Embed Size (px)
Citation preview

GENES, CHROMOSOMES 8 CANCER I1:40-50 (1994)
Detection of Numerical and Structural Chromosome Abnormalities in Pediatric Germ Cell Tumors by Means of In terphase Cytogenetics
Cornelia Stock, lnge M. Ambros, Thomas Lion, Oskar A. Haas, Andreas Zoubek, Helmut Gadner, and Peter F. Ambros
CCRI, Children's Cancer Research Institute, St. Anna Kinderspital (C.S., I.M.A., T.L., O.A.H., P.F.A.), St. Anna Kinderspital (O.A.H. A.Z., H.G.), and Inst. f. Clinical Pathol. (I.M.A.), Vienna, Austria
In contrast t o the cytogenetically well characterized testicular germ cell tumors (GCT) in adults, reports on cytogenetic studies in pediatric GCT are scarce. The presence of an i( I2p) and numerical abnormalities involving chromosome I 2 are the most frequent cytogenetic changes in GCT of adults. We have performed in situ hybridization (ISH) studies on paraffin sections and on isolated nuclei of I3 pediatric GCT with particular emphasis on those chromosome abnormalities that are common in adult GCT. These include numerical and structural abnormalities of chromosomes I and 12 as well as numerical deviations of chromosomes 8, 10, X, and Y. The histological subsets of the tumors investigated included two dysgerminomas (DGE), one seminoma (SE), two embryonal carcinomas (EC), four mixed and two pure yolk sac tumors (YST), and one undifferentiated (IT) and one differentiated teratoma (TD). Similar t o the GCT in adults, additional copies of chromosome 12 were the most frequently observed numerical abnormalities. In contrast t o the findings in adult GCT, changes in the size of the pericentromeric hybridization signals of chromosome 12, suggesting the presence of i( I2p) chromosomes, were found in only two cases. No chromosome abnormalities were found in the pure TD or in the TD cells of mixed tumors containing a YST component. In the YST portion, however, I p deletions and/or numerical chromosome changes were present. Surprisingly, deletions of the short arm of chromosome I, del( l)(p36.3), were frequent in pediatric GCT and were the sole abnormality detected in two cases. The I p36 deletions were present in all stage-lV EC and YST investigated and were absent in the relatively benign TD and in one YST stage4 Therefore, I p36 deletions may have value as a prognostic marker in pediatric GCT. Genes Chromosom Cancer I1:40-50 (1994). 0 1994 Wiley-Liss, Inc.
INTRODUCTION
Cytogenetic analyses of malignant germ cell tu- mors (GCT) have mainly been performed on tes- ticular G C T of adults. T h e most common finding has been an i(12p), seen in approximately 80% of all histologic subsets (Atkin and Baker, 1982, 1983, 1987; Delozier-Blanchet et al., 1985; Gibas et al., 1986; Castedo et al., 1989a-c; Samaniego et al., 1990). T h e high incidence of additional numerical and other structural abnormalities of chromosome 12 underlines the important role of 12p and of genes located in this region, such as the KRAS pro- tooncogene, in the development of G C T (Gibas e t al., 1986; Samaniego et al., 1990; Rodriguez et al., 1992; Suijkerbuijk et al., 1991; Atkin et al., 1993). Moreover, the copy number of i( 12p) seems to be a prognostic marker (Bod et al., 1989). Other fre- quently observed abnormalities in G C T include numerical and/or structural changes of the short or long arm of chromosome 1 (Atkin and Baker 1983, 1987; Castedo e t al., 1989a-c; Samaniego et al., 1990). Furthermore, chromosome 1 abnormalities have been reported in G C T cell lines (Murty et al., 1 990).
Reports on cytogenetic studies of pediatric G C T
are scarce. A few cases of differentiated teratomas ( T D ) (Hecht et al., 1984; Ishiwata et al., 1991; Speleman et al., 1992), undifferentiated teratomas (IT) (Ihara et a]., 1984; Surti et al., 1990), one I T with focal germinoma and embryonal carcinoma (EC) components (Shen et al., 1990), and one yolk sac tumor (YST) (Speleman et al., 1990) were an- alyzed. Two of the reports described an i(12p) (Speleman et al., 1990, 1992) and structural aber- rations of chromosome 1, respectively (Hecht et al., 1984; Shen et al., 1990). It is not clear whether pediatric G C T have a similar pathogenetic path- way as G C T in adults. Despite the markedly im- proved cure rate in gonadal and extragonadal G C T by combination chemotherapy (Hawkins, 1990), reliable prognostic markers could help identify pa- tients in whom postoperative chemotherapy may not be necessary.
Therefore, we performed in situ hybridization studies on paraffin embedded archival material to
Received November 1, 1993; accepted March 2, 1994. Address reprint requests to Cornelia Stock, PhD, CCRI, Chil-
dren's Cancer Research Inst., St. Anna Kinderspital, Kinderspital- gasse 6, A-1090 Vienna, Austria.
0 1994 Wiley-Liss, Inc.

CYTOGENETIC STUDIES IN PEDIATRIC GCT 41
study 1) the copy number of chromosomes 1 and 12, 2) the integrity of the short arm of chromosome 1, and 3) the integrity of the paracentromeric re- gion of chromosome 12. In addition, we investi- gated numerical changes of chromosomes 8 and X which are known to be overrepresented both in seminomatous (SGCT) and nonseminomatous (NSGCT) tumors, as well as numerical changes of chromosomes 10 and Y, both of which are known to be underrepresented in NSGCT of adult patients (Castedo et al., 1989a-c).
The numerical and/or structural chromosome ab- normalities were investigated with a peroxidase- based in situ hybridization (ISH) method which was performed directly on paraffin sections (Hop- man et al., 1991; Stock et al., 1993). For identifi- cation of the i( 12p) chromosome, a fluorescence in situ hybridization (FISH) technique was carried out on interphase nuclei which were isolated from formalin-fixed, paraffin-embedded tissues. Earlier reports had unambiguously demonstrated the pres- ence of an i(12p) in interphase tumor cell nuclei with the probe D12Z3 in cases with a nonpolymor- phic heterochromatin region of the normal chromo- somes 12. However, the diagnosis of a pericentro- meric anomaly on chromosome 12 with this probe is possible only in patients whose hybridization sig- nals on normal chromosomes 12 differ in size from those observed in the tumor cells. Thus, smaller or larger D12Z3 hybridization signals in tumor cells can be regarded as indicators for the presence of an i(12p) (Mukherjee et al., 1991; Rodriguez et al., 1992; own observation in the G C T cell line 27C4).
MATERIALS AND METHODS
We analyzed 26 paraffin-embedded specimens from 13 patients with primary G C T and one spec- imen from a patient with a sex cord/stromal tumor. The patients were treated at the St. Anna Chil- dren’s Hospital and the Mautner-Markhof Chil- dren’s Hospital in Vienna, between 1983 and 1992. T h e tumor samples consisted of different histo- logic types of G C T including two dysgerminomas (DGE), one seminoma (SE) from a patient with Klinefelter syndrome, two embryonal carcinomas (EC), six mixed or pure yolk sac tumors (YST), and one mature ( T D ) and one immature grade 2 ter- atoma ( IT) which mainly consisted of neural tissue (Table 1). Three of these patients (cases 4, 5, and 12) had a tumor recurrence, and two of them (cases 5 and 12) died despite high dose chemotherapy and surgery. All other patients have remained in com- plete remission during the entire observation pe- riod. With the exception of one patient (case 3),
the follow-up period has been over 5 years in all instances.
T h e human G C T cell line 27C4 (kindly pro- vided by MF Pera, CRC, Oxford, UK), displaying a variable number of isochromosomes i( 12p), was analyzed by conventional cytogenetic techniques and served as a control for detecting i(12p) in in- terphase nuclei.
In Situ Hybridization on Paraffin Embedded Sections
ISH on 6 k m tissue sections was used to deter- mine numerical and/or structural chromosome ab- normalities in interphase nuclei. T h e procedure was described previously (Hopman et al., 1991; Stock et al., 1993). Modifications were made only in the pretreatment of the nuclei before ISH to enable the penetration of the labeled probes and antibodies. Usually, application of 1 M sodium thiocyanate (NaSCN) for 5-12 min at 80°C and 0.4% pepsin in 0.2 N HCI (P7012, Sigma, St. Louis, MO) for 10-30 min at 37°C was adequate for T D and E C but not for IT, one DGE, and two YST. In the latter cases, digestion of the tissues with 0.1 or 0.2% proteinase K (P0390, Sigma) for 10-15 min instead of pepsin treatment resulted in distinct hybridization signals.
Hybridization was performed with six biotiny- lated paracentromeric probes specific for chromo- somes 1 (D121, Cooke and Hindley, 1979; kindly provided by Dr. Th. Cremer, Heidelberg, FRG), 8 (D822, Donlon et al., 1987), 10 (DlOZl, Devilee et al., 1988), 12 (D1223, Looijenga et al., 1990), X (DXZ1, Yang et al., 1982), and Y (DYZ3, Wolfe et al., 1985). All probes, with the exception of DlZ1, were obtained from the ATCC. T o analyze the integrity of the short arm of chromosome 1, the DNA probe D1Z2 (Buroker et al., 1987), specific for the subtelomeric region of l p (1~36 .3 ) (van Roy et al., 1993), was used. An identical VNTR probe DlS32 (described by Giannakudis et al., 1987; a gift from Giannakudis, Marburg, FRG), mapping to the same locus (Strehl and Ambros, 1993; van Roy et al., 1993), was applied to verify whether this probe reveals the same pattern of enlarged hy- bridization signals in the ciliated cells as seen with the probe DlZ2.
In each patient, the paraffin sections were also used to determine the size of the D12Z3 hybrid- ization signals in normal nuclei. For this purpose, fibrovascular septa, lymphocytes infiltrating the tu- mor tissue, or normal tissues surrounding the tu- mor (e.g., smooth muscle or adipose tissue) were analyzed.
42 STOCK ET AL.
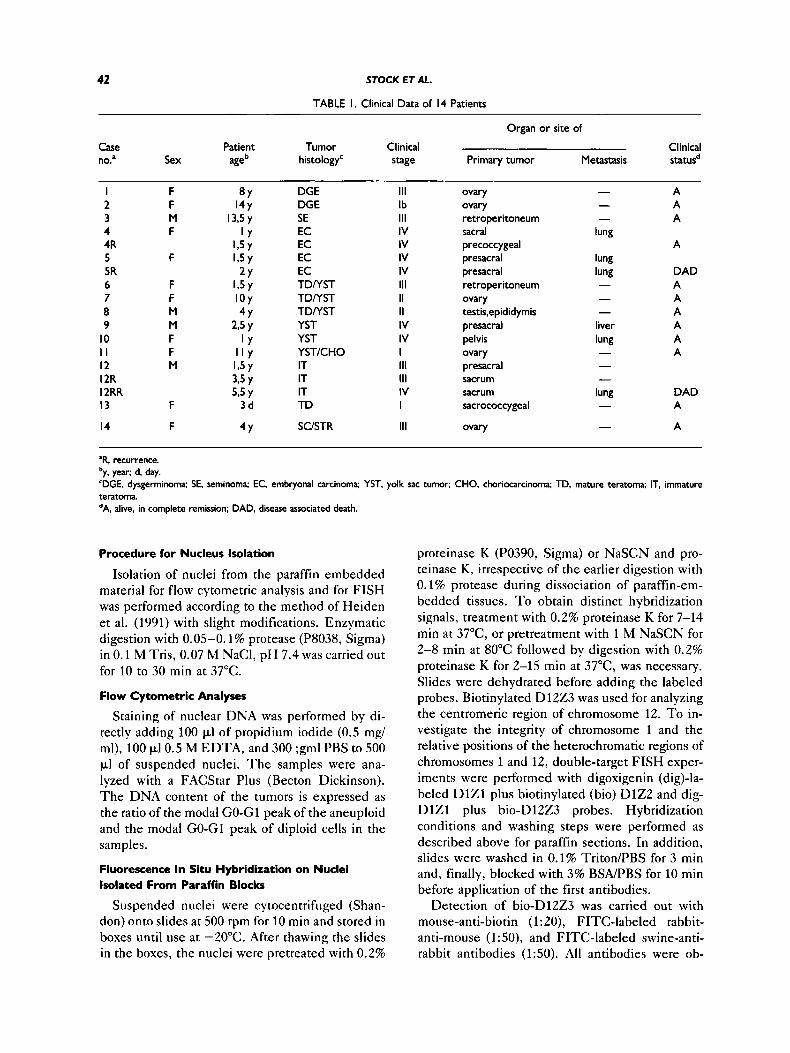
TABLE I. Clinical Data of I 4 Patients
Organ or site of
Case Patient Tumor Clinical Clinical no.' Sex age histology' stage Primary tumor Metastasis statusd
I 2 3 4 4R 5 5R 6 7 8 9
10 I I I 2 12R I 2RR 13
14
F F M F
F
F F M M F F M
F
F
8Y 14Y
13.5 y l Y
1,5y 1,5y
2Y l,5 y I O Y 4Y
2,5 y l Y
I l Y 1,5y 3.5 y 5,5 y
4Y
3 d
DGE DGE SE EC EC EC EC TDNST TDNST TDNST YST YST YSTICHO IT IT IT TD
SClSTR
111 Ib 111 IV IV IV IV 111 II II IV IV I 111 111 IV I
111
ovary ovary retroperitoneum sacral precoccygeal presacral presacral retroperitoneum ovary testis.epididymis presacral pelvis ovary presacral sacrum sacrum sacrococcygeal
ovary
- - -
lung
lung lung - - -
liver lung - - -
lung - -
A A A
A
DAD A A A A A A
DAD A
A
"R, recurrence. 'y, year; d, day. 'DGE, dysgerminoma; SE, seminoma; EC, embryonal carcinoma; YST, yolk sac tumor; CHO, choriocarcinoma; TD, mature teratoma; IT, immature teratoma. dA. alive. in complete remission; DAD, disease associated death.
Procedure for Nucleus Isolation
Isolation of nuclei from the paraffin embedded material for flow cytometric analysis and for FISH was performed according to the method of Heiden et al. (1991) with slight modifications. Enzymatic digestion with 0.05-0.1% protease (P8038, Sigma) in 0.1 M Tris, 0.07 M NaCI, pH 7.4 was carried out for 10 to 30 rnin at 37°C.
Flow Cytometric Analyses
Staining of nuclear DNA was performed by di- rectly adding 100 pl of propidium iodide (0.5 mg/ ml), 100 pI 0.5 M EDTA, and 300 ;gml PBS to 500 pl of suspended nuclei. T h e samples were ana- lyzed with a FACStar Plus (Becton Dickinson). T h e DNA content of the tumors is expressed as the ratio of the modal GO-G1 peak of the aneuploid and the modal GO-G1 peak of diploid cells in the samples.
Fluorescence In Situ Hybridization on Nuclei Isolated From Paraffin Blocks
Suspended nuclei were cytocentrifuged (Shan- don) onto slides at 500 rpm for 10 rnin and stored in boxes until use at -20°C. After thawing the slides in the boxes, the nuclei were pretreated with 0.2%
proteinase K (P0390, Sigma) or NaSCN and pro- teinase K, irrespective of the earlier digestion with 0.1% protease during dissociation of paraffin-em- bedded tissues. T o obtain distinct hybridization signals, treatment with 0.2% proteinase K for 7-14 min at 37"C, or pretreatment with 1 M NaSCN for 2-8 rnin at 80°C followed by digestion with 0.2% proteinase K for 2-15 min at 37"C, was necessary. Slides were dehydrated before adding the labeled probes. Biotinylated D12Z3 was used for analyzing the centromeric region of chromosome 12. T o in- vestigate the integrity of chromosome 1 and the relative positions of the heterochromatic regions of chromosomes 1 and 12, double-target FISH exper- iments were performed with digoxigenin (dig)-la- beled D l Z l plus biotinylated (bio) D1Z2 and dig- D l Z l plus bio-D1223 probes. Hybridization conditions and washing steps were performed as described above for paraffin sections. In addition, slides were washed in 0.1% Triton/PBS for 3 min and, finally, blocked with 3% BSNPBS for 10 min before application of the first antibodies.
Detection of bio-D1223 was carried out with mouse-anti-biotin (1:20), FITC-labeled rabbit- anti-mouse (1 :50), and FITC-labeled swine-anti- rabbit antibodies (1:SO). All antibodies were ob-

CYTOGENETIC STUDIES IN PEDIATRIC GCT 43
tained from Dakopatts, Denmark, and were diluted in 3% BSNPBS. In the double-target FISH experiments, immunocytochemical detection of the bio- and dig-labeled probes was performed si- multaneously by successive incubation with a mouse anti-bio antibody (1:20, Dakopatts) plus FITC-conjugated sheep-anti-dig antibody (1:80, Boehringer-Mannheim) for 45-60 min at 37°C. T h e second step antibody consisted of TRITC- conjugated rabbit F(ab'),-anti-mouse (1:20, Dako- patts) plus FITC-conjugated rabbit-anti-sheep (1: 80, Dakopatts) antibodies which were allowed to bind for 45-60 min at 37°C. After each step of antibody binding, washing steps were carried out in 0.1% BSAPBS for 10 min at 42°C each. Coun- terstaining was performed with 4'6-diamidino-2- phenylindole (DAPI). A Leitz fluorescence micro- scope equipped with a double bandpass filter (Omega Optical, Inc., U.S.A.) was used for simul- taneous visualization of the F ITC and T R I T C sig- nals.
FISH on Metaphase Spreads and lnterphase Nudei of the Germ Cell Tumor Line 27C4
Metaphase cells were prepared according to standard protocols. We used bio-DlZZ3 for evalu- ating numerical and structural anomalies of chro- mosome 12 in metaphase and interphase cells. In the bicolor double FISH experiment with dig- D t Z l and bio-DlZ2, the integrity of chromosome 1 was investigated. T h e hybridization conditions were essentially as reported previously (Stock et al., 1993). We used two different labeled probes, and detection of the hybridized probes was per- formed as described above.
Evaluation of ISH Data
T h e evaluation procedure for the number of hy- bridization signals on paraffin-embedded sections was reported previously (Hopman et al., 1991; Stock et al., 1993). Table 2 summarizes the esti- mated copy number of the chromosomes analyzed, which we deduced from the percentage of nuclei displaying a certain number of hybridization signals with different probes. ISH with D l Z l on 6 pm normal tissue sections and tumor tissues revealed 50-65% of 8-10 pm sized nuclei of normal cells displaying two hybridization signals, 40-50% sim- ilar-sized nuclei of a trisomic tumor tissue exhibit- ing three signals, and only 25-30% nuclei of a tet- rasomic tissue showing four hybridization spots. T h e low frequency of nuclei with the maximum number of hybridization signals was due to the presence of truncated nuclei. In larger nuclei, the
average number of hybridization signals per nu- cleus was even lower (e.g., in 18 pm sized tetra- somic nuclei, only 12% exhibited four hybridiza- tion spots). These data were derived from experiments performed on sections of paraffin-em- bedded neuroblastoma tissues. Since the ISH re- sults correlated well with those obtained by con- ventional cytogenetics (Stock et al., 1993), we extended these experiments to the analysis of pe- diatric GCT.
T h e problem of truncation could be overcome by isolating the nuclei from paraffin blocks. Nuclei isolated from a normal lymph node displayed two hybridization signals in 85.0% (k3.6) and one hy- bridization spot in 12.6% (k3.1). These results are in accordance with those reported by others (e.g., Wolman et al., 1993). Hence, for detection of rnonosomic clones, we defined 22% as the cutoff value (mean + 3 x SD: 12.6 + 3 x 3.1). In accordance with this definition, a minimum of 22% of the nuclei had to display different-sized hybrid- ization signals in order to be considered positive for i( 12p) or a der(l2).
RESULTS
FISH on Metaphase Cells and lnterphase Nuclei of the GCT Cell Line 27C4
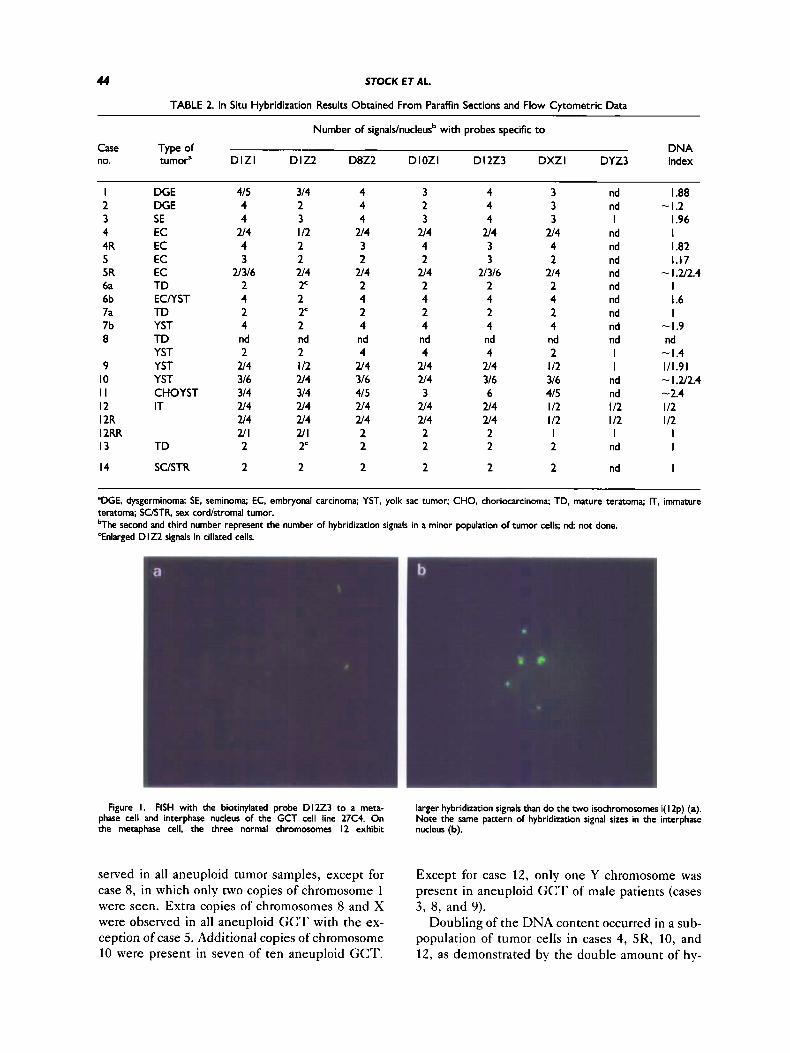
Conventional cytogenetic analysis revealed the presence of cells with either three copies of normal chromosomes 12 and one, two, or three copies of i(12p), or two normal chromosomes 12 and one or two copies of i( 12p) with an additional marker chro- mosome. T h e D l 2 2 3 hybridization signals of i( 12p) were distinctly smaller than those of normal chromosomes 12 (Fig. la), T h e same pattern of signals was unambiguously detected in interphase nuclei (Fig. lb).
In the double-target experiments using dig- D l Z l and bio-D1Z2 as probes, mostly one normal and one rearranged chromosome 1 were observed. T h e short arm of one homolog was translocated to marker chromosomes (data not shown).
ISH Results on Paraffin Sections
T h e ISH results on paraffin sections and the DNA indices are summarized in Table 2 . T h e only histologic subtype of GCT without numerical or structural chromosome abnormalities was the T D , either pure (case 13) or mixed with a YST compo- nent (cases 6a and 7a). All other subtypes of G C T showed numerical and/or structural chromosome abnormalities with the probes used. An increased copy number of chromosomes 1 and 12 was ob-
44 STOCK ET AL.
TABLE 2. In Situ Hybridization Results Obtained From Paraffin Sections and Flow Cytometric Data
Number of signals/nucleusb with probes specific t o
Case Type of DNA no. tumoP D l Z l D IZ2 D8Z2 DlOZl D12Z3 DXZl DYZ3 index
I 2 3 4 4R 5 5R 6a 6b 7a 7b 8
9 10 I I I 2 12R I2RR 13
14
DGE DGE SE EC EC EC EC TD ECMST TD YST TD YST YST YST CHOYST IT
TD
SUSTR
415 4 4
214 4 3
2l316 2 4 2 4 nd 2
214 316 314 214 2 4 2l1 2
2
314 2 3 I12 2 2 214 2c 2 2c 2 nd 2 I12 214 314 2/4 214 2l1 2'
2
4 4 4
214 3 2
2f4 2 4 2 4 nd 4
2f4 316 415 214 2t4 2 2
2
3 2 3
214 4 2
2l4 2 4 2 4 nd 4
2/4 214 3
214 214 2 2
2
4 4 4
2l4 3 3
2l316 2 4 2 4 nd 4
2 4 316 6
214 2/4 2 2
2
3 3 3
2f4 4 2
214 2 4 2 4 nd 2 I12 316 415 I12 I12 I 2
2
nd nd I
nd nd nd nd nd nd nd nd nd I I
nd nd I12 I 12 I
nd
nd
I .88 -1.2
I .96 I I .82 1.17 - I Z2.4 I I .6 I
-1.9 nd - I .4 111.91 - I X2.4 -2.4 I12 112
I I
I ~ ~~
'DGE. dysgerminoma: SE, seminoma: EC, embryonal carcinoma; YST, yolk sac tumor; CHO, choriocarcinoma; TD, mature teratoma; IT, immature teratoma; SC/STR, sex cord/stromal tumor. bThe second and third number represent the number of hybridization signals in a minor population of tumor cells; nd not done. 'Enlarged D I22 signals in ciliated cells.
Figure I. FISH with the biotinylated probe D I223 to a meta- phase cell and interphase nucleus of the GCT cell line 27C4. On the metaphase cell, the three normal chromosomes I2 exhibit nucleus (b).
larger hybridization signals than do the two isochromomnes i( I2p) (a). Note the same pattern of hybridization signal sizes in the interphase
served in all aneuploid tumor samples, except for case 8, in which only two copies of chromosome 1 were seen. Extra copies of chromosomes 8 and X were observed in all aneuploid G C T with the ex- ception of case 5. Additional copies of chromosome 10 were present in seven of ten aneuploid GCT.
Except for case 12, only one Y chromosome was present in aneuploid G C T of male patients (cases 3, 8, and 9).
Doubling of the DNA content occurred in a sub- population of tumor cells in cases 4, 5R, 10, and 12, as demonstrated by the double amount of hy-

CYTOGENETIC STUDIES IN PEDIATRIC GCT 45
bridization signals with all DNA probes used. In case 9, however, a single hybridization signal with the DYZ3 probe showed the presence of only one Y chromosome, although the double amount of hy- bridization spots with all other probes indicated a doubling of the DNA.
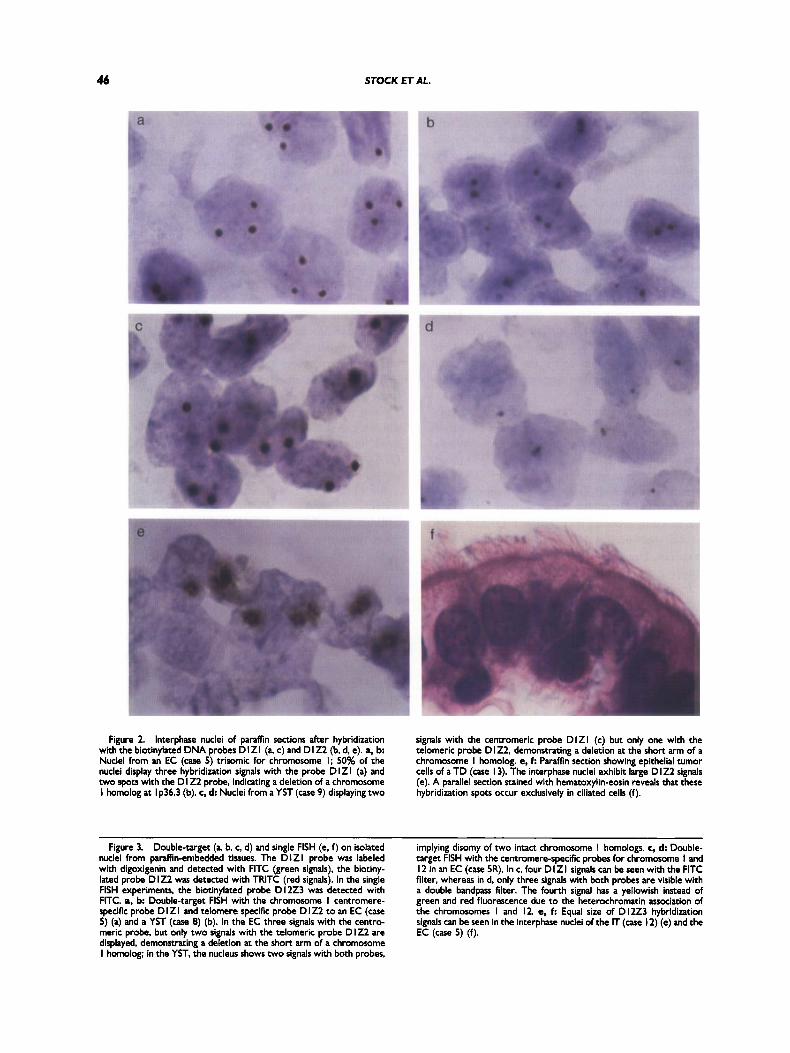
With the probe DlZ2, we detected deletions at 1p36.3 in 11 tumor samples from nine patients. For example, 50% of the nuclei of a primary tumor (case 5) displayed three hybridization signals with the centromere specific probe DlZ1. On parallel section, however, only 6% of the cells showed three D1Z2 hybridization signals and, on an aver- age, 60% of the tumor cells exhibited two D1Z2 hybridization spots, thus demonstrating the pres- ence of a deleted chromosome 1 (Fig 2a,b). In cases 4 and 9 (EC and YST), the 1p36.3 deletion was the only abnormality detected with the probes used (Fig. 2c,d).
In general, only minor size differences between T h e DlZ2 hybridization signals were seen within
both normal and tumor tissues in different individ- uals (Fig. 2b,d). However, in the three T D (cases 6a, 7a, and 13), significantly larger hybridization signals were present in nuclei of certain epithelial cells (Fig. 2e). Parallel sections stained with hema- toxilin-eosin revealed that the large D1Z2 hybrid- ization spots occurred exclusively in ciliated cells (Fig. 2f). T h e same phenomenon was observed by applying the VNTR probe DlS32.
In the sex cord/stromal tumor, no abnormalities were detected with the probes employed.
DNA Flow Cytometry (FCM)
T h e DNA indices of the G C T ranged from 1.0 to 2.44 (Table 2). In contrast to the TD, which were all diploid, no correlation was found between specific histologic types and the DNA indices. T h e small clone with additional chromosomes observed in case 4 was not detected with FCM. Further- more, FCM analysis of the tumor 5R could not distinguish between the diploid and near-diploid cell populations.
FISH on Isolated Nuclei
T h e FISH results on isolated nuclei are pre- sented in Table 3. With respect to lp36 deletions, an excellent correlation with the results obtained by ISH on paraffin sections was seen (Fig. 3a,b).
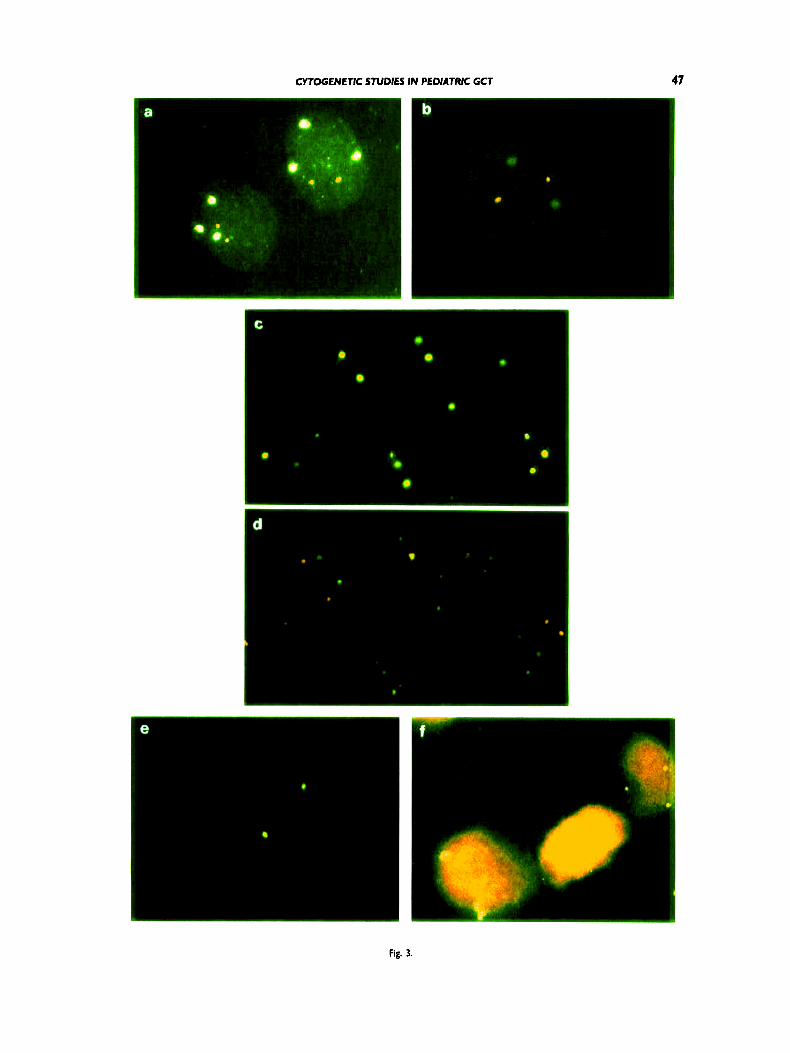
T h e double-target FISH with the probes D l Z l and D12Z3 unambiguously revealed four cell pop- ulations in the recurrent tumor 5R: a predominant population with two copies of chromosomes 1 and 12, two minor populations with three chromosomes
1 and two to three chromosomes 12, and, finally, a small population with four copies of chromosomes 1 and 12. Because of overlapping nuclei, the latter population could not be characterized on paraffin sections. On the other hand, tumor cells with hex- asomy for chromosomes 1 and 12 were only rarely observed in this case. An interesting observation in tumor SR was the association of at least one D l Z l and one D12Z3 signal in 17-37% of the nuclei. Occasionally, this association resulted in a yellow- ish fluorescence instead of green (FITC) or red (TRITC), respectively (Fig. 3d). Heterochromatic associations between chromosomes 1 and 12 were seen in cases 3 and 12 in approximately 20% of the nuclei, but in most instances the F ITC and T R I T C signals were distinct. In all other GCT, this phenomenon was seen in a maximum of 12% (range: 4-12) of the cases. T h e lowest frequency of these associations was observed in the TD (case 13) and the YST (case 10). To determine the sig- nificance of this observation, we analyzed lympho- cytes from normal individuals. Associations of the D l Z l and D l 2 2 3 hybridization signals were ob- served in 10 to 20% of normal nuclei.
An i(12p) or a der(l2) was noted only in two instances (cases 6 and 11). In all other tumors, no additional smaller or larger D12Z3 hybridization spots indicative of an i(12p) were present (Fig. 3e,f).
DISCUSSION
T h e aim of this study was to investigate the copy number of various chromosomes in pediatric G C T and to evaluate particular structural abnormalities of chromosomes 1 and 12. In the tumors analyzed, only chromosome 12 was numerically overrepre- sented (t-test, P < 0.05). Gains of chromosomes 8, X, and 1, in decreasing order, were observed in a number of tumor cells, whereas numeric abnormal- ities involving chromosomes 10 and Y were under- represented in the aneuploid GCT. However, dif- ferences in size between the Dl223 hybridization signals, which are considered indicators of abnor- mal centromeric regions of chromosome 12, in par- ticular of i(12p), were observed in only two cases. Similar to the germ cell tumor line 27C4, the hy- bridization spots in both tumors were smaller than those of the normal chromosomes 12. Whether these signals represented an i( 12p) or another chro- mosome 12 abnormality cannot be detected with the methods used.
Structural abnormalities of the short and long arms of chromosome 1 have been described fre- quently in cytogenetic studies of malignant G C T
46 STOCK ET AL.
Figure 2. Interphase nuclei of paraffin sections after hybridization with the biotinylated DNA probes D I Z I (a c) and D I Z2 (b, d, e). a, b Nuclei from an EC (case 5) trisomic for chromosome I; 50% of the nuclei display three hybridization signals with the probe D l Z l (a) and two spots with the D I Z2 probe, indicating a deletion of a chromosome I homolog at I p36.3 (b). c, d Nuclei from a YST (case 9) displaying two
signals with the centromeric probe D l Z l (c) but only one with the telomeric probe D 122. demonstrating a deletion a t the short arm of a chromosome I homolog. e, f: Paraffh section showing epithelial tumor cells of a TD (case I 3). The interphase nuclei exhibit large D I22 signals (e). A parallel section stained with hematoxylin-eosin reveals that these hybridization spots occur exclusively in ciliated cells (f).
Figure 3. Double-target (6 b, c, d) and single FISH (e, f) on isolated nuclei from paraffin-embedded tissues. The D l Z l probe was labeled with digoxigenin and detected with FlTC (green signals), the biotiny- lated probe DIZ2 was detected with TRlTC (red signals). In the single FISH experiments, the biotinylated probe D I2Z3 was detected with FITC. a, b Double-target FISH with the chromosome I centromere- specific probe D I Z I and telomere specific probe D 1Z2 to an EC (case 5) (a) and a YST (case 8) (b). In the EC three signals with the centro- meric probe, but only two signals with the telomeric probe D I22 are displayed, demonstrating a deletion at the short arm of a chromosome I homolog; in the YST, the nucleus shows two signals with both probes,
implying disomy of two intact chromosome I homologs. c, d: Double- target FISH with the centromere-specific probes for chromosome I and I2 in an EC (case SR). In c, four DlZl signals can be seen with the FlTC filter, whereas in d. only three signals with both probes are visible with a double bandpass filter. The fourth signal has a yellowish instead of green and red fluorescence due to the heterochromatin association of the chromosomes I and 12. e, f: Equal size of DIZZ3 hybridization signals can be seen in the interphase nuclei of the IT (case 12) (e) and the EC (case 5) (f).
CYlOGENETlC STUDIES IN PEDIATRIC GCT 47
Fig. 3.

48 STOCK ET AL.
TABLE 3. Results Obtained by Fluorescence In Situ Hybridization on Nuclei Isolated From
Paraffin-Embedded Tissue
N b of FISH-signals per nucleus'
Case no. DlZl D I Z 2 D12Z3 of different size
N b of D I223 signals
I 415 314 2 nd nd 3 514 413 4 u 4 I I2 4R 4 2 5 3 2 5R 21314 2/24 6a 2 2 6b 2/4 1 2 7a 2 2 7b 4 2 0 2 2 9 2 4 I I2
10 316 1 4 I I 314 314 I2 u 4 1 4 12R U 4 u 4 12RR 2/1 U I 13 2 2
14 2 2
The second and third numbers represent a subpopulation of tumor cells: nd: not done.
in adults (Atkin and Baker, 1983; Gibas et al., 1986; Oosterhuis et al., 1986; Castedo et al., 1988, 1989a-c). Castedo and co-workers even postulated that chromosome 1 is much more frequently in- volved in testicular G C T than in any other tumor. Their findings did not reveal an association be- tween structural aberrations of chromosome 1 and advanced tumor stages. In our study, however, all stage IV tumors, except for the IT, showed a de- letion at the short arm of chromosome 1. Although lp36 deletions were not seen in the T D (cases 6a, 7a, and 13), I T (case 12), YST (case 8), and CH/ YST (case l l ) , it is of interest to note that in the YST of a male patient (case 8), only two copies of chromosome 1 but four copies of chromosomes 8, 10, and 12 and two copies of chromosome X were present. In contrast, the mixed YST (case l l ) , which lacked a 1p36.3 deletion, was the only ex- tragonadal YST stage I in our study. Thus, the high frequency of 1p36.3 deletions observed in ad- vanced tumor stages suggests that this abnormality could play a role in the progression of pediatric GCT. In two tumors (cases 4 and 9), the l p dele- tions were the only abnormality detected with the probes used, except for a small population which
became polyploid after loss of 1~36.3. In the few reports of pediatric G C T available, numerical and/or structural chromosome 1 aberrations have also been described (Hecht et al., 1984; Shen et al., 1990). In one of them, the del (l)(p36) and structural abnormalities of l q were sole aberrations (Hecht et al., 1984). Hence, one may consider chromosome 1 aberrations to be involved in the pathogenesis of pediatric GCT.
In our study, the pure T D or the T D component in mixed tumors was the only subtype of G C T without numerical or structural chromosome changes. I t is not clear whether mixed tumors com- posed of a T D portion which lacks cytogenetic ab- normalities and of a YST portion which contains numerical and/or structural abnormalities evolve from a common or from two different cell clones.
Intact short arms of chromosome 1 are not only present in highly differentiated cells of TD. The same observation was made also in the highly dif- ferentiated ganglionic cells of maturing neuroblas- tomas (Ambros et al., 1994), despite the fact that lp36 deletions are very common in undifferenti- ated, aggressive neuroblastomas. Another study demonstrated the frequent loss of heterozygosity (LOH) at lq21-q24 in well-differentiated advanced gastric cancers and, in contrast, LOH at lp33-p35 in poorly differentiated gastric cancers (Sano et al., 1991). Based on these data one may hypothesize that genes located in the telomeric region of chro- mosome 1 are in some way involved in terminal cell differentiation.
Characteristic findings in the T D included the presence of two intact chromosomes 1 and, in a subset of these tumors, grossly enlarged D1Z2 or DlS32 hybridization signals. More importantly, the prominent D1Z2 or DlS32 signals were re- stricted to ciliated cells in these highly differenti- ated tumors. Possible explanations for the enlarged signals include the occurrence of associations be- tween the hybridization spots, a decondensation of this particular chromosome region, and the pres- ence of RNA tracks or amplified target sequences. However, RNAse treatment before hybridization did not alter this pattern and the associated signals of these or other probes never achieved a compa- rably large size. We are currently studying this puz- zling phenomenon.
Two of the patients in this study had tumors refractory to chemotherapy (cases 5 and 12). In cases 12 and 12R, the majority of the tumor cells were diploid and only a minor subpopulation was tetraploid. The last recurrent tumor in this case (12RR) consisted of diploid cells only, the tetra-

CMOGENETIC STUDIES IN PEDIATRIC GCT 49
ploid tumor cell population had disappeared as as- sessed by FACS analysis and ISH. T h e same phe- nomenon was observed in case 5 , as the recurrent tumor 5R was composed mainly of diploid and near-diploid cell populations as assessed by ISH analyses. T h e primary tumor in case 4 also con- sisted of a predominantly diploid and a small tet- raploid cell population. In contrast to the cases 5 and 12, the recurrent tumor 4R contained exclu- sively near-tetraploid cells, which responded well to chemotherapy. Moreover, the majority of the other G C T with a good response to chemotherapy, with the exception of T D , were near-triploid or tetraploid. A similar observation has been made in other childhood malignancies, e.g., in neuroblas- toma and acute lymphoblastic leukemia, where diploidy is often associated with an adverse prog- nosis (Hayashi et al., 1987; Pui et al., 1990).
Somatic pairing of the centromeric regions of the two homologous chromosomes 1 and 17 has been reported in normal human brain tissue (Arnoldus et al., 1989, 1991). In our study, centromeric associ- ations of chromosomes 1 and 12 were observed in a high proportion of tumor cells in the case 5R. Ad- ditional analyses, however, revealed that this phe- nomenon occurred also in normal cells. Although the significance of centromeric associations is un- known, it is intriguing that the heterochromatic associations occurred between the two chromo- somes most commonly involved in structural ab- normalities of adult GCT. Moreover, it is interest- ing to note that the performance of double-target FISH under low stringency revealed DNA se- quences in the centromeric region of chromosome 12 related to those of l q l l .
The correlation between the ISH on paraffin sections and the FISH results on isolated nuclei demonstrates the validity of both methods. How- ever, each method offers certain advantages. T h e i( 12p) should be assessed preferentially on intact nuclei, since, compared with signals on a normal chromosome 12, this abnormality can result in smaller hybridization signals. Furthermore, over- lapping nuclei cannot be analyzed on sections. T h e occurrence of mixed tumor cell populations with either three or four chromosomes 1 can cause prob- lems in the evaluation of l p deletions in paraffin sections. On the other hand, a disadvantage of the FISH method on isolated nuclei is the inability to distinguish normal cells from disomic tumor cells of equal size. This prevents the assessment of the precise percentage of monosomic cells within a predominantly diploid tumor. Hence, the presence of a monosomic tumor cell population defined by a
cutoff value of 222% tumor cells could not be de- termined by this technique. Most importantly, however, the histological context is completely lost when isolated nuclei are analyzed. For example, in patient 8, whose tumor cells were separated by fibrovascular septa, ISH analyses of tumor sections unambiguously revealed the occurrence of a 1p36.3 deletion exclusively in the YST cells. Further- more, the D1223 polymorphisms in small-sized nuclei of normal cells, or the grossly enlarged D122 signals which are restricted to ciliated cells, are easily detectable with the ISH technique on paraffin sections. These facts emphasize the ne- cessity of using both methods in order to obtain optimal information.
ACKNOWLEDGMENTS
We thank Andrea Zellner and Silvia Bauer for their excellent technical assistance. This work was supported by the “Osterreichische Kinderkrebs- hilfe.”
REFERENCES Ambros IM, Zellner A, Stock C , Amann G . Gadner H. Ambros PF
(1994) Proof of the reactive nature of the Schwann cell in neuro- blastoma and its clinical implications. In Evans AE, DAngio GJ, Knudson AG, Seeger RC (eds): Advances in Neuroblastoma, Re- search 4. New York: Wiley-Liss, Inc.
Arnoldus EPJ, Peters ACB, Bots GTAM, Raap AK, Van der Ploeg M (1989) Somatic pairing of chromosome 1 centromeres in inter- phase nuclei of human cerebellum. Hum Genet 83231-234.
Arnoldus EPJ, Noordermeer IA, Peten ACB, Raap AK, Van der Ploeg M ( 1 9 9 1 ) Interphase cytogenetics reveals somatic pairing of chromosome 17 cenrromeres in normal human brain tissue, bur no trisomy 7 or sex-chromosome loss. Cytogenet Cell Genet 56:214- 216.
Atkin NB, Baker MC (1982) Specific chromosome change, i(12p), in testicular rumours? Lancet 2:1349.
Atkin NB, Baker MC (1983) i(12p): Specific chromosomal marker in seminoma and malignant reratoma of the testis? Cancer Genet Cytogenet 10: 199-204.
Atkin NB, Baker MC (1987) Abnormal chromosomes including small metacentrics in 14 ovarian cancers. Cancer Genet Cvroee- , - net 26:355-361.
Atkin NB, Fox MF, Baker MC, Jackson Z (1993) Chromosome 12-containing markers, including 2 dicentrics, in three i(12p)- negative testicular germ cell rumors. Genes Chrom Cancer 6218-221.
Bosl GJ, Dmitrovskv E, Reuter VE, Samaniego F, Rodriguez E, Geller NL. Chaganti RSK (1989) lsochromosome of chromosome 12: Clinically useful marker for male germ cell tumors. J Natl Cancer lnst 81: 1874-1878.
Buroker N, Beswick R, Haight G, Magenis RE, Lirt M (1987) A hypervariable repeated sequence on human chromosome 1 p3h. Hum Genet 77:175-181.
Castedo SMMJ, de Jong B, Oosterhuis JW, Seruca R, Idenburg \’.I. Buist J, Sleijfer D T (1988) i(lZp)-Negarive testicular germ cell tumors. A different group? Cancer Genet Cytogenet 35: 17 1-1 78.
Castedo SMMJ, de Jong B, Oosterhuis JW, Seruca R, re Meerman GJ, p m A, Koops HS (198%) Cytogenetic analysis of ten human seminomas. Cancer Res 49:439-443.
Castedo SMMJ, de Jong B, Oosterhuis JW, Idenburg VJS, Seruca R, Buist J, te Meerman GJ, Koops HS, Sleijfer D T (l989b) Chro- mosomal changes in mature residual teratomas following poly- chemotherapy. Cancer Res 49:672-676.
Castedo SMMJ, de Jong B. Oosterhuis JW, Seruca R, Idenburg VJS, Dam A, re Meerman G, Koops HS, Sleijfer IIT (1989~) Chromosomal changes in human primary testicular nonsemi- nomatous germ cell tumors. Cancer Re5 49:5696-5701.

50 STOCK ET AL.
Cooke H, Hindley J (1979) Cloning of human satellite I11 DNA: Different components of different chromosomes. Nucleic Acid Res 6:3177-3197.
Delozier-Blanchet CD, Engel E, Walt H (1985) Isochrome 12p in malignant testicular tumors. Cancer Genet Cytogenet 15375- 376.
Devilee P, Kievits T, Waye JS, Pearson PL, Willard H F (1988) Chromosome-specific alpha satellite DNA: Isolation and mapping of a polymorphic alphoid repeat from human chromosome 10. Genomics 3: 1-7.
Donlon TA, Burns GA, Latt SA, Mulholland J, Wyman AR (1987) A chromosome 8-enriched alphoid repeat. Cytogenet Cell Genet 46:607.
Giannakudis J, Vortkamp A, Kabisch R, Schmitt-Linger E, Grze- schik KH (1987) A hypervariable region located near Ipter. Cy- togenet Cell Genet 46:619-620.
Gibas 2, Prout GR, Pontes JE, Sandberg AA (1986) Chromosome changes in germ cell tumors of the testis. Cancer Genet Cytoge- net 19:245-252.
Hawkins EP (1990) Pathology of germ cell tumors in children. On- col Hematol 10165-177.
Hayashi Y, Hanada R, Yamamoto K, Bessho F (1987) Chromosome findings and prognosis in neuroblastoma. Cancer Genet Cytoge- net 29:175-177.
Hecht F , Grix A Jr, Hecht BK, Berger C, Bixenman H, Szucs S, OKeeffe D, Finberg HJ (1984) Direct prenatal chromosome di- agnosis of a malignancy. Cancer Genet Cytogenet l l : 107-1 l l .
Heiden T , Wang N, Tribukait B (1991) An improved Hedley method for preparation of paraffin-embedded tissues for flow cy- tometric analysis of ploidy and S-phase. Cytometry 12614-621.
Hopman AHN, Van Hooren E, Van de Kaa CA, Vooijs GP, Ra- maekers FCS (1991) Detection of numerical chromosome aberra- tions using in situ hybridization in paraffin sections of routinely processed bladder cancers. Modern Pathol 4:503-513.
Ihara T , Ohama K, Satoh H, Fujii T , Nomura K, Fujiwara A (1984) Histologic grade and karyotype of immature teratoma of the ovary. Cancer 542988-2994.
Ishiwata I, Ishiwata C, Soma M, Ono I. Nakaguchi T, Joh K, Fu- rusato M, Ishikawa H (1991) N-my gene amplification and neuron specific enolase production in immature teratomas. Virchows Ar- chiv A Pathol Anat 418:333-338.
Looijenga LHJ, Smit VTHBM, Wessels JW, Mollevanger P, Oost- erhuis JW, Cornelisse CJ, Devilee P (1990) Localization and poly- morphism of a chromosome 12-specific a satellite DNA se- quence. Cytogenet Cell Genet 53916-218.
Mukherjee AB, Murty VWS, Rodriguez E, Reuter VE, Bosl GJ, Chaganti RSK (1991) Detection and analysis of i(lZp), a diagnos- tic marker of human male germ cell tumors, by fluorescence in situ hybridization. Genes Chromosom Cancer 3:300-307.
Murty VWS, Dmitrovsky E, Bosl G, Chaganti RSK (1990) Non- random chromosome abnormalities in testicular and ovarian germ cell tumor cell lines. Cancer Genet Cytogenet 5067-73.
Oosterhuis JW, de Jong B, Cornelisse CJ, Molenaar IM, Meiring A, Idenburg V, Schraffordt Koops H, Sleijfer D T (1986) Karyotyping and DNA flow cytometry of mature residual teratoma after inten- sive chemotherapy of disseminated nonseminomatous germ cell tumor of the testis: A report of two cases. Cancer Genet Cytoge- net 22149-157.
Pui CH, Crist WM, Look AT (1990) Biology and clinical signifi- cance of cytogenetic abnormalities in childhood acute lympho- blastic leukemia. Blood 76:1449-1463.
Rodriguez E, Mathew S, Mukherjee AB, Reuter VE, Bosl GJ, Chaganti RSK (1992) Analysis of chromosome 12 aneuploidy in interphase cells from human male germ cell tumors by fluores- cence in situ hybridization. Genes Chromosom Cancer 521-29.
Samaniego F, Rodriguez E, Houldsworth J , Murty WVS, Ladanyi M, Lele KP, Chen Q, Dmitrovsky E, Geller NL, Reuter V, Jhanwar SC, Bosl GJ, Chaganti RSK (1990) Cytogenetic and mo- lecular analysis of human male germ cell tumors: Chromosome 12 '
abnormalities and gene amplification. Genes Chromosom Cancer
Sano T, Tsujino T , Yoshida K, Nakayama H, Haruma K, Ito H, Nakamura Y, Kajiyama G, Tahara E (1991) Frequent loss of het- erozygosity on chromosomes lq, 5q, and 17p in human gastric carcinomas. Cancer Res 51:2926-2931.
Shen V, Chaparro M, Choi BH, Young R, Bernstein R (1990) Ab- sence of isochromosome 12p in a pineal region malignant germ cell tumor. Cancer Genet Cytogenet 50: 153-160.
Speleman F, De Potter C, Dal Cin P, Mangelschots K, Ingelaere H, Laureys G, Benoit Y, Leroy J , Van Den Berghe H (1990) i(12p) in a malignant ovarian tumor. Cancer Genet Cytogenet 45:49-53.
Speleman F, Laureys G, Benoit Y, Cuvelier C, Suijkerbuijk R, de Jong B (1992) i( 12p) in a near-diploid mature ovarian teratoma. Cancer Genet Cytogenet 60:216-218.
Stock C, Ambros IM, Mann G, Gadner H, Amann G, Ambros PF (1993) Detection of lp36 deletions in paraffin sections of neuro- blastoma tissues. Genes Chromosom Cancer 6: 1-9.
Strehl S, Ambros P F (1993) Fluorescence in situ hybridization com- bined with immunohistochemistry for highly sensitive detection of chromosome 1 aberrations in neuroblastoma. Cytogenet Cell Genet 63:24-28.
Suijkerbuijk RF, van de Veen AY, van Echten J, Buys CHCM, de Jong B, Oosterhuis JW, Warburton DA, Cassiman JJ, Schonk D, Geurts van Kessel A (1991) Demonstration of the genuine iso-12p character of the standard marker chromosome of testicular germ cell tumors and identification of further chromosome 12 aberra- tions by competitive in situ hybridization. Am J Hum Genet 48:
Surti U, Hoffner L, Chakravarti A, Ferrell RE (1990) Genetics and biology of human ovarian teratomas. I. Cytogenetic analysis and mechanism of origin. Am J Hum Genet 47:635-643.
van Roy N, Laureys G, Versteeg R, Opdenakker G, Speleman F (1993) High-resolution fluorescence mapping of 46 DNA markers to the short arm of human chromosome 1. Genomics 18:71-78.
Wolfe J, Darling SM, Erickson RP, Craig IW, Buckle VJ, Rigby PWJ, Willard HF, Goodfellow PN (1985) Isolation and character- ization of an alphoid centromeric repeat family from the human Y chromosome. J Mol Biol 182477-485.
Wolman SR, Waldman FM, Balazs M (1993) Complementarity of interphase and metaphase chromosome analysis in human renal tumors. Genes Chromosom Cancer 6: 17-23.
Yang TP, Hansen SK, Oishi KK, Ryder OA, Hamkalo BA (1982) Characterization of a cloned repetitive DNA sequence concen- trated on the human X chromosome. Proc Natl Acad Sci USA 79:6593-6597.
1:289-300.
269-273.





























