Embed Size (px)
Citation preview

Expression of Human Apolipoprotein A-I/C-III/A-IV GeneCluster in Mice Induces Hyperlipidemia but
Reduces AtherogenesisLaurent Vergnes, Nadine Baroukh, Maria A. Ostos, Graciela Castro, Nicolas Duverger,
M. Nazeem Nanjee, Jamila Najib, Jean-Charles Fruchart, Norman E. Miller,Mario M. Zakin, Alberto Ochoa
Abstract—The apolipoprotein (apo)A-I/C-III/A-IV gene cluster is involved in lipid metabolism and atherosclerosis.Overexpression of apoC-III in mice causes hypertriglyceridemia and induces atherogenesis, whereas overexpression ofapoA-I or apoA-IV increases cholesterol in plasma high density lipoprotein (HDL) and protects against atherosclerosis.Each gene has been studied alone in transgenic mice but not in combination as the entire cluster. To determine whichphenotype is produced by the expression of the entire gene cluster, transgenic mice were generated with a 33-kb humanDNA fragment. The results showed that the transgene contained the necessary elements to direct hepatic and intestinalexpression of the 3 genes. In the pooled data, plasma concentrations were 25769, 7.160.5, and 1.060.2 mg/dL forhuman apoA-I, apoC-III, and apoA-IV, respectively (mean6SEM). Concentrations of these apolipoproteins were higherin males than in females. Human apoA-I and apoC-III concentrations were positively correlated, suggesting that theyare coregulated. Transgenic mice exhibited gross hypertriglyceridemia and accumulation of apoB48–containingtriglyceride-rich lipoproteins. Plasma triglyceride and cholesterol concentrations were correlated positively with humanapoC-III concentration, and HDL cholesterol was correlated with apoA-I concentration. In an apoE-deficientbackground, despite being markedly hypertriglyceridemic, cluster transgenic animals compared with nontransgenicanimals showed a 61% reduction in atherosclerosis. This suggests that apoA-I and/or apoA-IV can protect againstatherosclerosis even in the presence of severe hyperlipidemia. These mice provide a new model for studies of theregulation of the 3 human genes in combination.(Arterioscler Thromb Vasc Biol. 2000;20:2267-2274.)
Key Words: transgenic micen hypertriglyceridemian cholesteroln lipoproteinsn atherosclerosis
Plasma lipoproteins influence the development of athero-sclerosis, and their concentrations are associated with the
risk of coronary heart disease.1 The genes for 3 apolipopro-teins, apoA-I, apoC-III, and apoA-IV, are grouped together ina cluster on 17 kb of human chromosome 11.2
ApoA-I is the major protein component of HDL. Throughits ability to promote cholesterol efflux from cultured cellsand to activate lecithin:cholesterol acyltransferase (LCAT), itis involved in reverse cholesterol transport (RCT).1 Expres-sion of the human apoA-I (hapoA-I) gene decreases thedevelopment of fatty lesions in cholesterol-fed transgenic(Tg) mice3 and in apoE-deficient (apoE2/2) mice.4 ApoA-Ideficiency in mice did not increase atherogenesis in a normalbackground5 but did so in hypercholesterolemic mice ex-pressing human apoB.6
ApoC-III is a component of HDL and triglyceride-richlipoproteins (TGRLs). Plasma apoC-III concentration is pos-
itively correlated with triglyceride concentration. By inhibit-ing TGRL catabolism, apoC-III induces hypertriglyceridemiain Tg mice.7 Expression of the gene was not atherogenic inapoE2/2 mice.8 In contrast,hapoC-III expression, in normal orin LDL receptor–deficient mice, resulted in increasedatherosclerosis.9,10
ApoA-IV, involved in RCT-like apoA-I,1 also has anti-atherogenic properties, inasmuch as overexpression ofapoA-IV reduced aortic lesions.11,12 The foregoing observa-tions indicate that the apoA-I/C-III/A-IV gene cluster is ofmajor interest in relation to atherogenesis.
A series of DNA elements seems to coordinately regulatethe transcription of all 3 genes.13–15 Indeed, analyses ofintestinal mRNA levels have shown that the expression of the3 genes is coregulated in this organ in vivo.16 Tg mice havealready been developed for apoA-I,3,13,17 apoC-III,8,13,18 andapoA-IV11,12,19,20and have provided valuable information on
Received November 3, 1999; revision accepted February 14, 2000.From the Unite d’Expression des Genes Eucaryotes, Institut Pasteur (L.V., N.B., M.A.O., M.M.Z., A.O.), Paris, France; U 325 Inserm Institut Pasteur
(G.C., J.N., J.-C.F.), Lille, France; Rhone-Poulenc Rorer (N.D.), Vitry sur Seine, France; Institut National de la Recherche Agronomique (A.O.)Jouy-en-Josas, France; and St. Bartholomew’s and the Royal London School of Medicine and Dentistry (M.N.N., N.E.M.), London, UK.
L.V. and N.B. contributed equally to this work.Correspondence to Alberto Ochoa, Unite d’Expression des Genes Eucaryotes, Institut Pasteur, 28 rue du Dr. Roux, 75724, Paris cedex 15, France.
E-mail [email protected]© 2000 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol.is available at http://www.atvbaha.org
2267
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from

the properties and functions of each gene in isolation.However, none of these animal models is suitable for studiesof the effect of the coordinated expression of the 3 genes incombination. Moreover, new therapeutic agents that increaseapoA-I concentration are currently being sought. Whethersuch agents will change the expression of the other genes ofthe cluster and produce an atherogenic or a nonatherogeniceffect is not known. A model expressing the entire humanapoA-I/C-III/A-IV cluster would be of considerable value forthe investigation of its properties and functions as an inte-grated unit. In the present study, we report the generation andcharacterization of such Tg mice.
MethodsAnimalsThe transgene was obtained from the DMPC-HFF#1-1412-B12cosmid isolated from a human genomic library (DuPont MerckPharmaceutical Co) screened for the presence of apoA-IV sequencesby polymerase chain reaction. A 33-kbNotI/SalI fragment, contain-ing 8.3 kb of the 59region of apoA-I, the 17-kb entire apoA-I/C-III/A-IV gene cluster, and 7.5 kb of the 39region of apoA-IV, wasmicroinjected into fertilized mouse eggs. Tg mice were identified bythe presence of the 3 human genes. Three Tg lines were establishedin the C57Bl/6 genetic background (Tg11, Tg12, and Tg21). Trans-gene copy numbers were determined by Southern blotting.20 Mice,housed in a temperature-controlled room with alternating 12-hourlight (7:00AM to 7:00PM) and dark (7:00PM to 7:00AM) periods, hadaccess to regular mouse chow and water ad libitum. All proceduresinvolving animal handling and care were conducted in accordancewith Pasteur Institute Guidelines for Husbandry of Laboratory Mice.Tg mice in an apoE2/2 background were established by backcrossingTg12 animals with apoE2/2 mice.21 For blood collection, food wasremoved from mice at 9:00AM, and 8 hours later, samples werecollected from the retro-orbital sinus into tubes containing Na2-EDTA (final concentration 5 mmol/L). Plasma was promptly sepa-rated from cells by centrifugation at 5000g at 4°C.
Northern Blot ExperimentsAfter an overnight fast, the animals were killed by cervical disloca-tion, and tissues (liver, small intestine, spleen, kidney, lung, brain,and stomach) were removed. Total RNAs were extracted withRNA-plus (Quantum), purified, and electrophoresed (20mg totalRNA per lane) on 1% agarose gels containing formaldehyde.Samples were blotted onto Hybond-N1 membranes (AmershamPharmacia Biotech) and hybridized with32P-labeled cDNA probes,as previously described.7,11,22
Apolipoprotein QuantificationHuman apolipoproteins were quantified by immunoelectrophoresiswith specific polyclonal antibodies (Hydragels SEBIA); mouseapolipoproteins did not cross-react. Mouse plasma apoA-I concen-trations were measured by immunonephelometry with the use ofrabbit polyclonal antibodies. These antibodies showed no cross-species reactivity.23
Human apolipoproteins in high-performance–sized exclusionchromatographic (HP-SEC) fractions were quantified by liquid-phase double-antibody–based radioimmunoassays, with the use ofTween 20 (0.2% [vol/vol]) to expose cryptic epitopes and polyeth-ylene glycol 8000 (3% [wt/vol]) to enhance reaction kinetics. Theprimary antisera were polyclonal IgG against delipidated humanapolipoproteins (goat anti–apoA-I and goat anti–apoC-III, providedby International Immunology Corp; rabbit anti–apoA-IV11) thatdisplayed insignificant cross-reactivity with their mouse counter-parts. Donkey anti-goat or donkey anti-rabbit IgGs were used asprecipitating antibodies (Chemicon). Radioactivity in antibody-bound pellets (1500g, 30 minutes, 4°C) was quantified to,0.1%counting error. Radioiodinated tracers were prepared by use ofchloramine T treatment of either delipidated human apolipoproteins(A-I and C-III, Intracel Corp) or recombinant apoA-IV. All assays
were standardized by using appropriate dilutions of the pure apoli-poproteins. All plasma samples were analyzed in duplicate withcoefficients of variation,12%.
ApoB48 and apoB100 were separated by SDS-PAGE in 4% to 15%gradient polyacrylamide PhastGels (Amersham Pharmacia Biotech).Gels were silver-stained, scanned, and analyzed by use of a videodensitometer scanner and NIH image software. Analyses wererepeated 3 times for each different group of animals.
Lipid and Lipoprotein AnalysesLipoproteins were separated into density (d) fractions ofd,1.006g/mL, d51.006 to 1.019 g/mL, andd51.019 to 1.063 g/mL bysequential isopycnic ultracentrifugation with use of KBr to adjustdensities. Each spin was carried out for 2 hours at 435 000gat 15°Cin a fixed-angle rotor.24
HP-SEC was used to separate lipoproteins according to size.Plasma aliquots (50mL) were chromatographed at ambient temper-ature through Superdex 200 and Superdex 75 columns connected inseries (HR 10/30, Amersham Pharmacia Biotech). A solution of50 mmol/L Tris-HCl (pH 7.4), 150 mmol/L NaCl, 1 mg/mL sodiumazide, and 1 mg/mL Na2-EDTA was used as eluant (pumped at 0.75mL/min). After the void volume was excluded, 603225-mL fractionswere collected. Recoveries were always.90%. Apparent hydratedsizes of lipoprotein subclasses were evaluated by comparing theirelution times with those of protein markers (Amersham PharmaciaBiotech).
HDL particle size and heterogeneity were evaluated by electro-phoresing whole plasma in commercially available nondenaturingpolyacrylamide gradient gels (Lipogel Lipoprotein FractionationSystem, Zaxis Inc), according to the manufacturer’s instructions.After electrophoretic separation for'1000 volts/hour in a Tris-borate-EDTA buffer system, the distribution of neutral lipids wasdetected by staining with Sudan black B.
Nondenaturing 2D electrophoresis and Western blotting wereperformed as previously described.25 After the 2 electrophoreses,lipoproteins were electrotransferred on to nitrocellulose membranes.The filters were immunodecorated with rabbit anti-mouse or anti-human apoA-I, apoC-III, or apoA-IV antibodies, followed by sec-ondary peroxidase–labeled antibodies directed against rabbit IgG.The antisera directed against human or mice apolipoproteins werehighly specific and did not show any cross-species reactivity, withthe exception of the anti-mouse apoC-III, which recognized humanapoC-III. Membranes were developed by use of a Chemilumines-cence Western blotting detection kit (ECL System, AmershamPharmacia Biotech). Experiments were repeated 3 times with sam-ples from different control and Tg animals.
Triglyceride concentrations were determined by an enzymaticGPO-PAP method, and total and HDL cholesterol concentrationswere determined by enzymatic CHOD-PAP methods with commer-cial kits (Boehringer-Mannheim). Precinorm L (Boehringer-Mannheim) was used as a calibrator.
Cholesterol Efflux and Esterification StudiesCellular cholesterol efflux was determined with rat Fu5AH hepatomacells incubated with 5% diluted serum.23 Values are reported as theaverage of at least 3 different determinations. Purified human HDLsamples were included in each assay as positive controls. LCATactivity was determined by use of the exogenous proteoliposomesubstrate method with 25mL of plasma from female mice.23
Experiments were repeated twice with 3 and 5 different pools ofsamples for control and Tg mice, respectively (coefficient of varia-tion ,5%). Cholesterol esterification rate was determined as thedecrease in unesterified cholesterol mass (measured enzymatically)during in vitro incubation of whole plasma at 37°C for periods up to4 hours. Only female mouse plasma was analyzed, and all measure-ments were made in triplicate (coefficient of variation,1.5%).
Evaluation of Atherosclerotic LesionsThe spontaneous atherosclerosis model used in the present study wasthe apoE2/2 mouse.21 Fatty streak lesions were quantified by evalu-ating cross-sectional lesion sizes in the aortic sinus.26 A randomlychosen subset of 10 Tg and 11 control female mice, both with the
2268 Arterioscler Thromb Vasc Biol. October 2000
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from

apoE2/2 background, were euthanized at 20 weeks of age. Hearts andproximal aortas were removed, cleaned of pericardial fat, and fixedin PBS formalin solution (5%) for at least 48 hours. Then the heartswere cut directly under and parallel to the aortic leaflet, and theupper portions were embedded in Tissue-Tek O.C.T. compound(Sakura Finetek) and frozen at280°C. Ten-micron-thick sectionswere cut through the aortic sinus, recognized by 3 valve cusps at thejunction of the left ventricle and the aorta. Eighty sections per animalwere stained for lipids with oil red O and counterstained withhematoxylin. Cross-sectional areas were analyzed by computerizedplanimetry. Statistical analysis was performed on 4 different cross-sectional lesion areas per animal, separated by 160mm. The firstsection analyzed for each animal corresponded to the lower origin ofthe aortic sinus.
Statistical AnalysesResults are reported as mean6SEM In all experiments non-Tglittermates were used as controls. Statistical differences in apoli-poprotein and lipid levels between different groups of animals wereevaluated by ANOVA. Other statistical differences were tested bythe Studentt test or Mann-WhitneyU test for nonparametricanalysis. Group differences or correlations withP,0.05 wereconsidered statistically significant.
ResultsApoA-I/C-III/A-IV Transgene ExpressionTg mice were generated with a 33-kb human genomicfragment containing the entire cluster and flanking sequences.Three Tg lines, Tg11, Tg12, and Tg21 containing'9, 1, and2 copies, respectively, of the transgene, were established. AllTg lines showed hepatic and intestinal expression of the 3human genes (Figure 1). Human mRNA levels were higher in
the liver than in the intestine. No expression was detected inother analyzed tissues, with the exception of traces of all 3human mRNAs in the kidney, ofhapoA-I mRNA in the brain,spleen, and stomach, and ofhapoA-IV mRNA in the stomach(data not shown). Because apoA-IV expression was reportedto be mainly intestinal in humans and inhapoA-IV Tg mice,we have compared by Northern blot analysis the mRNAlevels in hapoA-IV Tg mice20 with those in the cluster Tgmice. Although the hepatic levels were similar in both strains,the intestinal mRNA levels were 2.3-fold lower in the clusterTg mice (15.563.2 arbitrary units forhapoA-IV Tg miceversus 6.664.8 arbitrary units for cluster Tg mice,P,0.05),suggesting intestinal downregulation in the latter.
Human Plasma ApolipoproteinsAll 3 human apolipoproteins were present in plasma fromeach line, and their concentrations were higher in Tg malesthan in Tg females (Table 1). The means in the different lineswere very similar. AveragehapoA-I levels ranged from 241 to283 mg/dL, with an overall mean for the 3 lines of 25769mg/dL (n547); averagehapoC-III levels were between 4.8and 8.8 mg/dL, with an overall mean of 7.160.5 mg/dL; and
hapoA-IV concentrations averaged 0.8 to 1.4 mg/dL, with anoverall mean of 1.060.2 mg/dL. The plasma concentration of
hapoA-I was positively correlated with that ofhapoC-III(r50.67, P,0.001). Because of the low concentration of
hapoA-IV, it was difficult to assess the strength of itsassociation with the concentrations ofhapoA-I or hapoC-III.
Figure 1. Human apoA-I/C-III/A-IV clus-ter transgene used to generate Tg miceand Northern blot analyses. A, Schemeof human genomic DNA fragment con-taining apoA-I/C-III/A-IV gene clusterand its 59 and 39 flanking regions usedto generate Tg mice. B, Average levelsof each human mRNA in Tg lines Tg11,Tg12, and Tg21. Open and filled barscorrespond to liver and intestinal levels,respectively. Values are reported asarbitrary units normalized to b-actinmRNA levels. In the apoC-III panel, theright scale corresponds to the intestinalmRNA levels. Means were obtainedfrom RNA preparations from at least 3different mice for each Tg line.
TABLE 1. Human Apolipoprotein Concentrations in Plasma of Tg Mice
Strain
Concentration, mg/dL
Human ApoA-I Human ApoC-III Human ApoA-IV
All Males Females All Males Females All Males Females
Tg11 241614 279615* 197612 4.860.5 6.260.6† 3.260.4 0.860.1 0.760.1 1.060.2
Tg12 247620 336629* 199611 8.861.1 13.561.5* 6.360.6 1.160.2 2.460.3* 0.660.1
Tg21 283608 300611 263609 7.360.4 8.260.3 6.360.7 1.460.5 2.560.8* 0.560.3
Tg12/apoE2/2 163613‡ 202644*‡ 10869‡ 17.462.6§ 26.261.3*‡ 5.360.5 1.960.4 2.960.5* 0.560.1
Values are mean6SEM (n54 to 11).*P,0.001 and †P,0.01 vs females; ‡P,0.001 and §P,0.01 vs Tg12.
Vergnes et al Human ApoA-I/C-III/A-IV Cluster Tg Mice 2269
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Lipid ConcentrationsMean plasma triglyceride and total cholesterol concentrationswere higher in Tg males than in Tg females (Table 2). Asimilar sex effect was seen in control littermates. Expressionof the cluster produced a 4- to 10-fold increase in triglycerideconcentration relative to those in control animals and amoderate increase in total cholesterol concentration duemostly to an increase in the non-HDL subclass. However,HDL cholesterol was also significantly raised in 3 of the 3 Tglines (Tg11 and Tg12). Plasma triglyceride concentrations inindividual mice correlated with non-HDL cholesterol concen-trations (r50.92,P,0.001). In addition,hapoC-III concentra-tions were positively correlated with triglyceride (r50.80,P,0.001) and non-HDL cholesterol (r50.64,P,0.001) con-centrations. These associations suggest that the increasedconcentrations of triglycerides and cholesterol in Tg micewere mainly dependent on the concentration ofhapoC-III.Relatively low concentrations ofhapoC-III were sufficient toinduce marked changes in the lipid profiles (compare data inTables 1 and 2). Finally,hapoA-I concentration was corre-lated with HDL cholesterol concentration (r50.66,P,0.001). Triglycerides showed a low but significant corre-lation with hapoA-I (r50.36, P50.012). The correlationbetween HDL cholesterol andhapoC-III was not significant(r50.14, P50.92), nor was the correlation between HDLcholesterol and triglyceride concentrations (r50.16,P50.28).
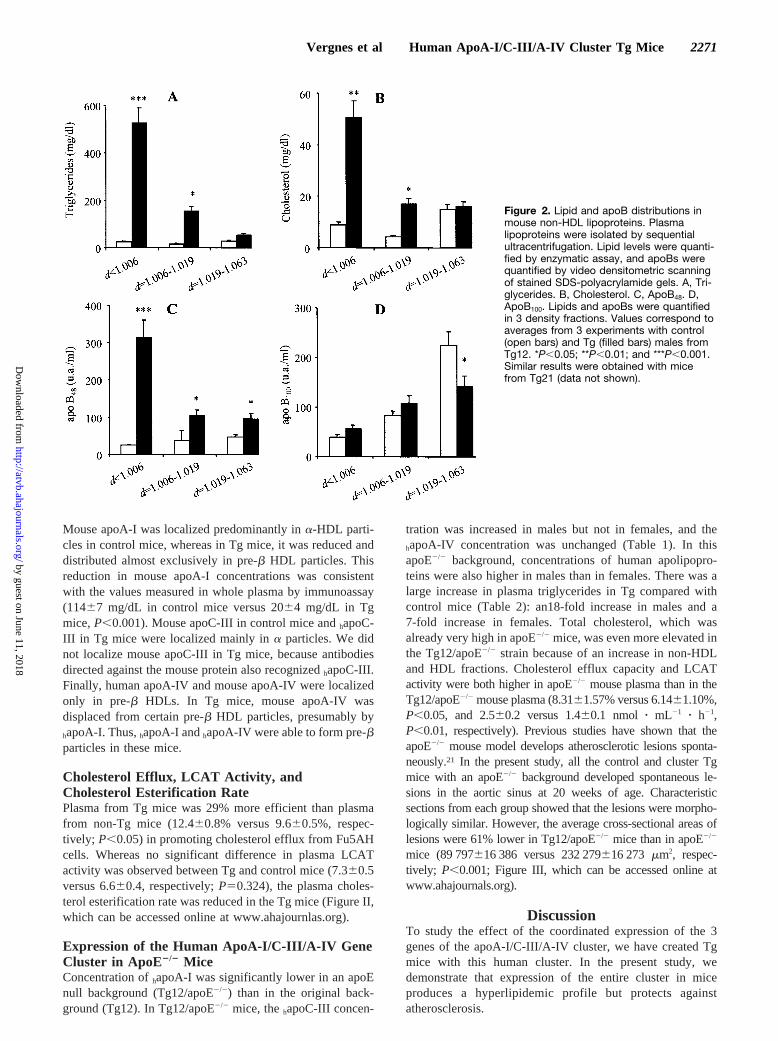
Lipid and ApoB Distributions in LipoproteinDensity FractionsLipoproteins were isolated by sequential ultracentrifugationfrom plasma samples from Tg mice of line 12 and controllittermates. Increases of triglycerides and cholesterol in Tgmice were observed in lipoproteins ofd,1.006 g/mL andd51.006 to 1.019 g/mL (Figure 2A and 2B). Analysis ofapoB showed a major increase in total apoB48, whereas totalapoB100 remained unchanged. The apoB48 increase in the Tgmice was detected in all fractions analyzed; in particular, an11-fold increase was observed in particles ofd,1.006 g/mL(Figure 2C). Although the total apoB100 level remainedunchanged, lipoprotein fraction analysis showed a significantdecrease in thed51.019 to 1.063 g/mL fraction in Tg mice(Figure 2D). Thus, the increases in triglycerides and choles-terol observed in Tg mice were due to an accumulation of
apoB48-rich lipoproteins ofd,1.006 g/mL. Similar resultswere obtained with mice from line 21.
Cholesterol and Apolipoprotein Distributions inSize Subclasses of LipoproteinsThe distributions of the cholesterol and human apolipopro-teins in size subclasses of lipoproteins were analyzed byHP-SEC of whole plasma. Results for representative maleand female animals are shown in Figure 3.hApoC-III andhapoA-IV were present in TGRLs plus LDLs (fractions 1 to14, molecular mass.670 kDa) as well as in lipid-rich HDLs(fractions 15 to 35, 100 to 670 kDa), whereashapoA-I wasalmost exclusively present in HDL particles of 70 to 350 kDa(fractions 20 to 40). In addition, a minor proportion ofhapoA-I and a substantial proportion ofhapoA-IV werepresent in small lipid-poor particles (fractions 40 to 50 of 35to 70 kDa and fractions 35 to 45 of 50 to 100 kDa). Male micehad significantly higher concentrations ofhapoA-IV in allfractions compared with females (0.8460.22 versus0.1060.03 mg/dL, respectively, in TGRLs plus LDLs,P,0.001; 5.5564.18 versus 0.8260.35 mg/dL, respectively,in lipid-rich HDLs, P,0.07; and 3.6160.75 versus 2.2760.55mg/dL, respectively, in lipid-poor forms,P,0.04). The higherconcentrations ofhapoA-I, hapoC-III, and cholesterol character-istic of males were also observed in the different lipoproteinfractions, although an insufficient number of animals wasstudied for statistical comparison.
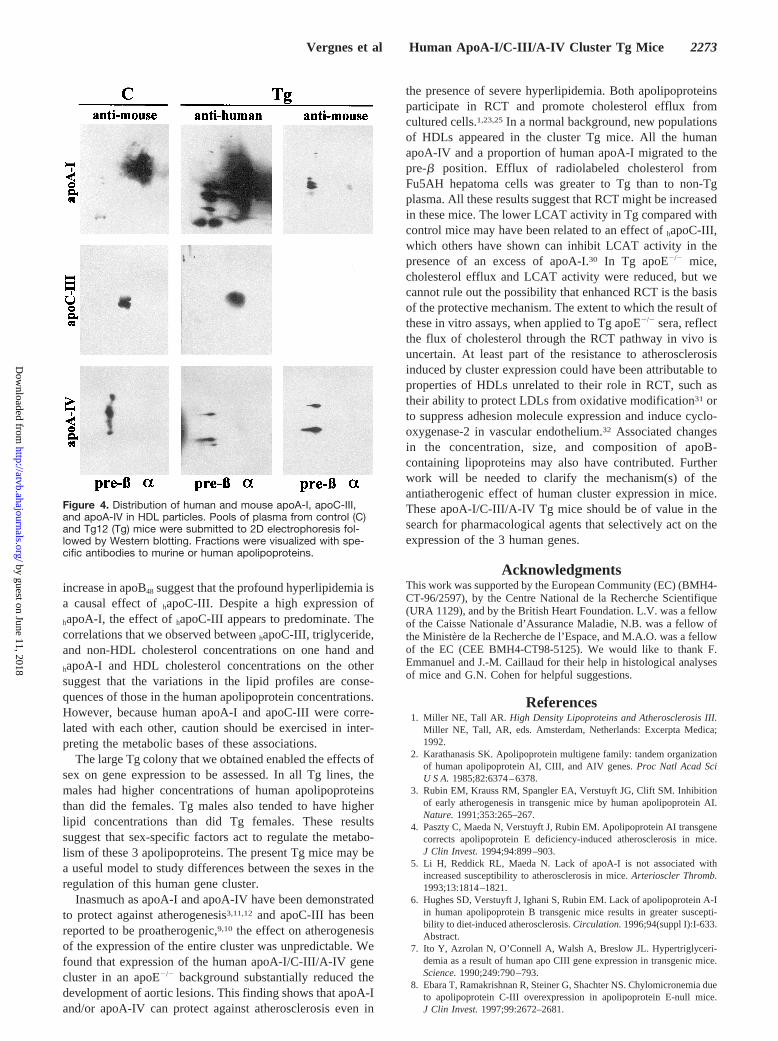
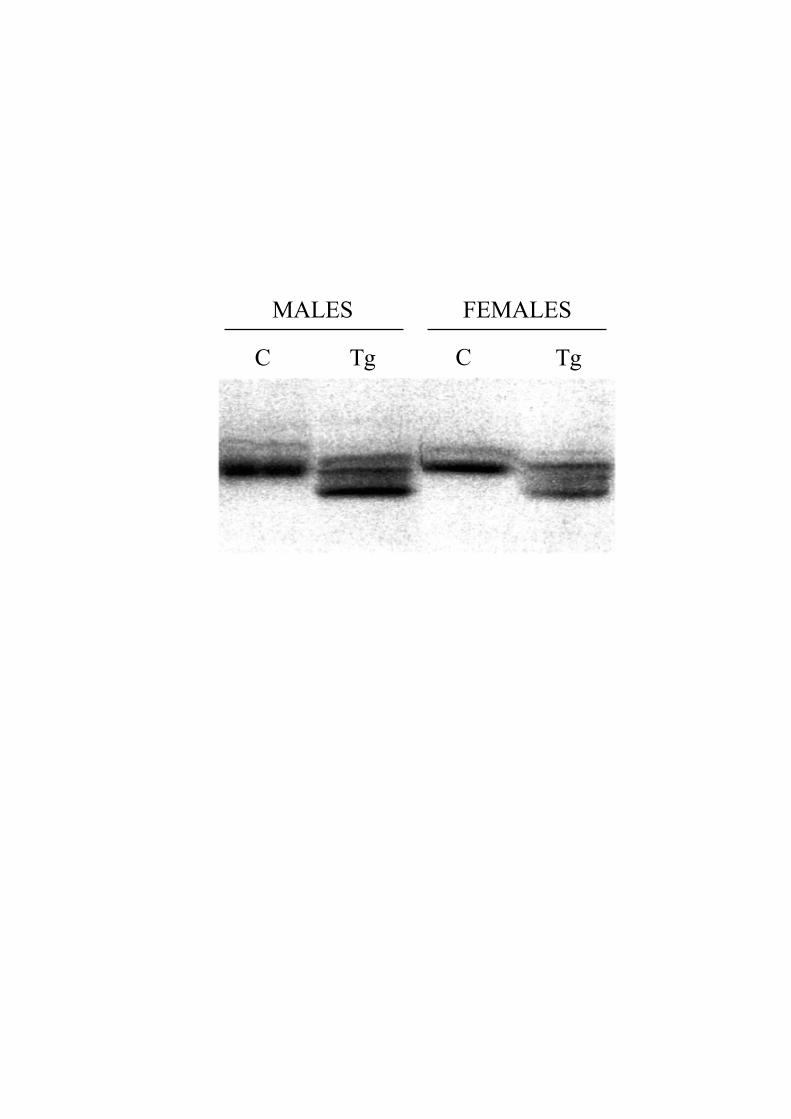
Human and Mouse Apolipoprotein Distribution inHDL ParticlesIn addition to the quantitative differences in non-HDL andHDL cholesterol between Tg and non-Tg animals demon-strated by precipitation assays, there were marked differencesbetween the 2 groups in the number and sizes of HDLsubpopulations analyzed in nondenaturing polyacrylamidegradient gels. An essentially monodisperse population oflarge HDLs present in the control mice was replaced bymultiple HDL subspecies in the cluster Tgs, accompanied bya redistribution of core lipids into small HDLs (Figure I,which can be accessed online at www.ahajournals.org).
Bidimensional electrophoresis and Western blotting withspecies-specific anti-apolipoprotein antisera (Figure 4)showed thathapoA-I was contained mostly ina-migratingparticles and to a lesser degree in multiple pre-b particles.
TABLE 2. Lipid Concentrations in Control and Tg Mice
Strain
Concentration, mg/dL
Triglycerides Total Cholesterol HDL Cholesterol
All Males Females All Males Females All Males Females
Control 78602 86603* 71603 87602 100602† 7662 5962 7062† 5162
Tg11 374640‡ 517637*‡ 247629§ 14164‡ 14864‡ 13566‡ 8862‡ 8763‡ 8864‡
Tg12 683677‡ 8506190*‡ 597648‡ 14867‡ 169616†‡ 13766‡ 8063‡ 9064*‡ 7463‡
Tg21 443638‡ 497648‡ 381656‡ 11563‡ 12263§ 10764‡ 6162 6463 5762
apoE2/2 122615 150618 7169 603644 689648* 444647 2062 1361† 3261
Tg12/E2/2 16656265\¶ 24916234\¶† 530665# 812669¶# 1001673¶**† 553641¶ 5063\¶ 4063\¶† 6162\††
Values are mean6SEM (n56 to 25).*P,0.01 and †P,0.001 vs females; ‡P,0.001 and §P,0.01 vs control mice; #P,0.05, **P,0.01, and \P,0.001 vs apoE2/2 mice; and ††P,0.01 and
¶P,0.001 vs Tg12.
2270 Arterioscler Thromb Vasc Biol. October 2000
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
Mouse apoA-I was localized predominantly ina-HDL parti-cles in control mice, whereas in Tg mice, it was reduced anddistributed almost exclusively in pre-b HDL particles. Thisreduction in mouse apoA-I concentrations was consistentwith the values measured in whole plasma by immunoassay(11467 mg/dL in control mice versus 2064 mg/dL in Tgmice,P,0.001). Mouse apoC-III in control mice andhapoC-III in Tg mice were localized mainly ina particles. We didnot localize mouse apoC-III in Tg mice, because antibodiesdirected against the mouse protein also recognizedhapoC-III.Finally, human apoA-IV and mouse apoA-IV were localizedonly in pre-b HDLs. In Tg mice, mouse apoA-IV wasdisplaced from certain pre-b HDL particles, presumably byhapoA-I. Thus,hapoA-I andhapoA-IV were able to form pre-bparticles in these mice.
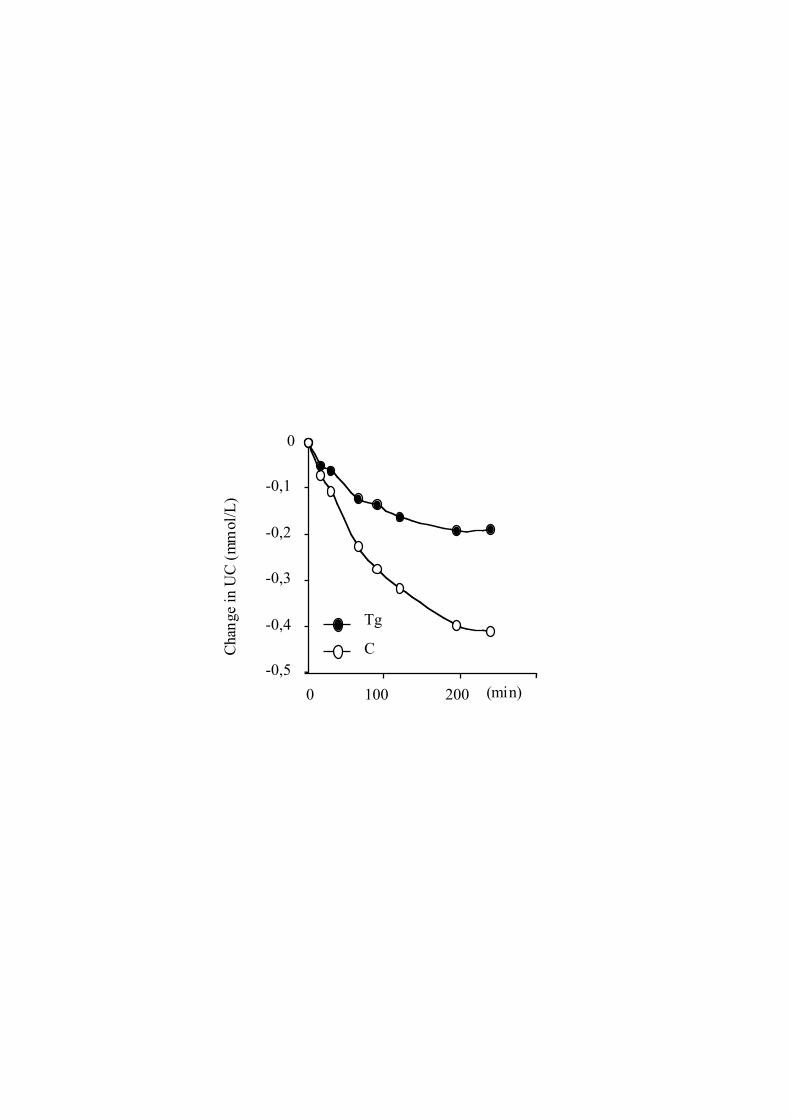
Cholesterol Efflux, LCAT Activity, andCholesterol Esterification RatePlasma from Tg mice was 29% more efficient than plasmafrom non-Tg mice (12.460.8% versus 9.660.5%, respec-tively; P,0.05) in promoting cholesterol efflux from Fu5AHcells. Whereas no significant difference in plasma LCATactivity was observed between Tg and control mice (7.360.5versus 6.660.4, respectively;P50.324), the plasma choles-terol esterification rate was reduced in the Tg mice (Figure II,which can be accessed online at www.ahajournlas.org).
Expression of the Human ApoA-I/C-III/A-IV GeneCluster in ApoE2/2 MiceConcentration ofhapoA-I was significantly lower in an apoEnull background (Tg12/apoE2/2) than in the original back-ground (Tg12). In Tg12/apoE2/2 mice, thehapoC-III concen-
tration was increased in males but not in females, and thehapoA-IV concentration was unchanged (Table 1). In thisapoE2/2 background, concentrations of human apolipopro-teins were also higher in males than in females. There was alarge increase in plasma triglycerides in Tg compared withcontrol mice (Table 2): an18-fold increase in males and a7-fold increase in females. Total cholesterol, which wasalready very high in apoE2/2 mice, was even more elevated inthe Tg12/apoE2/2 strain because of an increase in non-HDLand HDL fractions. Cholesterol efflux capacity and LCATactivity were both higher in apoE2/2 mouse plasma than in theTg12/apoE2/2 mouse plasma (8.3161.57% versus 6.1461.10%,P,0.05, and 2.560.2 versus 1.460.1 nmol z mL21 z h21,P,0.01, respectively). Previous studies have shown that theapoE2/2 mouse model develops atherosclerotic lesions sponta-neously.21 In the present study, all the control and cluster Tgmice with an apoE2/2 background developed spontaneous le-sions in the aortic sinus at 20 weeks of age. Characteristicsections from each group showed that the lesions were morpho-logically similar. However, the average cross-sectional areas oflesions were 61% lower in Tg12/apoE2/2 mice than in apoE2/2
mice (89 797616 386 versus 232 279616 273 mm2, respec-tively; P,0.001; Figure III, which can be accessed online atwww.ahajournals.org).
DiscussionTo study the effect of the coordinated expression of the 3genes of the apoA-I/C-III/A-IV cluster, we have created Tgmice with this human cluster. In the present study, wedemonstrate that expression of the entire cluster in miceproduces a hyperlipidemic profile but protects againstatherosclerosis.
Figure 2. Lipid and apoB distributions inmouse non-HDL lipoproteins. Plasmalipoproteins were isolated by sequentialultracentrifugation. Lipid levels were quanti-fied by enzymatic assay, and apoBs werequantified by video densitometric scanningof stained SDS-polyacrylamide gels. A, Tri-glycerides. B, Cholesterol. C, ApoB48. D,ApoB100. Lipids and apoBs were quantifiedin 3 density fractions. Values correspond toaverages from 3 experiments with control(open bars) and Tg (filled bars) males fromTg12. *P,0.05; **P,0.01; and ***P,0.001.Similar results were obtained with micefrom Tg21 (data not shown).
Vergnes et al Human ApoA-I/C-III/A-IV Cluster Tg Mice 2271
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Regulatory regions of the 3 genes have been localizedthroughout the entire gene cluster, and all known regulatoryelements were present in their natural positions in thegenomic fragment used to generate the Tg animals. Northernblot analyses showed the presence of the 3 human mRNAs inthe liver and intestine. These results suggest that the trans-gene contains all the regulatory regions required to direct atissue-specific expression of the 3 genes. Hepatic and intes-tinal expressions ofhapoA-I and hapoC-III were roughlysimilar to those observed in humans.27 ApoA-IV expressionwas mainly hepatic, in contrast to the predominant intestinalexpression observed in humans or inhapoA-IV Tg mice.19,20
Because the latter were generated with a DNA fragmentcontained within that used to generate the cluster Tg mice, theapoA-IV expression observed in the present study does notseem to be due to the absence of a DNA regulatory element.Comparison between intestinally expressinghapoA-IV Tgmice20 and cluster Tg mice showed that hepatic mRNA levelswere similar in both types of mice, whereas intestinal mRNAlevels were much lower in the cluster Tg mice. We hypoth-esize that the overexpression ofhapoA-I orhapoC-III genes orthe subsequent alterations induced in the lipid profile mightreduce intestinal apoA-IV expression in cluster Tg mice.
All Tg lines showed the presence of the 3 human apoli-poproteins in plasma. The mean plasma concentrations ofeach human apolipoprotein were rather similar in the 3 lines,suggesting that their concentrations were precisely controlled
in these animals. This control seems to be independent oftransgene copy number, inasmuch as Tg11, Tg12, and Tg21lines contained 9, 1, and 2 copies, respectively, of thetransgene. The control also seems independent of the inser-tion site, inasmuch as the 3 lines were independentlygenerated.
Concentrations ofhapoA-I andhapoC-III in plasma were'2-fold greater than the values in humans. In contrast, muchlower concentrations ofhapoA-IV were detected in the clusterTg mice, probably because of the low level of intestinalexpression observed. Size exclusion chromatography demon-strated that mosthapoA-I was present in particles of a sizesimilar to that of cholesterol-rich HDLs in humans. Minoramounts ofhapoA-I were present in larger particles as well asin small lipid-poor species. Human apoC-III and apoA-IVwere found in cholesterol-rich HDLs and in the non-HDLfraction. A substantial proportion ofhapoA-IV was alsopresent in small lipid-poor subclasses. These profiles werevery similar to those observed in normal humans.28,29
Analysis of the lipid profile of the Tg mice showed grosshypertriglyceridemia and a moderate increase in plasmacholesterol, mainly in the non-HDL fraction. This lipidprofile and the changes in apoB48 and apoB100 concentrationsin lipoprotein subclasses show that the major increases inplasma lipids in the Tg mice reflect an accumulation ofapoB48-rich particles. The observed association between thetriglyceride, cholesterol, andhapoC-III concentrations and the
Figure 3. Distribution of hapoA-I, hapoC-III, and hapoA-IV in size subclasses of lipoproteins. Lipoproteins were separated into size sub-classes by tandem Superdex HP-SEC. Representative male (F) and female (E) mice from the Tg12 line are shown. The distribution oftotal cholesterol is presented for comparative purposes. A, Total cholesterol. B, hApoA-I. C, hApoC-III. D, hApoA-IV. Fractions 1 to 14represent TGRLs plus LDLs, and fractions 15 to 60 represent lipid-rich and lipid-poor forms of HDLs.
2272 Arterioscler Thromb Vasc Biol. October 2000
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
increase in apoB48 suggest that the profound hyperlipidemia isa causal effect ofhapoC-III. Despite a high expression ofhapoA-I, the effect ofhapoC-III appears to predominate. Thecorrelations that we observed betweenhapoC-III, triglyceride,and non-HDL cholesterol concentrations on one hand andhapoA-I and HDL cholesterol concentrations on the othersuggest that the variations in the lipid profiles are conse-quences of those in the human apolipoprotein concentrations.However, because human apoA-I and apoC-III were corre-lated with each other, caution should be exercised in inter-preting the metabolic bases of these associations.
The large Tg colony that we obtained enabled the effects ofsex on gene expression to be assessed. In all Tg lines, themales had higher concentrations of human apolipoproteinsthan did the females. Tg males also tended to have higherlipid concentrations than did Tg females. These resultssuggest that sex-specific factors act to regulate the metabo-lism of these 3 apolipoproteins. The present Tg mice may bea useful model to study differences between the sexes in theregulation of this human gene cluster.
Inasmuch as apoA-I and apoA-IV have been demonstratedto protect against atherogenesis3,11,12 and apoC-III has beenreported to be proatherogenic,9,10 the effect on atherogenesisof the expression of the entire cluster was unpredictable. Wefound that expression of the human apoA-I/C-III/A-IV genecluster in an apoE2/2 background substantially reduced thedevelopment of aortic lesions. This finding shows that apoA-Iand/or apoA-IV can protect against atherosclerosis even in
the presence of severe hyperlipidemia. Both apolipoproteinsparticipate in RCT and promote cholesterol efflux fromcultured cells.1,23,25In a normal background, new populationsof HDLs appeared in the cluster Tg mice. All the humanapoA-IV and a proportion of human apoA-I migrated to thepre-b position. Efflux of radiolabeled cholesterol fromFu5AH hepatoma cells was greater to Tg than to non-Tgplasma. All these results suggest that RCT might be increasedin these mice. The lower LCAT activity in Tg compared withcontrol mice may have been related to an effect ofhapoC-III,which others have shown can inhibit LCAT activity in thepresence of an excess of apoA-I.30 In Tg apoE2/2 mice,cholesterol efflux and LCAT activity were reduced, but wecannot rule out the possibility that enhanced RCT is the basisof the protective mechanism. The extent to which the result ofthese in vitro assays, when applied to Tg apoE2/2 sera, reflectthe flux of cholesterol through the RCT pathway in vivo isuncertain. At least part of the resistance to atherosclerosisinduced by cluster expression could have been attributable toproperties of HDLs unrelated to their role in RCT, such astheir ability to protect LDLs from oxidative modification31 orto suppress adhesion molecule expression and induce cyclo-oxygenase-2 in vascular endothelium.32 Associated changesin the concentration, size, and composition of apoB-containing lipoproteins may also have contributed. Furtherwork will be needed to clarify the mechanism(s) of theantiatherogenic effect of human cluster expression in mice.These apoA-I/C-III/A-IV Tg mice should be of value in thesearch for pharmacological agents that selectively act on theexpression of the 3 human genes.
AcknowledgmentsThis work was supported by the European Community (EC) (BMH4-CT-96/2597), by the Centre National de la Recherche Scientifique(URA 1129), and by the British Heart Foundation. L.V. was a fellowof the Caisse Nationale d’Assurance Maladie, N.B. was a fellow ofthe Ministere de la Recherche de l’Espace, and M.A.O. was a fellowof the EC (CEE BMH4-CT98-5125). We would like to thank F.Emmanuel and J.-M. Caillaud for their help in histological analysesof mice and G.N. Cohen for helpful suggestions.
References1. Miller NE, Tall AR. High Density Lipoproteins and Atherosclerosis III.
Miller NE, Tall, AR, eds. Amsterdam, Netherlands: Excerpta Medica;1992.
2. Karathanasis SK. Apolipoprotein multigene family: tandem organizationof human apolipoprotein AI, CIII, and AIV genes.Proc Natl Acad SciU S A.1985;82:6374–6378.
3. Rubin EM, Krauss RM, Spangler EA, Verstuyft JG, Clift SM. Inhibitionof early atherogenesis in transgenic mice by human apolipoprotein AI.Nature.1991;353:265–267.
4. Paszty C, Maeda N, Verstuyft J, Rubin EM. Apolipoprotein AI transgenecorrects apolipoprotein E deficiency-induced atherosclerosis in mice.J Clin Invest.1994;94:899–903.
5. Li H, Reddick RL, Maeda N. Lack of apoA-I is not associated withincreased susceptibility to atherosclerosis in mice.Arterioscler Thromb.1993;13:1814–1821.
6. Hughes SD, Verstuyft J, Ighani S, Rubin EM. Lack of apolipoprotein A-Iin human apolipoprotein B transgenic mice results in greater suscepti-bility to diet-induced atherosclerosis.Circulation.1996;94(suppl I):I-633.Abstract.
7. Ito Y, Azrolan N, O’Connell A, Walsh A, Breslow JL. Hypertriglyceri-demia as a result of human apo CIII gene expression in transgenic mice.Science.1990;249:790–793.
8. Ebara T, Ramakrishnan R, Steiner G, Shachter NS. Chylomicronemia dueto apolipoprotein C-III overexpression in apolipoprotein E-null mice.J Clin Invest.1997;99:2672–2681.
Figure 4. Distribution of human and mouse apoA-I, apoC-III,and apoA-IV in HDL particles. Pools of plasma from control (C)and Tg12 (Tg) mice were submitted to 2D electrophoresis fol-lowed by Western blotting. Fractions were visualized with spe-cific antibodies to murine or human apolipoproteins.
Vergnes et al Human ApoA-I/C-III/A-IV Cluster Tg Mice 2273
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from

9. Hayek T, Masucci-Magoulas L, Jiang X, Walsh A, Rubin E, Breslow JL,Tall AR. Decreased early atherosclerosis lesions in hypertriglyceridemicmice expressing cholesteryl ester transfer protein transgene.J Clin Invest.1995;96:2071–2074.
10. Masucci-Magoulas L, Goldberg IJ, Bisgaier CL, Serajuddin H, FranconeOL, Breslow JL, Tall AR. A mouse model with features of familialcombined hyperlipidemia.Science.1997;275:391–394.
11. Duverger N, Tremp G, Caillaud JM, Emmanuel F, Castro G, Fruchart JC,Steinmetz A, Denefle P. Protection against atherogenesis in micemediated by human apolipoprotein A-IV.Science.1996;273:966–968.
12. Cohen RD, Castellani LW, Qiao JH, Van Lenten BJ, Lusis AJ, Reue K.Reduced aortic lesions and elevated high density lipoprotein levels intransgenic mice overexpressing mouse apolipoprotein A-IV.J ClinInvest.1997;99:1906–1916.
13. Walsh AM, Azrolan N, Wang K, Marcigliano A, O’Connell A, BreslowJL. Intestinal expression of the human ApoAI gene in transgenic mice iscontrolled by a DNA region 39to the gene in the promoter of the adjacentconvergently transcribed ApoCIII gene.J Lipid Res.1993;34:617–623.
14. Vergnes L, Taniguchi T, Omori K, Zakin MM, Ochoa A. The apoli-poprotein A-I/C-III/A-IV gene cluster: apoC-III and apoA-IV expressionis regulated by two common enhancers.Biochim Biophys Acta.1997;1348:299–310.
15. Le Beyec J, Chauffeton V, Kan H-Y, Janvier P-L, Cywiner-Golenzer C,Chatelet F-P, Kalopissis AD, Zannis V, Chambaz J, Pincon-Raymond M,Cardot P. The -700/-310 fragment of the apolipoprotein A-IV genecombined with the -890/-500 apolipoprotein C-III enhancer is sufficientto direct a pattern of gene expression similar to that for the endogenousapolipoprotein A-IV gene.J Biol Chem.1999;274:4954–4961.
16. Metzger S, Levy Y, Arnon R, Cajek-Shaul T. Co-regulation of apo A-I,apo C-III and apo A-IV gene expression in human intestinal biopsies.EurJ Clin Invest.1996;26:71–75.
17. Walsh A, Ito Y, Breslow JL. High levels of human apolipoprotein A-I intransgenic mice result in increased plasma levels of small high densitylipoprotein (HDL) particles comparable to human HDL3. J Biol Chem.1989;264:6488–6494.
18. de Silva HV, Lauer SJ, Wang J, Simonet WS, Weisgraber KH, Mahley RW,Taylor JM. Overexpression of human apolipoprotein C-III in transgenic miceresults in an accumulation of apolipoprotein B48 remnants that is corrected byexcess apolipoprotein E.J Biol Chem.1994;269:2324–2335.
19. Aalto-Seta¨la K, Bisgaier CL, Ho A, Kieft KA, Traber MG, Kayden HJ,Ramakrishnan R, Walsh A, Essenburg AD, Breslow JL. Intestinal expression ofhuman apolipoprotein A-IV in transgenic mice fails to influence dietary lipidabsorption or feeding behavior.J Clin Invest.1994;93:1776–1786.
20. Baralle M, Vergnes L, Muro AF, Zakin MM, Baralle FE, Ochoa A.Regulation of the human apolipoprotein A-IV gene expression intransgenic mice.FEBS Lett.1999;445:45–52.
21. Zhang SH, Reddick RL, Piedrahita JA, Maeda N. Spontaneous hyper-cholesterolemia and arterial lesions in mice lacking apolipoprotein E.Science.1992;258:468–471.
22. Sharpe CR, Sidoli A, Shelley CS, Lucero MA, Shoulders CC, Baralle FE.Human apolipoproteins AI, AII, CII and CIII: cDNA sequences andmRNA abundance.Nucleic Acids Res.1984;12:3917–3932.
23. Castro G, Parmentier Nihoul L, Dengremont C, de Geitere C, Delfly B,Tailleux A, Fievet C, Duverger N, Denefle P, Fruchart JC, Rubin EM.Cholesterol efflux, lecithin-cholesterol acyltransferase activity, and pre-b
particle formation by serum from human apolipoprotein A-I and apoli-poprotein A-I/apolipoprotein A-II transgenic mice consistent with thelatter being less effective for reverse cholesterol transport.Biochemistry.1997;36:2243–2249.
24. Brousseau T, Clavey V, Bard J-M, Fruchart J-C. Sequential ultracentrif-ugation micromethod for separation of serum lipoproteins and assays oflipids, apolipoproteins, and lipoprotein particles.Clin Chem.1993;39:960–964.
25. Castro G, Fielding C. Early incorporation of cell-derived cholesterol intopre-beta-migrating high-density lipoprotein.Biochemistry.1988;27:25–29.
26. Paigen B, Morrow A, Holmes PA, Mitchell D, Williams RA. Quantitativeassessment of atherosclerotic lesions in mice.Atherosclerosis.1987;68:231–240.
27. Lenich C, Brecher P, Makrides S, Chobanian A, Zannis VI. Apoli-poprotein gene expression in the rabbit: abundance, size, and distributionof apolipoprotein mRNA species in different tissues.J Lipid Res.1988;29:755–764.
28. Nanjee MN, Doran JE, Lerch PG, Miller NE. Acute effects of intravenousinfusion of apoA1/phosphatidycholine discs on plasma lipoproteins inhumans.Arterioscler Thromb Vasc Biol.1999;19:979–989.
29. Kondo K, Allan C, Fidge N. Quantitation of apolipoprotein A-IV inhuman plasma using a competitive enzyme-linked immunosorbent assay.J Lipid Res.1989;30:939–944.
30. Nishida HI, Nakanishi T, Yen EA, Arai H, Yen FT, Nishida T. Nature ofthe enhancement of lecithin-cholesterol acyltransferase reaction byvarious apolipoproteins.J Biol Chem.1986;261:12028–12035.
31. Watson AD, Berliner JA, Hama SY, La Du BN, Faull KF, Fogelman AM,Navab M. Protective effect of high density lipoprotein associated para-oxonase: inhibition of the biological activity of minimally oxidized lowdensity lipoproteins.J Clin Invest.1995;96:2882–2891.
32. Cockerill GW, Saklatvala J, Ridley SH, Yarwood H, Miller NE, Oral B,Nithyanathan S, Taylor G, Haskard DO. High-density lipoproteins dif-ferentially modulate cytokine-induced expression of E-selectin and cy-clooxygenase-2.Arterioscler Thromb Vasc Biol.1999;19:910–917.
2274 Arterioscler Thromb Vasc Biol. October 2000
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Alberto OchoaNazeem Nanjee, Jamila Najib, Jean-Charles Fruchart, Norman E. Miller, Mario M. Zakin and
Laurent Vergnes, Nadine Baroukh, Maria A. Ostos, Graciela Castro, Nicolas Duverger, M.Hyperlipidemia but Reduces Atherogenesis
Expression of Human Apolipoprotein A-I/C-III/A-IV Gene Cluster in Mice Induces
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2000 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.20.10.22672000;20:2267-2274Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/20/10/2267World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from

http://atvb.ahajournals.org/content/suppl/2000/09/27/20.10.2267.DC4 http://atvb.ahajournals.org/content/suppl/2000/09/27/20.10.2267.DC3 http://atvb.ahajournals.org/content/suppl/2000/09/27/20.10.2267.DC2 http://atvb.ahajournals.org/content/suppl/2000/09/27/20.10.2267.DC1
Data Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 11, 2018http://atvb.ahajournals.org/
Dow
nloaded from
C Tg C Tg
MALES FEMALES
Cha
nge
in U
C (
mm
ol/L
)
Tg
C-0,5
-0,4
-0,3
-0,2
-0,1
0
0 100 200 (min)

apoE-/- Tg12/apoE
-/-
Mea
n cr
oss-
sect
iona
l are
aof
lesi
ons
(105
µm2 )
0
1
2
3
4
***





























