Embed Size (px)
Citation preview

FEBS Letters 580 (2006) 3462–3468
CHOP is involved in neuronal apoptosis induced by neurotrophicfactor deprivation
Seiji Tajiria,b, Shigetoshi Yanob,*, Motohiro Moriokab, Jun-ichi Kuratsub,Masataka Moria, Tomomi Gotoha,*
a Department of Molecular Genetics, Graduate School of Medical Sciences, Kumamoto University, Honjo 1-1-1, Kumamoto 860-8556, Japanb Department of Neurosurgery, Graduate School of Medical Sciences, Kumamoto University, Honjo 1-1-1, Kumamoto 860-8556, Japan
Received 10 March 2006; revised 28 April 2006; accepted 6 May 2006
Available online 15 May 2006
Edited by Vladimir Skulachev
Abstract Neurotrophic factors are essential for the survival ofneurons. We found that the endoplasmic reticulum (ER) stress-C/EBP homologues protein (CHOP) pathway to be activatedduring neurotrophic factor deprivation-induced apoptosis inPC12 neuronal cells and in primary cultured neurons, and thisapoptosis was suppressed in the neurons from chop�/� mice. Inaddition, we found that CHOP is expressed in the subventricularzone (SVZ) and striatum of the young adult mouse brain. Thenumber of apoptotic cells in the SVZ decreased in chop�/� mice.These results indicate that the ER stress-CHOP pathway plays arole in neuronal apoptosis during the development of the brain.� 2006 Federation of European Biochemical Societies. Publishedby Elsevier B.V. All rights reserved.
Keywords: Apoptosis; C/EBP homologues protein;Endoplasmic reticulum stress; Neurotrophic factor;Subventricular zone
1. Introduction
Apoptosis, observed during either synaptogenesis or neuro-
genesis in the paranatal or postnatal stage of the mammalian
brain, is thought to be induced by competition for the limited
amount of neurotrophic factors, which are essential for sur-
vival [1–3]. The precise control of the neuronal cell number
is critical for the morphological development, and also the nor-
mal function of the adult brain [4]. However, the mechanisms
of those neuronal apoptosis are still not well known.
Nerve growth factor (NGF) is needed for the differentiation
and the survival of PC12 cells. NGF binds to TrkA receptor on
the cell surface, and induces phosphorylation of the receptor.
The signal transduction pathway downstream of the TrkA
receptor phosphorylation includes the activations of several
molecules including MAP kinase. MAP kinase pathway is in-
volved in the NGF-induced differentiation of PC12 cells [5].
Abbreviations: CHOP, C/EBP homologues protein; ER, Endoplasmicreticulum; GAPDH, glyceraldehyde 3-phosphate dehydrogenase;LDH, lactate dehydrogenase; MAP2, microtubule associated protein2; NGF, nerve growth factor; SVZ, subventricular zone; TUNEL,terminal transferase-mediated dUTP-biotin nick end labeling
*Corresponding authors. Fax: +81 96 373 5145 (T. Gotoh); +81 96 3718064 (S. Yano).E-mail addresses: [email protected] (S. Yano),[email protected] (T. Gotoh).
0014-5793/$32.00 � 2006 Federation of European Biochemical Societies. Pu
doi:10.1016/j.febslet.2006.05.021
However, the inhibition of the MAP kinase pathway has been
reported to not induce apoptosis in differentiated PC12 cells [5]
and synaptic neurons [6]. NGF also binds to a low affinity
receptor p75NTR while inducing neuronal cell death. This type
of neuronal apoptosis is observed during development [7].
The endoplasmic reticulum (ER) is an organelle where
newly-synthesized secretory and cell membrane proteins are
modified and folded. ER is highly developed in neuronal cells
[8]. Protein modification and folding in the ER is impaired
under various physiological or pathological conditions [9,10].
Recent studies have revealed that the perturbation of the ER
functions, which is called ER stress, activates the ER stress
response pathways, including translational attenuation, the
induction of ER chaperones such as BiP, and the degradation
of unfolded proteins by the system called ERAD, to protect
cells [11]. However, when the ER functions are severely im-
paired, apoptosis occurs. This apoptosis is mediated by factors
including C/EBP homologues protein (CHOP)/GADD153
[12]. CHOP is a transcription factor which belongs to the C/
EBP family [13]. CHOP heterodimerizes with other C/EBP
family members, and CHOP-C/EBP dimer binds to the CHOP
site which is distinct from the C/EBP site, and thus induces
apoptosis. The target gene(s) of CHOP, which is directly in-
volved in the apoptosis pathway, is still not known. Therefore,
the apoptosis pathway downstream of CHOP is also not well
known. CHOP is induced at the transcription level in response
to ER stress. We found that the ER stress-CHOP pathway is
activated by various stresses including brain ischemia-reperfu-
sion and neuronal hypoxia-reoxygenation [14–17]. We also
found that CHOP-induced apoptosis is mediated by the trans-
location of a proapoptotic Bcl-2 family member Bax from
cytosol to mitochondria [18].
The withdrawal of NGF activates the mitochondria-medi-
ated apoptosis pathway in differentiated PC12 neuronal cells
[19,20]. Bax has been reported to be involved in the apoptosis
of neurons in the adult brain [21–23]. As mentioned above,
neuronal apoptosis in the paranatal or postnatal brain is
thought to be induced by the competition for limited amounts
of neurotrophic factors. We therefore investigated whether
neurotrophic factor deprivation-induced apoptosis is mediated
by the ER stress pathway including CHOP.
We herein report that NGF deprivation in differentiated
PC12 cells and neurotrophic factor deprivation in primary
cultured neurons activate the ER stress-CHOP-mediated
apoptosis. We also describe that the number of apoptotic
cells in subventricular zone (SVZ) and striatum is reduced
blished by Elsevier B.V. All rights reserved.

S. Tajiri et al. / FEBS Letters 580 (2006) 3462–3468 3463
in young adult chop�/� mice in comparison to wild-type mice.
These results suggest that CHOP plays a crucial role in the
development and maintenance of the postnatal brain.
2. Materials and methods
2.1. AntibodiesPolyclonal antibodies against mouse, human GRP78/BiP, CHOP,
glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and monoclo-nal antibody against mouse CHOP were obtained from Santa CruzBiotechnology Inc. (Santa Cruz, CA, USA). Monoclonal antibodyagainst mouse neuron-specific microtubule associated protein 2(MAP2) was obtained from the Sigma Chemical Co. (St. Louis, MO,USA).
2.2. Cell culture and neurotrophic factor deprivationUndifferentiated PC12 cells were cultured at 37 �C in Dulbecco’s
modified Eagle medium (DMEM) containing 4500 mg/l glucose, sup-plemented with 4 mM LL-glutamine, 5% fetal calf serum (FCS) and10% horse serum. The cells cultured (1 · 104 cells) were grown until50% confluent in 60 mm-diameter collagen-coated plates, and were in-duced to undergo differentiation by treating them with 50 ng/ml NGF(Wako Pure Chemical Industries, Osaka, Japan) in DMEM/F-12 med-ium with 2% FCS and 0.25% insulin–transferrin–selenium-X supple-ment (Invitrogen, Carlsbad, CA, USA) for 7 days. Then the cellswere cultured in the medium (DMEM/F-12 medium with 2% FCSand 0.25% insulin–transferrin–selenium-X supplement) without NGFfor the indicated periods to trigger cell death.
Primary cultured cells were prepared from the cortex of embryonicwild-type or chop�/� mice at days 16–18 of gestation (E16–18) as de-scribed [17]. The cells from cerebral cortexes were suspended in a Neu-robasal medium (Invitrogen) with 0.5 mM LL-glutamine, 25 lMglutamate, 100 units/ml penicillin, 50 lg/ml streptomycin and 2% B27supplement containing neurotrophic factors (Invitrogen). The cellswere plated at a density of 1 · 105 cells/well on poly-DD-lysine-coateddishes (Clontech Laboratories, Inc., Palo Alto, CA, USA) and culturedfor 10 days either with cytosine arabinocide (Ara-C). Cell death wasthen triggered by removal of B27 supplement from the medium.
2.3. Quantitative real-time RT-PCR and semi-quantitative RT-PCR
analysisTotal RNA from either differentiated PC12 cells or primary cultured
cortical neurons from wild-type and chop�/� mice was isolated usingthe acid guanidium thiocyanate–phenol–chloroform extraction proce-dure [24]. A real-time RT-PCR analysis was performed with iCyclerreal-time PCR system (Bio-Rad Laboratories Inc., Hercules, CA,USA) using QuantiTect SYBR Green RT-PCR kit (Qiagen, Hilden,Germany). The amplified products were verified as single expected sizebands by agarose gel electrophoresis. The results obtained with 10–105
copies of GAPDH cDNA were used as a standard curve, and all resultswere standardized to the levels of GAPDH. The primers used for PCRwere as follows: sense primer, 5 0-GAGATTGTTCTGGTTGGCG-GATCTACTC-3 0 and antisense primer, 5 0-CCATATGCTA-CAGCCTCATCTGGGTT-3 0 for rat BiP (GenBank Accession No.M14050); sense primer, 5 0-CCTGAAAGCAGAAACCGGTC-3 0 andantisense primer, 5 0-CCTCATACCAGGCTTCCAGC-30 for ratCHOP (GenBank Accession No. NM_024134); sense primer, 5 0-TCTGTATGAGCCCTGAGTCCTACCT-3 0 and antisense primer,5 0-GGTCATAAGGTTTGGGTCGAGAACCAC-3 0 for rat ATF4(GenBank Accession No. NM_024403); sense primer, 5 0-CCTCTG-GAGGTGCCAAGGAG-3 0 and antisense primer, 5 0-TTGGATCC-TGCTGGGCGCTGTGCTGTC-3 0 for rat TrkA (GenBank AccessionNo. NM_012731) [25]; sense primer, 5 0-GTCGTGGGCCTTGTGG-CC-3 0 and antisense primer, 5 0-CTGTGAGTTCACACTGGGG-3 0
for rat p75NTR (GenBank Accession No. NM_012610) [25]; senseprimer, 5 0-AAACAGAGTAGCAGCGCAGACTGC-3 0 and antisenseprimer, 5 0-GGATCTCTAAAACTAGAGGCTTGGTG-30 for mouseXBP1 (GenBank Accession No. NM_013842) [26], sense primer,5 0-GAACATCATCCCTGCATCCA-3 0 and antisense primer, 5 0-CC-AGTGAGCTTCCCGTTCA-3 0 for rat GAPDH (GenBank AccessionNo. NW_047622); sense primer, 5 0-GAAAGGATGGTTAATGATG-
CTGAGAAG-3 0 and antisense primer, 5 0-GTCTTCAATGTCCGC-ATCCTG-30 for mouse BiP (GenBank Accession No. NM_022310);sense primer, 5 0-CATACACCACCACACCTGAAAG-3 0 and anti-sense primer, 5 0-CCGTTTCCTAGTTCTTCCTTGC-3 0 for mouseCHOP (GenBank Accession No. NT_081856); sense primer, 5 0-CAC--GAAATCCAGCAGCAGTGT-3 0 and antisense primer, 5 0-GGCTG-CAAGAATGTAAAGGGG-3 0 for mouse ATF4 (GenBank AccessionNo. BC085169); sense primer, 5 0-TGGCACAGTCAAGGCTGAGA-3 0 and antisense primer, 5 0-CTTCTGAGTGGCAGTGATGG-3 0 formouse GAPDH (GenBank accession number BC083149). All expectedPCR products, except for rat and mouse GAPDH, contained pluralexons. For a semiquantitative analysis, RT-PCR was performed usingthe above primer sets and the Superscript One-Step RT-PCR System(Invitrogen).
2.4. Immunoblot analysisDifferentiated PC12 cells and primary cultured neurons were
homogenized in 50 mM Tris–HCl containing 150 mM NaCl, 1% Tri-ton X-100. After centrifugation, the supernatants were used for animmunoblot analysis. Immunodetection was performed as described[18].
2.5. Immunocytochemical and immunohistochemical analysisFor the immunocytochemical analysis of PC12 cells or primary cul-
tured cortical cells were performed as described [17].For the immunohistochemical analysis, mice were anesthetized by
them giving sodium pentobarbital (50 ml/g, i.p.), then the whole brainwas removed. Immunohistochemical analysis using anti-rabbit CHOPpolyclonal antibody as the first antibody and the second antibody con-jugated with peroxidase (Amersham Pharmacia Biotech) was per-formed as described [17]. The peroxidase activity was visualized byincubation with a 3,3 0-diaminobenzidine solution.
2.6. Detection of apoptosisLactate dehydrogenase (LDH) release from damaged cells was mea-
sured as an index of cell destruction, using a LDH cell damage assay kit(Wako Pure Chemical Industries). The disruption of the mitochondrialmembrane potential is characteristic of apoptotic cells. To analyzemitochondrial membrane depolarization, the cells were stained with amitochondrial membrane potential-indicating dye DePsipher (TrevigenInc., Gaithersburg, MD, USA) as previously described [14]. To analyzethe morphological changes of nuclei, the cells were stained with Hoe-chst dye 33258 as previously described [15]. The detection of apoptosisin vivo was performed by the terminal transferase-mediated dUTP-bio-tin nick end labeling (TUNEL) method using an in situ apoptosis detec-tion kit (Takara, Otsu, Japan) as previously described [17]. TUNELsignals were amplified with Streptavidin Alexa Fluora 488 conjugate(Molecular Probes) using the TSA Biotin System (Perkin–Elmer LifeSciences Inc., Boston, MA, USA). Nuclear staining with propidiumiodide (PI) was performed as previously described [17].
2.7. Statistical analysisQuantitative results were expressed as the means ± S.D. and ana-
lyzed using Student’s t-test. The significance in difference was assignedat a level of less than a 5% probability (P < 0.05).
3. Results
3.1. NGF deprivation-induced apoptosis in differentiated PC12
cells
Differentiation of PC12 cells were induced by treatment with
NGF for 7 days. Differentiation was confirmed by the morpho-
logical changes, such as the outgrowth of neurites and the flat-
tening of cell bodies. Twenty-four hours after NGF deprivation
from the medium of differentiated cells, morphological apopto-
tic changes, such as the disappearance of neurites, round-
shaped cells, shrunken cells and detachment from the culture
bottom, were observed in phase-contrast image (Fig. 1A).
LDH release from the cells was used as an index of cell death

Fig. 1. Induction of NGF deprivation-induced apoptosis in differen-tiated PC12 cells. (A) Differentiation of PC12 cells were induced bytreatment with NGF (50 ng/ml) for 7 days. After differentiation, thecells were cultured with (a) or without (b) NGF for 24 h. Phase-contrast images are shown. Bar, 20 lm. (B) Differentiated PC12 cellswere cultured with or without NGF (50 ng/ml), or with tunicamycin(Tm, 1 lg/ml) plus NGF (50 ng/ml) for the indicated periods. TheLDH activity of the culture medium was measured, and the results areshown as the means ± S.D. (n = 4). The LDH activity of cell extractbefore treatment is set at 100%. (C) Differentiated PC12 cells weretreated as in (A), and then were stained with a mitochondrialmembrane potential-indicating dye DePsipher. The red fluorescencerepresents the intact potential, while the green fluorescence representsthe disrupted potential. The arrowhead indicates a cell negative formembrane potential. Bars, 20 lm. (D) Differentiated PC12 cells weretreated as in (A), then were stained with Hoechst dye 33258. Thefluorescence images and phase-contrast images of the same fields areshown. The arrowheads indicate cells showing apoptotic changes. Bar,20 lm.
Fig. 2. Induction of ER stress-associated mRNAs in differentiatedPC12 cells treated with the deprivation of NGF. (A) DifferentiatedPC12 cells were cultured without NGF for the indicated periods on thetop, and total RNAs were subjected to the RT-PCR analysis forCHOP, BiP, ATF4 and GAPDH. (B) The results of (A) and two otherindependent experiments were calculated, and were standardized to thelevels of GAPDH. The results at 0 h are set as 1.0, and are shown asthe means ± S.D. (C) The differentiated PC12 cells were culturedwithout NGF for the indicated periods on the top, then the RT-PCRanalysis was performed using a primer set encompassing the spliced-out region in XBP1 mRNA. The PCR products were resolved on 3%agarose gel to separate the unspliced (XBP1u) and spliced (XBP1s)XBP1 mRNAs. The differentiated PC12 cells treated with tunicamycin(Tm, 1 lg/ml) for 6 h were used as a positive control. (D) A RT-PCRanalysis for TrkA, p75NTR mRNAs and GAPDH was performed usingtotal RNAs from differentiated PC12 cells cultured without NGF forthe indicated periods. (E) Differentiated PC12 cells were culturedwithout NGF for the indicated periods on the top, and whole cellextracts were subjected to an immunoblot analysis using antibodiesagainst CHOP (diluted 1000-fold), BiP (diluted 1000-fold) andGAPDH (diluted 1000-fold). Whole cell extracts of PC12 cells treatedwith tunicamycin (Tm, 1 lg/ml) for 12 h were used as a positivecontrol.
3464 S. Tajiri et al. / FEBS Letters 580 (2006) 3462–3468
(Fig. 1B). When the cells were cultured without NGF, LDH re-
lease began at 6 h and increased thereafter. LDH release in-
duced by NGF deprivation was delayed compared to the cells
treated with an ER stress-inducer tunicamycin, and reached
to about 80% of that induced by tunicamycin at 24 h. Apopto-
sis was confirmed by staining with a mitochondria membrane
potential indicating dye (Fig. 1C) and nuclear staining
(Fig. 1D). When the cells were cultured without NGF for
24 h, most cells (70%) lost mitochondrial membrane potential,
thus indicating cell death. Nuclear condensation, nuclear frag-
mentation and chromatin condensation, which are characteris-
tic changes of apoptotic cells, were observed in the nuclear
staining of the cells cultured without NGF. These results show
that NGF deprivation induces apoptosis in differentiated PC12
cells.
3.2. Activation of the ER stress-CHOP pathway by NGF
deprivation
Next we examined whether the ER stress pathway is acti-
vated in NGF deprivation-induced apoptosis (Fig. 2). CHOP
mRNA was detected at a low level before deprivation, induced
by about 4-fold at 3–6 h, and gradually decreased thereafter
(Fig. 2A and B). BiP, an ER-resident Hsp70 family member,
is known to be induced by ER stress. BiP mRNA was present
at a low level before deprivation, and it was induced by about
3-fold at 3–6 h, and thereafter it gradually decreased. mRNA
for ATF4 which is one of the transcriptional activator of the
chop gene, was detected at a low level before deprivation,
and it was induced by about 4-fold at 3 h, and then it rapidly
decreased thereafter, and finally reached a level close to the
control level at 12 h. XBP1 is a member of the CREB/ATF
family of transcription factors, which is specifically induced
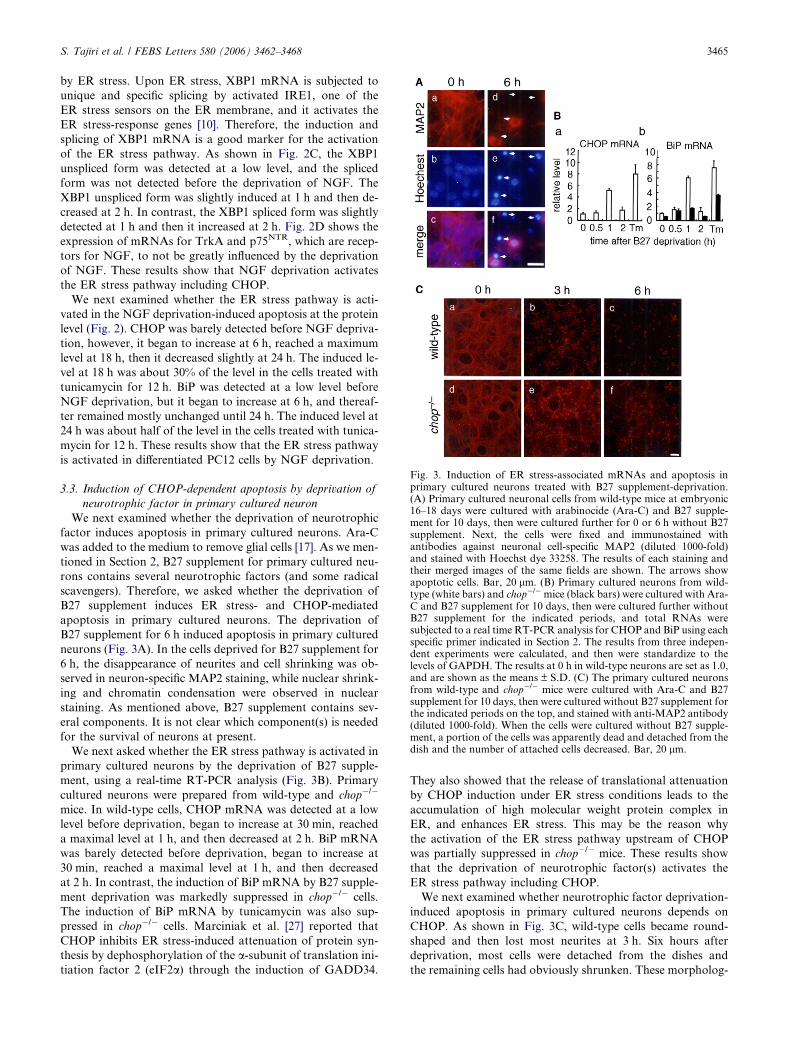
Fig. 3. Induction of ER stress-associated mRNAs and apoptosis inprimary cultured neurons treated with B27 supplement-deprivation.(A) Primary cultured neuronal cells from wild-type mice at embryonic16–18 days were cultured with arabinocide (Ara-C) and B27 supple-ment for 10 days, then were cultured further for 0 or 6 h without B27supplement. Next, the cells were fixed and immunostained withantibodies against neuronal cell-specific MAP2 (diluted 1000-fold)and stained with Hoechst dye 33258. The results of each staining andtheir merged images of the same fields are shown. The arrows showapoptotic cells. Bar, 20 lm. (B) Primary cultured neurons from wild-type (white bars) and chop�/�mice (black bars) were cultured with Ara-C and B27 supplement for 10 days, then were cultured further withoutB27 supplement for the indicated periods, and total RNAs weresubjected to a real time RT-PCR analysis for CHOP and BiP using eachspecific primer indicated in Section 2. The results from three indepen-dent experiments were calculated, and then were standardize to thelevels of GAPDH. The results at 0 h in wild-type neurons are set as 1.0,and are shown as the means ± S.D. (C) The primary cultured neuronsfrom wild-type and chop�/� mice were cultured with Ara-C and B27supplement for 10 days, then were cultured without B27 supplement forthe indicated periods on the top, and stained with anti-MAP2 antibody(diluted 1000-fold). When the cells were cultured without B27 supple-ment, a portion of the cells was apparently dead and detached from thedish and the number of attached cells decreased. Bar, 20 lm.
S. Tajiri et al. / FEBS Letters 580 (2006) 3462–3468 3465
by ER stress. Upon ER stress, XBP1 mRNA is subjected to
unique and specific splicing by activated IRE1, one of the
ER stress sensors on the ER membrane, and it activates the
ER stress-response genes [10]. Therefore, the induction and
splicing of XBP1 mRNA is a good marker for the activation
of the ER stress pathway. As shown in Fig. 2C, the XBP1
unspliced form was detected at a low level, and the spliced
form was not detected before the deprivation of NGF. The
XBP1 unspliced form was slightly induced at 1 h and then de-
creased at 2 h. In contrast, the XBP1 spliced form was slightly
detected at 1 h and then it increased at 2 h. Fig. 2D shows the
expression of mRNAs for TrkA and p75NTR, which are recep-
tors for NGF, to not be greatly influenced by the deprivation
of NGF. These results show that NGF deprivation activates
the ER stress pathway including CHOP.
We next examined whether the ER stress pathway is acti-
vated in the NGF deprivation-induced apoptosis at the protein
level (Fig. 2). CHOP was barely detected before NGF depriva-
tion, however, it began to increase at 6 h, reached a maximum
level at 18 h, then it decreased slightly at 24 h. The induced le-
vel at 18 h was about 30% of the level in the cells treated with
tunicamycin for 12 h. BiP was detected at a low level before
NGF deprivation, but it began to increase at 6 h, and thereaf-
ter remained mostly unchanged until 24 h. The induced level at
24 h was about half of the level in the cells treated with tunica-
mycin for 12 h. These results show that the ER stress pathway
is activated in differentiated PC12 cells by NGF deprivation.
3.3. Induction of CHOP-dependent apoptosis by deprivation of
neurotrophic factor in primary cultured neuron
We next examined whether the deprivation of neurotrophic
factor induces apoptosis in primary cultured neurons. Ara-C
was added to the medium to remove glial cells [17]. As we men-
tioned in Section 2, B27 supplement for primary cultured neu-
rons contains several neurotrophic factors (and some radical
scavengers). Therefore, we asked whether the deprivation of
B27 supplement induces ER stress- and CHOP-mediated
apoptosis in primary cultured neurons. The deprivation of
B27 supplement for 6 h induced apoptosis in primary cultured
neurons (Fig. 3A). In the cells deprived for B27 supplement for
6 h, the disappearance of neurites and cell shrinking was ob-
served in neuron-specific MAP2 staining, while nuclear shrink-
ing and chromatin condensation were observed in nuclear
staining. As mentioned above, B27 supplement contains sev-
eral components. It is not clear which component(s) is needed
for the survival of neurons at present.
We next asked whether the ER stress pathway is activated in
primary cultured neurons by the deprivation of B27 supple-
ment, using a real-time RT-PCR analysis (Fig. 3B). Primary
cultured neurons were prepared from wild-type and chop�/�
mice. In wild-type cells, CHOP mRNA was detected at a low
level before deprivation, began to increase at 30 min, reached
a maximal level at 1 h, and then decreased at 2 h. BiP mRNA
was barely detected before deprivation, began to increase at
30 min, reached a maximal level at 1 h, and then decreased
at 2 h. In contrast, the induction of BiP mRNA by B27 supple-
ment deprivation was markedly suppressed in chop�/� cells.
The induction of BiP mRNA by tunicamycin was also sup-
pressed in chop�/� cells. Marciniak et al. [27] reported that
CHOP inhibits ER stress-induced attenuation of protein syn-
thesis by dephosphorylation of the a-subunit of translation ini-
tiation factor 2 (eIF2a) through the induction of GADD34.
They also showed that the release of translational attenuation
by CHOP induction under ER stress conditions leads to the
accumulation of high molecular weight protein complex in
ER, and enhances ER stress. This may be the reason why
the activation of the ER stress pathway upstream of CHOP
was partially suppressed in chop�/� mice. These results show
that the deprivation of neurotrophic factor(s) activates the
ER stress pathway including CHOP.
We next examined whether neurotrophic factor deprivation-
induced apoptosis in primary cultured neurons depends on
CHOP. As shown in Fig. 3C, wild-type cells became round-
shaped and then lost most neurites at 3 h. Six hours after
deprivation, most cells were detached from the dishes and
the remaining cells had obviously shrunken. These morpholog-

3466 S. Tajiri et al. / FEBS Letters 580 (2006) 3462–3468
ical changes suggested apoptosis. In the case of chop�/� cells,
some cells still preserved neurites and many more attached
cells were observed at 6 h after deprivation. These results show
that the apoptosis induced by the neurotrophic factor depriva-
tion in primary cultured neuron is CHOP-dependent.
3.4. Induction of apoptosis by CHOP in neurons of SVZ and
striatum of young adult mice
In the brain, an excess number of neurons are produced dur-
ing development, and those excess neurons later disappear
through apoptosis, due to competition for limited amounts
of neurotrophic factors [2,3]. Recently, it was revealed that
neuronal progenitor cells exist in the SVZ of the cerebrum even
in adult mice [28]. These progenitor cells always need stimula-
tion by neurotrophic factors for their survival. As mentioned
above, neurotrophic factor deprivation induces ER stress-med-
iated apoptosis in primary cultured neuronal cells. Therefore,
we investigated whether the apoptosis of neuronal progenitor
cells and/or striatum neurons in adult mice is mediated by
the ER stress-CHOP pathway. Immunostaining of CHOP
was performed in the SVZ and striatum of young adult
(P14) mice (Fig. 4). Many neuronal cells, whose nuclei were
positive for CHOP, were observed in the SVZ and striatum
of wild-type mice (Fig. 4A and B). Most CHOP-positive cells
showed nuclear shrinking, thus suggesting apoptosis. In
chop�/� mice, no such cells were observed (Fig. 4C and D).
We next examined apoptosis using the TUNEL method
(Fig. 5). In the SVZ of wild-type mice, many TUNEL-positive
cells were detected, and these cells showed nuclear shrinking in
Fig. 4. Expression of CHOP in the SVZ and the striatum in the mousebrain at postnatal day 14. Coronal slices (5 lm) of wild-type (A and B)and chop�/� mice (C and D) brain at postnatal day 14 wereimmunostained with anti-mouse CHOP monoclonal antibody (diluted200-fold). Representative photographs of the SVZ (A and C) and thestriatum (B and D) are shown. Magnified images of the squares arealso shown. The arrowheads show cells positive for CHOP. Bar,20 lm.
Fig. 5. Suppression of apoptosis of the SVZ in chop�/� mice. (A) Inthe SVZ of wild-type and chop�/� mice brain at postnatal day 21,apoptosis was detected with the TUNEL method. The magnifiedimages of the squares are also shown. The results of TUNEL staining,nuclear staining with propidium iodide (PI) and their merged images ofthe same fields are shown. Bars, 20 lm. (B) TUNEL-positive cells inthe SVZ of wild-type and chop�/� mice in each hemisphere werecounted and were shown as the means ± S.D. (*; P < 0.05, n = 4). (C)The total brain weight of male wild-type and chop�/� mice, femalewild-type and chop�/�mice were measured on postnatal day 14 and 42,and the results are shown as the means ± S.D. (n = 7).
PI staining (Fig. 5A). In contrast, TUNEL-positive cells were
only rarely observed in the SVZ of chop�/� mice. The number
of TUNEL-positive cells per cerebral hemisphere in chop�/�
mice was about 25% of that in the wild-type mice (Fig. 5B).
The brain weight was higher in chop�/� mice than in wild-type
animals (Fig. 5C). These results strongly suggest that the apop-
tosis of neuronal cells in adult brain is mediated by the ER
stress-CHOP pathway, thus supporting the idea that apoptosis
is involved in the development or maintenance of the brain
after birth by eliminating any excess neurons.
4. Discussion
In this study, we found that NGF deprivation induces ER
stress-mediated apoptosis in differentiated PC12 neuronal cells.
We also found that the removal of B27 supplement from
the medium induces CHOP-dependent apoptosis in primary
cultured neurons. B27 supplement is used as an alternative

S. Tajiri et al. / FEBS Letters 580 (2006) 3462–3468 3467
for fetal calf serum, and it contains several hormones (cortico-
sterone, progesterone, triodo-1-thyronine and insulin), several
radical scavengers and some other components [29]. It is not
known about the anti-ER stress-induced apoptosis effects of
these hormones in neuronal cells at present. There have been
several reports regarding the mechanism of NGF depriva-
tion-induced apoptosis [5,30]. We need to investigate which
composition(s) of B27 supplement is crucial for the survival.
Reactive oxygen species can be the inducer of ER stress [27].
Therefore, we also need to consider about the anti-apoptotic
functions of radical scavengers. Deckwerth and Johnson [31]
reported that glucose uptake in differentiated PC12 cells falls
rapidly to 35% of the control level within 6 h after NGF depri-
vation. A decrease in the glucose uptake can disturb protein
modification and cause ER stress. Then the expression of ER
stress-related molecules including CHOP is induced [32,33].
In fact, we found that chop�/� mouse-derived neurons are
resistant to neurotrophic factor deprivation-induced apopto-
sis. Aoki et al. [34] showed that BiP mRNA is induced by
NGF deprivation in PC12 cells. Under our conditions, neuro-
trophic factor deprivation-induced apoptosis was not com-
pletely suppressed in the chop�/� primary cultured neurons.
At least three pathways of ER stress-induced apoptosis are
present. They are the CHOP pathway, the TRAF2-ASK1-
JNK pathway and the caspase 12 pathway [10]. We speculate
that other two pathways except the CHOP pathway are in-
volved in the late apoptosis of chop�/� neurons. The precise
mechanism of how neurotrophic factor deprivation induces
ER stress remains to be investigated.
We also found that CHOP is expressed in SVZ and the
striatum of young adult mice, and that the nuclei of CHOP-
expressing showed apoptotic changes. Apoptosis was sup-
pressed in the SVZ of chop�/� mice. From our present results,
we speculate that the ER stress-CHOP pathway is therefore
involved in the development and/or maintenance of the brain
after birth, by its role in the removal of excess neurons.
Acknowledgments: We thank Dr. Shizuo Akira (Osaka University, Ja-pan) for the chop�/� mice. We thank our colleagues for valuable sug-gestions and discussions. We also thank R. Shindo and M. Obata fortechnical assistance, and Y. Indo for secretarial assistance. This workwas supported in part by Grants-in-Aid (Nos. 14037257 and 17390096to M.M., 16590233 to T.G. and 16591448 to S.Y.) from the Ministry ofEducation, Culture, Sports, Science and Technology of Japan, and agrant (to T.G.) from the Inamori Foundation.
References
[1] Kuan, C.Y., Roth, K.A., Flavell, R.A. and Rakic, P. (2000)Mechanisms of programmed cell death in the developing brain.Trends Neurosci. 23, 291–297.
[2] Levi-Montalcini, R. and Angeletti, P.U. (1968) Nerve growthfactor. Physiol. Rev. 48, 534–569.
[3] Alvarez-Buylla, A., Garcia-Verdugo, J.M. and Tramontin, A.D.(2001) A unified hypothesis on the lineage of neural stem cells.Nat. Rev. Neurosci. 2, 287–293.
[4] Gould, E., McEwen, B.S., Tanapat, P., Galea, L.A. and Fuchs, E.(1997) Neurogenesis in the dentate gyrus of the adult tree shrew isregulated by psychosocial stress and NMDA receptor activation.J. Neurosci. 17, 2492–2498.
[5] Yao, R. and Cooper, G.M. (1995) Requirement for phosphati-dylinositol-3 kinase in the prevention of apoptosis by nervegrowth factor. Science 267, 2003–2006.
[6] Greenlund, L.J., Deckwerth, T.L. and Johnson Jr., E.M. (1995)Superoxide dismutase delays neuronal apoptosis: a role for
reactive oxygen species in programmed neuronal death. Neuron14, 303–315.
[7] Frade, J.M., Rodriguez-Tebar, A. and Barde, Y.A. (1996)Induction of cell death by endogenous nerve growth factorthrough its p75 receptor. Nature 383, 166–168.
[8] Majdan, M., Lachance, C., Gloster, A., Aloyz, R., Zeindler, C.,Bamji, S., Bhaker, A., Belliveau, D., Fawcett, J., Miller, F.D. andBarker, P.A. (1997) Transgenic mice expressing the intracellulardomain of the p75 neurotrophin receptor undergo neuronalapoptosis. J. Neurosci. 17, 6988–6998.
[9] Kaufman, R.J., Scheuner, D., Schroder, M., Shen, X., Lee, K.,Liu, C.Y. and Arnold, S.M. (2002) The unfolded protein responsein nutrient sensing and differentiation. Nat. Rev. Mol. Cell Biol. 3,411–421.
[10] Oyadomari, S., Araki, E. and Mori, M. (2002) Endoplasmicreticulum stress-mediated apoptosis in pancreatic b-cells. Apop-tosis 7, 335–345.
[11] Yoshida, H., Matsui, T., Hosokawa, N., Kaufman, R.J., Nagata,K. and Mori, K. (2003) A time-dependent phase shift in themammalian unfolded protein response. Dev. Cell 4, 265–271.
[12] Ferri, K.F. and Kroemer, G. (2001) Organelle-specific initiationof cell death pathways. Nat. Cell Biol. 3, E255–E263.
[13] Ron, D. and Habener, J.F. (1992) CHOP, a novel developmen-tally regulated nuclear protein that dimerizes with transcriptionfactors C/EBP and LAP and functions as a dominant-negativeinhibitor of gene transcription. Genes Dev. 6, 439–453.
[14] Oyadomari, S., Takeda, K., Takiguchi, M., Gotoh, T., Matsum-oto, M., Wada, I., Akira, S., Araki, E. and Mori, M. (2001) Nitricoxide-induced apoptosis in pancreatic b cells is mediated by theendoplasmic reticulum stress pathway. Proc. Natl. Acad. Sci.USA 98, 10845–10850.
[15] Gotoh, T., Oyadomari, S., Mori, K. and Mori, M. (2002) Nitricoxide-induced apoptosis in RAW 264.7 macrophages is mediatedby endoplasmic reticulum stress pathway involving ATF6 andCHOP. J. Biol. Chem. 277, 12343–12350.
[16] Oyadomari, S., Koizumi, A., Takeda, K., Gotoh, T., Akira, S.,Araki, E. and Mori, M. (2002) Targeted disruption of the Chopgene delays endoplasmic reticulum stress-mediated diabetes. J.Clin. Invest. 109, 525–532.
[17] Tajiri, S., Oyadomari, S., Yano, S., Morioka, M., Gotoh, T.,Hamada, J-I., Ushio, Y. and Mori, M. (2004) Ischemia-inducedneuronal cell death is mediated by the endoplasmic reticulumstress pathway involving CHOP. Cell Death Differ. 11, 403–415.
[18] Gotoh, T., Terada, K., Oyadomari, S. and Mori, M. (2004)hsp70-DnaJ chaperone pair prevents nitric oxide- and CHOP-induced apoptosis by inhibiting translocation of Bax to mito-chondria. Cell Death Differ. 11, 390–402.
[19] Putcha, G.V., Deshmukh, M. and Johnson Jr., E.M. (1999) BAXtranslocation is a critical event in neuronal apoptosis: regulationby neuroprotectants, BCL-2, and caspases. J. Neurosci. 19, 7476–7485.
[20] Vyas, S., Juin, P., Hancock, D., Suzuki, Y., Takahashi, R.,Triller, A. and Evan, G. (2004) Differentiation-dependent sensi-tivity to apoptogenic factors in PC12 cells. J. Biol. Chem. 279,30983–30993.
[21] Deckwerth, T.L., Elliott, J.L., Knudson, C.M., Johnson Jr., E.M.,Snider, W.D. and Korsmeyer, S.J. (1996) BAX is required forneuronal death after trophic factor deprivation and duringdevelopment. Neuron 17, 401–411.
[22] Lindsten, T., Golden, J.A., Zong, W.X., Minarcik, J., Harris,M.H. and Thompson, C.B. (2003) The proapoptotic activities ofBax and Bak limit the size of the neural stem cell pool. J.Neurosci. 23, 11112–11119.
[23] Sun, W., Winseck, A., Vinsant, S., Park, O.H., Kim, H. andOppenheim, R.W. (2004) Programmed cell death of adult-generated hippocampal neurons is mediated by the proapoptoticgene Bax. J. Neurosci. 24, 11205–11213.
[24] Chomczynski, P. and Sacchi, N. (1997) Single-step method ofRNA isolation by acid guanidinium thiocyanate–phenol–chloro-form extraction. Ann. Biochem. 162, 156–159.
[25] Gascon, E., Vutskits, L., Zhang, H., Barral-Moran, M.J., Kiss,P.J., Mas, C. and Kiss, J.Z. (2005) Sequential activation of p75and TrkB is involved in dendritic development of subventricularzone-derived neuronal progenitors in vitro. Eur. J. Neurosci. 21,69–80.

3468 S. Tajiri et al. / FEBS Letters 580 (2006) 3462–3468
[26] Calfon, M., Zeng, H., Urano, F., Till, J.H., Hubbard, S.R.,Harding, H.P., Clark, S.G. and Ron, D. (2002) IRE1 couplesendoplasmic reticulum load to secretory capacity by processingthe XBP-1 mRNA. Nature 415, 92–96.
[27] Marciniak, S.J., Yun, C.Y., Oyadomari, S., Nvoa, I., Zhang, Y.,Jungreis, R., Nagata, K., Harding, H.P. and Ron, D. (2004)CHOP induces death by promoting protein synthesis and oxida-tion in the stressed endoplasmic reticulum. Genes Dev. 18, 3066–3077.
[28] Alvarez-Buylla, A. and Garcia-Verdugo, J.M. (2002) Neurogen-esis in adult subventricular zone. J. Neurosci. 22, 629–634.
[29] Brewer, G.J., Torricelli, J.R., Evege, E.K. and Price, P.J. (1993)Optimized survival of hippocampal neurons in B27-supplementedNeurobasal, a new serum-free medium combination. J. Neurosci.Res. 35, 567–576.
[30] Chao, M.V. and Bothwell, M. (2002) Neurotrophins: to cleave ornot to cleave. Neuron 33, 9–12.
[31] Deckwerth, T.L. and Johnson Jr., E.M. (1993) Temporal analysisof events associated with programmed cell death (apoptosis) ofsympathetic neurons deprived of nerve growth factor. J. Cell Biol.123, 1207–1222.
[32] Kaufman, R.J. (2002) Orchestrating the unfolded proteinresponse in health and disease. J. Clin. Invest. 110, 1389–1398.
[33] Oyadomari, S. and Mori, M. (2004) Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ.11, 381–389.
[34] Aoki, T., Koike, T., Nakano, T., Shibahara, K., Kondo, S.,Kikuchi, H. and Honjo, T. (1997) Induction of Bip mRNA uponprogrammed cell death of differentiated PC12 cells as well as ratsympathetic neurons. J. Biochem. (Tokyo) 121, 122–127.









![Original Article MiRNA-214 ameliorates neuronal apoptosis ... · MiRNA-214 inhibits neuronal apoptosis 6294 Int J Clin Exp Med 2017;10(4):6293-6302 repression [11]. Recently, it has](https://img.dokumen.tips/doc/110x75/5fea33618b8dc9208a0e1027/original-article-mirna-214-ameliorates-neuronal-apoptosis-mirna-214-inhibits.jpg)



















