Embed Size (px)
Citation preview

1
New recombinant BCG expression vectors: improving genetic control over 1
mycobacterial promoters 2
3
4
Alex I. Kannoab, Cibelly Goulartab, Henrique K. Rofattoab, Sergio C. Oliveirac, Luciana 5
C. C. Leitea, Johnjoe McFaddend# 6
7
Centro de Biotecnologia, Instituto Butantan, São Paulo, SP, Brazila; Programa de Pós 8
Graduação Interunidades em Biotecnologia USP – IPT – IB, São Paulo, Brazilb, 9
Departamento de Bioquímica e Imunologia, Universidade Federal de Minas Gerais, Belo 10
Horizonte, MG, Brazilc; Department of Microbial and Cellular Sciences, Faculty of 11
Health and Medical Sciences, University of Surrey, Guildford, Surrey, United Kingdomd. 12
13
Running Head: New recombinant BCG expression vectors 14
15
16
17
18
19
20
# Address correspondence to Johnjoe McFadden, [email protected] 21
AEM Accepted Manuscript Posted Online 5 February 2016Appl. Environ. Microbiol. doi:10.1128/AEM.03677-15Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
Abstract 22
23
The expression of many antigens, stimulatory molecules or even metabolic pathways in 24
mycobacteria such as Mycobacterium bovis BCG or M. smegmatis was made possible 25
through the development of shuttle-vectors and several recombinant vaccines have been 26
constructed. However, gene expression in any of these systems relied mostly on selection 27
of natural promoters expected to provide the required level of expression by trial and 28
error. To establish a systematic selection of promoters with a range of strengths, we 29
generated a library of mutagenized promoters through error-prone PCR of the strong PL5 30
promoter, originally from the mycobacteriophage L5. These promoters were cloned 31
upstream of the enhanced green fluorescent protein (eGFP) gene reporter and 32
recombinant M. smegmatis were identified exhibiting a wide range of fluorescence. A set 33
of promoters was selected and identified as high (pJK-F8), intermediate (pJK-B7, pJK-34
E6, pJK-D6) and low (pJK-C1) promoter strengths in both M. smegmatis and M. bovis 35
BCG. The sequencing of the promoter region demonstrated it was extensively modified 36
(6-11%) in all selected plasmids. To test the functionality of the system two different 37
expression vectors were demonstrated to allow corresponding expression levels of the 38
Schistosoma mansoni antigen, Sm29, in BCG. The approach conducted here can be 39
useful for adjusting expression levels for synthetic and/or systems biology studies or for 40
vaccine development to maximize the immune response. 41
42
Keywords: Recombinant BCG, promoters, random mutagenesis, vaccine. 43
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
Introduction 44
45
BCG is currently the world’s most widely used vaccine and has been given to 46
more than three billion people, making it a very attractive prospect for development of a 47
live recombinant BCG (rBCG) multivaccine (1). The first generation of rBCG vaccines 48
was developed in the 1990’s as rBCG strains that expressed homologous and 49
heterologous antigens from a wide range of pathogens (2-5). Stability of the heterologous 50
(or native) gene(s) in BCG was usually obtained by cloning it on a plasmid or 51
chromosomally integrative vector with expression achieved by placing it under the 52
control of a range of mycobacterial promoters (2). The heat shock protein promoters 53
(PHSP60 from BCG and PHSP70 from M. tuberculosis), the IS900 ORF promoter (PAN from 54
M. paratuberculosis) and the mutated β-lactamase promoter (PBlaF* from M. fortuitum) 55
are the most used ones, as reviewed by (2). However, the lack of knowledge on the 56
transcriptional mechanisms regulating mycobacterial promoters, results in unpredictable 57
gene expression levels. Yet level of antigen expression is likely to be a crucial factor in 58
both the strength and the pattern of subsequent immune responses. For example, a study 59
in which expression of the M. tuberculosis Antigen 85B (Ag85B) in rBCG was placed 60
under the control of a limited set of promoters, found that increasing promoter activity 61
caused a skewing of the immune response to Ag85B in mice from a mixed Th1/Th2 to a 62
predominantly Th1 response (6). In another study, increasing expression of M. 63
tuberculosis 19 kDa lipoprotein was found to lead to a complete abrogation of the 64
protective efficacy of BCG by polarizing the host immune responses to the Th2 subtype 65
(7). Another important factor for any vaccine vector is stability and the associated 66
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
metabolic burden, which can lead to loss of antigen expression and/or premature 67
elimination of the vector in the host due to loss of fitness (8). Indeed, loss of vector 68
during replication in the host is thought to have been responsible for the failure of a 69
rBCG vaccine expressing the outer surface protein A (OspA) of Borrelia burgdorferi to 70
generate an appropriate immune response in a clinical trial, despite the apparent efficacy 71
of the vaccine in mice (9). Our own earlier studies similarly demonstrated vector 72
instability and a lack of correlation of immune response in mice with expression levels in 73
vitro (10). 74
To improve the genetic control on gene expression in mycobacteria, different 75
tools and strategies can be used, such as inducible promoters (11), synthetic genes with 76
codon optimized sequences (12, 13), substitution of the initiation codon (14), addition of 77
Shine-Dalgarno sequences (15), increase in plasmid copy number (16) or integration into 78
the mycobacterial genome (17). However, once again, expression levels are often 79
unpredictable and may be further complicated by regulatory influences on expression 80
levels, particularly when growing in the host environment. In this work, as a first step 81
towards a more rational approach to vaccine vector design we engineered the 82
mycobacteriophage promoter PL5 (18). Our aim was to generate a set of mycobacteria 83
promoters that can be used to obtain a predictable range of gene expression levels in both 84
M. smegmatis and M. bovis BCG. 85
86
Materials and Methods 87
88
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
Ethics Statement. This study was conducted in accordance with the recommendations and 89
approval by the Committee of Ethical in Animal Experiments of the Butantan Institute 90
under the protocol 594/09. 91
Bacterial strains and plasmids. E. coli DH5α (19) (Invitrogen) was used in all cloning 92
steps using lysogeny broth (LB) or LB plates with kanamycin (20 µg/mL) for selection of 93
transformants. M. smegmatis MC2 155 strain (20) and M. bovis BCG strain Pasteur (21) 94
were grown at 370C in Difco Middlebrook 7H9 broth (Becton, Dickinson) enriched with 95
10% (vol/vol) oleic acid-albumin-dextrose-catalase (OADC), 0.5% glycerol and 0.05% 96
Tween 80 (MB7H9) containing kanamycin when necessary. Eletrocompetent M. smegmatis 97
and BCG were transformed by electroporation as previously described (22) and 98
transformants selected in Middlebrook 7H10 agar plates with OADC (MB7H10) and 99
kanamycin. All primers were purchased from Eurofins MWG Operon (Ebersberg, 100
Germany) and all restriction enzymes from New England Biolabs (Hitchin, UK). GoTaq 101
DNA polymerase (Promega, Southampton, UK) was used in all PCR reactions. The 102
expression cassette of the pBRL8 plasmid (kindly provided by Dr. William Jacobs Jr., 103
Yeshiva University, NY) containing the PL5 promoter and egfp gene was PCR amplified 104
using primers 5’-TAGGGTACCTCTAGAGGAAACAGCTATGACCAT-3’ (Kpn I and 105
Xba I restriction sites underlined and in bold) and 5’-106
TAGATCGATCAGCTGTTACTTGTACAGCTCGT-3’ (Cla I and Pvu II restriction sites 107
underlined and in bold). This fragment was cloned into pJH152 (kindly provided by Dr. 108
Stewart T. Cole, Ecolé Polytechnique Fédérale de Lausanne, France) between restriction 109
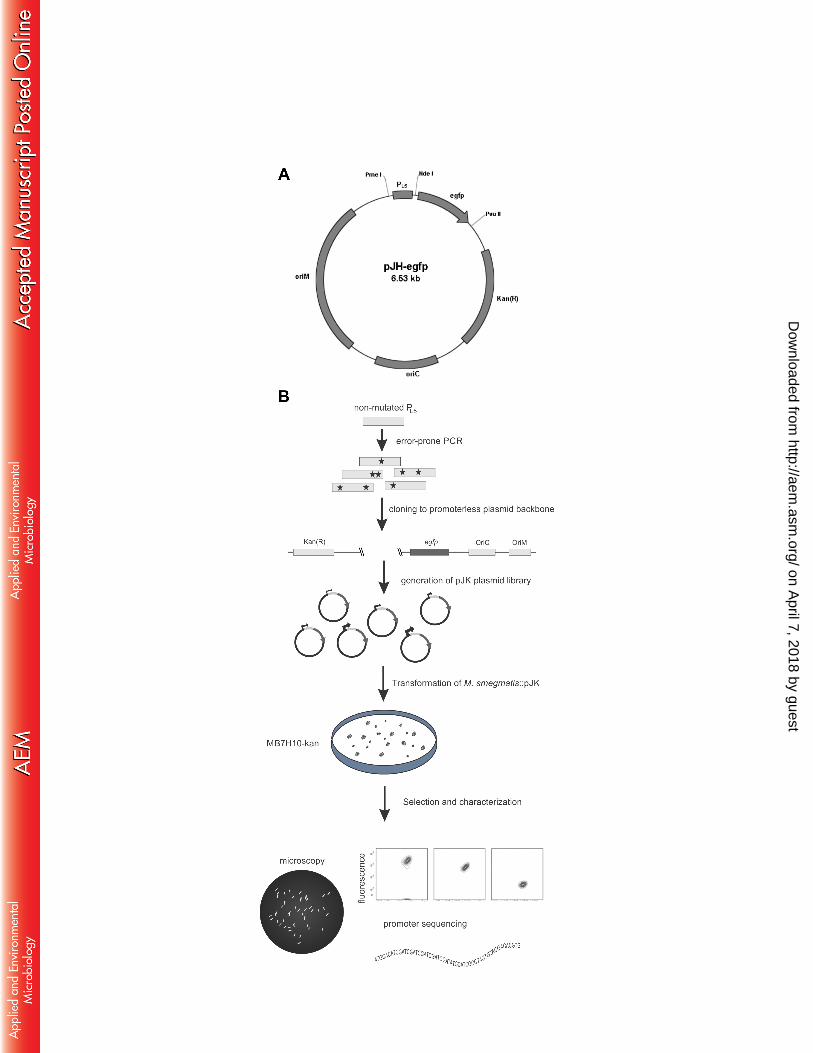
sites Kpn I and Pvu II, thus generating pJH-egfp (Figure 1A). Next, error-prone PCR of the 110
promoter sequence was carried out in the presence of 25 µM of 8-oxo-dGTP and 5 µM of 111
dPTP (Jena Bioscience, Germany), along with 400 µM of natural dNTPs (Figure 1B). 112
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
According to manufacturer’s instructions, 20 rounds of amplification were performed using 113
primers 5’-TAGGTTTAAACAAACGGAAACAGCTATGACCAT-3’ (Pme I restriction 114
site in bold) and 5’-TAGCATATGCGATCTCCCTTTCCCGT-3’ (Nde I restriction site in 115
bold). Plasmid pJH-egfp and the PCR product were digested with Pme I and Nde I, resolved 116
in an agarose gel electrophoresis to remove the original cassette and clean up the PCR 117
reaction, and further purified using QIAquick PCR Purification Kit (Qiagen, Hilden, 118
Germany). Before ligation, the digested vector was treated with CIAP (Promega), 119
according to the manufacturer instructions for the dephosphorylation of 5’ overhangs and 120
purified once again. Digested fragments were allowed to ligate using T4 DNA ligase 121
(Promega) at 16°C for 16 h and used to transform E. coli DH5α. Plasmid DNA from 122
approximately 2,000 colonies was extracted in pool generating the pJK plasmid library, 123
which in turn was used in the electroporation of M. smegmatis MC2 155 using a BioRad 124
Gene Pulser II apparatus (Hercules, USA). Transformants were allowed to grow for 3-5 125
days in MB7H10-kanamycin plates (Figure 1B). The plasmid pJH137 was digested using 126
Xba I and Nde I restriction enzymes to remove the heat shock promoter PHsp60 and insert in 127
place of PL5 at pJH-egfp generating pJHsp60. Plasmid pET-21 containing the Sm29 128
sequence to produce rSm29 protein was previously constructed (23). 129
Screening fluorescent M. smegmatis. After eye-visualization of M. smegmatis 130
transformants, approximately 200 colonies were isolated and grown in MB7H9-kanamycin. 131
Samples were allowed to grow for 16 h and screened in 96-well black plates in duplicates 132
using a Perkin Elmer Victor 3 fluorescence multiplate reader (Waltham, USA) setup to 485 133
nm/535 nm (excitation/emission). Also, the O.D.620 of each sample was measured. The 134
fluorescence of the strains was initially screened by fluorescence units (F.U.) and then, 135
following normalization by O.D., 5 strains were selected based on measurements of relative 136
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
fluorescence units (R.F.U.). The plasmid in each of these transformants was extracted using 137
illustra plasmidPrep Mini Spin kit (GE Healthcare Life Sciences, Freiburg, Germany) and 138
PL5 promoters sequenced using the primer 5’-TCAGCTTGCCGTAGGTGGCA-3’. 139
Characterization of GFP expression and in vitro growth of selected mutants. Shaking 140
flasks with MB7H9-kanamycin were inoculated with selected M. smegmatis::pJK in 141
triplicates to an O.D. of ~ 0.1. Culture turbidity and fluorescence were monitored every 1 h. 142
For comparison, the wild-type M. smegmatis mc2 155 strain grown in MB7H9 was also 143
analyzed. A FACSCanto II flow cytometer (Becton, Dickinson) was used to determine the 144
single-cell fluorescence of gated bacterial cells based on the median of FITC (using 488 145
excitation/500-560 emission filter). Turbidity was measured using a Ultrospec 2000 146
spectrophotometer (Pharmacia Biotech, Sweden). To compare the newly generated 147
promoters with a widely used one the PHsp60 was inserted in place of PL5. M. bovis BCG 148
containing these plasmids were analyzed and compared with M. smegmatis constructs. Late 149
log-phase cultures of recombinant M. smegmatis strains were disrupted on ice using a GE 150
100 Ultrasonic Processor at half-maximum constant output and protein concentration 151
determined in culture lysates by the DC Protein Assay (Biorad) using bovine serum 152
albumin as standard. Approximately 10 µg of soluble protein extracts were separated by 153
SDS-PAGE and stained with Coomassie Brilliant Blue (Biorad). The predicted molecular 154
weight of GFP is 28.3 kDa (http://web.expasy.org/compute_pi/). Following destaining the 155
band corresponding to GFP’s molecular weight was analyzed by densitometry using 156
ImageJ software. 157
Microscopy analysis of selected strains. M. smegmatis::pJK strains were observed under a 158
microscope. Late log-phase cultures were washed twice with PBS and placed on 159
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
microscope slides. Images were acquired on a LSM-510 META confocal system (Zeiss, 160
Germany) using 488 nm laser for excitation and BP500-550 filter for emission. 161
Fine-tuned expression of foreign antigen. To assess the usefulness of the pJK library 162
regardless of the downstream gene under its regulation, we used three plasmids containing 163
the mutagenized promoters, pJK-F8, pJK-B7 and pJK-C1. For this model, we removed the 164
egfp gene by digestion with Nde I and Pvu II and cloned the partial sequence of the 165
Schistosoma mansoni gene, sm29 (GenBank accession no. AF029222) codon-optimized for 166
expression in mycobacteria, as this antigen is currently considered a vaccine candidate (23), 167
creating pJK-F8.sm29, pJK-B7.sm29 and pJK-C1.sm29. The codon optimization was 168
performed by the DNA 2.0 company (CA, USA) according to their patented algorithm. 169
Codon-optimized sm29 had an increased GC content (from 37% to 51%). BCG was 170
transformed with these constructs and allowed to grow as described previously. 171
Kanamycin-resistant clones were grown until late-log phase, disrupted on ice and 172
quantified as previously described. Approximately 25 µg of total protein extracts were 173
separated by SDS-PAGE, proteins electrotransferred to a PVDF membrane (GE 174
Healthcare) and blocked for 16 h at 4oC in a 5% nonfat dry milk solution. Western blotting 175
analysis of Sm29 expression used polyclonal mouse anti-rSm29 antibody (1:1,000) and 176
horseradish peroxidase (HRP)-conjugated anti-mouse antibody adsorbed for human serum 177
proteins (KPL, Maryland, USA) (1:3,000). The chemiluminescent signal was detected 178
using the Immobilon Western Chemiluminescent HRP Substrate (Merck, Darmstadt, 179
Germany) and images acquired using the LAS4000 digital imaging system (GE). 180
Recombinant Sm29 protein and anti-rSm29 antibody production. Recombinant Sm29 181
was expressed in E. coli BL21 DE3 (Invitrogen) transformed with pET-21 containing the 182
Sm29 cDNA sequence and purified using metal affinity chromatography as previously 183
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
described (24). To generate the primary antibody, 3 BALB/c mice were immunized 184
subcutaneously with 3 doses of rSm29 (25 µg) adsorbed in aluminum hydroxide (250 µg) 185
with a 2 weeks interval between doses. Fifteen days after the last dose a cardiac puncture of 186
anesthetized mice was performed for blood collection and serum processing. 187
Statistical analysis. For statistical analysis, the software GraphPad Prism 5 (La Jolla, CA, 188
USA) was used. The relationship between pJH-egfp and pJK library in the initial screening 189
used Two-tailed Spearman’s Rank Correlation. The fluorescence induced by pJK in M. 190
smegmatis or BCG was analyzed by one-way ANOVA and Tukey’s post-test. Correlation 191
of interspecies fluorescence was tested using a non-parametric Spearman test. The growth 192
curves were compared using non-linear regression. 193
194
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
Results 195
196
Generation of the PL5 promoter library. First we used error-prone PCR (25) to generate a 197
promoter library of PL5 and assess variant sequences in their strengths by cloning the 198
promoters to drive expression of the enhanced green fluorescent protein, eGFP (Figure 1B). 199
The wild-type PL5 promoter and egfp gene were cloned into pJH152 generating pJH-egfp 200
(Figure 1A), which was used to transform M. smegmatis, thus representing the fluorescence 201
induced by the original promoter, which correlated with bacterial growth (Figure 2A). The 202
pJK plasmid library was used to transform M. smegmatis and the approximately 200 203
colonies named pJK-XA1 to -YH12 were screened by fluorimetry and demonstrated a wide 204
variation in fluorescence, ranging between 103 and 106 fluorescence units (F.U.) (Figure 205
2B). 206
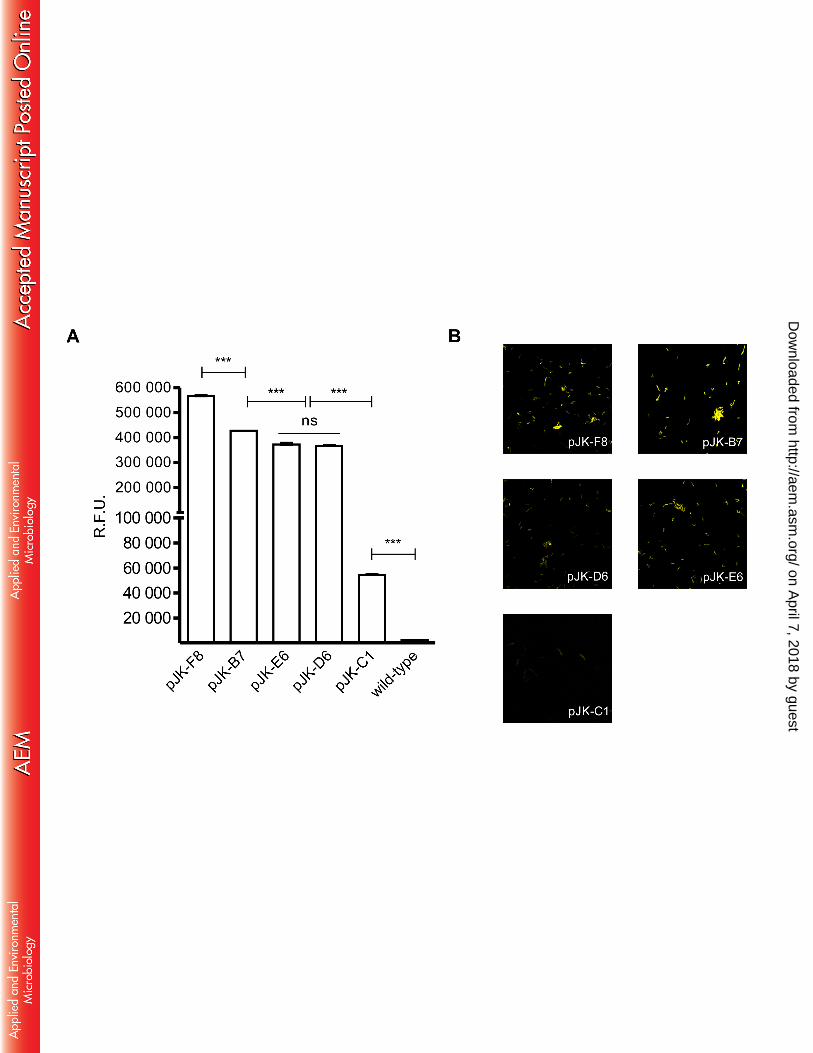
Characterization of selected M. smegmatis::pJK. Based on the initial screening, 5 strains 207
were selected covering the fluorescence range for further analysis. First, the sequencing of 208
selected promoters revealed multiple mutations with modification in 6 to 11% of their 209
sequence (Figure 3). Since the O.D. of M. smegmatis::pJK library mutants varied between 210
0.3 to 1.5 the data was normalized and fluorescence plotted by relative units (R.F.U.) 211
(Figure 4A). Likewise, confocal microscopy confirmed the differential expression of GFP 212
where the fluorescence was distributed throughout the bacillus cell (Figure 4B). To exclude 213
the possibility that the differential fluorescence would be a consequence of impaired growth 214
or a distinct rate of GFP expression, selected M. smegmatis::pJK were evaluated for single-215
cell fluorescence during growth. Constant fluorescence is observed during growth in each 216
mutant strain (not shown). The M. smegmatis::pJK-C1 was the only one demonstrated to be 217
significantly different from the growth of the wild-type strain (Supplementary Figure 1). 218
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
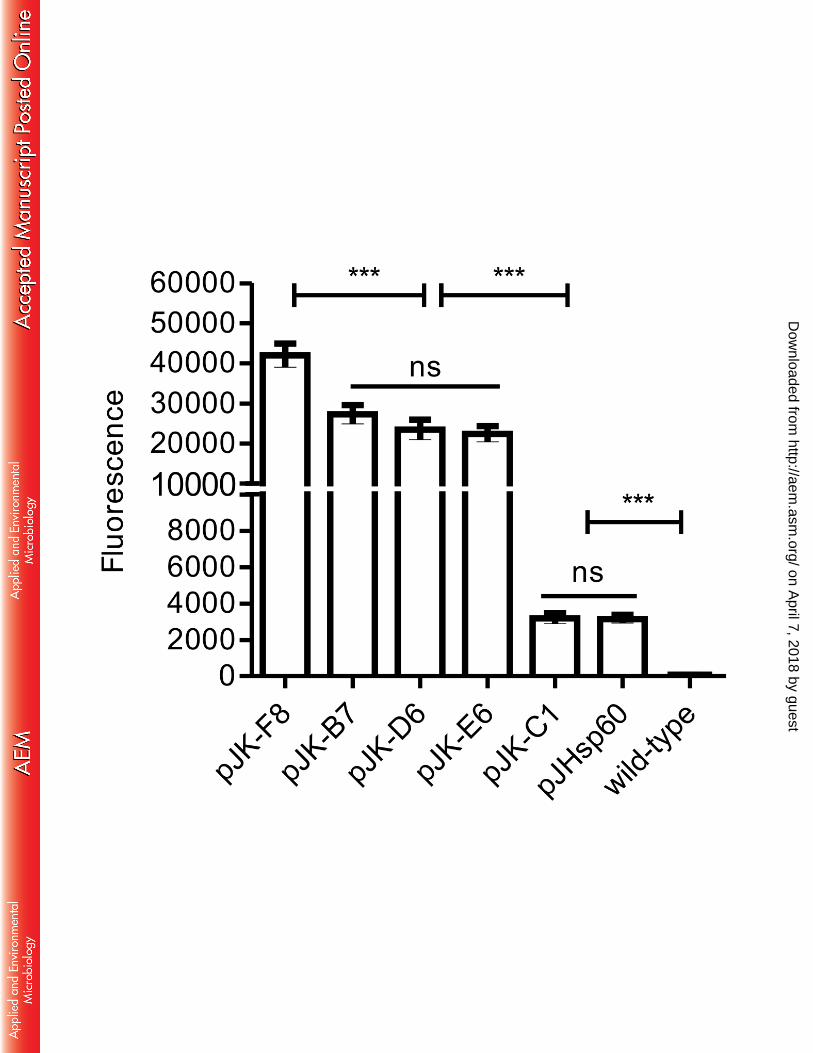
Flow cytometry of M. smegmatis and M. bovis BCG::pJK strains. The selected pJK 219
plasmids were transformed into M. bovis BCG and late-log phase cultures analyzed 220
alongside recombinant M. smegmatis by flow cytometry. After gating the bacilli’s 221
population, fluorescence was determined by the median of FITC value. M. smegmatis 222
transformed with pJK-F8 showed high fluorescence; pJK-B7, pJK-E6 and pJK-D6 were 223
intermediate and pJK-C1 was strikingly low (Figure 5A). Recombinant M. smegmatis 224
strains were also submitted to SDS-PAGE and the bands corresponding to GFP were 225
quantified by densitometry (Figure 5B). The results demonstrated that the fluorescence 226
intensity measured by flow cytometry correlated with the amount of protein produced (r 227
value = 0.96) according to the strength of each promoter. BCG transformed with the same 228
plasmids revealed a high correlation with the fluorescence observed for M. smegmatis (r 229
value = 0.94) (not shown) where all mutants maintained their referred high, intermediate or 230
low strength in both species. pJHsp60 demonstrated a strength comparable to the lowest 231
expression vector pJK-C1 (Figure 6). 232
Mutant promoters induce differential antigen expression. To investigate the usefulness 233
of these promoters for the expression of other heterologous genes, we replaced the egfp 234
gene from pJK-F8, pJK-B7 and pJK-C1 with the codon optimized sm29 gene from S. 235
mansoni. Since, pJK-F8 produced no colonies after transformation, further work was 236
carried on using pJK-B7 and pJK-C1. Recombinant BCG clones of these strains were 237
grown to late-log phase and lysed by sonication. Total protein extracts were analyzed by 238
Western blot using specific anti-rSm29 antibodies. As predicted, we observed a more 239
intense immunoblot in the rBCG::pJK-B7.Sm29 extracts (higher promoter activity) as 240
compared to the reaction in the rBCG::pJK-C1.Sm29 extracts (lower promoter activity) at 241
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
the same molecular weight as the recombinant protein rSm29, but no immunoreaction is 242
observed in the wild type BCG protein extracts (Figure 7). 243
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
Discussion 244
245
Here we used the enhanced GFP (eGFP) as gene reporter to measure the differential 246
expression of this protein induced by mutations in the regulatory region of the 247
mycobacteriophage L5 PL5 promoter sequence. The PL5 promoter regulatory sequence was 248
subjected to random mutagenesis and a library of promoters exhibiting different strengths 249
measured by fluorescence was selected. The GFP has been one of the most used gene 250
reporters to characterize promoter strength among many organisms in a quantitative fashion 251
(26-28), including in mycobacterial species for promoter discovery, strength and trafficking 252
in the host (11, 29-32). 253
The observed 6-11% of mutations in the regulatory sequence is consistent with the 254
expected rate of 10-15% mutagenesis using equimolar concentrations of altered and natural 255
dNTPs (25). A higher degree of mutations can span a wider range in downstream 256
expression; however, it can also abolish the expression by altering important motifs for 257
transcriptional factor binding. During the in vitro growth the selected M. smegmatis::pJK 258
mutants showed a stable level of fluorescence. The PHsp60 promoter placed in the same 259
plasmid backbone as the pJK series induced a fluorescence intensity comparable to the 260
weakest PL5 promoter, thus indicating the high strength of PL5. A heat-shock treatment did 261
not increase the fluorescence of M. smegmatis::pJK or pJHsp60 (not shown). It could be 262
argued that fluorescence may not serve as a good parameter to determine protein expression 263
and ultimately promoter activity. However, here we have demonstrated that the 264
fluorescence determined by flow cytometry correlates with the total amount of GFP 265
expression in SDS-PAGE even though only 6 promoters were used. 266
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
In this study, we observed mutations throughout the whole sequence, including the -267
10 and -35 regions for the promoters, exhibiting high, medium or low strength. 268
Interestingly, all mid-strength promoters (pJK-B7, -E6 and -D6) showed a mutation at the 269
first position of the -10 region (C → T). Also, the weakest promoter that we identified, in 270
pJK-C1, is the only one with a mutation at the predicted transcriptional starting site (A → 271
G). 272
Heterologous expression in recombinant bacteria can be deleterious or lethal. Given 273
the mycobacterial relatively slow growth rate, where faster-growing strains like M. 274
smegmatis and M. fortuitum have a doubling-time of 2-3 hours and M. bovis BCG and M. 275
tuberculosis 20-24 hours (33), the eGFP synthesis undoubtedly represents a metabolic 276
burden and stress through consumption of ATP, nucleotides, aminoacids and usage of 277
ribosomes and tRNAs that may displace the expression of endogenous proteins and hence 278
inhibit the bacillus’s growth. GFP itself can also be toxic to the host (26). Nevertheless, in 279
this study, the only strain significantly different from the growth of the wild-type was M. 280
smegmatis::pJK-C1. This strain appears to have a lag in early time points but reaching a 281
similar O.D. at later time points. However, it is unlikely that a low and not a high 282
expression of eGFP could inhibit growth as observed by others (34). 283
To evaluate the utility of PL5 promoters for driving foreign antigen expression in 284
BCG, the sm29 gene from S. mansoni was cloned in place of the egfp gene downstream of a 285
strong (pJK-F8), an intermediate (pJK-B7) and a weak (pJK-C1) mutant promoter. 286
Recombinant BCG expressed Sm29 at different levels, which corresponded to their eGFP-287
measured promoter strength. pJK-F8 was not evaluated since no colonies of rBCG were 288
obtained even after several attempts. 289
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
rBCG::pJK-B7 clones were 9-fold more fluorescent than rBCG::pJK-C1 clones 290
(average FITC value of 27,300 and 3,200, respectively). To achieve such a difference, 291
Bourn used an enrichment method to select for high-copy number variants of pORI101 292
plasmid which were able to express GFP with a 7-fold increase compared to the original 293
(16). Although this strategy can be interesting to obtain a significant expression of cloned 294
genes, some disadvantages such as metabolic burden and plasmid instability may reduce 295
the usefulness of the system. 296
Since pJK plasmids are extrachromossomal copy number could be partially 297
responsible for the differences observed in fluorescence or Sm29 expression among 298
mycobacterial hosts. The origin of replication of pJH-egfp and pJK is derived from 299
pAL5000 of M. fortuitum, which is widely used in mycobacterial shuttle-plasmids such as 300
pMV261, pLA71 and pMIP12. Its copy number in M. smegmatis was determined as three 301
by Southern Blot (35), eight by antibiotic resistance (36) and twenty three by quantitative 302
PCR (37). The variation in the copy number between these studies can be explained by the 303
different methods of quantification, but also by the differences in plasmid structure. Despite 304
using the same pAL5000-derived origin of replication, other features such as resistance 305
marker, size and some other elements are also different. In our study all features are same, 306
with exception of a few nucleotides at the promoter sequence. Thus, we expect to minimize 307
the effect of plasmid copy number in the differential expression. Besides that, although 308
there are minor variations in the fluorescence during growth of recombinant M. smegmatis 309
their fluorescence values do not intersect with each other. 310
In conclusion, this study demonstrates the utility of the rational design of systems to 311
obtain a range of expression of target proteins in mycobacteria. The systems described 312
present an alternative approach to obtain expression of foreign genes in mycobacteria and 313
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

16
to design a new generation of recombinant BCG vaccines that optimize the host immune 314
response towards protection. 315
316
Acknowledgments 317
318
This work was supported by FAPESP grant 2008/04631-7, Santander through its program 319
Santander Universities and Fundação Butantan. 320
321
We thank S.T. Cole for the generous gift of pJH152 and G.F. Hatfull for pBRL8. 322
323
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

17
References 324
325
1. Bloom BR, Snapper SB, Kieser T, Jacobs WR. 1990. Development of 326 recombinant BCG vaccines. Seminars in Virology 1:7. 327
2. Bastos RG, Borsuk S, Seixas FK, Dellagostin OA. 2009. Recombinant 328 Mycobacterium bovis BCG. Vaccine 27:6495-6503. 329
3. Fuerst TR, de la Cruz VF, Bansal GP, Stover CK. 1992. Development and 330 analysis of recombinant BCG vector systems. AIDS Res Hum Retroviruses 331 8:1451-1455. 332
4. Ohara N, Yamada T. 2001. Recombinant BCG vaccines. Vaccine 19:4089-333 4098. 334
5. Dale JW, Dellagostin OA, Norman E, Barret ADT, McFadden J. 1993. 335 Multivalent BCG vaccines. In D. T. O'Hagan e (ed), Novel delivery systems for 336 oral vaccines. CRC Press, Boca Raton. 337
6. Dhar N, Rao V, Tyagi AK. 2004. Immunogenicity of recombinant BCG vaccine 338 strains overexpressing components of the antigen 85 complex of Mycobacterium 339 tuberculosis. Med Microbiol Immunol 193:19-25. 340
7. Dhar N, Rao V, Tyagi AK. 2003. Skewing of the Th1/Th2 responses in mice due 341 to variation in the level of expression of an antigen in a recombinant BCG system. 342 Immunol Lett 88:175-184. 343
8. Galen JE, Levine MM. 2001. Can a 'flawless' live vector vaccine strain be 344 engineered? Trends Microbiol 9:372-376. 345
9. Edelman R, Palmer K, Russ KG, Secrest HP, Becker JA, Bodison SA, Perry 346 JG, Sills AR, Barbour AG, Luke CJ, Hanson MS, Stover CK, Burlein JE, 347 Bansal GP, Connor EM, Koenig S. 1999. Safety and immunogenicity of 348 recombinant Bacille Calmette-Guerin (rBCG) expressing Borrelia burgdorferi 349 outer surface protein A (OspA) lipoprotein in adult volunteers: a candidate Lyme 350 disease vaccine. Vaccine 17:904-914. 351
10. Varaldo PB, Leite LC, Dias WO, Miyaji EN, Torres FI, Gebara VC, Armoa 352 GR, Campos AS, Matos DC, Winter N, Gicquel B, Vilar MM, McFadden J, 353 Almeida MS, Tendler M, McIntosh D. 2004. Recombinant Mycobacterium 354 bovis BCG expressing the Sm14 antigen of Schistosoma mansoni protects mice 355 from cercarial challenge. Infect Immun 72:3336-3343. 356
11. Ehrt S, Guo XV, Hickey CM, Ryou M, Monteleone M, Riley LW, 357 Schnappinger D. 2005. Controlling gene expression in mycobacteria with 358 anhydrotetracycline and Tet repressor. Nucleic Acids Res 33:e21. 359
12. Kanekiyo M, Matsuo K, Hamatake M, Hamano T, Ohsu T, Matsumoto S, 360 Yamada T, Yamazaki S, Hasegawa A, Yamamoto N, Honda M. 2005. 361 Mycobacterial codon optimization enhances antigen expression and virus-specific 362 immune responses in recombinant Mycobacterium bovis bacille Calmette-Guerin 363 expressing human immunodeficiency virus type 1 Gag. J Virol 79:8716-8723. 364
13. Varaldo PB, Miyaji EN, Vilar MM, Campos AS, Dias WO, Armoa GR, 365 Tendler M, Leite LC, McIntosh D. 2006. Mycobacterial codon optimization of 366 the gene encoding the Sm14 antigen of Schistosoma mansoni in recombinant 367
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

18
Mycobacterium bovis Bacille Calmette-Guerin enhances protein expression but 368 not protection against cercarial challenge in mice. FEMS Immunol Med 369 Microbiol 48:132-139. 370
14. Fan XY, Ma H, Guo J, Li ZM, Cheng ZH, Guo SQ, Zhao GP. 2009. A novel 371 differential expression system for gene modulation in Mycobacteria. Plasmid 372 61:39-46. 373
15. Andreu N, Zelmer A, Fletcher T, Elkington PT, Ward TH, Ripoll J, Parish 374 T, Bancroft GJ, Schaible U, Robertson BD, Wiles S. 2010. Optimisation of 375 bioluminescent reporters for use with mycobacteria. PLoS One 5:e10777. 376
16. Bourn WR, Jansen Y, Stutz H, Warren RM, Williamson AL, van Helden PD. 377 2007. Creation and characterisation of a high-copy-number version of the 378 pAL5000 mycobacterial replicon. Tuberculosis (Edinb) 87:481-488. 379
17. Hatfull GF, Sarkis GJ. 1993. DNA sequence, structure and gene expression of 380 mycobacteriophage L5: a phage system for mycobacterial genetics. Mol 381 Microbiol 7:395-405. 382
18. Nesbit CE, Levin ME, Donnelly-Wu MK, Hatfull GF. 1995. Transcriptional 383 regulation of repressor synthesis in mycobacteriophage L5. Mol Microbiol 384 17:1045-1056. 385
19. Taylor RG, Walker DC, Mcinnes RR. 1993. Escherichia-Coli Host Strains 386 Significantly Affect the Quality of Small-Scale Plasmid DNA Preparations Used 387 for Sequencing. Nucleic Acids Res 21:1677-1678. 388
20. Snapper SB, Melton RE, Mustafa S, Kieser T, Jacobs WR, Jr. 1990. Isolation 389 and characterization of efficient plasmid transformation mutants of 390 Mycobacterium smegmatis. Mol Microbiol 4:1911-1919. 391
21. Brosch R, Gordon SV, Garnier T, Eiglmeier K, Frigui W, Valenti P, Dos 392 Santos S, Duthoy S, Lacroix C, Garcia-Pelayo C, Inwald JK, Golby P, Garcia 393 JN, Hewinson RG, Behr MA, Quail MA, Churcher C, Barrell BG, Parkhill J, 394 Cole ST. 2007. Genome plasticity of BCG and impact on vaccine efficacy. Proc 395 Natl Acad Sci U S A 104:5596-5601. 396
22. Parish T, Stoker NG. 1998. Electroporation of mycobacteria. Methods Mol Biol 397 101:129-144. 398
23. Cardoso FC, Macedo GC, Gava E, Kitten GT, Mati VL, de Melo AL, Caliari 399 MV, Almeida GT, Venancio TM, Verjovski-Almeida S, Oliveira SC. 2008. 400 Schistosoma mansoni tegument protein Sm29 is able to induce a Th1-type of 401 immune response and protection against parasite infection. PLoS Negl Trop Dis 402 2:e308. 403
24. Cardoso FC, Pacifico RN, Mortara RA, Oliveira SC. 2006. Human antibody 404 responses of patients living in endemic areas for schistosomiasis to the tegumental 405 protein Sm29 identified through genomic studies. Clin Exp Immunol 144:382-406 391. 407
25. Zaccolo M, Williams DM, Brown DM, Gherardi E. 1996. An approach to 408 random mutagenesis of DNA using mixtures of triphosphate derivatives of 409 nucleoside analogues. J Mol Biol 255:589-603. 410
26. Alper H, Fischer C, Nevoigt E, Stephanopoulos G. 2005. Tuning genetic 411 control through promoter engineering. Proc Natl Acad Sci U S A 102:12678-412 12683. 413
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

19
27. Davis JH, Rubin AJ, Sauer RT. 2011. Design, construction and characterization 414 of a set of insulated bacterial promoters. Nucleic Acids Res 39:1131-1141. 415
28. Qin JY, Zhang L, Clift KL, Hulur I, Xiang AP, Ren BZ, Lahn BT. 2010. 416 Systematic comparison of constitutive promoters and the doxycycline-inducible 417 promoter. PLoS One 5:e10611. 418
29. Gall K, Barker LP. 2006. Differential green fluorescent protein expression from 419 mycobacterial promoter constructs in Escherichia coli and Mycobacterium 420 marinum. FEMS Microbiol Lett 255:301-307. 421
30. Kremer L, Baulard A, Estaquier J, Poulain-Godefroy O, Locht C. 1995. 422 Green fluorescent protein as a new expression marker in mycobacteria. Mol 423 Microbiol 17:913-922. 424
31. Muller-Taubenberger A, Anderson KI. 2007. Recent advances using green and 425 red fluorescent protein variants. Appl Microbiol Biotechnol 77:1-12. 426
32. Triccas JA, Britton WJ, Gicquel B. 2001. Isolation of strong expression signals 427 of Mycobacterium tuberculosis. Microbiology 147:1253-1258. 428
33. Dziadek J, Madiraju MV, Rutherford SA, Atkinson MA, Rajagopalan M. 429 2002. Physiological consequences associated with overproduction of 430 Mycobacterium tuberculosis FtsZ in mycobacterial hosts. Microbiology 148:961-431 971. 432
34. Luo Y, Szilvasi A, Chen X, DeWolf WC, O'Donnell MA. 1996. A novel 433 method for monitoring Mycobacterium bovis BCG trafficking with recombinant 434 BCG expressing green fluorescent protein. Clin Diagn Lab Immunol 3:761-768. 435
35. Ranes MG, Rauzier J, Lagranderie M, Gheorghiu M, Gicquel B. 1990. 436 Functional analysis of pAL5000, a plasmid from Mycobacterium fortuitum: 437 construction of a "mini" mycobacterium-Escherichia coli shuttle vector. J 438 Bacteriol 172:2793-2797. 439
36. Gavigan JA, Ainsa JA, Perez E, Otal I, Martin C. 1997. Isolation by genetic 440 labeling of a new mycobacterial plasmid, pJAZ38, from Mycobacterium 441 fortuitum. J Bacteriol 179:4115-4122. 442
37. Huff J, Czyz A, Landick R, Niederweis M. 2010. Taking phage integration to 443 the next level as a genetic tool for mycobacteria. Gene 468:8-19. 444
445
446
447
448
449
450
451
452
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

453
Figure 1. Schematic representation of pJH-egfp and the methodology for the 454
generation of a functional promoter library. (A) pJH-egfp contains the PL5 promoter 455
upstream the egfp sequence, a kanamycin resistance marker Kan(R), and origins of 456
replication in E. coli (oriC) and mycobacteria (oriM). (B) The PL5 promoter was 457
submitted to error-prone PCR and used to drive the expression of egfp. M. smegmatis 458
transformants were selected by kanamycin resistance and functional promoters screened 459
within fluorescent colonies. Selected colonies spanning a wide range of fluorescence 460
were further analyzed by microscopy, specific promoter mutations and single cell 461
fluorescence by flow cytometry (Adapted from (26)). 462
463
464
Figure 2. Screening of PL5 promoter mutants strength. (A) Clones of M. 465
smegmatis::pJH-egfp with the non-mutated PL5 promoter show a correlation between 466
growth (O.D.620) and fluorescence (F.U.log10) (r = 0.79). (B) M. smegmatis transformed 467
with pJK plasmid library show a variation in fluorescence independently of growth (r = 468
0.26). Wild-type M. smegmatis (no egfp gene) and M. smegmatis::pJH-egfp are indicated 469
by a black triangle and a star, respectively. The mutants selected for further analysis are 470
indicated as black circles. Statistical analysis used Two-tailed Spearman’s Rank 471
Correlation. 472
473
474
20
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

21
Figure 3. Sequence alignment of wild-type and mutated PL5 promoters in pJK 475
plasmids. The promoter sequence containing the selected pJK plasmids was sequenced 476
and compared to the original. From 257 bp comprising the promoter region, there are 6 – 477
11% of altered bp within mutants, which also localized to the predicted -10 and -35 478
regions. pJK-C1 showed a mutation at the transcriptional starting site (TSS). The 479
initiation codon is the last 3 bp in the sequence showed. 480
481
482
Figure 4. The error-prone PCR change the strength of PL5 promoter. (A) Relative 483
fluorescence (R.F.U.) of selected transformants comprising the fluorescence range. (B) 484
Microscopy of M. smegmatis::pJK demonstrate the differential fluorescence as referred 485
of promoter strength with little variation among the bacilli population. Statistical analysis 486
by one-way ANOVA (*** p < 0.001, ns = not significant) 487
488
489
Figure 5. pJK plasmid series induce differential expression in M. smegmatis. Late 490
log-phase cultures of M. smegmatis were transformed with selected pJK plasmids or 491
pJHsp60. (A) Fluorescence of recombinant M. smegmatis harboring pJK plasmid series 492
analyzed by flow cytometry. (B) Relative density of the GFP band in the SDS-PAGE 493
analyzed by densitometry. Statistical analysis by one-way ANOVA (*** p < 0.001, ** p 494
< 0.01, ns = not significant). 495
496
497
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

22
Figure 6. pJK plasmids series induce differential fluorescence in M. bovis BCG. Late 498
log-phase cultures of BCG were transformed with pJK plasmid series or pJHsp60 and 499
fluorescence analyzed by flow cytometry. Statistical analysis by one-way ANOVA (*** 500
p < 0.001, ns = not significant). 501
502
503
Figure 7. Western blot of rBCG strains differentially expressing Sm29 antigen 504
through mutated PL5 promoters. Total protein extracts (25 µg) of wild-type BCG (Lane 505
2) or rBCG::pJK-C1.Sm29 (Lanes 3-4) and pJK-B7.Sm29 (Lanes 5-6). Two 506
independently-isolated clones were analyzed by Western blot using anti-rSm29 antibodies. 507
300 ng of rSm29 protein (Lane 1) was used as positive control. Molecular weight markers 508
standard are indicated on the left 509
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from