Embed Size (px)
Citation preview

88:3452-3468, 2002. doi:10.1152/jn.00331.2002 JNPierre A. Sylvestre and Kathleen E. Cullen During Disjunctive Saccades Dynamics of Abducens Nucleus Neuron Discharges
You might find this additional information useful...
56 articles, 22 of which you can access free at: This article cites http://jn.physiology.org/cgi/content/full/88/6/3452#BIBL
1 other HighWire hosted article: This article has been cited by
[PDF] [Full Text] [Abstract]
, August 1, 2003; 90 (2): 739-754. J NeurophysiolP. A. Sylvestre, J. T. L. Choi and K. E. Cullen
Disjunctive Saccades and FixationDischarge Dynamics of Oculomotor Neural Integrator Neurons During Conjugate and
on the following topics: http://highwire.stanford.edu/lists/artbytopic.dtlcan be found at Medline items on this article's topics
Medicine .. Eye Movement Biochemistry .. Motor Neurons
including high-resolution figures, can be found at: Updated information and services http://jn.physiology.org/cgi/content/full/88/6/3452
can be found at: Journal of Neurophysiologyabout Additional material and information http://www.the-aps.org/publications/jn
This information is current as of March 21, 2005 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the
publishes original articles on the function of the nervous system. It is published 12 times a yearJournal of Neurophysiology
on March 21, 2005
jn.physiology.orgD
ownloaded from

Dynamics of Abducens Nucleus Neuron Discharges DuringDisjunctive Saccades
PIERRE A. SYLVESTRE AND KATHLEEN E. CULLENAerospace Medical Research Unit, McGill University, Montreal, Quebec H3G 1Y6, Canada
Received 6 May 2002; accepted in final form 19 August 2002
Sylvestre, Pierre A. and Kathleen E. Cullen. Dynamics of abdu-cens nucleus neuron discharges during disjunctive saccades. J Neu-rophysiol 88: 3452–3468, 2002; 10.1152/jn.00331.2002. In this re-port, we provide the first characterization of abducens nucleus neuron(ABN) discharge dynamics during horizontal disjunctive saccades.These movements function to rapidly transfer the visual axes betweentargets located at different eccentricities and depths. Our primaryobjective was to determine whether the signals carried by ABNsduring these movements are appropriate to drive the motion of the eyeto which they project. We also asked whether ABNs encode eyemovements similarly during disjunctive saccades and disjunctive fix-ation. To address the first objective we 1) assessed whether we couldpredict the discharge dynamics of individual neurons during disjunc-tive saccades based on their discharge properties during conjugatesaccades and 2) directly estimated the sensitivity of individual neu-rons to either the ipsilateral/contralateral eye or the conjugate/ver-gence position and velocity using bootstrap statistics. Our main find-ing was that during disjunctive saccades in the direction ipsilateral tothe recording site (ON-direction), the majority of ABNs preferentiallyencoded the velocity and the position of the ipsilateral eye. Theremaining neurons predominantly encoded the conjugate motion ofthe eyes (i.e., were equally sensitive to the motion of both eyes).Generally, ipsilateral/contralateral eye based models better describedneuronal discharges than conjugate/vergence based models, yet bothmodel structures yielded similar conclusions. Moreover, the preferredeye of individual neurons based on their position and velocity sensi-tivities were generally well matched. We also found that for saccadesin the OFF-direction, the pausing behavior of ABNs was similar duringconjugate and disjunctive saccades, with the exception that for move-ments of small amplitudes, more ABNs paused during conjugatesaccades. Finally, we found that putative motoneurons and internu-clear neurons encoded ON- and OFF-direction disjunctive saccades in asimilar manner. To address our second objective, we compared thedischarge properties of individual ABNs during disjunctive saccadesand disjunctive fixation. Good coherence was observed between thepreferred eye of individual ABNs during the two behaviors. Takentogether, our results indicate that although individual ABNs canencode the motion of both eyes to various degrees, the populationdrive of ABNs accounts for most of the movement of the ipsilateraleye during disjunctive saccades and disjunctive fixation.
I N T R O D U C T I O N
To optimize visual perception, it is essential for foveatedanimals to precisely align their two visual axes on targets ofinterest. Therefore it is not surprising that the oculomotor
system of these animals has developed sophisticated mecha-nisms to ensure the tight control of binocular positioning. Morethan a century ago, Hering (1868) proposed the elegant “theoryof equal innervation” as a conceptual framework for the studyof binocular control. When applied to eye movements betweentwo immobile targets, for example, Hering’s theory suggeststhat two separate neural subsystems should exist that controldifferent aspects of these movements (Fig. 1). On the one hand,a conjugate saccadic subsystem would rapidly yoke the eyes ina given direction to generate movements between targets lo-cated at a constant depth but at different horizontal eccentric-ities. On the other hand, a slower and separate vergence sub-system would rotate the eyes by the same angle but in oppositedirections to generate eye movements between targets locatedat different depths but at constant eccentricities. To date, theneural basis of these two subsystems has been extensivelystudied in isolation. Under these conditions, neuronal circuitrythat are involved in generating conjugate saccades (reviewed inMoschovakis et al. 1996; Scudder et al. 2002) or slow sym-metric vergence shifts (reviewed in Gamlin 1999; Mays 1995a)have been well characterized.
However, during our normal daily activities these two sub-systems do not always function in isolation; we generate si-multaneous conjugate and vergence eye movements anytimewe rapidly reorient our eyes between targets located at differ-ent eccentricities and depths. During such eye movements,termed disjunctive saccades, the two eyes rotate by differentangles and with different trajectories. Accordingly, a questionthat naturally arises is: does Hering’s theory hold true duringdisjunctive saccades?
In fact, although Hering’s theory is attractive in its simplic-ity and in the anatomical and physiological correlates thatsupport it during pure conjugate or vergence movements, itcannot account for a number of observations made duringdisjunctive saccades. For example, a number of studies haveclearly demonstrated that the straightforward linear summationof the conjugate and vergence components of eye motionpredicted by the theory of equal innervation does not occurduring disjunctive saccades (human: Collewijn et al. 1995,1997; Enright 1984; Erkelens et al. 1989; Kenyon et al. 1980;Ono et al. 1978; Oohira 1993; Zee et al. 1992; monkey:Maxwell and King 1992). Rather, it was shown that the ver-gence component of the movement is dramatically accelerated
Address for reprint requests: K. E. Cullen, 3655 Prom. Sir William Osler,Rm. 1220, Montreal, Quebec H3G 1Y6, Canada (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked ‘‘advertisement’’in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
J Neurophysiol 88: 3452–3468, 2002;10.1152/jn.00331.2002.
3452 0022-3077/02 $5.00 Copyright © 2002 The American Physiological Society www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

when compared with a control pure vergence shift, while thesaccadic movement is slowed down in comparison to controlconjugate saccades, suggesting central interactions between theconjugate and vergence neural pathways. We have recentlyfurthered the evidence supporting the central coupling betweenthe conjugate and vergence premotor circuitry by describingsynchronized oscillations in the conjugate and transient ver-gence of conjugate saccades and gaze shifts (Sylvestre et al.2002).
Recent reports have also indicated that some neural struc-tures previously assumed to form the conjugate saccadic sys-tem do not carry purely conjugate information during disjunc-tive saccades. For example, electrical perturbations of thesuperior colliculus during disjunctive saccades were shown tomodify both the conjugate and the vergence trajectories(Chaturvedi and VanGisbergen 1999, 2000). Also, premotorsaccadic burst neurons that are active only during saccadic eyemovements (Sylvestre and Cullen 1999b; Zhou and King 1998)and nuclei prepositus/vestibular neurons (McConville et al.1994; Zhou and King 1996) were found to preferentially en-code the velocity and position of one of the two eyes (i.e., donot encode the conjugate eye position) during disjunctive sac-cades and disjunctive fixation, respectively. Thus these studieshave provided convincing evidence that Hering’s law is vio-lated at the premotor level during disjunctive saccades. It islikely that these neurophysiological observations represent thesubstrate for the saccadic facilitation of vergence, where thefaster vergence velocities are supplied through the saccadiccircuitry (see Sylvestre et al. 2002).
Although we are now beginning to better understand thepremotor mechanisms of binocular control during disjunctive
saccades, surprisingly nothing is known about the actual sig-nals that are generated by extraocular motoneurons to drivethese eye movements. To date, all of our knowledge on motorpatterns during disjunctive movements was obtained duringslow, nonsaccadic eye movements (Gamlin et al. 1989; Gamlinand Mays 1992; Keller 1973; Keller and Robinson 1972; Kingand Zhou 2000; King et al. 1994; Mays and Porter 1984; Zhouand King 1996, 1998). Most of these studies were conductedon neurons in the abducens nucleus (ABNs), which containstwo subpopulations of neurons: motoneurons (AMN) thatproject to the ipsilateral lateral rectus and internuclear neurons(AIN) that project to medial rectus motoneurons (OMNs) in thecontralateral oculomotor nucleus (see Fig. 1; Delgado-Garcia1986a,b). It was found that nearly all ABNs, including identi-fied AMNs and AINs (Gamlin et al. 1989), encode similarsignals during slow vergence eye movements. When the eyessymmetrically converge (i.e., both eyes move nasally), thedischarges of ABNs decrease, while they increase during di-vergence (i.e., both eyes move temporally). In contrast, OMNsthat drive the medial rectus muscles increase their dischargeswhen the eyes converge (Gamlin and Mays 1992; King et al.1994; Mays and Porter 1984). Thus the discharge patterns ofAMNs and OMNs during slow disjunctive eye movements aremodulated appropriately to drive the eye muscles to which theyproject, but those of AINs are modulated inappropriately todrive the contralateral eye to which they project. These results,overall, are consistent with Hering’s theory of equal innerva-tion (Mays 1998).
In the present study, our primary objective was to determinewhether the signals carried by ABNs during disjunctive sac-cades are appropriate to drive the motion of the eye to whichthey project. Since AMNs ultimately drive the extraocularmuscles of the ipsilateral eye, the conjugate and vergence-related premotor inputs that they receive during disjunctivesaccades might be combined, on a neuron-by-neuron basis, togenerate motor signals that are exclusively related to the move-ments of that eye. Alternatively, single neurons might encodemixed signals that get sorted out at the population level, suchthat the overall ABN drive to the ipsilateral eye is appropriate.Finally, the convergence of conjugate and vergence-relatedpremotor signals on ABNs might be incomplete or inappropri-ate, such that the discharge patterns of AMNs would notaccount entirely for the movements of the ipsilateral eye. Inthis scheme, additional mechanisms at or downstream to theabducens nucleus, for example co-contraction of the agonistand antagonist muscles, would be required to fine tune the eyemovements. A secondary goal of this study was to determinewhether ABNs encode conjugate and vergence signals simi-larly during disjunctive saccades and disjunctive fixation. Aswe described above, there is evidence that the source of thevergence-related premotor signals differs in these two condi-tions. Consequently, there are no a priori reasons to assumethat, for example, a neuron that encodes conjugate signalsduring disjunctive saccades will also encode conjugate signalsduring disjunctive fixation. Some of the results have beenreported in abstract form (Sylvestre and Cullen 1999b).
M E T H O D S
Two rhesus monkeys (Macaca mulatta) were prepared for chronicextracellular recording using the aseptic surgical procedures described
FIG. 1. Schematic representation of Hering’s theory of equal innervation.Drive from the conjugate subsystem is sent directly to the abducens nucleusand indirectly to the oculomotor nucleus via abducens internuclear neurons. Inturn, drive from the vergence subsystem is sent directly to both the abducensnucleus and the oculomotor nucleus. During disjunctive saccades, these 2drives would summate. OMN, oculomotor motoneurons; AIN, abducens in-ternuclear neuron; AMN, abducens motoneurons; III and VI, 3rd and 6thcranial nuclei; MR and LR, medial and lateral recti.
3453ABDUCENS NEURONS DURING DISJUNCTIVE SACCADES
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

elsewhere (Sylvestre and Cullen 1999a). To briefly summarize, astainless steel post that allowed the complete immobilization of theanimal’s head was attached to the animal’s skull with stainless steelscrews and dental acrylic. Two stainless steel recording chambersoriented stereotaxically toward the abducens nucleus on the right andleft side of the brain stem, respectively, were also anchored in theimplant. An eye coil (3 loops of Teflon coated stainless steel wire,18–19 mm diam) was implanted in each eye (Judge et al. 1980) toallow recordings of binocular eye movements with the magneticsearch coil technique (Fuchs and Robinson 1966). All procedureswere approved by the McGill University Animal Care Committee andwere in compliance with the guidelines of the Canadian Council onAnimal Care.
Behavioral paradigms
Both monkeys were trained to follow a target light in a dimly litroom for a juice reward. Only eye movements restricted to thehorizontal plane will be discussed in the present report. To elicitconjugate eye movements, a red HeNe laser target was projected viaa system of two galvanometer controlled mirrors onto a cylindricalscreen located 55 cm away from the monkey’s eyes (isovergent,�3.5° convergence). Ipsilaterally and contralaterally directed conju-gate saccades (�5–30°) were elicited by stepping the target betweenhorizontal positions in a predictable and an unpredictable sequence. Inaddition, smooth pursuit eye movements were obtained using a sinu-soidally moving target (40°/s peak velocity, 0.5 Hz).
An array of 16 computer-controlled red light emitting diodes(LEDs; with intensities comparable to that of the laser target) wereutilized to elicit different types of vergence eye movements. First,symmetric (pure) vergence eye movements were obtained using fourLEDs (convergence angles: 17°, 12°, 8°, and 6°) and a laser target thatwere aligned with the monkey’s mid-sagittal plane. Second, disjunc-tive saccades were generated using a variety of paradigms. In a firstconfiguration, the target jumped from one of the mid-sagittal LEDsdescribed above to an eccentric laser target (i.e., right or left of themid-sagittal plane). During this paradigm, monkeys made disjunctivesaccades with conjugate components 5–30° in amplitude in bothdirections and converging or diverging vergence components withamplitudes 4–13°. Disjunctive saccades were also obtained usingLEDs that were positioned in a configuration similar to the Mullerparadigm (see Ramat et al. 1999 for examples). More specifically,four LEDs were aligned with the left eye at an angle of �45° to theright of the mid-sagittal plane, and four other LEDs were aligned withthe right eye at an angle of �45° to the left of the mid-sagittal plane.This paradigm elicited disjunctive saccades during which the left orthe right eye barely moved, respectively. Finally, to enrich the varietyof disjunctive eye movements in our data set (and the monkey’sviewing experience), we also performed trials in which any of theLEDs and laser targets were randomly presented.
Data acquisition procedures
During the experiment, the head-restrained monkey was comfort-ably seated in a primate chair. The monkey’s head was restrained forthe duration of the experiment. The horizontal and vertical positionsof both eyes were recorded using the magnetic search coil technique(Fuchs and Robinson 1966). Extracellular single unit activity wasrecorded using enamel insulated tungsten microelectrodes (7–10 M�impedance, Frederick Haer; for details, see Sylvestre and Cullen1999a). Targets, data acquisition, and on-line data displays werecontrolled using real-time experimentation system (REX), a QNX-based real-time acquisition system (Hayes et al. 1982).
The abducens nucleus was identified as previously described (Syl-vestre and Cullen 1999a). Because of the invasiveness of implantingan electrode in the abducens nerve for antidromic activation (Del-gado-Garcia et al. 1986a,b) and/or a recording electrode in the lateral
rectus for spike triggered averaging (Fuchs et al. 1988), we elected tophysiologically identify putative AMNs and AINs using an approachmodified from Sylvestre and Cullen (1999a) (see also Broussard et al.1995). Specifically, Fuchs et al. (1998) found that identified AINs andAMNs formed fairly distinct clusters when their eye velocity sensi-tivities during sinusoidal smooth pursuit were plotted as a function oftheir eye position thresholds (see Fig. 8 of Fuchs et al. 1988). In fact,only a small area of their scatter plot showed overlap of the twoneuron types. This area of overlap can be easily defined using anupper border (R � 2.0 � 0.033 � Threshold) and a lower border (R �1.4 � 0.033 � Threshold). Here, we obtained a similar scatter plot forthe neurons in our sample and used the borders described above toseparate putative AMNs from AINs. Neurons that were plotted abovethe top border and below the lower border were labeled as putativeAINs and AMNs, respectively, while those that were plotted betweenthe upper and lower borders could not be identified and were labeledas ABNs.
When a neuron was properly isolated, unit activity, horizontal andvertical positions of the right and left eyes, target position, and tablevelocity were recorded on a digital audio tape (DAT). The isolationof each unit was re-evaluated off-line during playback. An abducensneuron was considered to be adequately isolated only when individualaction potential waveforms could be discriminated using a windowingcircuit (BAK) during saccades (e.g., see Fig. 1 in Sylvestre andCullen 1999a), during fixation and during smooth pursuit. Right eye,left eye, and target position signals were low-pass filtered at 250 Hz(analog 8-pole Bessel filter) and sampled at 1 kHz. Subsequent anal-ysis was performed using custom algorithms (Matlab, The Math-Works).
Coordinate conventions
The eyes are referred to as either ipsilateral or contralateral basedon their location relative to the recording site. Positive and negativevalues indicate eye positions that are to the right and left of the centralposition (i.e., straight ahead), respectively. Each eye was calibratedseparately by having the monkey fixate monocularly (i.e., one eyemasked) on a variety of targets at different eccentricities and depths.
The motion of the eyes is also reported in terms of conjugate andvergence coordinates
Conjugate ��Left Eye � Right Eye�
2(1a)
Vergence � �Left Eye � Right Eye� (1b)
The left eye and right eye inputs to Eq. 1 could be either position orvelocity signals. For the conjugate position signal (Eq. 1a), positiveand negative values correspond to the right and left of the mid-sagittalplane, respectively. For the vergence position signal (Eq. 1b), largerpositive values indicate greater angles of convergence. Note thatvergence position signals are always positive, but that vergence ve-locity signals can be either positive (during convergence) or negative(during divergence).
Analysis of abducens neuron discharges
Before analysis, recorded eye position signals were digitally filteredwith a 51st order finite-impulse-response (FIR) filter with a Hammingwindow, using a cutoff at 125 Hz. The position signals were digitallydifferentiated to produce eye velocity profiles. Zero-phase forwardand reverse digital filtering was employed to prevent phase distortion.A spike density function in which a Gaussian function was convolvedwith the spike train (SD of 5 ms for saccades, 10 ms for smoothpursuit and fixation) was utilized to represent the neuronal discharges(Cullen and Guitton 1997; Cullen et al. 1996; Sylvestre and Cullen1999a,b).
Horizontal saccades were defined as having vertical amplitudes
3454 P. A. SYLVESTRE AND K. E. CULLEN
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

10% of their horizontal amplitudes. Conjugate saccades hadchanges in vergence angles 2.5°, and were directed either ipsilater-ally (“ON” direction) or contralaterally (“OFF” direction) to the record-ing site. Disjunctive saccades during which both eyes moved either inthe direction ipsilateral or contralateral to the recording site, and forwhich one eye moved at least twice more than the other (meanvergence: 6.2 � 1.3°), were selected for the analysis. Note that foreach neuron analyzed, the numbers of converging and divergingdisjunctive saccades were matched. Fixation periods were defined astime intervals having peak conjugate and vergence velocities 10°/s.All analyzed fixation intervals had conjugate positions ipsilateral tothe neuron’s threshold.
The dynamic eye position and velocity sensitivities of a neuronduring saccades were estimating using linear optimization techniquesthat have been described in detail elsewhere (Sylvestre and Cullen1999a). The rationale for using this technique as opposed to a moreconventional metric-based analysis approach is described in the AP-PENDIX. The specific linear regression models utilized in this study aredescribed in RESULTS. The goodness-of-fit of a given model to the datawere quantified using the Variance-Accounted-For {VAF �1 � [var(mod � fr)/var (fr)], where mod represents the modeled firing rate andfr represents the actual firing rate}. For the estimation of linear models(like those utilized in this report), the VAF is mathematically equiv-alent to the correlation coefficient R2. A VAF value of 1 indicates aperfect fit to the data, and a value of 0 indicates a fit that is equivalentto a mean value. Note that the VAF can be utilized for the directcomparison of the goodness-of-fit of model estimations and predic-tions. The dynamic lead time of individual neurons (td) was deter-mined during conjugate saccades as described in Sylvestre and Cullen(1999a).
Statistical analysis of model parameters
In this study, the residuals of the multiple regression model utilizedfor the analysis of the discharge dynamics of ABNs during disjunctivesaccades (see model Est-ic-all in RESULTS) were not always normallydistributed. Therefore standard statistical tests could not be performedon the parameter estimates because the assumptions inherent to thesetests were invalid. To compensate for this limitation, we estimated theprobability distribution of the model parameters in Est-ic-all (and alsoEst-cv-all, see RESULTS) using a nonparametric bootstrap approach.This analysis method is described in Carpenter and Bithell (2000). Itis particularly well suited for small samples with unknown probabilitydistributions (Carpenter and Bithell 2000; Press et al. 1997; Richmondet al. 1987).
Briefly, the final model parameters in model Est-ic-all (see RESULTS)were estimated from an original data set of N (usually �40) disjunc-tive saccades (where 1⁄2 were divergent and 1⁄2 were convergent; botheyes moved in the “ON” direction). Then 1,999 “new data sets” of Nsaccades were obtained by randomly re-sampling with replacementfrom the original data set. Every new data set differed from theoriginal due to saccade repetitions and omissions, and from the othernew data sets due to the randomness of the re-sampling process.Preliminary tests conducted on 10 neurons selected randomly indi-cated that 1,999 re-samplings were sufficient to obtain stable distri-butions (i.e., yielded the same mean and SD as when using 2,999 or3,999 re-samplings). The model parameters were then estimated oneach of the new data sets.
Following the re-sampling process, 95% confidence intervals werecomputed for each model parameter (as well as for more complexstatistics such as the VAF; Sokal and Rohlf 1995) using the parametervalues obtained across the 1999 iterations (Bca method, Carpenter andBithell 2000). Parameters with 95% confidence intervals that over-lapped with zero were not statistically significant and were removedfrom the model (e.g., see Fig. 4). Parameters with 95% confidenceintervals that overlapped with one another were statistically identicaland were replaced by conjugate parameters in the model (e.g., see Fig.
7). Note that the parameters were removed one at a time, starting withthe parameter(s) that showed the most overlap, and that the parametersof the reduced model were estimated after each removal. This ap-proach prevented removing important parameters whose numericalvalues were biased by the inappropriate parameters included in theoriginal model.
R E S U L T S
The discharge dynamics of 50 abducens nucleus neurons (36from Monkey B, 14 from Monkey J) were analyzed duringconjugate and disjunctive saccades. Our analysis approach wasas follows: first, we assessed whether we could predict thedischarge dynamics of individual neurons during disjunctivesaccades based on their discharge properties during conjugatesaccades; second, we directly estimated the sensitivity of indi-vidual neurons to the velocity and position of either the right/left eyes or the conjugate/vergence traces on the same data setof disjunctive saccades. Based on this analysis, the neuronswere sorted in five categories according to the type of eyevelocity–related signals that they encoded during disjunctivesaccades: monocular with a preference for the ipsilateral eye,monocular with a preference for the contralateral eye, binoc-ular with a preference for the ipsilateral eye, binocular with apreference for the contralateral eye, or conjugate (i.e., equallyencoding the motion of both eyes). The eye velocity sensitivitywas chosen as the criterion because velocity signals are dom-inant during saccades (Sylvestre and Cullen 1999a).
In the following sections, we begin by demonstrating ouranalysis approach on a typical monocular ABN that preferen-tially encoded movements of the ipsilateral eye. We thencontrast the results with those of a typical conjugate ABN.Next, we describe in detail the distribution of our sample ofneurons across the categories described above. We also char-acterize the responses of ABNs during OFF direction disjunctivesaccades. Finally, we compare the discharge properties ofindividual ABNs during disjunctive saccades and disjunctivefixation.
Example monocular ABN with ipsilateral eye preference
We first estimated a neuron’s sensitivity to eye movementsduring conjugate saccades. Recall that during these move-ments, the two eyes rotate by the same amplitude and movewith highly comparable trajectories. The bias, conjugate eyeposition, and velocity sensitivities of the neurons were esti-mated using the following dynamic model, which we havepreviously shown provides an adequate description of ABNdischarge dynamics during conjugate saccades (Sylvestre andCullen 1999a)
FR�t� � bCS � kCSCJ�t � td� � rCSCJ�t � td� �model Est-CS�
where FR(t) is the neuron’s instantaneous firing rate, bCS, kCS,and rCS are constants and represent the neuron’s firing rate ateye position zero, the neuron’s conjugate eye position, and eyevelocity sensitivities, respectively (CS refers to conjugate sac-cades), CJ(t) and CJ(t) are instantaneous conjugate positionand velocity, respectively, and td is the neuron’s dynamic leadtime.
The model fits obtained for a typical ABN, unit B72_2, areshown in Fig. 2 for two conjugate saccades. This first-ordermodel of eye position provided a good fit of the neuron’s firing
3455ABDUCENS NEURONS DURING DISJUNCTIVE SACCADES
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

rate (Fig. 2, Est-CS; VAFEst-CS � 0.58; mean populationVAFEst-CS � SD � 0.68 � 0.12, see Table 1).
We next determined whether the conjugate model estimatedabove could be utilized to predict the neuron’s activity duringdisjunctive saccades. During converging disjunctive saccades,the contralateral eye moves more than the ipsilateral eye (e.g.,Fig. 3A), while the ipsilateral eye moves more than the con-tralateral eye during diverging saccades (e.g., Fig. 3B). Notethat during these movements, not only do the velocity profilesof the ipsilateral and contralateral eyes peak at different values,but often they also exhibit differences in their dynamics.Therefore a good fit from the conjugate predictions would
indicate that the neuron equally encodes the motion of botheyes (i.e., encodes conjugate eye movements).
The first indication that unit B72_2 did not encode conjugateeye movements came from the poor conjugate predictionsshown in the top row of Fig. 3 (Pred-CS; VAFPred-CS � 0.45).Such low prediction VAFs were observed for all monocularunits (e.g., mean VAFPred-CS � 0.45 � 0.20, for the monocularipsilateral eye preference category; see Table 1). Another char-acteristic of monocular units was that the conjugate predictionstended to overshoot the firing rate when the preferred eye (in
FIG. 2. Discharge patterns of unit B72_2 during conjugate saccades. Neu-ron’s firing rate is shown as the gray shaded area (top row). Model fit obtainedusing Est-CS is shown as the thick black curve superimposed on the firing rate.Note the good fit to data. Also shown are ipsilateral eye (IE), contralateral eye(CE), and conjugate (CJ) velocity (2nd row) and position (4th row) traces, andvergence (VG) velocity (3rd row) and position (bottom row) traces. Note thedifferent line styles utilized for each trace. Vertical dotted lines denote saccadeonsets and offsets (20°/s criterion), and horizontal dotted lines represent 0velocity.
TABLE 1. Average conjugate, prediction, and estimation VAFs
Category n VAFCS VAFPred VAFEst-all VAFEst-red
Monocular, ipsilateral eye preference 17 0.67 � 0.11 0.45 � 0.20 0.60 � 0.15 0.59 � 0.16Monocular, contralateral eye preference 5 0.69 � 0.15 0.49 � 0.17 0.58 � 0.17 0.57 � 0.19Binocular, ipsilateral eye preference 10 0.72 � 0.09 0.65 � 0.09 0.74 � 0.07 0.73 � 0.08Binocular, contralateral eye preference 3 0.78 � 0.13 0.60 � 0.17 0.74 � 0.11 0.74 � 0.11Conjugate 15 0.64 � 0.14 0.51 � 0.16 0.58 � 0.11 0.57 � 0.11Population 50 0.68 � 0.12 0.52 � 0.18 0.63 � 0.14 0.62 � 0.14
Values are averages � SDs.
FIG. 3. Discharge patterns of monocular unit B72_2 during (A) 2 converg-ing disjunctive saccades and (B) 2 diverging disjunctive saccades. Top row:neuron’s firing rate with the model fit obtained with Pred-CS. Note theparticularly poor fit to data. 2nd row: same firing rate traces (duplicated forclarity), but with the model fits obtained using Est-ic-all (black curve) andEst-ic-red (dark gray curve; equation is also shown). Both fits were virtuallyidentical and modeled equally well the firing rate. 3rd–6th rows: velocity andposition traces recorded during these disjunctive saccades.
3456 P. A. SYLVESTRE AND K. E. CULLEN
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

this example the ipsilateral eye) moved less (Fig. 3A), and toundershoot the firing rate when the preferred eye moved more(Fig. 3B). Thus the conjugate-based prediction analysis sug-gested that unit B72_2 did not encode the conjugate move-ments of the eyes but rather that it exhibited a marked prefer-ence for the movements of the ipsilateral eye.
To directly quantify the sensitivity of individual ABNs dur-ing disjunctive saccades, we used the following two ap-proaches. First, we described neuronal discharges as a functionof the movements of each eye. This approach was motivated byrecent studies of premotor neurons in the saccadic burst gen-erator (Sylvestre and Cullen 1999b; Zhou and King 1998).Second, we utilized a conjugate/vergence based model to de-scribe the activity of the same neurons during the same dis-junctive saccades. This model structure follows from the pro-posal of Hering (1868) and is described in a subsequentsection.
When applied to unit B72_2, the ipsilateral/contralateral eyemovements-based approach first involved estimating the pa-rameters of the following model on the sample of disjunctivesaccades gathered for this neuron
FR�t� � bDS � ki�DSIE�t � td� � kc�DSCE�t � td� � . . .
. . . ri�DSIE�t � td� � rc�DSCE�t � td� �model Est-ic-all�
where bDS, ki-DS, kc-DS, ri-DS, and rc-DS are the bias, ipsilateraleye position, contralateral eye position, ipsilateral eye velocity,and contralateral eye velocity sensitivities of the neuron, re-spectively (DS, i, and c refer to disjunctive saccades, ipsilateraleye, and contralateral eye, respectively; ic in the model nameindicates the ipsilateral/contralateral eye based approach), andIE(t), CE(t), IE(t), and CE(t) are instantaneous ipsilateral andcontralateral eye positions and instantaneous ipsilateral andcontralateral eye velocities, respectively. This model is thebinocular expansion of Est-CS. Model fits obtained using Est-ic-all for unit B72_2 are shown in the second row of Fig. 3, Aand B (thick black curve). Clearly, this model fit was farsuperior to the conjugate model predictions (VAFEst-ic-all �0.66 vs. VAFPred-CS � 0.45; mean VAFEst-ic-all � 0.60 � 0.15vs. mean VAFPred-CS � 0.45 � 0.20, for the monocular ipsi-lateral eye preference category, Table 1). Although this obser-vation strongly supports the idea that unit B72_2 did notencode conjugate eye movements, it does not provide enoughinformation to determine if it solely encoded the movements ofone eye or a weighted mixture of both eyes’ movements.
To address this limitation, we estimated 95% confidenceintervals for each of the model parameters in Est-ic-all usingthe bootstrap technique described in METHODS. Figure 4 showsthe parameter estimates (vertical arrows) of Est-ic-all for unitB72_2 (left, eye velocity parameters; right, eye position pa-rameters), as well as the bootstrap distributions (histograms)and the 95% confidence intervals (thick horizontal bars) foreach parameter. Two important observations can be made fromthe 95% confidence intervals. First, for both the velocity andthe position parameters, the parameter values estimated for theipsilateral (ri-DS and ki-DS) and contralateral (rc-DS and kc-DS)eyes were statistically different (i.e., the confidence intervalsdid not overlap). This confirmed that unit B72_2 did notencode conjugate signals. Second, both the position and veloc-ity parameters for the contralateral eye had confidence intervalsthat overlapped with zero (i.e., were not statistically different
from 0). Therefore these parameters played no significant rolein modeling the neuron’s discharge dynamics.
When the position and velocity terms relating to the con-tralateral eye (rc-DS and kc-DS) were removed from Est-ic-alland the remaining model parameters were estimated for thisreduced model
FR�t� � bDS � ki�DSIE�t � td� � ri�DSIE�t � td� �model Est-ic-red�
the obtained fit was nearly identical to that of the full ipsilat-eral/contralateral eye-based model (Fig. 3, A and B, 2nd row,Est-ic-red, thick gray curves). Indeed, the goodness-of-fit ofthis reduced monocular model was the same as that of the fullipsilateral/contralateral eye-based model (VAFEst-ic-red �VAFEst-ic-all � 0.66). The model parameters of Est-ic-red forunit B72_2 are shown in Table 2, in the monocular ipsilateraleye category. Note that the parameter values estimated for the“meaningless” eye were appropriately replaced by zeros. Alsonote that Est-ic-red will not be the same for all neurons (seeExample conjugate ABN). Altogether, the prediction-based andestimation-based analyses clearly demonstrated that duringdisjunctive saccades, unit B72_2 encoded signals related to themotion of the ipsilateral eye only.
Example conjugate ABN
Figures 5–7 show the results of the same analysis of a typicalconjugate ABN, unit B27_1. This neuron discharged a vigor-ous burst of action potentials during conjugate saccades thatcould be well described using Est-CS (Fig. 5; VAFest-CS �0.69). However, in marked contrast to unit B72_2, the conju-gate predictions of the neuron’s discharge during disjunctivesaccades provided a fairly good fit to the data (Fig. 6, A and B,top rows, Pred-CS, thick black curve; VAFPred-CS � 0.54).This result, which was consistent across the category of con-jugate ABNs (mean VAFPred-CS � 0.51 � 0.16; Table 1),provided strong indications that unit B27_1 encoded conjugate
FIG. 4. Results of the bootstrap analysis for monocular unit B72_2. Left:results for eye velocity sensitivity of this neuron. Right: results for eye positionsensitivity. Histograms represent the distribution of parameter values obtainedwith the bootstrap analysis using Est-ic-all for the ipsilateral (black bars) andcontralateral (white bars) eye. Vertical arrows indicate mean value for eachparameter. Thick horizontal bars below histograms indicate the 95% confi-dence intervals associated with each parameter (black bar, ipsilateral eye;white bar, contralateral eye).
3457ABDUCENS NEURONS DURING DISJUNCTIVE SACCADES
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from
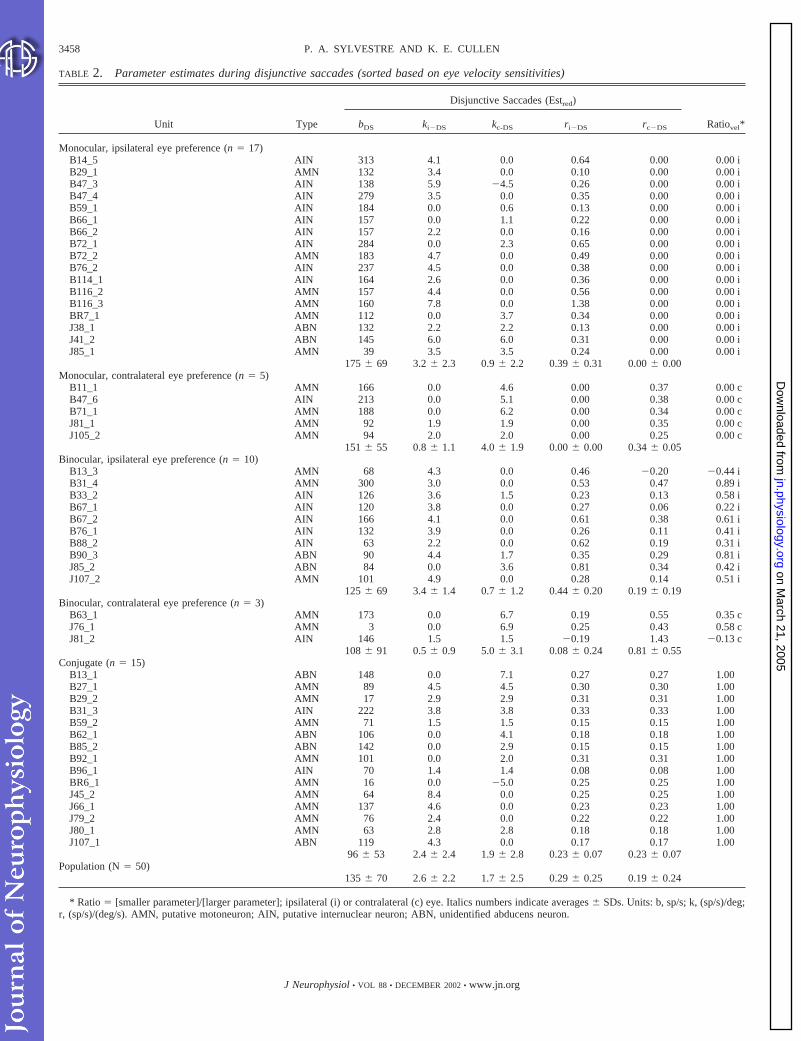
TABLE 2. Parameter estimates during disjunctive saccades (sorted based on eye velocity sensitivities)
Unit Type
Disjunctive Saccades (Estred)
Ratiovel*bDS ki�DS kc-DS ri�DS rc�DS
Monocular, ipsilateral eye preference (n � 17)B14_5 AIN 313 4.1 0.0 0.64 0.00 0.00 iB29_1 AMN 132 3.4 0.0 0.10 0.00 0.00 iB47_3 AIN 138 5.9 �4.5 0.26 0.00 0.00 iB47_4 AIN 279 3.5 0.0 0.35 0.00 0.00 iB59_1 AIN 184 0.0 0.6 0.13 0.00 0.00 iB66_1 AIN 157 0.0 1.1 0.22 0.00 0.00 iB66_2 AIN 157 2.2 0.0 0.16 0.00 0.00 iB72_1 AIN 284 0.0 2.3 0.65 0.00 0.00 iB72_2 AMN 183 4.7 0.0 0.49 0.00 0.00 iB76_2 AIN 237 4.5 0.0 0.38 0.00 0.00 iB114_1 AIN 164 2.6 0.0 0.36 0.00 0.00 iB116_2 AMN 157 4.4 0.0 0.56 0.00 0.00 iB116_3 AMN 160 7.8 0.0 1.38 0.00 0.00 iBR7_1 AMN 112 0.0 3.7 0.34 0.00 0.00 iJ38_1 ABN 132 2.2 2.2 0.13 0.00 0.00 iJ41_2 ABN 145 6.0 6.0 0.31 0.00 0.00 iJ85_1 AMN 39 3.5 3.5 0.24 0.00 0.00 i
175 � 69 3.2 � 2.3 0.9 � 2.2 0.39 � 0.31 0.00 � 0.00Monocular, contralateral eye preference (n � 5)
B11_1 AMN 166 0.0 4.6 0.00 0.37 0.00 cB47_6 AIN 213 0.0 5.1 0.00 0.38 0.00 cB71_1 AMN 188 0.0 6.2 0.00 0.34 0.00 cJ81_1 AMN 92 1.9 1.9 0.00 0.35 0.00 cJ105_2 AMN 94 2.0 2.0 0.00 0.25 0.00 c
151 � 55 0.8 � 1.1 4.0 � 1.9 0.00 � 0.00 0.34 � 0.05Binocular, ipsilateral eye preference (n � 10)
B13_3 AMN 68 4.3 0.0 0.46 �0.20 �0.44 iB31_4 AMN 300 3.0 0.0 0.53 0.47 0.89 iB33_2 AIN 126 3.6 1.5 0.23 0.13 0.58 iB67_1 AIN 120 3.8 0.0 0.27 0.06 0.22 iB67_2 AIN 166 4.1 0.0 0.61 0.38 0.61 iB76_1 AIN 132 3.9 0.0 0.26 0.11 0.41 iB88_2 AIN 63 2.2 0.0 0.62 0.19 0.31 iB90_3 ABN 90 4.4 1.7 0.35 0.29 0.81 iJ85_2 ABN 84 0.0 3.6 0.81 0.34 0.42 iJ107_2 AMN 101 4.9 0.0 0.28 0.14 0.51 i
125 � 69 3.4 � 1.4 0.7 � 1.2 0.44 � 0.20 0.19 � 0.19Binocular, contralateral eye preference (n � 3)
B63_1 AMN 173 0.0 6.7 0.19 0.55 0.35 cJ76_1 AMN 3 0.0 6.9 0.25 0.43 0.58 cJ81_2 AIN 146 1.5 1.5 �0.19 1.43 �0.13 c
108 � 91 0.5 � 0.9 5.0 � 3.1 0.08 � 0.24 0.81 � 0.55Conjugate (n � 15)
B13_1 ABN 148 0.0 7.1 0.27 0.27 1.00B27_1 AMN 89 4.5 4.5 0.30 0.30 1.00B29_2 AMN 17 2.9 2.9 0.31 0.31 1.00B31_3 AIN 222 3.8 3.8 0.33 0.33 1.00B59_2 AMN 71 1.5 1.5 0.15 0.15 1.00B62_1 ABN 106 0.0 4.1 0.18 0.18 1.00B85_2 ABN 142 0.0 2.9 0.15 0.15 1.00B92_1 AMN 101 0.0 2.0 0.31 0.31 1.00B96_1 AIN 70 1.4 1.4 0.08 0.08 1.00BR6_1 AMN 16 0.0 �5.0 0.25 0.25 1.00J45_2 AMN 64 8.4 0.0 0.25 0.25 1.00J66_1 AMN 137 4.6 0.0 0.23 0.23 1.00J79_2 AMN 76 2.4 0.0 0.22 0.22 1.00J80_1 AMN 63 2.8 2.8 0.18 0.18 1.00J107_1 ABN 119 4.3 0.0 0.17 0.17 1.00
96 � 53 2.4 � 2.4 1.9 � 2.8 0.23 � 0.07 0.23 � 0.07Population (N � 50)
135 � 70 2.6 � 2.2 1.7 � 2.5 0.29 � 0.25 0.19 � 0.24
* Ratio � [smaller parameter]/[larger parameter]; ipsilateral (i) or contralateral (c) eye. Italics numbers indicate averages � SDs. Units: b, sp/s; k, (sp/s)/deg;r, (sp/s)/(deg/s). AMN, putative motoneuron; AIN, putative internuclear neuron; ABN, unidentified abducens neuron.
3458 P. A. SYLVESTRE AND K. E. CULLEN
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

position and velocity signals and hence was equally sensitive tothe motion of both eyes.
The estimation of Est-ic-all confirmed this conclusion. First,the goodness-of-fit provided by Est-ic-all was only marginallybetter than that provided by the conjugate predictions (Fig. 6,A and B, 2nd row, Est-ic-all, thick black curve; VAFEst-ic-all �0.57; mean VAFEst-ic-all � 0.58 � 0.11, for the conjugatecategory; Table 1). Second, as is shown in Fig. 7, for both theeye velocity (left) and eye position (right) sensitivities of theneuron, the estimated parameter values (vertical arrows) werevery similar for the ipsilateral and contralateral eyes. Indeed,the bootstrap distributions (histograms) and the 95% confi-dence intervals (thick horizontal bars) overlapped for the ipsi-lateral and contralateral eye parameters (i.e., ri-DS � rc-DS, andki-DS � kc-DS, at 95% confidence), but did not overlap withzero. It can be concluded from these results that unit B27_1was equally sensitive to the position and velocity of the twoeyes, and hence that it encoded conjugate position and velocitysignals.
As for the monocular unit described previously, this conclu-sion was strengthened by replacing the monocular position andvelocity signals in Est-ic-all by conjugate signals [i.e., Est-ic-red for this neuron: FR(t) � bCS � kCSCJ(t � td) � rCS CJ(t �td)]. This reduced model (Fig. 6, A and B, 2nd rows, thickgray curve) provided a goodness-of-fit that was identical tothat of Est-ic-all (VAFEst-ic-red � VAFEst-ic-all � 0.57; meanVAFEst-ic-red � 0.57 � 0.11, for the conjugate category; Table1). Furthermore, the parameter values (Table 2) that were
estimated using Est-ic-red were comparable to those estimatedduring conjugate saccades (P � 0.5, paired t-test). Note that inTable 2, the parameter values for neurons with conjugateocular preferences are represented as their monocular equiva-lent (see Eq. 2). Thus, when taken together, the prediction-based and estimation-based analyses clearly demonstrated thatunit B27_1 encoded conjugate signals during disjunctive sac-cades.
Population distributions
The parameter values obtained for each ABN in our sampleusing Est-ic-red are shown in Table 2. Neurons are grouped infive categories according to the eye velocity-based criteriadescribed below. Average parameter values (�SDs) are in-cluded for each category and for the entire sample. To quantifythe “ocular-preference” of a given neuron, ratios of ipsilateral
FIG. 6. Discharge patterns of conjugate unit B27_1 during (A) 2 convergingdisjunctive saccades and (B) 2 diverging disjunctive saccades. Note the goodfit to data provided by PredCS (top row). Also shown are the model fitsobtained using Est-ic-all (black curve) and Est-ic-red (dark gray curve; equa-tion is also shown). For this neuron, Est-ic-red had the same structure asPred-CS.
FIG. 5. Discharge patterns of a 2nd example neuron, unit B27_1, duringconjugate saccades. Model fit obtained using Est-CS, which provided a good fitto data, is shown as the thick black curve superimposed on firing rate.
3459ABDUCENS NEURONS DURING DISJUNCTIVE SACCADES
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

and contralateral eye velocity (Ratiovel) and eye position(Ratiopos) parameters were calculated as follows using theparameter values from Table 2 (Est-ic-red)
Ratio � �smaller parameter value�/�larger parameter value�
To indicate which eye yielded the larger parameter (par.) value,each Ratio value is accompanied by an i or a c, for theipsilateral (ipsi.) or contralateral (contra.) eye, respectively.The Ratiovel values were utilized to sort neurons in the cate-gories shown in Tables 1 and 2, and can be interpreted asfollows
Hence, monocular unit B72_2 shown in Figs. 2– 4 had aRatiovel � 0i, while conjugate unit B27_1 shown in Figs.5–7 had a Ratiovel � 1 (note that in this case, the i or c
character was omitted because both parameters had equalvalues). This Ratio index was chosen because its interpre-tation is more intuitive than that of other indexes utilized inprevious studies. For example, for the index utilized in thepresent study, values of 0.1i and 0.5i simply represent ratiosof ipsilateral to contralateral eye parameters of 10:1 and 2:1,respectively. On the other hand, the same values of 0.1 and0.5 for a common index ([ipsi. eye � contra. eye]/[ipsi.eye � contra. eye]; see for example Zhou and King 1998)represent ratios of ipsilateral to contralateral eye parametersof 1.22:1 and 3:1, respectively.
A graphical summary of the Ratio values is presented in Fig.8, where the distributions of Ratiovel (Fig. 8A) and Ratiopos
(Fig. 8B) for our sample of neurons are shown. With respect tothe eye velocity sensitivity of ABNs during disjunctive sac-cades, many ABNs in our sample (44%; light and dark redbars, Fig. 8A) exhibited monocular velocity sensitivities (i.e.,
FIG. 7. Results of the bootstrap analysis for conjugate unit B27_1. Left:results for eye velocity sensitivity of this neuron. Right: results for eye positionsensitivity, using the same conventions as for Fig. 4.
FIG. 8. A: distribution of Ratiovel during disjunctive saccades for our sam-ple of abducens nucleus neurons (ABNs). Columns marked with asterisksindicate neurons that had a vergence sensitivity greater than one-half theirconjugate sensitivity. Red, blue, and green bars denote monocular, conjugate,and binocular units, respectively. B: distribution of Ratiopos during disjunctivesaccades for our sample of ABNs. The conventions utilized were the same asin A. C: coherence of the preferred eyes for the position and velocity sensitivitiesof ABNs during disjunctive saccades. The x and y axes plot the 3 preferred eyecategories (ipsilateral eye, Ipsi; contralateral eye, Contra; or conjugate, Conj) forthe position and velocity sensitivities, respectively. Black columns indicate coher-ence (i.e., the eye position and the eye velocity preferred eyes of a neuron were thesame), and gray bars indicate absence of coherence.
Monocular, ipsi. eye preference Ratio � 0, Subscript � “i”.� contra. par. � 0
Monocular, contra. eye preference Ratio � 0, Subscript � “c”.� ipsi. par. � 0
Binocular, ipsi. eye preference 1 � Ratio � 0 (same signs),� ipsi. par. � contra. par. 0 � Ratio � �1 (diff. signs),� ipsi. par. contra. par. 0 Subscript � “i”.
Binocular, contra. eye preference 1 � Ratio � 0 (same signs),� contra. par. � ipsi. par. 0 � Ratio � �1 (diff. signs),� ipsi. par. contra. par. 0 Subscript � “c”.
Conjugate Ratio � 1, no subscript.� ipsi. par. � contra. par. 0
3460 P. A. SYLVESTRE AND K. E. CULLEN
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

Ratiovel � 0). Of these monocular ABNs, 73% preferred theipsilateral eye (light red bars, Fig. 8A). Furthermore, 30% ofthe ABNs in our sample equally encoded the velocity of botheyes (i.e., conjugate, Ratiovel � 1; blue bar, Fig. 8A). Theremaining 26% of ABNs encoded the motion of both eyes(light and dark green bars, Fig. 8A), of which 77% favored theipsilateral eye (light green bars, Fig. 8A). Only two neuronsencoded opposite ON directions for the two eyes (Ratiovel 0).The distribution of Ratiopos (Fig. 8B) was similar to that ofRatiovel. Most neurons in our sample (70%) exhibited monoc-ular preferences. Of these monocular ABNs, 63% preferred theposition of the ipsilateral eye. In addition, 24% of all ABNstested were equally sensitive to the position of both eyes. Onlyone neuron had a negative Ratiopos. The main difference be-tween the distributions of Ratiopos and Ratiovel was thatslightly more units were binocular with respect to their velocitysensitivity than to their position sensitivity (24% vs. 6%, ve-locity and position sensitivities, respectively).
We conclude that during disjunctive saccades, our sample ofABNs is dominated by a subpopulation of neurons that mo-nocularly encodes the motion of the ipsilateral eye (i.e., “mon-ocular with ipsilateral eye preference”; Fig. 8) and by a secondless pronounced subpopulation that encodes the conjugate mo-tion of the eyes (i.e., “conjugate”; Fig. 8). As a result of thisdistribution, the average sensitivity to the velocity of the ipsi-lateral eye for our sample of ABNs was 1.5 times larger thanthat for the contralateral eye. The average eye position sensi-tivity of our sample of ABNs to the ipsilateral eye was also 1.5times larger than that of the contralateral eye.
Coherence of the “preferred eye” for the position andvelocity coefficients
For each neuron in our sample, our analysis approach iden-tified a “preferred eye” (defined as the eye that yielded thelargest parameter value) for both the position and the velocitysensitivities. Here, we asked whether the preferred eye for theposition and velocity sensitivities of ABNs were matched on aneuron-by-neuron basis. To do so, we regrouped our data underthree general categories: ipsilateral eye preference category(grouping the “monocular with ipsilateral eye preference” and“binocular with ipsilateral eye preference” cell types; Table 2),contralateral eye preference category (grouping the “monocu-lar with contralateral eye preference” and “binocular withcontralateral eye preference” cell types; Table 2), and conju-gate category (Table 2). Hence, a total of nine permutationsrepresent all the possible combinations of preferred eyes for theposition and velocity sensitivities.
The fraction of neurons that fell within each of the ninepossible categories are illustrated in Fig. 8C, where the x andy axes represent the three preferred eye categories for theposition and velocity sensitivities, respectively, and the z axisrepresents the percentage of neurons that fell within eachcategory. As is shown by the black columns, the majority ofneurons (58%) exhibited coherence between their preferred eyefor the position and velocity sensitivities (i.e., had the samepreferred eye). Of those neurons, 62% preferred the ipsilateraleye, 17% preferred the contralateral eye, and 21% were con-jugate. With the exception of noncoherent neurons that en-coded ipsilateral position/conjugate velocity eye preferencesand were equally numerous as those that exhibited conjugate
coherence, no trend could be identified for the other categoriesof noncoherent neurons; they were approximately uniformlydistributed over the remaining five combinations of preferredeyes (gray columns). Thus during disjunctive saccades, a ma-jority of ABNs exhibited coherence in their preferred eye forthe position and velocity sensitivities.
Testing the alternative conjugate/vergence approach
In our second approach, we utilized a conjugate/vergencebased model to describe the activity of the same neurons
FR�t� � bDS � kcj�DSCJ�t � td� � kvg�DSVG�t � td� � . . .
. . . rcj�DSCJ�t � td� � rvg�DSVG�t � td� �model Est-cv-all�
where cj and vg refer to conjugate and vergence-related param-eters, respectively, cv in the model name indicates the conju-gate/vergence based approach, and CJ(t), VG(t), CJ(t), andVG(t) are instantaneous conjugate and vergence eye positionsand velocities, respectively. As for Est-ic-all, we estimated theparameters of this model on our entire data set of neuronalactivities and computed bootstrap confidence intervals for allof the parameters. The latter were then used to reduce themodel to its simplest form (Est-cv-red; note that this model canvary from neuron to neuron).
In its nonreduced form, Est-cv-all is mathematically equiv-alent to Est-ic-all. Accordingly, the VAF values obtained withboth models were identical on a neuron-by-neuron basis. Fur-thermore, when the parameters of Est-cv-all were converted tothose of Est-ic-all using the following relationships (shown fora neuron recorded to the left of the midline)
ki�DS �kcj�DS
2� kvg�DS; kc�DS �
kcj�DS
2� kvg�DS (2a)
ri�DS �rcj�DS
2� rvg�DS; rc�DS �
rcj�DS
2� rvg�DS (2b)
the parameters obtained with either model were all statisticallyidentical (paired t-tests, P � 0.05). However, because Est-ic-all and Est-cv-all are not always equivalent after one or moreparameters have been removed and because the parameters inthese models can take markedly different numerical values andhave an inherent variability (i.e., have confidence intervalswith nonnegligible widths), it was not possible to utilize Eq. 2to derive Est-cv-red from Est-ic-red. Stated differently, wecould not assume that if the bootstrap confidence intervals ofthe ipsilateral and contralateral eye parameters in Est-ic-alloverlapped slightly (which we interpreted as conjugacy), theconfidence interval of the vergence term in Est-cv-all wouldautomatically overlap with zero. Hence, we properly evaluatedEst-cv-red by independently computing bootstrap confidenceintervals for the conjugate/vergence based model Est-cv-all onall the neurons in our sample.
Figure 9 shows the results of this conjugate/vergence anal-ysis for our population of neurons. Note that to allow directcomparisons of these results with those described in the pre-vious sections, we processed the parameters of Est-cv-red withEq. 2 to obtain the equivalent parameter values in ipsilateral/contralateral eye coordinates, and then computed Ratiovel andRatiopos indexes as described above. As illustrated in Fig. 9 bythe axis labels between square brackets, a conjugate unit (ver-
3461ABDUCENS NEURONS DURING DISJUNCTIVE SACCADES
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

gence-related parameters nonsignificant) will yield a Ratioindex of 1, a vergence unit (conjugate-related parameters non-significant) will yield a Ratio of �1, and a “monocular” unit(conjugate-related parameters twice bigger than the absolutevergence parameters; see Eq. 2) will yield a Ratio of 0. Thedistribution in Fig. 9A shows the Ratiovel values calculatedusing Est-cv-red. This distribution can be directly comparedwith that obtained with Est-ic-red (Fig. 8A). A first importantobservation was that the main features of the two distributionswere similar in that both showed two predominant peaks [i.e.,monocular (Ratio � 0) and conjugate (Ratio � 1) peaks].Similar results were also observed for the Ratiopos values(compare Figs. 9B and 8B). However, the distributions ob-tained with the two types of reduced models differed in that thenumber of monocular versus conjugate units was less based onthe conjugate/vergence approach. The number of units withbinocular tuning for their eye position and velocity sensitivitieswas also higher using the conjugate/vergence approach.
Because the analyses using models Est-ic-all and Est-cv-allyielded slightly different results, we sought to determine whichof the two provided the most appropriate description of ABNdischarges. To do so, we analyzed the VAF values generatedby these two models. For 64% of the neurons in our sample,Est-ic-red yielded VAF values that were clearly larger than
those obtained with Est-cv-red (7 � 11%). In contrast, for theremaining neurons, the VAF values obtained with Est-cv-redwere only slightly larger than those obtained with Est-ic-red(2 � 2%). Hence, for almost two-thirds of the neurons in oursample, model Est-ic-red provided markedly better goodness-of-fits than model Est-cv-red, while the latter model onlyprovided marginally (if at all) better fits for the remainingneurons. Furthermore, and consistent with these results, theVAF values obtained with Est-ic-red were, on average, only1% smaller than those obtained with Est-ic-all, while thoseobtained with Est-cv-red were 5% smaller than those obtainedwith Est-cv-all [recall that VAF(Est-ic-all) � VAF(Est-cv-all)]. Thus removing conjugate or vergence parameters fromEst-cv-all (based on the bootstrap statistics) was far moredetrimental to the goodness-of-fit than removing ipsilateral orcontralateral eye parameters from Est-ic-all. We conclude thatipsilateral/contralateral eye based models were better suited forour analysis than conjugate/vergence based models.
Responses during off-direction disjunctive saccades
In good agreement with our previous findings (Sylvestre andCullen 1999a), the majority of ABNs in our sample (82%)were driven into inhibitory cutoff (i.e., “paused”) during all OFF
direction conjugate saccades. Similarly, most ABNs (64%)were also driven into inhibitory cutoff during all OFF directiondisjunctive saccades. Whether the saccade was divergent orconvergent did not affect the pausing behavior of these ABNs.Discharge patterns from a representative neuron in this cate-gory, unit J66_1, are shown in Fig. 10A during converging anddiverging OFF-direction saccades. For the remaining ABNs, theamplitude of the conjugate movement appeared to be the maindeterminant of their pausing behaviors, since the neurons’discharges were comparable during converging and divergingsaccades. Of these neurons, the majority (67%) paused com-pletely for disjunctive saccades with conjugate components�10°. In turn, 33% paused only for disjunctive saccades withconjugate amplitudes �20°. Figure 10B shows example dis-junctive saccades from a neuron in this latter category (unitB76_1). Note that the neuron clearly paused for large ampli-tude converging and diverging saccades (right). Also note that,as for all neurons that did not always reach inhibitory cutoff,there was nevertheless a significant decrease in firing rate whenthe neuron did not pause. Thus the pausing behavior of ABNsis generally similar during conjugate and disjunctive saccades,with the exception that for movements of small amplitudes,slightly more ABNs pause during conjugate saccades.
Comparison of disjunctive saccades and disjunctive fixation
We next addressed whether individual ABNs retain the samepreferred eye during disjunctive saccades and disjunctive fix-ation. For each neuron, we fitted its average firing rate as afunction of the average ipsilateral and contralateral eye posi-tions during intervals of disjunctive fixation. We next com-puted a RatioFIX value for each neuron using the same proce-dure as defined above for calculating RatioPOS duringdisjunctive saccades.
Figure 11A shows the distribution of RatioFIX during dis-junctive fixation. This distribution was similar to the distribu-tion of Ratiopos observed during disjunctive saccades (compare
FIG. 9. A: distribution of Ratiovel during disjunctive saccades obtained withEst-cv-red for our sample of ABNs. Ratio indexes are equivalent to those inFig. 8 (see text for details). The axis labels between square brackets illustratethe relationship between the Ratio values and the conjugate/vergence param-eter values. Conventions for red, blue, and green bars are as in Fig. 8. B:distribution of Ratiopos during disjunctive saccades obtained with Est-cv-redfor our sample of ABNs. The conventions utilized were the same as in A.
3462 P. A. SYLVESTRE AND K. E. CULLEN
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

Figs. 11A and 8A). The main difference between the twodistributions was that a greater proportion of ABNs encodedthe position of both eyes during disjunctive fixation versusdisjunctive saccades. As a consequence, during disjunctivefixation, fewer ABNs (48%) encoded the position of a singleeye (of which 75% preferred the ipsilateral eye), while acomparable number of ABNs (26%) encoded the conjugateposition of the eyes. Thus at the population level, ABNsgenerally encode the position of the two eyes in a similarmanner during disjunctive saccades and disjunctive fixation.For our sample of ABNs, the average sensitivity to the ipsilat-eral eye position during disjunctive fixation was 3.1 timeslarger than that of the contralateral eye. This ratio is larger thanthat observed during disjunctive saccades (1.5) because theparameter values estimated during fixation were larger thanthose estimated during disjunctive saccades. This result isconsistent with our previous finding that the eye positionsensitivities of ABNs decrease as the eye velocity increases(Sylvestre and Cullen 1999a). We attributed this observation tothe changes in antagonist/agonist muscle interactions that oc-cur at different eye velocities.
On a neuron-by-neuron basis, ABNs exhibited good coher-ence between their preferred eye (eye position sensitivity)during disjunctive saccades and disjunctive fixation. This isillustrated in Fig. 11B, where for each neuron in our sample,the preferred eye during disjunctive fixation was plotted versusthe preferred eye (position sensitivity) during disjunctive sac-cades. The majority of ABNs (60%) exhibited coherencebetween their preferred eye during these two behavioralconditions (black columns). No consistent pattern could berecognized for the remaining neurons. Thus these resultsclearly demonstrate that ABNs have similar ocular preferencesduring fixation and saccadic behaviors.
Putative motoneurons versus internuclear neurons
Figure 12 shows the relative distribution of preferred eyeposition and velocity sensitivities for the putative AMNs (ab-ducens motoneurons) and AINs (internuclear neurons) in oursample. Note that eight neurons (labeled ABN in Table 2)could not be classified as AINs or AMNs using the identifica-tion criteria described in METHODS and were excluded from thefollowing analysis. With respect to the eye position sensitivi-
FIG. 11. A: distribution of Ratiopos during disjunctive fixation for oursample of ABNs. Conventions for red, blue, and green bars are as in Fig. 8. B:coherence of the preferred eyes for the position sensitivities of ABNs duringdisjunctive saccades and disjunctive fixation. The x and y axes plot the 3preferred eye categories for the eye position sensitivities during fixation andsaccades, respectively. The other conventions are the same as for Fig. 8C.
FIG. 10. ABN discharge patterns during OFF-direction disjunctive saccades.A: 2 smaller (left pair; first is diverging, second is converging) and larger (rightpair; first is diverging, second is converging) disjunctive saccades for a typicalABN, unit J66_1. The same conventions as Fig. 2 were utilized (note that forclarity, only position traces are shown). As for most units in our sample, thisneuron completely ceased firing (paused) for disjunctive saccades of all am-plitudes. Whether the saccade was divergent or convergent did not affect thepausing behavior (e.g., compare left and right small amplitude saccades). B:example saccades from another neuron, unit B76_1, that only paused for largeramplitude disjunctive saccades. Note that the neuron’s firing rate decreasedduring smaller amplitude disjunctive saccades (left), but not sufficiently toreach cutoff.
3463ABDUCENS NEURONS DURING DISJUNCTIVE SACCADES
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

ties (Fig. 12, left), our results suggest that a slightly greaterproportion of AINs than AMNs (61% vs. 50%) preferentiallyencoded the position of the ipsilateral eye. In turn, a greaterproportion of AMNs encoded the conjugate position of botheyes (29% vs. 17%, AMNs vs. AINs, respectively). Thesetrends were more pronounced for the eye velocity sensitivities(Fig. 12, right). However, for our samples of putative AMNsand AINs, their relative distributions across the five categoriesof preferred eye shown in Table 2 were not statistically sig-nificant (�2 test on a 2 � 5 contingency table; P � 0.50 andP � 0.10, for position and velocity sensitivities, respectively).Thus we conclude that putative AMNs and AINs encode eyemovements in a similar manner during disjunctive saccades.
D I S C U S S I O N
In this report, we provide the first characterization of abdu-cens nucleus neuron discharges during disjunctive saccades.The analysis approach that we utilized allowed us, for eachneuron, to reduce a generic ipsilateral/contralateral eye-basedmodel of ABNs firing rate (i.e., Est-ic-all) to a model that onlyincluded the terms that significantly modulated the neuron’sdischarge dynamics (i.e., Est-ic-red). Our main finding wasthat most ABNs preferentially encoded the motion and theposition of the eye ipsilateral to the recording site duringdisjunctive saccades; the remaining neurons predominantlyencoded the conjugate motion of the eyes. At the populationlevel, the average eye position and eye velocity sensitivities ofabducens neurons were approximately 50% larger for the ip-silateral eye than for the contralateral eye. When the analysiswas repeated using a conjugate/vergence based model, thegoodness-of-fit obtained for the reduced models (Est-cv-red)was in most cases lower or equivalent to that obtained whenusing an eye-based model (Est-ic-red). This finding indicatesthat ABN discharges, during disjunctive saccades, are betterdescribed using ipsilateral and contralateral eye movementparameters. We also found that the OFF-direction responses ofABNs during conjugate and disjunctive saccades were gener-ally similar. Moreover, there were no significant differences inthe physiological properties of putative AMNs and AINs. Thislatter result agrees with, and complements, the findings ofprevious studies which have shown that during disjunctivefixation, AINs carry a signal that is “inappropriate” to drive the
medial rectus of the contralateral eye (Gamlin et al. 1989).Finally, we found that individual ABNs generally encode theposition of the same eye during disjunctive saccades and fix-ation.
Implications for the motor drive to the agonist lateral rectus
During conjugate saccades, there is good evidence that neg-ligible co-contraction occurs (i.e., simultaneous contraction ofthe agonist and antagonist muscles for a given eye; Fuchs andLuschei 1970; Robinson 1970; Schiller 1970). Therefore themovement of an abducting eye reflects accurately the motorcommand carried by AMNs to the lateral rectus of that eye, asthe medial rectus only provides passive resistance to the move-ment. However, we found that during disjunctive saccades thedischarge dynamics of ABNs, on a neuron-by-neuron basis, didnot exclusively reflect the motion of the ipsilateral eye. Thequestion therefore arises as to whether ABNs, at the populationlevel, encode signals that are sufficient to control the move-ments of the ipsilateral eye.
To address this question, we performed a simple simulationof the population drive to the lateral rectus of the ipsilateral eye(in this case, the right eye) that would have been generatedduring a conjugate and a disjunctive saccade. First, we selecteda typical conjugate saccade from our data set (Fig. 13A, #1).Next, we utilized the dynamic models estimated on the actualdata to reconstruct the firing rate that each neuron in oursample would have had during this saccade (FR1:FRN; n � 50).To determine whether we could predict the activity of ourpopulation of neurons during both conjugate and disjunctivesaccades based on the sensitivity of each neuron to ipsilateraland contralateral eye movement, model Est-ic-red (Table 2)was selected for this analysis. The resulting N firing rates wereaveraged to provide an estimate of the population drive duringthis particular conjugate saccade (Fig. 13A, #3, solid curve).We then performed a comparable simulation using a hypothet-ical disjunctive saccade for which the right eye’s motion wasidentical to that of the conjugate saccade in #1, but the lefteye’s motion was markedly reduced (Fig. 13A, #2). If oneassumes that the agonist drive alone shapes the motion of theright eye during all saccades, then the two population drivesproduced in our simulation should be identical since the move-ment of the right eye was the same for both saccades. Indeed,as is illustrated in #3 of Fig. 13A, the population drives gen-erated in both cases were quite similar. Thus the populationdrive generated by ABNs can, by itself, account for most of theipsilateral eye movement during disjunctive saccades. Notethat identical conclusions were drawn when we repeated thisexercise using the parameter values obtained from the conju-gate/vergence-based analysis (Est-cv-red).
Although the population drive computed above could pro-vide most of the drive necessary to move the ipsilateral eye, thearea under the population drive during the disjunctive saccadewas nevertheless approximately 15% smaller than that com-puted for the conjugate saccade (Fig. 13A, #3, gray shadedarea; see legend). This is not surprising given that a significantpercentage (66%) of the neurons in our sample were sensitiveto the contralateral eye, which in our simulation moved lessduring the disjunctive saccade. To account for these appar-ently inappropriate signals at the level of ABNs, it is im-portant to note that a number of simplifications were made
FIG. 12. Distribution of eye position (left) and eye velocity (right) sensi-tivities of putative abducens motoneurons (AMNs; top row) and abducensinternuclear neurons (AINs; bottom row). Conventions for red, blue, and greenbars are as in Fig. 8.
3464 P. A. SYLVESTRE AND K. E. CULLEN
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

in this simple simulation. First, it was postulated that allneurons in our sample were AMNs, while we know that oursample contained many putative AINs (Fig. 13B, #1). Sec-ond, all neurons in our simulation were assumed to haveequal synaptic weights, although unequal weighting of theAMN projections to the lateral rectus most certainly exists(Fig. 13B, #2). Finally, the simulation did not take intoconsideration the possibility of co-contraction between thelateral and medial recti during disjunctive saccades (Fig.13B, #3). The physiological relevance of these mechanismsis discussed in the following text.
It is possible that the neurons in our sample that exhibitedstronger tuning to the motion of the ipsilateral eye were all
AMNs, while those that encoded the motion of the contralat-eral eye or the conjugate eye motion were AINs (Fig. 13B, #1).Based on this scenario, the motor drive sent to the lateral rectusduring all saccades would be appropriate to move the ipsilat-eral eye, without further inputs required to the oculomotorplant. However, when we repeated the simulation shown inFig. 13A using only the putative AMNs in our sample (seeTable 2), our conclusions remained unchanged: the populationdischarge was still markedly smaller (approximately 18% de-crease in total area) during the disjunctive saccade. We alsoobserved experimentally that the physiological properties ofputative AMNs and AINs during disjunctive saccades did notdiffer significantly. The small nonsignificant differences thatwe measured still could not account for the results of oursimulation. For example, more AMNs encoded conjugate sig-nals than AINs (Fig. 12), while the opposite might be expected.Thus, although we cannot completely rule out that the dis-charge properties of AMNs and AINs are selectively tuned tothe motion of the eye to which they project, we argue that thismechanism plays a minor role during disjunctive saccades.
It is also conceivable that a sampling bias is responsible forthe apparently inappropriate signals at the level of ABNsdescribed above. Recent studies have identified two morpho-logically distinct classes of abducens neurons that appear toserve different physiological roles (Buttner-Ennever et al.2001, 2002). The first class is composed of neurons with largesomas distributed throughout the nucleus and that are dividedinto motoneurons that innervate twitch fibers (Buttner-Enneveret al. 2001) and internuclear neurons that project to the con-tralateral oculomotor nucleus (Destombes et al. 1979; McCreaet al. 1986; Spencer and Sterling 1977; Steiger and Buttner-Ennever 1978). The discharge properties of these neurons areconsistent with them mediating all types of eye movements(Delgado-Garcia et al. 1986a,b; Fuchs et al. 1988; Mays andPorter 1984). In contrast, the second class is formed of small tomedium-size motoneurons located in a shell-like structurearound the medial edge of the abducens nucleus (Buttner-Ennever et al. 2001). These peri-abducens motoneurons inner-vate nontwitch muscle fibers whose function remains ill de-fined and receive premotor signals from the neural integrator,the smooth pursuit, and the vergence premotor areas, but notfrom the premotor saccadic burst neurons (Buttner-Ennever etal. 2001, 2002). Thus, given that 1) we were more likely tosample the activity of large neurons with our single unit re-cording techniques and 2) all of the neurons in our sampledischarged in relation to saccadic eye movements, we concludethat our sample contained few if any of the smaller neurons ofthe nucleus shell. Whether the smaller neurons contribute tooffset the observed discrepancy during disjunctive saccadesremains to be determined.
Selective weighting of AMNs at the level of the lateralrectus could also be utilized to further “monocularize” thesignals carried to this muscle (Fig. 13B, #2). For example, unitsthat better encode the motion of the ipsilateral eye could makemore efficient synapses, while units that better encode themovements of the contralateral eye would make weaker syn-apses. This proposed mechanism is consistent with the resultsof our simulation, where the population drive of ABNs duringthe disjunctive saccade was too small to generate the move-ment of the right eye (Fig. 13A, #3). Selectively increasing theweight of the motoneurons that are better tuned to the move-
FIG. 13. A: simulation of abducens population drive during a conjugatesaccade and a disjunctive saccade. Example shown here is discussed for theright abducens nucleus (see text for other assumptions). Right and left eyeposition and velocity traces utilized for the conjugate saccade are shown in #1.Disjunctive saccade shown in #2 was derived from #1 to ensure that the righteye motion was identical for both saccades. To do so, the left eye’s motion wasscaled down to 25% of its original amplitude. N parameter sets shown in Table2 (i.e., Est-ic-red) were utilized to reconstruct the firing rates that each neuronin our sample would have generated during these 2 saccades. Note that asimilar procedure was repeated with Est-cv-red. The average firing ratesobtained with this approach are shown in #3. Solid curve is for the conjugatesaccade; dotted curve is for the disjunctive saccade. Gray shaded area empha-sizes difference between the 2 average firing rates. Its relative area, calculatedas (�conjugate population drive � �disjunctive population drive)/(�conjugatepopulation drive), represents the percent difference in the amplitude of the 2population drives (15% in this example). B: suggested mechanisms. In #1, theproperties of AINs and AMNs are distinct: the former, which project to thecontralateral eye, encode signals related to this eye, while the latter, whichproject to this ipsilateral lateral rectus, encode signals related to the motion ofthis eye. In #2, the weighting of AMNs that preferentially carry signals relatedto the motion of the ipsilateral eye are selectively increased. Finally, in #3,co-contraction of the lateral and medial recti modulates the impact of theagonist muscle contraction on the eye movement.
3465ABDUCENS NEURONS DURING DISJUNCTIVE SACCADES
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

ment of the ipsilateral eye could compensate for this deficit.We conclude that the selective weighting of AMN projectionsis most certainly a predominant mechanism during disjunctivesaccades. Another way to increase the population drive duringdisjunctive saccades would be to have neurons in the abducensnucleus that are more sensitive to the vergence than to theconjugate eye movements. However, our sample of ABNs didnot contain any neurons with such properties. In fact, only twoneurons in our sample had vergence sensitivities more thanone-half their conjugate sensitivities (see asterisk in Fig. 8).
Finally, co-contraction of the medial and lateral recti of agiven eye could also occur during disjunctive saccades (Fig.13B, #3). This mechanism, however, would impede the move-ment of the eye during the saccade (i.e., it would compete withthe agonist muscle in a push-pull manner). As a result, co-contraction would not compensate for the lack of agonist driveillustrated in Fig. 13A, but rather would accentuate it. More-over, our experimental results suggest that there is negligibleco-contraction during most disjunctive saccades (Fig. 10). Forexample, during saccades, the OMNs that control the medialrectus of the right eye (i.e., the antagonist muscle of that eye)receive strong inputs from the AINs on the left side of the brainstem. Because our analysis demonstrated that virtually all ofthese left AINs would pause completely during most largeamplitude disjunctive saccades to the right (i.e., their OFF-direction is to the right), we can infer that the majority ofOMNs are disfacilitated during these movements, and thatminimal co-contraction occurs. This conclusion might differfor smaller amplitude disjunctive saccades, since approxi-mately one-third of ABNs do not pause completely duringthese movements (e.g., Fig. 10B). As a consequence, OMNswould be less disfacilitated, and the medial rectus of the righteye would actively contract to oppose the drive from theabducens nucleus to the lateral rectus of this same eye. Recall,however, that co-contraction of the medial and lateral rectiwould be counter-productive to the ongoing eye movement.
Implications for the motor drive to the agonist medial rectus
Another implication of our results is that the signals sent byAINs to the OMNs that drive the motion of the contralateraleye (re the recording side) do not encode the monocular posi-tion and velocity of that eye. Such an observation has also beenmade on identified internuclear neurons during disjunctivefixation (Gamlin et al. 1989). To ensure the proper movementof that eye, additional mechanisms at or downstream to thelevel of the oculomotor motoneurons must be used to comple-ment the signal carried by AINs. Of course, the mechanismsdescribed above for the drive to the lateral rectus (Fig. 13B)can also be applied to the drive to the medial rectus. Inaddition, OMNs may receive additional vergence-related pre-motor commands from neurons located nearby the oculomotornucleus (see review by Gamlin 1999; Mays and Gamlin1995a). Some of these neurons, the vergence-velocity neurons,are known to encode the velocity of pure vergence shifts (Mayset al. 1986) and have also been described to burst duringdisjunctive saccades (Mays and Gamlin 1995b). It has beenproposed that vergence-velocity neurons project to the OMNsvia the near response cells (Mays and Gamlin 1995a). Theselatter neurons encode vergence position and velocity signalsduring pure vergence shifts (Judge and Cumming 1986; Mays
1984; Zhang et al. 1992) and project monosynaptically to theoculomotor nucleus (Zhang et al. 1991, 1992). This mechanismmust be exclusive to the medial rectus motoneurons, however,as no vergence-related neurons that project to the abducensnucleus have been identified (Gamlin 1999).
Implications for the premotor drive to the abducens nucleus
During conjugate saccades, most of the premotor drive thatis related to the velocity of the eyes originates from thesaccadic burst neurons, while the drive related to the positionof the eyes comes from neurons in the neural integrator (re-viewed by Moschovakis et al. 1996). What remains to beelucidated is the source of the vergence signals carried byABNs during disjunctive saccades. On the one hand, previousstudies have suggested that during disjunctive eye movements,both saccadic burst neurons (Sylvestre and Cullen 1999b; Zhouand King 1998) and neurons in the prepositus hypoglossi/vestibular nuclei (McConville et al. 1994; Zhou and King1996) carry signals related to the movements of both eyes, andoften to the motion of a single eye. Furthermore, in all of thesestructures, neurons that preferred either the ipsilateral or thecontralateral eye were found. These neural pathways couldtherefore provide ABNs with drives that are consistent with ourfinding that a large proportion of ABNs encode signals relatedto the motion of a single eye. On the other hand, neither thevergence-related neurons near the IIIrd nucleus, nor any otheridentified pure vergence-related neurons, project to the abdu-cens nucleus (Gamlin 1999). Furthermore, if premotor neuronsencoding vergence movements were to drive ABN dischargestogether with premotor neurons encoding conjugate move-ments, as proposed by Hering’s theory, then one would expectABNs with properties ranging from pure conjugate to purevergence sensitivities to be found. However, although we re-corded ABNs that encoded the conjugate movements of theeyes, we never encountered any neuron that solely encodedvergence movements (see Figs. 8 and 11). In light of theseresults, we propose that the saccadic burst neurons and theneurons in the neural integrator provide the binocular informa-tion that is required to generate the discharge patterns that werecorded in the abducens nucleus during disjunctive saccades.
Another striking result was that individual ABNs generallyencoded the position of the same eye(s) during disjunctivesaccades and disjunctive fixation (i.e., exhibited good coher-ence). In the neural integrator, neurons that encode the positionof either eye can be found (McConville et al. 1994; Zhou andKing 1996). Hence, our finding strongly suggests that the eyeposition–related signal on any given ABN originate from thesame premotor neurons in the neural integrator during disjunc-tive saccades and disjunctive fixation. An interesting implica-tion of this proposal is that premotor neurons in the neuralintegrator should encode the motion of the same eye duringdisjunctive saccades and disjunctive fixation. Indeed, prelimi-nary results indicate that prepositus hypoglossi neurons dogenerally encode the position of the same eye(s) during dis-junctive saccades and disjunctive fixation (unpublished obser-vations).
General conclusions
The primary objective of this study was to determinewhether the signals carried by ABNs during disjunctive sac-
3466 P. A. SYLVESTRE AND K. E. CULLEN
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

cades are appropriate to drive the motion of the eye to whichthey project. We found that although individual ABNs canencode the motion of both eyes to various degrees (rangingfrom monocular with a preference for either eye to conjugate),the population drive of ABNs still accounts for most (approx-imately 85%) of the movements of the ipsilateral eye. Wepropose that the selective weighting of the AMN projections tothe lateral rectus fine tunes the motion of the ipsilateral eye bycompensating for the differences in discharge properties thatwere observed among individual neurons. The organized tun-ing of AINs versus AMNs, or the co-contraction of the medialand lateral recti, likely play minor roles during disjunctivesaccades. The second goal of this study was to determinewhether ABNs encode conjugate and vergence signals simi-larly during disjunctive saccades and disjunctive fixation. In-deed, we found that individual ABNs generally encode similareye position-related signals during these two conditions. Thisresult suggests that the premotor structures responsible forthese signals are the same in both conditions.
A P P E N D I X
Classically, saccade-related bursting activity in the brain stem hasbeen characterized using metric-based relationships, like the numberof spikes in a burst (NOS) versus the amplitude of the correspondingeye movement (E). The primary assumption inherent to such a“NOS-based analysis” is that the NOS is proportional to the Eduring a saccade
NOS � E, which implies that �FR � �E, and therefore that FR � E (A1)
However, during saccades, the most simplified description of ABNdischarge dynamics is FR(t) � b � kE(t) � rE(t), and not FR(t) �b � rE(t) (Sylvestre and Cullen 1999a). It is therefore clear that theassumption described in Eq. A1 is false under these conditions (i.e.FR �/ E), which invalidates the use of a NOS-based analysis for ABNsduring saccades.
To further emphasize the potential risks of using a NOS-basedanalysis on ABN discharges, we re-analyzed our dataset of disjunctivesaccades using the following model
NOS � b � nIEIE � nCECE (A2)
where IE and CE are the changes in ipsilateral and contralateraleye positions, respectively. A particularly striking result was obtainedwith this analysis for our example monocular ABN, unit B72_2 (seeFigs. 5–7). It is obvious from the raw data in Fig. 6 (and from thedynamic analysis) that this neuron did not encode the conjugatemotion of the eyes. Nevertheless, the NOS-based analysis yielded theconclusion that nIE � nCE (at 95% confidence), suggesting that unitB72_2 encoded conjugate eye movements. Similar misleading con-clusions were obtained for many other neurons. This example com-plements that of our previous studies (Cullen and Guitton 1997a,b;Cullen et al. 2001) in demonstrating that NOS-based analyses oftenyield misleading results.
We thank J. E. Roy, S. Sadeghi Ghandehari, and E. Cervoni, for commentson the manuscript. We also thank W. Kucharski for the instrumental role in thedevelopment of our experimental setup, E. Moreau for taking great care of ouranimals, F. Rouah for feedback on the statistical analysis, and J. Knowles andA. Smith for technical assistance.
This study was supported by the Canadian Institutes of Health Research(CIHR), the Natural Science and Engineering Research Council of Canada(NSERC), and the Fonds de la Recherche en Sante du Quebec (FRSQ).
REFERENCES
BROUSSARD DA, DECHARMS RC, AND LISBERGER SG. Inputs from the ipsilat-eral and contralateral vestibular apparatus to behaviorally characterizedabducens neurons in rhesus monkey. J Neurophysiol 74: 2445–2459, 1995.
BUTTNER-ENNEVER JA, HORN AKE, SCHERBERGER H, AND D’ASCANIO P.Motoneurons of twitch and nontwitch extraocular muscle fibers in theabducens, trochlear, and oculomotor nuclei of monkeys. J Comp Neurol438: 318–335, 2001.
BUTTNER-ENNEVER JA, HORN AKE, GRAF W, AND UGOLINI G. Modern con-cepts of brainstem anatomy. From extraocular motoneurons to propriocep-tive pathways. Ann NY Acad Sci 956: 75–84, 2002.
CARPENTER J AND BITHELL J. Bootstrap confidence intervals: when, which,what? A practical guide for medical statisticians. Stat Med 19: 1141–1164,2000.
CHATURVEDI V AND VAN GISBERGEN JAM. Perturbation of combined saccade-vergence movements by microstimulation in monkey superior colliculus.J Neurophysiol 81: 2279–2296, 1999.
CHATURVEDI V AND VAN GISBERGEN JAM. Stimulation in the rostral pole ofmonkey superior colliculus: effects on vergence eye movements. Exp BrainRes 132: 72–78, 2000.
COLLEWIJN H, ERKELENS CJ, AND STEINMAN RM. Voluntary binocular gaze-shifts in the plane of regard: dynamics of version and vergence. Vision Res35: 3335–3358, 1995.
COLLEWIJN H, ERKELENS CJ, AND STEINMAN RM. Trajectories of the humanbinocular fixation point during conjugate and non-conjugate gaze-shifts.Vision Res 37: 1049–1069, 1997.
CULLEN KE, GALIANA HL, AND SYLVESTRE PA. Comparing extraocular mo-toneuron discharges during head-restrained saccades and head-unrestrainedgaze shifts. J Neurophysiol 83: 630–637, 2001.
CULLEN KE AND GUITTON D. Analysis of primate IBN spike trains usingsystem identification techniques. I. Relationship to eye movement dynamicsduring head-fixed saccades. J Neurophysiol 78: 3259–3282, 1997a.
CULLEN KE AND GUITTON D. Analysis of primate IBN spike trains usingsystem identification techniques. II. Relationship to gaze, eye, and headmovement dynamics during head-free gaze shifts. J Neurophysiol 78: 3283–3306, 1997b.
CULLEN KE, REY CD, GUITTON D, AND GALIANA HL. The use of systemidentification techniques in the analysis of oculomotor burst neuron spiketrain dynamics. J Comput Neurosci 3: 347–368, 1996.
DELGADO-GARCIA JM, DEL POZO F, AND BAKER R. Behavior of neurons in theabducens nucleus of the alert cat. I. Motoneurons. Neuroscience 17: 929–952, 1986a.
DELGADO-GARCIA JM, DEL POZO F, AND BAKER R. Behavior of neurons in theabducens nucleus of the alert cat. II. Internuclear neurons. Neuroscience 17:953–973, 1986b.
DESTOMBES J, GOGAN P, AND ROUVIERE A. The fine structure of neurons andcellular relationships in the abducens nucleus in the cat. Exp Brain Res 35:249–267, 1979.
ENRIGHT JT. Changes in vergence mediated by saccades. J Physiol (Lond) 350:9–31, 1984.
ERKELENS CJ, STEINMAN RM, AND COLLEWIJN H. Ocular vergence undernatural conditions. II. Gaze shifts between real targets differing in distanceand direction. Proc R Soc Lond 236: 441–465, 1989.
FUCHS AF AND LUSCHEI ES. Firing patterns of abducens neurons of alertmonkeys in relationship to horizontal eye movement. J Neurophysiol 33:382–392, 1970.
FUCHS AF AND ROBINSON DA. A method for measuring horizontal and verticaleye movements in the monkey. J Physiol (Lond) 191: 609–631, 1966.
FUCHS AF, SCUDDER CA, AND KANEKO CRS. Discharge patterns and recruit-ment order of identified motoneurons and internuclear neurons in the mon-key abducens nucleus. J Neurophysiol 60: 1874–1895, 1988.
GAMLIN PDR. Subcortical neural circuits for ocular accommodation and ver-gence in primates. Ophthal Physiol Opt 19: 81–89, 1999.
GAMLIN PDR, GNADT JW, AND MAYS LE. Abducens internuclear neurons carryan inappropriate signal for ocular convergence. J Neurophysiol 62: 70–81,1989.
GAMLIN PDR AND MAYS LE. Dynamic properties of medial rectus motoneu-rons during vergence eye movements. J Neurophysiol 67: 64–74, 1992.
HAYES AV, RICHMOND BJ, AND OPTICAN LM. A UNIX-based multiple processsystem for real-time data acquisition and control. WESCON Conf Proc 2:1–10, 1982.
HELMHOLTZ JA. Treatise on Physiological Optics. New York: Dover, 1910.
3467ABDUCENS NEURONS DURING DISJUNCTIVE SACCADES
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from

HERING E. Lehre vom Binokularen Sehen, edited by Bridgeman B and Stark L.New York: Plenum Press, 1977.
JUDGE SJ AND CUMMING BG. Neurons in monkey midbrain with activity relatedto vergence eye movement and accommodation. J Neurophysiol 55: 915–930, 1986.
JUDGE SJ, RICHMOND BJ, AND CHU FC. Implantation of magnetic search coilsfor measurement of eye position: an improved method. Vision Res 20:535–538, 1980.
KELLER EL. Accommodative vergence in the alert monkey. Motor unit anal-ysis. Vision Res 13: 1565–1575, 1973.
KELLER EL AND ROBINSON DA. Abducens unit behavior in the monkey duringvergence movements. Vision Res 12: 369–382, 1972.
KENYON RV, CIUFFREDA KJ, AND STARK L. Unequal saccades during vergence.Am J Opt Physiol Opt 57: 586–594, 1980.
KING WM AND ZHOU W. New ideas about binocular coordination of eyemovements: is there a chameleon in the primate family tree? New Anat 261:153–161, 2000.
KING WM, ZHOU W, TOMLINSON RD, MCCONVILLE KM, PAGE WK, PAIGE GD,AND MAXWELL JS. Eye position signals in the abducens and oculomotornuclei of monkeys during ocular convergence. J Vestib Res 4: 401–408,1994.
MAXWELL JS AND KING WM. Dynamics and efficacy of saccade-facilitatedvergence eye movements in monkeys. J Neurophysiol 68: 1248–1260, 1992.
MAYS LE. Neural control of vergence eye movements: convergence anddivergence neurons in midbrain. J Neurophysiol 51: 1091–1108, 1984.
MAYS LE. Has Hering been hooked? Nature Med 4: 889–890, 1998.MAYS LE AND GAMLIN PDR. Neuronal circuitry controlling the near response.
Curr Opin Neurobiol 5: 763–768, 1995a.MAYS LE AND GAMLIN PDR. A neural mechanism subserving saccade-ver-
gence interactions. In: Eye Movement Research: Mechanisms, Processesand Applications, edited by Findlay J, Walker R, and Kentridge RW.Amsterdam: Elsevier, 1995b: p.215–223.
MAYS LE AND PORTER JD. Neural control of vergence eye movements: activityof abducens and oculomotor neurons. J Neurophysiol 52: 743–761, 1984.
MAYS LE, PORTER JD, GAMLIN PDR, AND TELLO CA. Neural control ofvergence eye movements: neurons encoding vergence velocity. J Neuro-physiol 56: 1007–1021, 1986.
MCCONVILLE K, TOMLINSON RD, KING WM, PAIGE G, AND NA E-Q. Eyeposition signals in the vestibular nuclei: consequences for models of inte-grator function. J Vest Res 4: 391–400, 1994.
MCCREA RA, STRASSMAN A, AND HIGHSTEIN SM. Morphology and physiologyof abducens motoneurons and internuclear neurons intracellularly injectedwith horseradish peroxidase in alert squirrel monkeys. J Comp Neurol 243:291–308, 1986.
MOSCHOVAKIS AK, SCUDDER CA, AND HIGHSTEIN SM. The microscopic anat-omy and physiology of the mammalian saccadic system. Prog Neurobiol 50:133–254, 1996.
ONO H, NAKAMIZO S, AND STEINBACH MJ. Nonadditivity of vergence andsaccadic eye movement. Vision Res 18: 735–739, 1978.
OOHIRA A. Vergence eye movements facilitated by saccades. Jpn J Ophtalmol37: 400–413, 1993.
PRESS WH, TEUKOLSKY SA, VETTERLING WT, AND FLANNERY BP. NumericalRecipes in C. The Art of Scientific Computing (2nd ed.). New York:Cambridge, 1997, p. 689–693.
RAMAT S, DAS VE, SOMERS T, AND LEIGH RJ. Tests of two hypotheses toaccount for different-sized saccades during disjunctive gaze shifts. ExpBrain Res 129: 500–510, 1999.
RICHMOND BJ, OPTICAN LM, PODELL M, AND SPITZER H. Temporal encoding oftwo-dimensional patterns by single units in primate inferior temporal cortex.I. Response characteristics. J Neurophysiol 57: 132–146, 1987.
ROBINSON DA. The mechanics of human saccadic eye movement. J Physiol(Lond) 174: 245–264, 1964.
ROBINSON DA. Oculomotor unit behavior in the monkey. J Neurophysiol 33:393–404, 1970.
SCHILLER PH. The discharge characteristics of single units in the oculomotorand abducens nuclei of the unanesthetized monkey. Exp Brain Res 10:347–362, 1970.
SCUDDER CA, KANEKO CRS, AND FUCHS AF. The brainstem burst generator forsaccadic eye movements. A modern synthesis. Exp Brain Res 142: 439–462, 2002.
SOKAL RR AND ROHLF FJ. Biometry. The Principles and Practice of Statisticsin Biological Research (3rd ed.). New York: W.H. Freeman and Co., 1995,p. 820–825.
SPENCER RF AND STERLING P. An electron microscopy study of motoneuronesand interneurones in the cat abducens nucleus identified by retrogradeintraaxonal transport of horseradish peroxidase. J Comp Neurol 176: 65–86,1977.
STEIGER H-J AND BUTTNER-ENNEVER JA. Relationship between motoneuronsand internuclear neurons in the abducens nucleus: a double retrograde tracerstudy in the cat. Brain Res 148: 181–188, 1978.
SYLVESTRE PA AND CULLEN KE. Quantitative analysis of abducens neurondischarge dynamics during saccadic and slow eye movements. J Neuro-physiol 82: 2612–2632, 1999a.
SYLVESTRE PA AND CULLEN KE. Monocularly tuned discharge dynamics ofabducens and brainstem inhibitor burst neurons during disjunctive saccades.Soc Neurosci Abstr 25: 1652, 1999b.
SYLVESTRE PA, GALIANA HL, AND CULLEN KE. Conjugate and vergenceoscillations during saccades and gaze shifts: implications for integratedcontrol of binocular motion. J Neurophysiol 87: 257–272, 2002.
ZEE DS, FITZGIBBON EJ, AND OPTICAN LM. Saccade-vergence interactions inhumans. J Neurophysiol 68: 1624–1641, 1992.
ZHANG Y, GAMLIN PDR, AND MAYS LE. Antidromic identification of midbrainnear response cells projecting to the oculomotor nucleus. Exp Brain Res 84:525–528, 1991.
ZHANG Y, MAYS LE, AND GAMLIN PDR. Characteristics of near response cellsprojecting to the oculomotor nucleus. J Neurophysiol 67: 944–960, 1992.
ZHOU W AND KING WM. Ocular selectivity of units in oculomotor pathways.Ann NY Acad Sci 781: 724–728, 1996.
ZHOU W AND KING WM. Premotor commands encode monocular eye move-ments. Nature 393: 692–695, 1998.
3468 P. A. SYLVESTRE AND K. E. CULLEN
J Neurophysiol • VOL 88 • DECEMBER 2002 • www.jn.org
on March 21, 2005
jn.physiology.orgD
ownloaded from














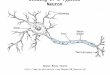














![VH405 - 3bscientific.com · 11 VI. Par craneal [Nervio abducens] 12 VIII. par craneal [Nervio abducens] VII. Par cráneal [Nervio facial] Parte no motora del VII. Par craneal [nervio](https://img.dokumen.tips/doc/110x75/5bb3322209d3f249438e4177/vh405-11-vi-par-craneal-nervio-abducens-12-viii-par-craneal-nervio-abducens.jpg)