Embed Size (px)
Citation preview

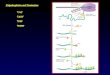
Molecular Model of αCaMKII-CPEB Interaction Loop
Protein Synthesis
Protein Synthesis
Degradation (rate - )
Degradation (rate - )
CaCaM
Gillespie Algorithm
Initialize the system and simulation parameters
Pick a type of molecule randomly
Is cumulative propensity positive?
Pick a reaction of thistype of molecule randomly and generate a random time interval
NO
YES
Update time by the stepgenerated previously, and population according to the picked reaction
Is desired time reached?
STOP
YES
NO
0.15 0.16 0.17 0.18 0.19 0.2 0.21 0.22 0.23 0.24 0.250
20
40
60
80
100
120
140
160
180
CPEB Deterministic Bifurcation Curve
Degradation Rate
Tota
l Am
ount
of C
aMKI
I (mol
)
Bifurcation Algorithm Initialize the system and simulation parameter
NO
YES
STOP
Is convergence possible?
Find all equilibria under this parameter
Generate a new parameter and find new equilibria
Is shift small enough or corrector reached max iteration?
Is desired number of data points reached?
YES
YES
NO
Modify predictions
NO
Cytoplasmic polyadenylation switching mechanism A comparison beteween deterministic and stochastic approaches
Xueyao Liu 1, Naveed Aslam 2, Harel Z. Shouval1 Department of Mathematics, University of California - Berkeley, Berkeley, CA 2 Department of Neurobiology and Anatomy, University of Texas Medical School, Houston, TX
2
Abstract
Translation efficiency of certain mRNAs is regulated through a cytoplasmic polyadenylation process at pre-initiation phase. A translation factor regulates the polyadenylation process through its posttranscriptional modification e.g., phosphorylation. The cytoplasmic polyadenylation binding protein (CPEB1) is one such translation factor which regulates the translation of mRNAs through cytoplasmic polyadenylation element (CPE). The cytoplasmic polyadenylation process can be turned on or off by the phosphorylation or dephosphorylation state of CPEB1. The phosphorylated form of CPEB1 increases the translational activity of an otherwise dormant mRNA. A physiological instantiation could be the regulation of αCaMKII mRNA stability through the phosphorylation- dephosphorylation cycle of CPEB1. Here, we show that CPEB1 mediated translation of αCaMKII mRNA through polyadenylation is regulated through a bistable switching mechanism. The simple translation switch for regulating the polyadenylation is based on two state model αCaMKII-CPEB1 molecular pair. Here, the de-novo synthesis of αCaMKII is modeled through an active/inactive form of αCaMKII mRNA. Based on elementary biochemical kinetics a highdimensional system of non-linear ordinary differential equations can describe the dynamic characteristics of the polyadenylation loop. We used deterministic and stochastic approaches to analyze the feasibility of CaMKII translation switching mechanism. We also developed the one parameter bifurcation diagram to show the numerical robustness of proposed switching mechanism.
The University of California, Berkeley
CPEB Stochastic Dynamics Curves ( = .16)
0 500 1000 1500 2000 2500 3000 3500 4000 4500 50002600
2800
3000
3200
3400
3600
3800
4000
Time t
Total
Amou
nt of
CaMK
II (m
olecu
les)
0 10 20 30 40 502920
2930
2940
2950
2960
2970
2980
2990
3000
Time t
Total
Amou
nt of
CaMK
II (m
olecu
les)
CPEB Deterministic Dynamics Curves ( = .16)
Time t
Total
Amou
nt of
CaMK
II (m
ol)
0 500 1000 1500 2000 2500 3000 3500 4000 4500 50000
50
100
150
Time t 0 20 40 60 80 100 120 140 160 180 200
10
20
30
40
50
60
70
80
90
100
Total
Amou
nt of
CaMK
II (m
ol)
DiscussionThis work proposes the general hypothesis that a feedback loop between a plasticity related kinase and its translation factor can act as a bistable switch thatstabilizes long term memory. The one parameter bifurcation diagram proves the existence of such a bistable switching system. In the deterministic dynamicsgraph, with a set of appropriate parameter, we also see two clearly separated steady states. However, because the concentration of translation machinery is extremely low in necks of spines and spines, it is more reasonable to assume that the translation process happens stochastically. Here, we observed differences between results from deterministic and stochastic approaches. The initial condition that merged to the lower steady state in deterministic approach seems to rise to the upper steady state in stochastic approach, and does so faster than the other condition that rose to the upper steady state in deterministic approach. This discrepancy happened either because there was an error in the implementation of Gillespie Algorithm or because stochasticity disturbs the system to an extent that bistability is impossible and so do the systems in naturebehave. Further investigations are needed to arrive at a more detailed conclusion.
Acknowledgements and References1. Allgower, E.L., Georg, K. (1990). Numerical Continuation Methods: An Introduction.Springer-Verlag, New York 2. Ullah, M., Schmidt, H., Cho, K.H. and Wolkenhauer, O.(2006).Deterministic modelling and stochastic simulation of biochemical pathways usingMATLAB. IEE Proceedings-Systems Biology153, 53-60
This work was partially supported by NSF REU Grant DMS-0755294