Embed Size (px)
Citation preview

30
HASIL DAN PEMBAHASAN
Karakteristik Kultur Starter Koumiss dan Bakteri Patogen
Kultur starter koumiss yang digunakan dalam penelitian ini terdiri atas bakteri
Lc. lactis D-01, Lb. acidophilus Y-01 dan khamir S. cereviceae. Bakteri patogen
yang digunakan berupa S. Typhimurium ATCC 14028 dan M. tuberculosis H37RV.
Pemeriksaan kultur starter koumiss bertujuan untuk mempelajari karakteristik
dinding sel (pewarnaan Gram), morfologi (bentuk dan susunan), sifat katalase dan
keberadaan kontaminasi dengan mikroorganisme lain yang tidak diinginkan.
Hasil pemeriksaan memperlihatkan bahwa kultur starter koumiss dan bakteri
patogen memiliki koloni yang homogen dan menunjukkan kesesuaian karakteristik
morfologi dari masing-masing bakteri dan khamir serta tidak terdapat kontaminasi.
Bakteri Lc. lactis D-01 dan Lb. acidophilus Y-01 berdasarkan pewarnaan Gram
termasuk ke dalam kelompok bakteri asam laktat Gram positif, sesuai dengan
Chandan et al. (2008) dan Holt et al. (1994) yang menyatakan bahwa bakteri Lc.
lactis dan Lb. acidophilus tergolong ke dalam bakteri Gram positif.
Lc. lactis D-01 dan Lb. acidophilus Y-01 tergolong ke dalam bakteri Gram
positif karena dapat mempertahankan warna ungu dari kristal violet setelah diberikan
pewarna tandingan safranin. Bakteri Gram positif memiliki dinding sel yang lebih
tebal dari pada bakteri Gram negatif, sehingga membuat zat pewarna kristal violet
tidak dapat keluar dari sel. Dinding sel bakteri Gram positif terdiri atas lapisan
peptidoglikan 90% dengan ketebalan 18-80 nm dan lapisan tipis yakni asam teikoat
10% yang merupakan polimer dari ribitol fosfat yang dihubungkan dengan N
asetilglukosamin sehingga mampu menyerap warna ungu lugol dan tetap
mempertahankan warna tersebut ketika dicuci dengan alkohol (Fardiaz, 1992).
Bakteri S. Typhimurium ATCC 14028 termasuk ke dalam kelompok Gram
negatif karena tidak dapat mempertahankan warna ungu kristal violet sehingga
menyerap pewarna tandingan safranin. Bakteri Gram negatif tidak dapat
mempertahankan zat pewarna kristal violet disebabkan ketika ditetesi dengan alkohol
95%, komponen lipid dari dinding sel terekstraksi, pori-pori sel mengembang
sehingga membuat zat pewarna kristal violet keluar dari sel dan menjadi tidak
berwarna. Sel bakteri yang tidak bewarna tersebut akan menyerap zat pewarna
safranin sebagai pewarna tandingan sehingga akan tampak berwarna merah (Pelczar

31
dan Chan, 2007). Karakteristik kultur starter koumiss dan bakteri patogen disajikan
pada Tabel 5.
Tabel 5. Karakteristik Kultur Starter Koumiss dan Bakteri Patogen
Mikroorganisme Pewarnaan Morfologi Sifat katalase
Keberadaan kontaminan
Lc. lactis D-01
Gram positif bulat, berantai, pendek negatif
tidak ada, koloni
homogen
Lb. acidophilus Y-01
Gram positif batang,
berantai, pendek
negatif tidak ada,
koloni homogen
Sc. Cereviceae
- Oval tunggal td tidak ada,
koloni homogen
S. Typhimurium ATCC 14028
Gram negatif batang, tunggal,
berkoloni positif
tidak ada, koloni
homogen
M. tuberculosis H37RV
Ziehl-Neelsenbatang
bengkok, berkoloni
td tidak ada,
koloni homogen
keterangan: td = tidak diuji

32
Dinding sel bakteri Gram negatif terdiri atas peptidoglikan dengan ukuran
10-15 nm sehingga dinding selnya lebih tipis. Bakteri Gram negatif ini dikelilingi
membran luar yang terpisah dengan suatu ruang periplasmik yang terdiri atas bagian
dalam fosfolipid dan bagian luar lipopolisakarida (Fardiaz, 1992).
Morfologi kultur starter koumiss berupa Lc. lactis D-01 menunjukkan bahwa
bakteri ini berbentuk bulat, berantai pendek sesuai dengan pernyataan Surono (2004)
yang menjelaskan bahwa Lc. lactis termasuk ke dalam famili Streptococcaceae yang
memiliki bentuk bulat berantai pendek. Lb. acidophilus Y-01 berbentuk batang
berantai pendek. Khamir Sc. cereviceae berbentuk oval dan tunggal, sesuai dengan
Fardiaz (1992) yang menyatakan bahwa Sc. cereviceae memiliki bentuk oval dengan
serta memiliki ukuran yang lebih besar dibandingkan bakteri.
Morfologi bakteri S. Typhimurium ATCC 14028 mempunyai bentuk batang
pendek dan berkoloni sesuai dengan Holt et al. (1994) bahwa S. Typhimurium
merupakan bakteri yang berbentuk batang pendek, Gram negatif, anaerob fakultatif
dan memiliki flagela peritrikat. Pemeriksaan untuk bakteri M. tuberculosis H37RV
berbeda dengan pemeriksaan starter koumiss dan S. Typhimurium ATCC 14028 yaitu
menggunakan pewarnaan Ziehl-Neelsen. Pewarnaan ini khusus untuk
mengidentifikasi bakteri yang tahan asam. M. tuberculosis H37RV memiliki bentuk
batang bengkok dan berkoloni, sesuai pernyataan Brooks et al. (2005) bahwa M.
tuberculosis berbentuk batang lurus atau agak bengkok.
Kultur starter bakteri asam laktat koumiss berupa Lc. lactis D-01 dan Lb.
acidophilus Y-01 memiliki sifat katalase negatif karena tidak melepaskan O2. Buckle
et al. (2007) menyatakan bahwa Lc. lactis D-01 dan Lb. acidophilus Y-01 termasuk
ke dalam katalase negatif yang berarti bakteri tersebut memiliki enzim peroksidase
yang dapat mencegah produksi gas O2. Bakteri patogen S. Typhimurium ATCC
14028 tergolong ke dalam katalase positif karena dapat melepaskan O2 sesuai dengan
Surono (2004).
Viabilitas Kultur Starter Koumiss dan Bakteri Patogen
Kultur starter BAL, Lb. acidophilus Y-01 mempunyai populasi 1,16 × 108
cfu/ml, Lc. lactis D-01 sebanyak 2,21 × 108 cfu/ml dan khamir Sc. cereviceae
1,42 × 109 cfu/ml. Populasi BAL telah memenuhi standar jumlah minimal yang
dibutuhkan dalam pembuatan susu fermentasi yaitu sebesar 108 cfu/ml (Makinen dan

33
Bigret, 1998). Populasi khamir sepuluh kali lebih banyak dari BAL, sehingga
penggunaannya sebagai kultur starter harus diencerkan terlebih dahulu. Populasi S.
Typhimurium ATCC 14028 dan M. tuberculosis H37RV sebesar 108 cfu/ml sesuai
dengan standar 0,5 Mc. Farland (NCCLS,1991) untuk pengujian Aktivitas
antimikroba.
Karakteristik Koumiss
Karakteristik Fisik Koumiss
Bahan utama pembuatan koumiss adalah susu kuda yang memiliki
karakteristik bewarna putih kebiruan, konsistensi cair dan rasa yang manis. Susu
kuda menurut SNI 01-6054-1999 memiliki karakteristik berwarna putih kebiruan,
beraroma khas dan berasa manis. Seydim et al. (2010) juga menyatakan bahwa susu
kuda memiliki konsistensi cair, berwarna putih dan mengandung laktosa tinggi
sehingga lebih manis bila dibandingkan dengan susu sapi atau susu kambing.
Koumiss yang dihasilkan memiliki karakteristik fisik berwarna putih, agak
kental, dengan sedikit gelembung gas CO2 hasil fermentasi dari khamir. Koumiss
juga memiliki rasa yang manis dan agak asam. Koumiss tetap memiliki rasa manis
walaupun agak asam yang berasal dari hasil metabolisme bakteri asam laktat. Enzim
β-galactosidase, glycolase dan lactate dehydrogenase yang diproduksi oleh kultur
starter BAL akan mengubah laktosa menjadi asam laktat sehingga dapat menurunkan
pH dan menyebabkan susu menjadi asam (Surono, 2004).
Nilai pH dan TAT Koumiss
Susu kuda pasteurisasi mengalami penurunan pH dan peningkatan TAT
selama proses fermentasi. Penurunan pH dan peningkatan TAT terjadi karena
akumulasi asam organik hasil metabolisme BAL. Penurunan pH disertai peningkatan
TAT terjadi pada koumiss segar (H0) hingga hari ke-4 (H4) penyimpanan. Asam
organik yang dihasilkan akan semakin tinggi karena laktosa terus dipecah menjadi
asam laktat oleh bakteri asam laktat (L. lactis D-01 dan L. acidophilus Y-01) yang
terkandung di dalam koumiss. Asam laktat adalah produk akhir dari fermentasi
karbohidrat oleh Lactobacillus sp. Pertumbuhan bakteri pembusuk dan patogen
terhambat akibat pH yang rendah dan konsentrasi asam organik yang tinggi (Cintas
et al., 2001).

34
Penurunan pH tidak terlalu tinggi karena koumiss disimpan pada suhu 4 oC.
Lee dan Wong (1998) menyatakan bahwa penyimpanan pada suhu rendah dapat
menghambat produksi asam hasil metabolisme dari bakteri asam laktat sehingga
mencegah produk susu fermentasi menjadi terlalu masam. Nilai pH dan TAT
disajikan pada Tabel 6.
Tabel 6. Nilai pH dan TAT Koumiss dengan berbagai Lama Penyimpanan pada Suhu 4 oC
Lama Penyimpanan pH TAT (% asam laktat)
H0 3,97 ± 0,004 1,17 ± 0,089
H2 3,92 ± 0,028 1,34 ± 0,166
H4 3,84 ± 0,005 1,73 ± 0,102
H6 3,88 ± 0,011 2,06 ± 0,083
H8 3,87 ± 0,004 1,77 ± 0,032
Susu kuda pasteurisasi 6,29 ± 0,014 0,35 ± 0,035
Keterangan: H0 = koumiss segar; H2 = koumiss disimpan selama dua hari; H4 = koumiss disimpan selama empat hari; H6 = koumiss disimpan selama enam hari; H8 = koumiss disimpan selama delapan hari
Nilai TAT pada koumiss pada berbagai lama penyimpanan yang diperoleh
pada penelitian ini (Tabel 6), masih memenuhi Standar Nasional Indonesia 01-2981-
1992 (1992) untuk susu fermentasi yaitu sebesar 0,5%-2%. TAT semakin meningkat
dengan penyimpanan yang semakin lama hingga hari ke-6 (H6).
Peningkatan pH diikuti dengan penurunan TAT terjadi pada koumiss
penyimpanan hari ke-8 (H8). Khamir tumbuh pada pH rendah dengan pertumbuhan
bakteri yang terhambat. Khamir tumbuh pada pH 2,5-8,5 dengan pH optimum
tumbuh 4-5 (Fardiaz, 1992). Aktivitas khamir dalam susu fermentasi menghasilkan
senyawa metabolit berupa alkohol, diasetil dan CO2 yang bersifat basa (Surono,
2004). Hal ini berakibat pada penurunan persentase asam laktat disertai dengan
peningkatan pH.
Karakteristik Mikrobiologis Koumiss
Karakteristik mikrobiologis koumiss yang diamati pada penelitian ini berupa
total koliform, total mikroorganisme (TPC), total bakteri asam laktat dan total
35
khamir. Pengamatan dilakukan pada susu kuda segar, susu kuda pasteurisasi, kultur
starter koumiss dan koumiss dengan berbagai lama penyimpanan. Perubahan
karakteristik mikrobiologis susu kuda segar, susu kuda pasteurisasi dan koumiss
segar, disajikan pada Tabel 7.
Tabel 7. Karakteristik Mikrobiologis Susu Kuda Segar, Susu Kuda Pasteurisasi dan Koumiss Segar
Produk TPC Koliform BAL Khamir
----------------------------(Log10 cfu/ml)--------------------
Susu kuda segar 6,27 >3 6,30 6,64
SNI Susu Segar 01-3141-1992
6 1,3
Susu kuda pasteurisasi LTLT 6,12 >3 6,03 6,01
SNI Susu Pasteurisasi 01-3951-1995 4,48 1
Koumiss segar 9,67 <1 10,13 9,72
Susu fermentasi 2981:2009 Min 7 1
Keterangan: TPC = Total Plate Count, BAL = Bakteri Asam Laktat
Karakteristik mikrobiologis susu kuda segar yang diamati, memiliki TPC
6,27 log10 cfu/ml, koliform > 3 log10 cfu/ml, BAL 6,30 log10 cfu/ml dan khamir 6,64
log10 cfu/ml; belum sesuai dengan Standar Nasional Indonesia 01-3141-1992 untuk
TPC yaitu sebesar 6 log10 cfu/ml dan total koliform sebesar 1,3 log10 cfu/ml. Total
koliform yang merupakan indikator sanitasi penanganan susu, kondisi pemerahan
dan pengelolaan yang kurang higienis; dapat meningkatkan pencemaran bakteri
koliform ke dalam susu. Hasil penelitian memperlihatkan bahwa penanganan susu
kuda segar oleh peternak kurang dikelola dengan baik sehingga pencemaran koliform
ditemukan masih tinggi.
Salah satu bentuk penanganan susu adalah dengan pemanasan (pasteurisasi).
Pemanasan dapat memberi daya tahan yang lebih lama terhadap susu dan menjamin
kelayakan untuk dikonsumsi. Karakteristik mikrobiologis susu kuda pasteurisasi
yaitu memiliki TPC 6,12 log10 cfu/ml, koliform > 3 log10 cfu/ml, BAL 6,03 log10
cfu/ml dan khamir 6,01 log10 cfu/ml. Nilai-nilai karakteristik mikrobiologis ini

36
mengalami penurunan jika dibandingkan dengan susu kuda segar, sehingga tidak
memenuhi Standar Nasional Indonesia 01-3951-1995 (1995). Standar Nasional
Indonesia 01-3951-1995 (1995) menyatakan bahwa TPC susu pasteurisasi
seharusnya mengalami penurunan dari 2 log10 cfu/ml menjadi 4,48 log10 cfu/ml.
Total koliform susu kuda pasteurisasi mengalami penurunan setelah dipasteurisasi,
tetapi belum memenuhi standar yang seharusnya turun menjadi 1 log10 cfu/ml,
sehingga perlakuan pasteurisasi LTLT (Low Temperature Low Time) belum mampu
untuk menekan jumlah bakteri koliform dan bakteri perusak lain.
Karakteristik mikrobiologis koumiss segar pada penelitian ini memiliki TPC
9,67 log10 cfu/ml, koliform > 1 log10 cfu/ml, BAL 10,13 log10 cfu/ml dan khamir 9,72
log10 cfu/ml; masih sesuai dengan Standar Nasional Indonesia 2981:2009 (2009).
Standar Nasional Indonesia 2981:2009 (2009) menyatakan bahwa koumiss segar
memiliki minimal total kultur starter 7 log10 cfu/ml dan total koliform 1 log10 cfu/ml.
Penurunan total koliform terjadi karena koliform tidak dapat bertahan hidup pada
koumiss dengan pH rendah. Wood (1999) menyatakan bahwa penurunan jumlah
koliform pada produk susu asam disebabkan penurunan pH akibat produksi asam
laktat. Akumulasi asam laktat dan asetat hasil metabolisme laktosa dari kultur starter
dapat merusak sel-sel bakteri koliform karena proses pengasaman sitoplasma oleh
difusi asam-asam terdisosiasi. Jumlah BAL dan khamir koumiss segar yang
teridentifikasi pada penelitian ini melebihi standar TPC, karena media yang
digunakan berbeda-beda sesuai dengan media tumbuh selektif pada masing-masing
bakteri dan khamir. BAL dapat tumbuh banyak pada media MRSA (de Man Rogosa
Sharpe Agar) dan khamir pada media PDA (Potato Dextrose Agar), tetapi tumbuh
sedikit pada media PCA (Plate Count Agar).
Jumlah koloni BAL semakin menurun hingga penyimpanan hari ke-4 (H4)
dan meningkat pada penyimpanan hari ke-6 (H6) dan hari ke-8 (H8). Hal tersebut
diilustrasikan pada Gambar 11. Penurunan koloni disebabkan penurunan pH hingga
3,84 ± 0,005 (Tabel 6) yang menghambat pertumbuhan BAL. Surono (2004)
menyatakan bahwa sejumlah besar asam laktat dalam bentuk tidak terdisosiasi akan
menjadi racun bagi banyak bakteri, khususnya bakteri asam laktat; pada pH rendah.
Koumiss mengandung BAL (Lc. lactis D-01 dan Lb. acidophilus Y-01) yang
memiliki tingkat toleransi terhadap asam laktat yang berbeda. Bakteri Lc. lactis

37
lebih cepat menghasilkan asam laktat, namun akan mati akibat asam tersebut.
Kondisi asam yang tinggi masih dapat ditoleransi oleh Lb. acidophilus sehingga
bakteri tetap hidup. Lb. acidophilus tidak dapat bertahan lama karena akumulasi
asam laktat yang semakin tinggi dan berakibat pada kematian bakteri tersebut
(Wijaningsih, 2008).
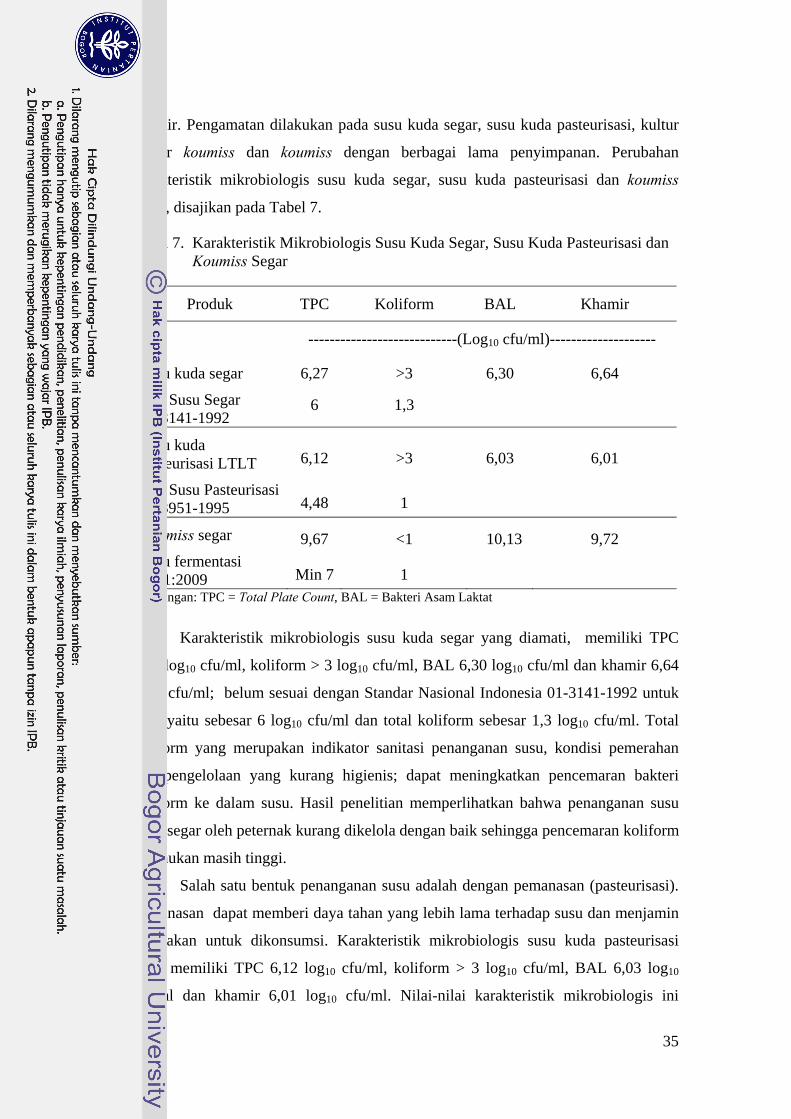
Jumlah koloni khamir semakin menurun hingga penyimpanan hari ke-4 (H4)
dan mengalami peningkatan pada penyimpanan hari ke-6 (H6) dan hari ke-8 (H8)
Hal tersebut diilustrasikan pada Gambar 12. Khamir lebih tahan hidup pada pH
asam dibanding bakteri (Fardiaz, 1992). Jumlah koloni khamir selama penyimpanan
berkisar 9-11 log10 cfu/ml. Jumlah ini lebih sedikit dibandingkan dengan jumlah
koloni BAL selama penyimpanan berkisar 8-12 log10 cfu/ml. BAL dan khamir
tumbuh bersama-sama membentuk simbiosis di dalam koumiss seperti kefir grain.
Khamir di dalam kefir grain berfungsi untuk memelihara integritas dan viabilitas
populasi mikroflora. Asam amino dan faktor esensial pertumbuhan bakteri asam
laktat diproduksi khamir, sedangkan senyawa metabolit dari BAL digunakan sebagai
sumber energi. Simbiosis antara BAL dan khamir ini membuat produk kefir menjadi
stabil (Farnworth dan Mainville, 2003).
10,138,92
8,01
12,0011,50
0
2
4
6
8
10
12
14
0 2 4 6 8
Jum
lah
Kol
oni L
og 1
0 cf
u/m
l
Lama Penyimpanan (hari)
Gambar 11. Pertumbuhan Koloni BAL ( ) di dalam Koumiss pada Berbagai Lama Penyimpanan
38
Aktivitas Daya Hambat Antimikroba Koumiss terhadap S. Typhimurium ATCC 14028
Pengujian daya hambat antimikroba koumiss terhadap S. Typhimurium
ATCC 14028 pada berbagai lama penyimpanan dilakukan dengan dua metode, yaitu
metode pour plate dan spread plate. Diameter penghambatan susu kuda pasteurisasi
selama penyimpanan adalah 0 mm. Susu kuda pasteurisasi memiliki total BAL dan
khamir berturut-turut sebesar 6,03 cfu/ml dan 6,01 cfu/ml (Tabel 7). Jumlah tersebut
tidak dapat menghambat populasi S. Typhimurium ATCC 14028 pada kedua metode.
Susu kuda memiliki antimikroba alami yaitu lisozim dan laktoferin sebesar 0,2-2
g/kg yang lebih tinggi dibandingkan susu sapi, unta, kambing dan kerbau (Sheng dan
Fang, 2009), tetapi belum dapat menghambat pertumbuhan S. Typhimurium ATCC
14028.
Filtrat adalah substrat antimikroba kasar dari BAL (L. lactis D-01 dan L.
acidophilus Y-01 hasil sentrifugasi koumiss 6.000 rpm selama 20 menit. Diameter
penghambatan filtrat terhadap S. Typhimurium ATCC 14028 pada metode pour
plate semakin besar selama penyimpanan filtrat. Diameter penghambatan susu kuda
pasteurisasi, filtrat dan koumiss pada metode pour plate dan berbagai lama
penyimpanan disajikan pada Tabel 8.
Senyawa metabolit yang bersifat antimikroba dan dihasilkan BAL terdiri atas
senyawa metabolit primer (asam laktat, asam asetat dan hidrogen peroksida) dan
9,729,17
7,94
10,2511,39
0
2
4
6
8
10
12
0 2 4 6 8
Jum
lah
Kol
oni L
og 1
0 cf
u/m
l
Lama Penyimpanan (hari)
Gambar 12. Pertumbuhan Koloni Khamir ( ) di dalam Koumiss pada Berbagai Lama Penyimpanan

39
senyawa metabolit sekunder (bakteriosin, diasetil dan CO2) (Surono, 2004). Lama
penyimpanan berpengaruh pada jumlah senyawa metabolit, yaitu semakin lama
penyimpanan filtrat maka senyawa metabolit yang dihasilkan semakin banyak dan
berakibat pada penghambatan pertumbuhan S. Typhimurium ATCC 14028 yang
semakin besar. Senyawa metabolit BAL menghambat berbagai bakteri patogen
seperti E. coli, S. Typhimurium dan C. perfringens (Thanh et al., 2010). Diameter
penghambatan koumiss terhadap S. Typhimurium ATCC 14028 pada metode pour
plate mengalami peningkatan hingga koumiss pada penyimpanan hari ke-2 (H2).
Tabel 8. Diameter Penghambatan Susu Kuda Pasteurisasi, Filtrat dan Koumiss terhadap S. Typhimurium ATCC 14028 dengan Metode Pour Plate dan Spread Plate pada Berbagai Lama Penyimpanan
Produk Diameter (mm)
H0 H2 H4 H8
Pour Plate (populasi S. Typhimurium 106 cfu/ml)
Susu kuda segar 0 0 0 0
Filtrat 5,77 ± 0,62 5,73 ± 0,97 6,04 ± 0,03 6,47 ± 0,37
Koumiss 7,99 ± 0,21 8,03 ± 0,19 7,52 ± 0,34 7,67 ± 0,66
Spread Plate (populasi S. Typhimurium 108 cfu/ml)
Susu kuda segar 0 0 0 0
Filtrat 4,66 ± 0,09 4,82 ± 0,06 3,97 ± 0,59 6,50 ± 0,95
Koumiss 5,48 ± 0,27 6,30 ± 0,05 5,45 ± 1,78 6,65 ± 0,65
Keterangan: H0 = koumiss segar; H2 = koumiss disimpan selama dua hari; H4 = koumiss disimpan selama empat hari; H6 = koumiss disimpan selama enam hari; H8 = koumiss disimpan selama delapan hari
Peningkatan diameter penghambatan hingga penyimpanan hari ke-2 (H2)
disebabkan kandungan subtrat antimikroba berupa asam laktat dan bakteriosin yang
dihasilkan BAL dalam koumiss. Penghambatan tersebut dipengaruhi temperatur,
faktor lingkungan antimikroba, kondisi keasaman dan ketersediaan makanan (Thanh,
2010). Produksi bakteriosin oleh bakteri asam laktat memperlihatkan efek
bakterisidal. Bakteriosin dari Lactobacillus dapat menembus membran terluar bakteri

40
dan meningkatkan inaktivasi bakteri Gram negatif. Bakteriosin mengubah
keseimbangan membran potensial dengan mengambil ion K+ sehingga sel tidak dapat
menyeimbangkan pH intraseluler (Sezer dan Guven, 2009).
Ray (2000) menyatakan bahwa aktivitas antimikroba terjadi akibat produksi
asam yang tidak terdisosiasi dan terdisosiasi. Bentuk asam yang tidak terdisosiasi
pada suatu komponen antimikroba akan mengakibatkan proton lebih cepat masuk ke
dalam sel. Nilai pH yang rendah akan mengakibatkan proton yang masuk ke dalam
sitoplasma sel semakin banyak, sehingga energi yang diperlukan semakin banyak
untuk mengeluarkan proton. Pengeluaran proton ini dilakukan untuk mencegah
pengasaman dan denaturasi komponen-komponen sel, sehingga bakteri yang tidak
memiliki cukup energi akan mati. Asam laktat yang dihasilkan dalam fermentasi
mampu menurunkan pH dan mengganggu aktivitas enzim sehingga sel tidak dapat
melakukan aktivitas metabolisme. Membran terluar bakteri Gram negatif menjadi
permeabel dan memungkinkan bakteri S. Typhimurium menjadi sensitif. Kombinasi
senyawa metabolit berupa bakteriosin dan asam laktat, memiliki penghambatan
lebih besar dibandingkan senyawa metabolit tunggal (Thu et al., 2011).
Penurunan diameter penghambatan terhadap S. Typhimurium ATCC 14028
terjadi pada koumiss pada penyimpanan hari ke-4 (H4). Penurunan terjadi karena
metabolisme antimikroba itu sendiri yaitu berupa enzim yang dapat menghidrolisis
protein dan karbohidrat sehingga berakibat pada penurunan daya hambat antimikroba
(Murray, 1997). Bakteri Gram negatif memiliki dinding sel lebih kompleks terutama
lapisan luar peptidoglikan dan lapisan yang terdiri atas fosfolipida, polisakarida dan
protein. Lipid dan polisakarida membentuk struktur yang khas yang disebut dengan
lipopolisakarida atau LPS, sehingga mempunyai daya pertahanan yang lebih kuat
terhadap bahan asing yang menembus ke dalam sel bakteri (Lay dan Hastowo,
1992).
Peningkatan diameter penghambatan koumiss terhadap S. Typhimurium
ATCC 14028 terjadi pada hari penyimpanan ke-8 (H8). Koumiss tersebut memiliki
pH sebesar 3,87 ± 0,004 dan TAT 1,77 ± 0,032. Nilai pH optimum untuk
pertumbuhan S. Typhimurium adalah 6,5-7,5 (Cox, 2000), sehingga pertumbuhan S.
Typhimurium ATCC 14028 terhambat akibat pH koumiss yang terlalu rendah. Asam
ini merupakan akumulasi asam laktat hasil metabolisme BAL dan hasil metabolisme

41
khamir seperti asam asetat, sitrat, dan suksinat yang dapat dikeluarkan sel ke
medium (Walker, 1998).
Pertumbuhan S. Typhimurium ATCC 14028 dapat juga dihambat hidrogen
peroksida. Senyawa ini bereaksi dengan ion klorida di dalam sel dan membentuk
hipoklorit yang meracuni sel mikroba. Hidrogen peroksida dapat secara efektif
menghambat pertumbuhan mikroba bila tersedia dalam konsentrasi yang sangat
tinggi dan melakukan kontak dengan mikroba dalam waktu yang cukup lama (Cords
dan Dychdala, 1993). Efek dari asam laktat dalam menghambat S. Typhimurium
tidak hanya disebabkan pH yang rendah dalam produk tetapi juga karena aktivitas
intrasellular yang masuk ke dalam sel (Yessilik et al., 2011).
Diameter penghambatan filtrat koumiss terhadap S. Typhimurium ATCC
14028 dengan metode spread plate mengalami peningkatan pada penyimpanan hari
ke-2 (H2). Hal tersebut disajikan pada Tabel 8. Peningkatan penghambatan filtrat
karena akumulasi senyawa metabolit yang bersifat antimikroba berakibat pada
diameter penghambatan terhadap S. Typhimurium ATCC 14028 semakin besar.
Bakteriosin adalah senyawa metabolit hasil metabolisme BAL, yang bersifat
antimikroba. Senyawa ini diproduksi saat fase pertumbuhan logaritmik bakteri dan
berhenti pada fase stationer (Savadogo et al., 2006). Penurunan diameter
penghambatan terhadap S. Typhimurium ATCC 14028 terjadi pada filtrat koumiss
dengan penyimpanan hari ke-4 (H4). Hurst dan Hoover (1993) menyatakan bahwa
semakin lama penyimpanan maka efektivitas bakteriosin semakin menurun.
Peningkatan diameter kembali terjadi pada filtrat koumiss dengan penyimpanan hari
ke-8 (H8). Peningkatan diameter karena jumlah substrat antimikroba kasar selama
penyimpanan hari ke-8 (H8) lebih banyak dibandingkan dengan penyimpanan hari
ke-6 (H6).
Diameter penghambatan koumiss terhadap S. Typhimurium ATCC 14028
dengan metode spread plate mengalami peningkatan hingga penyimpanan hari ke-2
(H2). Peningkatan diameter penghambatan terjadi karena peran dari senyawa
metabolit yang bersifat antimikroba, khususnya asam laktat. Tingkat keefektifan
asam laktat lebih besar dibandingkan asam sitrat sehingga mampu memberikan
aktivitas antimikroba yang membuat membran bakteri patogen menjadi tidak stabil
(Ray, 2000). Diameter penghambatan koumiss terhadap S. Typhimurium ATCC

42
14028 dengan metode spread plate lebih kecil dibandingkan metode pour plate.
Jumlah populasi S. Typhimurium ATCC 14028 pada metode spread plate yaitu 108
cfu/ml, sedangkan metode pour plate 106 cfu/ml. Aktivitas antimikroba yang dapat
diamati pada uji difusi sumur dipengaruhi beberapa faktor, seperti: (1) tipe dan
ukuran cawan, (2) tipe agar, pH dan kandungan garam, (3) kemampuan zat untuk
berdifusi ke dalam agar, (4) karakteristik media dan (5) jenis bakteri uji yang
digunakan (Branen, 1993).
Penggunaan antibiotika pada manusia dilakukan untuk penyembuhan
penyakit tipus yang disebabkan bakteri S. Typhimurium. Antibiotika yang dapat
menghambat pertumbuhan S. Typhimurium adalah kloramfenikol dan ampisilin.
Kloramfenikol merupakan antibiotika yang mempunyai spektrum luas dan aktif
terhadap bakteri Gram positif dan Gram negatif, bakteriostatik. Kloramfenikol dapat
melekat pada ribosom bakteri serta mengganggu pengikatan asam amino baru pada
rantai peptida. Ampisilin merupakan antibiotika yang termasuk ke dalam golongan
penisilin. Ampisilin tidak membunuh bakteri secara langsung tetapi dengan cara
mencegah bakteri membentuk semacam lapisan kapsul yang melekat pada tubuh
bakteri. Lapisan ini berfungsi untuk melindungi tubuh bakteri. Dosis antibiotik
kloramfenikol sebesar 40 mg/kg/hari digunakan pada orang dewasa, sedangkan
dosis ampisilin sebesar dua g/hari (Kurniawan, 2007).
Konsentrasi kloramfenikol dan ampisilin sebesar 30 µg memiliki zona
sensitivitas terhadap bakteri Gram negatif masing-masing sebesar 21-27 mm dan 16-
22 mm (WH0, 2006). Zona penghambatan ini berada di atas rentang zona hambat
antimikroba filtrat dan koumiss sebesar 5,7-6,4 mm dan 7,5-8,0 mm. Kategori zona
hambat antimikroba secara umum adalah diameter > 5 mm yang dinyatakan lemah,
5-10 mm sedang (intermediet) dan >10-20 mm kuat serta >20-30 mm sangat kuat
(sensitif) (Morales et al., 2002). Penghambatan filtrat dan koumiss berdasarkan
kategori Morales et al. (2002) termasuk ke dalam kategori intermediet. Kategori
intermediet adalah kategori dengan respon penghambatan terhadap bakteri patogen
yang perlu ditangani dengan dosis antimikroba tinggi. Salah satu cara adalah dengan
melakukan terapi berkala dengan menggunakan antimikroba tersebut.

43
Penetapan Konsentrasi Koumiss untuk Penghambatan M. tuberculosis H37RV
Penetapan konsentrasi koumiss untuk penghambatan M. tuberculosis H37RV
adalah untuk mencari konsentrasi koumiss yang sesuai dan tepat dalam menghambat
M. tuberculosis H37RV sebelum pengujian penghambatan koumiss pada berbagai
lama penyimpanan. Konsentrasi yang dimaksud adalah persentase penambahan
koumiss ke dalam media Lowenstein Jensen sebagai media pertumbuhan M.
tuberculosis H37RV. Penghambatan koumiss terhadap M. tuberculosis H37RV
diilustrasikan pada Gambar 13.
Gambar 13. Penghambatan Koumiss pada kontrol, 5%, 7%, 10%, 12%, 14% dan 20% terhadap M. tuberculosis H37RV
Koumiss yang ditambahkan ke dalam media Lowenstein Jensen yang
dimodifikasi yaitu sebesar 5%, 7%, 10%, 12%, 14% dan 20% (v/v). Pertumbuhan M.
tuberculosis H37RV diamati setelah bakteri diinkubasi selama empat minggu.
Bakteri M. tuberculosis H37RV dapat tumbuh pada media yang ditambahkan
koumiss 5%, 7% dan 10%. Penghambatan terjadi pada media pertumbuhan M.
tuberculosis H37RV yang mendapat perlakuan penambahan koumiss sebanyak 14%
dan 20%.
Kontrol adalah media yang tidak mendapat penambahan koumiss sehingga
pertumbuhan M. tuberculosis H37RV ditemukan lebih banyak yaitu empat koloni,
sedangkan sebanyak 3 dan 2 koloni ditemukan pada media dengan penambahan
koumiss sebanyak 5% dan 7%. Hal ini menjelaskan bahwa penambahan koumiss
dapat menghambat pertumbuhan M. tuberculosis H37RV, tetapi persentase tersebut
tidak dikehendaki karena bakteri masih tumbuh.
k 7% 10%5 % 12% 14% 20%

44
Media pertumbuhan dengan penambahan koumiss sebanyak 10% dan 12%
memperlihatkan pertumbuhan M. tuberculosis H37RV masing-masing sebanyak 4
dan 1 koloni kecil. Pertumbuhan M. tuberculosis H37RV tidak ditemukan pada
media dengan penambahan koumiss sebanyak 14% dan 20%. Konsentrasi yang
digunakan pada pengujian selanjutnya adalah penambahan koumiss sebanyak 14%.
Penambahan koumiss dengan persentase 20% walaupun tidak memperli-
hatkan pertumbuhan M. tuberculosis H37RV, membuat media Lowenstein Jensen
hancur sehingga tidak digunakan dalam pengujian. Media yang baik untuk
penanaman M. tuberculosis H37RV adalah media yang tidak kering, tidak mudah
hancur dan tidak mengandung gelembung udara (Sjahrurachman, 2008).
Penghambatan Koumiss terhadap M. tuberculosis H37RV pada berbagai Lama Penyimpanan
Penghambatan koumiss pada berbagai lama penyimpanan dilakukan dengan
melihat ada tidaknya pertumbuhan M. tuberculosis H37RV. Pengamatan dilakukan
pada minggu ke-4, ke-6 dan ke-8. Bakteri M. tuberculosis tumbuh lambat, karena
koloni tampak setelah lebih kurang dua minggu bahkan setelah 6-8 minggu
pengamatan (Brooks et al., 2005). Sjahrurachman (2008) juga menyatakan bahwa
pembacaan untuk pengujian resistensi M. tuberculosis dilakukan pada hari ke-28,
apabila resisten (ditemukan pertumbuhan) maka pembacaan ulang tidak perlu
dilakukan. Pembacaan dilanjutkan hingga hari ke-42 sampai hari ke-56 apabila tidak
ditemukan pertumbuhan bakteri M. tuberculosis. Pengamatan pada minggu ke-6 dan
ke-8 berfungsi sebagai alat kontrol. Hasil pertumbuhan M. tuberculosis H37RV pada
kontrol dan berbagai lama penyimpanan koumiss pada pengamatan minggu ke-4.
Kontrol memperlihatkan pertumbuhan M. tuberculosis dengan proporsi 1,00
yang berarti M. tuberculosis H37RV tumbuh pada media Lowenstein Jensen kontrol
yang merupakan media asli pertumbuhan M. tuberculosis. Koumiss pada lama
penyimpanan 0, 2, 4, 6 dan 8 hari tidak memperlihatkan pertumbuhan (hasil proporsi
0,00). Hasil analisis Cohran juga menjelaskan bahwa media kontrol sangat berbeda
nyata (P<0,01) terhadap media yang telah mendapat penambahan koumiss pada
berbagai lama penyimpanan. Koumiss dapat menghambat pertumbuhan M.
tuberculosis H37RV pada pengamatan minggu ke-4, tetapi pengamatan harus tetap

45
dilakukan pada minggu ke-6 dan ke-8 untuk memastikan penghambatan terhadap
pertumbuhan M. tuberculosis H37RV.
Pertumbuhan M. tuberculosis H37RV memperlihatkan hasil yang sangat
nyata (P<0,01) antara kontrol dengan koumiss pada berbagai lama penyimpanan
untuk pengamatan minggu ke-6. Kontrol memperlihatkan proporsi sebesar 1,00
yang berarti bakteri M. tuberculosis H37RV mengalami pertumbuhan. Bakteri
M. tuberculosis H37RV tumbuh lebih banyak pada minggu ke-4.
Hasil analisis Cohran menyatakan bahwa koumiss pada peyimpanan H0, H2,
H4, H6, dan H8 tidak berbeda (P>0,05). Koumiss dapat menghambat pertumbuhan
M. tuberculosis H37RV. Penghambatan terjadi karena kandungan antimikroba di
dalam koumiss. Kandungan antimikroba tersebut berupa asam organik, bakteriosin
dan alkohol (Surono, 2004). Koumiss dengan lama penyimpanan 4, 6 dan 8 hari
masing-masing memiliki pH 3,84;3,88 dan 3,87 dengan total asam tertitrasi (TAT)
berturut-turut sebesar 1,74%; 2,06%; 1,3%.
Penghambatan pertumbuhan M. tuberculosis H37RV diduga terjadi karena
kandungan asam organik yang tinggi. Efek antimikroba dari asam organik
merupakan akibat dari nilai pH yang menurun dan bentuk molekul asam organik
yang tidak terdisosiasi (Surono, 2004). Asam laktat merupakan senyawa metabolit
utama yang dihasilkan susu fermentasi. Sejumlah besar asam laktat dalam bentuk
tidak terdisosiasi meracuni banyak bakteri, terutama bakteri patogen.
Terapi konsumsi koumiss memberikan hasil penurunan gejala penyakit
tuberkolosis terbaik saat diberikan yaitu gejala pengurusan, nafas pendek, kelelahan,
batuk berdahak, haemoptisis, keringat dingin, diare dan kelesuan (Burt, 2000).
Koumiss dapat meningkatkan fungsi sirkulasi, metabolisme, sistem syaraf,
pembentukan sel darah merah, fungsi ginjal, kelenjar endokrin dan sistem kekebalan
tubuh. Koumiss efektif digunakan untuk terapi tuberkolosis paru dan sistem
urogenital, kelelahan dan anemia. (Wang et al., 2008). Koumiss memiliki kandungan
Ca : P yaitu 2 : 1 dan kandungan vitamin A,C, E, B1, B2, B12 serta antimikroba yang
lebih tinggi dibandingkan susu kuda segar (Ping dan Li, 2009).
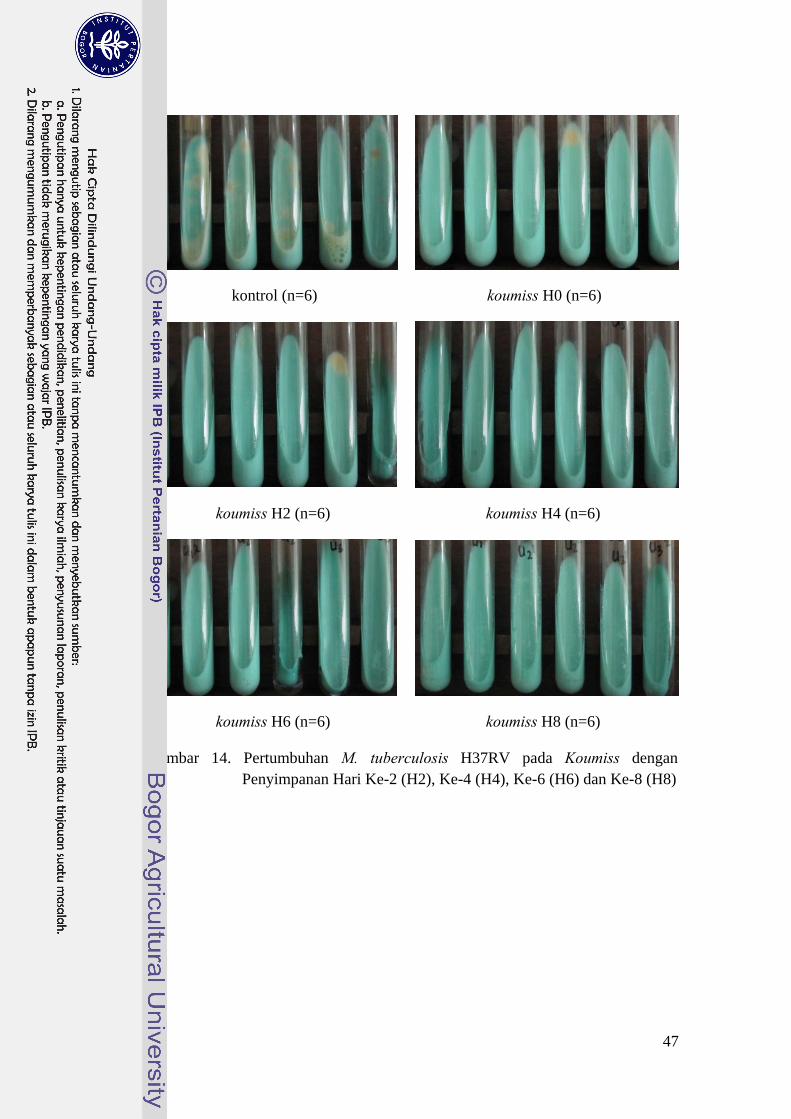
Pertumbuhan M. tuberculosis H37RV pada pengamatan minggu ke-8 untuk
perlakuan kontrol sangat nyata (P<0,01) dengan perlakuan penambahan koumiss
pada lama penyimpanan nol hari (H0), empat hari (H4), enam hari (H6) dan delapan

46
hari (H8). Aditama (1999) menyatakan bahwa sifat penghambatan senyawa hasil
fermentasi hanya bersifat bakteriostatik, karena M. tuberculosis masih dapat
berkembang ketika sifat keasaman dihilangkan hingga mencapai pH netral.
Pertumbuhan M. tuberculosis H37RV pada pengamatan minggu ke-8 disajikan pada
Gambar 14.
Perlakuan koumiss pada lama penyimpanan dua hari memperlihatkan hasil
yang tidak berbeda. Bakteri M. tuberculosis H37RV merupakan bakteri tahan asam
sehingga dapat tumbuh pada media yang asam walaupun terjadi secara lambat.
Koumiss perlakuan H2 yang memiliki pH 3,92 dan TAT sebesar 1,34% ternyata
belum dapat menghambat pertumbuhan M. tuberculosis H37RV secara signifikan.
Penelitian Rijatmoko (2003) menyatakan secara umum bahwa pertumbuhan
M. tuberculosis pada media dengan penambahan susu kuda terfermentasi dengan pH
netral membentuk koloni lebih sedikit dibandingkan media yang ditambahkan obat
INH (isoniazid) ataupun Rifampisin. Pernyataan tersebut menjelaskan bahwa tidak
hanya antimikroba koumiss berupa asam organik yang dapat menghambat
pertumbuhan M. tuberculosis H37RV.
Bakteriosin yang merupakan hasil metabolisme BAL ikut berperan dalam
menghambat pertumbuhan M. tuberculosis H37RV. Bakteriosin merupakan suatu
peptida yang kebanyakan bersifat bakterisidal yaitu membunuh bakteri patogen yang
tidak hanya menghambat pertumbuhan bakteri tersebut (Surono, 2004).
BAL yang digunakan dalam pembuatan koumiss pada penelitian ini yaitu Lb.
acidophilus Y-01. BAL tersebut menghasilkan bakteriosin berupa asidofilin dan
bakteri Lc. lactis D-01 penghasil nisin. Khamir pada koumiss menghasilkan senyawa
natamisin yang berfungsi sebagai fungisidal. Hurst dan Hoover (1993) menyatakan
bahwa nisin bekerja dengan cara melepaskan materi sitoplasmik sel sehingga terjadi
lisis. Nisin memiliki efek penghambatan dalam melawan bakteri Gram negatif
dengan merusak bagian luar membran sel bakteri (Rodriguez, 1996). Bakteriosin lain
bekerja dengan cara menyebabkan membran sitoplasma kehilangan asam amino dan
ion dengan cepat. Hal ini secara drastis akan menurunkan transport membran dari
seluruh sel (Barefoot dan Nettles, 1993).
47
kontrol (n=6) koumiss H0 (n=6)
koumiss H2 (n=6) koumiss H4 (n=6)
koumiss H6 (n=6) koumiss H8 (n=6)
Gambar 14. Pertumbuhan M. tuberculosis H37RV pada Koumiss dengan Penyimpanan Hari Ke-2 (H2), Ke-4 (H4), Ke-6 (H6) dan Ke-8 (H8)







![Modulationofadenylatecyclasetoxin productionasBordetella ... · BGmedium. Bacterial cells wereharvestedfromthe plates andresuspended in RPMI-1640andadjustedto anOD620of 0.5 [109colony-formingunits(cfu)/ml]](https://img.dokumen.tips/doc/110x75/5e2072e4b6beba411e4e979e/modulationofadenylatecyclasetoxin-productionasbordetella-bgmedium-bacterial.jpg)











![2017_Annual_Report_Congress · Web view[most probable number (MPN)/100 L or colony-forming units (CFU)/100 mL] elevation (m) emergent vegetation survey Enterococci (MPN/100 L or CFU/100](https://img.dokumen.tips/doc/110x75/5d1c0b8188c993d66e8c2d98/2017annualreportcongress-web-viewmost-probable-number-mpn100-l-or-colony-forming.jpg)









