Embed Size (px)
Citation preview

HAL Id: hal-00902441https://hal.archives-ouvertes.fr/hal-00902441
Submitted on 1 Jan 1996
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Changes in intestinal intra-epithelial and systemic T-cellsubpopulations after an Eimeria infection in chickens:comparative study between E acervulina and E tenella
M Bessay, Y Le Vern, D Kerbœuf, P Yvoré, P Quéré
To cite this version:M Bessay, Y Le Vern, D Kerbœuf, P Yvoré, P Quéré. Changes in intestinal intra-epithelial andsystemic T-cell subpopulations after an Eimeria infection in chickens: comparative study between Eacervulina and E tenella. Veterinary Research, BioMed Central, 1996, 27 (4-5), pp.503-514. <hal-00902441>

Original article
Changes in intestinal intra-epithelial and systemicT-cell subpopulations after an Eimeria infection
in chickens: comparative studybetween E acervulina and E tenella
M Bessay Y Le Vern D Kerbœuf P Yvoré P Quéré
1 Laboratoire de protozoologie;2 Service commun de cytométrie;
3 Laboratoire de virologie aviaire et d’oncologie, station de pathologie aviaire et parasitologie,Inra, 37380 Nouzilly, France
(Received 29 November 1995; accepted 11 April 1996)
Summary ― During chicken coccidiosis, the growth of the parasite in the intestinal epithelium cells leadsto the development of host immune response. Cell-mediated immune mechanisms appear to be mainlyresponsible for the acquired resistance to disease. The action of two species of Eimeria, with two dif-ferent intestinal localizations, on T-lymphocyte subsets was followed by fluorescent antibody cell-sorter analysis, locally at the intestinal site of the parasitic development and systemically in spleen andblood. An Eimeria acervulina infection, localized in duodenum, induced a significant increase in the pro-portion of CD4+ (up to 15%), CD8+ (up to 12%) and TCR7/6 (up to 6%) in the duodenal intraepithelialleucocytes (IEL) from day 4 to day 8 Pi, and an increase in the proportion of IgM+ cells (12%) on day8. At the same time, the proportion of CD8+ cells dropped significantly in the blood and spleen (-5 to- 10%) on days 4 and 6 PI and then increased with the proportion of CD4+ cells on day 8. An E tenellainfection, localized in caecum, increased the proportion of CD4+ cells on day 8 PI (20%) and of CD8+cells (10°/ ) on days 6 and 8 PI in caecal IEL. A negative or zero effect on the proportion of TCR7/5+ cellswas observed as well as on the IgM+ cells. At the same time, the proportion of CD4+ cells dropped inthe spleen on day 8 Pi (-10%) and that of CD8+ cells dropped in the blood on day 6 (-15%). In con-clusion, Eimeria infection seems to rapidly induce, locally at the site of the parasite development, a dra-matic modification of the proportion of T-cell subsets in IEL, accompanied by systemic variations thatare generally opposing, in the lymphocyte populations. The timing of the changes seems to followthe phases of the parasitic cycle for the Eimeria species considered.
chicken / Eimeria acervulina l Eimeria tenella l intraepithelial leukocyte / immunity
*
Correspondence and reprints

Résumé ― Modifications des sous-populations de cellules T intraépithéliales intestinales et sys-témiques après infection chez le poulet. Comparaison entre E acervulina et E tenella. Les coc-cidies aviaires se développent dans les cellules épithéliales de l’intestin. La multiplication parasitaireconduit à l’apparition d’une réponse immune chez l’hôte, dont les mécanismes d’immunité à médiationcellulaire paraissent être les principaux responsables de la résistance acquise. L’effet de l’infection pardeux espèces d’Eimeria à localisations intestinales différentes, sur les populations lymphocytaires T,a été suivi par cytométrie de flux, localement au lieu de multiplication parasitaire, dans le sang etdans la rate. L’infection par E acervulina, se développant dans le duodénum, induit une augmentationsignificative du pourcentage des lymphocytes CD4+ (jusqu’à 15 °/), CD8+ (jusqu’à 12 °/) et TCR)118+(jusqu’à 6 °/) dans les leucocytes intraépithéliaux du duodénum du quatrième au huitième jour suiv-ant l’inoculation, de même qu’une augmentation des cellules IgM+ (12 %) au huitième jour. Paral-lèlement, le pourcentage de lymphocytes CD8+ chute significativement dans la rate et le sang (-5 à 10 %)du quatrième au sixième jour suivant l’inoculation, puis s’accroît avec le pourcentage de lymphocytesCD4+ au huitième jour. Par comparaison, l’infection par E tenella induit une augmentation du pourcentagedes lymphocytes CD4+ au huitième jour (20°l) et CDB+ du sixième au huitième jour suivant l’inocula-tion (10%) dans les leucocytes intraépithéliaux du caecum. Un effet négatif ou pas d’effet n’est observésur le pourcentage des cellules TCRylô+ ou IgM+, Parallèlement, le pourcentage de lymphocytesCD4+ chute dans la rate au huitième jour (-10 %) et celui des lymphocytes CD8+ chute dans le sangau sixième jour suivant l’inoculation (-15 % En conclusion, l’infection par Eimeria modifie rapide-ment et fortement la proportion des sous-populations T dans les lymphocytes intraépithéliaux du lieudu développement parasitaire, obtenus par une technique d’isolement. Elle s’accompagne d’une mod-ification, en général inverse, de ces mêmes populations au niveau systémique (sang et rate). Lachronologie de ces modifications suit les phases du cycle parasitaire (période prépatente).
pouletlEimeria acervulina /Eimeria tenella / leucocyte intraépithéliallimmunité
INTRODUCTION
Avian coccidiosis in chickens is caused byintracellular protozoan parasites from sevenspecies of the genus Eimeria (E acervulina,E tenella, E praecox, E maxima, E brunetfi,E mitis and E necatrix) that develop in theintestinal epithelium. Host responses to coc-cidian parasites are complex and involveboth humoral and cellular mechanisms.
Studies on the protective immunity of chick-ens to coccidiosis underline the preponder-ant role of cell-mediated immune responses(Rose and Hesketh, 1982; Lillehoj, 1987;Lillehoj and Trout, 1993). Nevertheless, thepaucity of chicken immune reagents avail-able does not permit a clear understandingof the immune reactions and cytokine net-work such as is possible in mammals. Abetter understanding of the immune mech-anisms and the cells involved in the pro-tection against coccidiosis is necessary asa crucial step toward the development ofadapted vaccines, especially those using
recombinant proteins developed usinggenetic engineering.
Sporozoites enter the intestinal mucosaby penetrating villus epithelial cells, but thencomplete the first generation schizogony inthe crypt epithelial cells, with the exceptionof E brunetti and E praecox. During theirmigration through the mucosa, the sporo-zoites encounter cells from the immune sys-tem. Attention has focused in particular onthe possible involvement of cells from theimmune system such as T-lymphocytes andmacrophages in possibly transporting thesporozoites (Lawn and Rose, 1982; Lille-hoj and Chung, 1992; Trout and Lillehoj,1993; Vervelde et al, 1995).
Host immune responses are triggeredduring the sporozoite migration as the par-asite cycle progresses, and a strong pro-tective immunity generally takes place aftera primary infection (for a review, see Shirley,1992). Intraepithelial (IEL) and lamina pro-pria (LPL) leucocytes are the first line ofdefence in the intestine. Little information

is available as yet on the chicken intestinalimmune system. But the phenotypic char-acterization of cells from the chicken intesti-
nal immune system reveals an organizationsimilar to that in mammals (Bucy et al, 1988;Chai and Lillehoj, 1988; HoggenmOeller etal, 1993).
The purpose of our study was to followthe changes in T-cell subsets in IEL follow-ing a primary infection with Eimeria at thesite of parasite multiplication. Moreover, thegeneral impact of infection was also stud-ied on the systemic immune system (bloodand spleen) and other areas of the intes-tine. A comparison was performed betweentwo species that localize in two distinct areasof the intestine: E acervulina, which devel-
ops in the duodenum, and E tenella which
develops in the caeca.
MATERIALS AND METHODS
Chickens and parasites
Male White Leghorn chickens of histocompatibleline (GB1) were hatched at INRA, Nouzilly, andraised on wire-floored cages under specificpathogen free (SPF) conditions until use. Feedand water were provided ad libitum. At eightweeks of age, they were randomly distributedinto treatment groups according to the experi-mental design. Two species of Eimeria were used:E acervulina (strain PAPa46) and E tenella (strainPAPt36). These two strains were maintained bysuccessive passages in vivo.
Cell preparation
Blood cells
Blood was removed by intracardiac puncture.After an incubation step (45 min, 37 °C), the bloodwas centrifuged at low speed (5 min, 45 g).Plasma was removed and the cells were con-
centrated after further centrifugation (10 min,400 g). Viable cells were numbered in the pres-ence of trypan blue.
Spleen cells
The spleens were removed and gently teasedthrough a 40 pm-mesh steel screen in 1.1 equivPBS (phosphate buffer saline, pH 7.4, Gibco).The cells were collected after centrifugation(20 min, 210 g), then resuspended in 1.6 mL PBS.Viable cells were numbered in the presence of
trypan blue.
lntraepithelial leukocytes
The preparation of intraepithelial leukocytes isbased on a modification of a previously describedtechnique (Chai and Lillehoj, 1988). The duode-num (c loop, 12-15 cm) and the caeca werewashed in a HBSS medium (Hank’s BalancedSalt Solution, Gibco) supplemented with glucose(4 g/L) and foetal calf serum (FCS, 2%, Gibco)(medium HBSS1). The intestinal portions werethen cut in HBSS1 medium and incubated for
10 min at room temperature in HBSS1 containingdithiothreitol (DTT, 2 mM, Gibco). This step elim-inates intestinal mucus. The supernatant was dis-carded and the small pieces of intestine wereincubated twice for 20 min at 41 °C in HBSS1 1
containing 2 mM DTT and 3 mM EDTA (ethylene-diaminetetraacetic acid, Gibco). The supernatantwas centrifuged for 2 min at 45 g. The mediumcontaining the leukocytes was passed through aglass-wool column. This step eliminates mostepithelial cells and cellular clusters. The cellswere then centrifuged on Ficoll (d= 1.077, 30 min,490 g) to remove red cells. The intraepithelialleukocytes were washed several times andcounted.
Flow cytometric analysis
Spleen or blood cells (2 x 106) and 20 000-800 000 intestinal cells were centrifuged for 3 minat 400 g. The supernatant was discarded and100 pL monoclonal antibody was added to thecells. The monoclonal antibodies used to mark
the chicken T-lymphocytes recognize the CD4or CD8 (Chan et al, 1988) or TCR7/6 (Bucy et al,1988) cell surface antigen and were kindly pro-vided by MD Cooper and CL Chen (Departmentsof Medicine, Pediatrics and Microbiology, Uni-versity of Alabama at Birmingham, AL). Themonoclonal antibody which recognizes class 11
molecules at the surface of B-lymphocytes, acti-vated T-lymphocytes and macrophages (Guille-

mot et al, 1986) was kindly provided by N LeDouarin (Collbge de France and CNRS, Institutd’embryologie, Nogent s/Marne, France). Themonoclonal anti-IgM antibody was purchasedfrom Janssen (Noisy-Le-Grand, France). The 9D4monoclonal antibody, kindly provided by F Coud-ert (Laboratoire de virologie et d’oncologie avi-aires, INRA, Tours-Nouzilly, France), stains byfluorescence 90% thymus cells, 45-50% bursacells, 90% blood cells (obtained by low speedcentrifugation) and 70% spleen cells, and is usedas a pan-lymphocyte marker. After an incubationstep (30 min, 4 °C) and two washes in PBS with2% FCS, a goat polyclonal antibody directedagainst mouse IgG was added. For the blood andspleen cells it was conjugated with fluorescein(used at a dilution of 1:40, Nordic) and for theintestinal leukocytes it was conjugated with phy-coerythrin (used at a dilution of 1:20, Caltag).After another incubation step, 100 pL of 1%paraformaldehyde was added to fix the cells. Thefluorescence analysis was performed with a FACSStar Plus (Becton Dickinson).
Experimental design
Two groups of chickens were used: a control
group (four chickens) and an infected group (eightchickens). The chickens were infected with 105oocysts of E acervulina or 2 x 104 oocysts ofE tenella. The blood, spleen, duodenum andcaeca were removed 2, 4, 6, 8 and 14 days after
infection. For practical reasons, a first experimentwas conducted for the analysis of the blood andspleen lymphocytes, and a second one for theanalysis of the duodenal and caecal lymphocytes.The results are expressed as:
% = % infected group - % control group
Statistical analysis
The Student’s ‘t’test was used to compare thevalues obtained in the control group with the val-ues obtained in the infected group.
RESULTS
Phenotypic characterization of IELpopulations obtained from duodenumor caecum of GB1-histocompatiblechickens
In the duodenum, as in the caecum, the per-centage of CD8+ cells was higher than thepercentage of CD4+ cells. We found a lower
percentage of TCRy/8+ cells in the duode-num than in the caecum. The percentageof [gM+ cells in the caecum was higher than
the percentage of the same cells in the duo-denum. For the class I I+ cells, the percent-age in the duodenum was about the same
as the percentage in the caecum. Theseresults are summarized in table I.
Changes in the total number of IELobtained from the duodenum
or caecum during infection with Eimeria
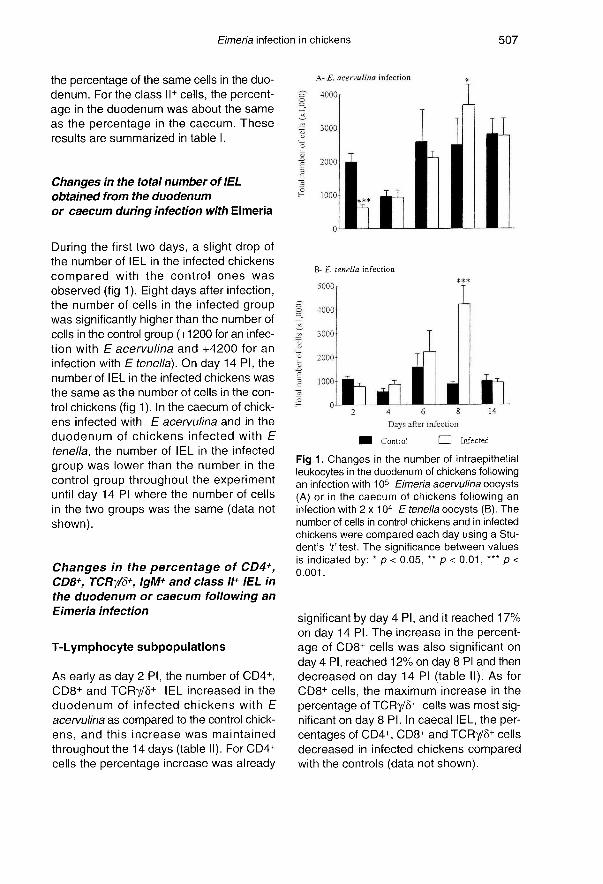
During the first two days, a slight drop ofthe number of IEL in the infected chickens
compared with the control ones wasobserved (fig 1 Eight days after infection,the number of cells in the infected groupwas significantly higher than the number ofcells in the control group (+1200 for an infec-tion with E acervulina and +4200 for an
infection with E tenella). On day 14 PI, thenumber of IEL in the infected chickens was
the same as the number of cells in the con-
trol chickens (fig 1 ). In the caecum of chick-ens infected with E acervulina and in the
duodenum of chickens infected with E
tenella, the number of IEL in the infected
group was lower than the number in the
control group throughout the experimentuntil day 14 PI where the number of cellsin the two groups was the same (data notshown).
Changes in the percentage of CD4+,CD8+, TCRy/6+, IgM+ and class I/+ IEL in
the duodenum or caecum following anEimeria infection
T-Lymphocyte subpopulations
As early as day 2 PI, the number of CD4+,CD8+ and TCRy/8+ IEL increased in the
duodenum of infected chickens with E
acervulina as compared to the control chick-ens, and this increase was maintained
throughout the 14 days (table II). For CD4+cells the percentage increase was already
significant by day 4 PI, and it reached 17%on day 14 PI. The increase in the percent-age of CD8+ cells was also significant onday 4 PI, reached 12% on day 8 PI and thendecreased on day 14 PI (table II). As forCD8+ cells, the maximum increase in the
percentage of TCRy/8+ cells was most sig-nificant on day 8 PI. In caecal IEL, the per-centages of CD4+, CD8+ and TCRy/8+ cells
decreased in infected chickens comparedwith the controls (data not shown).
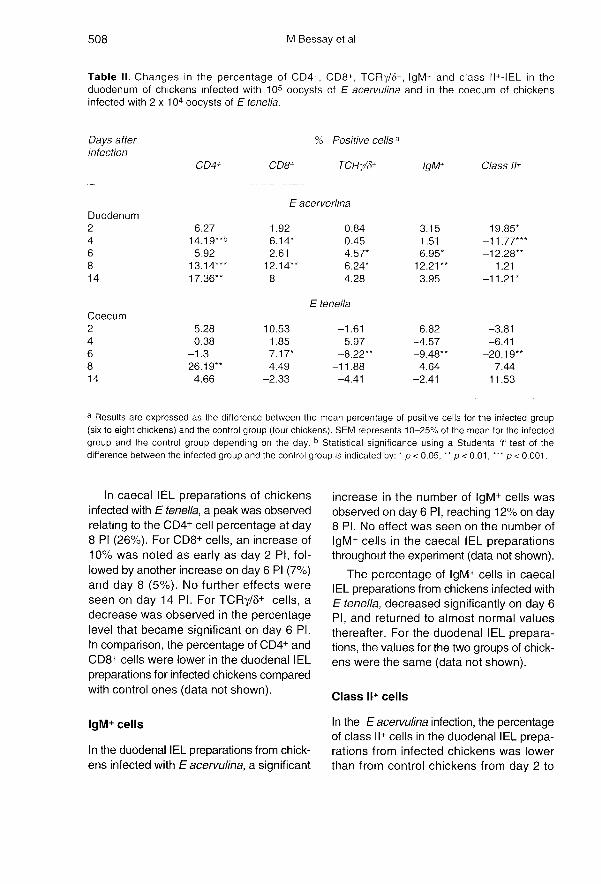
In caecal IEL preparations of chickensinfected with E tenella, a peak was observedrelating to the CD4+ cell percentage at day8 PI (26%). For CD8+ cells, an increase of10% was noted as early as day 2 PI, fol-lowed by another increase on day 6 Pi (7%)and day 8 (5%). No further effects wereseen on day 14 PI. For TCRy/8+ cells, adecrease was observed in the percentagelevel that became significant on day 6 PI.In comparison, the percentage of CD4+ and
CD8+ cells were lower in the duodenal IEL
preparations for infected chickens comparedwith control ones (data not shown).
IgM+ cells
In the duodenal IEL preparations from chick-ens infected with Eacervulina, a significant
increase in the number of IgM+ cells wasobserved on day 6 PI, reaching 12% on day8 PI. No effect was seen on the number of
IgM+ cells in the caecal IEL preparationsthroughout the experiment (data not shown).
The percentage of IgM+ cells in caecalIEL preparations from chickens infected withE tenella, decreased significantly on day 6Pi, and returned to almost normal valuesthereafter. For the duodenal IEL prepara-tions, the values for the two groups of chick-ens were the same (data not shown).
Class 11+ cells
In the E acervulina infection, the percentageof class II+ cells in the duodenal IEL prepa-rations from infected chickens was lowerthan from control chickens from day 2 to

day 6 PI, returned to normal levels on day 8PI and decreased again on day 14 PI. Noeffect was seen on the caecal IEL prepara-tions (data not shown).
After E tenella infection (table II), the per-centage of class ll+ cells was also lower incaecal IEL preparations from infected chick-ens from day 2 to day 6 PI and increased ondays 8 and 14. No effect was seen on theduodenal IEL preparations (data not shown).
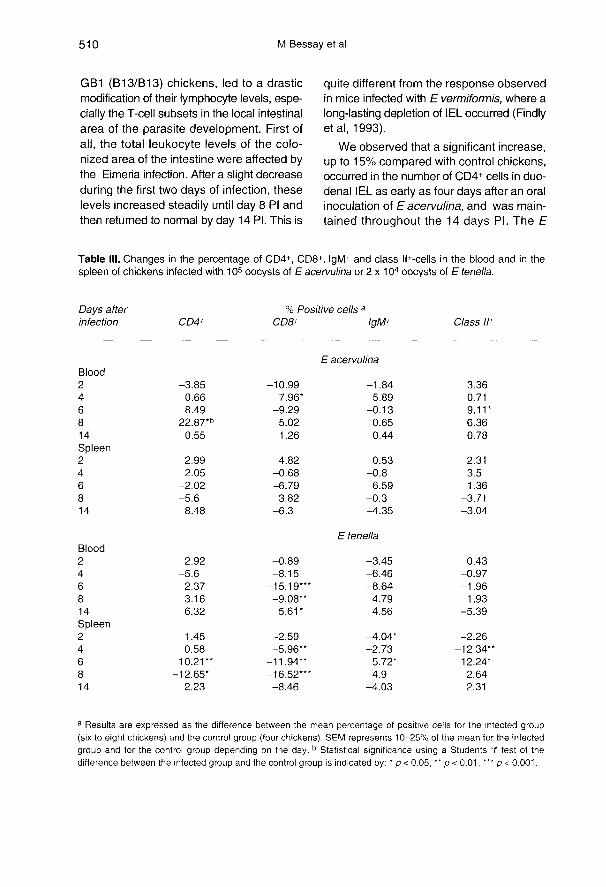
Effect of Eimeria infection
on the percentage of CD4+, CD8+, IgM+and class 11+ cells in blood and spleen
In the blood of chickens infected withE acervulina a peak in the level of CD4+lymphocytes was observed on day 8 Picompared to control (table 111). For the otherdays, the difference between the percent-age of cells in infected chickens and in con-trol chickens was not significant. For thelevels of CD8+ lymphocytes and IgM+ cells,the differences between infected and controlchickens were not significant except on day4 PI for the former. For the class II+ cells, anincrease was observed on day 6 PI. No sig-nificant effect was observed in the spleen,whatever the monoclonal antibody used.
In the blood of chickens infected with E
tenella, there were no significant differencesbetween infected and control chickens for
CD4+, IgM+ or class 11+ cell levels. For CD8+
lymphocytes, the minimal values wereobtained on days 6 and 8 Pi. On day 14 PI,the percentage of CD8+ cells was higher ininfected chickens than in control chickens. In
the spleen, an increase in CD4+ cells wasobtained on day 6 PI, followed by adecrease on day 8 Pl. For CD8+ cells, the
percentage in the infected group was lowerthan in the control group throughout theexperiment. No great variation in IgM+ cell
percentage was noted between the two
groups of chickens. On day 4 PI there wasa decrease in the percentage of class II+
cells in the infected chickens compared tothe control chickens. However, on day 8 PI,the percentage of these cells was higher forthe infected chickens than for the controls.
DISCUSSION
Because avian coccidia develop in theepithelial cells from the intestinal mucosa,the first line of host defence is constituted bythe leukocytes from this area, either locatedwithin the epithelium or in the lamina pro-pria. As in mammals, the majority of chickenIEL are T-lymphocytes (Arnaud-Battandieret al, 1980; Bucy et al, 1988; Vervelde andJeurissen, 1993). They are therefore capa-ble of initiating a cell-mediated immuneresponse following coccidia infection. Stud-ies on the migration of sporozoites in naivechickens have shown that they are firstdetected in the intestinal villus epitheliumafter the infection, then in the lamina pro-pria, and subsequently accumulate in thecrypt epithelium where they multiply(Vervelde et al, 1995). Leukocytes havebeen suggested to have a role in the trans-port of sporozoites to the crypt. Sporozoiteshave been identified inside such cells as
macrophages (Van Doorninck and Becker,1957; Trout and Lillehoj, 1993) or T-lym-phocytes (Vervelde et al, 1995), mainly ofthe CD8 phenotype (Trout and Lillehoj,1993, 1995).
In our study, we chose to test the effect ofan oral Eimeria infection on the intestinalimmune system, by considering isolated IELobtained after the dissociation of the intesti-
nal epithelium (Chai and Lillehoj, 1988;Schwager and Weber, 1992; HoggenmOelleret al, 1993). A final aim would be to anal-yse their local role in immunological func-tion studies.
Our results showed that a primary high-dose infection with Eimeria (100 000 oocystsfor E acervulina and 20 000 oocysts forE tenella) in two-month-old histocompatible
GB1 (B13/B13) chickens, led to a drasticmodification of their lymphocyte levels, espe-cially the T-cell subsets in the local intestinalarea of the parasite development. First ofall, the total leukocyte levels of the colo-nized area of the intestine were affected bythe Eimeria infection. After a slight decreaseduring the first two days of infection, theselevels increased steadily until day 8 PI andthen returned to normal by day 14 PI. This is
quite different from the response observedin mice infected with E vermiformis, where a
long-lasting depletion of IEL occurred (Findlyet al, 1993).We observed that a significant increase,
up to 15% compared with control chickens,occurred in the number of CD4+ cells in duo-
denal IEL as early as four days after an oralinoculation of E acervulina, and was main-tained throughout the 14 days PI. The E

acervulina infection induced a smallerincrease in the number of CD8+ cells, alsobeginning on day 4 Pi and culminating onday 8 Pi (up to 12%), and followed by adecrease as early as 14 days PI. The pat-tern of variation for TCRy/8+ cells in thiscase followed the pattern of CD8+ cells, witha significant increase in numbers reaching6% on day 8 PI, and a return to normal val-ues thereafter. When compared with thefew studies present in literature, the per-centage of CD8+ cells we obtained for con-trol chickens by isolating IEL (27%), wasless than that of Lillehoj and Bacon (1991)and Lillehoj (1994), 37-53%, but the per-centage of CD4+ cells was a little higher,17% compared to 10%. Nevertheless, thenumber of TCRy/8+ cells we found in thecontrol duodenum, about 10% of the totalIEL, was far lower than values in the litera-ture, 33-45% (Bucy et al, 1988; Lillehoj andChung, 1992). Literature on the subject hasunderlined the great variability in the per-centage of CD4+, CD8+ and TCRy/8+ cells inintestines. These differences are due to the
ages of the chickens or genetic differences(Bucy et ai, 1988; Lillehoj and Chung, 1992;Lillehoj, 1994). To sum up, compared withthe study of Lillehoj (1994), after a primaryE acervulina infection, the increase weobserved in the GB1 line in duodenal IELfor CD4+ cells followed the amplitude andthe pattern of the TK line, which is very sen-sitive to coccidiosis, but the one for CD8+cells and TCRy/8+ cells followed the ampli-tude and the pattern of the SC line, which ismore resistant to coccidiosis and in any casehas a maximum on day 7 Pl. So, for the pre-sent, it remains difficult to connect a phe-notype of resistance to the triggering of spe-cific intestinal T-cell subsets for E acervulina.
The primary oral infection with E tenella,led to somewhat different results regard-ing the variation in the percentage of CD4+,CD8+ and TCR7/6+ cells in the caecal IEL.The increase in CD8+ cells appeared ear-lier, at day 2 PI, then another peak (6-10%)was observed from days 6 to 8 Pl. No
increase in the level of TCRy/8+ cells wasnoted throughout the 14 days of the exper-iment. A large increase in the number ofCD4+ cells (up to 25%) occurred on day 8PI, followed by a severe decrease on day14 PI. The percentages of CD4+ cells (26%)and CD8+ cells (37%) were slightly higherthan those found in the literature for con-trol chickens (Lillehoj and Chung, 1992 ;Lillehoj, 1994). However the percentage ofTCRy/8+ cells (21 %) was lower. Our resultsare in agreement with those available inthe literature for E tenella infection usingthe same isolation technique (Schwagerand Weber, 1992). These authorsobserved an increase in the number ofCD4+ cells on days 5 to 6 PI, but no changefor CD8+ cells.
For other cells such as IgM+ cells, weobserved a rather low number in the duo-denal IEL (7%). This was a little higher inthe caecum (15%). Immunohistological stud-ies on chicken intestine underline the
scarcity of B-cells in epithelium (Jeurissen etal, 1989). The presence of some IgM+ cells
in our preparation could be a sign of possi-ble contamination with cells from the lam-ina propria. HoggenmOeller et al (1993)found 4-5% plasma cells in isolated IELfrom the small intestine using the same tech-nique. Others did not determine the num-ber of IgM+ cells in their duodenal IEL prepa-rations (Lillehoj, 1994). Nevertheless theCD4+/CD8+ cell ratio in our duodenal IELis in agreement with the literature (Bucy etal, 1988). No datum is available on isolatedcaecal IEL. After an Eacervulina infection,the number of IgM+ cells increased signifi-cantly by days 6 to 8 PI in the duodenal IELpreparations. But no such increase wasnoted after an oral E tenella infection in the
caecal IEL up to 14 days Pi. Moreover, noincrease in the number of class I I+ cells wasnoted after an oral E acervulina infection induodenal IEL throughout the 14 days Pi,but an increase was observed, in caecalIEL number after an oral E tenella infection,as seen by Schwager and Weber (1992).

Using a different technique, such asimmunohistochemical analysis studying thevariation of T-cell subsets ’in situ’ at the localsite of parasite multiplication, gave ratherdifferent results. Trout and Lillehoj (1995)found that an E acervulina infection inducedan increase in CD8+ cells in the duodenal
lamina propria as soon as 24 h PI but had lit-tle effect in the epithelium, and then veryfew changes occurred up to 72 h Pl. FewCD4+ cells were observed in the lamina pro-pria. On the other hand, Rothwell et al(1995) showed that an oral E maxima in thejejunum induced a very high accumulation ofCD4+ cells and TCRa/(3+ cells specificallyin the lamina propria concomitantly with anincrease of CD8+ cells and TCRy/6+ cellsin the epithelium 10 days PI (ie, at the end ofthe prepatency for this coccidia). In theirwork the IgM+ cells exhibited an earlyincrease in the days just following infection,and then another ten days later.
It is rather difficult at present to draw ageneral picture of which modifications of theimmune system appeared in the intestine,especially at the local area of parasite mul-tiplication, as a result of a primary infectionwith avian coccidia. The literature points todiscrepancies among authors, first of all dueto the different techniques used (isolationprocedures to obtain leukocytes from theintestinal epithelium or immunohistochemi-cal studies, but also due to the kind of chick-ens used, their genetic origin and age, andthe doses of coccidia used (high or low).Immune modifications taking place after aprimary infection are quite different fromthose following a second inoculation (Troutand Lillehoj, 1995; Vervelde et al, 1995;Rothwell et al, 1995). As a first line ofdefence against coccidiosis, NK cells (Chaiand Lillehoj, 1988; Lillehoj, 1989) andTCRy/8+ cells (Trout and Lillehoj, 1995;Rothwell et ai, 1995) have been consideredto be important immediately after the para-sitic invasion, as for the case of some intesti-nal parasites in mammals. For E acervulina
in isolated duodenal IEL, we found a parallelincrease in CD8+ cells and TCRy/8+ cellsfrom the middle to the end of the prepatentperiod, similar to what was observed injejunum epithelium for E maxima (Rothwellet al, 1995). No effect on TCRy/8+ cells wasnoted for E tenella in coecal IEL, despite aslight increase of CD8+ cells. This mightreflect a possible difference in the earlyimmune response involved according to theparasitized area, ie, the upper or lower partof the intestinal tract, but this hypothesisneeds confirmation. Moreover our results
place particular emphasis on CD4+ cells;during infection these seemed to be stronglytriggered at the local areas of parasite mul-tiplication, the maximum levels of these cellsbeing observed in IEL around the end ofthe prepatent period (4.5-5 days for Eacervulina and 6-8 days for E tenella). Thiscorroborates the observation of Rothwell etal (1995) with E maxima, but in lamina pro-pria. The end of the prepatent period coin-cides with oocystal excretion and the estab-lishment of immunity, greatly limiting furtherparasite multiplication.
Due to the lack of data in the literature,we were particularly interested to follow thevariations caused by the primary infectionwith Eimeria in chickens, in the T-cell sub-sets in other compartments of the immunesystem, such as an intestinal area differentfrom that colonized by the parasite, or thesystemic compartments (blood and spleen).In every case, the infection with Eimeriainduced a significant decrease in all T cell-subsets in the caecum for E acervulina andin the duodenum for E tenella (data notshown) corresponding to the prepatentperiod. At the same time, the number of sys-temic CD8+ cells was particularly depressedduring the four days preceding the end ofthe prepatent period for each Eimeria. Thiswas seen in blood for E acervulina, wheninfected chickens increased their duodenalCD8+ IEL, but even more in blood andspleen for E tenella despite the lower effect

of infection on coecal CD8+ IEL. Consider-
ing CD4+ cells, their number was firstdepressed and then increased around theprepatent period (significantly for Eacervulina at day 8 PI). The same resultswere obtained by Schwager and Weber(1992) with E tenella. It is noteworthy thatthe appearance of a systemic cell-mediatedimmune response, revealed by antigen-spe-cific proliferation studies (Lillehoj, 1986; Mar-tin et al, 1993, 1994), takes place as soon as7 days Pi for E acervulina and E tenella.Moreover the local antibody immuneresponse is significant as soon as 7 daysPi for E tenella (Zigterman et al, 1993) andE acervulina (personal observation) in theintestinal segments colonized by the para-site, but not in the other intestinal areas.This demonstrates an already high andeffective stimulation of the immune system,involving the stimulation of CD4+ cells for
antigen-specific lymphocyte proliferationand the antibody production. In fact, studiesin mice underline the essential role of CD4+cells in controlling a primary E vermiformisinfection (Rose et al, 1988, 1992). Com-pared with several histocompatible chickenlines, the GB1 line interestingly exhibitedthe highest specific antibody response inserum following an E tenella infection, con-comitant with the lowest oocyst production,despite showing the highest sensitivity tothe disease (M Naciri, personal communi-cation).
In conclusion we observed, with two dif-ferent Eimeria located at distinct intestinal
areas, duodenum and coeca, the same gen-eral pattern of lymphocyte changes follow-ing a primary infection of GB1 chickens. Atthe local area of the parasitic multiplication,T-lymphocytes seemed to be triggeredquickly during the middle to end of theprepatent period as was shown by theincrease in the total number of leukocytes,a little earlier for CD8+ cells and then espe-cially for CD4+ cells. In other intestinal areas,this positive effect was concomitant with (or
somewhat preceded by in the systemic com-partment), a negative effect seen on theCD4+ and especially CD8+ T-cell subsets.So the local changes in lymphocyte sub-sets in the intestinal area where the Eimeria
multiplication takes place are probably dueto a powerful local triggering of lymphocytestimulation and proliferation, but the imme-diate participation of cells from other intesti-nal areas, and also from the systemic com-partment, seems to be significant in the caseof a high level of infection. Further studies ofthe lymphocyte migration pattern are nec-essary to clarify this point.
REFERENCES
Arnaud-Battandier F, Lawrence EC, Blaese RM (1980)Lymphoid population of gut mucosa in chickens.Digest Dis 25, 252-259
Bucy RP, Chen CLH, Cihak J, L6sch U, Cooper MD(1988) Avian T cells expressing gamma delta recep-tor localize in the splenic sinusoids and the intestinalepithelium. J Immunol 141, 2200-2205
Chai JY, Lillehoj HS (1988) Isolation and functional char-acterization of chicken intestinal intra-epithelial lym-phocytes showing natural killer cell activity againsttumor target cells. Immunology 63, 111-117 7
Chan MM, Chen CLH, Ager LL, Cooper MD (1988) Iden-tification of the avian homologues of mammalianCD4 and CD8 antigens. J Immunol 140, 2133-2138
Findly RC, Roberts SJ, Hayday AC (1993) Dynamicresponse of murine gut intraepithelial T cells afterinfection by the coccidian parasite Eimeria. Eur J/mmuno/23, 2557-2564
Guillemot FP, Turmel P, Charron D, Le Douarin N, Auf-fray C (1986) Structure, biosynthesis and polymor-phism of chicken MHC Class II (B-L) antigens andassociated molecules. J lmmunol 137, 1251-1257
HoggenmUeller L, Wakenell PS, Schat KA (1993) Prepa-ration and characterization of chicken intraepitheliallymphocytes. Avian Pathol22, 509-523
Jeurissen SHM, Janse EM, Koch G, de Boer GF (1989)Postnatal development of mucosa-associated lym-phoid tissues in chickens. Cell Tissue Res 258, 119-124
Lawn AM, Rose ME (1982) Mucosal transport of Eime-ria tenella in the caecum of the chicken. J Parasitol68, 1117-1123
Lillehoj HS (1986) Immune response during coccidio-sis in SC and FP chickens I. In vitro assessment ofT-cell proliferation response to stage-specific para-

site antigens. Vet Immunol Immunopathol 13, 321-330
Lillehoj HS (1987) Effects of immunosuppression onavian coccidiosis: cyclosporin A but not hormonalbursectomy abrogates host protective immunity.Infect Immun 55, 1616-1621 1
Lillehoj HS (1989) Intestinal intraepithelial and splenicnatural killer cell responses to eimerian infections ininbred chickens. Infect Immun 57, 1879-1884
Lillehoj HS (1994) Analysis of Eimeria acervulinainduced changes in the intestinal T lymphocyte sub-populations in two chicken strains showing differ-ent levels of susceptibility to coccidiosis. Res VetSci 56, 1-7
Lillehoj HS, Bacon LD (1991) Increase of intestinalintraepithelial lymphocytes expressing CD8 antigenfollowing challenge infection with Eimeria acervulina.Avian Dis 35, 294-301 1
Lillehoj HS, Chung KS (1992) Postnatal developmentof T-lymphocyte subpopulations in the intestinalintraepithelium and lamina propria in chickens. VetImmunol Immunopathol 31, 347-360
Lillehoj HS, Trout JM, (1993) Coccidia: a review of recentadvances on immunity and vaccine development.Avian Pathol22, 3-21
Martin A, Lillehoj HS, Kaspers B, Bacon LD (1994) Anti-gen-specific T-cell proliferation following coccidiainfection. Poult Sci72, 2084-2094
Martin A, Lillehoj HS, Kaspers B, Bacon LD (1994) Mito-gen-induced lymphocyte proliferation and interferonproduction following coccidia infection. Avian Dis 38,262-268
Rose ME, Hesketh P (1982) Immunity to coccidia inchickens: adoptive transfer with peripheral bloodlymphocytes and spleen cells. Parasite Immuno154,171-185
Rose ME, Joysey HS, Hesketh P, Grencis RK, WakelinD (1988) Mediation of immunity to Eimeria vermi-formis in mice by L3T4+ T cells. Infect Immun 56,1760-1765
Rose ME, Hesketh P, Wakelin D (1992) Immune controlof murine coccidiosis: CD4+ and CD8+ T lympho-cytes contribute differentially in resistance to primaryand secondary infections. Parasitology 105, 349-354
Rothwell L, Graminski RA, Rose ME, Kaiser P (1995)Avian coccidiosis: changes in intestinal lymphocytepopulat’rons associated with the development of immu-nity to Eimeria maxima. Parasite Immunol 17, 525-533
Shirley MW (1992) Research on avian coccidia: anupdate. Br Vet J 148, 479-499
Schwager J, Weber G (1992) Immune response to Eime-ria tenella : analysis of cellular and molecular param-eters. In: Proceedings 4th Conference COST 89,Tours-INRA, 4-7 October, 1992 (P Loudert, ed)(abstract), 5
Trout JM, Lillehoj HS (1993) Evidence of a role for intesti-nal CD8+ lymphocytes and macrophages in transportof Eimeria acervulina sporozoites. J Parasitol79,790-792
Trout JM, Lillehoj HS (1995) Eimeria acervulina infection:evidence for the involvement of CD8+ lymphocytesin sporozoite transport and host protection. Poult Sci74, 1117-1125
Van Doorninck WM, Becker ER (1957) Transport ofsporozoite of Eimeria necatrix in macrophage. J Par-asitol43, 40-44
Vervelde L, Jeurissen SHM (1993) Postnatal develop-ment of intra-epithelial leukocytes in the chickendigestive tract: phenotypical characterization in situ.Cell Tissue Res 274, 295-301
Vervelde L, Vermeulen AN, Jeurissen SHM (1995) Eime-ria tenella: Sporozoites rarely enter leukocytes inthe cecal epithelium of the chicken (Gallus domes-ticus). Exp Paras/fo/81, 29-38
Zigterman GJWJ, van de Ven W, van Geffen C, LoeffenAHC, Panhuijzen JHM, Rijke EO, Vermeulen AN(1993) Detection of mucosal immune responses inchickens after immunization or infection. Vet Immunol
Immunopathol36, 281-291





























