Embed Size (px)
Citation preview

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
550
FATORES QUE AFETAM A LIBERAÇÃO E A EFICIÊNCIA DE PA RASITÓIDES NO
CONTROLE BIOLÓGICO DE INSETOS-PRAGA
Jacqueline Barbosa Nascimento
I) Doutoranda em Agronomia - Fitossanidade. Escola de Agronomia e Engenharia de Alimentos, Universidade Federal de Goiás (UFG), Goiânia, Goiás. Brasil.
*E-mail: [email protected] Data de recebimento: 07/10/2011 - Data de aprovação : 14/11/2011
RESUMO
Um dos limitadores enfrentados para o desenvolvimento do programa de controle biológico com o uso de parasitóide é a falta de informações sobre a tecnologia de liberação destes inimigos naturais nas diversas regiões do Brasil. Estudos devem ser realizados nesta área, pois o material com que se trabalha é invariavelmente sensível ao meio ambiente. É sabido que a eficiência das liberações destes parasitóides no campo depende basicamente de alguns fatores, tais como a capacidade de “busca”, preferência hospedeira, tolerância às condições climáticas, intervalo, número e época de liberações, número de insetos liberados, arquitetura da planta, entre outros. Portanto, o conhecimento desses fatores e suas interações possibilitam melhor manejo do controle natural de uma praga ou conjunto delas. O objetivo deste estudo é abordar e discutir os fatores que associados a liberação dos parasitóides em campo afeta sua eficiência de parasitismo. PALAVRAS-CHAVE: parasitismo, relação planta-inseto, nível tritrófico, COV’s
FACTORS AFFECTING THE RELEASE AND EFFICIENCY OF PAR ASITOIDS IN BIOLOGICAL CONTROL OF INSECT PESTS
ABSTRACT
One of the constraints faced in developing the program of biological control using parasitoid is the lack of information technology on the release of these natural enemies in different regions of Brazil. Studies should be conducted in this area because the material with which it works is invariably sensitive to the environment. It is known that the release efficiency of these parasitoids in the field depends largely on factors such as the ability to "search", host preference, tolerance to climatic conditions, range, number and timing of releases, number of released insects, plant architecture, among others. Therefore, knowledge of these factors and their interactions enable better management of natural control of pest or a set of them. The aim of this study is to address and discuss the factors associated with the release of parasitoids in the field of parasitism affects its efficiency. KEYWORDS: parasitism, plant-insect relationship, tritrophic level, VOC’s

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
551
1 INTRODUÇÃO O uso de defensivos agrícolas, nos últimos tempos, é o principal método de controle de insetos-praga em diversas culturas. Porém, eles são utilizados muitas vezes de forma abusiva na agricultura e traz consigo inúmeros impactos ao meio ambiente e a outros componentes do sistema, como a saúde do trabalhador e do consumidor. Também causam intoxicação de organismos benéficos, como os inimigos naturais e desequilibra ainda mais o agroecossistema (SILVA, 2007a). Neste cenário, tem-se o retorno de insetos herbívoros que se tornam pragas, atingindo níveis prejudiciais com grande rapidez, uma vez que seus predadores e parasitóides, ocupando categorias mais elevadas na teia alimentar, levam maior tempo para recompor os níveis populacionais (SILVA, 2007a). Os predadores e os parasitóides são elementos da fauna que controlam a população de insetos-praga e sua sobrevivência está relacionada com a diversificação de habitat que fornecem recursos essenciais para estes inimigos naturais (ALTIERI, 1999).
Dentro do grupo de inimigos naturais, os parasitóides representam grande valor econômico por regularem de forma eficiente os insetos-praga. Algumas características são inerentes a este grupo de insetos, tais como a utilização de apenas um tipo de hospedeiro para completar o seu ciclo (insetos monoécios), serem menores que seu hospedeiro, parasitar apenas na fase imatura, nos quais os adultos são de vida livre, alimentando-se de pólen e néctar. Neste sentido, a utilização deste grupo de insetos no controle biológico de populações de herbívoros ganha respaldo em diversas culturas agrícolas (GARCIA, 1991).
O controle biológico de insetos-praga é uma prática sustentável que pode diminuir o uso de inseticidas e vem ganhando representatividade para o controle de diversas pragas, em especial na cultura da cana-de-açúcar, trigo, citros, mandioca e em pastagens (NAVA, 2007). Para o sucesso de um programa de controle biológico utilizando parasitóides é importante observar alguns aspectos relacionados ao comportamento e a biologia destes insetos no campo (BUENO, 2008).
As variações comportamentais destes insetos estão relacionadas com fatores bióticos e abióticos, que de forma direta ou indireta, influenciam na eficiência do parasitismo. O hospedeiro, a adaptabilidade e a capacidade intrínseca da linhagem sob as condições de cada agroambiente são alguns fatores bióticos que proporcionam alterações comportamentais em parasitóides. Já, os fatores abióticos que podem ser descritos são as condições climáticas, as características dos cultivos e o impacto ambiental dos agrotóxicos (PRATISSOLI et al., 2002). Para a avaliação das características biológicas dos parasitóides, PARRA (1996) cita que é necessário seguir algumas etapas para se ter sucesso na utilização de Trichogramma sp. (Hymenoptera), um parasitóide de ovos de diversas pragas-chave de 34 culturas em mais de 30 países.
Um dos limitadores enfrentados para o desenvolvimento do programa de controle biológico com o uso de parasitóide é a falta de informações sobre a tecnologia de liberação destes inimigos naturais nas diversas regiões do Brasil. Estudos devem ser realizados nesta área, pois o material com que se trabalha é invariavelmente sensível ao meio ambiente (PINTO & PARRA, 2002).
É sabido que a eficiência das liberações destes parasitóides no campo depende basicamente de alguns fatores, tais como a capacidade de “busca”, preferência hospedeira, tolerância às condições climáticas, intervalo, número e época de liberações, número de insetos liberados, arquitetura da planta, entre outros. Portanto, o conhecimento desses fatores e suas interações possibilitam

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
552
melhor manejo do controle natural de uma praga ou conjunto delas (BROGLIO-MICHELETTI et al., 2007; BUENO, 2008). O objetivo deste estudo é abordar e discutir os fatores que associados a liberação dos parasitóides em campo afeta sua eficiência de parasitismo. 2 FATORES QUE AFETAM A LIBERAÇÃO E A EFICIÊNCIA DE PARASITÓIDES 2.1 INSETOS PARASITÓIDES
As espécies de insetos cujas larvas se desenvolvem no corpo de outro
artrópode, usualmente um inseto, ou em uma massa única ou gregária de hospedeiros, como ootecas ou massas de larvas galhadoras, acarretando a morte do hospedeiro ao final do seu desenvolvimento são denominados parasitóides. O organismo que é denominado parasita geralmente é menor que a sua presa e ao parasitá-lo não leva a sua morte (PARRA & BENTO, 2006).
Os parasitóides são organismos que possuem um comportamento intermediário entre os predadores e os parasitas, pois residem dentro de um hospedeiro vivo se alimentando de seus tecidos e matam seus hospedeiros de forma inevitável. Os adultos são de vida livre e se alimentam de néctar e pólen. Ao encontrar um hospedeiro, que pode ser um ovo, uma larva, uma pupa ou imagos de outros insetos, a fêmea deposita um ou vários ovos sobre ele e a larva do parasitóide se alimentará do tecido desse hospedeiro o qual morre nessa relação (MARCHIORI et al., 2006).
Os parasitóides de ovos matam o hospedeiro antes mesmo que este inicie seu ataque sobre a plantação. Já os parasitóides de larvas e de pupas reduzem o tamanho das populações hospedeiras apenas nas gerações seguintes. Do ponto de vista do controle biológico os parasitóides de ovos são mais importantes que os demais (CAMPOS & ARAÚJO, 1994).
Em função do local de deposição dos ovos, esses insetos podem ser classificados como endo ou ectoparasitóide, em que os ovos são depositados no interior ou sobre os hospedeiros, respectivamente. Eles também podem ser divididos quanto ao modo de ataque do hospedeiro após ser parasitado em idiobiontes e coinobiontes. Os primeiros cessam o desenvolvimento do hospedeiro e o segundo permitem que o hospedeiro permaneça vivo até o seu desenvolvimento (GARCIA, 1991). Os parasitóides podem ser ainda especialistas ou generalistas. Aqueles que utilizam exclusivamente uma determinada espécie de hospedeiro são ditos como especialistas. Estes dependem inteiramente de sua espécie hospedeira e comumente são endoparasitóides coinobiontes. Já os generalistas, utilizam uma gama variável de espécies hospedeiras distintas (CAMPOS & ARAÚJO, 1994).
As características reprodutivas dos parasitóides conferem a este grupo de insetos a capacidade de regular outros insetos que, num nível populacional alto, podem provocar prejuízos a agricultura e pecuária. Esta atividade reguladora desempenhada pelos parasitóides nos ecossistemas é tão importante que sem a mesma ocorreria um consumo alto de espécies vegetais por insetos herbívoros e, nesse caso, tornam-se essenciais para a sustentação do equilíbrio ecológico, contribuindo com a diversidade de outros organismos. Nestes novos tempos, os parasitóides vêm sendo estudados com o propósito de serem utilizados em programas de controle biológico de insetos (SCATOLINI & PENTEADO-DIAS, 2003).
Em vários países o interesse no controle biológico tem crescido como resposta aos efeitos adversos dos pesticidas químicos no ambiente e na

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
553
biodiversidade. Outro aspecto que favoreceu o seu crescimento está relacionado com as novas tendências internacionais na produção agrícola que envolve a utilização de meios alternativos que são menos agressivos ao meio ambiente e que favorecem a conservação e o uso sustentável da biodiversidade (MARCHIORI et al., 2007).
Os insetos parasitóides ocorrem nas ordens Coleoptera, Lepidoptera, Trichoptera, Neuroptera sendo mais comuns na ordem Hymenoptera, onde as estimativas atuais indicam que as vespas parasitóides podem constituir de 10 a 20% do total de insetos. Esta última ordem é composta de espécies que habitam os diferentes tipos de ambientes, com aproximadamente 115.000 representantes, e uma perspectiva aponta que existem cerca de 250.000 espécies em diferentes regiões do mundo. Os Hymenoptera endoparasitóide podem ser encontrados nas superfamílias Platygastroidea e Cynipoidea; nas famílias Monomachidae, Diapriidae e Heloridae; indivíduos ectoparasitóides são Evanioidea, Ceraphronoidea, Ichneumonoidea e Chalcidoidea (SANTOS, 2008).
Este grupo de insetos favoreceu nos últimos anos uma grande economia de recursos em programas de controle de pragas e são responsáveis pela maior parte dos benefícios econômicos e ambientais produzidos nestes. Porém, na região Neotropical estas espécies foram pouco estudadas e são pouco conhecidas (MARCHIORI et al., 2007). 2.2 PANORAMA DAS PESQUISAS SOBRE OS PARASITÓIDES ASSOCIADOS ÀS CULTURAS
O uso de insetos parasitóides no programa de controle biológico é uma
prática crescente em vários países. O controle biológico de pragas, em conjunto com outras táticas preservacionistas de controle, como o manejo sustentável dos recursos naturais e a preservação da biodiversidade, contribui com a sustentabilidade do agronegócio brasileiro (STEFANELO, 2002).
As pesquisas brasileiras voltadas para o controle biológico de insetos pragas tornam-se uma estratégia que pode auxiliar na qualidade da produção, principalmente numa cultura orgânica que tem como vantagens a diminuição nos custos de produção, impactos ambientais reduzidos e menor exposição a produtos químicos pelo homem, sendo possível reduzir o uso dos mesmos em até 60 % (SANTOS, 2008).
O programa de controle biológico com o parasitóide de ovos Trichogramma sp. (Hymenoptera: Trichogrammatidae), que envolveu desde a taxonomia até a avaliação da eficiência deste parasitóide, é um modelo seguido por outros programas de controle biológico no Brasil e na América Latina. Este projeto gerou inúmeras publicações, permitiu a formação de recursos humanos na área, abriu novas áreas de pesquisa e mostrou que o parasitóide pode ser usado no controle de pragas-chave do algodoeiro, cana-de-açúcar, grãos armazenados, hortaliças, milho, soja e tomateiro (PARRA & ZUCCHI, 2004).
O controle biológico utilizando parasitóides em outros países da América Latina como a Venezuela, o Peru, a Argentina e a Bolívia começou na década de 1930 e 1940 com a introdução dos parasitóides Aphelinus mali Haldeman (Hymenoptera: Aphelinidae) e Apanteles thurberiae Muesebeck (Hymenoptera: Braconidae) para o controle do pulgão Eriosoma lanigerum Hausmann (Homoptera: Aphididae) e a praga do algodão Sacadodes pyralis Dyar (Lepidoptera: Noctuidae), respectivamente. Nos últimos anos, o controle biológico de pragas vem sendo

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
554
utilizado de forma bem sucedida nas lavouras de cana, milho e sorgo, apresentando tanto valor econômico como ecológico (FERRER, 2001).
Na Europa o uso de insetos parasitóides tem sido uma estratégia bem sucedida para o controle biológico em estufas. Esta estratégia foi adotada pelos produtores por causa da resistência das pragas aos inseticidas e os aumentos do custo do controle químico. O programa foi originalmente construído em torno do uso do parasitóide Encarsia formosa Gahan (Hymenoptera: Aphelinidae) contra a mosca branca Bemisia tabaci biótipo B (Hemiptera: Aleyrodidae) e o ácaro predador Phytoseiulus persimilis Athias-Henriot (Acarina: Phytoseiidae) contra os dois ácaros rajados. Ao longo dos anos, mais parasitóides foram adicionados ao controle de outras pragas, como tripes, larvas, pulgões, lagartas e outras espécies de mosca-branca (STONER, 2011).
Na Ásia e na Austrália as pesquisas sobre parasitóides estão voltadas para o controle de uma das pragas mais destrutivas, a traça das crucíferas Plutella xylostella L. (Lepidoptera: Plutellidae). O endoparasitóide larval Diadegma semiclausum Hellén (Hymenoptera: Ichneumonidae) vem sendo utilizado com sucesso nestes dois continentes para o controle desta praga (GICHINI et al., 2008). Na África, a broca Hypothenemus hampei Ferrari (Coleoptera: Curculionidae) é um dos insetos mais devastador do café neste continente. Nos dias atuais, o controle biológico com os parasitóides é o mais utilizado para o controle desta praga (JARAMILLO et al., 2009). 2.3 O COMPLEXO TRITRÓFICO E A SUA INTERFERÊNCIA NA LIBERAÇÃO E EFICIÊNCIA DE PARASITÓIDES
As plantas e os insetos mantêm uma relação de mútua dependência na busca de alimentos e reprodução. Juntos, eles formam os dois maiores grupos de organismos que ocupam diferentes habitats na natureza. As plantas fornecem recursos alimentares, abrigo e locais de reprodução para os insetos. As abelhas, vespas, borboletas, mariposas e moscas são alguns exemplos de insetos que contribuem com os processos reprodutivos das plantas atuando como agentes polinizadores, o que favorece a fecundação cruzada e incrementa a diversidade genética de várias espécies vegetais. Neste sentido, o conhecimento e a conservação das relações entre os organismos pertencentes a um sistema tritrófico, ou seja, a interação planta-herbívoro-inimigo natural, servem não somente para manter a dinâmica do ecossistema natural, como também para ser aproveitada economicamente pelo homem no controle biológico de insetos-pragas em agroecossistemas (HEIL, 2007). 2.3.1 Interações com a planta e a comunidade vegeta l
As propriedades individuais ou da comunidade de plantas podem
influenciar nas interações de insetos-praga e seus inimigos naturais. O teor de nitrogênio, água e outros nutrientes, bem como a presença de substâncias do metabolismo secundário podem afetar o crescimento, a sobrevivência e a reprodução de insetos herbívoros e de seus inimigos naturais (CAMPOS & ARAÚJO, 1994).
As interações das plantas com os demais níveis tróficos estão relacionadas com a liberação de certas substâncias do metabolismo secundário ou compostos orgânicos voláteis (COV). A liberação de COV é induzida pela

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
555
alimentação do inseto, sendo uma resposta mediada por um hormônio na planta, o ácido jasmônico (Figura 1) e que, na maioria das vezes, relacionam-se a defesa do vegetal (HEIL, 2007).
FIGURA 1. Efeitos biológicos e interações de compostos orgânicos
voláteis liberados pelas plantas como mecanismo de defesa (Fonte: HEIL, 2007).
Os COV’s apresentam-se em quantidades variáveis, de acordo com os
diferentes estágios da vida das plantas, podendo variar, ainda, segundo a localização, grau, tempo e tipo da injúria ocasionada pelo inseto-praga. É hoje amplamente aceito que estes compostos orgânicos podem atrair artrópodes predadores e/ou repelir herbívoros e, assim, servir como meio de resistência de plantas (HEIL, 2007; ARRUDA, 2011).
A cobertura vegetal e a arquitetura da planta são outros aspectos importantes que podem influenciar a vida dos insetos-pragas e a eficiência de controle de seus inimigos naturais. Já o policultivo aumenta a diversidade de habitats e fornece abrigo, alimento alternativo para os parasitóides na sua fase adulta e são indispensáveis à sobrevivência e reprodução deste grupo de insetos (GINGRAS et al., 2002).
O policultivo é facilmente incorporado em hortas e até mesmo em pequena escala em plantações comerciais, não sendo comum em produção agrícola em grande escala. Em monocultivos, a ausência de recursos alimentares adicionais inviabiliza a permanência ou a fixação dos parasitóides nos agroecossistemas agrícolas (ALTIERI, 1999). Em trabalhos de ROMERO et al. (1984) citado por BASTOS et al. (2003) observou-se uma maior porcentagem de parasitismo em ovos de Empoasca kraemeri Ross & Moore (Homoptera: Cicadellidae) por Anagrus sp. (Hymenoptera: Mymaridae) no cultivo do feijão consorciado com o milho que no cultivo exclusivo. 2.3.2 Interação com o hospedeiro
A relação entre o parasitóide e seu hospedeiro está relacionada com aspectos endócrinos, fisiológicos e comportamentais. Nesta interação, o parasitóide segue algumas etapas, tais como a localização do habitat do vegetal e do seu

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
556
hospedeiro, aceitação, adequação e regulação do hospedeiro, que favorecem o sucesso do seu parasitismo. Neste sentido, os estudos sobre as respostas sensoriais dos insetos são importantes para compreender as interações entre a capacidade de busca e o comportamento dos parasitóides (CAMPOS & ARAÚJO, 1994; SILVA et al., 2006; FATOUROS et al., 2008).
A localização do habitat do hospedeiro pelo parasitóide é favorecida pela liberação de sinais importantes que podem vir da planta, do hospedeiro, da relação planta-hospedeiro ou uma combinação destes fatores. SIQUEIRA & FARIAS (2003) verificaram a atração de E. formosa a estímulos provenientes da planta de tomate (Lycopersicom esculentum Mill.), ao hospedeiro B. tabaci biótipo B e ao complexo planta-hospedeiro. Os autores verificaram que as fêmeas do parasitóide estudado respondem positivamente aos odores emitidos do complexo planta-hospedeiro (Tabela 1). TABELA 1. Tempo médio e porcentagem do tempo total utilizados pelas fêmeas de
E. formosa quando expostas a diferentes odores em olfatômetro de quatro vias a 26°C e UR de 74% (Siqueira e Farias, 2003).
Campos de odores Tempo médio (seg)
Média de tempo
Ar filtrado 1 47,75 14,40 Ar filtrado 2 40,04 12,15 Folhas de tomate infestadas com ninfas de Bemisia tabaci biótipo B
197,87 60,06
Folhas de tomate sem ninfas 44,08 13,38
Depois de localizar o hábitat do hospedeiro, o parasitóide direciona sua busca pelo indivíduo onde vai depositar seus ovos e tem de aprender a discriminar muitas pistas diferentes relacionadas à presença do hospedeiro, especialmente pistas químicas (cairomônios) que podem ser liberadas tanto pelos ovos como pelos adultos (BOIÇA JR. et al., 1997; PACHECO et al., 2000; FATOUROS et al., 2008).
SILVA et al. (2006) avaliaram a resposta sensorial do parasitóide de ovos Telenomus podisi Ashmead (Hymenoptera: Scelionidae) a estímulos provenientes do percevejo Euschistus heros Fabricius (Hemiptera: Pentatomidae) e concluíram que o parasitóide utiliza estímulos sensoriais do hospedeiro masculino e a longas distâncias é fracamente atraído por fêmeas e massas de ovos do percevejo.
2.4 FATORES QUE AFETAM A LIBERAÇÃO E A EFICIÊNCIA D E PARASITÓIDES
A liberação de um parasitóide é o ato de soltura deste inimigo natural no campo para que este venha reduzir o índice populacional de um determinado inseto-praga (PINTO & PARRA, 2002). Segundo WILLIAMS & LEPPLA (1992) citado por PARRA & BENTO (2006), as formas de liberação de inimigos naturais são inoculativa, inundativa e inoculativa estacional ou sazonal que são dependentes do sistema visado.
A liberação inoculativa é comum em controle biológico clássico, pois é feita em locais abertos e com baixa variabilidade temporal e principalmente em culturas perenes ou semiperenes e florestais. O segundo tipo de liberação é para culturas com alta variabilidade temporal como as anuais. Já a liberação inundativa estacional é feita para cultivos de curta duração, sendo comum em casa de vegetação (PARRA & BENTO, 2006).

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
557
Alguns fatores, tais como o número de pontos, a técnica, a frequência e o intervalo de liberações, a arquitetura e idade da planta e as condições climáticas prejudicam a eficiência destes tipos de liberação e, consequentemente, a eficiência dos parasitóides no campo. A eficiência do parasitóide pode ser prejudicada, além destes fatores, pela presença de outros inimigos naturais, preferência hospedeira, número de insetos liberados, entre outros (BUENO, 2008; PINTO & PARRA, 2002). Neste sentido, o sucesso do programa de controle biológico depende de pesquisas que avaliem tais fatores.
2.4.1 Número de pontos de liberação
O número de pontos de liberação por unidade de área é determinado a partir da capacidade de dispersão do parasitóide no campo. Esta dispersão pode ocorrer de forma ativa ou passiva, sendo influenciada pelos fatores abióticos, tais como o vento, a temperatura e a umidade, pela arquitetura da planta e/ou espécie do parasitóide. A dispersão ativa é estimulada pela temperatura e inibida pela densidade de ovos do hospedeiro. Já o segundo tipo é controlado pela ação do vento (CANTO-SILVA et al., 2006). O conhecimento da capacidade de dispersão de parasitóide é uma ferramenta importante na determinação do número de pontos de liberação e, consequentemente, na elaboração de metodologias que viabilizem a eficiência deste grupo de insetos no campo, principalmente em liberação inundativas. Já a determinação do número de pontos de liberação por unidade de área possui papel fundamental na adoção do programa de liberação de um agente de controle biológico, uma vez que os custos de liberação estão diretamente relacionados a este número (ZACHRISSON & PARRA, 1998).
Em trabalhos realizados por SILVA (2007b) a capacidade de dispersão do parasitóide Telenomus remus Nixon (Hymenoptera: Scelionidae) foi determinada em condições de laboratório e em cultivo de algodoeiro (Gossypium hirsutum L.), sendo avaliada a partir de diferentes variáveis meteorológicas, do desenvolvimento de plantas e da disponibilidade de hospedeiros. Este inseto parasita ovos de Spodoptera spp. (Lepidoptera: Noctuidae), uma praga representativa desta cultura. A direção eólica, o tamanho das plantas e a disponibilidade de hospedeiros influenciaram a movimentação das fêmeas T. remus, e, consequentemente, o seu padrão de dispersão. Ao concluir seu trabalho, o autor determinou o número ideal de pontos de liberação por unidade de área (25 pontos por hectare) e guiou a elaboração de esquemas de distribuição dos pontos de liberação, na cultura estudada. A quantidade de ponto de liberação estabelecida para este parasitóide chega a ser até quatro vezes menor do que às indicadas para o parasitóide Trichogramma pretiosum Riley (100 e 130 pontos/ha), por exemplo, nas culturas do milho e da soja, para controle de Helicoverpa zea Boddie e Anticarsia gemmatalis Hübner (Lepidoptera: Noctuidae), respectivamente, segundo os dados obtidos por SÁ et al. (1993) e ZACHRISSON & PARRA (1998).
2.4.2 Técnica de liberação
No momento da liberação do parasitóide no campo alguns aspectos importantes, tais como o tipo de recipiente para liberação, fase de desenvolvimento em que o parasitóide é liberado e o modo de liberação, devem ser considerados para que se tenha sucesso no programa de controle biológico. O recipiente utilizado para a soltura do parasitóide deve ser de material atóxico, economicamente viável e de fácil manuseio, transporte e distribuição pela área. Ele tem como função principal proteger os inimigos naturais de outros agentes biológicos e adversidade climáticas.
ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
558
Os modos de liberação dos parasitóides no campo podem ser manual ou mecanicamente, de forma aérea ou terrestre, em pontos fixos ou em vários pontos dentro de uma área. Porém, todos estes modos de liberação devem ser testados para que a eficiência do parasitóide não seja diminuída (PINTO & PARRA, 2002).
Na cultura de cana-de-açúcar o controle da broca Diatraea saccharalis Fabricius (Lepidoptera: Crambidae) é feito com o parasitóide larval Cotesia flavipes Cameron (Hymenoptera: Braconidae) que é obtido pelos produtores na fase pupal em copos de plásticos contendo cerca de 1.500 indivíduos cada um e a liberação do tipo inundativa é realizada após 12 horas do início da emergência dos adultos. Recomenda-se que entre no talhão, no sentido das linhas de cana, caminhando com o copo aberto e a cada 41 passos (± 33m) colocá-lo, na bainha da cana, em posição horizontal. Neste sentido, serão formados quatro pontos por hectare e quando o caminhamento interno é difícil, como em canaviais muito fechados com cana-de-açúcar acamadas, é recomendável fazer as liberações circundando os mesmos e penetrando cerca de 25m os carreadores. O momento ideal de liberação do parasitóide é quando as lagartas começam a fazer galerias nos colmos, observando a presença de dejetos e quando elas estiverem com 1 a 1,5 cm de comprimento (PINTO et al., 2006).
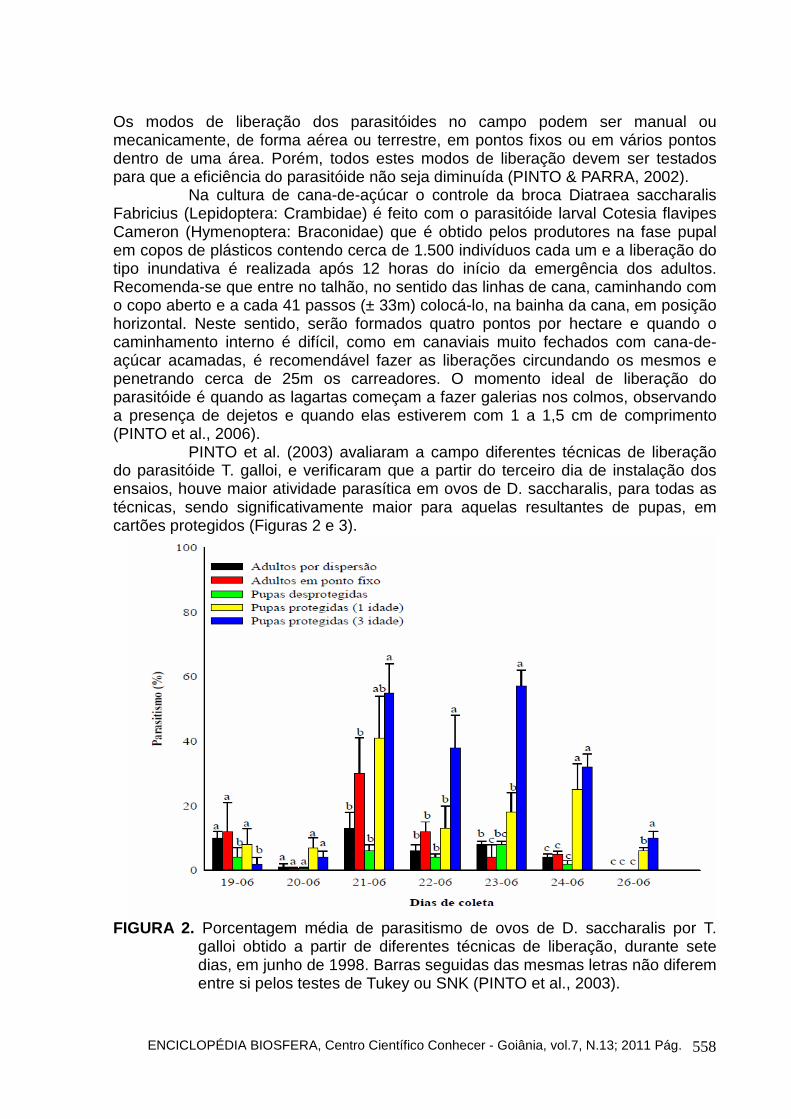
PINTO et al. (2003) avaliaram a campo diferentes técnicas de liberação do parasitóide T. galloi, e verificaram que a partir do terceiro dia de instalação dos ensaios, houve maior atividade parasítica em ovos de D. saccharalis, para todas as técnicas, sendo significativamente maior para aquelas resultantes de pupas, em cartões protegidos (Figuras 2 e 3).
FIGURA 2. Porcentagem média de parasitismo de ovos de D. saccharalis por T.
galloi obtido a partir de diferentes técnicas de liberação, durante sete dias, em junho de 1998. Barras seguidas das mesmas letras não diferem entre si pelos testes de Tukey ou SNK (PINTO et al., 2003).
ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
559
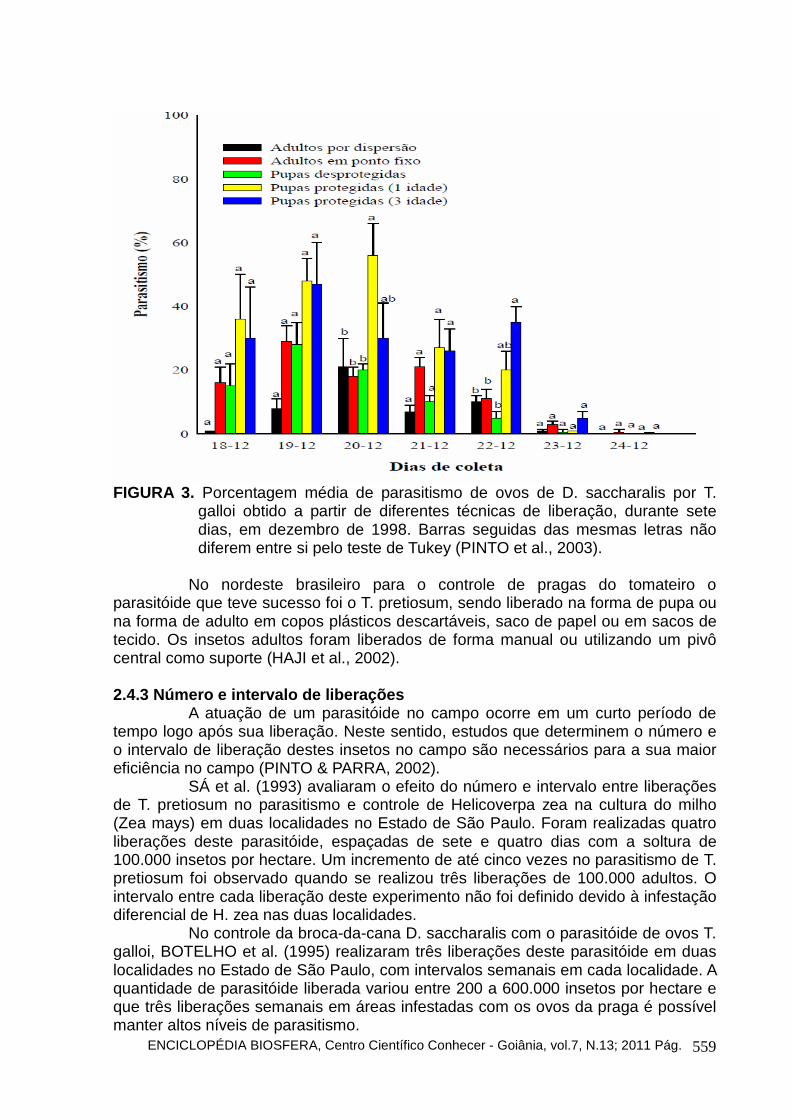
FIGURA 3. Porcentagem média de parasitismo de ovos de D. saccharalis por T.
galloi obtido a partir de diferentes técnicas de liberação, durante sete dias, em dezembro de 1998. Barras seguidas das mesmas letras não diferem entre si pelo teste de Tukey (PINTO et al., 2003).
No nordeste brasileiro para o controle de pragas do tomateiro o
parasitóide que teve sucesso foi o T. pretiosum, sendo liberado na forma de pupa ou na forma de adulto em copos plásticos descartáveis, saco de papel ou em sacos de tecido. Os insetos adultos foram liberados de forma manual ou utilizando um pivô central como suporte (HAJI et al., 2002). 2.4.3 Número e intervalo de liberações
A atuação de um parasitóide no campo ocorre em um curto período de tempo logo após sua liberação. Neste sentido, estudos que determinem o número e o intervalo de liberação destes insetos no campo são necessários para a sua maior eficiência no campo (PINTO & PARRA, 2002).
SÁ et al. (1993) avaliaram o efeito do número e intervalo entre liberações de T. pretiosum no parasitismo e controle de Helicoverpa zea na cultura do milho (Zea mays) em duas localidades no Estado de São Paulo. Foram realizadas quatro liberações deste parasitóide, espaçadas de sete e quatro dias com a soltura de 100.000 insetos por hectare. Um incremento de até cinco vezes no parasitismo de T. pretiosum foi observado quando se realizou três liberações de 100.000 adultos. O intervalo entre cada liberação deste experimento não foi definido devido à infestação diferencial de H. zea nas duas localidades.
No controle da broca-da-cana D. saccharalis com o parasitóide de ovos T. galloi, BOTELHO et al. (1995) realizaram três liberações deste parasitóide em duas localidades no Estado de São Paulo, com intervalos semanais em cada localidade. A quantidade de parasitóide liberada variou entre 200 a 600.000 insetos por hectare e que três liberações semanais em áreas infestadas com os ovos da praga é possível manter altos níveis de parasitismo.
ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
560
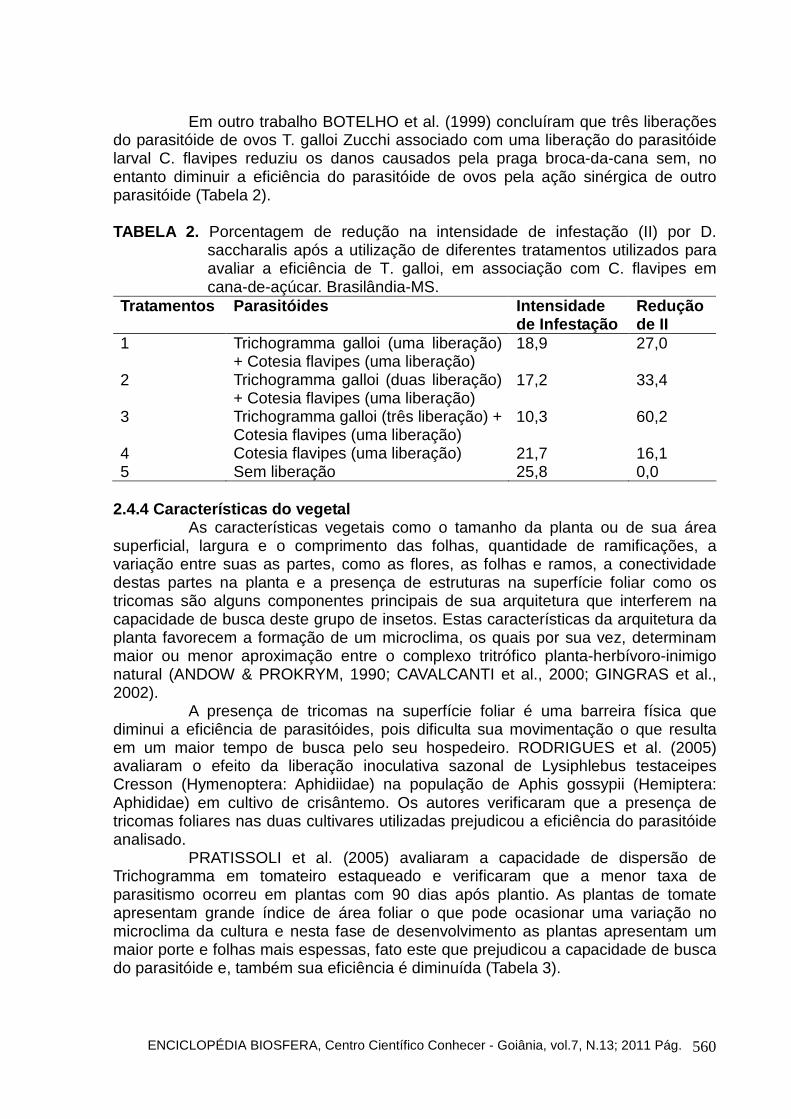
Em outro trabalho BOTELHO et al. (1999) concluíram que três liberações do parasitóide de ovos T. galloi Zucchi associado com uma liberação do parasitóide larval C. flavipes reduziu os danos causados pela praga broca-da-cana sem, no entanto diminuir a eficiência do parasitóide de ovos pela ação sinérgica de outro parasitóide (Tabela 2).
TABELA 2. Porcentagem de redução na intensidade de infestação (II) por D.
saccharalis após a utilização de diferentes tratamentos utilizados para avaliar a eficiência de T. galloi, em associação com C. flavipes em cana-de-açúcar. Brasilândia-MS.
Tratamentos Parasitóides Intensidade de Infestação
Redução de II
1 Trichogramma galloi (uma liberação) + Cotesia flavipes (uma liberação)
18,9 27,0
2 Trichogramma galloi (duas liberação) + Cotesia flavipes (uma liberação)
17,2 33,4
3 Trichogramma galloi (três liberação) + Cotesia flavipes (uma liberação)
10,3 60,2
4 Cotesia flavipes (uma liberação) 21,7 16,1 5 Sem liberação 25,8 0,0
2.4.4 Características do vegetal
As características vegetais como o tamanho da planta ou de sua área superficial, largura e o comprimento das folhas, quantidade de ramificações, a variação entre suas as partes, como as flores, as folhas e ramos, a conectividade destas partes na planta e a presença de estruturas na superfície foliar como os tricomas são alguns componentes principais de sua arquitetura que interferem na capacidade de busca deste grupo de insetos. Estas características da arquitetura da planta favorecem a formação de um microclima, os quais por sua vez, determinam maior ou menor aproximação entre o complexo tritrófico planta-herbívoro-inimigo natural (ANDOW & PROKRYM, 1990; CAVALCANTI et al., 2000; GINGRAS et al., 2002).
A presença de tricomas na superfície foliar é uma barreira física que diminui a eficiência de parasitóides, pois dificulta sua movimentação o que resulta em um maior tempo de busca pelo seu hospedeiro. RODRIGUES et al. (2005) avaliaram o efeito da liberação inoculativa sazonal de Lysiphlebus testaceipes Cresson (Hymenoptera: Aphidiidae) na população de Aphis gossypii (Hemiptera: Aphididae) em cultivo de crisântemo. Os autores verificaram que a presença de tricomas foliares nas duas cultivares utilizadas prejudicou a eficiência do parasitóide analisado.
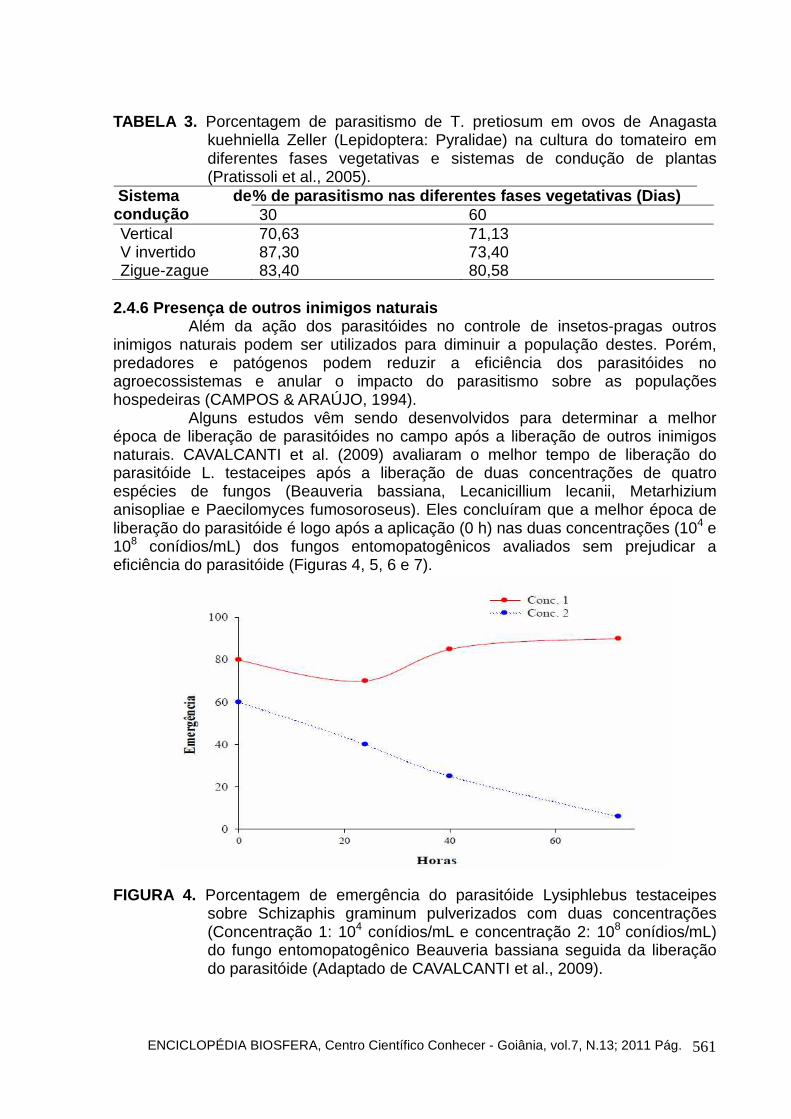
PRATISSOLI et al. (2005) avaliaram a capacidade de dispersão de Trichogramma em tomateiro estaqueado e verificaram que a menor taxa de parasitismo ocorreu em plantas com 90 dias após plantio. As plantas de tomate apresentam grande índice de área foliar o que pode ocasionar uma variação no microclima da cultura e nesta fase de desenvolvimento as plantas apresentam um maior porte e folhas mais espessas, fato este que prejudicou a capacidade de busca do parasitóide e, também sua eficiência é diminuída (Tabela 3).
ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
561
TABELA 3. Porcentagem de parasitismo de T. pretiosum em ovos de Anagasta kuehniella Zeller (Lepidoptera: Pyralidae) na cultura do tomateiro em diferentes fases vegetativas e sistemas de condução de plantas (Pratissoli et al., 2005).
% de parasitismo nas diferentes fases vegetativas ( Dias) Sistema de condução 30 60 Vertical 70,63 71,13 V invertido 87,30 73,40 Zigue-zague 83,40 80,58
2.4.6 Presença de outros inimigos naturais
Além da ação dos parasitóides no controle de insetos-pragas outros inimigos naturais podem ser utilizados para diminuir a população destes. Porém, predadores e patógenos podem reduzir a eficiência dos parasitóides no agroecossistemas e anular o impacto do parasitismo sobre as populações hospedeiras (CAMPOS & ARAÚJO, 1994).
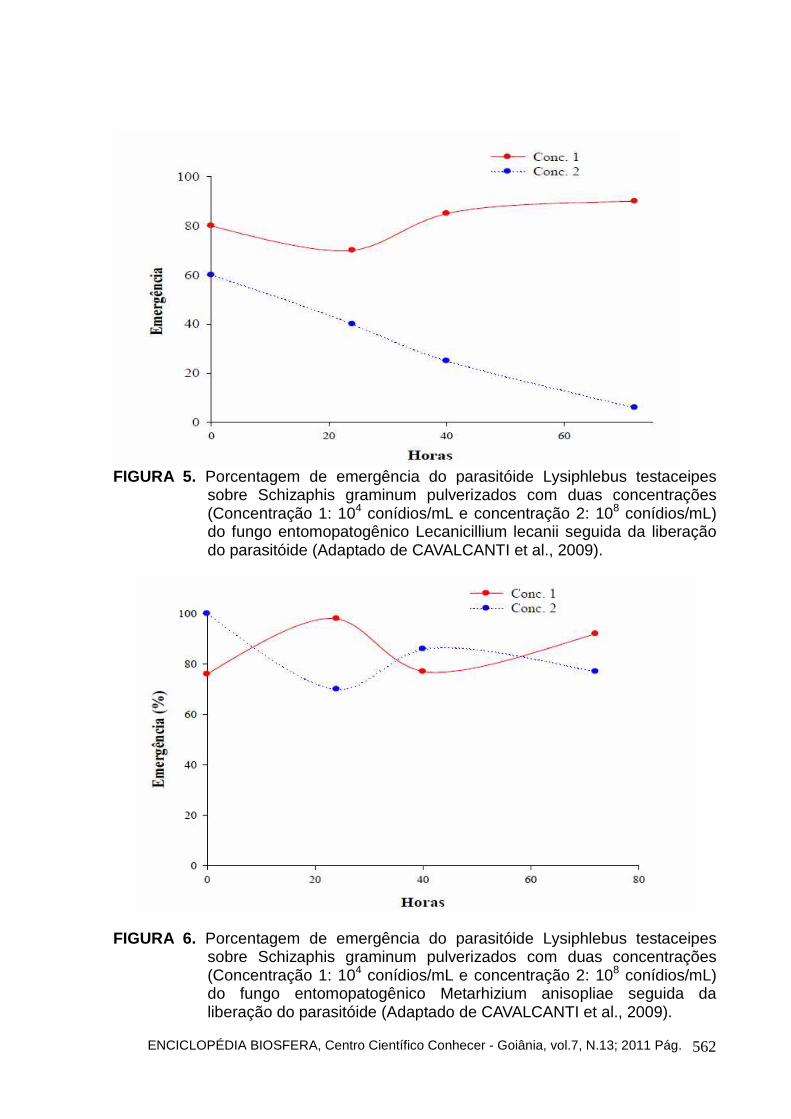
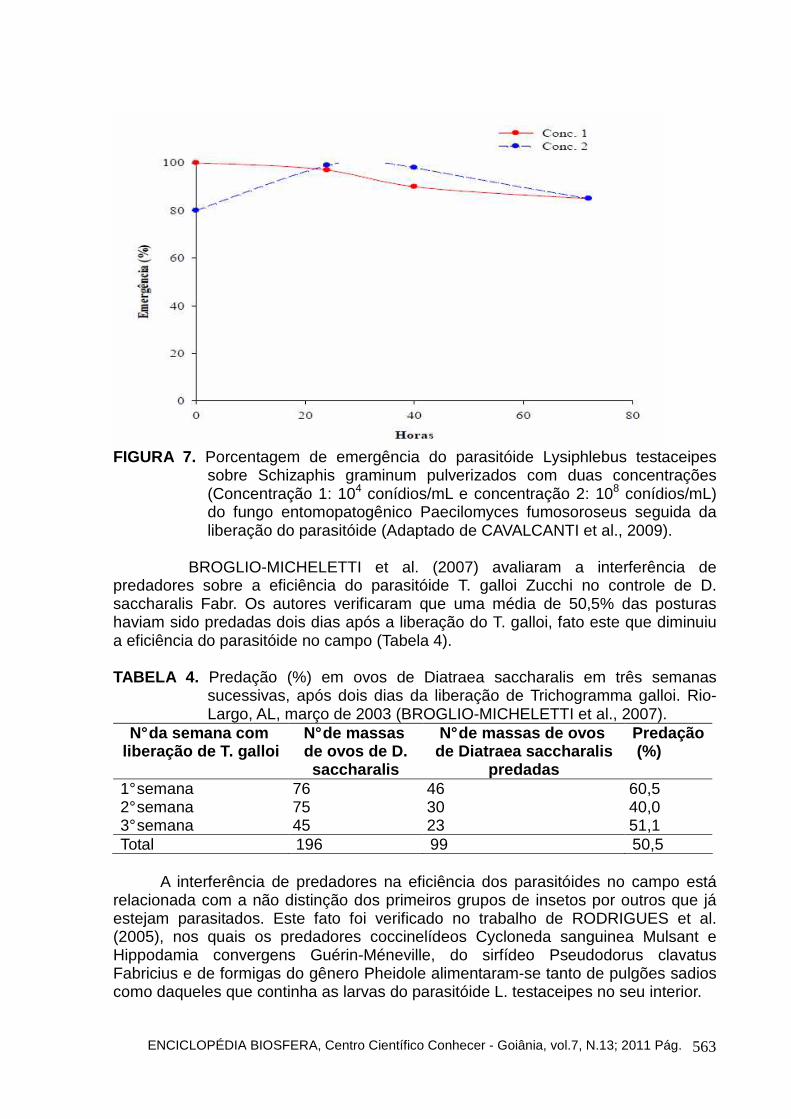
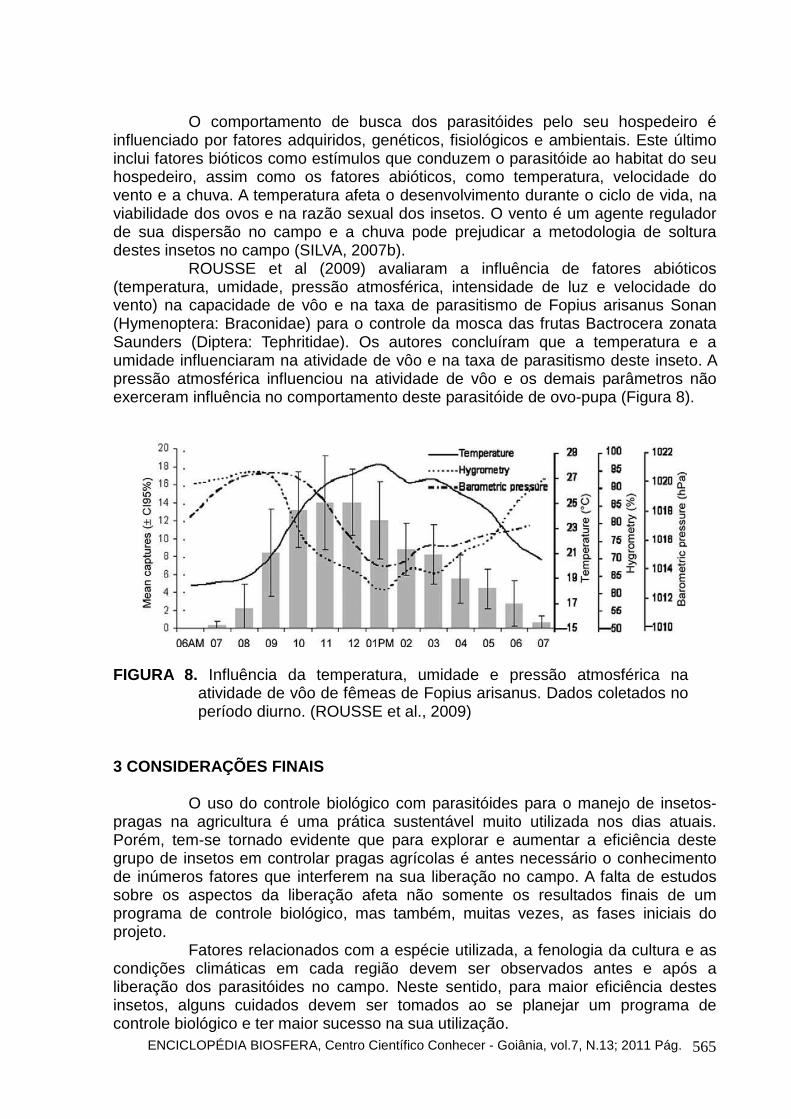
Alguns estudos vêm sendo desenvolvidos para determinar a melhor época de liberação de parasitóides no campo após a liberação de outros inimigos naturais. CAVALCANTI et al. (2009) avaliaram o melhor tempo de liberação do parasitóide L. testaceipes após a liberação de duas concentrações de quatro espécies de fungos (Beauveria bassiana, Lecanicillium lecanii, Metarhizium anisopliae e Paecilomyces fumosoroseus). Eles concluíram que a melhor época de liberação do parasitóide é logo após a aplicação (0 h) nas duas concentrações (104 e 108 conídios/mL) dos fungos entomopatogênicos avaliados sem prejudicar a eficiência do parasitóide (Figuras 4, 5, 6 e 7).
FIGURA 4. Porcentagem de emergência do parasitóide Lysiphlebus testaceipes sobre Schizaphis graminum pulverizados com duas concentrações (Concentração 1: 104 conídios/mL e concentração 2: 108 conídios/mL) do fungo entomopatogênico Beauveria bassiana seguida da liberação do parasitóide (Adaptado de CAVALCANTI et al., 2009).
ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
562
FIGURA 5. Porcentagem de emergência do parasitóide Lysiphlebus testaceipes
sobre Schizaphis graminum pulverizados com duas concentrações (Concentração 1: 104 conídios/mL e concentração 2: 108 conídios/mL) do fungo entomopatogênico Lecanicillium lecanii seguida da liberação do parasitóide (Adaptado de CAVALCANTI et al., 2009).
FIGURA 6. Porcentagem de emergência do parasitóide Lysiphlebus testaceipes
sobre Schizaphis graminum pulverizados com duas concentrações (Concentração 1: 104 conídios/mL e concentração 2: 108 conídios/mL) do fungo entomopatogênico Metarhizium anisopliae seguida da liberação do parasitóide (Adaptado de CAVALCANTI et al., 2009).
ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
563
FIGURA 7. Porcentagem de emergência do parasitóide Lysiphlebus testaceipes
sobre Schizaphis graminum pulverizados com duas concentrações (Concentração 1: 104 conídios/mL e concentração 2: 108 conídios/mL) do fungo entomopatogênico Paecilomyces fumosoroseus seguida da liberação do parasitóide (Adaptado de CAVALCANTI et al., 2009).
BROGLIO-MICHELETTI et al. (2007) avaliaram a interferência de
predadores sobre a eficiência do parasitóide T. galloi Zucchi no controle de D. saccharalis Fabr. Os autores verificaram que uma média de 50,5% das posturas haviam sido predadas dois dias após a liberação do T. galloi, fato este que diminuiu a eficiência do parasitóide no campo (Tabela 4).
TABELA 4. Predação (%) em ovos de Diatraea saccharalis em três semanas
sucessivas, após dois dias da liberação de Trichogramma galloi. Rio-Largo, AL, março de 2003 (BROGLIO-MICHELETTI et al., 2007).
N° da semana com liberação de T. galloi
N° de massas de ovos de D. saccharalis
N° de massas de ovos de Diatraea saccharalis
predadas
Predação (%)
1° semana 76 46 60,5 2° semana 75 30 40,0 3° semana 45 23 51,1 Total 196 99 50,5
A interferência de predadores na eficiência dos parasitóides no campo está relacionada com a não distinção dos primeiros grupos de insetos por outros que já estejam parasitados. Este fato foi verificado no trabalho de RODRIGUES et al. (2005), nos quais os predadores coccinelídeos Cycloneda sanguinea Mulsant e Hippodamia convergens Guérin-Méneville, do sirfídeo Pseudodorus clavatus Fabricius e de formigas do gênero Pheidole alimentaram-se tanto de pulgões sadios como daqueles que continha as larvas do parasitóide L. testaceipes no seu interior.

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
564
2.4.7 Preferência hospedeira
A especificidade dos parasitóides na escolha de seu hospedeiro está relacionada com a maior dependência da fisiologia e desenvolvimento de seus hospedeiros, principalmente os coinobiontes (PARRA & BENTO, 2006). Neste sentido, cada espécie de parasitóide possui uma preferência por um tipo de hospedeiro, a partir da qual estes desenvolveram mecanismos que os guiam até esses hospedeiros, através de sinais químicos, físicos e visuais. Estes mecanismos são fundamentais para a localização, a aceitação e reconhecimento dos hospedeiros (CORDEIRO, 2006).
A preferência por hospedeiro depende do alimento que este pode oferecer para as suas larvas, pois o tipo de hospedeiro pode alterar o ciclo de vida do parasitóide. E esta preferência pode ser refletida no comportamento de busca e tolerância às condições ambientais (CORDEIRO, 2006). O parasitóide Glyptapanteles muesebecki Blanchard (Hymenoptera: Braconidae) parasita preferencialmente a lagarta do trigo Mythimna (Pseudaletia) sequax Franclemont (Lepidoptera: Noctuidae) (FOERSTER et al., 2001). Já o parasitóide L. testaceipes Cresson demonstra preferência hospedeira por A. gossypii (RODRIGUES et al., 2005) 2.4.8 Número de insetos liberados
Os fatores que determinam o número de parasitóide a ser liberado no campo estão relacionados com a fenologia da planta, espécie ou linhagem do parasitóide, condições climáticas, bem como a intensidade de infestação da praga no campo e densidade de plantio (SÁ & PARRA, 1993).
Segundo PEREIRA (2007) o número do Telenomus remus (Hymenoptera: Scelionidae) a ser liberado no campo é 3000 indivíduos por hectare, sendo considerado suficiente para diminuir ou pelo menos manter a população de Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) em índices baixos na cultura do milho. As liberações massivas deste parasitóide nesta cultura devem ser feitas quando 10% das folhas estiverem infestadas com massas de ovos ou quando o cultivo estiver na fase de duas a três folhas. Se as infestações forem superiores a 10%, recomenda-se uma liberação em grande escala ou inundativa dos parasitóides.
Trabalhos desenvolvidos por PASTORI et al. (2008) foi observado o parasitismo diário de ovos de Bonagota salubricola Meyrick (Lepidoptera: Tortricidae) em função do número de Trichogramma pretiosum Riley (Hymenoptera: Trichogramamatidae) em pomares de macieiras. Os autores concluíram que a liberação de 150.000 parasitóides por hectare resultava na maior taxa de parasitismo diário de ovos. 2.4.9 Condições climáticas
Os insetos são seres pecilotérmicos, ou seja, seu metabolismo sofre influência das condições climáticas. Consequentemente, a sua sobrevivência é drasticamente afetada pelos fatores abióticos. Muitas vezes, se as condições climáticas forem favoráveis à sobrevivência do agente biológico, a técnica de liberação não interfere na taxa de parasitismo (PINTO & PARRA, 2002). Muitos autores recomendam que a liberação dos parasitóides no campo deva ocorrer nos períodos mais frescos do dia, evitando-se dias chuvosos ou com muito vento (BOTELHO & MACEDO, 2002; PINTO et al., 2006; LEAL, 2007; BUENO, 2008).
ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
565
O comportamento de busca dos parasitóides pelo seu hospedeiro é influenciado por fatores adquiridos, genéticos, fisiológicos e ambientais. Este último inclui fatores bióticos como estímulos que conduzem o parasitóide ao habitat do seu hospedeiro, assim como os fatores abióticos, como temperatura, velocidade do vento e a chuva. A temperatura afeta o desenvolvimento durante o ciclo de vida, na viabilidade dos ovos e na razão sexual dos insetos. O vento é um agente regulador de sua dispersão no campo e a chuva pode prejudicar a metodologia de soltura destes insetos no campo (SILVA, 2007b).
ROUSSE et al (2009) avaliaram a influência de fatores abióticos (temperatura, umidade, pressão atmosférica, intensidade de luz e velocidade do vento) na capacidade de vôo e na taxa de parasitismo de Fopius arisanus Sonan (Hymenoptera: Braconidae) para o controle da mosca das frutas Bactrocera zonata Saunders (Diptera: Tephritidae). Os autores concluíram que a temperatura e a umidade influenciaram na atividade de vôo e na taxa de parasitismo deste inseto. A pressão atmosférica influenciou na atividade de vôo e os demais parâmetros não exerceram influência no comportamento deste parasitóide de ovo-pupa (Figura 8).
FIGURA 8. Influência da temperatura, umidade e pressão atmosférica na
atividade de vôo de fêmeas de Fopius arisanus. Dados coletados no período diurno. (ROUSSE et al., 2009)
3 CONSIDERAÇÕES FINAIS
O uso do controle biológico com parasitóides para o manejo de insetos-pragas na agricultura é uma prática sustentável muito utilizada nos dias atuais. Porém, tem-se tornado evidente que para explorar e aumentar a eficiência deste grupo de insetos em controlar pragas agrícolas é antes necessário o conhecimento de inúmeros fatores que interferem na sua liberação no campo. A falta de estudos sobre os aspectos da liberação afeta não somente os resultados finais de um programa de controle biológico, mas também, muitas vezes, as fases iniciais do projeto.
Fatores relacionados com a espécie utilizada, a fenologia da cultura e as condições climáticas em cada região devem ser observados antes e após a liberação dos parasitóides no campo. Neste sentido, para maior eficiência destes insetos, alguns cuidados devem ser tomados ao se planejar um programa de controle biológico e ter maior sucesso na sua utilização.

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
566
REFERÊNCIAS BIBLIOGRÁFICAS
ALTIERI, M. A. The ecological role of biodiversity in agroecosystems. Agriculture, Ecosystems and Environment, v. 74, p. 19-31, 1999. ANDOW, D. A.; PROKRYM, D. R. Plant structural complexity and host-finding by a parasitoid. Oecologia, Berlin, v. 82, p. 162-165, 1990. ARRUDA, L. H. Compostos orgânicos voláteis (VOCs) de plantas nas interações inseto-planta. 2009. Artigo em PDF. Disponível em: <www.genetica.esalq.usp.br/pub/LHArruda-200901-Resumo.pdf>. Acesso em: 04 set. 2011. BASTOS, C. S.; GALVÃO, J. C. C.; PICANÇO, M. C.; CECON, P. R.; PEREIRA, P. R. G. Incidência de insetos fitófagos e de predadores no milho e no feijão cultivados em sistema exclusivo e consorciado. Ciência Rural , Santa Maria, v. 33, n. 3, p. 391- 397, 2003. BOIÇA JR., A. L.; LARA, F. M.; BELLODI, M. P. Influência de variedades de cana-de-açúcar, incorporadas em dieta artificial, no desenvolvimento de Diatraea saccharalis (Fabr.) e no seu parasitismo por Cotesia flavipes (Cam.). Anais da Sociedade Entomológica , Londrina, v. 26, n. 3, p. 537- 542, 1997. BOTELHO, P. S. M.; MACEDO, N. Cotesia flavipes para o controle de Diatraea saccharalis. In: PARRA, J. R. P.; BOTELHO, P. S. M.; CORRÊA-FERREIRA, B. S.; BENTO, J. M. S. (Ed.). Controle biológico no Brasil: parasitóides e predadores. Barueri, SP: Manole, 2002. cap. 25, p. 409-425. BOTELHO, P. S. M.; PARRA, J. R. P.; CHAGAS NETO, J. F.; OLIVEIRA, C. P. B. Associação do parasitóide de ovos Trichogramma galloi Zucchi (Hymenoptera: Trichogrammatidae) e do parasitóide larval Cotesia flavipes (Cam.) (Hymenoptera: Braconidae) no controle de Diatraea saccharalis (Fabr.) (Lepidoptera: Crambidae) em cana-de-açúcar. Anais da Sociedade Entomológica do Brasil , Londrina, v. 28, n. 3, p. 491-496, 1999. BOTELHO, P. S. M.; PARRA, J. R. P.; MAGRINI, E. A.; HADDAD, M. L.; RESENDE, L. C. L. Efeito do número de liberações de Trichogramma galloi (Zucchi, 1988) no parasitismo de ovos de Diatraea saccharalis (Fabr., 1794). Scientia Agricola , Piracicaba, v. 52, n. 1, p. 65- 69, 1995. BROGLIO-MICHELETTI, S. M. F.; PEREIRA-BARROS, J. L.; SANTOS, A. J. N.; CARVALHO, L. W. T.; CARVALHO, L. H. T.; OLIVEIRA, C. J. T. Efeito do número de adultos de Trichogramma galloi Zucchi, 1988 (Hymenoptera: Trichogrammatidae) liberados em semanas sucessivas, para o controle de Diatraea saccharalis (Fabricius, 1794) (Lepidoptera: Crambidae). Ciência e Agrotecnologia , v. 31, n. 1, p. 53-58, 2007. BUENO, R. C. O. F. Bases biológicas para a utilização de Trichogramma pretiosum Riley, 1879 (Hymenoptera: Trichogrammatidae) para o controle de

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
567
Pseudoplusia includens (Walker, 1857) e Anticarsia gemmatalis Hübner, 1818 (Lepidoptera: Noctidae) em soja. 119 f. Tese (Doutorado em Ciências: Entomologia) - Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, Piracicaba, 2008. CAMPOS, W. G.; ARAÚJO, E. R. Ecologia de insetos parasitóides e o controle biológico de pragas. Vertentes , n. 4, p. 79-93, 1994. CANTO-SILVA, C. R.; KOLBERG, R.; ROMANOWSKI, H. P.; REDAELLI, L. R. Dispersal of the egg parasitoid Gryongallardoi (Brethes) (Hymenoptera: Scelionidae) in tobacco crops. Brazilian Journal of Biology, São Carlos, v. 66, n. 1A, p. 9-17, 2006. CAVALCANTI, M. DAS G.; VILELA, E. F.; EIRAS, A. E.; ZANUNCIO, J. C.; PICANÇO, M. C. Interação tritrófica entre Podisus nigrispinus (Dallas) (Heteroptera: Pentatomidae), Eucalyptus e lagartas de Thyrinteina arnobia (Stoll) (Lepidoptera: Geometridae): I Visitação. Anais da Sociedade Entomológica do Brasil , Londrina, v. 29, n. 4, p. 697- 703, 2000. CAVALCANTI, R. S.; MOINO JUNIOR, A.; LOUREIRO, E. S.; MENDONÇA, L. A.; ARNOSTI, A. Determinação da época de liberação de Lysiphlebus testaceipes (Cresson, 1880) (Hymenoptera: Aphidiidae) após aplicação de diferentes fungos entomopatogênicos para o controle integrado de Schizaphis graminum (Rond., 1852) (Hemíptera: Aphididae). Arquivo do Instituto Biológico , São Paulo, v.76, n.2, p.251-257, 2009. CORDEIRO, E. S. Liberação de linhagens de Trichogramma pretiosum para controle biológico de Spodoptera frugiperda na cultura do milho. 63 f. Dissertação (Mestrado em Agronomia) - Centro de Ciências Agrárias, Universidade Estadual do Oeste do Paraná, Marechal Cândido Rondon, 2006. FATOUROS, N. E.; DICKE, M..; MUMM, R.; MEINERS, T.; HILKER, M. Foraging behavior of egg parasitoids exploiting chemical information. Behavioral Ecology, New York, v. 19, n. 3, p. 677-689, 2008. FERRER, F. Biological control of agricultural insect pests in Venezuela; advances, achievements, and future perspectives. Biocontrol News and Information , v. 22, n. 3, p. 67-74, 2001. FOERSTER, L. A.; DOETZER, A. K.; AVANCI, M. R. F. Parasitóides larvais de Mythimna (Pseudaletia) sequax (Franclemont) e capacidade de parasitismo de Glyptapanteles muesebecki (Blanchard) em relação ao tempo de exposição, temperatura e densidade de hospedeiros. Acta Biológica Paranaense, Curitiba, v. 30, nos. 1, 2, 3, 4; p. 139-149, 2001. GARCIA, M. A. Ecologia nutricional de parasitóides e predadores terrestres. In: PANIZZI, A. R.; PARRA, R. P. Ecologia nutricional de insetos e suas implicações no manejo de pragas. São Paulo, SP: Manole, 1991. cap. 8, p. 289-311.

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
568
GICHINI, G.; LOHRA, B., ROSSBACHB, A. NYAMBOA, B.; GATHU, B. Can low release numbers lead to establishment and spread of an exotic parasitoid: The case of the diamondback moth parasitoid, Diadegma semiclausum (Hellén), in East Africa. Crop Protection , v. 27, p. 906-914, 2008. GINGRAS, D.; DUTILLEUL, P.; BOIVIN, G. Modeling the impact of plant structure on host-finding behavior of parasitoids. Oecologia , Marburg, v. 130, p. 396-402, 2002. HAJI, F. N. D.; PREZOTTI, L.; CARNEIRO, J. S.; ALENCAR, J. A. Trichogramma pretiosum para o controle de pragas no tomateiro industrial. In: PARRA, J. R. P., P. S. M. BOTEHO, B. S. CORRÊA-FERREIRA E J. M. S. BENTO (Eds). Controle biológico no Brasil : parasitóides e predadores. São Paulo: Manole, 2002. cap. 28, p. 477- 494. HEIL, M. Indirect defence via tritrophic interactions. New Phytologist, Chicago, v. 178, p. 41-61, 2007. JARAMILLO, J.; CHABI-OLAYE, A.; BORGEMEISTER, C.; KAMONJO, C.; POEHLING, H. M.; VEJA, F. E. Where to sample? Ecological implications of sampling strata in determining abundance and impact of natural enemies of the coffee berry borer, Hypothenemus hampei. Biological Control , v. 49, p. 245-253, 2009. LEAL, W. C. O. Controle biológico da broca-da-cana. 2007. Disponível em: <www.criareplantar.com.br/noticia/ler/?idNoticia>. Acesso em: 24 set. 2011 MARCHIORI, C. H.; LELES, A. S.; BARBARESCO, L. F.; FERREIRA, M. M. Parasitóides de dípteros coletados em Itumbiara, GO, e Tupaciguara, MG, Brasil. Arquivo Instituto Biológico , v.73, n.3, p.371-374, 2006. MARCHIORI, C. H.; LUSSARI, M. A.; ROSA, D. C.; PENTEADO-DIAS, A. M. Parasitoid Hymenoptera collected during the diurnal and nocturnal periods in Itumbiara, Goiás. Brazilian Journal of Biology, v. 67, n. 3, p. 581-582, 2007. NAVA, D. E. Controle biológico de insetos-praga em frutíferas d e clima temperado : uma opção viável, mas desafiadora (Documentos 2008). Pelotas: Embrapa Clima Temperado, 2007. 20 p. PACHECO, D. J. P.; CORRÊA-FERREIRA, B. S. Parasitismo de Telenomus podisi Ashmead (Hymenoptera: Scelionidae) em populações de percevejos pragas da soja. Anais da Sociedade Entomológica do Brasil , Londrina, v. 29, n. 2, p. 295-302, 2000. PARRA, J. R. P.; BENTO, J. M. S. Parasitóides e predadores no controle biológico de insetos-praga. In: OLIVEIRA-FILHO, E. C.; MONNERAT, R. G. Fundamentos para a regulação de semioquímicos, inimigos naturai s e agentes microbiológicos de controle de pragas. Planaltina, DF: Embrapa Cerrados, 2006. cap. 5, p. 103-119. PARRA, J. R. P.; ZUCCHI, R. A. Trichogramma in Brazil: Feasibility of use after twenty years of research. Neotropical Entomology , v.33, n.3, p. 271-281, 2004.

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
569
PARRA, J. R. P. Etapas básicas para programas de controle biológico com Trichogramma no Brasil. In: SIMPÓSIO DE CONTROLE BIOLÓGICO, 5., 1996, Foz do Iguaçu. Anais … Foz do Iguaçu. p. 353- 354. PASTORI, P. L.; MONTEIRO, L. B.; BOTTON, M.; SOUZA, A.; POLTRONIERI, A. S.; SCHUBER, J. M. Parasitismo de ovos da lagarta-enroladeira-da-maçã em função do número de Trichogramma pretiosum Riley (Hymenoptera: Trchogrammatidae) liberado. Scientia Agraria , Curitiba, v. 9, n. 4, p. 497-504, 2008. PEREIRA, L. G. B. Táticas de controle da lagarta-do-cartucho do milho , Spodoptera frugiperda. Fundação Centro Tecnológico de Minas Gerais: Dossiê Técnico, 2007. 28 p. PINTO, A. S.; PARRA, J. R. P. Liberação de inimigos naturais. In: PARRA, J. R. P.; BOTELHO, P. S. M.; CORRÊA-FERREIRA, B. S.; BENTO, J. M. S. (Ed.). Controle biológico no Brasil: parasitóides e predadores. Barueri, SP: Manole, 2002. cap. 19, p. 325-342. PINTO, A. S.; PARRA, J. R. P.; OLIVEIRA, H. N.; ARRIGONI, E. D. B. Comparação de técnicas de liberação de Trichogramma galloi Zucchi (Hymenoptera: Trichogrammatidae) para o controle de Diatraea saccharalis (Fabricius) (Lepidoptera: Crambidae). Neotropical Entomology, Londrina, v. 32, n. 2, p. 311-318, 2003. PINTO, A. S.; GARCIA, J. F.; OLIVEIRA, H. N.; ALMEIDA, J. E. M; ALVES, S. B. Manejo das principais pragas da cana-de-açúcar. In: SEGATO, S. V.; PINTO, A. S.; JENDIROBA, E.; NÓBREGA, J. C. M. (Ed.). Atualização em produção de cana-de-açúcar. Piracicaba, SP: ESALQ/USP, 2006. cap. 15, p. 257-280. PRATISSOLI, D.; FORNAZIER, M. J.; HOLTZ, A. M.; GONÇALVES, J. R.; CHIORAMITAL, A. B.; ZAGO, H. Ocorrência de Trichogramma pretiosum em áreas comerciais de tomate, no Espírito Santo, em regiões de diferentes altitudes. Horticultura Brasileira , v. 21, n. 1, p. 73-76, 2002. PRATISSOLI, D.; VIANNA, U. R.; ZAGO, H. B.; PASTORI, P. L. Capacidade de dispersão de Trichogramma em tomateiro estaqueado. Pesquisa agropecuária brasileira , Brasília, v. 40, n. 6, p.613-616, 2005. RODRIGUES, S. M. M.; BUENO, V. H. P.; SAMPAIO, M. V. Efeito da liberação inoculativa sazonal de Lysiphlebus testaceipes (Hymenoptera: Aphidiidae) na população de Aphis gossypii (Hemiptra: Aphididae) em cultivo de crisântemo em casa de vegetação comercial. Boletín de Sanidad Vegetal y Plagas , Madrid, v. 31, p. 199-207, 2005. ROUSSE, P.; GOURDON, F.; ROUBAUD, M.; CHIROLEU, F.; QUILICI, S. Biotic and abiotic factors affecting the flight activity of Fopius arisanus, an egg-pupal parasitoid of fruit fly pests. Environmental Entomology , Lanham, v. 38, n. 3, p. 896-903, 2009.

ENCICLOPÉDIA BIOSFERA, Centro Científico Conhecer - Goiânia, vol.7, N.13; 2011 Pág.
570
SÁ, L. A. N.; PARRA, J. R. P. Efeito do número e intervalo entre liberações de Trichogramma pretiosum Riley no parasitismo e controle de Helicoverpa zea (Boddie), em milho. Scientia Agricola , Piracicaba, v. 50, n. 3, p. 355- 359, 1993. SÁ, L. A. N.; PARRA, J. R. P.; SILVEIRA NETO, S. Capacidade de dispersão de Trichogramma pretiosum (Riley, 1879) para controle de Helicoverpa zea (Boddie, 1850) em milho. Scientia Agricola , Piracicaba, v. 50, n. 2, p. 226-231, 1993. SANTOS, M. C. P. Diversidade de vespas parasitóides (Himenóptera: Pa rasítica) em áreas de cultivo de café ( Coffea arabica) em uma área de vegetação nativa localizada nos municipios de Piatã, Chapada Diamant ina, Bahia. 69 f. Dissertação (Mestrado em Agronomia: Fitotecnia) - Universidade Estadual do Sudoeste da Bahia, Vitória da Conquista, 2008. SCATOLINI, D.; PENTEADO-DIAS, A. M. Análise faunística de Braconidae (Hymenoptera) em três áreas de mata nativa do Estado do Paraná, Brasil. Revista Brasileira de Entomologia , v. 47, n. 2, p. 187-195, 2003. SILVA, A. F. Seleção, biologia e exigências térmicas de Trichogramma sp. criados em ovos de Diaphania nitidalis Cramer (Lepidoptera: Pyralidae). 44 f. Dissertação (Mestrado em Produção Vegetal) – Centro de Ciências Agrárias, Universidade Federal do Espírito Santo, Alegre, 2007a. SILVA, C. S. B. Dispersão do parasitóide de ovos Telenomus remus Nixon (Hymenoptera: Scelionidae) e sua interação com algu mas variáveis ambientais em agroecossistemas de algodoeiro (Gossypium hirsut um L). 139 f. Dissertação (Mestrado em Agronomia: Entomologia Agrícola) - Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal, 2007b. SILVA, C. C.; MORAES, M. C. B.; LAUMANN, R. A.; BORGES, M. Sensory response of the egg parasitoid Telenomus podisi to stimuli from the bug Euschistus heros. Pesquisa Agropecuária Brasileira , Brasília, v. 41, n. 7, p.1093-1098, 2006. SIQUEIRA, K. M. M.; FARIAS, A. M. I. Resposta de fêmeas de Encarsia formosa Gahan (Hymenoptera: Aphelinidae) aos odores do hospedeiro e da planta-hospedeira em olfatômetro de quatro vias. Bragantia , Campinas, v. 62, n.3, p.447-450, 2003. STEFANELO, E. L. Agronegócio Brasileiro: Propostas e tendências. FAE BUSINESS, Curitiba, n. 3, p. 10-13, 2002. STONER, K. Approaches to the Biological Control of Insects. Bulletin 7144. Disponível em: <http://www.umext.maine.edu/onlinepubs/htmpubs/7144.htm>. Acesso em: 04 set. 2011. ZACHRISSON, B.; PARRA J. R. P. Capacidade de dispersão de Trichogramma pretiosum Riley, 1879 para o controle de Anticarsia gemmatalis Hübner, 1818 em soja. Scientia Agrícola , Piracicaba, v. 55, n. 1, 1998.





























