Embed Size (px)
Citation preview

5
II. TINJAUAN PUSTAKA
Botani Tanaman Mangga
Secara taksonomis, mangga termasuk dalam Famili Anarcadiaceae, Ordo
Sapindales, Genus Mangifera, dan Spesies Mangifera indica. Famili
Anarcadiaceae (mangga-manggaan) terdiri dari sekitar 500 spesies, sedangkan
genus Mangifera meliputi 62 spesies (Pracaya, 2011). Arifin (2009) menyatakan
bahwa tanaman mangga berkerabat dekat dengan pakel (M. foetida), kweni (M.
odorata), dan kemang (M. caesia).
Menurut Pracaya (2011), tanaman mangga berasal dari negara India dan
menyebar ke wilayah Asia Tenggara pada abad ke-4 dan ke-5 Sebelum Masehi.
Penanaman mangga dimulai di Filipina dan Indonesia (sekitar Maluku) pada
tahun 1600-an. Bangsa Portugis menyebarkan tanaman mangga ke Barat pada
abad ke-18 dan Afrika pada abad ke-19. Keberadaan mangga di Meksiko
dilaporkan pada tahun 1779. kemudian mulai ditanam di Florida, Amerika Serikat
(1833), Queensland, Australia (1870), dan Italia bagian selatan (1905).
Tanaman mangga tumbuh tegak, bercabang banyak, dan bertajuk
rindang serta hijau sepanjang tahun. Tinggi tanaman dapat mencapai 10-40
meter dan berumur lebih dari seratus tahun. Buah mangga tergolong buah
berdaging dengan bentuk beragam sesuai dengan varietas. Warna buah hijau,
kuning, merah atau campuran. Ujung buah melancip ataupun membengkok.
Daging buah tebal atau tipis, berserat atau tidak, serta berair ataupun tidak
(Pracaya 2011).
Setiap varietas mangga mempunyai karakteristik yang berbeda.
Contohnya perbandingan buah mangga gedong gincu dan arumanis. Bobot buah
mangga arumanis biasanya lebih besar dibandingkan mangga gedong gincu.
Namun demikian, aroma mangga gedong gincu lebih harum menyengat
dibandingkan dengan mangga arumanis. Pangkal buah mangga gedong gincu
berwarna merah keunguan pada saat matang, sedangkan mangga arumanis
berwarna hijau kekuningan. Perbedaan lain yang terlihat adalah bentuk buah.
Mangga gedong gincu berbentuk bulat. Mangga arumanis berbentuk jorong
dengan pucuk meruncing.
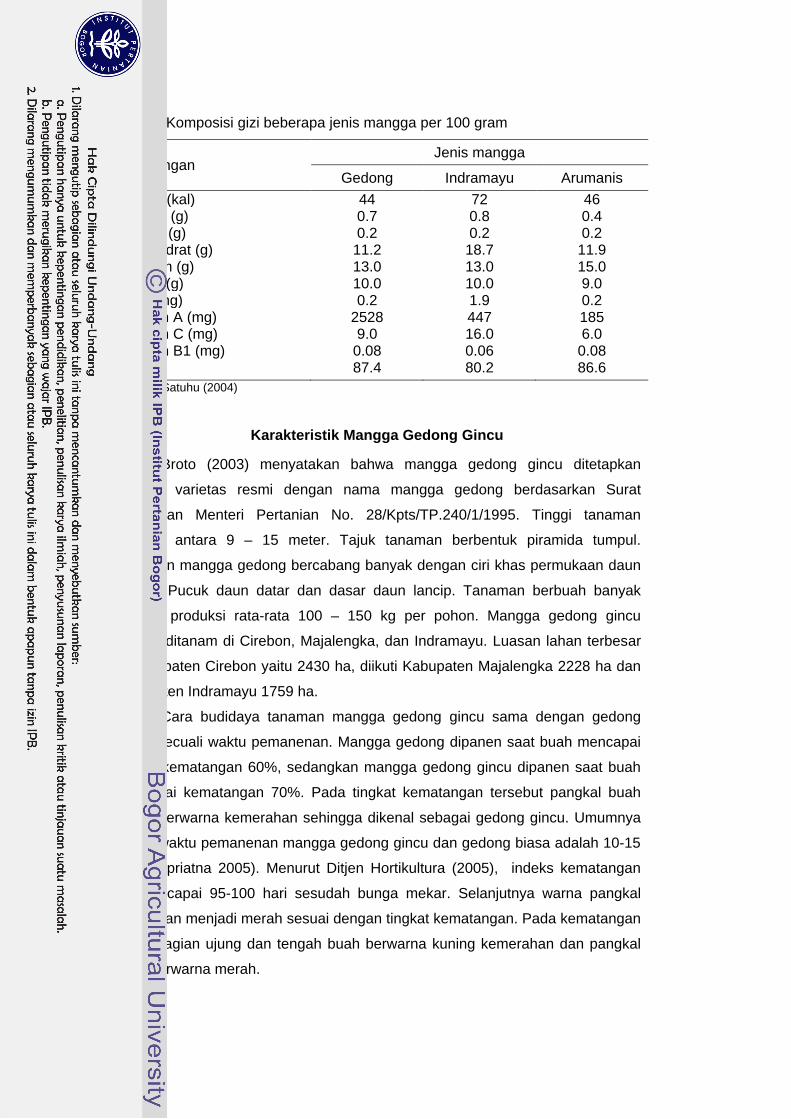
Karakteristik fisikokimia dan kandungan nutrisi buah mangga berbeda
untuk masing-masing varietas. Tabel 1 menunjukkan perbandingan kandungan
nutrisi antara mangga gedong, arumanis, dan Indramayu.
6
Tabel 1 Komposisi gizi beberapa jenis mangga per 100 gram
Kandungan Jenis mangga
Gedong Indramayu Arumanis Energi (kal) 44 72 46 Protein (g) 0.7 0.8 0.4 Lemak (g) 0.2 0.2 0.2 Kabohidrat (g) 11.2 18.7 11.9 Kalsium (g) 13.0 13.0 15.0 Fosfor (g) 10.0 10.0 9.0 Besi (mg) 0.2 1.9 0.2 Vitamin A (mg) 2528 447 185 Vitamin C (mg) 9.0 16.0 6.0 Vitamin B1 (mg) 0.08 0.06 0.08 Air (g) 87.4 80.2 86.6
Sumber: Satuhu (2004)
Karakteristik Mangga Gedong Gincu
Broto (2003) menyatakan bahwa mangga gedong gincu ditetapkan
sebagai varietas resmi dengan nama mangga gedong berdasarkan Surat
Keputusan Menteri Pertanian No. 28/Kpts/TP.240/1/1995. Tinggi tanaman
berkisar antara 9 – 15 meter. Tajuk tanaman berbentuk piramida tumpul.
Tanaman mangga gedong bercabang banyak dengan ciri khas permukaan daun
sempit. Pucuk daun datar dan dasar daun lancip. Tanaman berbuah banyak
dengan produksi rata-rata 100 – 150 kg per pohon. Mangga gedong gincu
banyak ditanam di Cirebon, Majalengka, dan Indramayu. Luasan lahan terbesar
di Kabupaten Cirebon yaitu 2430 ha, diikuti Kabupaten Majalengka 2228 ha dan
Kabupaten Indramayu 1759 ha.
Cara budidaya tanaman mangga gedong gincu sama dengan gedong
biasa, kecuali waktu pemanenan. Mangga gedong dipanen saat buah mencapai
tingkat kematangan 60%, sedangkan mangga gedong gincu dipanen saat buah
mencapai kematangan 70%. Pada tingkat kematangan tersebut pangkal buah
sudah berwarna kemerahan sehingga dikenal sebagai gedong gincu. Umumnya
selisih waktu pemanenan mangga gedong gincu dan gedong biasa adalah 10-15
hari (Supriatna 2005). Menurut Ditjen Hortikultura (2005), indeks kematangan
70% tercapai 95-100 hari sesudah bunga mekar. Selanjutnya warna pangkal
buah akan menjadi merah sesuai dengan tingkat kematangan. Pada kematangan
100% bagian ujung dan tengah buah berwarna kuning kemerahan dan pangkal
buah berwarna merah.

7
Mangga varietas gedong gincu mempunyai karakteristik yang menarik
bagi konsumen. Menurut Pracaya (2007), buah mangga gedong gincu memiliki
warna daging merah kekuningan. Bentuk buah gedong agak bulat dengan
pangkal agak datar. Tangkai buah kuat dan terletak di tengah buah. Bobot buah
200 – 300 gram. Ukuran buah 8 cm x 7 cm x 6 cm. Permukaan kulit buah halus
dan kadangkala berbintik putih kehijauan. Daging buah tebal, berserat halus,
manis, berair, dan beraroma keras. Karakteristik aroma harum menyengat
menjadi keunggulan mangga gedong gincu dibandingkan dengan mangga lokal
lain di Indonesia. Konsumen luar negeri juga tertarik dengan warna kulit buah
mangga yang berwarna merah menyala saat matang.
Gambar 1 Buah mangga gedong gincu pada, (a) indeks kematangan awal (70%), (b) indeks kematangan 80-85%.
Hama Tanaman Mangga
Beragam organisme pengganggu golongan hama dilaporkan menyerang
tanaman mangga di lapangan. Tandon et al. (1976 dalam Kumar 2009)
menyatakan terdapat 492 spesies serangga yang menyerang tanaman mangga.
Hama utama mangga di lapangan meliputi Sternochetus mangiferae, Noorda
albizonalis, dan beberapa spesies lalat buah (Panhwar 2005). Menurut
Kalshoven (1981), terdapat 13 genus serangga yang menyerang tanaman
mangga di Indonesia, diantaranya: Rastrococcus, Sternochetus, Noorda,
Philotroctis, Rhytidodera, dan Orthaga.
Sternochetus mangiferae merupakan hama penting mangga yang
termasuk dalam Famili Curculionidae. Serangga ini termasuk dalam organisme
pengganggu tumbuhan karantina untuk pemasukan mangga dari Hawaii ke
Amerika Serikat daratan (Follett dan Gabbard 2000). Stadia dewasa berukuran
tubuh 10 mm, berwarna hitam abu-abu, dan mempunyai ciri moncong (snout)
yang jelas. Telur diletakkan serangga betina di permukaan buah. Induk
memanfaatkan sap yang keluar dari jaringan buah sebagai perekat telur di
a b

8
permukaan buah. Setelah menetas, larva masuk ke biji buah dan melakukan
aktivitas makan hingga masa pupasi. Serangga dewasa yang muncul memakan
lapisan biji buah. Pengendalian dilakukan dengan aplikasi insektisida sebelum
pembungaan ataupun memusnahkan buah bergejala yang jatuh ke tanah (Chin
et al. 2010).
Philotroctis eutraphera merupakan hama penggerek buah mangga,
terutama buah muda. Larva berwarna kemerahan dan berkembang menjadi biru
gelap saat menjelang pupasi. Pupa dalam kokon dapat dijumpai di tanah.
Ngengat berumur 6 – 7 hari dan betina menghasilkan telur sebanyak 125 – 450
telur di permukaan dan tangkai buah (Kalshoven 1981).
Autocharis albizonalis atau Noorda albizonalis yang merupakan hama
utama mangga di India. Serangga melewati lima tahap instar larva selama 11-13
hari. Tingkat keparahan tertinggi diakibatkan oleh serangan oleh larva instar
kedua atau ketiga. Kerusakan akibat serangan N. albizonalis dilaporkan
mencapai 10 – 52% di Benggal Barat, India (Sahoo dan Jha 2009). Di Indonesia,
N. albizonalis menyerang berbagai stadia perkembangan buah mangga
(Kalshoven 1981).
Wereng daun merupakan hama penting yang menyerang daun tanaman
mangga. Serangga dewasa dan nimfa memakan jaringan tanaman dengan cara
menghisap sap tanaman. Daun terserang menunjukkan gejala distorsi dan
mengeriting. Salah satu spesies wereng daun pada pertanaman mangga di
Australia adalah Idioscopus nitidulus (Chin et al. 2010), sedangkan di Indonesia
banyak dijumpai I. niveoparsus dan I. clypealis (Kalshoven 1981).
Kutu putih yang menyerang daun mangga adalah Rastrococcus spinosus.
Salah satu ciri khas R. Spinosus adalah rambut berlilin panjang di seluruh
tubuhnya. Betina berbentuk pipih, oval, lebar, dan dilapisi lapisan lilin tebal. Kutu
putih ini telah dilaporkan di Jawa, Malaysia, Filpina, dan Taiwan. Selain mangga,
spesies ini banyak ditemukan di permukaan bawah daun pada jeruk dan kopi.
Kutu putih mendapatkan makanan dengan menghisap sap jaringan tanaman
(Kalshoven 1981).
Orthaga euadrusalis merupakan hama yang memakan daun tanaman
mangga. Salah satu ciri khas serangannya adalah keberadaan jaring yang
menutup daun dan ranting. Larva berwarna ungu dengan garis lateral hitam.
Larva O. euadrusalis mengakibatkan kerusakan serius pada daun dan tunas
(Kalshoven 1981).

9
Salah satu hama penggerek batang mangga adalah Rhytidodera
simulans. Menurut Kalshoven (1981), serangga ini telah menyebar luas di Asia
Tenggara kecuali daerah Nusa Tenggara. Gejala serangan berupa lubang
menyerupai terowongan pada cabang yang terserang. Apabila cabang telah
terbuka dan terbelah menjadi terowongan yang lebih besar maka umumnya akan
ditempati semut. R. simulans juga diketahui menyerang M. odorata, M. foetida,
dan Elaeocarpus grandifloris. Selain hama-hama di atas, hama penting lain yang
menjadi musuh petani mangga di seluruh dunia adalah lalat buah (Subbab 2.4).
Lalat Buah sebagai Hama
Biologi dan Morfologi Lalat buah termasuk ke dalam Filum Arthropoda, Klas Insekta, Ordo
Diptera, Subordo Brachycera, dan Famili Tephritidae. Lalat buah mengalami
metamorfosis sempurna dengan melalui stadia telur, larva, pupa, dan imago.
Telur diletakkan secara berkelompok dan dalam waktu 2 hari akan menetas.
Menurut Vijaysegaran dan Drew (2006), satu ekor betina B. dorsalis dapat
menghasilkan 1200 – 1500 butir telur. Larva lalat buah mengalami tiga tahap
perkembangan instar. Larva bernafas dengan spirakel yang terdapat di bagian
anterior dan posterior. Khusus larva instar pertama, anterior spirakel belum
menunjukkan perkembangan. Lama stadia larva adalah 6 – 9 hari. Setelah itu
larva akan berkembang menjadi pupa berbentuk oval dan berwarna cokelat.
Pupa umumnya berukuran sekitar 5 mm dan berlangsung sekitar 10 hari. Setelah
itu stadia dewasa mudah dikenali karena mempunyai sayap dengan pola unik
dan bervariasi.
Stadia hidup lalat buah yang berpotensi terbawa dalam lalu lintas buah
mangga adalah telur dan larva. Telur lalat buah umumnya berwarna putih atau
krem kekuningan. Warna telur semakin gelap seiring umur telur semakin tua.
Ukuran dan bentuk telur bemacam-macam sesuai jenis spesies. Ceratitis
capitata dan B. tryoni mempunyai telur memanjang dan menyempit secara
bertahap, sedangkan Urophora solsitialis mempunyai telur dengan ujung
membulat dan meruncing pada ujung lainnya (White dan Elson-Harris 1994).
Tubuh larva terdiri dari bagian kepala, thoraks, dan abdomen. Thoraks
meliputi 3 segmen, yaitu prothoraks, mesothoraks, dan metathoraks, sedangkan
abdomen terdiri dari 8 segmen. Batas antara kepala dan thoraks tidak tampak
jelas. Kepala tidak mengalami sklerotisasi. Bagian tubuh larva yang mengalami

10
pengerasan adalah cephalopharyngeal skeleton dimana terdapat mulut kait
berwarna hitam atau cokelat (JFTA 1996). Keberadaan larva di dalam buah
merupakan penyebab utama penolakan buah ekspor di negara tujuan (Frias et
al. 2006).
Bentuk dan ukuran larva bervariasi sesuai dengan spesies dan nutrisi
dalam makanannya. Sebagian larva cenderung berbentuk silindris dan membulat
ataupun menyerupai bentuk terpotong pada kedua ujung tubuh. Larva berwarna
krem keputihan, namun dapat juga berwarna lebih gelap sesuai dengan jenis
makanannya (White dan Elson-Harris 1994).
Gambar 2 Perkembangan hidup lalat buah: (a) telur, (b) larva, (c) pupa, (d) imago
Larva instar pertama berukuran sangat kecil. Cephalopharyngeal skeleton
masih kurang tersklerotisasi. Mulut kait berwarna kuning atau lebih gelap. Mulut
kait mempunyai 1 atau lebih preapical teeth berukuran besar. Anterior spirakel
tampak seperti pori yang hanya dapat dilihat di bawah mikroskop elektron. Pada
posterior spirakel hanya terdapat 2 spiracular slit (JFTA 1996; White dan Elson-
Harris 1994).
Larva instar kedua mempunyai karakteristik hampir sama dengan larva
instar ketiga namun berukuran lebih kecil. Larva berwarna krem keputihan atau
sesuai makanan dalam saluran pencernaanya. Cephalopharyngeal skeleton
menyerupai instar ketiga namun terdapat 1 atau lebih preapical teeth. Anterior
dan posterior spirakel telah berkembang dengan baik. Posterior spirakel terdiri
a b
c d

11
dari 3 buah spiracular slit dan dikelilingi oleh rimae yang tersklerotisasi. Terdapat
4 rumpun spiracular hair namun jumlah rambut lebih sedikit dibanding larva instar
ketiga (JFTA 1996).
Larva instar ketiga merupakan instar dengan periode hidup paling lama di
dalam inang. Pada instar tersebut, larva mempunyai sepasang anterior spirakel
pada bagian prothoraks dan sepasang posterior spirakel di caudal segment.
Posterior spirakel dikelilingi oleh banyak spiracular tubules. Pada sisi ventral
caudal segment terdapat anus. Larva instar ketiga mempunyai kemampuan
melenting. Umumnya hal ini dilakukan saat larva akan mengalami pupasi (JFTA
1996).
Dampak Serangan Lalat buah merupakan masalah utama petani buah di dunia. Dampak
serangannya dirasakan dalam pemeliharaan tanaman di lapang maupun dalam
upaya ekspor ke negara lain. Kerusakan buah dimulai saat lalat buah betina
meletakkan telur di dalam jaringan inang. Larva menyebabkan kerusakan buah
secara cepat. Selain itu, kulit buah yang luka dapat menjadi tempat masuk
bakteri pembusuk. Pada aspek perdagangan, keberadaan lalat buah
menimbulkan kesulitan suatu negara untuk memasarkan produk buah segar ke
negara lain (Drew 2001).
Serangan lalat buah menimbulkan kehilangan hasil yang bervariasi. Lalat
buah dilaporkan menimbulkan kerusakan mangga hingga kisaran 10 – 50% di
Benin (Vayssieres et al. 2005). Dhillon et al. (2005) menyatakan kerusakan B.
cucurbitae pada tanaman cucurbit mencapai 100% dan pare sebesar 95%. B.
dorsalis merupakan hama penting pada tanaman mangga ‘Namdokmai Si Thong’
di Thailand (Varith et al. 2006) dan mengakibatkan kerusakan serius pada jambu
(Psidium guajava) dan jambu stroberi (Psidium cattleianum) di Hawaii (Vargas et
al. 2007). Beberapa informasi kehilangan hasil tanaman akibat serangan lalat
buah di kawasan Asia Pasifik dapat dilihat pada Tabel 2.
Pencegahan penyebaran lalat buah melalui lalulintas perdagangan
komoditas pertanian telah dilakukan oleh banyak negara. Hal ini dilakukan
karena lalat buah mampu hidup dan berkembang dengan cepat di daerah baru di
luar sebaran asalnya (Armstrong et al. 2009). Bahkan Siwi et al. (2006)
menyatakan bahwa lalat buah eksotik yang telah masuk ke daerah baru dan
berhasil berkolonisasi mempunyai daya rusak lebih tinggi dibanding lalat buah

12
lokal. Salah satu contoh adalah kasus masuknya 8 spesies baru lalat buah di
California yang mengakibatkan kehilangan hasil sebesar 910 juta dollar AS.
Biaya pengendalian akibat masuknya lalat buah eksotik sangat tinggi.
Drew (2001) menyatakan program eradikasi B. papayae di North Queensland
menelan biaya sebesar 35 juta dollar AS. Eradikasi B. dorsalis dengan male
annihilation method di Kepulauan Okinawa, Miyako, dan Yaeyama Jepang juga
dilaporkan sangat tinggi, yaitu mencapai 2.575 milyar Yen (OPPPC 2006). Biaya
tinggi juga diperlukan untuk pengembangan teknik serangga mandul. Teknik ini
telah berhasil dikembangkan untuk pengendalian B. dorsalis di Ogasawara Island
dan Kume Island (Jepang), serta C. capitata di Hawaii, California, Meksiko,
Nikaragua, Kostarika, Peru, Italia, Spanyol, dan Tunisia (White dan Elson-Harris
1992).
Tabel 2 Kehilangan hasil tanaman akibat serangan lalat buah pada beberapa
negara Asia Pasifik
Negara Tanaman Kehilangan Hasil Lalat Buah Penyebab
Malaysia Belimbing 100% B. carambolae Thailand Mangga 65% B. dorsalis Cantaloupe 94% B. cucurbitae Vietnam Persik 65% B. pyrifoliae Jambu air 76% B. correcta Palau Pisang 80% B. dorsalis complex Nauru Mangga 95% B. dorsalis / B. frauenfeldi Vanuatu Jambu 95% B. trilineola
Sumber: Drew (2001).
Lalat buah mampu berkembang di daerah baru dan menghasilkan
kerusakan. Beberapa kasus diantaranya adalah B. philippinensis yang menyebar
ke Palau (Vueti dan Leblanc 2002), B. tryoni ke Papua New Guinea (Purea et al.
1997), B. carambolae ke Suriname (Sauers-Mullers 2005), dan B. cucurbitae ke
beberapa negara kepulauan di Samudera Hindia (Vayssieres et al. 2008). Lalat
buah Anastrepha juga mulai menginvasi daerah Amerika Utara dari Amerika
Selatan. Salah satunya A. suspensa yang menyebar ke Florida, Amerika Serikat
(Weems et al. 2001). De Meyer et al. (2009) melaporkan bahwa beberapa
negara Afrika di kawasan tropis telah terintroduksi B. invadens yang awalnya
hanya ditemukan di Kenya.
Peningkatan lalulintas perdagangan membuka peluang penyebaran lalat
buah ke negara lain. Saat ini Amerika Serikat menghadapi ancaman masuknya

13
lalat buah Anastrepha dari kawasan Amerika Selatan dan Bactrocera dari
kawasan Asia. Pada tahun 1999 – 2009 beberapa spesies lalat buah eksotik
ditemukan di Florida dan California, diantaranya B. correcta, B. dorsalis, B.
latifrons, B. oleae, dan B. zonata. Jepang juga waspada terhadap penyebaran
lalat buah melalui komoditas pertanian impor. Iwaizumi (2004 dalam Ebina dan
Ohto 2006) melaporkan bahwa otoritas karantina Jepang berhasil mengintersepsi
beberapa spesies lalat buah pada komoditas pertanian impor, meliputi B.
dorsalis, B. carambolae, B. papayae, B. occipitalis, dan B. philippinensis.
Bactrocera dorsalis complex
Bactrocera merupakan genus lalat buah dari daerah sekitar khatulistiwa.
Sebagian besar lalat buah berasosiasi dengan buah-buahan tropis. Menurut
Hardy (1977 dalam Siwi et al. 2006), terdapat 160 genus dalam Tephritidae dan
180 spesies Bactrocera di kawasan Asia. Dari jumlah tersebut terdapat beberapa
spesies yang mempunyai kemiripan morfologis yang dikenal dengan Bactrocera
dorsalis complex.
Spesies dalam kelompok B. dorsalis complex dibedakan berdasarkan
karakteristik tertentu, seperti panjang aculeus dan pola warna di bagian thoraks.
Menurut Drew (2004), hasil survei di kawasan Asia Tenggara, Papua New
Guinea, Australia, dan Pasifik Selatan berhasil mendeskripsikan 80 spesies
dalam B. dorsalis complex. Namun demikian, Drew dan Hancock (1994)
menyatakan bahwa hanya sebagian kecil spesies dalam B. dorsalis complex
yang merugikan secara ekonomi. Beberapa spesies penting dalam B. dorsalis
complex, meliputi: B. carambolae, B. caryae, B. dorsalis, B. kandiensis, B.
occipitalis, B. papayae, B. philippinensis, dan B. pyrifoliae.
Sebagian besar spesies B. dorsalis complex mempunyai daerah sebaran
yang terbatas. Namun demikian, B. dorsalis, B. carambolae, B. papayae, dan B.
philippinensis dikenal sebagai jenis lalat buah yang mampu menyebar dan
menjadi masalah di tempat yang baru. B. dorsalis ditemukan di Hawaii pada
tahun 1946 dan menjadi hama penting pada berbagai buah. B. carambolae
masuk ke Suriname pada tahun 1975, sedangkan B. philippinensis telah
menyebar hingga Palau di Pasifik. Oleh karena itu, upaya eradikasi telah
dilakukan untuk mengendalikan lalat buah pendatang dari area lain. Beberapa
eradikasi yang berhasil adalah B. dorsalis di Okinawa, Guam, dan Kepulauan
Mariana (Drew dan Hancock 1994).

14
Bactrocera carambolae. Spesies ini merupakan salah satu spesies lalat
buah yang berpotensi sebagai hama potensial tanaman buah dan hortikultura di
Indonesia (Soesilohadi et al. 2003). Spesies lalat buah ini menyerang berbagai
macam buah-buahan di daerah tropis dan temperate hangat, terutama pepaya
(Carica papaya), mangga (Mangifera indica), dan belimbing (Averrhoa
carambola). Tanaman inang lainnya meliputi: kluwih (Artocarpus altilis), jambu air
(Syzgium jambos dan S. aqueum), jambu bol (S. malaccense), cabai (Capsicum
annuum), jambu biji (Psidium guajava), tomat (Lycopersicon esculentum),
nangka (Artocarpus heterophyllus), Terminalia setappa, Solanum ferox, dan
Lepisanthes fruticosa (Siwi et al. 2006). B. carambolae termasuk dalam kelompok B. dorsalis complex dan
sebelumnya sempat disebut Bactrocera sp. near B. dorsalis. Ciri morfologis
sayap adalah pita hitam pada garis costa dan garis anal (anal streak). Pola sayap
bagian ujung berbentuk seperti pancing. Skutum berwarna hitam dengan pita
kuning di kedua sisi lateral (lateral post sutural vittae). Postpronotal berwarna
kuning atau oranye. Pada lalat betina terdapat spot berwarna hitam pada femur
tungkai depan, sedangkan abdomen berwarna coklat oranye dengan pola-pola
yang jelas (Siwi et al. 2006).
Lalat buah betina meletakkan telur di bawah kulit buah. Kemudian larva
yang muncul dari telur melakukan aktivitas makan dari dalam buah. Lubang yang
dibuat oleh larva dalam jaringan daging buah juga merangsang masuknya
cendawan dan bakteri patogen tanaman. Buah yang diserang B. carambolae
menunjukkan gejala perubahan warna kulit di sekitar tanda sengatan. Selain itu,
buah juga dapat mengalami pembusukan secara cepat. Larva hidup dalam buah
sampai instar akhir, kemudian melenting ke tanah untuk pupasi (Siwi et al. 2006).
Bactrocera papayae. Spesies ini dikenal juga sebagai B. conformis. B.
papayae merupakan spesies yang berkembang luas di kawasan Asia Tenggara,
terutama: Thailand, Malaysia, Singapura, dan Indonesia (Siwi et al. 2006). B.
papayae merupakan polifag yang menyerang banyak jenis buah dan sayuran.
Inang utama meliputi beberapa tanaman buah tropis, seperti: pisang (Musa x
parasidiaca), mangga (Mangifera indica), pepaya (Carica papaya), dan rambutan
(Nephelium lappaceum). B. papayae diketahui telah menyerang mangga
‘Kensington’ di Australia (Jacobi dan Giles 1997). Toraks B. papayae mempunyai ciri berwarna hitam dominan pada skutum
dan mempunyai rambut supra alar di sisi anterior. Sebagaimana B. carambolae,

15
skutum ditandai dengan pita berwarna kuning di sisi lateral (lateral postsutural
vittae). Sayap B.papayae dicirikan dengan pita hitam pada garis costa dan garis
anal, serta sel bc tampak jelas. Sedangkan abdomen terbagi dalam ruas-ruas
yang jelas dimana terdapat pekten pada tergit ketiga. Selain itu, tergit ketiga juga
dicirikan dengan garis melintang. Lalat betina dewasa tidak mempunyai spot
hitam femur tungkai depan sebagaimana dijumpai pada B. carambolae dewasa
(Siwi et al. 2006).
Perlakuan Karantina Tumbuhan
Aturan karantina diperlukan untuk pencegahan penyebaran suatu
organisme pengganggu tumbuhan. Salah satunya dengan perlakuan terhadap
komoditas yang akan dikirim (Jang dan Moffit 1994). Perlakuan karantina
bertujuan untuk membunuh, membuang, ataupun mencegah perkembangbiakan
organisme tertentu yang tidak dikehendaki pada komoditas pertanian. Perlakuan
yang dikembangkan diharapkan mampu mengakibatkan mortalitas yang tinggi
pada serangga target. Umumnya yang dipersyaratkan adalah probit 9 atau
mortalitas mencapai 99.9968% (Mangan dan Hallmann 1998).
Perlakuan karantina tumbuhan terdiri dari perlakuan kimiawi dan fisik.
Teknik perlakuan kimiawi yang umum digunakan adalah fumigasi. Pada tahun
1980-an etilen dibromida (EDB) merupakan fumigan paling populer, namun
kemudian ditinggalkan karena bersifat karsinogenik (Mangan dan Hallmann
1998). Metil bromida menjadi fumigan paling populer berikutnya karena daya
kerja cepat dan berspektrum luas (Fields dan White 2002). Fumigan ini juga
efektif untuk nematoda dan patogen tumbuhan. Namun sesuai Protokol Montreal
tahun 1997, metil bromida dilarang digunakan karena menyebabkan kerusakan
ozon. Loaharanu (1999) menyatakan bahwa negara-negara maju telah
menghentikan penggunaan metil bromida sejak 2005, sedangkan negara
berkembang dijadualkan tahun 2015. Penggunaan metil bromida untuk
selanjutnya hanya diperbolehkan untuk keperluan karantina dan tindakan darurat
tertentu.
Alternatif pengganti metil bromida mulai dikembangkan sebagai tindakan
karantina, baik kimiawi maupun fisik. Menurut Fields dan White (2002), bahan
aktif sebagai pengganti metil bromida adalah fosfin. Fosfin merupakan fumigan
yang dapat digunakan untuk komoditas yang tidak direkomendasikan difumigasi
dengan metil bromida. Komoditas tersebut meliputi benih, produk makanan, serta

16
biji-bijian yang mengandung lemak dan protein tinggi. Barantan (2007)
menyatakan bahwa fosfin relatif tidak menimbulkan residu pada komoditas.
Bahan kimia lain yang sudah dikembangkan adalah sulfuryl fluoride untuk
pengendalian hama kayu (Yu et al. 2010) dan carbonyl sulfide (Fields dan White
2002). Perlakuan non-kimiawi yang dikembangkan sebagai perlakuan karantina
meliputi vacuum-steam-vacuum (Fuester et al. 2004), irradiasi (Mitcham 1999),
atmosfer terkendali (Hallmann 1994), radio frequency (Tang et al. 2000),
perlakuan dingin (Gould dan Hennesey 1997; Wilink et al. 2006) dan perlakuan
panas (Shellie dan Mangan 2000; Shellie dan Mangan 1994; Neven 2000; Neven
1998).
Perlakuan temperatur dingin merupakan teknik populer sebelum digeser
fumigasi. Perlakuan digunakan terhadap buah yang toleran terhadap temperatur
kurang dari 2oC, seperti belimbing, jeruk, leci, manggis, apel, dan pir. Awalnya
perlakuan ini digunakan untuk disinfestasi C. capitata pada apel pada tahun
1907. Selanjutnya perlakuan dingin telah banyak dikaji untuk beberapa telur
serangga gudang, seperti Ephestia cautella, Sitotroga cerealella, Tribolium
confusum, Plodia interpunctella, dan Oryzaephilus surinamensis. Perlakuan juga
digunakan untuk membunuh stadia telur dan larva Grapholita molesta dan Cydia
pomonella. Dalam pengendalian lalat buah, perlakuan dingin 1.1oC selama 20,
12 atau 18 hari digunakan untuk Anastrepha ludens, C. capitata, dan B. tryoni
(Armstrong 1994).
Perlakuan karantina dengan temperatur tinggi telah dikembangkan pada
banyak komoditas. Awalnya panas digunakan untuk mengendalikan serangga
gudang. Mahroof (2007) menyatakan bahwa panas untuk mengendalikan hama
pertamakali dilakukan oleh Duhamel du Monceau dan Tillet pada tahun 1762 di
Perancis bagian barat. Pada masa itu temperatur 69oC selama 3 hari digunakan
untuk mengendalikan Sitotroga cerealella pada komoditas biji-bijian. Pada tahun
1835 perlakuan panas digunakan untuk Sitophilus oryzae pada gandum di
Amerika Serikat (Fields dan White 2002). Perlakuan panas sebagai tindakan
karantina dilakukan pertamakali tahun 1929 untuk mencegah penyebaran C.
capitata dari Florida. Selanjutnya tahun 1930-an digunakan untuk mendisinfestasi
lalat buah yang menyerang jeruk, alpukat, mangga, dan jambu. Saat ini metode
perlakuan panas untuk persyaratan karantina tumbuhan negara tujuan meliputi
hot water treatment, force-air treatment atau hot air treatment, dan vapor heat
treatment (Shellie dan Mangan 2000; Follett 2004).

17
Metode hot water treatment (HWT) merupakan metode paling tua
diantara semua metode perlakuan panas. Prinsipnya mencelupkan komoditas ke
dalam air panas pada temperatur dan waktu tertentu. HWT merupakan salah
satu persyaratan perlakuan untuk buah-buahan yang masuk ke Amerika Serikat,
seperti buah mangga, leci, longan, dan jeruk lemon (APHIS 2011). Metode ini
terutama digunakan untuk mencegah penyebaran C. capitata dan Anastrepha
spp. dari Amerika Tengah, Karibia, dan Amerika Selatan (Sharp 1988). Selain
untuk pengendalian hama, metode HWT juga dapat digunakan untuk
mengendalikan penyakit pascapanen. Perendaman mangga pada air temperatur
53oC selama 3 menit dilaporkan mampu mengendalikan penyakit antraknose
(Klein dan Lurie 1992). Namun demikian metode HWT tidak cocok untuk buah-
buah tertentu karena dapat merusak buah. Drake et al. (2005) menyebutkan
bahwa metode air panas 48 – 55oC hingga 14 menit tidak cocok digunakan untuk
buah ceri varietas Bing dan Sweetheart.
Perlakuan udara panas merupakan alternatif lain perlakuan karantina.
Prinsipnya pemanasan komoditas dengan udara pada temperatur 40-50oC
selama waktu tertentu untuk mengendalikan lalat buah. Perlakuan udara panas
dapat digunakan untuk mangga, anggur, jeruk, belimbing, dan pepaya. APHIS
(2011) mensyaratkan perlakuan ini terhadap jeruk impor asal Meksiko, Hawaii,
dan daerah Amerika Serikat yang tidak bebas lalat buah tertentu. Perlakuan jeruk
dari Meksiko untuk mendisinfestasi Anastrepha spp. adalah udara panas 44oC
selama 100 menit dengan waktu kondisioning 90 menit. Sedangkan jeruk asal
Hawaii dengan lalat buah target C. capitata, B. dorsalis, dan B. cucurbitae harus
diberi perlakuan 47.2oC selama 5 menit dengan waktu kondisioning 4 jam.
Perlakuan udara panas juga mampu menekan penyakit pada buah. Shellie dan
Skaria (1998) melaporkan bahwa perlakuan udara panas pada 46o
Perlakuan uap panas atau Vapor Heat Treatment (VHT) merupakan
metode disinfestasi OPT dengan uap basah pada temperatur 43 – 50
C selama 300
menit dapat menghambat pertumbuhan Penicillium digitatum.
Perlakuan Uap Panas
oC.
Armstrong (1994) menyebutkan bahwa panas ditransfer dari udara ke buah
dengan kondensasi uap air pada permukaan buah. Pada VHT, peningkatan
temperatur secara perlahan lebih diharapkan untuk mencegah kerusakan
jaringan buah. Perlakuan uap panas dan udara panas mempunyai prinsip kerja

18
menggunakan udara yang dipanaskan untuk menaikkan temperatur buah yang
dapat mematikan serangga target. Perbedaan keduanya terletak pada tingkat
kelembaban yang digunakan dimana perlakuan uap panas lebih tinggi
dibandingkan perlakuan udara panas (APHIS 2011).
Fasilitas perlakuan uap panas dirancang untuk dapat mempertahankan
temperatur dan kelembaban tertentu dalam chamber. Temperatur dan
kelembaban dikendalikan dengan kendali elektrik temperatur dan kelembaban.
Mesin perlakuan uap panas dilengkapi pencatat rekaman
temperatur/kelembaban selama perlakuan berlangsung dengan sensor yang
dapat diatur posisinya dalam chamber perlakuan. Saat ini perlakuan uap panas
banyak diaplikasikan terhadap buah dengan OPT serangga sebagai target.
Kelembaban yang tinggi dapat mencegah evaporasi berlebihan pada buah. Uap
yang terdistribusi sempurna melalui sirkulasi udara memungkinkan kondisi uap
panas yang terkendali dalam chamber perlakuan (JFTA 1996).
Penelitian mengenai perlakuan uap panas dilakukan sejak tahun 1910-an
untuk membunuh telur dan larva lalat buah. Selanjutnya perlakuan uap panas
digunakan pertamakali pada tahun 1929 dalam dunia karantina tumbuhan. Pada
masa itu perlakuan dilakukan terhadap buah yang dikirim dari Florida ke daerah
lain di Amerika Serikat untuk mencegah penyebaran C. capitata. Pada awalnya
standar perlakuan uap panas yang berlaku adalah temperatur 43.3-46oC selama
delapan jam. Namun keterbatasan kemampuan peralatan masa itu
menyebabkan kesulitan untuk mengendalikan temperatur secara akurat selama
perlakuan sehingga terjadi kerusakan pada buah. Beberapa hama yang diuji
dengan perlakuan uap panas adalah lalat buah Anastrepha, thrips, pink
bollworm, dan cigarette beetle (JFTA 1996).
Pada tahun 1960-an teknologi perlakuan uap panas makin berkembang,
terutama oleh negara Jepang. VHT juga merupakan salah satu metode
perlakuan yang dipersyaratkan bagi ekspor buah ke beberapa negara. Salah
satunya persyaratan pepaya varietas Solo (Hawaii) ke Jepang. Jenis perlakuan
perlakuan uap panas mempunyai beberapa kelebihan dibandingkan dengan
metode konvensional lain, diantaranya kerusakan akibat panas pada komoditas
pertanian lebih rendah daripada metode perendaman air panas serta periode
pemaparan panas dapat lebih singkat daripadaperlakuan panas kering. Selain
itu, aplikasi perlakuan uap panas juga dapat mencegah adanya residu bahan
kimia pada buah segar yang diberi perlakuan, sehingga aman untuk dikonsumsi.

19
Tabel 3 Perlakuan uap panas untuk memenuhi aturan karantina tumbuhan
Komoditas (asal) Negara tujuan Jenis perlakuan
Mangga (Filipina) Australia, Amerika Serikat
Temperatur 46oC selama 10 menit untuk target B. cucurbitae, B. philippinensis, dan B. occipitalis
Mangga ‘Kensington’ (Australia)
Jepang Temperatur 47o
C selama 15 menit untuk target B. tryoni
Pepaya ‘Solo’ (Filipina)
Jepang Temperatur 46o
Yellow pitaya (Kolombia)
C selama 10 menit untuk target B. cucurbitae.
Amerika Serikat Temperatur 46o
Jeruk (Meksiko)
C selama 20 menit untuk target C. capitata dan Anastrepha fraterculus.
Amerika Serikat Temperatur 43.3oC selama 6 jam untuk target Anastrepha spp.
Sumber: JFTA (1996) dan APHIS (2011)
Saat ini fasilitas komersial perlakuan uap panas telah beroperasi di
Jepang, Thailand, Filipina, Australia, dan Amerika Serikat (Monck dan Pearce
2007; Hansen et al. 1992). Jepang, Amerika Serikat, dan beberapa negara lain
menerapkan aturan perlakuan untuk komoditas buah-buahan yang berasal dari
kawasan yang tidak bebas B. dorsalis, B. cucurbitae, dan C. capitata. Salah satu
metode yang dipersyaratkan adalah perlakuan uap panas. Sedangkan Australia
mengharuskan penerapan perlakuan uap panas 46o
C selama 10 menit untuk
mendisinfestasi B. cucurbitae, B. occipitalis, dan B. philippinensis. Beberapa
contoh perlakuan uap panas sebagai aturan karantina tumbuhan dapat dilihat
pada Tabel 3.
Perlakuan uap panas dengan kelembaban lebih dari 90% digunakan oleh
APHIS-USDA untuk mendisinfestasi mexican fruitfly pada buah clementine,
anggur, jeruk, dan mangga. Uap panas juga dilaporkan efektif untuk
mendisinfestasi mealybug pada Pisum sativum (Follett et al. 2004) dan thrips
pada bunga potong (Hansen et al. 1992).
Perlakuan uap panas dimulai dengan periode pemanasan dimana lama
waktu yang dibutuhkan tergantung dari jenis komoditas yang diberi perlakuan.
Waktu perlakuan berdasarkan temperatur internal dari komoditas telah mencapai
temperatur yang diinginkan untuk membunuh serangga target. Selanjutnya
perlakuan uap panas biasanya diikuti pendinginan dengan air atau udara (Lurie
1998).





























