridizations in this study show that the sensitivity of the nonradioactive in situ hybrid- ization on clinical material is at the single copy gene level. When small probes of direct pathoge- netic significance become available, these too can be implemented, and interphase cytogenetics will help to narrow the gap between molecular genetics and classical cytogenetics in solid tumors.
ACKNOWLEDGMENTS
T h e authors thank F.C. De Beer, H. Van Dulken, R.T.W.M. Thomeer, and M.J.H.M. Herpers from the Department of Neurosurgery of the Academic Hospital Leiden for the supply of clinical material; G.C. Grosveld from the Depart- ment of Cell Biology and Genetics of the Erasmus University, Rotterdam, for the gift of the two cosmid probes; H.F. Willard for a number of sat- ellite DNA probes; and G.C. Beverstock from the Department of Human Genetics, Leiden Univer- sity, for critical review of the manuscript.
REFERENCES Al Saadi A, Latimer F, Madercic M, Robbins T (1987) Cytogenetic
studies of human brain tumors and their clinical significance. 11. Meningioma. Cancer Genet Cytogen 2 6 127-141.
Arnoldus EPJ, Noordermeer IA, Peters ACB, Raap AK, Van der Ploeg M (1990a) Interphase cytogenetics reveals somatic pairing of chromosome 17 centromeres in normal human brain tissue, but no trisomy 7 or sex-chromosome loss. Cytogenet Cell Genet (in press).
Arnoldus EPJ, Peters ACB, Bots GTAM, Raap AK, Van der Ploeg M (1989) Somatic pairing of chromosome 1 centromeres in in- terphase nuclei of human cerebellum. Hum Genet 83:231-234.
Arnoldus EPJ, Wiegant J, Noordermeer IA, Wessels JW, Bever- stock GC, Grosveld GC, Van der Ploeg M, Raap AK (1990b) Detection of the Philadelphia chromosome in interphase nuclei. Cytogenet Cell Genet 54: 108-1 11.
Bigner SH, Mark J, Bullard DE, Mahaley Jr MS, Bigner D D (1986) Chromosomal evolution in malignant human gliomas starts with specific and usually numerical deviations. Cancer Genet Cyto- genet 22:121-135.
Bigner SH, Mark J, Burger PC, Mahaley MS Jr, Bullard DE, Muhlbaier LH. Biener D D (1988) Specific chromosomal abnor- malities in malignant human gliomk. Cancer Res 48405-411.
Bigner SH, Mark J, Bigner D D (1990) Cytogenetics of human brain tumors. Cancer Genet Cytogenet 47: 141-154.
Casalone R, Granata P, Simi P, Tarantino E, Butti G, Buonaguidi R, Faggionato F, Knerich R, Solero L (1987) Recessive cancer genes in meningiomas? An analysis of 31 cases. Cancer Genet Cytogenet 27: 14S159.
Casalone R, Sirni P, Granata P, Minelli E, Giudici A, Butti G, Solero CL (1990) Correlations between cytogenetic and histo- pathological findings in 65 human meningiomas. Cancer Genet Cytogeiet 45:237-243.
Cooke HI. Hindlev J (1979) Cloning of human satellite I11 DNA: different components are on diff&ent chromsomes. Nucleic Ac- ids Res 6:3177-3197.
Cooke HJ, Schmidtke J, Gosden JR (1982) Characterization of a human Y chromosome repeated sequence and related sequences in higher primates. Chromosoma 87:491-502.
Cornelisse CJ, Tanke HJ (1990) Flow cytometry applied to cytopa- thology. In Bibbo M (ed): Comprehensive Cytopathology. Phil- adelphia: W.B. Saunders, in press.
Cremer T , Landegent J, Bruckner A, Scholl HP, Schardin M, Hager MD, Devilee P, Pearson P, Van der Ploeg M (1986) Detection of chromosome aberrations in the human interphase nucleus by visualization of specific target DNAs with radioactive and non-radioactive in situ hybridization techniques: Diagnosis of trisorny 18 with L1.84. Hum Genet 74:34&352.
Devilee P, Crerner T, Slagboom P, Bakker E, Scholl H, Hager H, Stevenson A, Cornelisse C, Pearson P (1986) Two subsets of human alphoid repetitive DNA show distinct preferential local- ization in the pericentric regions of chromosomes 13, 18 and 21. Cytogenet Cell Genet 41:193-202.
Devilee P, Kievits T, Waye JS, Pearson PL, Willard H F (1988a): Chromsome-specific alpha satellite DNA: Isolation and map- ping of a polymorphic alphoid repeat from human chromosome 10. Genomics 3: 1-7.
Devilee P, Thierry RF, Kievits T , Kolluri R, Hoprnan AHN, Willard HF, Pearson PL, Cornelisse CJ (1988b) Detection of chromosome aneuploidy in interphase nuclei from human pri- mary breast tumors using chromosome-specific repetitive DNA probes. Cancer Res 48:5825-5830.
Heim S, Mandabl N, Yin Y, Stromblad S, Lindstrom E, Salford LG, Mitelman F (1989) Trisomy 7 and sex chromosome loss in hu- man brain tissue. Cytogenet Cell Genet 52136-138.
Heisterkamp N, Groffen J, Stephenson JR (1983) T h e human v-061 cellular homologue. J Mol App Genet 2:57-68.
Hermans A, Selleri L , Gow J, Wiedemann L, Grosveld GC (1989) Molecular analysis of the Philadelphia translocation in Chronic myelogenous and acute lymphoblastic leukemia. Cancer Cells 7:21-26 (in Molecular Diagnostics of Human Cancer, from Cold Spring Harbor Laboratory).
Hopman AHN, Poddighe PJ, Smeets AWGB, Moesker 0, Beck JLM, Vooijs GP, Ramaekers FCS (1989) Detection of numerical chromosome aberrations in bladder cancer by in situ hybridiza- tion. Am J Pathol 135: 1105-1 117.
Hopman AHN, Ramaekers FCS, Raap AK, Beck JLM, Devilee P, Van der Ploeg M, Vooijs GP (1988) In situ hybridization as a tool to study numerical chromosome aberrations in solid bladder tu- mors. Histochemistry 89:307-3 16.
Jabs EW, Wolf SF, Migeon BR (1984) Characterization of a cloned DNA sequence that is present at centromeres of all human au-

INTERPHASE CYTOGENETICS OF BRAIN TUMORS 107
tosomes and the X chromosome and shows polymorphic varia- tion. Proc Natl Acad Sci USA 81:48844888.
Jenkins RB, Kimmel DW, Moertel CA, Schultz CG, Scheithauer BW, Kelly PJ, Dewald GW (1989) A cytogenetic study of 53 human gliomas. Cancer Genet Cytogenet 3932.53-279.
Maltby EL, Ironside JW, Battersby RD (1988) Cytogenetic studies in 50 meningiomas. Cancer Genet Cytogenet 31:199-210.
Mark J (1973) Karyotype patterns in human meningiomas. A com- parison between studies with G- and Q-banding techniques. Heriditas 75213-220.
Mendelsohn ML, Mayall BH, Bogart E, Moore DH 11, Perry BH (1973) DNA content and DNA-based centromeric index of the 24 human chromosomes. Science 179: 1126-1129.
Metzdorf R, Gottert E, Blin N (1988): A novel centromeric repet- itive DNA from human chromosome 22. Chromosoma 97:154- 158.
Nederlof PM, Van der Flier S, Raap AK, Tanke HJ, Van der Ploeg M, Kornips F, Geraedts JPM (1989) Detection of chromosome aberrations in interphase tumor nuclei by non-radioactive in situ hybridization. Cancer Genet Cytogenet 4287-98.
Raap AK, Dirks RW, Jiwa NM, Nederlof PM, Van der Ploeg M (1990) In situ hybridization with hapten-modified DNA probes. In Racz P, Haase AT, Gluckman JC (eds): Modern Pathology of AIDS and Other Retroviral Infections. Basel: Karger, pp 17-28.
Raap AK, Van der Ploeg M, Hopman AHN, Landegent JE , Van Duijn P (1987) Localization of DNA sequences by non- radioactive in situ hybridization. In Burger G, Ploem JS, Goertt- ler K (eds): Clinical cytometry and histometry. San Diego: Ac- ademic Press, pp 221-226.
Rey JA, Bello MJ, De Campos JM, Kusak ME, Moreno S (1987a) Chromosomal composition of a series of 22 human low-grade gliomas. Cancer Genet Cytogenet 29:223-237.
Rey JA, Bello MJ, De Campos JM, Kusak ME, Ramos C, Benitez J (1987b) Chromosomal patterns in human malignant astrocyto- mas. Cancer Genet Cytogenet 29:201-221.
Smit VTHBM, Wessels JW, Mollevanger P , Schrier PI, Raap AK, Beverstock GC, Cornelisse CJ (1990) Combined GTG-banding and non-radioactive in situ hybridization improves characteriza- tion of complex karyotypes. Cytogenet Cell Genet 54:ZO-23.
Teyssier JR (1989) T h e chromosomal analysis of human solid tu- mors: A triple challenge. Cancer Genet Cytogenet 37: 103-125.
Van Dekken H, Pizzolo JG, Kelsen DP, Melamed MR (1990) Tar- geted cytogenetic analysis of gastric tumors by in situ hybridiza- tion with a set of chromosome-specific DNA probes. Cancer 66:491-497.
Walt H, Emmerich P, Cremer T, Hofmann M-C, Bannwart F (1989) Supernumerary chromosome 1 in interphase nuclei of atypical germ cells in paraffin-embedded human seminiferous tubules. Lab Invest 61:527-531.
Waye JS, Creeper LA, Willard H F (1987a) Organization and evo- lution of alpha satellite DNA from human chromosome 1 1 . Chromosoma 95: 182-188.
Waye JS, England SB, Willard H F (1987b) Genomic organization of alpha satellite DNA on human chromosome 7: Evidence for two distinct alphoid domains on a single chromosome. Mol Cell Biol 7:349-356.
Waye JS, Willard H F (1986) Molecular analysis of a deletion poly- morphism in alpha satellite of human chromosome 17: Evidence for homologous unequal crossing-over and subsequent fixation. Nucleic Acids Res 14:6915-6927.
Willard HF, Smith KD, Sutherland J (1983) Isolation and charac- terization of a major tandem repeat family from the human X chromosome. Nucleic Acids Res 11:2017-2033.
Zang KD (1982) Cytological and cytogenetical studies on human meningioma. Cancer Genet Cytogenet 27: 145-159.
Zang KD, Singer H (1967) Chromosomal constitution of meningi- omas. Nature 216:84-85.
Ziilch KJ (1979) Histological classification of the tumors of the C.N.S. International classification of tumors, vol 21. Geneva: World Health Organization.