Embed Size (px)
Citation preview

BASE Biotechnol. Agron. Soc. Environ.201014(4),667-680 Focus on:
GeneexpressionandgeneticanalysisduringhigherplantsembryogenesisGhassenAbid(1,2),Jean-MarieJacquemin(2),KhaledSassi(3),YordanMuhovski(2),AndréToussaint(1),Jean-PierreBaudoin(1)(1)Univ.Liege-GemblouxAgro-BioTech.UnitofTropicalCropHusbandryandHorticulture.PassagedesDéportés,2.B-5030Gembloux(Belgium).E-mail:[email protected](2)WalloonAgriculturalResearchCentre.DepartmentofBiotechnology.ChausséedeCharleroi,234.B-5030Gembloux(Belgium).(3)InstitutNationalAgronomiquedeTunisie.DepartmentofAgronomyandPlantBiotechnology.LaboratoryofAgronomy.AvenueCharlesNicolle,43.TU-1082Tunis-Mahrajène(Tunisia).
ReceivedonNovember21,2009;acceptedonMarch30,2010.
Thisreviewdescribesanddiscussesrecentattemptstoanalyzetheembryogenesisprocessinhigherplants,throughcombinationofdescriptive,experimental,andgeneticapproach.Analysisofgeneexpressionprofileshaspermitted tobuildhypothesisconcerningtheinducedmechanismsinearlyphasesofembryogenesisinhigherplants.Suchmechanismsinvolvespecifictranscriptionalandpost-transcriptionalregulatorypathwaysaswellasdiversesignaltransductionprocessesateachstageofplantdevelopment.Keywords.Plantembryogenesis,molecularregulation,geneexpression.
Analyse génétique et expression de gènes au cours de l’embryogenèse chez les plantes supérieures. Cetterevuedécritet discute les récentes études ciblant l’analysedesprocessusde l’embryogenèse chez lesplantes supérieuresgrâce àunecombinaisond’approchesdescriptive,expérimentaleetgénétique.L’étuded’expressiondegènesapermisdedégagerdeshypothèsessur lesmécanismes induitsaucoursdesphasesprécocesde l’embryogenèsechez lesplantessupérieures.Cesmécanismesimpliquentnotammentunerégulationtranscriptionelleetpost-transcriptionellefineetspécifiqueetdiversesvoiesdetransductiondusignalàchacundesstadesdedéveloppement.Mots clés.Embryogenèsechezlesplantessupérieures,régulationmoléculaire,expressiondegène.
1. IntroductIon
In flowering plant, embryogenesis begins with adoublefertilizationduringwhichthetwospermcellsof the pollen grainmerge with the egg cell and thecentralcelltoproducethezygoteandtheendosperm,respectively.Thezygote thendividesasymmetricallygiving rise to two cells of different size and fate:the small apical cell forms the major part of theembryo proper whereas the big basal cell developsinto the suspensor. Subsequently, the embryo growsaccording to two different axes, the apical-basal andtheradialaxes,toestablishdifferentorgansandtissues(Umehara et al., 2007). This developmental switchinvolves not only differential gene expression, butvarious signal transduction pathways for activating/repressingnumerousgenesets,manyofwhichareyetto be identified and characterized. Several processesoffloweringplantembryogenesisarenowwellknown
although limited genetic information is availableduringtheearlystagesofseeddevelopment.Molecularandphysiologicaleventsleadingtotheseedformationare still far to be completely understood.MutagenicsilencingofgenesandcharacterizationofmodelplantmutantssuchasArabidopsis thaliana L.withabnormalembryo development have been used as genetic tooltounderstandgeneregulationofplantembryogenesis(McElver et al., 2001). SeveralArabidopsis mutantswhich are altered during early or late embryodevelopment have been described and several geneshavebeenidentifiedthatplayasignificantroleduringcelldivisionandcellwallformationatvariousstagesofembryodifferentiation(Tzafriretal.,2004).Moreover,large-scalesingle-passsequencingofcDNApreparedfrom tissues with specific metabolism activity hasprovidedanefficientandrapidmeantowardsisolationof newgenes (Suh et al., 2003).The present reviewis thus an effort to collate information on various

668 Biotechnol. Agron. Soc. Environ. 201014(4),667-680 AbidG.,JacqueminJ.-M.,SassiK.etal.
embryo-specificgenesthathavebeenidentifiedduringfloweringplantembryogenesis.Italsofocusesonthepotentialbenefitsandlimitationsofmutantsanalysisinrelationwithplantembryodevelopment.Inparticular,wediscusssomeoftheprocessesinvolvedinembryoformationanditsgeneticcontrol;thesignalingsystemsthatregulatetheexpressionofrelevantgenesarealsoexamined.
2. GenetIc AnAlysIs oF plAnt eMbryoGenesIs
The growth and development of higher plants arecharacterized by the execution of cell division,expansion and differentiation along two axes: theapical-basalandtheradialaxis.Geneticapproachwasdeveloped to investigateembryogenesisdevelopmentin plant by isolating and characterizing mutantsexhibiting abnormal embryo development. Thesemutants represent an important tool to characterizethe corresponding genes and to elucidate their roles.Analysesofseedlinglethalmutantsledtothehypothesisthatearlydicotyledonousembryoisdividedintothreedomains:apical,centralandbasal(Lauxetal.,2004).The apical embryo domain will generate the shootmeristem and most of the cotyledons; the centralembryodomainwillformthehypocotylandrootsandcontribute to cotyledons and the root meristem; thebasalembryodomainwillgiverisetothedistalpartsoftherootmeristem,andthestemcellsofthecentralrootcap. Two meristematic cell populations arise duringembryogenesis: the Shoot Apical Meristem (SAM)generating the aerial part of the plant, and the RootApicalMeristem (RAM)generating theundergroundpart.
2.1. the apical domain of the embryo
Phytohormones such as auxin and several groups ofgenes cooperate to balance meristem maintenanceandorganproduction (Figure 1).Mutations ingenesinvolvedintheapicalembryodomainspecificationarepredictedtodisruptbothcotyledonandshootmeristemdevelopment.SHOOTMERISTEMLESS(STM)geneisrequiredforSAMformationduringembryogenesisandisexpressedbyonlyaspecificsetofcellswithinthe embryo apex. STM has been shown to code fora class I KOTTED-like homeodomain containingprotein. In the mutants stm organs originating fromthe shoot meristems are fused, which indicates arole of STM in restricting cells and organ formation(Long et al., 1998). The WUSCHEL (WUS) genehas been shown to encode homeobox genes family,which involves important regulatory genes in thespeciationofcellfateandbodyplantattheearlystage
ofembryogenesisinhigherorganism.Moreover,WUSand STM play complementary roles in maintainingshootmeristem.However,mutantwusandstmstudiessuggestedthatcombinedexpressionofbothWUSandSTMcantriggertheinitiationofectopicmeristemandorganogenesiseven indifferentiated tissues(Lenhardet al., 2002). The cell differentiation is controlledby the genes CLAVATA (CLV). The CLV genes arerequired to limit themeristem size (DeYoung et al.,2008).CLVandSTMgenesareknowntoplayoppositeor competitive roles in the regulation of meristemactivity.The interaction between theWUS andCLVpathwaysestablishesanegativefeedbackloopbetweenthe stem cell and the meristematic regions. Indeed,WUS expression is sufficient to induce meristemcell identity, and expression ofCLV genes repressesmeristemmaintenance andWUSactivity.The apicalregionof theglobularembryoispartitionedinto twodomains:aperipheraldomainofSTM-expressingcellsand a central domain showing expression of CUP-SHAPED COTYLEDON (CUC) genes. CUC genesregulateboththeformationoftheembryonicSAMandtheseparationofcotyledonsandfloralorgans(Takadaetal.,2001).Fujitaetal.(2008)showedthatembryotreatmentswithvariouspolarauxininhibitorsstoppedcotyledon andSAMdevelopment.This indicates theroleofauxinsignalinginthepatterningoftheapicalembryodomain.Mutations inotherhomeoboxgenesfamilies important for embryonic shoot meristemdevelopment, such as homeodomain LEUCINEZIPPER(HD-ZIP)andhomeodomainFINGER(PHD-FINGER)familiesrevealthatthesegenesnotonlyarerequired for stem cell initiation but also tomaintainthem. Functional analyses of zll mutants and geneexpressionindicatethatZWILLE(ZLL)isrequiredforstemcellfatewithinthedevelopingembryonicshootmeristem(Moussainetal.,1998).STMgeneinteractswithZLL,whichisexpressedatthelateglobularstageintheapicaldomain.Moreover,ZLLgeneappearstoplayasimilarroleasSTM,andmayberegulatingthespatial expression pattern of STM (Moussain et al.,1998). GURKE (GK) is assumed to ensure correctorganization of the shoot apical region (Torres-Ruizet al., 1996). Indeed, mutants gk showed apicaldefects in seedlings but also reduced the efficiencyof seed germination and seed growth. Likewise, thePASTICCINO(PAS)geneswhichareinvolvedinthecontrolofcelldivisionanddifferentiationarerequiredfor normal organization of the apical region in theembryo (Faure et al., 1998). PASTICCINO mutantssuch as pas1 and pas2 produce abnormal embryoswithalteredcotyledonprimordialleadingtoaflatapex(Bellecetal.,2002).Morerecently,Baudetal.(2004)identifiedlethalacc1mutantsaffectedinacethyl-CoAcarboxylase 1 (ACCase 1) that displays a similarembryo phenotype to GURKE and PASTICCINO

Embryogenesisinhigherplants 669
mutants. Genetic analyses have identified multiplegene functions involved in embryonic pattern;among themamember of theWOXgenes family isdifferentially expressed in the small apical cell afterthe asymmetric division of the zygote.WOXs genesencodeaWUSCHEL-relatedhomeoboxgene family.WOX2hasaputativezincfingerdomaindownstreamofthehomeodomain.Transcriptsareexpressedintheegg cell, the zygote and the apical cell lineage.Thisgeneisnecessaryforcelldivisionsthatformtheapicalembryodomain(Haeckeretal.,2003).PreviousstudieshaveshownthatSTIMPY(STIP)/WOX9arerequiredformaintainingcelldivisionandpreventingprematuredifferentiationinemergingseedlings.CompletelossofSTIP activity results in early embryonic arrest,mostlikelyduetoafailureincelldivision.STIMPY-LIKE(STPL)/WOX8 share redundant functions with amorerelatedmemberoftheWOXgenesfamilysuchasWOX2, in regulating embryonic apical patterning
(Kieffer et al., 2006). These findings show thatcombinatorial action of WOX transcription factorsis essential for embryonic development.Mutation inLATERAL ORGAN BOUNDARIES (LOB) leadsto alterations in size and shape of leaves and floralorgans and causes male and female sterility (Shuaietal.,2002).Characterizationofmutantslobsuggestsa potential role for this gene in organ separation orlateral organ development. Other genes requiredfor shoot apical meristem formation are defined byALTEREDMERISTEMPROGRAM1(AMP1)whichplaysaroleinbalancingandrestrictingthemeristem-promotingactivityofauxin signaling.Mutantsamp1showeddiversemorphologicalabnormalitiesincludingsupernumerary cotyledons (Vidaurre et al., 2007).Characterization ofamp1 andmpmutants suggestedthatAMP1andMONOPTEROS(MP)antagonisticallyregulate embryo and meristem development inArabidopsis.
Figure 1. Diagram of interactions between the regulators involved in cotyledons and shoot meristem formationdevelopment—Schéma d’interactions entre des régulateurs impliqués dans la formation et le développement de cotylédons et de méristèmes de la tige.
Activationisindicatedbyanarrowandrepressionbyabluntedline—L’activation est indiquée par une flèche et la répression par une ligne émoussée;BDL:BODENLOS;CLV:CALVATA;CAN:CORONA;HAN:HANABATARANU;CUC:CUP-SHAPEDCOTYLEDON;CK:CYTOKININ;GA:GIBBERELLIN;GK:GURKE;KAN:KANADI;LTP1:LIPIDTRANSFERPROTEIN1;LOB:LATERALORGANBOUNDARIES;MP:MONOPTEROS;PHB:PHABULOSA;PAS:PASTICCINO;PIN:PIN-FORMED;PHV:PHAVOLUTA;REV:REVOLUTA;STM:SOOTMERISTEMLESS;SYD:SPLAYED;TPL:TOPLESS;TPR1:TOPLESS-RELATED1;ULT1:ULTRAPETALA1;WOS:WUS-RELATEDHOMEOBOX;WUS:WUSCHEL;YAB:YABBY;ZLL:YAB.

670 Biotechnol. Agron. Soc. Environ. 201014(4),667-680 AbidG.,JacqueminJ.-M.,SassiK.etal.
To sustain the post-embryonic developmentof the shoot, it is crucial that the balance betweenthe indeterminacy of the SAMand the determinategrowth of lateral organs is maintained. Thetranscription factor of the homeobox-LEUCINEZIPPER (HD-ZIP) proteins such as PHABULOSA(PHB), PHAVOLUTA (PHV), and REVOLUTA(REV) play critical roles in this process (Priggeet al., 2005). It is likely that PHB, PHV, andREVregulate embryonic SAM formation by positivelyregulating STM expression.Mutants phb, phv, andrev display strongly reduced auxin polar transportto inflorescence, stems and hypocotyls. Thisalterationresultsindisruptedcelldifferentiationandmorphology.A fundamental aspectof the transitionphase of embryogenesis, when apical and basalorgan fates are established, is the repression ofroot-promoting gene cascades in the apical embryodomain, which is provided by the activity of theTOPLESS (TPL) and TOPLESS-RELATED (TPR)proteins,co-repressorswhichfunctiontogetherwithAUX/IAAproteinsinauxin-mediatedtranscriptionalrepression(Szemenyeietal.,2008).
2.2. the central and basal domains of the embryo
TogetherthecentralandbasaldomainsoftheembryocontributetoRAMformation.Defectsinspecificationsignal in both regions disrupt organization ofprovascularcells.BothauxinandethylenehormoneexpressintheRAM.Auxinplaysaconspicuousrolein the specification and maintenance of stem celland meristematic activity, while ethylene has beendemonstrated to participate in the regulation of thecelldivisionactivityof theQuiescentCenter (QC),a group of non dividing cells,whichmaintains theundifferentiatedstateof thesurroundingstemcells.Alteration of the ethylene pathway by genetic orchemicaltoolsaffectsthedivisionactivityoftheQCsuggestingitsfunctionsinmaintenanceofstemcellnichebyregulatingthebalancebetweenproliferationandquiescenceofstemcells(Ortega-Martinezetal.,2007).ItisnowclearthatactivityofsomeregulatorfactorsandnormalauxinsignallingarerequiredforQCidentityandthusformeristemmaintenanceandrootgrowth(Figure 2).StudiesofgenesinvolvedinauxinresponseandembryoproperdevelopmentsuchasBODENLOS(BDL),MONOPTEROS(MP),andGNOM (GN), suggest that early root developmentdepends on these gene activities. GN proteins areinvolved in the specificationof apical-basal patternformationandwererecentlyconsideredasessentialfor cell division, expansion, and adhesion (Robertet al., 2008).Mutantgn embryos showed a loss ofapical-basalpolarity andalteration in radial pattern
that displays abnormal intracellular traffic oftranscriptionregulators(Mayeretal.,1993).Mutantsbdlandmparedisruptedatthetwo-cellstage,whentheapicalcelldivideshorizontallyrathervertically.ThesedatasuggestthatMPandBDLareimplicatedin auxin-mediated apical-basal patterning process.The cells of the central domain divide abnormallyandthehypophysisfailstoproducethelens-shapedcell, resulting in seedlings devoid of the root, therootmeristem,andthehypocotyls.MPandBDLareexpressedonlyinembryoproperandareco-expressedintheearlyglobularembryo;theycaninteracttoformheterodimers(Hardtkeetal.,2004).IncontrasttoMPandBDL,thefirstdefectsinAUXINRESISTANT6(AXR6)embryoareaberrantlyorientedcelldivisionsin the derivatives of the basal daughter cell of thezygote; subsequently, no root meristem is formedand axr6 mutant seedlings are arrested in theirdevelopment soon after germination (Hobbie et al.,2000).Mutantsmp,bdl,andaxr6interferetovaryingdegreeswith the formation of the embryo axis andarerecognizedbylessorienteddivisionsinthebasaldomain. Contrarily to mutant mp gene, mutationsinHOBBIT (HBT) gene affect the development ofthebasal,butnotthecentralregion,resultinginthelackofafunctionalrootmeristem,andthereforerootformation.Incontrasttompseedlings,hbtseedlingsinitiate but do not develop adventitious roots fromtheir hypocotyls because they are unable to formnewrootmeristem.Itisclearthatrootdevelopmentand organogenesis require not only transcriptionalhormonalsignalingnetworkbutalsokeycomponentsregulators such as sterols which are the majorcomponentsmodulatingthefluidityandpermeabilityof eukaryotic cell membranes and in additionserve as precursor for steroid-signaling moleculesthat are known to elicit a variety of physiologicalprocesses (Kepinski, 2006). Functional roles forthe campesterol-derived brassinosteroids (BRs)are established for integrating light signals in theregulationofpostembryonicplantgrowth.AnalysesofFACKEL(FK)activities indicate importantrolesof sterols for proper embryodevelopment.Mutantsfk provoke failure of cells in the central embryodomain fated to form vascular primordium. Laterduring development, mutant embryos also showabnormalities in the apical and basal domains andoftenformmultipleshootmeristemsandcotyledons(Schrick et al., 2000). Phenotypic abnormality ischaracterized by seedlings inwhich the hypocotylsismissing and by cotyledons attached directly to ashort root.Thesedatasuggest thatFKgenehas thespatially restricted task of hypocotyls specificationcontrarily to MP and BDL genes characterized bymutants identified with impaired capacities for theproductionofbothhypocotylsandroot.

Embryogenesisinhigherplants 671
3. GenetIc AnAlysIs oF seed MAturAtIon
Seed maturation is an important phase of seeddevelopmentduringwhichembryogrowthceasesandstorageproductsaccumulate.Thespatialandtemporalregulationofalltheseprocessesrequirestheconcertedaction of several signaling pathways that integrateinformationfromgeneticprograms,andbothhormonalandmetabolicsignals(Figure 3).
The LEAFY COTYLEDON 2 (LEC2), LEAFYCOTYLEDON 1 (LEC1), FUSCA3 (FUS3), andABSCISICACIDINSENSITIVE3(ABI3)havebeenidentified as master regulators of seed maturation(Santos Mendoza et al., 2005). Interestingly, studyofabi3, fus3, lec1,and lec2single,doubleandtriplemutantsshowthatthesedifferentregulatorsinteracttocontrol thevarious seedmaturation features.Ectopicexpression of LEC1 has demonstrated that this geneis required for the maintenance of the suspensorcell fate, the specification of cotyledon identity, the
initiationandmaintenanceofthematurationphase,thepromotionofembryoniccellidentityanddivision,andthesuppressionofgermination(Cassonetal.,2006).Moreover,seedstorageprotein(SSR)geneexpressioniscontrolledbyLEC1throughtheregulationofABI3and FUS3 expression (Kagaya et al., 2005). FUS3expressionisregulatedbyLEC2andFUS3itselfintheroottip,byLEC2andABI3intheembryoaxis,andbythefourregulators(LEC1,LEC2,ABI3,andFUS3)incotyledons.TheexpressionofABI3isalsocontrolledby the four regulatoryproteins incotyledons (SantosMendozaetal.,2005).Indeed,bothpost-transcriptionaland post-translational regulatory mechanisms arefound to control ABI3 expression. However, ABI3appears to be involved in auxin signaling as wellas lateral root development. Additionally, ectopicexpression of LEC2 suggests that auxin can inducethis gene expression.Moreover, FUS3 expression isup-regulatedbyauxin in theembryowhere it furthercoordinates ABA-GA action by feedback loopsassociatedwithAbscisicAcid(ABA)andGibberillins
Figure 2. Diagramofinteractionsbetweentheregulatorsinvolvedinrootmeristemformationanddevelopment—Schéma d’interactions entre des régulateurs impliqués dans la formation et le développement de méristème racinaire.
Activationisindicatedbyanarrowandrepressionbyabluntedline—L’activation est indiquée par une flèche et la répression par une ligne émoussée;BDL:BODENLOS;HBT:HOBBIT;MP:MONOPTEROS;PLT1:PLETHORA1;PIN:PIN-FORMED;PLT2:PLETHORA2;RAM:ROOTAPICALMERISTEM;RML1:ROOTMERISTEMLESS1;QC:QUIESCENTCENTRE;SCR:SCARECROW;SHR:SHORTROOT;WOX5:WUSCHEL-RELATEDHOMEOBOX5.

672 Biotechnol. Agron. Soc. Environ. 201014(4),667-680 AbidG.,JacqueminJ.-M.,SassiK.etal.
Figure 3.Proposedmodelofgeneticandmolecularinteractionsintheregulatorynetworkinvolvedinthecontrolofplantseeddevelopmentandmaturation—Proposition d’un modèle d’interactions génétique et moléculaire au sein du réseau de régulation impliqué dans le contrôle du développement et de la maturation des graines chez les plantes.
Activationisindicatedbyanarrowandrepressionbyabluntedline—L’activation est indiquée par une flèche et la répression par une ligne émoussée;ABA:ABSCISICACID;GA:GIBERRELLIN;LEC1:LEAFYCOTYLEDON1;LEC2:LEAFYCOTYLEDON2;FUS3:FUSCA3;ABI3:ABSCISICACIDINSENSITIVE3.
(GA)synthesis(Gazzarinietal.,2004).Furthermore,FUS3hasbeenshowntoregulateSSRgeneexpressionby two ABA-independent mechanisms that seemdifferent from that ofABI3 (Kagaya et al., 2005).Otherimportantregulatorsforseeddevelopmentand/ormaturation have been identified (Gallardo et al.,2008).PICKEL(PKL)achromatin-remodelingfactor(CHD3)actsinconcertwithGAtorepressembryonictraits during and after germination, including theexpression of LEC1, LEC2, and FUS3 in roots (Liet al., 2005). Several pkl mutants which exhibitreduced GA transport cause alteration in embryopatterning.Yamagishietal.(2005)isolatedTANMEI(TAN)which controls various aspects of both earlyandlatephasesofembryodevelopment.Analysisoftan mutants indicates that this gene plays a role inboth the morphogenesis and maturation phases ofembryogenesis.Moreover, tanmutants sharesmanycharacteristics with lec mutants, suggesting thatTANhasoverlappingfunctionswiththeLECgenes.For example, both tan-1 and lec mutations cause
desiccation intolerance, defects in storage proteinand lipid accumulation, and trichome formationon cotyledons. Together, these results indicate thatTANandLECgenes,particularlyLEC1andLEC2,interact synergistically. Moreover, TAN and LECgenesmayoperateincloselyrelatedpathwayduringembryogenesis(Yamagishietal.,2005).
4. eMbryo-deFectIve MutAnts
A vast number of genes represented by embryo-defective mutants may be required for normalembryogenesis,andtheymaybemasterregulatorsofthisprocess(table 1).Severalembryo-lethalmutantsin Arabidopsis have been identified and analyzed(McElveretal.,2001).Theseauthorsestimatedthat750genes canbe easilymutated to give an embryolethal phenotype. On the other side, Tzafrir et al.(2004)showedthat250genesareessentialtocontrolnormalphenotypeinArabidopsisseeddevelopment.

Embryogenesisinhigherplants 673
table 1.Comparativeanalysisofembryo-defectivemutantphenotypeandcorrespondinggenes—Analyse comparative d’embryons mutants et de leurs gènes correspondant.Normalstagesofembryodevelopment—étapes normales du développement de l’embryon :A:embryoattheglobularstage—embryon au stade globulaire;b:embryoattheheartstage—embryon au stade cordiforme;c:embryoatthetorpedostage—embryon au stade torpille;d:embryoatthecotyledonstage—embryon au stade cotylédonaire.
Genes Mutantphenotypes
sites ofexpression
Gene function embryonic defects of gene mutants
reference
TOPLESS(TPL)
Lossofapicalstructuresortransformationoftheapicalpoleoftheembryointoaroot.Mutantseedlingsdevelopasinglecotyledonbutdonotshowshootapicalmeristem
Apicaldomain(shootapicalmeristem),expressedatheartstage
Embryonicpatternformation
Szemenyeietal.,2008
ALTEREDMERISTEMPROGRAM1(AMP1)encodingglutamatecarboxypeptidase
Mutantsshowdiversemorphologicalabnormalitiesincludingsupernumerarycotyledons
Apicaldomainat4,8,16-cellandglobularstage
Stabilizessuspensorcellfateandrestrictscellnumberinembryosandnegativelyregulates meriste-maticactivitiesinshootsandroots
Vidaurreetal.,2007
Schlepperless(SLP)encodingchaperonin-60αsubunitprotein
slpembryoismorphologicallynormaluptoheartstagebutitsdevelopmentisslowerthanthatofnormalembryo
Upperhypocotylsregionandthecotyledons,expressedattheearlycotyledonstage
Highlightstheimportanceofthechaperonin-60αproteinforchloroplastdevelopmentandsubsequentlyfortheproperdevelopmentoftheplantembryo
Apuyaetal.,2001
FASS(FS) fassmutationsaffectcellelongationandorientationofcellwallsresultinginaseedlingthatisstronglycompressedintheapical-basalaxisandenlargedinthehypocotyl
Apicalbasalpattern
Elaborationofcellshapeandcorrectorientationofcellallsfromveryearlystagesofembryogenesistotheadultstage
Torres-Ruizetal.,1994
TITAN6(TTN6)encodingisopepti-saseTclassofthedeubiquitinatingenzymesthatrecycle polyubi-quitinchainsfollowingproteindegradation
Mutationresultsinembryoniclethality
Earlystageofembryodevelopment
Unknown Tzafriretal.,2004
A b c d
A b c d
A b c d
A b c d
A
A b c d

674 Biotechnol. Agron. Soc. Environ. 201014(4),667-680 AbidG.,JacqueminJ.-M.,SassiK.etal.
table 1 (continued 2).Comparativeanalysisofembryo-defectivemutantphenotypeandcorrespondinggenes—Analyse comparative d’embryons mutants et de leurs gènes correspondant.Genes Mutant
phenotypessites ofexpression
Gene function embryonic defects of gene mutants
reference
Suspensor(sus) Mutantsshowanabnormalproliferationofthesuspensorcellsresultingfromadefectinthetransitionfromtheglobulartotheheartstageoftheembryoproper
Earlystageofembryodevelopment
Promotingnormalmorphogenesisandmaintainingthesuspensorcellidentity
Zhangetal.,1997
LEAFYCOTYLEDON1(LEC1),encodingaproteinrelatedtoatranscriptionfactorsubunitoftheHAP3genefamily
LEC1 mutationcauses defects insuspensormorphologyinearly-stageembryo;mutantembryosareintolerantofdesiccationandshowdefectsintheexpressionofsome,butnotallmaturationspecificgenes
LEC1isactiveduringbothearlyandlateseeddevelopment
Inducingboththemorphogenesisandmaturationphasesofembryogenesis
Mendozaetal.,2005
LEAFYCOTYLEDON2(LEC2)encodingaproteinwithsimilaritytotheB3domainoftheABI3/VP1transcriptionfactorfamily
Mutantseedsarelesssensitivetodesiccationandonlyslightlyaffectedintheaccumulationofstoragecompounds
Expressedduringearlyembryodevelopmentandoccasionallydetectedinvegetativetissues
Initiateandcontrolseedmaturation
Mendozaetal.,2005
FUSCA(FUS3)encodingaproteinwithsimilaritytotheB3domainoftheABI3/VP1transcriptionfactorfamily
Mutationinfus3generesultsinalterationincotyledonidentity,inabilitytocompletelateseedmaturationprocesses,andprematureactivationofapicalandrootembryonicmeristem
Expressedintheprotodermofglobularandheartstageembryo
Requiredforcorrectspecificationoffuturecotyledonparenchymacells
Mendozaetal.,2005
GURK(GK)encodingACC1,anacetyl-CoAcarboxylasewhichcatalyzesmalonyl-CoAsynthesis
Gkmutationscauseapicaldefectsinseedlingsandalsoreducedtheefficiencyofseedgerminationandseedlinggrowth
Earlystageofembryodevelopment
Involvedintheorganizationofapicalregionintheembryoandalsoapossibleroleduringpostembryonicdevelopment
Torres-Ruizetal.,1996
A b c d
A b c d
A b c d
A b c d
A b c d
Embryogenesisinhigherplants 675
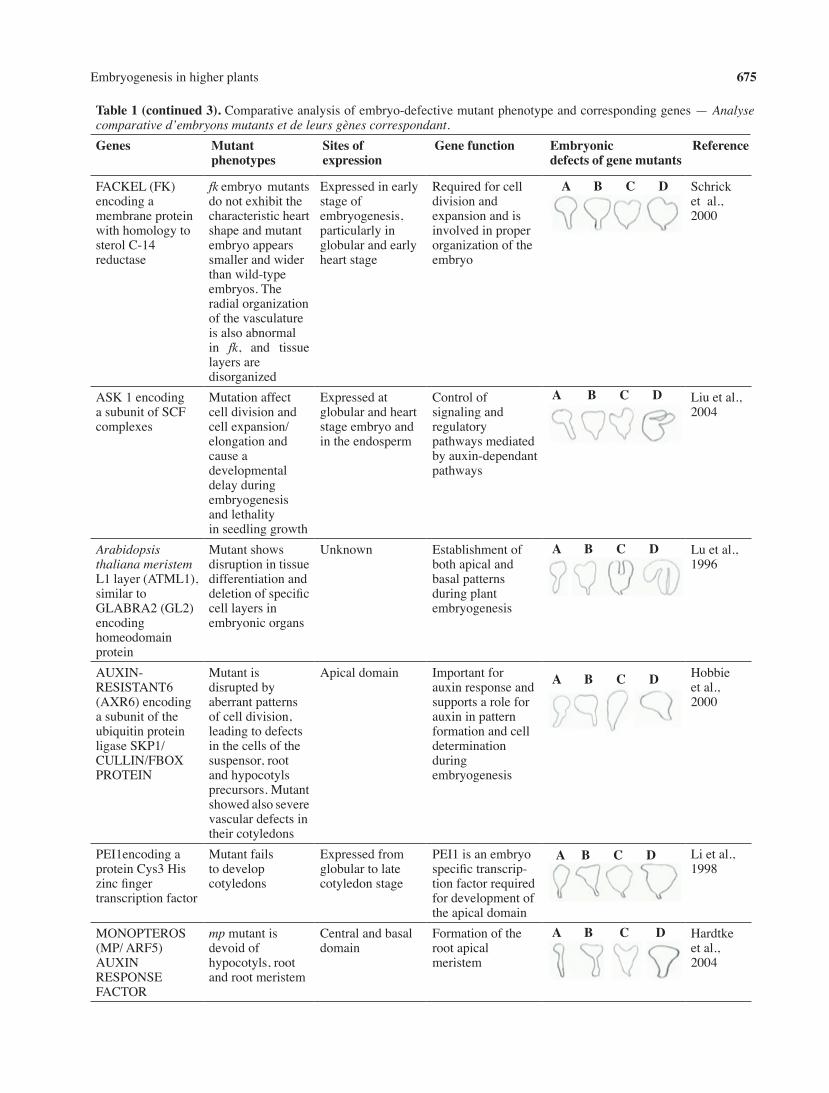
table 1 (continued 3).Comparativeanalysisofembryo-defectivemutantphenotypeandcorrespondinggenes—Analyse comparative d’embryons mutants et de leurs gènes correspondant.Genes Mutant
phenotypessites ofexpression
Gene function embryonic defects of gene mutants
reference
FACKEL(FK)encodingamembraneproteinwithhomologytosterolC-14reductase
fkembryomutantsdonotexhibitthecharacteristicheartshapeandmutantembryoappearssmallerandwiderthanwild-typeembryos.Theradialorganizationofthevasculatureisalsoabnormalin fk, and tissuelayersaredisorganized
Expressedinearlystageofembryogenesis,particularlyinglobularandearlyheartstage
Requiredforcelldivisionandexpansionandisinvolvedinproperorganizationoftheembryo
Schricketal.,2000
ASK1encodingasubunitofSCFcomplexes
Mutationaffectcelldivisionandcellexpansion/elongationandcauseadevelopmentaldelayduringembryogenesisandlethalityinseedlinggrowth
Expressedatglobularandheartstageembryoandintheendosperm
Controlofsignalingandregulatorypathwaysmediatedbyauxin-dependantpathways
Liuetal.,2004
Arabidopsisthaliana meristem L1layer(ATML1),similartoGLABRA2(GL2)encodinghomeodomainprotein
Mutantshowsdisruptionintissuedifferentiationanddeletionofspecificcelllayersinembryonicorgans
Unknown Establishmentofbothapicalandbasalpatternsduringplantembryogenesis
Luetal.,1996
AUXIN-RESISTANT6(AXR6)encodingasubunitoftheubiquitinproteinligaseSKP1/CULLIN/FBOXPROTEIN
Mutantisdisruptedbyaberrantpatternsofcelldivision,leadingtodefectsinthecellsofthesuspensor,rootandhypocotylsprecursors.Mutantshowedalsoseverevasculardefectsintheircotyledons
Apicaldomain Importantforauxinresponseandsupportsaroleforauxininpatternformationandcelldeterminationduringembryogenesis
Hobbieetal.,2000
PEI1encodingaproteinCys3Hiszincfingertranscriptionfactor
Mutantfailstodevelopcotyledons
Expressedfromglobulartolatecotyledonstage
PEI1isanembryospecifictranscrip-tionfactorrequiredfordevelopmentoftheapicaldomain
Lietal.,1998
MONOPTEROS(MP/ARF5)AUXINRESPONSEFACTOR
mpmutantisdevoidofhypocotyls,rootandrootmeristem
Centralandbasaldomain
Formationoftherootapicalmeristem
Hardtkeetal.,2004
A b c d
A b c d
A b c d
A b c d
A b c d
A b c d

676 Biotechnol. Agron. Soc. Environ. 201014(4),667-680 AbidG.,JacqueminJ.-M.,SassiK.etal.
Mutant study showed that disruption of thenormally regular patterns of cell division in theArabidopsis embryo does not necessarily interferewithpropercellfatedetermination.Thesedatasuggestthat precise cell division is not required for patternformation. Moreover, several embryo mutants showgeneticdisruptionsforpolarauxintransportandauxinresponse.
Mutations inFASS (FS)geneproducedisorderedpatterns of cell division. The fass mutants showinability to form microtubule preprophase bands,whichpredictthesiteoffuturecellwalldepositioninplantcells(Torres-Ruizetal.,1994).Howeverthefullrangeofappropriatecelltypesispresentandcompleteinthefassmutants,albeitdisorderedandseedlingsareproduced.These data suggest that lineage-dependentsegregation of cell fate determinants does not playa crucial role in patterning. Other mutants, such asKnolle(Kn),andKeule(Keu)alsodisruptcelldivisionand shape.At the cellular level,mutant embryos arecharacterizedbyincompletecrosswallsandenlargedcellswithpolyploidynuclei.KNOLLE(KN)encodesamemberofthesyntaxinfamily,aclassofmembrane-bound receptor required for docking and fusion ofvesiclesatthetargetmembrane(Lukowitzetal.,1996),whereasKEULE(KEU)geneencodesamemberoftheSec1proteinfamilywhichregulatesvesicletraffickingand interacts with syntaxins (Assaad et al., 2001).Additional mutation in HINKEL gene, a Kinesin-like protein apparently involved in reorganization ofphragmoplast microtubules (Strompen et al., 2002),show similar embryo-lethal phenotypes as KN andKEUgenes.
TheArabidopsisKIESEL (KIS) encodesTubilin-FoldingCofactors (TFCs) involved in the control oftheα/β-tubilinmonomer balancewhich is importantduringmicrotubulebiogenesis.Inplant,microtubuleshaveanimportantfunctioninchromosomeseparation,cell division, and the establishment of cell polarity.Mutationsinkisaffectthebiogenesisofmicrotubulesandresultinlethalembryoswithoneorfewenlargedcellsandfewmicrotubules(Kiriketal.,2002).
Several genes regulate a wide range of cellularprocesses in eukaryotes and theirmutations result insevere cell division defects during endosperm andembryodevelopment.TheTITAN(TTN)genesencodechromosome scaffold proteins of the condensingand cohesion classes named STRUCTURALMAINTENANCE OF CHROMOSOME (SMC)proteins (condensins and cohesions) required forchromosome function at mitosis. Development ofttn1 and ttn2 embryos is arrested at 1-2 cell stage,whereas ttn3canproducefertilehomozygousmutantplants. Embryo development is stopped shortly afterfertilizationinmost ttnmutantsandinsomecases isaccompaniedbydramaticcellenlargement.Moreover,embryo growth is arrested in several mutants withdefectsinembryoandendospermatdifferentstagesofembryodevelopment(Liuetal.,1998).
Disruption in RPN1 gene encodes subunit of the26Sproteasome(consideredasnegativeregulatorsofauxin signaling for ubiquitin-mediatedproteolyse) inArabidopsis and provokes embryo lethality (Brukhinetal.,2005).Theregulatoryparticle(RP)consistsof17subunits,whichformtwosubparticles,thelidandthebase.Thebasecontainsthreenon-ATPasesubunits
A b c d
A b c d
table 1 (continued 4).Comparativeanalysisofembryo-defectivemutantphenotypeandcorrespondinggenes—Analyse comparative d’embryons mutants et de leurs gènes correspondant.Genes Mutant
phenotypessites ofexpression
Gene function embryonic defects of gene mutants
reference
GNOM(GN)encodingmembraneassociatedADPribosylationGTPexchangefactor
gn mutantshowslossofapicalbasalpolarityanddisorganizedgrowth
Earlystageofembryodevelopment
Requiredforstablemaintenanceofplantcellpolarityingeneral
Mayeretal.,1993
STIMPY-LIKE(STPL/WOX8)ahomeodomaintranscription
stpl1mutantfailstoelongatealongtheapical-basalaxis,andembryoshowsreducedshootmeristem.Abnormaldivisionpatternsseeninthesuspensorregion
Expressedearlyinfirstzygoticdivisioninbothapicalandbasalcellsandofembryosatglobularandheartstages
Requiredformaintainingcelldivisioninboththeembryoandsuspensorandalsoinpost-embryonicproliferatingtissues
Kiefferetal.,2006

Embryogenesisinhigherplants 677
(RPN1,RPN2,andRPN10)andsixATPasesubunits(RPT1 to RPT6), which contact the α-subunit ringand likely assist in substrate unfolding and transport(Brukhin et al., 2005).The lidbinds to thebase andcontainstheotherninesubunits(RPN3,RPN5toRPN9,andRPN11toRPN13).Embryosofrpn1mutantsstoptheirgrowthat theglobular stagewithdefects in theformationof the embryonic root, theprotoderm, andprocambium.Mutant phenotypes analyses show thatRPN12a subunit ofArabidopsis thaliana participatesin cytokinin responses (Smalle et al., 2002). Thesedata suggest the indirectmechanism of auxin actioncontrastingwiththedirecteffectsofcytokininonbudgrowth;thereisgoodevidencethatauxincanregulatecytokininbiosynthesis.
The ARP7 gene encodes actin-related proteinsinvolved in chromatin dynamics and transcriptionalregulation.Asubstantialknockdownofitsexpressionseverely affects plant architecture and considerablydelays the abscission of floral organs.Mutants arp7produce morphologically aberrant embryos that arearrestedatorbeforethetorpedostageofdevelopment(Kandasamy et al., 2005). Genes that affect thefunctionand/ordevelopmentoforganellesmayalsobeplaying similarly significant roles in embryogenesis.A mutation in EDD1 encodes a plastedic form ofglycyl-tRNAsynthase (Uwer et al., 1998), and leadsto embryo arrest between globular and heart stages.Ontheotherside,embryomutantsSchlepperless(slp),a plastid chaperonin-60α subunit protein involved inchloroplastdevelopment,showlesioninthechloroplastanddelayofembryodevelopmentbeforeheartstage.The embryonic cotyledons of thismutant are highlyreduced(Apuyaetal.,2001).
ArrestofembryogrowthaccompaniedbyabnormalmitoticdivisioniscommonlyobservedinabortedseedsofsomemutantssuchasTROMOZ(TOZ).Indeed,tozmutation affects cell division during embryogenesisand mutant embryos stop at globular stage (Griffithet al., 2007). Mutations severely disrupt cytokinesis(specific phases lead to cytoplasm partitions of adividingcellandsynthesisofanewplasmamembrane)during embryo and endosperm formation (Sorensenetal.,2002).ThesedatasuggestthatTOZaffectsearlyembryonicdevelopment.
In Arabidopsis thaliana, genes that regulatesuspensordevelopmenthavebeenisolatedbystudiesofmutants,andthefunctionsofthesegeneshavebeencharacterized (Zhang et al., 1997). Indeed in somemutants, suspensorceases togrowatdifferentstagesofembryodevelopment;thisarrestisaccompaniedbyabnormalactivityofthesuspensor.
TWN2andTWN1areconsideredasgenesinvolvednotonlyformaintainingsuspensoridentitybutalsoforregulatingorganogenesisintheembryoapex(Vernonetal.,2002).Mutanttwn2showdegeneratesatanearly
stage; however suspensor cells initiate embryonicdevelopmentandeventuallyformoneormoreembryo.Mutanttwn1showsdefectsbothinthesuspensorandthe cotyledons (Vernon et al., 2002). Similarly, theabnormalraspberry1(rsy1)andraspberry2(rsy2)arecharacterizedbyamorphologicallyabnormalembryothat fails to form axis and cotyledons, and have anenlargedsuspensor(Yadegarietal.,1994).RSY3geneislocalizedinchloroplasts,andithasbeensuggestedthatRSY3proteinisrequiredforchloroplastdifferentiationand embryonic development (Apuya et al., 2002).The raspberrymutant rsy3 shows abnormality in theembryoproperandisarrestedintheglobularembryostageofdevelopment.Thesedatasuggestthattwnandrsymutations are affected components of the apical-basalcellsignalingpathway.
Mutants with abnormal suspensors have beenidentified and explored in part through the analysisofembryo-defectivemutantsinArabidopsis thaliana.Mutantsuspensors(sus1,sus2,andsus3)arebothlargerand wider than normal suspensors and the embryo-properconsistentlyisarrestedatthepreglobularstageofdevelopment.AccordingtoZhangetal.(1997),asrsyandsusmutationscausethearrestoftheembryo-proper,itcouldbearguedthattheabnormalsuspensordevelopmentinthesemutantsresultsintheabsenceofaviableembryo-proper.
5. conclusIon And perspectIves
Embryogenesisrepresentsakeystepofplantlifecycleduring which embryo develops the main structuresof the future individual according to a well-definedmorphologicalpattern.
During embryo development, different post-transcriptional regulation mechanisms appear to berequired to control the genetic programs induced ateachstage.
Although histological, morphological, molecular,and biochemical studies have provided descriptiveinformation about embryogenesis, a mechanisticunderstandingoftheprocessesthatunderlieembryonicdevelopmentislacking.
Important clues about the process involvedin embryo development may be obtained fromanalysesofembryonicmutant.Theidentificationandcharacterization of embryo-defective mutants havebeenshowntobepotentiallyusefulindissectingeventsinembryodevelopmentasageneticapproach.
Differential gene expression is a commonly usedapproach for interpreting gene expression profilesduringembryodevelopment.Manygenesinvolvedindiverseprocessesofembryodevelopmenthavealreadybeencharacterizedthroughawiderangeoftechniquessuchasdifferentialscreeningapproachesormutational

678 Biotechnol. Agron. Soc. Environ. 201014(4),667-680 AbidG.,JacqueminJ.-M.,SassiK.etal.
screening.Seeddevelopmentproceedsthroughaseriesofspatiallyandtemporallyregulatedgeneexpressionsteps.Elucidationofgeneexpressionpatternsassociatedwithaspecificstageofseeddevelopmentiscriticalforunderstanding themolecularandbiochemicalevents,whichmaybecharacteristicofthatstage.
Recently,verysensitivetechniquesadaptedtothestudyof transcriptionalactivitiesofscarcebiologicaltissues and allowing genome-wide gene expressionanalysis, such as in silico expressed sequence tags(EST) analysis, SAGE (Serial Analysis of GeneExpression) or microarrays technology, have beendeveloped (Kasai et al., 2005). These techniquesgivetheopportunitytoobtainglobalgeneexpressionpatternsduring seeddevelopment infloweringplant,particularly inArabidopsis thaliana and a few otherwell-characterized plant species such as Helianthus annuus L., Medicago truncatula L., and Brassica napus L.(Whiteetal.,2000).
More recently novel factors affecting plantembryogenesis have been identified (Spartz et al.,2008); cross-linking between phytohormone andtranscription factors is likely to display a part ofembryogenesis. However, the mechanism of plantembryogenesis remains partially unclear. Particularlyresults of epigenetic approaches suggest that DNAmethylation and chromatin modification are alsoimportant factors for plant embryogenesis (Xiaoetal.,2006).Futurestudieswillclarifyandhighlightthe interaction of these factors, thereby revealingthe entire embryogenesis regulatory mechanism.Therefore,holisticapproachessuchastheintegrationof genomics, proteomics, and metabolic, from theperspectiveofsystemsbiology,haveagreatpotentialinrevealingtheinteractionbetweendifferentsignalingcascades involved in triggering embryogenesis infloweringplant.
bibliography
ApuyaN.R. et al., 2001.TheArabidopsis embryomutantSchlepperlesshasadefectintheChaperonin-60αgene.Plant Physiol.,126,717-730.
ApuyaN.R.etal.,2002.RASPBERRAY3geneencodesanovelproteinimportantforembryodevelopment.Plant Physiol.,129,691-705.
AssaadF.F., HuetY., MayerU. & JurgenG., 2001. Thecytokenesis geneKEULE encodes a Sec1 protein thatbindsthesyntaxinKNOLLE.J. Cell Biol.,152,531-543.
BaudS.etal.,2004.Gurkeandpasticcino3mutantsaffectedin embryo development are impaired in acetyl-CoAcarboxylase.EMBO Reports,5,515-520.
BellecY. et al., 2002. Pasticcino2 is a protein tyrosinephosphatase-like involved in cell proliferation anddifferentiationinArabidopsis. Plant J.,32,713-722.
BrukhinV. et al., 2005. The RPN1 subnit of the 26SproteasomeinArabidopsisisessentialforembryogenesis.Plant Cell,17,2723-2737.
CassonS.A. & LindseyK., 2006. The turnip mutant ofArabidopsis reveals that LEAFY COTYLEDON1expressionmediates the effectsof auxin and sugars topromote embryonic cell identity. Plant Physiol., 142,526-541.
DeYoungB.J.&ClarksE.,2008.BAMreceptors regulatestemcell specificationandorgandevelopment throughcomplexinteractionwithCLAVATAsignaling.Genetics,180,895-904.
FaureJ.D. et al., 1998. The PASTICCINO genes ofArabidopsis thalianaareinvolvedinthecontrolofcelldivision and differentiation. Development, 125, 909-918.
FujitaT.etal.,2008.Convergentevolutionofshootinlandplants: lack of auxin polar transport in moss shoots.Evol. Dev.,10,176-186.
GallardoK., ThompsonR. & BurstinJ., 2008. Reserveaccumulationinlegumeseeds.C.R. Biol.,331,755-762.
GazzarriniS.etal.,2004.ThetranscriptionfactorFUSCA3controls developmental timing inArabidopsis throughthehormonesgibberellinsandabscisicacid.Dev. Cell,7,373-385.
GriffithM.E. et al., 2007.TheTORMOZ gene encodes anucleolarproteinrequiredforregulateddivisionplanesandembryodevelopmentinArabidopsis.Plant Cell,19,2246-2263.
HaeckerA. et al., 2003. Expression dynamics of WOXgenesmark cell fate decisionsduring early embryonicpatterning inArabidopsis thaliana.Development,131,657-668.
HardtkeC.S. et al., 2004.Overlapping andnon-redundantfunctions of the Arabidopsis auxin response factorsMONOPTEROS and NONPHOTOTROPICHYPOCOTYLA.Development,131,1089-1100.
HobbieL. et al., 2000. The axr6 mutants of Arabidopsis thaliana defineagene involved in auxin responseandearlydevelopment.Development,127,23-32.
KagayaY. et al., 2005. LEAFY COTYLEDON1 controlsseed storage protein genes through its regulation ofFUSCA3andABSCISICACIDINSENSITIVE3.Plant Cell Physiol.,46,399-406.
KandasamyM.K., McKinneyE.C., DealR.B. &MeagherR.B.,2005.ArabidopsisARP7isanessentialactin-relatedproteinrequiredfornormalembryogenesis,plant architecture, and floral organ abscission. Plant Physiol.,138,2019-2032.
KasaiY.etal.,2005.5’SAGE:5’-endserialanalysisofgeneexpressiondatabase.Nucleic Acids Res.,33,550-552.
KepinskiS., 2006. Integrating hormone signaling andpatterning mechanisms in plant development. Curr. Opin. Plant Biol.,9,28-34.
KiefferM.etal.,2006.AnalysisofthetranscriptionfactorWUSCHELanditsfunctionalhomologueinAntirrhinum

Embryogenesisinhigherplants 679
revealsapotentialmechanismfortheirrolesinmeristemmaintenance.Plant Cell,18,560-573.
KirikV.etal.,2002.TheArabidopsisTUBULIN-FOLDINGCOFACTORgeneisinvolvedinthecontroloftheα/β-tubulinmonomerbalance.Plant Cell,14,2265-2276.
LauxT., WürschumT. & BreuningerH., 2004. Geneticregulationofembryonicformation.Plant Cell,16,190-202.
LenhardM.,JürgenG.&LauxT.,2002.TheWUSCHELandSHOUTMERISTEMLESS gene fulfill complementaryroles in Arabidopsis shoot meristem regulation.Development,129,3195-3206.
LiH.C. et al., 2005. PIKEL acts during germination torepress expression of embryonic traits. Plant J., 44,1010-1022.
LiZ.&ThomasT.L.,1998.PEI1,anembryo-specificzincfinger protein gene required for heart-stage embryoformationinArabidopsis.Plant Cell,10,383-398.
LiuC.M. & MeinkeD.W., 1998. The titan mutants ofArabidopsis are disrupted in mitosis and cell cyclecontrolduringseeddevelopment.Plant J.,16,21-31.
LiuF.etal.,2004.TheASK1andASK2genesareessentialforArabidopsisearlydevelopment.Plant Cell,16,5-20.
LongJ.A.&BartonM.K.,1998.Thedevelopmentofapicalembryonic pattern in Arabidopsis. Development, 125,3027-3035.
LuP., PoratR., NadeauJ.A. & O’NeillS.D., 1996.Identification of a meristem L1 layer-specific gene inArabidopsisthatisexpressedduringembryonicpatternformationanddefinesanewclassofhomeoboxgenes.Plant Cell,8,2155-2168.
LukowitzW.,MayerU.&JurgensG.,1996.Cytokinesisinthe Arabidopsis embryo involves the syntaxin-relatedKNOLLEgeneproduct.Cell,84,61-71.
MayerU., BüttnerG. & JürgensG., 1993. Apical basalpatternformationintheArabidopsisembryo:studiesontheroleofthegnomgene.Development,117,149-162.
McElverJ. et al., 2001. Insertional mutagenesis of generequiredforseeddevelopmentinArabidopsisthaliana.Genetic,159,1751-1763.
MendozaM. et al., 2005. LEAFY COTYLEDON 2activationissufficienttotriggertheaccumulationofoilandseedspecificmRNAsinArabidopsisleaves.FEBS Letter,579,4666-4670.
MoussainB.etal.,1998.RoleoftheZWILLEgeneintheregulation of central shoot meristem cell fate duringArabidopsisembryogenesis.EMBRO J.,17,1799-1809.
Ortega-MartenezO., PernasM., CarolR.J. & DolanL.,2007. Ethylen modulates stem cell division in theArabidopsis thalianaroot.Science,317,507-510.
PriggeM.J. et al., 2005. Class III homeodomain-leucinezipper gene family members have overlappingantagonistic, and distinct roles in Arabidopsisdevelopment.Plant Cell,17,61-76.
RobertS.etal.,2008.Endosidin1definesacompartementinvolved inendocytosisof thebrassinosteroid receptor
BRI1andtheauxintransportersPIN2andAUX1.Proc. Natl Acad. Sci. USA.,105,8464-8469.
SchrickK.etal.,2000.FACKELisasterolC-14reductaserequired for organized cell division and expansion inArabidopsis embryogenesis. Genes Dev., 14, 1471-1484.
ShuaiB., Reynaga-PenaC.G. & SpringerP.S., 2002. Thelateral organ boundaries gene defines a novel, plant-specificgenefamily.Plant Physiol.,129,747-761.
SmalleJ. et al., 2002. Cytokinin growth responses inArabidopsis involve 26S proteasome subunit RPN12.Plant Cell,14,17-32.
SorensenM.B.etal.,2002.CellularisationintheendospermofArabidopsis thalianaiscoupledtomitosisandsharesmultiple components with cytokinesis. Development,129,5567-5576.
SpartzA.K.&GrayW.M.,2008.Planthormonereceptors:newperception.Genes Dev.,22,2139-2148.
StrompenG. et al., 2002.TheArabidopsisHINKELgeneencodesakinesin-relatedproteininvolvedincytokinesisandisexpressedinacellcycle-dependentmanner.Curr. Biol.,12,153-158.
SuhM.C. et al., 2003.Comparative analysis of expressedsequence tags Sesamum indicum and Arabidopsis thaliana developing seeds.Plant Mol. Biol.,52, 1107-1123.
SzemenyeiH., HannonM.& LongJ.A., 2008. TOPLESSmediates auxin-dependent transcriptional repressionduringArabidopsisembryogenesis.Science,319,1384-1386.
TakadaS., HibaraK., IshibaT. & TassakaM., 2001. TheCUP-SHAPED COTYLEDON 1 gene ofArabidopsisregulatesshootapicalmeristemformation.Development,128,1127-1135.
TorrezRuizR.A.&JürgensG.,1994.MutationintheFASSgene uncouple pattern formation and morphogenesisinArabidopsis development.Development,120, 2967-2978.
Torres-RuizR.A., LohnerA. & JurgensG., 1996. TheGURKEgeneisrequiredfornormalorganizationoftheapical region of theArabidopsis embryo.Plant J.,10,1005-1016.
TzafrirI. et al., 2004. Identification of gene required forembryo development in Arabidopsis. Plant Physiol.,135,1206-1220.
UmeharaM., IkedaM. & KamadaH., 2007. Endogenousfactors that regulate plant embryogenesis: recentadvances.Jpn. J. Plant Sci.,1,1-6.
UwerU.,WillmitzerL.&AltmannT.,1998.Inactivationofglycyl-tRNAsynthaseleadstoanarrestinplantembryodevelopment.Plant Cell,10,1277-1294.
VernonD.M.,HannonM.J.,LeM.&ForsthoefelN.R.,2002.AnexpandedrolefortheTWN1geneinembryogenesis:defectsincotyledonpatternandmorphologyinthetwn1mutantofArabidopsis(Brassicaceae).Annu. J. Bot.,88,570-582.

680 Biotechnol. Agron. Soc. Environ. 201014(4),667-680 AbidG.,JacqueminJ.-M.,SassiK.etal.
VidaurreD.P.,PloenseS.,KroganN.T.&BerlethT.,2007.AMP1 and MP antagonistically regulate embryo andmeristem development in Arabidopsis. Development,134,2561-2567.
WhiteJ.A.etal.,2000.AnewsetofArabidopsisexpressedsequence tags from developing seeds. The metabolicpathwayfromcarbohydratestoseedsoil.Plant Physiol.,124,1582-1594.
XiaoW. et al., 2006. DNA methylation is critical forArabidopsis embryogenesis and seed viability. Plant Cell,18,805-814.
YadegariR. et al., 1994. Cell differentiation andmorphogenesisareuncoupledinArabidopsisraspberryembryo.Plant Cell,6,1713-1729.
YamagishiK. et al., 2005.TANMEI/EMB2757 encodes aWDrepeatproteinrequiredforembryodevelopmentinArabidopsis.Plant Physiol.,139,163-173.
ZhangJ.Z. & SomervilleC.R., 1997. Suspensor-derivedpolyembryony caused by altered expression of valyl-tRNA synthetase in the twn2 mutant of Arabidopsis.Dev. Biol.,94,7349-7355.
(59ref.)





























