Embed Size (px)
Citation preview

CD46 and Dlg4 interact. 1
1
A functional interaction between CD46 and Dlg4: a role for Dlg4 in epithelial
polarization.
1Mandy J. Ludford-Menting, 1Suzanne J. Thomas, Blessing Crimeen, 2Lisa J. Harris, 2Bruce
E. Loveland, 3Margaret Bills, Sarah Ellis, 4Sarah M. Russell
1These authors contributed equally to the work.
Peter MacCallum Cancer Institute, Trescowthick Research Laboratories, St Andrew’s Place,
East Melbourne, Victoria 3002, Australia.
2Austin Research Institute, Studley Rd, Heidelberg, Victoria 3084, Australia.
3Current address: St. Vincent's Institute of Medical Research, 9 Princes St, Fitzroy, Vic 3065,
Australia.
4To whom correspondence should be addressed at:
Peter MacCallum Cancer Institute, Trescowthick Research Laboratories, St Andrew’s Place,
East Melbourne, Victoria 3002, Australia.
Tel 613 9656 3727, Fax 613 9656 1411
E-mail: [email protected]
Running Title:
CD46 and Dlg4 interact.
Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on November 19, 2001 as Manuscript M108479200 by guest on M
ay 28, 2018http://w
ww
.jbc.org/D
ownloaded from

CD46 and Dlg4 interact. 2
2
SUMMARY
Using a Yeast Two Hybrid screen, we identified a physical interaction between CD46
and Dlg4. CD46 is a ubiquitous human cell surface receptor for the complement
components C3b and C4b, and for measles virus and human herpes virus 6. Dlg4 is a
scaffold protein important for neuronal signaling, and is homologous to the Drosophila
tumor suppressor, Dlg. We show that an interaction between CD46 and Dlg4 is
important for polarization in epithelial cells. Specifically we show (i) biochemical
evidence for an interaction between CD46 and Dlg4, (ii) that this interaction is specific
for the Cyt1 and not the Cyt2 domain of CD46, (iii) that both CD46 and an alternatively
spliced isoform of Dlg4 are polarized in normal human epithelial cells, and (iv) that the
polarized expression of CD46 in epithelial cells requires the Dlg4 binding domain, and
alters with expression of a truncated form of Dlg4. This is the first identification of a
direct, and cytoplasmic domain-specific interaction between CD46 and an intracellular
signaling molecule, and provides a molecular mechanism for the polarization of CD46.
These data also indicate that, in addition to the known role for Dlg4 in neuronal cells,
Dlg4 may be important for polarization in epithelial cells.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 3
3
INTRODUCTION
CD46 (or Membrane Cofactor Protein) is a receptor for the complement
components C3b and C4b, and for several pathogens, including the measles virus and
human herpes virus 6 (1-5). CD46 is expressed on all human cells except erythrocytes,
and protects cells from complement-mediated destruction by acting as a cofactor for the
factor-I mediated breakdown of C3b and C4b (1, 6). The expression of CD46 is
upregulated on many tumor cells, and different alternatively spliced isoforms of CD46 are
expressed on both malignant cells and activated T cells (6). When expressed in the canine
epithelial cell line, MDCK, CD46 is polarized to the basolateral surface (7-9).
In addition to the effects of CD46 on the activity of complement, it has recently
become clear that signaling through CD46 can have important effects on both innate and
acquired immunity. For instance, signaling through CD46 can affect the production of
interleukin 12, macrophage inflammatory protein-1a, and nitric oxide by macrophages,
the production of interleukin 6 by astrocytes, IgE class switching in B cells, and
proliferation of T cells (10-16). CD46 is alternatively spliced to give two different
cytoplasmic domains (Cyt1 and Cyt2, Fig1 A), each of which is expressed in every cell
type examined to date (6). The cytoplasmic domains have different signaling
characteristics: (i) expression of Cyt1 causes a preferential increase in the production of
nitric oxide by macrophages (13), and (ii) only the Cyt2 domain can be phosphorylated on
tyrosine, and coprecipitates with a 38 kDa phosphorylated protein (17). The cytoplasmic
domains are only 28 and 35 amino acids long respectively, and are therefore likely to
signal via interactions with other intracellular proteins. CD46 has been
coimmunoprecipitated with SHP-1 in macrophages (18), and antibodies to CD46 can
cause the phosphorylation of p120cbl and LAT in T cells (15). These observations
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 4
4
describe cellular processes that can be affected by CD46, but as yet we cannot describe
the molecular pathways through which CD46 affects cell events. As a first step towards
elucidating these pathways, we sought to identify proteins that interact with the different
cytoplasmic domains by a Yeast Two-Hybrid screen.
We show here that the Cyt1 domain interacts with Dlg4, a member of the MAGUK
family of proteins that provides a scaffold for the assembly of signaling complexes. Dlg4,
also called Post-Synaptic Density (PSD)-95 or Synapse Associated Protein (SAP)-90, is one
of a family of four human proteins. This family shares a single homologue in Drosophila: a
tumor suppressor called Discs Large or Dlg (19, 20, 21). The Dlg proteins are thought to
have two primary functions: (i) to mediate the localization and clustering of receptors (22-
28), and (ii) to form a scaffold for the assembly of signaling proteins (27-36). Dlg proteins
have multiple protein-protein interaction motifs: three PDZ domains, an SH3 domain and a
guanylate kinase-like domain (GuK). By virtue of their PDZ domains, Dlg proteins are part
of a larger family of proteins that are polarized, and that recruit other proteins to a polarized
domain in neurons and epithelial cells (22, 37, 38). Thus, the interactions of PDZ domain-
containing proteins with each other, with membrane-bound receptors, and with the
cytoskeleton, serve to define and regulate polarization, and to coordinate signaling networks.
PDZ domains are 90 amino acid modules which classically interact with the C-terminal 4
residues of proteins with the sequence X-S/T-X-I/L/V. Dlg4 has previously been shown to
interact with receptors that are important for neuronal signaling, and is localized to the
postsynaptic density region of neurons. Results described herein indicate that Dlg4 interacts
with CD46, a protein not known to play a role in synaptic signaling. We show that the C-
terminal 4 residues of the CYT1 domain (FTSL) of CD46 interact with the PDZ3 domain of
Dlg4. We also show colocalization of CD46 and Dlg in epithelial tissues, and describe
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 5
5
evidence suggesting that this interaction is important for the polarization of CD46 in
epithelial cells.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 6
6
EXPERIMENTAL PROCEDURES
Yeast Two-Hybrid Screen. The sequence encoding the Cyt1 domain of CD46 was
fused in frame to the GAL4 binding domain (GAL4-BD) in the pAS2.1 vector (Clontech).
The vector was transformed into PJ09-4A yeast cells (39), selected on Trp-deficient medium,
and screened for expression of the bait by immunoblotting with an antibody to the GAL4-BD
(Santa Cruz). The yeast were further transformed with a library of the GAL4-AD fused to
cDNA from human peripheral blood lymphocytes activated with phytohaemagglutinin, and
selected in medium deficient in Trp, Leu and His with 3 mM 3-AT. Of 3 x 106 independent
clones screened, 112 colonies grew on both His-deficient and Ade-deficient medium. DNA
was extracted from each of these colonies and characterized by restriction digestion and
sequencing. DNA from one colony corresponded to a partial cDNA sequence of human
Dlg4.
To test for specificity, yeast were cotransformed with bait and prey vectors and selected on
plates deficient in Leu and Trp, then streaked onto both Ade- and His- deficient media and
assessed for growth. Control clones for these tests included GAL4-pLAM (Clontech) and
GAL4-ABP280 (40), and vectors expressing the GAL4-BD and GAL4-AD alone. At least 4
clones were tested for each transformation, and clones were analyzed by immunoblotting
with antibodies to the GAL4-AD and –BD respectively (Santa Cruz) to ensure equivalent
protein expression.
Cloning and construct derivation. The mutant constructs used in Yeast Two-
Hybrid specificity tests were derived from the original GAL4-BD-CYT1 fusion used in the
screen and the GAL4-AD-Dlg-4 fusion isolated from the screen. Mutations were made using
the Quick-Change Site-Directed Mutagenesis kit (Stratagene). The primer sequences were as
follows, with the underlined residue indicating a mutation from the original sequence:
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 7
7
GTAAAATTTACTTCTCGCTGAGAAGGAGAGATG and
CATCTCTCCTTCTCAGCGAGAAGTAAATTTTAC for the GAL4-BD-CYT1(L358R) and
for CD46-Cyt1L(358R), CAGCCGATTCGAGGCCTAGATCCACGACCTTCG and
CGAACCTCGTGGATCTAGGCCTCGAATCGGCTG for the GAL4-AD-Dlg4(trunc), and
CGAATTGTGATCCACGCGGGCTCCACGGGCCTG and
CAGGCCCGTGGAGCCCGCGTGGATCACAATTCG for the GAL4-AD-Dlg4mutPDZ3.
To generate an expression construct of Dlg4, the 5’ region was amplified from cDNA
from human spleen polyA+ RNA, and fused to the Dlg4 fragment isolated in the Yeast-Two
Hybrid screen using Splice Overlap Extension PCR. This splice variant of Dlg4 (which lacks
exon 20)( 44) was then subcloned into pcDNA3 (InVitrogen) and fully sequenced. The splice
variant of Dlg4 that encodes an intact GuK domain was created using an amplified GuK-
encoding domain from pooled human leukemic cells using RT-PCR. The PCR product
contained FseI and EcoRI restriction sites, which allowed insertion into the original
expression construct of Dlg4. Sequence coding for the PDZ3 domain of Dlg4 was amplified
from the original Yeast Two Hybrid clone and was fused in frame to the coding region of
EGFP in the EGFP-C2 vector (Clontech) using EcoR1 sites. GST fusion proteins were
created in the pGEX-4T-1 vector (Amersham) using CD46-Cyt1 and Cyt2 sequences
amplified from Gly 342 to beyond the stop codon.
Molecular Modelling. Molecular modelling of the peptide FTSL bound to PDZ3
domain was performed using the known X-ray crystallographic structure of PDZ3 of Dlg4 in
complex with its peptide ligand QTSV (41) (PDB entry 1BE9). Computer graphics program
“O” (42) was used to replace sidechains of QTSV to those of FTSL respectively; Cβ and
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 8
8
peptide mainchain atoms were unaltered. Using “O” rotamers, FTSL sidechains were
modelled for stereochemical fit and favorable contact with the PDZ domain.
Transfections. For transient expression, the canine kidney epithelial cell line,
MDCK or the human kidney epithelial cell line, 293, were transfected using FuGene6
(Roche), and analysed 2-3 days later. Stable transfectants of MDCK cells were selected in
G418 for resistance to neomycin, and then screened for expression of CD46 or Dlg4 by
immunofluorescence or flow cytometry. Antibodies used were K28/43 and K28/86.2 mouse
monoclonal antibodies to Dlg4 (Upstate Biotechnology). The 1839 polyclonal rabbit
antibody was raised against the recombinant extracellular domain of CD46. Clones were
generated by limiting dilution. To obtain double transfectants, cells were transfected with
CD46, selected, cloned, and then transfected with EGFP-PDZ3 and enriched by flow
cytometry.
Immunofluorescence and analysis of polarization. MDCK cells were grown to
confluency on polyester transwell membranes (24mm, pore size 0.4 µm). The apical and
basolateral regions of the cells were stained by incubation with primary and secondary
antibody in the inside of the insert (FITC) or the base of the well (Fluor594) respectively.
The membranes were removed from the insert with a scalpel blade, and sealed on a slide with
ProLong antifade. Immunofluorescence was analyzed using a Bio-Rad (Bio-Rad
Laboratories; Hercules, CA) confocal scanning microscope (model MRC1000). The 488-nm
line from a krypton/argon laser was used to excite the FITC-tagged secondary antibody and
emission was detected through the 522-nm filter. The 568-nm line from a krypton/argon
laser was used to excite the Fluo594-tagged secondary antibody and emission was detected
through the 605-nm filter. Optical sections were recorded at 0.4-um vertical steps with each
image averaged for seven scans. All images were processed using the Comos program (Bio-
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 9
9
Rad) and Cas 4.02 (confocal assistant software, copyright Todd Clark Brelje). Staining
patterns are representative of the staining observed in at least three clones of each construct
transfected, and staining with subclass control antibodies indicated that the staining was
specific.
Immunohistochemistry. Sections of human kidney were fixed in methanol at 37°C for 18
hours, blocked in 5 % BSA and immunolabelled in 0.8 % BSA with primary antibody and
then biotinylated swine anti-mouse/rabbit secondary antibody. Signal was amplified with
streptavidin peroxidase and visualized using 3,3’Diaminobenzadine (Dako, USA). Bound
antibody was detected with streptavidin-coupled peroxidase and 3,3’Diaminobenzadine
(Dako, USA). Sections were counterstained with haemotoxylin.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 10
10
RESULTS
CD46 interacts with Dlg4. We screened for proteins interacting with the cytoplasmic
domains of CD46 by a Yeast Two Hybrid assay, using a library derived from activated human
peripheral blood lymphocytes. Two separate “baits” were used in the screen, in which the
cytoplasmic domains of CD46-Cyt1 and CD46-Cyt2 (Fig1 A, Bi & Bii) were fused to the
GAL4 binding domain (GAL4-BD-Cyt1, Fig1 Biv) and GAL4-BD Cyt2, Fig1 Bvi). After
screening with the GAL4-BD-Cyt1 clone, a partial cDNA clone encoding Dlg4 (PSD-95) was
isolated (43). Full length Dlg4 includes 5 domains known to be important for protein-protein
interactions, namely 3 PDZ domains, one SH3 domain and a guanylate kinase like domain
(Fig1 Ci). The clone we isolated corresponded to a previously described alternatively spliced
variant (a deletion of exon 20, which causes a frame shift leading to a premature termination,
44) in which the GuK domain was truncated by 102 amino acids (termed Dlg4-DelGuK, Fig1
Cii). Protein encoded by the yeast clone (termed GAL4-AD-Dlg4, Fig1 Ciii) contained the
GAL4-AD fused in frame to the C-terminal 45 amino acids of PDZ2 (starting at nucleotide
1590 within the PDZ2 domain (43)), the complete PDZ3 domain, the SH3 domain and the N-
terminal part of the GuK domain. Thus, the only intact modules were the PDZ3 and the SH3
domains.
To map the domains of each protein that were important for the interaction, we compared the
growth of yeast transformed with CD46 and either the Dlg4 fusion from the Yeast Two
Hybrid screen (GAL4-AD-Dlg4), or mutants thereof (see Fig1 B & C). Cotransformation of
GAL4-BD-Cyt1 and GAL4-AD-Dlg4 allowed growth on selective media, indicating that
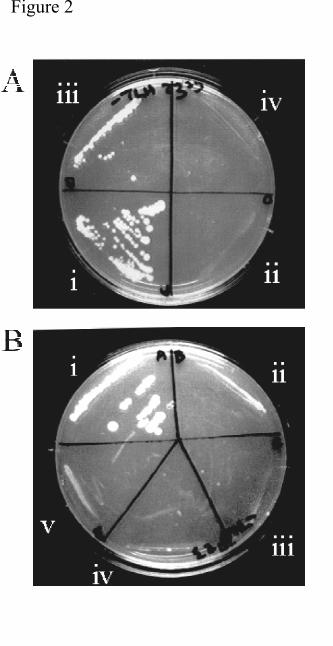
these proteins interact (Fig2, Ai). However, mutation of R318 adjacent to the GLGF domain
of PDZ3 (critical for binding to other C-terminal peptides (41), GAL4-AD-Dlg4mutPDZ3,
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 11
11
Fig1 Cv) completely abrogated growth, indicating that a functional PDZ domain was required
for interaction of the fusion protein (Fig2 Aii). Truncation of the Gal4-Dlg4 clone at the end
of PDZ3 (GAL4-AD-Dlg4(trunc)) allowed growth, although at a consistently reduced rate
(Fig2 Aiii). This clone contains only the C-terminal 45 amino acids of the PDZ2 domain and
the complete PDZ3 domain. Coexpression of GAL4-BD-Cyt1 with an irrelevant GAL4-AD
fusion protein (GAL4-AD-ABP280) did not confer growth (Fig2 Aiv), indicating that the
interactions between CD46 and Dlg4 fusion proteins were specific. These data indicate that
GAL4-AD-Dlg4 interacts with the Cyt1 domain via the PDZ3 domain, but suggest that the C-
terminal region of the molecule might enhance binding.
We next investigated the regions of CD46 that were important for binding to Dlg4.
Cotransformation of GAL4-AD-Dlg4 with GAL4-BD-Cyt1 gave the expected growth (Fig2
Bi), but cotransformation of GAL4-AD-Dlg4 with GAL4-BD-Cyt2 did not confer growth
(Fig2 Bii). This indicated that the binding of Dlg4 was specific for the Cyt1 isoforms of
CD46. As the C-terminal 4 residues of the Cyt1 domain (FTSL) fits the consensus for
binding to PDZ domains, we tested whether these residues interacted with the PDZ3 domain
of Dlg4. Mutation of the C-terminal leucine residue of CD46 (GAL4-BD-Cyt1(L358R), Fig1
Biii) completely abrogated growth (Fig2 Biii), indicating that the C-terminus of CD46 is
important for binding to Dlg4. Cotransformation with two irrelevant GAL4-BD fusion
proteins did not confer growth (Fig2 Biv & v). These data show that the CD46 Cyt1, but not
Cyt2, domain binds to Dlg4 through its C-terminus, and are consistent with the hypothesis
that the consensus Dlg4 binding site within CD46-Cyt1 (the FTSL sequence) is necessary for
the interaction.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 12
12
CD46 can interact with Dlg4 expressed in mammalian cells. We tested whether the
interaction between CD46 and Dlg4 could be replicated in a system other than the Yeast Two
Hybrid system, using Dlg4 expressed in mammalian cells. The transformed human kidney
cell line, 293T, was transfected with vector alone or either alternatively spliced variant of
Dlg4 (Dlg4 and Dlg4-delGuK, Fig1 Ci & ii). Transfected cells were lysed, and incubated
with beads coupled to GST or a GST-Cyt1 (Fig1 Bvii) or GST-Cyt2 (Fig1 Bviii) fusion
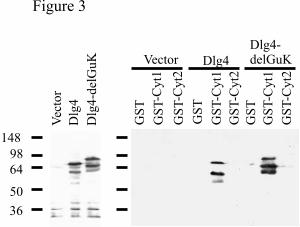
protein. Transfection of the two splice variants of Dlg4 gave equivalent levels of expression,
and the expected difference in size between the splice variants (Fig3 A). A band of lower
molecular weight was also seen when Dlg4 was transfected, consistent with previous reports
(45). It is thought that this band might be generated by proteolytic cleavage of the N-
terminus (45). In each case the GST-Cyt1 fusion protein interacted with Dlg4, but neither
GST alone nor GST-Cyt2 interacted. Interestingly, the lower molecular weight form of Dlg4
consistently bound more strongly to the GST-Cyt1, perhaps indicating that the modification
causing this form affects the binding of Dlg4 to CD46.
We attempted to coimmunoprecipitate CD46 and Dlg from the transfected 293T cells, using
the endogenous CD46. We were unable to do this, and in fact found that CD46 and Dlg were
not colocalised in these cells (Dlg expression was predominantly cytoplasmic, data not
shown). The intracellular localization of Dlg is regulated by complex mechanisms that may
be cell specific, and therefore it is possible that CD46 and Dlg are colocalised and able to
interact only in certain cell types. Other cell types, in which CD46 and Dlg do colocalize (see
below), expressed Dlg at levels too low to allow for efficient coimmunoprecipitation with the
available antibodies. However, our data with GST fusion proteins indicate that the Cyt1
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 13
13
domain of CD46 can interact with both alternatively spliced isoforms of Dlg4 expressed by
mammalian cells, but the Cyt2 domain does not interact with Dlg4.
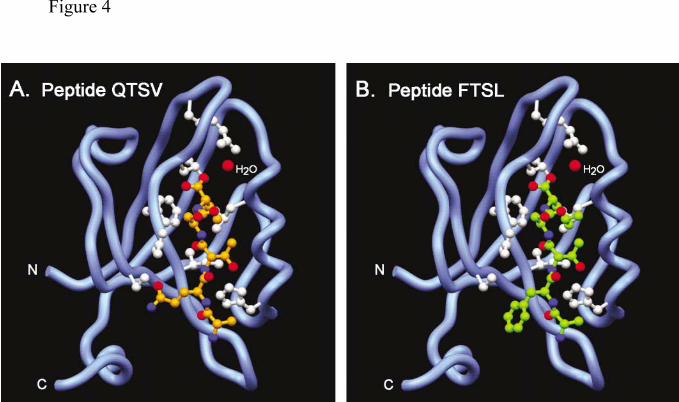
An interaction with Cyt1 is compatible with the structure of PDZ3. Although the C-
terminus of the Cyt1 domain contains the consensus sequence for PDZ binding (X-S/T-X-
I/L/V), the Phe at position –3 differs from the charged residue at this position in proteins
previously reported to bind to Dlg4. The crystal structure of the PDZ3 domain of Dlg4 has
been solved, both without peptide and with the C-terminus of CRIPT (an intracellular protein
that binds to PDZ3 of Dlg4 via the C-terminal sequence, QTSV) (32, 41). We used this
structure (Fig4 A) to model the C-terminus of CD46-Cyt1 in the groove of PDZ3 (Fig4 B).
The four-residue segment binds to the PDZ domain through antiparallel main chain
interactions with a β-sheet of the domain. The carboxylate group of the peptide is recognized
by a network of main chain amides provided by the loop sequence Gly322-Leu323-Gly324-
Phe325 as well as by the Arg318 sidechain. Additional side chain contacts and a
hydrophobic pocket determine the selectivity of the C-terminal consensus sequence. In this
model the Cyt1 domain fits in the groove with no obstructions, and although two hydrogen
bonds are lost compared with the CRIPT structure, this is unlikely to prevent the interaction.
A carboxyl terminal leucine in place of valine is possible with slight adjustment to the
hydrophobic pocket, considering local steric constraints and potential Van der Waal contacts.
Thus, the CD46 Cyt1 domain peptide sequence FTSL can be accommodated by a PDZ3
binding pocket.
The biological significance of an interaction between CD46 and Dlg4. Many PDZ-
containing proteins, including Dlg-1, are expressed in epithelial cells, where they regulate cell
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 14
14
polarization (37, 38). Both RNA and protein for Dlg4 have been identified in non-brain
tissues such as kidney (44). Interestingly, CD46 is polarized when ectopically expressed in
the MDCK epithelial cell line (7-9), so we next investigated whether CD46 and Dlg family
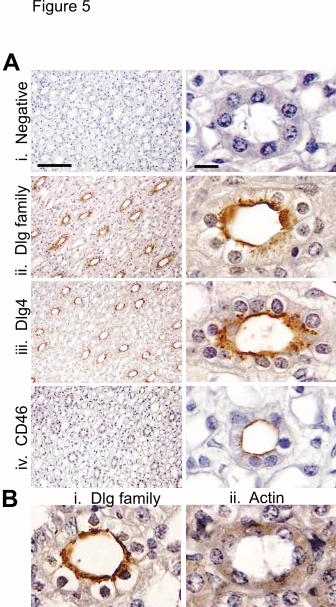
members were colocalized in epithelial tissue. An irrelevant antibody gave no
immunostaining in the epithelium of the kidney tubules, indicating that the staining was
specific (Fig5 Ai). Staining with antibodies to the Dlg family (Fig5 Aii), or specifically to
Dlg4 (Fig5 Aiii), showed apical polarization, Staining of normal human kidney sections with
antibodies to CD46 (Fig 5 Aiv), showed apical polarization, and indicated that CD46 and
Dlg4 are colocalized in epithelial cells. This polarized staining is not an artifact of the
immunohistochemistry procedure, as staining a different tissue section with Dlg gave the
same polarized staining (Fig5 Bi), but staining with an antibody to actin gave diffuse
reactivity over the whole cell (Fig5 Bii). These data are the first indication of a tissue in
which CD46 and Dlg4 might interact, and also suggested that Dlg4, as well as Dlg1, might be
involved in epithelial cell polarization. We also found polarization and colocalization of
CD46 in human gut epithelial tissue (data not shown) indicating that this role might extend to
other epithelial tissues.
Basolateral targeting of CD46 requires an interaction with PDZ-containing proteins.
The combined observations that (i) CD46 interacts with a PDZ domain, (ii) PDZ-containing
proteins are important for asymmetric targeting, and (iii) CD46 is polarized in epithelial cells
(above data and 7-9), suggested that this interaction might be important for intracellular
targeting of CD46. Further evidence for this hypothesis was that the removal of the FTSL
region of CD46 prevented basolateral targeting (8), however, the observation that the CD46
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 15
15
Cyt2 domain was also basolaterally targeted (7) suggested that additional mechanisms for
polarization were present in the CD46 sequence. We firstly investigated the expression of
Dlg family proteins in MDCK cells. Both mRNA and protein corresponding to Dlg4 were
detected in MDCK cells by RT-PCR and by immunoblots with an antibody specific for Dlg4
(data not shown). Other Dlg family members were expressed at higher levels as indicated by
immunoblotting with an antibody to the Dlg family, including Dlg1, which is known to be
important for epithelial polarization (37, 38). Dlg proteins are highly conserved amongst
mammalian species (human Dlg4 is 99% identical to mouse Dlg4, and the same four family
members have been identified in both mouse and human).
MDCK cells (which do not express CD46) were transfected with CD46 variants to
investigate the role of Dlg in the targeting of CD46. We firstly investigated the localization
of Dlg in these cells. MDCK cells were immunostained with antibodies to the Dlg family
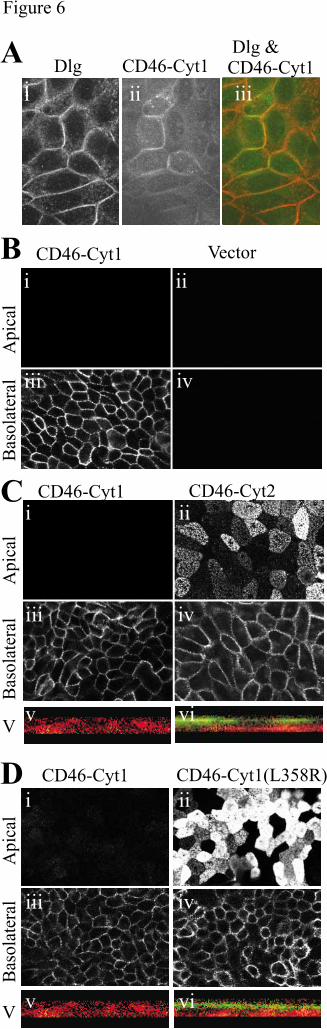
(Fig6 Ai), and to CD46 (Fig6 Aii). In both cases the staining was predominantly at the
periphery of the cells, and was colocalised (Fig6 Aiii). We assessed protein targeting by
seeding transfected MDCK cells onto transwell plates in which both upper and lower surfaces
were accessible to antibody. After allowing development of a completely confluent polarized
monolayer, we stained for CD46 using either a FITC-coupled secondary antibody (upper
surface), or a Fluo594 -coupled secondary antibody (lower surface). The slides were viewed
by confocal microscopy, with images taken horizontally, at the apical (upper) or basolateral
(lower) lever of the monolayer, and vertically. We found that CD46-Cyt1 was targeted to the
basolateral region of MDCK cells (Fig6 Bi & iii), in agreement with the results of Maisner et
al (7). The staining was specific for CD46 as cells transfected with vector alone showed no
staining (Fig6 Bii & iv). However, in contrast to CD46-Cyt1 (Fig6 Bi & iii), and to the
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 16
16
results of Maisner et al, CD46-Cyt2 was not polarized to the basolateral region, but was
distributed throughout the cell surface (Fig6 Cii & iv). Importantly, the mutation of leucine
358 of the Cyt1 domain (CD46-Cyt1L358R), that had abrogated GAL4-CtermDlg4 binding
in yeast two hybrid assays, prevented basolateral targeting (compare Fig6 Di & iii with Dii &
iv). Although CD46-Cyt1L358R could still be observed on the basolateral domain, protein
was also apical, indicating that the mechanism for localizing CD46-Cyt1 specifically to the
basolateral domain had been prevented. These observations suggest that the interaction with
Dlg is required for CD46 polarization.
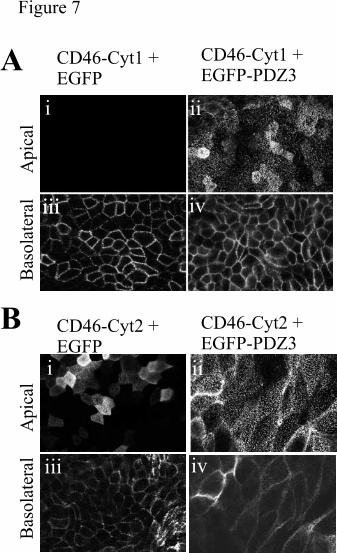
To further verify this hypothesis, we sought to block the interaction between CD46 and Dlg
and to investigate the effect of this block on CD46 polarization. We speculated that, if
CD46-Cyt1 was polarized via an interaction with a PDZ-containing protein, we would be
able to block that interaction by overexpression of the PDZ3 domain in isolation from the rest
of the molecule. We created a construct that encoded PDZ3 fused in frame to EGFP to
enable monitoring of expression. Because the EGFP emitted at similar wavelengths to FITC,
we labeled both upper and lower chambers with Fluo594 for these experiments. The
expression of CD46-Cyt1 gave the expected basolateral staining (Fig7 Ai & iii), but
coexpression of CD46-Cyt1 with the PDZ3 domain of Dlg4 fused to EGFP (EGFP-PDZ3)
prevented the basolateral polarization of CD46-Cyt1 (Fig7 Aii & iv). Thus, in the presence
of the EGFP-PDZ3, CD46-Cyt1 was expressed on both apical and basolateral surfaces,
resembling the localization of CD46-Cyt2. The expression of EGFP-PDZ3 had no effect on
the polarization of Cyt2 (compare Fig7 Bi & iii with Fig 7 Bii & iv). These data are
consistent with the model that overexpression of a PDZ3 domain can compete with wildtype
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 17
17
PDZ-containing domains to prevent the basolateral targeting of CD46. These data together
strongly suggest that CD46-Cyt1 is polarized via an interaction with Dlg proteins.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 18
18
DISCUSSION
We describe here a novel interaction between the Cyt1 cytoplasmic domain of CD46 (a
receptor for complement, measles virus and HHV6), and Dlg4 (an adaptor protein that
mediates signaling in the synapses of neurons), and demonstrate that Dlg4 or a similar protein
can regulate the intracellular trafficking of CD46 in epithelial cells. These studies highlight a
role for Dlg4 in the organization of epithelial cells, suggest the mechanism by which CD46
polarization is regulated, and provide a basis for the understanding of CD46 signal
transduction mechanisms.
A role for Dlg4 in epithelial polarization. The ubiquitous expression of Dlg4 has been
described previously (44), but attention has remained focused on a role for Dlg4 in nervous
tissue. We were interested in the possibility that CD46 and Dlg4 might interact in the brain,
because both measles virus and HHV6 (two pathogens that use CD46 as a receptor) are
associated with neuronal pathology (for instance subacute sclerosing panencephalitis caused
by measles, and febrile convulsions and encephalitis caused by HHV6, 46, 47). It is
conceivable that a component of this pathology could be due to effects on Dlg4 function, and
particularly on nitric oxide-mediated toxicity (48), caused by pathogen-receptor interactions.
In the absence of human brain sections of suitable quality for immunostaining, we looked for
colocalization of CD46 and Dlg in the brain of a mouse transgenic for CD46-Cyt1 (49).
These data suggest that CD46 and Dlg4 are not colocalized in neurons (data not shown), so
we have no evidence to suggest that CD46 and Dlg4 have a functional interaction in the
brain. However, our data indicate for the first time a possible role for Dlg4 in regulating the
polarization of CD46 in epithelial tissue. We identified mRNA transcripts for Dlg4 in
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 19
19
MDCK cells, and in human epithelial cell lines, using RT-PCR (data not shown). Scanning
of the EST databases and SAGE analysis indicates that Dlg4 is expressed in tissues such as
normal ovarian epithelium, normal ductal epithelial cells, adenomas and adenocarcinomas.
In addition, immunostaining with an antibody specific for Dlg4 showed expression in normal
human epithelial tissue. It is possible that Dlg4 and Dlg1 may both contribute to the
polarization of epithelial proteins.
Interestingly, the Dlg4 clone that we isolated during our yeast two hybrid screen contained a
110 base deletion, which changed the reading frame and therefore resulted in deletion of most
of the guanylate kinase-like domain. Clones with this deletion have been identified
previously from mammary gland and testicular tissue (44), and we have identified it in
numerous cell types by RT-PCR (data not shown) and searching of the GenBank EST
databases. Searching for EST sequences corresponding to this region indicated that the
splicing of Dlg4 is tissue specific. Of the 12 sequences that contained exon 20, 11 were
from brain or nervous tissue. By contrast, of the 8 sequences in which exon 20 was deleted,
all were from non-brain derived tissue such as breast, prostate and head and neck tumors, and
Wilm’s tumor. We show here that both splice variants of Dlg4 can bind to CD46. Recent
reports have suggested that Dlg4 adopts a tertiary conformation in which the interaction of
the GuK domain with other regions of the protein affect the capacity of Dlg4 to bind to and
cluster other proteins (50, 51). It is therefore likely that these splice variants might differ in
their ability to recruit proteins into a signaling complex. The tissue-specific splicing of Dlg4
may reflect a different function for the protein in different tissues. The data described above
indicate both that Dlg4 is expressed in epithelial tissues, and that Dlg4 in epithelial tissues is
an alternatively spliced form.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 20
20
A mechanism for the basolateral targeting of CD46. Three studies have described the
basolateral targeting of CD46 in the polarized MDCK epithelial cell line. The targeting of
CD46-Cyt1 isoforms was prevented by various mutations in the last 4 amino acids of the
CYT1 domain (7-9). The molecular mechanisms regulating this polarization had not been
elucidated, but it was thought that the FTSL sequence might operate in a similar manner to a
canonical YxxL basolateral targeting sequence. In light of the observed interaction between
CD46 and Dlg, we considered the alternative possibility that Dlg4 or a related family member
might regulate the polarization of CD46 in epithelial cells. We show: (i) that the L358R
mutant (which fails to bind Dlg4) is targeted differently than the wild-type CD46-Cyt1, (ii)
that expression of the PDZ3 domain of Dlg4 alone (PDZ3-EGFP) can affect the basolateral
targeting of CD46 (perhaps acting as a dominant negative inhibitor of Dlg family binding),
and (iii) that the two proteins are copolarized in normal human epithelial cells. A previous
report (7) had shown that an isoform containing the Cyt2 domain of CD46 (that does not bind
to Dlg4) is also basolateral in MDCK cells. However, in our hands this was not the case, and
three different clones expressing Cyt2 isoforms displayed a non-polarized distribution similar
to that of CD46(L358R) and of the Cyt1 isoform coexpressed with PDZ3-EGFP. We do not
know why our data are different to the published data (7), however our results fit with the
idea that basolateral targeting of CD46 requires an interaction between the FTSL region of
the Cyt1 domain and Dlg4 or related proteins. This study (7) also showed other mutations
that prevented basolateral targeting, and all of these mutations disrupted the consensus PDZ-
binding motif of CD46 and so are likely to prevent binding of CD46 to Dlg4. Although it
was suggested that the FTSL sequence mediates targeting by its resemblance to a YxxL
basolateral targeting sequence (8, 9), it is not usual for such a sequence to (i) remain
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 21
21
functional when the tyrosine is replaced by a phenylalanine, and (ii) be functional at the C-
terminus of a protein. Interestingly, there is a YxxL motif just upstream of the sequence that
Maisner et al define as the cytoplasmic domain (see Fig1 A). We mutated this motif (Y to A)
in both the Cyt1 and Cyt2 isoforms of CD46 and found no effect on the polarization of CD46
(data not shown). Furthermore, we found that CD46 expressed in MDCK cells could be
internalized irrespective of binding to Dlg, and that expression of Dlg did not chance the rate
or extent of internalization (data not shown). We suggest that the mechanism by which the
FTSL sequence regulates the basolateral distribution of CD46-Cyt1 is through an interaction
with the PDZ domain of Dlg4 or a similar molecule.
We were concerned that the polarization of CD46 in MDCK cells might not necessarily
reflect trafficking in normal human polarized cells, and so investigated the expression of both
CD46 and Dlg4 in human kidney sections. CD46 was clearly polarized in kidney (Fig5) and
gut (data not shown) epithelial cells, and colocalized with Dlg4 and the Dlg family. Both
proteins were apical in these cells, rather than basolateral as in the MDCK cells. The
localization of other polarized proteins has been shown to change depending on the cell in
which it is expressed (52, 53). Interestingly, our immunohistochemistry indicates that all of
the CD46 in human kidney epithelium is polarized, even though our data suggests that Cyt2
isoforms would not be polarized. Possible explanations for this observation include either
that the Cyt2 domain is not expressed in these tissues, or that it is “piggy-backed” by the Cyt1
domain when the isoforms are coexpressed. We favour the second hypothesis, as we have
seen significant expression of the Cyt2 isoforms in kidney tissue, and in epithelial cell lines
(54, data not shown).
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 22
22
The mechanisms by which CD46 transmits intracellular signals is not yet known. The known
role of Dlg4 as a scaffold for signaling complexes, and the wealth of literature showing
interactions between Dlg4 and other signaling molecules, provides a basis for the
investigation of interactions between CD46 and many of these signaling proteins. For
example, Dlg proteins interact with the src-family kinases, lck and fyn (34, 35), and CD46
can be phosphorylated by lck (17). It will be interesting to determine whether the
phosphorylation of CD46 by lck is mediated by an interaction through Dlg4. Importantly, we
have identified a single residue of the CD46 Cyt1 domain that can be mutated to prevent the
interaction with Dlg4. This can be used to determine the relevance of Dlg to cellular effects
mediated by CD46 signaling, such as effects on cytokine production.
These data described in this study indicate that the Cyt1 domain of CD46 is both physically
and functionally associated with Dlg4. This is the first identification of a direct, and isoform-
specific interaction between CD46 and an intracellular protein, forming a potential platform
for the assembly of a signaling complex. This interaction provides a molecular mechanism
for the polarization of CD46 in epithelial cells described previously (7-9). Furthermore, these
data suggest that, in addition to the documented role of Dlg4 in establishing neuronal
architecture, Dlg4 may contribute to epithelial cell polarization.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 23
23
REFERENCES
1. Oglesby, T.J., Allen, C.J., Liszewski, M.K., White, D.J., and Atkinson, J.P. (1992) J.
Exp. Med. 175, 1547-51.
2. Naniche, D., Varior-Krishan, G., Cervoni, F., Wild, T.F., Rossi, B., Rabourdin-Combe, C.,
and Gerlier, D. (1993) J.Virol. 67, 6025-6032.
3. Dorig, R.E., Marcil, A., Chopra, A. and Richardson, C.D. (1993) Cell, 75, 295-305.
4. Manchester, M., Eto, D.S., Valsamakis, A., Liton, P.B., Fernandez-Munoz, R., Rota, P.
A., Bellini, W.J., Forthal, D.N., and Oldstone, M.B. (2000) J Virol 74, 3967-74.
5. Santoro F, Kennedy PE, Locatelli G, Malnati MS, Berger EA, Lusso P. (1999) Cell.
99, 817-27.
6. Russell, S.M., Sparrow, R.L., McKenzie, I.F.C. and Purcell, D.F.J. (1992). Eur. J.
Imm. 22: 1513-1518.
7. Maisner, A., Liszewski, M. K., Atkinson, J. P., Schwartz-Albiez, R., and Herrler, G.
(1996) J Biol Chem 271, 18853-8.
8. Maisner, A., Zimmer, G., Liszewski, M. K., Lublin, D. M., Atkinson, J. P., and Herrler,
G. (1997) J Biol Chem 272, 20793-9.
9. Teuchert, M., Maisner, A., and Herrler, G. (1999) J Biol Chem 274, 19979-84.
10. Karp, C. L., Wysocka, M., Wahl, L. M., Ahearn, J. M., Cuomo, P. J., Sherry, B.,
Trinchieri, G., and Griffin, D. E. (1996). Science 273, 228-31.
11. Karp, C. L. (1999) Immunol Rev 168, 91-101.
12. Imani, F., Proud, D. and Griffin, D.E. (1999) J. Immunol. 162, 1597-602.
13. Hirano, A., Yang, Z., Katayama, Y., Korte-Sarfaty, J., and Wong, T. C. (1999) J Virol
73, 4776-85.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 24
24
14. Katayama, Y., Hirano, A., and Wong, T. C. (2000) J Virol 74, 1252-7.
15. Astier A, Trescol-Biemont MC, Azocar O, Lamouille B, Rabourdin-Combe C.
(2000) J. Immunol. 164, 6091-5.
16. Marie, J.C., Kehren, J., Trescol-Biemont, Evlashev, A., Valentin, H., Walzer, T.,
Tedone, R., Loveland, B., Bicholas, J.F., Rabourdin-Combe, C. and Horvat, B. (2001)
Immunity 14, 69-79.
17. Wang, G., Liszewski, M.K., Chan, A.C. and Atkinson, J.P. (2000) J. Immunol. 164,
1839-1846.
18. Kurita-Taniguchi M, Fukui A, Hazeki K, Hirano A, Tsuji S, Matsumoto M, Watanabe
M, Ueda S, Seya T. (2000) J Immunol. 165, 5143-52.
19. Woods, D.F & Bryant, P.J. (1991) Cell 66, 451-64.
20. Fujita, A. & Kurachi, Y. (2000) Biochem. Biophys. Res. Commun. 269, 1-6.
21. Dimitratos, S.D., Woods, D.F., Stathakis, D.G., and Bryant, P.J. (1999) BioEssays
21, 912-921.
22. Garner, C.C., Nash, J. & Huganir, R.L. (2000) Trends. Cell. Biol. 10, 274-80.
23. Kornau, HC., Schenker LT., Kennedy MB., and Seeburg, P.H. (1995) Science. 269,
1737–40.
24. Kim, E., Niethammer, M., Rothschild, A., Jan, Y. N., and Sheng, M. (1995) Nature
378, 85-8.
25. Irie, M., Hata, M., Ichtenko, K., Toyoda, A., Hirao, K., Takai, Y., Rosahl, T.W.,
Sudhof, T.C. (1997) Science, 277, 1511–5.
26. Huang Y.Z., Won, S., Ali, D.W., Wang, Q., Tanowitz, M., Du, Q.S., Pelkey, K.A.,
Yang, D.J., Xiong, W.C., Salter, M.W., and Lin Mei (2000). Neuron, 26. 443–455.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 25
25
27. Husi, H., Ward, M.A., Choudhary, J.S., Blackstock, W.P. and Grant, S.G.N. (2000)
Nat Neurosci. 3:661-9.
28. Sheng, M., and Sala, C. (2001). Annu Rev. Neuro. 24, 1-29.
29. Hanada, T., Lin, L., Chandy, K.G., Oh, S.S. & Chisti, A.H. (1997) J. Biol. Chem.
272, 26899.
30. Kim, E., Naisbitt, S., Hsueh, Y.-P., Rao, A., Rothschild, A., Craig, A.M., and Sheng, M.
(1997) J. Cell. Biol. 136, 669-678.
31. Kim, J.H., Liao, D., Lau, L.-F., & Huganir, R.L. (1998) Neuron 20, 683.
32. Niethammer, M., Valtschanoff, J.G., Kapoor, T.M., Allison, D.W., Weinberg, R.J.,
Craig, A.M. & Sheng, M. (1998) Neuron 20, 693.
33. Passafaro, M., Sala, C., Niethammer, M. and Sheng, M. (1999) Nature Neuroscience,
2, 1063-1069.
34. Tezuka, T., Umemori, H., Akiyama, T., Nakanishi, S., and Yamamoto, T. (1999) Proc.
Natl. Acad. Sci (USA) 96, 435-440.
35. Hanada, T., Lin, L., Tibaldi, E. V., Reinherz, E. L., and Chishti, A. H. (2000) J Biol
Chem. 275, 28774-84.
36. Savinainen A, Garcia, E.P., Dorow, D., Marshall, J. and Liu, Y.F. (2001) J. Biol.
Chem. 276,11382-6.
37. Kim, S. K. (1997) Curr Opin Cell Biol 9, 853-9.
38. Fanning, A. S., and Anderson, J. M. (1999) J Clin Invest 103, 767-72.
39. James, P., Halladay, J. & Craig, E.A. (1996) Genetics 144, 1425 – 1436.
40. Browne, K.A, Johnstone, R.W., Jans, D.A., and Trapani, J.A. (2000) J. Biol. Chem
275, 39262-6.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 26
26
41. Doyle, D.A., Lee, A., Lewis, J., Kim, E., Sheng, M. and MacKinnon, R. (1996) Cell 85,
1067-1076.
42. Jones, T.A. and Kjeldgaard, M. (1994) O - The Manual. Uppsala University Press,
Uppsala.
43. Stathakis, D.G., Hoover, K.B., You, Z., and Bryant, P.J.. (1997) Genomics, 44, 71–
82.
44. Stathakis, D.G., Udar, N, Sandgren, O., Andreasson, S., Bryant, P.J., Small, K., and
Forsman-Semb, K. (1999) J. Neurochem. 73, 2250-2265.
45. Topinka, J.R. and Bredt, D.S. (1998) Neuron 20, 125-134.
46. Scheider-Schaulies S, ter Meulen, V. (1999) Arch Virol. Suppl 15, 139-58.
47. Campadelli-Fiume, G., Mirandola, P. and Menotti, L. (1999) Emerging Infectious
Diseases 5, 353-366.
48. Sattler, R., Xiong, Z., Lu, W.-Y., Hafner, M., MacDonald, J.F., and Tymianski, M.
(1999). Science, 284, 1845-1848.
49. Thorley, B.R., Milland, J., Christiansen, D., Lanteri, M.B., McInnes, B., Moeller, I.,
Rivailler, P., Horvat, B., Rabourdin-Combe, C., Gerlier, D., McKenzie, I.F.C., and
Loveland, B.E. (1997) Eur. J. Immunol. 27, 726 – 734.
50. Wu, H., Reissner, C., Kuhlendahl, S., Coblent, B., Reuver, S., Kindler, S., Gundelfinger,
E.D., and Garner, C.C. (2000) EMBO J 19, 5740-5751.
51. McGee, A.W. and Bredt, D.S. (1999) J. Biol. Chem 274, 17431-17436.
52. Roush, D.L., Gottardi, C.J., Naim, H.Y., Roth, M.G., aand Caplan, M.J. (1998) J.
Biol. Chem 273, 26862-9.
53. Breton, S., Wiederhold, T., Marshansky, V., Nsumu, N.N., Ramesh, V. and Brown,
D. (2000) J. Biol. Chem 16, 18219-24.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 27
27
54. Johnstone, R.W., Russell, S.M., Loveland, B.E. and McKenzie, I.F. (1993) Mol Immunol
30, 1231-41.
55. Evans, S.V. (1993) J. Molec. Graphics 11, 134-138.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 28
28
FOOTNOTES.
We thank Phillip James (University of Wisconsin) for the PJ yeast strain, Peter Gunning
(New Children’s Hospital, NSW) for the antibody to β-actin, Caroline Farrelly and Manuella
Palatsides (Peter MacCallum Cancer Institute) for technical assistance, Alpha Yap
(University of Queensland) for helpful discussions and Ricky Johnstone and Joe Trapani
(Peter MacCallum Cancer Institute), and Julie Milland (Austin Research Institute) for critical
reading of the manuscript.
S. Russell is supported by a Wellcome Senior Research Fellowship in Medical Science in
Australia.
Abbreviations are : Dlg, Discs large; FITC, fluorescein isothiocyanate; MDCK, Madin-Darby
canine kidney; SAP, synapse-associated protein; MAGUK, membrane-associated Guanylate
kinase; PSD, Post-synaptic density.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 29
29
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 30
30
FIGURE LEGENDS
Figure 1. Structure of CD46 and Dlg4. (A) Sequence of the two alternatively spliced
cytoplasmic domains of CD46. Immediately following the hydrophobic transmembrane
domain is a common domain of 12 amino acids starting at amino acid 331, followed by
unique sequence of 15 or 23 amino acids for the Cyt1 and Cyt2 domains respectively.
Numbering of CD46 amino acid residues is according to Russell et al (6). (B) Schematic of
the domain structure of CD46 and constructs of CD46 used in this study. (i) & (ii) The
extracellular domain of CD46 is composed of four Short Consensus Repeat (SCR) domains
responsible for binding to ligands (filled ovals), and the glycosylated Ser/Thr/Pro (STP-rich
domain, bold line), the transmembrane domain (thin line) is common in all isoforms and is
followed by the cytoplasmic domain 1 (dotted oval) or 2 (hashed oval). (iii) The point
mutation (L358R) in the cytoplasmic domain of Cyt1, which prevents binding to Dlg4, is
indicated by an X. For yeast two hybrid analysis the GAL-4 activation domain (square) was
fused to the sequence corresponding to the Cyt1 domain of CD46 (iv), the Cyt1 domain with
a L358R mutation (v), or the Cyt2 domain (vi). The Cyt1 (vii) and Cyt2 (viii) domain was
also fused to glutathione-S-transferase (GST)(indicated by a triangle). (C) Schematic of the
domain structure of Dlg4 and constructs of Dlg4 used in this study. (i) Dlg4 is composed of 3
PDZ domains (triangles), an SH3 domain (rectangle) and a GuK (guanylate kinase) domain
(oval). (ii) An alternatively spliced isoform of Dlg4 in which most of the GuK domain is
deleted (circle). (iii) The clone isolated during Yeast Two Hybrid screening contains the
GAL4 activation domain (square) fused to Dlg4 beginning at residue 198 in PDZ 2, and
corresponds to the isoform in which most of the GuK domain is deleted. Constructs created
for this study include (iv) the original Yeast Two Hybrid clone with a stop codon
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 31
31
immediately after the PDZ3 domain, (v) the original Yeast Two Hybrid clone with a single
amino acid change in Arg 318 of PDZ3, and (vi) the PDZ3 domain of Dlg4 fused in frame to
EGFP (filled rectangle). Sequence numbering is from Stathakis et al (43).
Fig2. Dlg4 binds specifically to the Cyt1 domain of CD46 through PDZ3. (A). The
interaction between Dlg4 and CD46 requires PDZ3 of Dlg4. Yeast strain PJ69-4A was
cotransformed with (i) GAL4-AD-Dlg4, (ii) GAL4-AD-Dlg4mutPDZ3, (iii) GAL4-AD-
Dlg4(trunc) or (iv) GAL4-ABP280 together with GAL4-BD-Cyt1. Yeast colonies were
subjected to growth tests on selective medium deficient in Leu, Trp and His. (B) The
interaction between Dlg4 and CD46 is specific for the C-terminus of the Cyt1 domain of
CD46. Yeast strain PJ69-4A was cotransformed with isolated GAL4-AD-Dlg4 together with
(i) GAL4-BD-Cyt1, (ii) GAL4-BD-Cyt2, (iii) GAL4-BD-Cyt1(L358R), (iv) pLAM, and (v)
pAS2-1. Yeast colonies were subjected to growth tests on selective medium deficient in Leu,
Trp and His.
Fig. 3. CD46-Cyt1 interacts with Dlg4 expressed in mammalian cells. (A) Human 293T
cells were transfected with vector alone, Dlg4, or Dlg4-DelGuK (left panel). Cell lysates
were incubated with beads coupled to GST, GST-Cyt1, or GST-Cyt2 fusion protein. After
extensive washing the bound protein was electrophoresed under reducing conditions and
immunoblotted with antibody to Dlg4 (right panel).
Figure 4. The Cyt1 domain is compatible with the model of the PDZ3 domain. (A) X-ray
crystallographic structure of the third PDZ domain of Dlg4 (blue) in complex with its peptide
ligand QTSV (orange) (41). PDZ amino acids 308-402 are shown. PDZ sidechains (white)
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 32
32
interacting with the peptide specifically are Arg318, Gly 322, Leu323, Gly324, Phe325,
Asn326, Ile327, Gly329, Ser339, His372, Leu379, and Gly383. A well-ordered H2O molecule
(red) hydrogen bonds the peptide carboxylate group to Arg318 of PDZ. (B) Molecular model
of peptide FTSL (green) interacting with PDZ domain, based on the crystal structure of PSD-
95 complex seen in figure A. Images were generated with Setor (55).
Fig. 5. CD46 and Dlg4 are coexpressed and copolarized in normal human epithelial
cells. A. Sections of a normal human kidney were immunostained with antibodies to (i)
the murine Ly2.1 antigen (IgG2a antibody, negative control antibody), (ii) the Dlg family,
(iii) Dlg4, and (iv) CD46 (1840). B. To determine whether the apical staining was due to
inaccessibility of the basolateral regions to antibody, immunostaining with antibody to the
Dlg family (i) was compared with immunostaining with antibody to actin (ii). Sections
were counterstained with haemotoxylin, specific staining is represented by the brown
color, and scale bars represent 100 µm (A, left panels) and 10 µm (A, right panels and B).
Fig 6. The Dlg4-binding site of CD46 is required for basolateral targeting in polarized
MDCK cells. A. MDCK cells transfected with CD46-Cyt1 were grown to confluency on
trasnwell plates, fixed in methanol for 20 minutes and immunostained with antibodies to Dlg
family (followed by Fluo-594-labelled secondary antibody, panel i and red in panel iii) or to
CD46 (followed by FITC-labelled secondary antibody, panel ii and green in panel iii). An
optical section was taken through the middle of the cells. B. MDCK were transfected with
CD46-Cyt1 (i & iii) or with a vector control (ii & iv), and immunostained to detect apical (i
& ii) or basolateral (iii & iv) expression of CD46 with the 1840 rabbit antibody to CD46.
Cells were seeded onto transwell plates and grown to confluency. The upper and lower
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CD46 and Dlg4 interact. 33
33
surfaces of the membrane were stained with the 1840 antibody to CD46 and either FITC-
labeled (upper) or Fluo-594-labelled (lower) anti-rabbit secondary antibodies. The slides
were viewed by confocal microscopy, with images taken horizontally, at the apical (upper) or
basolateral (lower) level of the monolayer, and vertically (bottom images). C. MDCK cells
were transfected with CD46-Cyt1 (i, iii, v) and CD46-Cyt2 (ii, iv, vi) and cultured and
stained as for A. Vertical sections (v & vi) indicate both basolateral (green) and apical (red)
staining. D. MDCK cells were transfected with CD46-Cyt1 (i, iii, v) and CD46-
Cyt1(L358R) (ii, iv, vi) and cultured and stained as for A. Vertical sections (v & vi) indicate
both basolateral (green) and apical (red) staining.
Fig 7. Expression of the PDZ3 domain of Dlg4 prevents basolateral targeting of CD46-
Cyt1. MDCK cells were firstly transfected with CD46-Cyt1 (A) or CD46-Cyt2 (B), and then
were cotransfected with EGFP (i & iii) or EGFP-PDZ3 (ii & iv), sorted by flow cytometry for
EGFP fluorescence, and tested for polarization of CD46. Cells were seeded onto transwell
plates and grown to confluency. The upper and lower surfaces of the membrane were stained
with the 1840 antibody to CD46 and Fluo-594-labelled anti-rabbit secondary antibodies. The
slides were viewed by confocal microscopy, with images taken horizontally, at the apical
(upper) or basolateral (lower) lever of the monolayer.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Loveland, Margaret Bills, Sarah Ellis and Sarah M. RussellMandy J. Ludford-Menting, Suzanne J. Thomas, Blessing Crimeen, Lisa J. Harris, Bruce E.
polarisationA functional interaction between CD46 and Dlg4: a role for Dlg4 in epithelial
published online November 19, 2001J. Biol. Chem.
10.1074/jbc.M108479200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from