Embed Size (px)
Citation preview

BMJ Paediatrics Open is committed to open peer review. As part of this commitment we make the peer review history of every article we publish publicly available. When an article is published we post the peer reviewers’ comments and the authors’ responses online. We also post the versions of the paper that were used during peer review. These are the versions that the peer review comments apply to. The versions of the paper that follow are the versions that were submitted during the peer review process. They are not the versions of record or the final published versions. They should not be cited or distributed as the published version of this manuscript. BMJ Paediatrics Open is an open access journal and the full, final, typeset and author-corrected version of record of the manuscript is available on our site with no access controls, subscription charges or pay-per-view fees (http://bmjpaedsopen.bmj.com). If you have any questions on BMJ Paediatrics Open’s open peer review process please email
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
Peripheral blood RNA gene expression in children with
pneumococcal meningitis provides insights into host
response mechanisms
Journal: BMJ Paediatrics Open
Manuscript ID bmjpo-2017-000092
Article Type: Original article
Date Submitted by the Author: 23-May-2017
Complete List of Authors: Kulohoma, Benard ; Centre for Biotechnology and Bioinformatics,
University of Nairobi, Nairobi, Kenya;; University of Oxford, Wellcome Trust Centre for Human Genetics, Nuffield Department of Medicine, Marriage, Fiona; 3. Centre for Integrated Genomic Research, University of Manchester, Manchester, UK. Vasieva, Olga; University of Liverpool, Institute of Integrative Biology Nguyen, Kha; University of Liverpool, Institute of Translational Medicine Mankhambo, Limangeni; 2. Malawi-Liverpool-Wellcome Trust Clinical Research Programme, College of Medicine, Blantyre, Malawi. Molyneux, Malcolm; 2. Malawi-Liverpool-Wellcome Trust Clinical Research Programme, College of Medicine, Blantyre, Malawi Molyneux, ELizabeth; Paediatrics Day, Philip; 3. Centre for Integrated Genomic Research, University of
Manchester, Manchester, UK. Carrol, Enitan; University of Liverpool, Institute of Infection and Global Health
Keywords: HIV, Immunology, Infectious Diseases, Microbiology, Tropical Paediatrics
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open on June 3, 2020 by guest. P
rotected by copyright.http://bm
jpaedsopen.bmj.com
/bm
jpo: first published as 10.1136/bmjpo-2017-000092 on 31 A
ugust 2017. Dow
nloaded from

Confidential: For Review O
nly
1
Title: Peripheral blood RNA gene expression in children with 1
pneumococcal meningitis provide insights into host response 2
mechanisms 3
Benard W. Kulohoma 1 † *, Fiona Marriage 3 *, Olga Vasieva 4, Limangeni 4
Mankhambo 2, Kha Nguyen 5, Malcolm E Molyneux 2, Elizabeth M Molyneux 6, 5
Philip JR Day 3, Enitan D Carrol 2 5 6 ‡ 6
7
1. Centre for Biotechnology and Bioinformatics, University of Nairobi. 8
2. Malawi-Liverpool-Wellcome Trust Clinical Research Programme, College of 9
Medicine, Blantyre, Malawi. 10
3. Centre for Integrated Genomic Research, University of Manchester, 11
Manchester, UK. 12
4. Institute of Integrative Biology, University of Liverpool, Liverpool, UK. 13
5. Institute of Infection and Global Health, University of Liverpool, Liverpool, 14
UK. 15
6. Department of Paediatrics, University of Malawi College of Medicine, 16
Blantyre, Malawi. 17
18
*Contributed equally. 19
† Current address: Wellcome Trust Centre for Human Genetics, University of 20
Oxford. 21
‡ Corresponding author 22
23
Benard W. Kulohoma ([email protected]) 24
Fiona Marriage ([email protected]) 25
Olga Vasieva ([email protected] ) 26
Limangeni Mankhambo ([email protected]) 27
Kha Nguyen ([email protected]) 28
Malcolm Molyneux ([email protected]) 29
Elizabeth Molyneux ([email protected]) 30
Philip J. R. Day ([email protected]) 31
Enitan D. Carrol ([email protected]) - Corresponding author 32
Page 1 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
2
Abstract 33
Introduction: Invasive pneumococcal disease (IPD), caused by Streptococcus 34
pneumoniae, is a leading cause of pneumonia, meningitis and septicaemia 35
worldwide. Morbidity and mortality in pneumococcal meningitis are significantly 36
increased among children with underlying HIV infection. 37
Objectives: We hypothesised that peripheral blood expression profiles from 38
HIV-infected and un-infected children with pneumococcal meningitis would 39
provide insight into the host inflammatory response leading to poorer outcomes. 40
Design and setting: Prospective case control observational study in a tertiary 41
hospital in Malawi 42
Participants: Children up to the age of 16 years with a presumptive diagnosis 43
of meningitis or pneumonia. For the microarray analysis, we studied children 44
with pneumococcal meningitis (n=12) and matched HIV-uninfected controls 45
(n=3). For the real time qPCR, we studied 229 cases and 13 controls with IPD 46
Methods: We used the human genome HGU133A Affymetrix array to explore 47
differences in gene expression between cases and controls and between HIV-48
infected and uninfected cases, and validated gene expression profiles for 34 49
genes using real time qPCR in an independent set of cases and controls. 50
Pathway analysis was used to explore genes differentially expressed. 51
Results: Irrespective of underlying HIV infection, cases showed significant up-52
regulation of: S100 calcium binding protein A12 (S100A12); vanin-1 (VNN1); 53
arginase, liver (ARG1); matrix metallopeptidase 9 (MMP9); annexin A3 54
(ANXA3); interleukin 1 receptor, type II (IL1R2); CD177 molecule (CD177); 55
S100 calcium binding protein A9 (S100A9), cytoskeleton-associated protein 4 56
Page 2 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
3
(CKAP4); and glycogenin 1 (GYG1). The RT-PCR results were in agreement 57
with our microarray analysis findings. There was no differential gene expression 58
in HIV-infected compared to HIV-uninfected cases, but there was significant up-59
regulation of folate receptor 3 (FOLR3), endocytic adaptor protein (NUMB) and 60
S100A12 in survivors compared non-survivors. 61
Conclusion: Children with pneumococcal meningitis demonstrated increased 62
expression in genes regulating immune activation, oxidative stress, leucocyte 63
adhesion and migration, arginine metabolism, and glucocorticoid receptor 64
signalling. 65
66
(298 words) 67
68
Key words: Differential expression, children, invasive pneumococcal disease, 69
host response, pneumococcal meningitis 70
71
Key messages: 72
73
What is known about the subject 74
Invasive pneumococcal disease (IPD) caused by Streptococcus pneumoniae is 75
a leading cause of pneumonia, meningitis and septicaemia worldwide. Globally, 76
IPD is reported to cause about 11% (0.8 million) of all deaths in children less 77
than 5 years of age annually. The overall burden of IPD is 40-fold increased in 78
HIV-infected compared with HIV-uninfected children. However, mechanisms 79
involved in host response during IPD are not yet fully understood. 80
Page 3 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
4
81
What this study adds 82
We describe differences in transcriptional profiles between HIV-infected and 83
HIV-uninfected children with pneumococcal meningitis (as a homogeneous 84
disease entity of invasive pneumococcal disease) and healthy controls. We 85
demonstrate for the first time, increased expression in cases of genes regulating 86
the innate immune response, leucocyte migration, glucose homeostasis and 87
endothelial cell migration. 88
89
90
Introduction 91
92
Streptococcus pneumoniae infection is a leading cause of pneumonia, 93
meningitis and septicaemia worldwide, and results in approximately 1 million 94
deaths in children under the age of 5 years annually 1. The overall burden of 95
invasive pneumococcal disease (IPD) is 40-fold increased in HIV-infected 96
compared with HIV-uninfected children 2. Pneumococcal meningitis is a life-97
threatening disease with poor prognosis associated with neurologic 98
complications and a high case-fatality ratio in African children, which is further 99
increased by HIV co-infection 3. 100
101
Gene expression profiling during sepsis provides new insights into the host 102
response to invasive bacterial disease. Several sepsis studies have 103
demonstrated up-regulation of pathogen recognition receptors and signal 104
Page 4 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
5
transduction pathways, and a down-regulation of zinc homeostasis 4-6. The 105
intricate host inflammatory response is associated with neuronal and vascular 106
injury, even after cerebrospinal fluid (CSF) sterilization with antibiotics. 107
Adjunctive new therapies for bacterial meningitis have to date not shown any 108
conclusive benefit, prompting the need for an improved understanding of key 109
mechanisms that might reveal potential new therapeutic targets 7. Chronic co-110
infection may impact gene expression, even among asymptomatic patients. HIV 111
is a major risk factor for IPD, characterised by waning immunity and 112
dysregulated physiology among infected individuals, greatly escalating disease 113
susceptibility and mortality outcomes8 9, thereby influencing the host’s gene 114
expression. We examine differences in gene-expression using blood samples 115
from children with pneumococcal meningitis and matched healthy controls, and 116
also compare transcriptional profiles between children with pneumococcal 117
meningitis with and without underlying HIV infection. 118
119
Materials and methods 120
121
Patients and controls 122
Children (n=377) were recruited as part of a larger study investigating host 123
determinants of susceptibility to IPD conducted at Queen Elizabeth Central 124
Hospital, Blantyre, Malawi, between April 2004 and October 2006 10. Ethical 125
approval was granted from the College of Medicine Research Committee 126
(COMREC), Malawi and the Liverpool School of Tropical Medicine Local 127
Page 5 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
6
Research Ethics Committee. Written informed consent was given prior to 128
recruitment. 129
130
We excluded children (n=135) infected with other commonly prevalent microbes 131
(Salmonella typhimurium, Escherichia coli, Haemophilus influenzae b, Neisseria 132
meningitidis, Staphylococcus aureus, Streptococcus pyogenes) identified by 133
positive laboratory culture. Cases for the microarray analysis were HIV 134
treatment-naive children with confirmed pneumococcal meningitis defined as 135
abnormal CSF white blood cell (WBC) count, >10 WBC/µl plus one or more of 136
the following: CSF culture positive for pneumococci, CSF gram stain consistent 137
with pneumococci, CSF positive for pneumococcal polysaccharide antigen or 138
pneumococcal DNA. Cases for the RT-qPCR were children with confirmed IPD, 139
which was defined as: pneumococcal pneumonia (n=40) or pneumococcal 140
meningitis (n=189), confirmed by either culture, antigen test, or PCR. Controls 141
were healthy afebrile, aparasitaemic children from the same villages as the 142
index cases, and were as closely age matched as possible. Microarray analysis 143
was conducted on 15 samples (12 from cases with pneumococcal meningitis 144
and 3 from controls), and RT-PCR was conducted on 242 samples (229 from 145
cases with IPD and 13 from controls), which included all those used for 146
microarray analysis. 147
148
In the microarray discovery cohort, there were 12 children with pneumococcal 149
meningitis (6 male, 6 female, median age 1.1 years) and 3 controls (2 male, 1 150
female, median age 7 years). There were 3 cases in each of the following 151
Page 6 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
7
categories: HIV-infected survivors, HIV-infected non-survivors, HIV-uninfected 152
survivors and HIV-uninfected non-survivors (Supplementary Table 1). The RT-153
PCT validation cohort had a median age of 3.09 years. There were no known 154
co-infections apart from HIV, and we did not test for other viruses in the CSF, 155
like CMV or EBV, which have been reported to be associated with increased 156
mortality in Malawian adults with bacterial meningitis 11 12. 157
158
Samples 159
Whole blood was drawn at admission from consecutive children with IPD. The 160
methodology for downstream transcriptome analysis from small blood samples 161
has been previously described 13. 162
163
RNA extraction, quantification and hybridization 164
Total RNA was extracted from whole blood using an optimised method for the 165
PAXgeneTM blood RNA kit (Qiagen, West Sussex, UK), as previously described 166
13. The total RNA concentration (ng/µl) and ratios (260/280 and 260/230) were 167
measured using a NanoDrop ND-100 UV-vis spectrophotometer (NanoDrop 168
Technologies, Delaware, USA) and the RNA integrity was assessed using the 169
Agilent 2100 BioAnalyser (Agilent Technologies, Edinburgh, UK) before and 170
after concentration. 171
172
RNA (2µg) was reverse transcribed into cDNA using the Superscript double-173
stranded cDNA synthesis kit (Invitrogen, Paisley, UK) according to the 174
manufacturers’ guidelines. The double stranded cDNA was purified using a 175
Page 7 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
8
GeneChipTM sample clean-up module (Invitrogen, Paisley, UK). The purified 176
cDNA was then biotin labelled with the ENZO BioArray high yield RNA transcript 177
labelling kit (Affymetrix, High Wycombe, UK) and cleaned with a cRNA clean-up 178
module (Invitrogen, Paisley, UK). Aliquots of labelled cRNA (20µg) were 179
fragmented at 94°C for 35 minutes and then hybridized to a Human Genome 180
U133A GeneChipTM array for 16 hours rotating at 60rpm at 45°C in a 181
GeneChipTM Hybridization Oven 640 (Affymetrix, High Wycombe, UK). Each 182
chip was washed and stained on a GeneChipTM Fluidics Station 450 (Affymetrix, 183
High Wycombe, UK) and scanned using a GeneChipTM Scanner 3000 184
(Affymetrix, High Wycombe, UK) employing standard recommended protocols 185
(Affymetrix, High Wycombe, UK). 186
187
Microarray data analysis 188
Microarray experiment data were analysed using R and Biocondutor packages 189
14. Briefly, the human genome HGU133A array (Affymetrix, High Wycombe, UK) 190
scans output was pre-processed using the affy package 15. The limma package 191
was used to evaluate differential expressed genes 16 17. Gene annotations were 192
performed using hgu133a.db, KEGG.db database packages 18-21. Genes were 193
considered differentially expressed if they had a Benjamini and Hochberg (BH) 194
adjusted p-value < 1.5e-3 and > ±2-fold change in gene expression. Bonferroni 195
p-value adjustments were also performed for comparison. Canonical pathways 196
and functional networks that involve the differently expressed genes play were 197
determined using the Ingenuity Pathway Analysis (IPA) catalogue of well-198
characterized metabolic and cell-signaling cascades. Expression data can be 199
accessed using accession number GSE47172 at the NCBI GEO database. 200
Page 8 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
9
201
Reverse transcription for quantitative PCR 202
RNA samples were DNAse (Ambion, Warrington, UK) treated to remove any 203
contaminating genomic DNA. RNA (1µg) was reverse transcribed with 204
SuperScript IITM RNase H- Reverse Transcriptase and oligo (dT)12-18 (0.5µg/µL) 205
following the manufacturer’s guidelines. The cDNA was stored at -40°C until 206
required. 207
208
Real-Time quantitative PCR measurement of target genes 209
The Human Universal Probe Library (UPL, Roche, Switzerland) employing 210
proprietary locked nucleic acid (LNA) analogues was used to design qPCR 211
assays to measure expression levels in genes of interest. Using the Roche 212
Online Assay Design Centre, specific primers and an associated probe were 213
selected for the reference and target transcripts. Gene expression was 214
determined using real time qPCR on a Roche LightCycler 480 (Supplementary 215
Table 2). 216
217
The following 34 genes were identified from literature and prioritised for real-218
time qPCR differential expression analysis between cases and controls: ACSL1, 219
ANXA3, ATP, BAG1, BPGM, C3AR1, CA4, CD177, CD55, CD59, CEACAM, 220
CFLAR, FOLR3, GNAI3, GNLY, GYG1, IL1R2, IL1RN, ITGAM, KLRF1, LCK, 221
LCN2, LTF, MAPK14, MMP9, NUMB, OLAH, PSEN1, RETN, S100A12, 222
SAMSN, SERPINA1, SUB1, and VNN1. We used a previously described real-223
time qPCR data normalization method 22. 224
Page 9 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
10
225
Results 226
227
Transcriptional profiles among the cases and controls 228
We examined whether global transcriptional profiles of peripheral blood from 229
children with pneumococcal meningitis (n=12) were distinct from those of 230
healthy controls (n=3) randomly selected from a larger dataset by microarray 231
expression profile analysis 10. In general, there was a marked distinction in 232
differential expression between cases and controls (Supplementary Figure 1). 233
We identified 10 significantly differentially expressed genes (Benjamini & 234
Hochberg (BH) adjusted p-value < 1.5e-3 and > ±2-fold change) (Figure 1). We 235
observed significant up-regulation of: S100 calcium binding protein A12 236
(S100A12); vanin-1 (VNN1); arginase, liver (ARG1); matrix metallopeptidase 9 237
(gelatinase B, 92kDa gelatinase, 92kDa type IV collagenase) (MMP9); annexin 238
A3 (ANXA3); interleukin 1 receptor, type II (IL1R2); CD177 molecule (CD177); 239
S100 calcium binding protein A9 (S100A9), cytoskeleton-associated protein 4 240
(CKAP4); and glycogenin 1 (GYG1) supporting previous findings that have 241
highlighted the important roles of some of these genes during host pathogen 242
response. These genes have important interconnected functions for cellular 243
interactions and response to infection (Figure 3A and Supplementary Table 3), 244
and play crucial roles during host immune response; cell regulatory processes 245
such as apoptosis and differentiation; and metabolic processes such as amino 246
acid and glucose metabolism. 247
248
249
Page 10 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
11
Differential expression, underlying HIV infection and disease outcomes 250
We established the effect of HIV on host transcriptional profiles during 251
meningitis by comparing HIV-infected cases (n=6) and HIV-uninfected cases 252
(n=6). We also compared transcriptional profiles among survivors (n=6) and 253
non-survivors (n=6) to establish whether differential expression might account 254
for these two different disease outcomes. We did not find any significantly 255
differentially expressed genes for any of these comparisons (Supplementary 256
Figure 2). 257
258
RT qPCR validation 259
RT qPCR validation was performed for a set of 34 prioritised genes selected 260
from literature for cases (n=229) and controls (n=13). The RT-PCR results are in 261
agreement with our microarray analysis findings. All the genes are significantly 262
differentially expressed between the two groups with the exception of guanine 263
nucleotide-binding protein subunit alpha-13 (GNA13), protein numb (NUMB), 264
and presenilin-1 (PSEN1) (Figure 2). There was no significantly increased gene 265
expression in HIV-infected compared to HIV-uninfected cases (Supplementary 266
Figure 3). Interestingly, there was significant up-regulation of folate receptor 3 267
(FOLR3), NUMB and S100A12 in survivors compared non-survivors 268
(Supplementary Figure 4). There was wide variability in relative gene expression 269
in cases, but not controls. 270
271
Pathway analysis 272
Networks were reconstructed using IPA software for the genes differentially 273
expressed between cases and controls in the microarray experiment, and the 274
Page 11 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
12
combined microarray and RT qPCR experiments (Figure 3A and 3B). For the 275
genes defined by the microarray experiments the networks were predominantly 276
related to granulocyte function, but also including antimicrobial and endothelial 277
activation responses (Figure 3A). The merged set demonstrated wider networks 278
involving immune cell activation as well as leucocyte migration and adhesion 279
(Figure 3B). The canonical pathways mapped by genes defined in the 280
microarray experiment demonstrated greatest changes in the arginine and 281
granulocyte pathways. Canonical pathways in the combined microarray and RT 282
qPCR experiments demonstrated greatest change in leucocytes, and especially 283
neutrophils activation and migration, Notch and glucocorticoid receptor 284
signalling pathways (Supplementary Tables 3 and 4). 285
286
Discussion and conclusion 287
We have shown significant differences in RNA transcriptional profiles in children 288
with pneumococcal meningitis compared to controls. Children with 289
pneumococcal meningitis demonstrated increased expression in genes involved 290
in the inflammatory response, and glucose and L-arginine metabolic pathways. 291
Dysregulation of these pathways can lead to an impaired adaptive host 292
response to pneumococcal infection, thereby contributing to the increased 293
morbidity and mortality. Our findings in the initial cohort of pneumococcal 294
meningitis were validated in the larger cohort of all children with IPD, which 295
includes presentations with pneumonia as well as meningitis. These findings 296
add validity to the initial results from the microarray experiments, and suggest 297
that the host response observed is systemic, and not simply localised to the 298
Page 12 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
13
process of blood-brain barrier disruption per se. We observe wide variability in 299
relative gene expression in cases, which perhaps reflects differences in disease 300
onset and robustness of host response within this group. 301
302
Vanin-1 (VNN1) protein is expressed by human phagocytes, and involved in 303
leukocyte adhesion and migration 23. The VNN-1 knockout mice model has 304
provided clear evidence that VNN-1 modulates redox and immune pathways 24. 305
Exposure of human mononuclear cells to oxidative stress results in up-306
regulation of human VNN1 and down-regulation of PPARγ 25. IL1R2 and IL1RN 307
were up-regulated in cases, which is consistent with our previous report of the 308
IL-1Ra single-nucleotide polymorphism rs4251961 playing a key role in the 309
pathophysiology of IPD and in other human infections10. Recent reports also 310
demonstrate IL1R2 expression up-regulation in sepsis, being more pronounced 311
in Gram negative than Gram positive infections 26. 312
313
During infection, host production of the cytokine nitric oxide (NO) after non-314
opsonic phagocytosis, exerts microbicidal effects 27 28. ARG1 expression is 315
inducible in the lungs of mice in response to pneumococcal infection29. 316
Phagocytosis of pneumococci by macrophages also results in increased 317
production of nitric oxide synthase 2 (NOS2) dependent production of NO and 318
reactive nitrogen species 30. Increased plasma arginase activity depletes L-319
arginine concentrations, the substrate for NO synthesis, leading to vascular 320
dysfunction during severe sepsis and supressed NO-mediated microbicidal 321
effects 31. Increased ARG1 activity may also be a bacterial survival strategy to 322
escape the NO-dependent host antimicrobial immune response 30. 323
Page 13 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
14
324
Neutrophil-specific glycoprotein CD177 is expressed on a subset of human 325
neutrophils, and is involved in neutrophil transendothelial migration. A previous 326
microarray study of purified neutrophils from septic shock patients revealed 327
CD177 mRNA has the highest differential expression between cases and 328
controls 32. Consistent with our data, that study also demonstrated increased 329
expression of ARG1, ANXA3, CKAP4, IL1R2, MMP9, and VNN1. ANXA3 330
promotes endothelial cell junction integrity in animal models, and endothelial cell 331
motility in vitro. ANXA3 is up-regulated following neuronal injury, which may 332
explain the finding in pneumococcal meningitis 33. 333
334
S100A12 plays a prominent role in the regulation of proinflammatory processes 335
and immune response by recruiting leukocytes, promoting cytokine and 336
chemokine production, and regulating leukocyte adhesion and migration 34. The 337
S100A8/A9 heterodimer is expressed by myeloid cells, especially neutrophils, 338
and has a protective effect in the host response to pneumococcal infection by 339
increasing circulating neutrophils through increased granulocyte-colony 340
stimulating factor production 35. It is an antimicrobial peptide, but also plays an 341
important role in leukocyte migration 36. During infection, S100 proteins stimulate 342
the proinflammatory immune response through interaction with the 343
immunoglobulin family transmembrane pattern recognition receptors: Receptor 344
for advanced glycation endproducts (RAGE) and Toll-like receptor 4 (TLR4). 345
This leads to NF-κB-mediated pro-inflammatory response with production of 346
proinflammatory cytokines. This inflammatory response in turn leads to 347
increased expression of S100 proteins, and the start of a positive feedback loop. 348
Page 14 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
15
Although as antimicrobial proteins they protect against infection, they can also 349
have a negative detrimental effect on the host by amplifying the destructive pro-350
inflammatory responses. The increased expression in survivors may be 351
explained by this positive feedback loop 37. 352
353
Glucocorticoids also play a significant role in immune response regulation, by 354
supressing immune and inflammatory response, and modulating cytokines that 355
promote host immune responses 38. We speculate that these responses are 356
important in regulating immune responses to avoid host tissue damage. NUMB 357
negatively regulates Notch, which in turn attenuates proinflammatory cytokines 358
and increases anti-inflammatory cytokines, which may explain the increase in 359
NUMB expression in survivors 39 40. 360
361
Our results are consistent with previous studies on transcription profiling in other 362
bacterial diseases, suggesting that some of the mechanisms are not specific to 363
IPD. Although it could be argued that the lack of age matching in cases versus 364
controls could cause differential expression due to maturation of the immune 365
system in older children, the consistency of our results with other studies makes 366
this unlikely 41 42. A limitation of our study is that the microarray analysis was 367
performed using the Affymetrix Human Genome U133A array, and sample 368
number was constrained at the time by cost. The microarray dataset was not 369
large enough to allow all possible multifactorial models of co-morbidity and 370
disease outcomes to be exhaustively examined, but sought to validate our 371
findings using RT-qPCR in a larger cohort of 242 children. Further evaluation is 372
Page 15 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
16
in a larger cohort using whole genome sequencing would provide more insights 373
on the mechanisms of host response. 374
375
In conclusion, this comparative study of gene expression provides mechanistic 376
insight for IPD in children, and demonstrates significant and widespread immune 377
activation, with oxidative stress, recruitment of neutrophils, leucocyte adhesion 378
and migration, activation of antimicrobial peptides, and preservation of 379
endothelial cell junction integrity. 380
381
(2542 words) 382
383
Competing interests 384
None declared 385
386
Author contributions 387
FM, PJRD, EDC conceived and designed the study; LM, EDC collected the 388
clinical samples; FM performed experiments; BWK, FM, OV, KN, SR, PJRD, 389
EDC performed the analysis; BWK, FM, PJRD, EDC wrote the first draft of the 390
manuscript; BWK, FM, SR, LM, KN, OV, MM, EM, PJRD, EDC contributed to 391
writing the manuscript. 392
393
394
395
396
Page 16 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
17
Acknowledgements 397
398
The IPD Study Group (C Antonio, M Chinamale, L Jere, D Mnapo, V Munthali, 399
F Nyalo, J Simwinga, M Kaole, DL Banda, A Manyika, K Phiri) recruited 400
patients. We thank the children included in this study, and their parents and 401
guardians for giving consent for them to participate in the study. We also extend 402
thanks to the nursing and medical staff, at the Malawi-Liverpool-Wellcome Trust 403
Clinical Research Programme (MLW), Research Ward, for their contribution to 404
this study. EDC was supported by a Wellcome Trust Career Development Grant 405
(068026). BWK was supported by a Wellcome Trust Major Overseas 406
programme award (084679/Z/08/Z). The funding bodies had no role in 407
collection, analysis or interpretation of data or in writing the manuscript. 408
409 410 References 411 412 1. O'Brien KL, Wolfson LJ, Watt JP, et al. Burden of disease caused by 413
Streptococcus pneumoniae in children younger than 5 years: global 414 estimates. Lancet 2009;374(9693):893-902. doi: S0140-6736(09)61204-6 415 [pii] 416
10.1016/S0140-6736(09)61204-6 [published Online First: 2009/09/15] 417 2. Madhi SA, Petersen K, Madhi A, et al. Impact of human immunodeficiency 418
virus type 1 on the disease spectrum of Streptococcus pneumoniae in 419 South African children. Pediatr Infect Dis J 2000;19(12):1141-7. 420
3. Nyasulu P, Cohen C, De Gouveia L, et al. Increased risk of death in human 421 immunodeficiency virus-infected children with pneumococcal meningitis 422 in South Africa, 2003-2005. Pediatr Infect Dis J 2011;30(12):1075-80. doi: 423 10.1097/INF.0b013e31822cca05 [published Online First: 2011/07/30] 424
4. Wong HR, Cvijanovich N, Allen GL, et al. Genomic expression profiling 425 across the pediatric systemic inflammatory response syndrome, sepsis, 426 and septic shock spectrum. Crit Care Med 2009;37(5):1558-66. doi: 427 10.1097/CCM.0b013e31819fcc08 [published Online First: 2009/03/28] 428
5. Wong HR, Freishtat RJ, Monaco M, et al. Leukocyte subset-derived 429 genomewide expression profiles in pediatric septic shock. Pediatric 430 critical care medicine : a journal of the Society of Critical Care Medicine 431 and the World Federation of Pediatric Intensive and Critical Care 432
Page 17 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
18
Societies 2010;11(3):349-55. doi: 10.1097/PCC.0b013e3181c519b4 433 [published Online First: 2009/12/17] 434
6. Khoo SK, Petillo D, Parida M, et al. Host response transcriptional profiling 435 reveals extracellular components and ABC (ATP-binding cassette) 436 transporters gene enrichment in typhoid fever-infected Nigerian children. 437 BMC infectious diseases 2011;11:241. doi: 10.1186/1471-2334-11-241 438
1471-2334-11-241 [pii] [published Online First: 2011/09/15] 439 7. Hoffman O, Weber RJ. Pathophysiology and treatment of bacterial meningitis. 440
Ther Adv Neurol Disord 2009;2(6):1-7. doi: 10.1177/1756285609337975 441 [published Online First: 2010/12/25] 442
8. Glennie SJ, Williams NA, Heyderman RS. Mucosal immunity in resource-443 limited setting: is the battle ground different? Trends in microbiology 444 2010;18(11):487-93. doi: 10.1016/j.tim.2010.08.002 445
S0966-842X(10)00139-3 [pii] [published Online First: 2010/09/14] 446 9. Iwajomo OH, Finn A, Moons P, et al. Deteriorating pneumococcal-specific B-447
cell memory in minimally symptomatic African children with HIV infection. 448 The Journal of infectious diseases 2011;204(4):534-43. doi: jir316 [pii] 449
10.1093/infdis/jir316 [published Online First: 2011/07/28] 450 10. Carrol ED, Payton A, Payne D, et al. The IL1RN promoter rs4251961 451
correlates with IL-1 receptor antagonist concentrations in human infection 452 and is differentially regulated by GATA-1. J Immunol 2011;186(4):2329-453 35. doi: 10.4049/jimmunol.1002402 454
11. Kelly MJ, Benjamin LA, Cartwright K, et al. Epstein-barr virus coinfection in 455 cerebrospinal fluid is associated with increased mortality in Malawian 456 adults with bacterial meningitis. J Infect Dis 2012;205(1):106-10. doi: 457 10.1093/infdis/jir707 458
jir707 [pii] [published Online First: 2011/11/15] 459 12. Benjamin LA, Kelly M, Cohen D, et al. Detection of herpes viruses in the 460
cerebrospinal fluid of adults with suspected viral meningitis in Malawi. 461 Infection 2013;41(1):27-31. doi: 10.1007/s15010-012-0292-z [published 462 Online First: 2012/07/17] 463
13. Carrol ED, Salway F, Pepper SD, et al. Successful downstream application 464 of the Paxgene Blood RNA system from small blood samples in 465 paediatric patients for quantitative PCR analysis. BMC Immunol 466 2007;8:20. doi: 10.1186/1471-2172-8-20 467
14. Gentleman RC, Carey VJ, Bates DM, et al. Bioconductor: open software 468 development for computational biology and bioinformatics. Genome 469 biology 2004;5(10):R80.1-R80.16. 470
15. Irizarry R. From CEL files to annotated lists of interesting genes. In: 471 Gentleman R. CV, Dudoit S., Irizarry R., Huber W., ed. Bioinformatics 472 and Computational Biology Solutions using R and Bioconductor. New 473 York: Springer 2005:431-42. 474
16. Smyth GK. Limma: linear models for microarray data. In: Gentleman R. CV, 475 Dudoit S., Irizarry R., Huber W., ed. Bioinformatics and Computational 476 Biology Solutions using R and Bioconductor. New York: Springer 477 2005:397-420. 478
Page 18 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
19
17. Smyth GK. Linear models and empirical Bayes methods for assessing 479 differential expression in microarray experiments. Statistical Applications 480 in Genetics and Molecular Biology 2004;3(1):3. 481
18. hgu133a.db: Affymetrix Human Genome U133 Set annotation data (chip 482 hgu133a) [program]. R package version 2.7.1 version, 2009. 483
19. annotate: Annotation for microarrays. R package version 1.34.1. [program]. 484 R package version 1.34.1. version, 2008. 485
20. GO.db: A set of annotation maps describing the entire Gene Ontology 486 [program]. R package version 2.12 version, 2013. 487
21. KEGG.db: A set of annotation maps for KEGG [program]. R package version 488 2.7.1 version, 2009. 489
22. Vandesompele J, De Preter K, Pattyn F, et al. Accurate normalization of 490 real-time quantitative RT-PCR data by geometric averaging of multiple 491 internal control genes. Genome Biol 2002;3(7):RESEARCH0034. 492
23. Meghari S, Berruyer C, Lepidi H, et al. Vanin-1 controls granuloma formation 493 and macrophage polarization in Coxiella burnetii infection. Eur J Immunol 494 2007;37(1):24-32. doi: 10.1002/eji.200636054 495
24. Ferreira DW, Naquet P, Manautou JE. Influence of Vanin-1 and Catalytic 496 Products in Liver During Normal and Oxidative Stress Conditions. Curr 497 Med Chem 2015;22(20):2407-16. 498
25. Zhang B, Lo C, Shen L, et al. The role of vanin-1 and oxidative stress-499 related pathways in distinguishing acute and chronic pediatric ITP. Blood 500 2011;117(17):4569-79. doi: 10.1182/blood-2010-09-304931 501
26. Lang Y, Jiang Y, Gao M, et al. Interleukin-1 Receptor 2: A New Biomarker 502 for Sepsis Diagnosis and Gram-Negative/Gram-Positive Bacterial 503 Differentiation. Shock 2017;47(1):119-24. doi: 504 10.1097/SHK.0000000000000714 505
27. Ulett GC, Adderson EE. Nitric oxide is a key determinant of group B 506 streptococcus-induced murine macrophage apoptosis. The Journal of 507 infectious diseases 2005;191(10):1761-70. doi: 10.1086/429693 508 [published Online First: 2005/04/20] 509
28. Gross A, Spiesser S, Terraza A, et al. Expression and bactericidal activity of 510 nitric oxide synthase in Brucella suis-infected murine macrophages. 511 Infection and immunity 1998;66(4):1309-16. [published Online First: 512 1998/04/07] 513
29. Knippenberg S, Brumshagen C, Aschenbrenner F, et al. Arginase 1 activity 514 worsens lung-protective immunity against Streptococcus pneumoniae 515 infection. Eur J Immunol 2015;45(6):1716-26. doi: 10.1002/eji.201445419 516
30. Marriott HM, Ali F, Read RC, et al. Nitric oxide levels regulate macrophage 517 commitment to apoptosis or necrosis during pneumococcal infection. 518 FASEB J 2004;18(10):1126-8. doi: 10.1096/fj.03-1450fje 519
31. Darcy CJ, Woodberry T, Davis JS, et al. Increased plasma arginase activity 520 in human sepsis: association with increased circulating neutrophils. 521 Clinical chemistry and laboratory medicine : CCLM / FESCC 522 2014;52(4):573-81. doi: 10.1515/cclm-2013-0698 [published Online First: 523 2013/10/30] 524
32. Demaret J, Venet F, Plassais J, et al. Identification of CD177 as the most 525 dysregulated parameter in a microarray study of purified neutrophils from 526
Page 19 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
20
septic shock patients. Immunol Lett 2016;178:122-30. doi: 527 10.1016/j.imlet.2016.08.011 528
33. Chong KW, Chen MJ, Koay ES, et al. Annexin A3 is associated with cell 529 death in lactacystin-mediated neuronal injury. Neurosci Lett 530 2010;485(2):129-33. doi: 10.1016/j.neulet.2010.08.089 531
34. Goyette J, Geczy CL. Inflammation-associated S100 proteins: new 532 mechanisms that regulate function. Amino Acids 2011;41(4):821-42. doi: 533 10.1007/s00726-010-0528-0 534
35. De Filippo K, Neill DR, Mathies M, et al. A new protective role for S100A9 in 535 regulation of neutrophil recruitment during invasive pneumococcal 536 pneumonia. FASEB J 2014;28(8):3600-8. doi: 10.1096/fj.13-247460 537
36. Raquil MA, Anceriz N, Rouleau P, et al. Blockade of antimicrobial proteins 538 S100A8 and S100A9 inhibits phagocyte migration to the alveoli in 539 streptococcal pneumonia. J Immunol 2008;180(5):3366-74. 540
37. Zackular JP, Chazin WJ, Skaar EP. Nutritional Immunity: S100 Proteins at 541 the Host-Pathogen Interface. The Journal of biological chemistry 542 2015;290(31):18991-8. doi: 10.1074/jbc.R115.645085 543
38. Sakai A, Han J, Cato AC, et al. Glucocorticoids synergize with IL-1beta to 544 induce TLR2 expression via MAP Kinase Phosphatase-1-dependent dual 545 Inhibition of MAPK JNK and p38 in epithelial cells. BMC molecular 546 biology 2004;5:2. doi: 10.1186/1471-2199-5-2 [published Online First: 547 2004/05/06] 548
39. Kueanjinda P, Roytrakul S, Palaga T. A Novel Role of Numb as A Regulator 549 of Pro-inflammatory Cytokine Production in Macrophages in Response to 550 Toll-like Receptor 4. Scientific reports 2015;5:12784. doi: 551 10.1038/srep12784 552
40. Singla DK, Wang J, Singla R. Primary human monocytes differentiate into 553 M2 macrophages and involve Notch-1 pathway. Can J Physiol Pharmacol 554 2017;95(3):288-94. doi: 10.1139/cjpp-2016-0319 555
41. Langley RJ, Tsalik EL, Velkinburgh JC, et al. An integrated clinico-556 metabolomic model improves prediction of death in sepsis. Sci Transl 557 Med 2013;5(195):195ra95. doi: 10.1126/scitranslmed.3005893 558
5/195/195ra95 [pii] [published Online First: 2013/07/26] 559 42. Liu K, Chen L, Kaur R, et al. Transcriptome signature in young children with 560
acute otitis media due to Streptococcus pneumoniae. Microbes Infect 561 2012;14(7-8):600-9. doi: 10.1016/j.micinf.2012.01.006 562
S1286-4579(12)00035-4 [pii] [published Online First: 2012/02/07] 563
564
Page 20 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
21
Figure and Table legends 565
566
Figure 1: Distribution plot of the differentially expressed genes. The 567
significantly differentially expressed genes are shown in red colour. The 568
significance threshold (P<1.5e-3) is indicated by a dashed red line, and a fold 569
change threshold of more than 2 is shown by the dashed vertical lines. The 570
green line shows P<0.05. (A) Shows results for un-adjusted p-values, (B) 571
results for Benjamini and Hochberg (BH) adjusted p-values, and (C) stringent 572
Bonferroni adjusted p-values, which represents an over-correction. 573
574
Figure 2: Validation of RNA transcription profile differential expression 575
using Real Time quantitative PCR. Relative gene expression in cases 576
compared to controls. The black line shows the boxplot median. The red dot 577
shows the mean, and the Welch two-sample t-test p-value is shown on the top 578
right corner. 579
Figure 3: The gene network for significantly differentially expressed 580
genes in the cases. Networks reconstructed to support the most direct 581
connectivity between genes differentially expressed between cases and 582
controls. IPA (Qiagen) software's Core analysis application has been used to 583
perform an automatic graphical reconstruction of the network via utilisation of 584
IPA's Knowledge base database of protein interactions. The meaning of links 585
and shapes is explained in the inserted legend. Functional connections are 586
presented in blue, information connections to the associated categories in 587
pink and grey. (A) Network reconstructed only for the DE genes identified by 588
Page 21 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
22
microarray analysis. (B) Network reconstructed for the merged datasets of DE 589
genes identified via microarray (red) and PCR analysis (green). White blocks 590
correspond to nodes added by IPA's network editor automatically to ensure 591
network connectivity. 592
593
Supplementary Figure 1: Children with pneumococcal meningitis and 594
healthy controls have distinct transcriptional profiles. A heatmap of the 595
top 100 (P<0.05) differentially expressed genes compared in cases and 596
controls. The cases and controls were distinguished into two distinct clusters 597
highlighting their distinct transcriptional profiles. 598
599
600
Supplementary Figure 2: Comparison of microarray transcriptional 601
profiles of HIV infected and HIV-uninfected and survivors and non-602
survivors. There were no significantly differentially expressed genes for any 603
of these comparisons 604
605
Supplementary Figure 3: Comparison of RT-qPCT gene expression for 606
HIV infected and HIV-uninfected. There were no significantly differentially 607
expressed genes for all of these comparisons. 608
609
Supplementary Figure 4: Comparison of RT-qPCT gene expression for 610
survivors and non-survivors. There were three significantly differentially 611
expressed genes: FOLR3, NUMB, and S100A12. 612
Page 22 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
23
613
Supplementary Table 1: Clinical data on children with pneumococcal 614
meningitis and healthy controls. 615
616
Supplementary Table 2: Primer sequences and the UPL probe number 617
for each RT qPCR assay. 618
619
Supplementary Table 3: Canonical pathways of differentially expressed 620
genes between cases and controls in microarray experiment 621
622
Supplementary Table 4: Canonical pathways of differentially expressed 623
genes between cases and controls in combined microarray and RT- 624
qPCR experiments 625
626
627
628
629
Page 23 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
94x24mm (300 x 300 DPI)
Page 24 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
177x177mm (300 x 300 DPI)
Page 25 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
361x481mm (300 x 300 DPI)
Page 26 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
270x203mm (300 x 300 DPI)
Page 27 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
117x42mm (300 x 300 DPI)
Page 28 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
177x177mm (300 x 300 DPI)
Page 29 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
177x177mm (300 x 300 DPI)
Page 30 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from
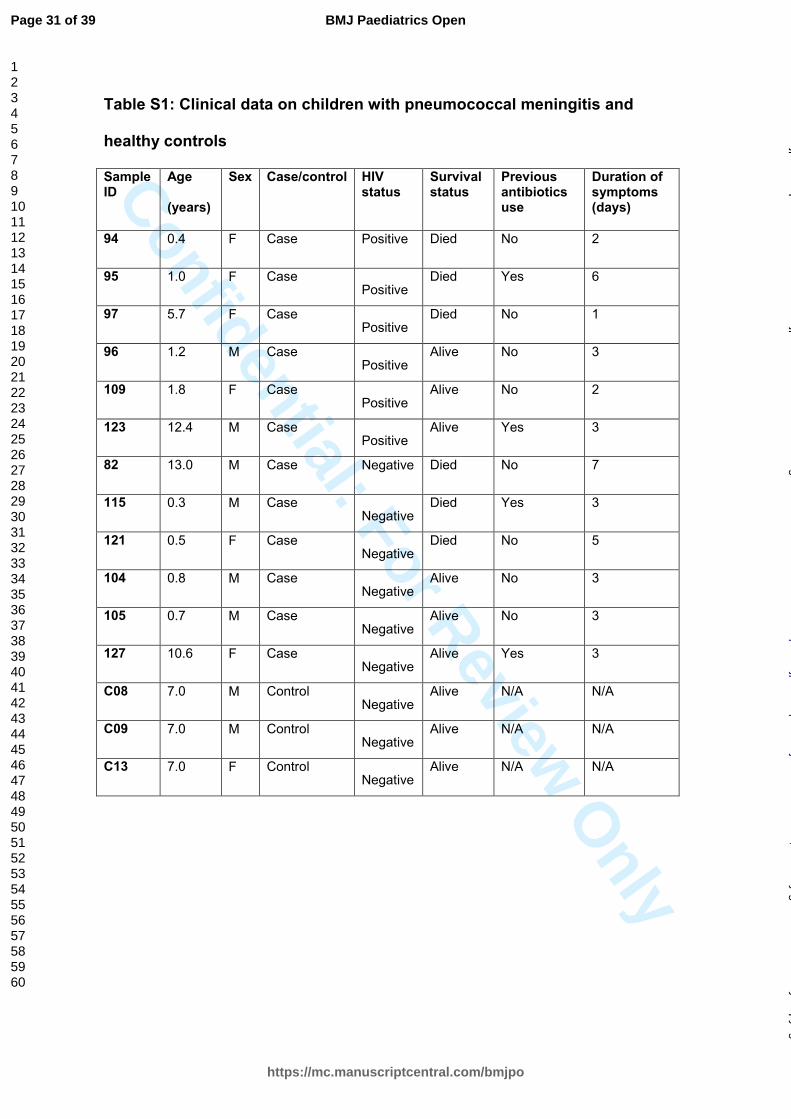
Confidential: For Review O
nlyTable S1: Clinical data on children with pneumococcal meningitis and
healthy controls
Sample ID
Age
(years)
Sex Case/control HIV status
Survival status
Previous antibiotics use
Duration of symptoms (days)
94 0.4 F Case Positive Died No 2
95 1.0 F Case Positive
Died Yes 6
97 5.7 F Case Positive
Died No 1
96 1.2 M Case Positive
Alive No 3
109 1.8 F Case Positive
Alive No 2
123 12.4 M Case Positive
Alive Yes 3
82 13.0 M Case Negative Died No 7
115 0.3 M Case Negative
Died Yes 3
121 0.5 F Case Negative
Died No 5
104 0.8 M Case Negative
Alive No 3
105 0.7 M Case Negative
Alive No 3
127 10.6 F Case Negative
Alive Yes 3
C08 7.0 M Control Negative
Alive N/A N/A
C09 7.0 M Control Negative
Alive N/A N/A
C13 7.0 F Control Negative
Alive N/A N/A
Page 31 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only1
Table S2: Primer sequences and the UPL probe number for each RT qPCR assay
Gene Symbol Gene Forward Primer (5'-3') Reverse Primer (5'-3') UPL Probe Accession Number
ACSL1 Acyl-CoA synthetase long-chain family member 1 ccactgtgctgacgttcct gcactctgtctgtccgtatcc 26 NM_001995
ANXA3 Annexin A3 ttgttaaggaatatcaagcagcat gccagagagatcacccttca 17 NM_005139
C4A Complement component 4A (Rodgers blood group) ggaccctgaagacgaaattg gaagccctcgttcacctg 75 NM_007293, NM_001252204
C3AR1 complement component 3a receptor 1
aagaaagcaaggcagtccatt
cgtgtgagctcctcactgaa
16
NM_004054
ILR2A ILR2A interleukin 1 receptor, type II tacgcaccacagtcaaggaa aagaaggccagtgaaagtgg 2 NM_004633, NM_173343
MAPK14
MAPK 14 mitogen-activated protein kinase 14
MAPK 14 mitogen-activated protein kinase 14
ccatgagatgggtcaccag
65
NM_139012, NM_139014, NM_001315, NM_139013
MMP9
matrix metallopeptidase 9 (gelatinase B, 92kDa gelatinase, 92kDa type IV collagenase)
gaaccaatctcaccgacagg
gaaccaatctcaccgacagg
6
NM_004994
CD177 CD177 molecule gcctctccctgatctcctaca caaggatcctgggtctgc 79 NM_020406
CD55 CD55 molecule, decay accelerating factor for complement (Cromer blood group) aataatgatgaaggagagtggagtg tggtgggaccttggaagtta 77 NM_000574
CEACAM1 Carcinoembryonic antigen-related cell adhesion molecule 1 (biliary glycoprotein) taggcctaagaggctttctcc tgagggaaagtatgcacagtccgtgtc 80
NM_001712, NM_001184816, NM_001184813, NM_001184815, NM_001205344, NM_001024912
GYG1 Glycogenin 1 ctgaaacagcacaggaccac atcgccactgtccaagacat 75 NM_004130
FOLR3 Folate receptor 3 (gamma) cacaaaggctggaattggac cttgaaggagtggctccaga 19 NM_000804
ITGAM Integrin, alpha M (complement component 3 receptor 3 subunit) tctgaagaccattcagaaccag ggagctgctacttcctgtctg 7 NM_000632
LTF Lactotransferrin ctaatctgaaaaagtgctcaacctc gccatcttcttcggttttacttc 79 NM_002343, NM_001199149
OLAH Oleoyl-ACP hydrolase accgagatcgctccactg ggagaatcaagtgagattttaatagga 19 NM_018324, NM_001039702
RETN Resistin tgcaggatgaaagctctctg catggagcacagggtcttg 45 NM_020415
Page 32 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only2
Gene Symbol Gene Forward Primer (5'-3') Reverse Primer (5'-3') UPL Probe Accession Number
S100A12 S100 calcium binding protein A12 tcatatccctggtagccattg acctactctttgtgggtgtggt 44 NM_005621
SERPINA1 Serpin peptidase inhibitor, clade A (alpha-1 antiproteinase, antitrypsin), member 1 aatggggctgacctctcc gtcagcacagccttatgcac 82 NM_001002235, NM_001002236,
NM_000295
SAMSN1 SAM domain, SH3 domain and nuclear localization signals 1 tgaaaatctgtctgacatggtaca tagttgggaatgcgtgttca 21 NM_022136, NM_001256370
VNN1 Vanin 1 tcctgaggtgttgctgagtg agcgtccgtcagttgacac 80 NM_004666
CD59 CD59 molecule, complement regulatory protein caaggagggtctgtcctgtt
agcactgcaggct[a or g]tgacc 66
NM_000611, NM_203331, NM_203330, NM_203329
CFLAR CASP8 and FADD-like apoptosis regulator gcaatccaaaagagtctcaagg
tgagcgccaagctgttcc 7 NM_003879
KLRF1 Killer cell lectin-like receptor subfamily F, member 1
tgatctccttgatcctgttgg
tcttcttgtgccattattcacttt 47 NM_016523
LCN2 Homo sapiens lipocalin 2 tcacctccgtcctgtttagg
aggtaactcgttaatccagggtaa 61 NM_005564
NUMB Numb homolog (Drosophila) gaggaagactgatttccccatt
gcacagggagctgatgctc 16
NM_001005745, NM_003744, NM_001005744, NM_001005743
Page 33 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Supplementary Table 3: Canonical pathways of differentially expressedgenes between cases and controls in microarray experiment Ingenuity Canonical Pathways -log(p-value) Ratio MoleculesLXR/RXR Activation 2.77 0.0165 IL1R2,MMP9Arginine Degradation I (Arginase Pathway) 2.69 0.25 ARG1Urea Cycle 2.51 0.167 ARG1Arginine Degradation VI (Arginase 2 Pathway) 2.51 0.167 ARG1Glycogen Biosynthesis II (from UDP-D-Glucose) 2.51 0.167 GYG1Granulocyte Adhesion and Diapedesis 2.51 0.0121 IL1R2,MMP9Hepatic Fibrosis / Hepatic Stellate Cell Activation 2.43 0.011 IL1R2,MMP9Airway Pathology in Chronic Obstructive Pulmonary Disease 2.38 0.125 MMP9Citrulline Biosynthesis 2.38 0.125 ARG1Role of IL-17A in Psoriasis 2.17 0.0769 S100A9Superpathway of Citrulline Metabolism 2.14 0.0714 ARG1Inhibition of Angiogenesis by TSP1 1.79 0.0312 MMP9Inhibition of Matrix Metalloproteases 1.71 0.0263 MMP9Neuroprotective Role of THOP1 in Alzheimer's Disease 1.69 0.025 MMP9IL-10 Signaling 1.46 0.0147 IL1R2Glioma Invasiveness Signaling 1.45 0.0143 MMP9Bladder Cancer Signaling 1.36 0.0116 MMP9PPAR Signaling 1.34 0.0111 IL1R2HIF1α Signaling 1.25 0.00885 MMP9p38 MAPK Signaling 1.23 0.00855 IL1R2Pancreatic Adenocarcinoma Signaling 1.23 0.00847 MMP9Atherosclerosis Signaling 1.21 0.00806 MMP9IL-6 Signaling 1.2 0.00787 IL1R2Ovarian Cancer Signaling 1.15 0.00699 MMP9Relaxin Signaling 1.13 0.00671 MMP9Hepatic Cholestasis 1.11 0.00637 IL1R2PPARα/RXRα Activation 1.08 0.00602 IL1R2Agranulocyte Adhesion and Diapedesis 1.06 0.00571 MMP9NF-κB Signaling 1.06 0.00565 IL1R2Regulation of the Epithelial-Mesenchymal Transition Pathway 1.03 0.00535 MMP9ILK Signaling 1.02 0.00521 MMP9IL-8 Signaling 1.01 0.0051 MMP9Leukocyte Extravasation Signaling 0.996 0.00488 MMP9LPS/IL-1 Mediated Inhibition of RXR Function 0.99 0.00481 IL1R2Role of Osteoblasts, Osteoclasts and Chondrocytes in Rheumatoid Arthritis 0.954 0.00441 IL1R2Colorectal Cancer Metastasis Signaling 0.928 0.00413 MMP9Glucocorticoid Receptor Signaling 0.862 0.00352 IL1R2Role of Macrophages, Fibroblasts and Endothelial Cells in Rheumatoid Arthritis 0.84 0.00333 IL1R2Axonal Guidance Signaling 0.686 0.00226 MMP9
Page 34 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Ingenuity Canonical Pathways -log(p-value) Ratio MoleculesComplement System 6.35 0.111 CD55,CD59,ITGAM,C3AR1Granulocyte Adhesion and Diapedesis 5.02 0.0303 IL1R2,GNAI3,ITGAM,IL1RN,MMP9LXR/RXR Activation 4.24 0.0331 IL1R2,IL1RN,SERPINA1,MMP9IL-10 Signaling 3.64 0.0441 IL1R2,MAPK14,IL1RNLeukocyte Extravasation Signaling 3.36 0.0195 GNAI3,MAPK14,ITGAM,MMP9p38 MAPK Signaling 2.95 0.0256 IL1R2,MAPK14,IL1RNAtherosclerosis Signaling 2.88 0.0242 IL1RN,SERPINA1,MMP9IL-6 Signaling 2.85 0.0236 IL1R2,MAPK14,IL1RNInhibition of Angiogenesis by TSP1 2.84 0.0625 MAPK14,MMP9Glucocorticoid Receptor Signaling 2.83 0.0141 IL1R2,MAPK14,BAG1,IL1RNIL-17A Signaling in Fibroblasts 2.77 0.0571 MAPK14,LCN2Notch Signaling 2.72 0.0541 NUMB,PSEN1Acute Phase Response Signaling 2.5 0.0179 MAPK14,IL1RN,SERPINA1Amyloid Processing 2.46 0.04 MAPK14,PSEN1Agranulocyte Adhesion and Diapedesis 2.45 0.0171 GNAI3,IL1RN,MMP9NF-κB Signaling 2.44 0.0169 IL1R2,LCK,IL1RNMolecular Mechanisms of Cancer 2.42 0.0109 GNAI3,MAPK14,CFLAR,PSEN1IL-8 Signaling 2.31 0.0153 GNAI3,ITGAM,MMP9LPS/IL-1 Mediated Inhibition of RXR Function 2.24 0.0144 IL1R2,IL1RN,ACSL1CCR5 Signaling in Macrophages 2.21 0.0299 GNAI3,MAPK14Chemokine Signaling 2.2 0.0294 GNAI3,MAPK14Caveolar-mediated Endocytosis Signaling 2.16 0.0282 CD55,ITGAMArginine Degradation I (Arginase Pathway) 2.15 0.25 ARG1Acetate Conversion to Acetyl-CoA 2.15 0.25 ACSL1Toll-like Receptor Signaling 2.14 0.0274 MAPK14,IL1RNRole of Osteoblasts, Osteoclasts and Chondrocytes in Rheumatoid Arthritis 2.14 0.0132 IL1R2,MAPK14,IL1RNIL-15 Signaling 2.1 0.0263 LCK,MAPK14Rapoport-Luebering Glycolytic Shunt 2.06 0.2 BPGMPEDF Signaling 2.02 0.0238 MAPK14,CFLARUrea Cycle 1.98 0.167 ARG1Arginine Degradation VI (Arginase 2 Pathway) 1.98 0.167 ARG1Glycogen Biosynthesis II (from UDP-D-Glucose) 1.98 0.167 GYG1PPAR Signaling 1.96 0.0222 IL1R2,IL1RNIL-1 Signaling 1.95 0.022 GNAI3,MAPK14Cholecystokinin/Gastrin-mediated Signaling 1.87 0.0198 MAPK14,IL1RNAirway Pathology in Chronic Obstructive Pulmonary Disease 1.86 0.125 MMP9Citrulline Biosynthesis 1.86 0.125 ARG1Corticotropin Releasing Hormone Signaling 1.82 0.0187 GNAI3,MAPK14Role of Macrophages, Fibroblasts and Endothelial Cells in Rheumatoid Arthritis 1.81 0.01 IL1R2,MAPK14,IL1RNPaxillin Signaling 1.8 0.0182 MAPK14,ITGAMHIF1α Signaling 1.78 0.0177 MAPK14,MMP9Role of Tissue Factor in Cancer 1.73 0.0168 LCK,MAPK14FXR/RXR Activation 1.69 0.016 IL1RN,SERPINA1CCR3 Signaling in Eosinophils 1.69 0.0159 GNAI3,MAPK14
Supplementary Table 4: Canonical pathways of differentially expressed genes between cases and controls in combined microarrayand RT- qPCR experiments
Page 35 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
GNRH Signaling 1.68 0.0157 GNAI3,MAPK14Role of IL-17A in Psoriasis 1.65 0.0769 S100A9Fatty Acid Activation 1.65 0.0769 ACSL1Superpathway of Citrulline Metabolism 1.61 0.0714 ARG1IL-12 Signaling and Production in Macrophages 1.58 0.0139 MAPK14,SERPINA1Parkinson's Signaling 1.56 0.0625 MAPK14Relaxin Signaling 1.55 0.0134 GNAI3,MMP9γ-linolenate Biosynthesis II (Animals) 1.53 0.0588 ACSL1Mitochondrial L-carnitine Shuttle Pathway 1.53 0.0588 ACSL1Oxidative Ethanol Degradation III 1.53 0.0588 ACSL1Hepatic Cholestasis 1.51 0.0127 IL1R2,IL1RNPPARα/RXRα Activation 1.47 0.012 IL1R2,MAPK14Tec Kinase Signaling 1.46 0.0119 GNAI3,LCKEthanol Degradation IV 1.44 0.0476 ACSL1Differential Regulation of Cytokine Production in Intestinal Epithelial Cells by IL-17A and IL-17F1.4 0.0435 LCN2Role of NFAT in Regulation of the Immune Response 1.4 0.0111 GNAI3,LCKHepatic Fibrosis / Hepatic Stellate Cell Activation 1.4 0.011 IL1R2,MMP9Dendritic Cell Maturation 1.39 0.011 MAPK14,IL1RNEndothelin-1 Signaling 1.39 0.0109 GNAI3,MAPK14IL-22 Signaling 1.38 0.0417 MAPK14Glycolysis I 1.38 0.0417 BPGMRegulation of the Epithelial-Mesenchymal Transition Pathway 1.37 0.0107 MMP9,PSEN1IL-17A Signaling in Gastric Cells 1.37 0.04 MAPK14Role of JAK family kinases in IL-6-type Cytokine Signaling 1.37 0.04 MAPK14Gluconeogenesis I 1.37 0.04 BPGMRole of NFAT in Cardiac Hypertrophy 1.36 0.0106 GNAI3,MAPK14Production of Nitric Oxide and Reactive Oxygen Species in Macrophages 1.35 0.0104 MAPK14,SERPINA1Clathrin-mediated Endocytosis Signaling 1.33 0.0102 NUMB,SERPINA1Thrombin Signaling 1.32 0.01 GNAI3,MAPK14Fatty Acid β-oxidation I 1.29 0.0333 ACSL14-1BB Signaling in T Lymphocytes 1.28 0.0323 MAPK14G Protein Signaling Mediated by Tubby 1.28 0.0323 LCKEthanol Degradation II 1.26 0.0312 ACSL1Systemic Lupus Erythematosus Signaling 1.26 0.00922 LCK,IL1RNCoagulation System 1.22 0.0286 SERPINA1Cardiac Hypertrophy Signaling 1.2 0.00862 GNAI3,MAPK14April Mediated Signaling 1.19 0.0263 MAPK14Inhibition of Matrix Metalloproteases 1.19 0.0263 MMP9Role of PKR in Interferon Induction and Antiviral Response 1.17 0.025 MAPK14B Cell Activating Factor Signaling 1.17 0.025 MAPK14Neuroprotective Role of THOP1 in Alzheimer's Disease 1.17 0.025 MMP9Stearate Biosynthesis I (Animals) 1.17 0.025 ACSL1Role of Hypercytokinemia/hyperchemokinemia in the Pathogenesis of Influenza 1.16 0.0244 IL1RNUVC-Induced MAPK Signaling 1.15 0.0238 MAPK14iNOS Signaling 1.14 0.0233 MAPK14Primary Immunodeficiency Signaling 1.13 0.0227 LCKGraft-versus-Host Disease Signaling 1.13 0.0227 IL1RN
Page 36 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Role of Cytokines in Mediating Communication between Immune Cells 1.06 0.0192 IL1RNRetinoic acid Mediated Apoptosis Signaling 1.01 0.0172 CFLARMSP-RON Signaling Pathway 1.01 0.0169 ITGAMCalcium-induced T Lymphocyte Apoptosis 0.999 0.0167 LCKIL-2 Signaling 0.973 0.0156 LCKUVB-Induced MAPK Signaling 0.96 0.0152 MAPK14Role of IL-17A in Arthritis 0.954 0.0149 MAPK14EGF Signaling 0.948 0.0147 MAPK14Melatonin Signaling 0.942 0.0145 GNAI3Glioma Invasiveness Signaling 0.936 0.0143 MMP9Role of MAPK Signaling in the Pathogenesis of Influenza 0.936 0.0143 MAPK14ErbB4 Signaling 0.936 0.0143 PSEN1GPCR-Mediated Integration of Enteroendocrine Signaling Exemplified by an L Cell 0.93 0.0141 GNAI3Role of PI3K/AKT Signaling in the Pathogenesis of Influenza 0.919 0.0137 GNAI3Ephrin B Signaling 0.919 0.0137 GNAI3STAT3 Pathway 0.919 0.0137 MAPK14BMP signaling pathway 0.913 0.0135 MAPK14CD40 Signaling 0.897 0.013 MAPK14IL-17A Signaling in Airway Cells 0.897 0.013 MAPK14ATM Signaling 0.887 0.0127 MAPK14FLT3 Signaling in Hematopoietic Progenitor Cells 0.872 0.0122 MAPK14Communication between Innate and Adaptive Immune Cells 0.872 0.0122 IL1RNAltered T Cell and B Cell Signaling in Rheumatoid Arthritis 0.867 0.012 IL1RNGPCR-Mediated Nutrient Sensing in Enteroendocrine Cells 0.862 0.0119 GNAI3α-Adrenergic Signaling 0.857 0.0118 GNAI3IL-17 Signaling 0.857 0.0118 MAPK14Neuregulin Signaling 0.852 0.0116 PSEN1LPS-stimulated MAPK Signaling 0.852 0.0116 MAPK14NF-κB Activation by Viruses 0.852 0.0116 LCKBladder Cancer Signaling 0.852 0.0116 MMP9TGF-β Signaling 0.848 0.0115 MAPK14G Beta Gamma Signaling 0.843 0.0114 GNAI3Factors Promoting Cardiogenesis in Vertebrates 0.838 0.0112 MAPK14FGF Signaling 0.834 0.0111 MAPK14Death Receptor Signaling 0.83 0.011 CFLARReelin Signaling in Neurons 0.825 0.0109 LCKErbB Signaling 0.804 0.0103 MAPK14CTLA4 Signaling in Cytotoxic T Lymphocytes 0.8 0.0102 LCKCDK5 Signaling 0.8 0.0102 MAPK14RANK Signaling in Osteoclasts 0.796 0.0101 MAPK14Antioxidant Action of Vitamin C 0.796 0.0101 MAPK14UVA-Induced MAPK Signaling 0.788 0.0099 MAPK14Virus Entry via Endocytic Pathways 0.784 0.0098 CD55SAPK/JNK Signaling 0.78 0.00971 LCKMouse Embryonic Stem Cell Pluripotency 0.772 0.00952 MAPK14Type I Diabetes Mellitus Signaling 0.769 0.00943 MAPK14T Cell Receptor Signaling 0.761 0.00926 LCK
Page 37 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Androgen Signaling 0.754 0.00909 GNAI3p53 Signaling 0.751 0.00901 MAPK14Axonal Guidance Signaling 0.741 0.00452 GNAI3,MMP9iCOS-iCOSL Signaling in T Helper Cells 0.73 0.00855 LCKPancreatic Adenocarcinoma Signaling 0.726 0.00847 MMP9Fc Epsilon RI Signaling 0.723 0.0084 MAPK14Renin-Angiotensin Signaling 0.723 0.0084 MAPK14Natural Killer Cell Signaling 0.72 0.00833 LCKfMLP Signaling in Neutrophils 0.72 0.00833 GNAI3Gαi Signaling 0.72 0.00833 GNAI3Sphingosine-1-phosphate Signaling 0.713 0.0082 GNAI3Type II Diabetes Mellitus Signaling 0.704 0.008 ACSL1CD28 Signaling in T Helper Cells 0.701 0.00794 LCKPKCθ Signaling in T Lymphocytes 0.698 0.00787 LCKCdc42 Signaling 0.692 0.00775 MAPK14p70S6K Signaling 0.689 0.00769 GNAI3Th1 Pathway 0.689 0.00769 PSEN1Role of Pattern Recognition Receptors in Recognition of Bacteria and Viruses 0.686 0.00763 C3AR1HMGB1 Signaling 0.686 0.00763 MAPK14P2Y Purigenic Receptor Signaling Pathway 0.686 0.00763 GNAI3Synaptic Long Term Depression 0.66 0.00714 GNAI3Ovarian Cancer Signaling 0.652 0.00699 MMP9Th2 Pathway 0.644 0.00685 PSEN1Regulation of eIF4 and p70S6K Signaling 0.624 0.00649 MAPK14Dopamine-DARPP32 Feedback in cAMP Signaling 0.614 0.00633 GNAI3CXCR4 Signaling 0.6 0.0061 GNAI3Mitochondrial Dysfunction 0.598 0.00606 PSEN1Gap Junction Signaling 0.595 0.00602 GNAI3Germ Cell-Sertoli Cell Junction Signaling 0.589 0.00592 MAPK14RhoGDI Signaling 0.582 0.00581 GNAI3Ephrin Receptor Signaling 0.582 0.00581 GNAI3Sertoli Cell-Sertoli Cell Junction Signaling 0.58 0.00578 MAPK14Th1 and Th2 Activation Pathway 0.565 0.00556 PSEN1CREB Signaling in Neurons 0.561 0.00549 GNAI3B Cell Receptor Signaling 0.561 0.00549 MAPK14RAR Activation 0.551 0.00535 MAPK14AMPK Signaling 0.549 0.00532 MAPK14NRF2-mediated Oxidative Stress Response 0.545 0.00526 MAPK14ILK Signaling 0.541 0.00521 MMP93-phosphoinositide Biosynthesis 0.539 0.00518 LCKBreast Cancer Regulation by Stathmin1 0.523 0.00495 GNAI3Integrin Signaling 0.505 0.00472 ITGAMcAMP-mediated signaling 0.492 0.00455 GNAI3Superpathway of Inositol Phosphate Compounds 0.481 0.00441 LCKPhospholipase C Signaling 0.47 0.00427 LCKColorectal Cancer Metastasis Signaling 0.458 0.00413 MMP9Signaling by Rho Family GTPases 0.453 0.00407 GNAI3
Page 38 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Protein Ubiquitination Pathway 0.441 0.00394 BAG1Xenobiotic Metabolism Signaling 0.422 0.00372 MAPK14G-Protein Coupled Receptor Signaling 0.42 0.0037 GNAI3Protein Kinase A Signaling 0.313 0.00266 GNAI3
Page 39 of 39
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
Peripheral blood RNA gene expression in children with
pneumococcal meningitis; a prospective case control study
Journal: BMJ Paediatrics Open
Manuscript ID bmjpo-2017-000092.R1
Article Type: Original article
Date Submitted by the Author: 20-Jul-2017
Complete List of Authors: Kulohoma, Benard ; Centre for Biotechnology and Bioinformatics, University of Nairobi, Nairobi, Kenya;; University of Oxford, Wellcome Trust Centre for Human Genetics, Nuffield Department of Medicine, Marriage, Fiona; 3. Centre for Integrated Genomic Research, University of Manchester, Manchester, UK. Vasieva, Olga; University of Liverpool, Institute of Integrative Biology
Nguyen, Kha; University of Liverpool, Institute of Translational Medicine Mankhambo, Limangeni; 2. Malawi-Liverpool-Wellcome Trust Clinical Research Programme, College of Medicine, Blantyre, Malawi. Molyneux, Malcolm; 2. Malawi-Liverpool-Wellcome Trust Clinical Research Programme, College of Medicine, Blantyre, Malawi Molyneux, ELizabeth; Paediatrics Day, Philip; 3. Centre for Integrated Genomic Research, University of Manchester, Manchester, UK. Carrol, Enitan; University of Liverpool, Institute of Infection and Global Health
Keywords: HIV, Immunology, Infectious Diseases, Microbiology, Tropical Paediatrics
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open on June 3, 2020 by guest. P
rotected by copyright.http://bm
jpaedsopen.bmj.com
/bm
jpo: first published as 10.1136/bmjpo-2017-000092 on 31 A
ugust 2017. Dow
nloaded from

Confidential: For Review O
nlyPeripheral blood RNA gene expression in children with pneumococcal meningitis; a prospective case control study
Journal: BMJ Paediatrics Open
Manuscript ID bmjpo-2017-000092
Article Type: Original article
Date Submitted by the
Author: 23-May-2017
Complete List of Authors:
Kulohoma, Benard ; Centre for Biotechnology and Bioinformatics,
University of Nairobi, Nairobi, Kenya;; University of Oxford, Wellcome Trust
Centre for Human Genetics, Nuffield Department of Medicine,
Marriage, Fiona; 3. Centre for Integrated Genomic Research,
University of
Manchester, Manchester, UK. Vasieva, Olga; University of Liverpool, Institute of Integrative Biology
Nguyen, Kha; University of Liverpool, Institute of Translational Medicine
Mankhambo, Limangeni; 2. Malawi-Liverpool-Wellcome Trust Clinical
Research Programme, College of Medicine, Blantyre, Malawi.
Molyneux, Malcolm; 2. Malawi-Liverpool-Wellcome Trust Clinical Research
Programme, College of Medicine, Blantyre, Malawi Molyneux, ELizabeth; Paediatrics
Day, Philip; 3. Centre for Integrated Genomic Research, University of
Manchester, Manchester, UK.
Carrol, Enitan; University of Liverpool, Institute of Infection and Global Health
Keywords: HIV, Immunology, Infectious Diseases, Microbiology, Tropical Paediatrics
Page 1 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyTitle: Peripheral blood RNA gene expression in children with pneumococcal meningitis: a prospective case control study
Benard W. Kulohoma 1 † *, Fiona Marriage 3 *, Olga Vasieva 4, Limangeni
Mankhambo 2, Kha Nguyen 5, Malcolm E Molyneux 2, Elizabeth M Molyneux 6, Philip
JR Day 3, Enitan D Carrol 2 5 6 ‡
1. Centre for Biotechnology and Bioinformatics, University of Nairobi.
2. Malawi-Liverpool-Wellcome Trust Clinical Research Programme, College of
Medicine, Blantyre, Malawi.
3. Centre for Integrated Genomic Research, University of Manchester, Manchester,
UK.
4. Institute of Integrative Biology, University of Liverpool, Liverpool, UK.
5. Institute of Infection and Global Health, University of Liverpool, Liverpool, UK.
6. Department of Paediatrics, University of Malawi College of Medicine, Blantyre,
Malawi.
*Contributed equally.
† Current address: Wellcome Trust Centre for Human Genetics, University of
Oxford.
‡ Corresponding author
Benard W. Kulohoma ([email protected])
Fiona Marriage ([email protected])
Olga Vasieva ([email protected] )
Limangeni Mankhambo ([email protected])
Kha Nguyen ([email protected])
Malcolm Molyneux ([email protected])
Elizabeth Molyneux ([email protected])
Philip J. R. Day ([email protected])
Enitan D. Carrol ([email protected]) - Corresponding author
Page 2 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyAbstract
Introduction: Invasive pneumococcal disease (IPD), caused by Streptococcus
pneumoniae, is a leading cause of pneumonia, meningitis and septicaemia
worldwide, with increased morbidity and mortality in HIV-infected children.
Objectives: We aimed to compare peripheral blood expression profiles between
HIV-infected and uninfected children with pneumococcal meningitis and controls,
and between survivors and non-survivors, in order to provide insight into the host
inflammatory response leading to poorer outcomes.
Design and setting: Prospective case control observational study in a tertiary
hospital in Malawi
Participants: Children aged 2 months to 16 years with pneumococcal meningitis or
pneumonia.
Methods: We used the human genome HGU133A Affymetrix array to explore
differences in gene expression between cases with pneumococcal meningitis (n=12)
and controls, and between HIV-infected and uninfected cases, and validated gene
expression profiles for 34 genes using real time qPCR (RT qPCR) in an independent
set of cases with IPD (n=229) and controls (n=13). Pathway analysis was used to
explore genes differentially expressed.
Results: Irrespective of underlying HIV infection, cases showed significant up-
regulation compared to controls of: S100 calcium binding protein A12 (S100A12);
vanin-1 (VNN1); arginase, liver (ARG1); matrix metallopeptidase 9 (MMP9); annexin
A3 (ANXA3); interleukin 1 receptor, type II (IL1R2); CD177 molecule (CD177);
S100 calcium binding protein A9 (S100A9), cytoskeleton-associated protein 4
(CKAP4); and glycogenin 1 (GYG1). RT qPCR confirmed differential expression in
keeping with microarray results. There was no differential gene expression in HIV-
infected compared to HIV-uninfected cases, but there was significant up-
regulation of folate receptor 3 (FOLR3), endocytic adaptor protein (NUMB) and
S100A12 in survivors compared non-survivors.
Conclusion: Children with IPD demonstrated increased expression in genes
regulating immune activation, oxidative stress, leucocyte adhesion and migration,
arginine metabolism, and glucocorticoid receptor signalling.
(280 words)
Page 3 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyKey words: Differential expression, children, invasive pneumococcal disease, host
response, pneumococcal meningitis
Key messages:
What is known about the subject
Invasive pneumococcal disease (IPD) caused by Streptococcus pneumoniae is
a leading cause of pneumonia, meningitis and septicaemia worldwide. Globally,
IPD is reported to cause about 11% (0.8 million) of all deaths in children less
than 5 years of age annually. The overall burden of IPD is increased 40-fold in
HIV-infected compared with HIV-uninfected children. However, mechanisms
involved in host response during IPD are not yet fully understood.
What this study adds
We demonstrate for the first time, differences in transcriptional profiles between HIV-
infected and HIV-uninfected children with pneumococcal meningitis (as a
homogeneous disease entity of invasive pneumococcal disease) and healthy
controls. We demonstrate increased expression in cases of genes regulating the
innate immune response, leucocyte migration, glucose homeostasis and endothelial
cell migration.
Introduction
Streptococcus pneumoniae infection is a leading cause of pneumonia, meningitis
and septicaemia worldwide, and results in approximately 1 million deaths in children
under the age of 5 years annually 1. The overall burden of invasive pneumococcal
disease (IPD) is increased 40-fold in HIV-infected compared with HIV-uninfected
children 2. Pneumococcal meningitis is a life-threatening disease with poor prognosis
associated with neurologic complications and a high case-fatality ratio in African
children, which is further increased by HIV co-infection 3.
Gene expression profiling during sepsis provides new insights into the host response
to invasive bacterial disease. Several sepsis studies have demonstrated up-
regulation of pathogen recognition receptors and signal transduction pathways, and
Page 4 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlya down-regulation of zinc homeostasis 4-6. The intricate host inflammatory response
is associated with neuronal and vascular injury, even after cerebrospinal fluid (CSF)
sterilization with antibiotics. Adjunctive new therapies for bacterial meningitis have to
date not shown any conclusive benefit, prompting the need for an improved
understanding of key mechanisms that might reveal potential new therapeutic targets
7. Chronic co-infection may impact gene expression, even among asymptomatic
patients. HIV is a major risk factor for IPD, characterised by waning immunity and
dysregulated physiology among infected individuals, greatly escalating disease
susceptibility and mortality outcomes 8 9, thereby influencing the host’s gene
expression. We examine differences in gene-expression using blood samples from
children with pneumococcal meningitis and matched healthy controls, and also
compare transcriptional profiles between children with pneumococcal meningitis with
and without underlying HIV infection.
Materials and methods
Patients and controls
Children (n=377) were recruited as part of a larger study investigating host
determinants of susceptibility to IPD conducted at Queen Elizabeth Central Hospital,
Blantyre, Malawi, between April 2004 and October 2006 10. Ethical approval was
granted from the College of Medicine Research Committee (COMREC), Malawi and
the Liverpool School of Tropical Medicine Local Research Ethics Committee. Written
informed consent was given prior to recruitment.
We excluded children (n=135) infected with other commonly prevalent microbes
(Salmonella typhimurium, Escherichia coli, Haemophilus influenzae b, Neisseria
meningitidis, Staphylococcus aureus, Streptococcus pyogenes) identified by positive
laboratory culture of blood, CSF or lung aspirate. Cases for the microarray analysis
were HIV treatment-naive children with confirmed pneumococcal meningitis defined
as abnormal CSF white blood cell (WBC) count, >10 WBC/∝l plus one or more of the
following: CSF culture positive for pneumococci, CSF gram stain consistent with
pneumococci, CSF positive for pneumococcal polysaccharide antigen or
pneumococcal DNA. Cases for the RT-qPCR were children with confirmed IPD,
Page 5 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlywhich was defined as: pneumococcal pneumonia (n=40) or pneumococcal meningitis
(n=189), confirmed by either culture, antigen test, or PCR. Controls were healthy
afebrile, malaria aparasitaemic children from the same villages as the index cases,
and were as closely age matched as possible. Microarray analysis was conducted
on 15 samples (12 from cases with pneumococcal meningitis and 3 from controls),
and RT-PCR was conducted on 242 samples (229 from cases with IPD and 13 from
controls), which included all those used for microarray analysis.
There were no known co-infections apart from HIV, and we did not test for other
viruses in the CSF, like CMV or EBV, which have been reported to be associated
with increased mortality in Malawian adults with bacterial meningitis 11 12.
Samples
Whole blood was drawn at admission from consecutive children with IPD. The
methodology for downstream transcriptome analysis from small blood samples has
been previously described 13.
RNA extraction, quantification and hybridization
Total RNA was extracted from whole blood using an optimised method for the
PAXgeneTM blood RNA kit (Qiagen, West Sussex, UK), as previously described 13.
The total RNA concentration (ng/µl) and ratios (260/280 and 260/230) were
measured using a NanoDrop ND-100 UV-vis spectrophotometer (NanoDrop
Technologies, Delaware, USA) and the RNA integrity was assessed using the
Agilent 2100 BioAnalyser (Agilent Technologies, Edinburgh, UK) before and after
concentration.
RNA (2µg) was reverse transcribed into cDNA using the Superscript double-stranded
cDNA synthesis kit (Invitrogen, Paisley, UK) according to the manufacturers’
instructions. The double stranded cDNA was purified using a GeneChipTM sample
clean-up module (Invitrogen, Paisley, UK). The purified cDNA was then biotin
labelled with the ENZO BioArray high yield RNA transcript labelling kit (Affymetrix,
High Wycombe, UK) and cleaned with a cRNA clean-up module (Invitrogen, Paisley,
UK). Aliquots of labelled cRNA (20µg) were fragmented at 94°C for 35 minutes and
then hybridized to a Human Genome U133A GeneChipTM array for 16 hours rotating
Page 6 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyat 60rpm at 45°C in a GeneChipTM Hybridization Oven 640 (Affymetrix, High
Wycombe, UK). Each chip was washed and stained on a GeneChipTM Fluidics
Station 450 (Affymetrix, High Wycombe, UK) and scanned using a GeneChipTM
Scanner 3000 (Affymetrix, High Wycombe, UK) employing standard recommended
protocols (Affymetrix, High Wycombe, UK).
Microarray data analysis
Microarray experiment data were analysed using R and Biocondutor packages 14.
Briefly, the human genome HGU133A array (Affymetrix, High Wycombe, UK) scans
output was pre-processed using the affy package 15. The limma package was used
to evaluate differential expressed genes 16 17. Gene annotations were performed
using hgu133a.db, KEGG.db database packages 18-21. Genes were considered
differentially expressed if they had a Benjamini and Hochberg (BH) adjusted p-value
< 1.5e-3 and > ±2-fold change in gene expression. Bonferroni p-value adjustments
were also performed for comparison. Canonical pathways and functional networks
that involve the differently expressed genes play were determined using the
Ingenuity Pathway Analysis (IPA) catalogue of well characterized metabolic and cell
signalling cascades. Expression data can be accessed using accession number
GSE47172 at the NCBI GEO database.
Reverse transcription for quantitative PCR
RNA samples were DNAse (Ambion, Warrington, UK) treated to remove any
contaminating genomic DNA. RNA (1µg) was reverse transcribed with SuperScript
IITM RNase H- Reverse Transcriptase and oligo (dT)12-18 (0.5µg/µL) following the
manufacturer’s guidelines. The cDNA was stored at -40°C until required.
Real-Time quantitative PCR measurement of target genes
The Human Universal Probe Library (UPL, Roche, Switzerland) employing
proprietary locked nucleic acid (LNA) analogues was used to design qPCR assays to
measure expression levels in genes of interest. Using the Roche Online Assay
Design Centre, specific primers and an associated probe were selected for the
reference and target transcripts. Gene expression was determined using real time
qPCR on a Roche LightCycler 480 (Supplementary Table 2).
Page 7 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyThe following 34 genes were identified from literature and prioritised for real-time
qPCR differential expression analysis between cases and controls: ACSL1, ANXA3,
ATP, BAG1, BPGM, C3AR1, CA4, CD177, CD55, CD59, CEACAM, CFLAR, FOLR3,
GNAI3, GNLY, GYG1, IL1R2, IL1RN, ITGAM, KLRF1, LCK, LCN2, LTF, MAPK14,
MMP9, NUMB, OLAH, PSEN1, RETN, S100A12, SAMSN, SERPINA1, SUB1, and
VNN1. We used a previously described real-time qPCR data normalization method
22.
Statistical analysis of genes prioritised for real-time qPCR differential
expression analysis
First, we derived the relative gene expression in cases compared to controls for the
34 genes under assessment. We used a previously described real-time qPCR data
normalization method 22. Briefly, the amounts of target genes expressed in a sample
were normalized to the average of the three endogenous controls. This is given by
∆Cq, where ∆Cq is determined by subtracting the average endogenous gene Cq
value from the average target gene Cq value:
Cq target gene – Cq average endogenous gene = ∆Cq.
The calculation of relative expression, ∆∆Cq, involves subtraction of ∆Cq value for the
controls from the ∆Cq value for the cases:
∆Cq target gene(case) – ∆Cq target gene(control) = ∆∆Cq.
2-∆∆Cq is the relative expression of the target gene in cases compared to controls.
Next, we examined statistically significant differences in relative gene expression
between cases and controls using the Welch two-sample t-test implemented in the R
package. We generate boxplots to visualise the mean and media relative expression
in cases and controls separately, and the Welch two-sample t-test p-value to show
statistically significant differences in relative gene expression between these two
groups.
Results
Page 8 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
Transcriptional profiles among the cases and controls
In the microarray discovery cohort, there were 12 children with pneumococcal
meningitis (6 male, 6 female, median age 1.1 years) and 3 controls (2 male, 1
female, median age 7 years). The breakdown was as follows: HIV-infected survivors
(n=3), HIV-infected non-survivors (n=3), HIV-uninfected survivors (n=3) and HIV
uninfected non-survivors (n=3) (Supplementary Table 1). The RT-PCT validation
cohort had a median age of 3.09 years.
We examined whether global transcriptional profiles of peripheral blood from children
with pneumococcal meningitis (n=12) were distinct from those of healthy controls
(n=3) randomly selected from a larger dataset by microarray expression profile
analysis 10. In general, there was a marked distinction in differential expression
between cases and controls (Supplementary Figure 1). We identified 10 significantly
differentially expressed genes (Benjamini & Hochberg (BH) adjusted p-value < 1.5e
3 and > ±2-fold change) (Figure 1). We observed significant up-regulation of: S100
calcium binding protein A12 (S100A12); vanin-1 (VNN1); arginase, liver (ARG1);
matrix metallopeptidase 9 (gelatinase B, 92kDa gelatinase, 92kDa type IV
collagenase) (MMP9); annexin A3 (ANXA3); interleukin 1 receptor, type II (IL1R2);
CD177 molecule (CD177); S100 calcium binding protein A9 (S100A9), cytoskeleton
associated protein 4 (CKAP4); and glycogenin 1 (GYG1).
Differential expression, underlying HIV infection and disease outcomes
In the microarray discovery cohort, we did not find any significantly differentially
expressed genes for comparisons between HIV-infected cases and HIV-uninfected
cases or between survivors and non-survivors (Supplementary Figure 2).
RT qPCR validation
RT qPCR validation was performed for a set of 34 prioritised genes selected from
literature for cases (n=229) and controls (n=13). The RT-PCR results are in
agreement with our microarray analysis findings. All the genes are significantly
differentially expressed between the two groups with the exception of guanine
nucleotide-binding protein subunit alpha-13 (GNA13), protein numb (NUMB), and
presenilin-1 (PSEN1) (Figure 2). There was no significantly increased gene
Page 9 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyexpression in HIV-infected compared to HIV-uninfected cases (Supplementary
Figure 3). Interestingly, there was significant up-regulation of folate receptor 3
(FOLR3), NUMB and S100A12 in survivors compared non-survivors (Supplementary
Figure 4). There was wide variability in relative gene expression in cases, but not
controls.
Pathway analysis
Networks were reconstructed using IPA software for the genes differentially
expressed between cases and controls in the microarray experiment, and the
combined microarray and RT qPCR experiments (Figure 3A and 3B). For the genes
defined by the microarray experiments the networks were predominantly related to
granulocyte function, but also including antimicrobial and endothelial activation
responses (Figure 3A). The merged set demonstrated wider networks involving
immune cell activation as well as leucocyte migration and adhesion (Figure 3B). The
canonical pathways mapped by genes defined in the microarray experiment
demonstrated greatest changes in the arginine and granulocyte pathways. Canonical
pathways in the combined microarray and RT qPCR experiments demonstrated
greatest change in leucocytes, and especially neutrophil activation and migration,
Notch and glucocorticoid receptor signalling pathways (Supplementary Tables 3 and
4).
Discussion and conclusion
We have shown significant differences in RNA transcriptional profiles in children with
pneumococcal meningitis compared to controls. Children with pneumococcal
meningitis demonstrated increased expression in genes involved in the inflammatory
response, and glucose and L-arginine metabolic pathways. Dysregulation of these
pathways can lead to an impaired adaptive host response to pneumococcal
infection, thereby contributing to the increased morbidity and mortality. Our findings
in the initial cohort of pneumococcal meningitis were validated in the larger cohort of
all children with IPD, which includes presentations with pneumonia as well as
meningitis. These findings add validity to the initial results from the microarray
experiments, and suggest that the host response observed is systemic, and not
simply localised to the process of blood-brain barrier disruption per se. We observe
Page 10 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlywide variability in relative gene expression in cases, which perhaps reflects
differences in disease onset and robustness of host response within this group.
Elevation of S100A12, VNN, ARG1, MMP9, ANXA3, IL1R2, CD177, S100A9,
CKAP4 and GYG1 in pneumococcal meningitis supports previous findings that have
highlighted the important roles of some of these genes during host pathogen
response. These genes have important interconnected functions for cellular
interactions and response to infection (Figure 3A and Supplementary Table 3), and
play crucial roles during host immune response; cell regulatory processes such as
apoptosis and differentiation; and metabolic processes such as amino acid and
glucose metabolism.
Vanin-1 (VNN1) protein is expressed by human phagocytes, and involved in
leukocyte adhesion and migration 23. The VNN-1 knockout mice model has provided
clear evidence that VNN-1 modulates redox and immune pathways 24. Exposure of
human mononuclear cells to oxidative stress results in up-regulation of human VNN1
and down-regulation of PPAR 25. IL1R2 and IL1RN were up-regulated in cases,
which is consistent with our previous report of the IL-1Ra single-nucleotide
polymorphism rs4251961 playing a key role in the pathophysiology of IPD and in
other human infections 10. Recent reports also demonstrate IL1R2 expression up
regulation in sepsis, being more pronounced in Gram negative than Gram positive
infections 26.
During infection, host production of the cytokine nitric oxide (NO) after non-opsonic
phagocytosis, exerts microbicidal effects 27 28. ARG1 expression is inducible in the
lungs of mice in response to pneumococcal infection 29. Phagocytosis of
pneumococci by macrophages also results in increased production of nitric oxide
synthase 2 (NOS2) dependent production of NO and reactive nitrogen species 30.
Increased plasma arginase activity depletes L-arginine concentrations, the substrate
for NO synthesis, leading to vascular dysfunction during severe sepsis and
supressed NO-mediated microbicidal effects 31. Increased ARG1 activity may also be
a bacterial survival strategy to escape the NO-dependent host antimicrobial immune
response 30.
Page 11 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
Neutrophil-specific glycoprotein CD177 is expressed on a subset of human
neutrophils, and is involved in neutrophil transendothelial migration. A previous
microarray study of purified neutrophils from septic shock patients revealed CD177
mRNA has the highest differential expression between cases and controls 32.
Consistent with our data, that study also demonstrated increased expression of
ARG1, ANXA3, CKAP4, IL1R2, MMP9, and VNN1. ANXA3 promotes endothelial cell
junction integrity in animal models, and endothelial cell motility in vitro. ANXA3 is up
regulated following neuronal injury, which may explain the finding in pneumococcal
meningitis 33.
S100A12 plays a prominent role in the regulation of proinflammatory processes
and immune response by recruiting leukocytes, promoting cytokine and chemokine
production, and regulating leukocyte adhesion and migration 34. The S100A8/A9
heterodimer is expressed by myeloid cells, especially neutrophils, and has a
protective effect in the host response to pneumococcal infection by increasing
circulating neutrophils through increased granulocyte-colony stimulating factor
production 35. It is an antimicrobial peptide, but also plays an important role in
leukocyte migration 36. During infection, S100 proteins stimulate the proinflammatory
immune response through interaction with the immunoglobulin family
transmembrane pattern recognition receptors: Receptor for advanced glycation end
products (RAGE) and Toll-like receptor 4 (TLR4). This leads to NF-κB-mediated pro-
inflammatory response with production of proinflammatory cytokines. This
inflammatory response in turn leads to increased expression of S100 proteins, and
the start of a positive feedback loop. Although as antimicrobial proteins they protect
against infection, they can also have a negative detrimental effect on the host by
amplifying the destructive pro-inflammatory responses. The increased expression in
survivors may be explained by this positive feedback loop 37.
Glucocorticoids also play a significant role in immune response regulation, by
supressing immune and inflammatory responses, and modulating cytokines that
promote host immune responses 38. We speculate that these responses are
important in regulating immune responses to avoid host tissue damage. NUMB
negatively regulates Notch, which in turn attenuates proinflammatory cytokines and
Page 12 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyincreases anti-inflammatory cytokines, which may explain the increase in NUMB
expression in survivors 39 40.
Our results are consistent with previous studies on transcription profiling in other
bacterial diseases, suggesting that some of the mechanisms are not specific to IPD.
Although it could be argued that the lack of age matching in cases versus controls
could cause differential expression due to maturation of the immune system in older
children, the consistency of our results with other studies makes this unlikely 41 42. A
limitation of our study is that the microarray analysis was performed using the
Affymetrix Human Genome U133A array, and sample number was constrained at
the time by cost. The microarray dataset was not large enough to allow all possible
multifactorial models of co-morbidity and disease outcomes to be exhaustively
examined, but sought to validate our findings using RT-qPCR in a larger cohort of
children. Further evaluation is in a larger cohort using whole genome sequencing
would provide more insights on the mechanisms of host response.
In conclusion, this comparative study of gene expression provides mechanistic
insight for IPD in children, and demonstrates significant and widespread immune
activation, with oxidative stress, recruitment of neutrophils, leucocyte adhesion and
migration, activation of antimicrobial peptides, and preservation of endothelial cell
junction integrity.
(2736 words)
Competing interests
None declared
Author contributions
FM, PJRD, EDC conceived and designed the study; LM, EDC collected the clinical
samples; FM performed experiments; BWK, FM, OV, KN, SR, PJRD, EDC
performed the analysis; BWK, FM, PJRD, EDC wrote the first draft of the manuscript;
BWK, FM, SR, LM, KN, OV, MM, EM, PJRD, EDC contributed to writing the
manuscript.
Page 13 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyAcknowledgements
The IPD Study Group (C Antonio, M Chinamale, L Jere, D Mnapo, V Munthali, F
Nyalo, J Simwinga, M Kaole, DL Banda, A Manyika, K Phiri) recruited patients. We
thank the children included in this study, and their parents and guardians for giving
consent for them to participate in the study. We also extend thanks to the nursing
and medical staff, at the Malawi-Liverpool-Wellcome Trust Clinical Research
Programme (MLW), Research Ward, for their contribution to this study. EDC was
supported by a Wellcome Trust Career Development Grant (068026). BWK was
supported by a Wellcome Trust Major Overseas programme award (084679/Z/08/Z).
The funding bodies had no role in collection, analysis or interpretation of data or in
writing the manuscript.
References
1. O'Brien KL, Wolfson LJ, Watt JP, et al. Burden of disease caused by Streptococcus pneumoniae in
children younger than 5 years: global estimates. Lancet 2009;374(9693):893-902. doi: S0140-
6736(09)61204-6 [pii]
10.1016/S0140-6736(09)61204-6 [published Online First: 2009/09/15]
2. Madhi SA, Petersen K, Madhi A, et al. Impact of human immunodeficiency virus type 1 on the
disease spectrum of Streptococcus pneumoniae in South African children. The Pediatric
infectious disease journal 2000;19(12):1141-7.
3. Nyasulu P, Cohen C, De Gouveia L, et al. Increased risk of death in human immunodeficiency virus-
infected children with pneumococcal meningitis in South Africa, 2003-2005. The Pediatric
infectious disease journal 2011;30(12):1075-80. doi: 10.1097/INF.0b013e31822cca05
4. Wong HR, Cvijanovich N, Allen GL, et al. Genomic expression profiling across the pediatric
systemic inflammatory response syndrome, sepsis, and septic shock spectrum. Crit Care Med
2009;37(5):1558-66. doi: 10.1097/CCM.0b013e31819fcc08
5. Wong HR, Freishtat RJ, Monaco M, et al. Leukocyte subset-derived genomewide expression
profiles in pediatric septic shock. Pediatric critical care medicine : a journal of the Society of
Critical Care Medicine and the World Federation of Pediatric Intensive and Critical Care
Societies 2010;11(3):349-55. doi: 10.1097/PCC.0b013e3181c519b4
6. Khoo SK, Petillo D, Parida M, et al. Host response transcriptional profiling reveals extracellular
components and ABC (ATP-binding cassette) transporters gene enrichment in typhoid fever-
infected Nigerian children. BMC infectious diseases 2011;11:241. doi: 10.1186/1471-2334-
11-241
7. Hoffman O, Weber RJ. Pathophysiology and treatment of bacterial meningitis. Ther Adv Neurol
Disord 2009;2(6):1-7. doi: 10.1177/1756285609337975
8. Glennie SJ, Williams NA, Heyderman RS. Mucosal immunity in resource-limited setting: is the
battle ground different? Trends in microbiology 2010;18(11):487-93. doi:
10.1016/j.tim.2010.08.002
S0966-842X(10)00139-3 [pii] [published Online First: 2010/09/14]
9. Iwajomo OH, Finn A, Moons P, et al. Deteriorating pneumococcal-specific B-cell memory in
minimally symptomatic African children with HIV infection. The Journal of infectious diseases
2011;204(4):534-43. doi: jir316 [pii]
Page 14 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly10.1093/infdis/jir316 [published Online First: 2011/07/28]
10. Carrol ED, Payton A, Payne D, et al. The IL1RN promoter rs4251961 correlates with IL-1 receptor
antagonist concentrations in human infection and is differentially regulated by GATA-1. J
Immunol 2011;186(4):2329-35. doi: 10.4049/jimmunol.1002402
11. Kelly MJ, Benjamin LA, Cartwright K, et al. Epstein-barr virus coinfection in cerebrospinal fluid is
associated with increased mortality in Malawian adults with bacterial meningitis. The Journal
of infectious diseases 2012;205(1):106-10. doi: 10.1093/infdis/jir707
12. Benjamin LA, Kelly M, Cohen D, et al. Detection of herpes viruses in the cerebrospinal fluid of
adults with suspected viral meningitis in Malawi. Infection 2013;41(1):27-31. doi:
10.1007/s15010-012-0292-z
13. Carrol ED, Salway F, Pepper SD, et al. Successful downstream application of the Paxgene Blood
RNA system from small blood samples in paediatric patients for quantitative PCR analysis.
BMC Immunol 2007;8:20. doi: 10.1186/1471-2172-8-20
14. Gentleman RC, Carey VJ, Bates DM, et al. Bioconductor: open software development for
computational biology and bioinformatics. Genome biology 2004;5(10):R80.1-R80.16.
15. Irizarry R. From CEL files to annotated lists of interesting genes. In: Gentleman R. CV, Dudoit S.,
Irizarry R., Huber W., ed. Bioinformatics and Computational Biology Solutions using R and
Bioconductor. New York: Springer 2005:431-42.
16. Smyth GK. Linear models and empirical Bayes methods for assessing differential expression in
microarray experiments. Statistical Applications in Genetics and Molecular Biology
2004;3(1):3.
17. Smyth GK. Limma: linear models for microarray data. In: Gentleman R. CV, Dudoit S., Irizarry R.,
Huber W., ed. Bioinformatics and Computational Biology Solutions using R and
Bioconductor. New York: Springer 2005:397-420.
18. hgu133a.db: Affymetrix Human Genome U133 Set annotation data (chip hgu133a) [program]. R
package version 2.7.1 version, 2009.
19. annotate: Annotation for microarrays. R package version 1.34.1. [program]. R package version
1.34.1. version, 2008.
20. GO.db: A set of annotation maps describing the entire Gene Ontology [program]. R package
version 2.12 version, 2013.
21. KEGG.db: A set of annotation maps for KEGG [program]. R package version 2.7.1 version, 2009.
22. Vandesompele J, De Preter K, Pattyn F, et al. Accurate normalization of real-time quantitative RT-
PCR data by geometric averaging of multiple internal control genes. Genome Biol
2002;3(7):RESEARCH0034.
23. Meghari S, Berruyer C, Lepidi H, et al. Vanin-1 controls granuloma formation and macrophage
polarization in Coxiella burnetii infection. Eur J Immunol 2007;37(1):24-32. doi:
10.1002/eji.200636054
24. Ferreira DW, Naquet P, Manautou JE. Influence of Vanin-1 and Catalytic Products in Liver During
Normal and Oxidative Stress Conditions. Curr Med Chem 2015;22(20):2407-16.
25. Zhang B, Lo C, Shen L, et al. The role of vanin-1 and oxidative stress-related pathways in
distinguishing acute and chronic pediatric ITP. Blood 2011;117(17):4569-79. doi:
10.1182/blood-2010-09-304931
26. Lang Y, Jiang Y, Gao M, et al. Interleukin-1 Receptor 2: A New Biomarker for Sepsis Diagnosis and
Gram-Negative/Gram-Positive Bacterial Differentiation. Shock 2017;47(1):119-24. doi:
10.1097/SHK.0000000000000714
27. Ulett GC, Adderson EE. Nitric oxide is a key determinant of group B streptococcus-induced
murine macrophage apoptosis. The Journal of infectious diseases 2005;191(10):1761-70. doi:
10.1086/429693
28. Gross A, Spiesser S, Terraza A, et al. Expression and bactericidal activity of nitric oxide synthase in
Brucella suis-infected murine macrophages. Infection and immunity 1998;66(4):1309-16.
Page 15 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly29. Knippenberg S, Brumshagen C, Aschenbrenner F, et al. Arginase 1 activity worsens lung-
protective immunity against Streptococcus pneumoniae infection. Eur J Immunol
2015;45(6):1716-26. doi: 10.1002/eji.201445419
30. Marriott HM, Ali F, Read RC, et al. Nitric oxide levels regulate macrophage commitment to
apoptosis or necrosis during pneumococcal infection. FASEB J 2004;18(10):1126-8. doi:
10.1096/fj.03-1450fje
31. Darcy CJ, Woodberry T, Davis JS, et al. Increased plasma arginase activity in human sepsis:
association with increased circulating neutrophils. Clinical chemistry and laboratory medicine
: CCLM / FESCC 2014;52(4):573-81. doi: 10.1515/cclm-2013-0698
32. Demaret J, Venet F, Plassais J, et al. Identification of CD177 as the most dysregulated parameter
in a microarray study of purified neutrophils from septic shock patients. Immunol Lett
2016;178:122-30. doi: 10.1016/j.imlet.2016.08.011
33. Chong KW, Chen MJ, Koay ES, et al. Annexin A3 is associated with cell death in lactacystin-
mediated neuronal injury. Neurosci Lett 2010;485(2):129-33. doi:
10.1016/j.neulet.2010.08.089
34. Goyette J, Geczy CL. Inflammation-associated S100 proteins: new mechanisms that regulate
function. Amino Acids 2011;41(4):821-42. doi: 10.1007/s00726-010-0528-0
35. De Filippo K, Neill DR, Mathies M, et al. A new protective role for S100A9 in regulation of
neutrophil recruitment during invasive pneumococcal pneumonia. FASEB J 2014;28(8):3600-
8. doi: 10.1096/fj.13-247460
36. Raquil MA, Anceriz N, Rouleau P, et al. Blockade of antimicrobial proteins S100A8 and S100A9
inhibits phagocyte migration to the alveoli in streptococcal pneumonia. J Immunol
2008;180(5):3366-74.
37. Zackular JP, Chazin WJ, Skaar EP. Nutritional Immunity: S100 Proteins at the Host-Pathogen
Interface. The Journal of biological chemistry 2015;290(31):18991-8. doi:
10.1074/jbc.R115.645085
38. Sakai A, Han J, Cato AC, et al. Glucocorticoids synergize with IL-1beta to induce TLR2 expression
via MAP Kinase Phosphatase-1-dependent dual Inhibition of MAPK JNK and p38 in epithelial
cells. BMC molecular biology 2004;5:2. doi: 10.1186/1471-2199-5-2
39. Kueanjinda P, Roytrakul S, Palaga T. A Novel Role of Numb as A Regulator of Pro-inflammatory
Cytokine Production in Macrophages in Response to Toll-like Receptor 4. Scientific reports
2015;5:12784. doi: 10.1038/srep12784
40. Singla DK, Wang J, Singla R. Primary human monocytes differentiate into M2 macrophages and
involve Notch-1 pathway. Can J Physiol Pharmacol 2017;95(3):288-94. doi: 10.1139/cjpp-
2016-0319
41. Langley RJ, Tsalik EL, van Velkinburgh JC, et al. An integrated clinico-metabolomic model
improves prediction of death in sepsis. Sci Transl Med 2013;5(195):195ra95. doi:
10.1126/scitranslmed.3005893
42. Liu K, Chen L, Kaur R, et al. Transcriptome signature in young children with acute otitis media due
to Streptococcus pneumoniae. Microbes Infect 2012;14(7-8):600-9. doi:
10.1016/j.micinf.2012.01.006
Figure and Table legends
Page 16 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyFigure 1: Distribution plot of the differentially expressed genes. The significantly
differentially expressed genes are shown in red colour. The significance threshold
(P<1.5e-3) is indicated by a dashed red line, and a fold change threshold of more
than 2 is shown by the dashed vertical lines. The green line shows P<0.05. (A)
Shows results for un-adjusted p-values, (B) results for Benjamini and Hochberg (BH)
adjusted p-values, and (C) stringent Bonferroni adjusted p-values, which represents
an over-correction.
Figure 2: Validation of RNA transcription profile differential expression using
Real Time quantitative PCR. Relative gene expression in cases compared to
controls for 34 genes assessed. The black line shows the boxplot median. The red
dot shows the mean, and the Welch two-sample t-test p-value is shown on the top
right corner.
Figure 3: The gene network for significantly differentially expressed
genes in the cases. Networks reconstructed to support the most direct connectivity
between genes differentially expressed between cases and controls. IPA (Qiagen)
software's Core analysis application has been used to perform an automatic
graphical reconstruction of the network via utilisation of IPA's Knowledge base
database of protein interactions. The meaning of links and shapes is explained in the
inserted legend. Functional connections are presented in blue, information
connections to the associated categories in pink and grey. (A) Network reconstructed
only for the DE genes identified by microarray analysis. (B) Network reconstructed
for the merged datasets of DE genes identified via microarray (red) and PCR
analysis (green). White blocks correspond to nodes added by IPA's network editor
automatically to ensure network connectivity.
Supplementary Figure 1: Children with pneumococcal meningitis and healthy
controls have distinct transcriptional profiles. A heatmap of the top 100 (P<0.05)
differentially expressed genes compared in cases and controls. The cases and
controls were distinguished into two distinct clusters highlighting their distinct
transcriptional profiles.
Page 17 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlySupplementary Figure 2: Comparison of microarray transcriptional profiles of
HIV infected and HIV-uninfected and survivors and non- survivors. There were
no significantly differentially expressed genes for any of these comparisons.
Supplementary Figure 3: Comparison of RT-qPCT gene expression for HIV
infected and HIV-uninfected. Relative gene expression in cases compared to
controls for 34 genes assessed. There were no significantly differentially expressed
genes for all of these comparisons. Relative gene expression in cases compared to
controls. The black line shows the boxplot median. The red dot shows the mean, and
the Welch two-sample t-test p-value is shown on the top right corner.
Supplementary Figure 4: Comparison of RT-qPCT gene expression for
survivors and non-survivors. Relative gene expression in cases compared to
controls for 34 genes assessed. There were three significantly differentially
expressed genes: FOLR3, NUMB, and S100A12. Relative gene expression in cases
compared to controls. The black line shows the boxplot median. The red dot shows
the mean, and the Welch two-sample t-test p-value is shown on the top right corner.
Supplementary Table 1: Clinical data on children with pneumococcal meningitis and healthy controls. Supplementary Table 2: Primer sequences and the UPL probe number for each RT qPCR assay. Supplementary Table 3: Canonical pathways of differentially expressed genes between cases and controls in microarray experiment Supplementary Table 4: Canonical pathways of differentially expressed genes between cases and controls in combined microarray and RT- qPCR experiments
Page 18 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
94x24mm (300 x 300 DPI)
Page 19 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
177x177mm (300 x 300 DPI)
Page 20 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
361x481mm (300 x 300 DPI)
Page 21 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
270x203mm (300 x 300 DPI)
Page 22 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
117x42mm (300 x 300 DPI)
Page 23 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
177x177mm (300 x 300 DPI)
Page 24 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nly
177x177mm (300 x 300 DPI)
Page 25 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review O
nlyTable S1: Clinical data on children with pneumococcal meningitis and
healthy controls
Sample ID
Age
(years)
Sex Case/control HIV status
Survival status
Previous antibiotics use
Duration of symptoms (days)
94 0.4 F Case Positive Died No 2
95 1.0 F Case Positive
Died Yes 6
97 5.7 F Case Positive
Died No 1
96 1.2 M Case Positive
Alive No 3
109 1.8 F Case Positive
Alive No 2
123 12.4 M Case Positive
Alive Yes 3
82 13.0 M Case Negative Died No 7
115 0.3 M Case Negative
Died Yes 3
121 0.5 F Case Negative
Died No 5
104 0.8 M Case Negative
Alive No 3
105 0.7 M Case Negative
Alive No 3
127 10.6 F Case Negative
Alive Yes 3
C08 7.0 M Control Negative
Alive N/A N/A
C09 7.0 M Control Negative
Alive N/A N/A
C13 7.0 F Control Negative
Alive N/A N/A
Page 26 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only1
Table S2: Primer sequences and the UPL probe number for each RT qPCR assay
Gene Symbol Gene Forward Primer (5'-3') Reverse Primer (5'-3') UPL Probe Accession Number
ACSL1 Acyl-CoA synthetase long-chain family member 1 ccactgtgctgacgttcct gcactctgtctgtccgtatcc 26 NM_001995
ANXA3 Annexin A3 ttgttaaggaatatcaagcagcat gccagagagatcacccttca 17 NM_005139
C4A Complement component 4A (Rodgers blood group) ggaccctgaagacgaaattg gaagccctcgttcacctg 75 NM_007293, NM_001252204
C3AR1 complement component 3a receptor 1
aagaaagcaaggcagtccatt
cgtgtgagctcctcactgaa
16
NM_004054
ILR2A ILR2A interleukin 1 receptor, type II tacgcaccacagtcaaggaa aagaaggccagtgaaagtgg 2 NM_004633, NM_173343
MAPK14
MAPK 14 mitogen-activated protein kinase 14
MAPK 14 mitogen-activated protein kinase 14
ccatgagatgggtcaccag
65
NM_139012, NM_139014, NM_001315, NM_139013
MMP9
matrix metallopeptidase 9 (gelatinase B, 92kDa gelatinase, 92kDa type IV collagenase)
gaaccaatctcaccgacagg
gaaccaatctcaccgacagg
6
NM_004994
CD177 CD177 molecule gcctctccctgatctcctaca caaggatcctgggtctgc 79 NM_020406
CD55 CD55 molecule, decay accelerating factor for complement (Cromer blood group) aataatgatgaaggagagtggagtg tggtgggaccttggaagtta 77 NM_000574
CEACAM1 Carcinoembryonic antigen-related cell adhesion molecule 1 (biliary glycoprotein) taggcctaagaggctttctcc tgagggaaagtatgcacagtccgtgtc 80
NM_001712, NM_001184816, NM_001184813, NM_001184815, NM_001205344, NM_001024912
GYG1 Glycogenin 1 ctgaaacagcacaggaccac atcgccactgtccaagacat 75 NM_004130
FOLR3 Folate receptor 3 (gamma) cacaaaggctggaattggac cttgaaggagtggctccaga 19 NM_000804
ITGAM Integrin, alpha M (complement component 3 receptor 3 subunit) tctgaagaccattcagaaccag ggagctgctacttcctgtctg 7 NM_000632
LTF Lactotransferrin ctaatctgaaaaagtgctcaacctc gccatcttcttcggttttacttc 79 NM_002343, NM_001199149
OLAH Oleoyl-ACP hydrolase accgagatcgctccactg ggagaatcaagtgagattttaatagga 19 NM_018324, NM_001039702
RETN Resistin tgcaggatgaaagctctctg catggagcacagggtcttg 45 NM_020415
Page 27 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from
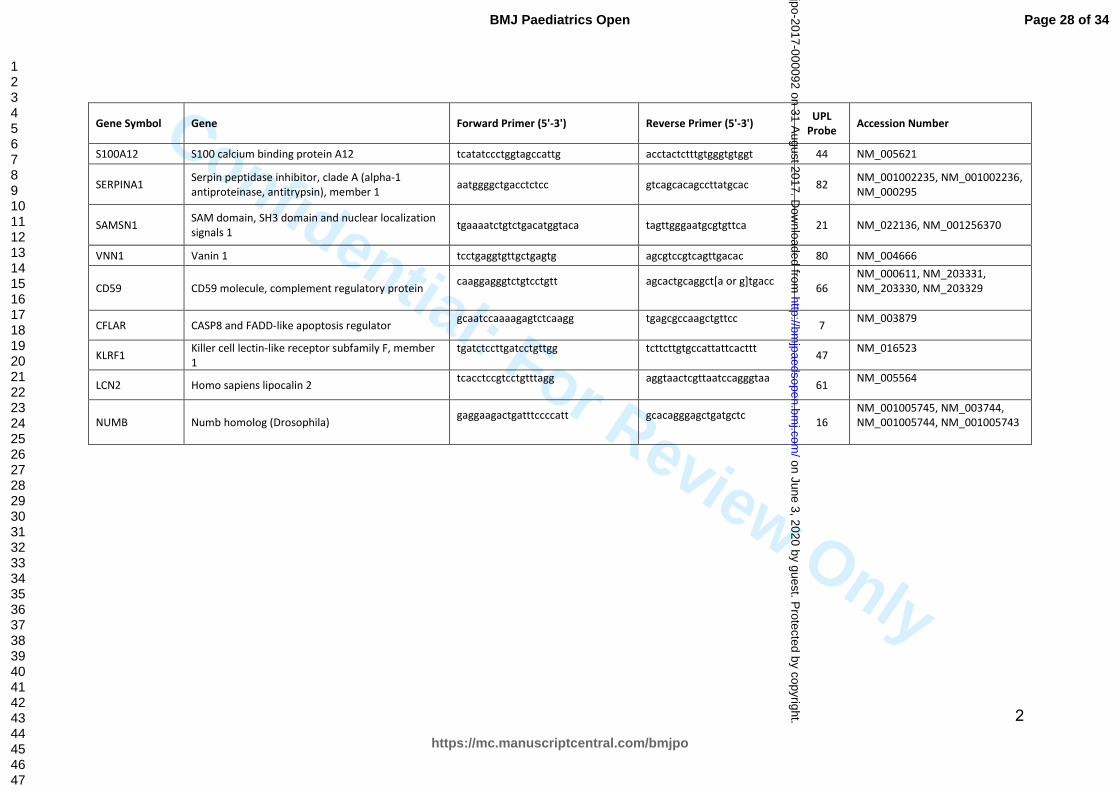
Confidential: For Review Only2
Gene Symbol Gene Forward Primer (5'-3') Reverse Primer (5'-3') UPL Probe Accession Number
S100A12 S100 calcium binding protein A12 tcatatccctggtagccattg acctactctttgtgggtgtggt 44 NM_005621
SERPINA1 Serpin peptidase inhibitor, clade A (alpha-1 antiproteinase, antitrypsin), member 1 aatggggctgacctctcc gtcagcacagccttatgcac 82 NM_001002235, NM_001002236,
NM_000295
SAMSN1 SAM domain, SH3 domain and nuclear localization signals 1 tgaaaatctgtctgacatggtaca tagttgggaatgcgtgttca 21 NM_022136, NM_001256370
VNN1 Vanin 1 tcctgaggtgttgctgagtg agcgtccgtcagttgacac 80 NM_004666
CD59 CD59 molecule, complement regulatory protein caaggagggtctgtcctgtt
agcactgcaggct[a or g]tgacc 66
NM_000611, NM_203331, NM_203330, NM_203329
CFLAR CASP8 and FADD-like apoptosis regulator gcaatccaaaagagtctcaagg
tgagcgccaagctgttcc 7 NM_003879
KLRF1 Killer cell lectin-like receptor subfamily F, member 1
tgatctccttgatcctgttgg
tcttcttgtgccattattcacttt 47 NM_016523
LCN2 Homo sapiens lipocalin 2 tcacctccgtcctgtttagg
aggtaactcgttaatccagggtaa 61 NM_005564
NUMB Numb homolog (Drosophila) gaggaagactgatttccccatt
gcacagggagctgatgctc 16
NM_001005745, NM_003744, NM_001005744, NM_001005743
Page 28 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Supplementary Table 3: Canonical pathways of differentially expressedgenes between cases and controls in microarray experiment Ingenuity Canonical Pathways -log(p-value) Ratio MoleculesLXR/RXR Activation 2.77 0.0165 IL1R2,MMP9Arginine Degradation I (Arginase Pathway) 2.69 0.25 ARG1Urea Cycle 2.51 0.167 ARG1Arginine Degradation VI (Arginase 2 Pathway) 2.51 0.167 ARG1Glycogen Biosynthesis II (from UDP-D-Glucose) 2.51 0.167 GYG1Granulocyte Adhesion and Diapedesis 2.51 0.0121 IL1R2,MMP9Hepatic Fibrosis / Hepatic Stellate Cell Activation 2.43 0.011 IL1R2,MMP9Airway Pathology in Chronic Obstructive Pulmonary Disease 2.38 0.125 MMP9Citrulline Biosynthesis 2.38 0.125 ARG1Role of IL-17A in Psoriasis 2.17 0.0769 S100A9Superpathway of Citrulline Metabolism 2.14 0.0714 ARG1Inhibition of Angiogenesis by TSP1 1.79 0.0312 MMP9Inhibition of Matrix Metalloproteases 1.71 0.0263 MMP9Neuroprotective Role of THOP1 in Alzheimer's Disease 1.69 0.025 MMP9IL-10 Signaling 1.46 0.0147 IL1R2Glioma Invasiveness Signaling 1.45 0.0143 MMP9Bladder Cancer Signaling 1.36 0.0116 MMP9PPAR Signaling 1.34 0.0111 IL1R2HIF1α Signaling 1.25 0.00885 MMP9p38 MAPK Signaling 1.23 0.00855 IL1R2Pancreatic Adenocarcinoma Signaling 1.23 0.00847 MMP9Atherosclerosis Signaling 1.21 0.00806 MMP9IL-6 Signaling 1.2 0.00787 IL1R2Ovarian Cancer Signaling 1.15 0.00699 MMP9Relaxin Signaling 1.13 0.00671 MMP9Hepatic Cholestasis 1.11 0.00637 IL1R2PPARα/RXRα Activation 1.08 0.00602 IL1R2Agranulocyte Adhesion and Diapedesis 1.06 0.00571 MMP9NF-κB Signaling 1.06 0.00565 IL1R2Regulation of the Epithelial-Mesenchymal Transition Pathway 1.03 0.00535 MMP9ILK Signaling 1.02 0.00521 MMP9IL-8 Signaling 1.01 0.0051 MMP9Leukocyte Extravasation Signaling 0.996 0.00488 MMP9LPS/IL-1 Mediated Inhibition of RXR Function 0.99 0.00481 IL1R2Role of Osteoblasts, Osteoclasts and Chondrocytes in Rheumatoid Arthritis 0.954 0.00441 IL1R2Colorectal Cancer Metastasis Signaling 0.928 0.00413 MMP9Glucocorticoid Receptor Signaling 0.862 0.00352 IL1R2Role of Macrophages, Fibroblasts and Endothelial Cells in Rheumatoid Arthritis 0.84 0.00333 IL1R2Axonal Guidance Signaling 0.686 0.00226 MMP9
Page 29 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Ingenuity Canonical Pathways -log(p-value) Ratio MoleculesComplement System 6.35 0.111 CD55,CD59,ITGAM,C3AR1Granulocyte Adhesion and Diapedesis 5.02 0.0303 IL1R2,GNAI3,ITGAM,IL1RN,MMP9LXR/RXR Activation 4.24 0.0331 IL1R2,IL1RN,SERPINA1,MMP9IL-10 Signaling 3.64 0.0441 IL1R2,MAPK14,IL1RNLeukocyte Extravasation Signaling 3.36 0.0195 GNAI3,MAPK14,ITGAM,MMP9p38 MAPK Signaling 2.95 0.0256 IL1R2,MAPK14,IL1RNAtherosclerosis Signaling 2.88 0.0242 IL1RN,SERPINA1,MMP9IL-6 Signaling 2.85 0.0236 IL1R2,MAPK14,IL1RNInhibition of Angiogenesis by TSP1 2.84 0.0625 MAPK14,MMP9Glucocorticoid Receptor Signaling 2.83 0.0141 IL1R2,MAPK14,BAG1,IL1RNIL-17A Signaling in Fibroblasts 2.77 0.0571 MAPK14,LCN2Notch Signaling 2.72 0.0541 NUMB,PSEN1Acute Phase Response Signaling 2.5 0.0179 MAPK14,IL1RN,SERPINA1Amyloid Processing 2.46 0.04 MAPK14,PSEN1Agranulocyte Adhesion and Diapedesis 2.45 0.0171 GNAI3,IL1RN,MMP9NF-κB Signaling 2.44 0.0169 IL1R2,LCK,IL1RNMolecular Mechanisms of Cancer 2.42 0.0109 GNAI3,MAPK14,CFLAR,PSEN1IL-8 Signaling 2.31 0.0153 GNAI3,ITGAM,MMP9LPS/IL-1 Mediated Inhibition of RXR Function 2.24 0.0144 IL1R2,IL1RN,ACSL1CCR5 Signaling in Macrophages 2.21 0.0299 GNAI3,MAPK14Chemokine Signaling 2.2 0.0294 GNAI3,MAPK14Caveolar-mediated Endocytosis Signaling 2.16 0.0282 CD55,ITGAMArginine Degradation I (Arginase Pathway) 2.15 0.25 ARG1Acetate Conversion to Acetyl-CoA 2.15 0.25 ACSL1Toll-like Receptor Signaling 2.14 0.0274 MAPK14,IL1RNRole of Osteoblasts, Osteoclasts and Chondrocytes in Rheumatoid Arthritis 2.14 0.0132 IL1R2,MAPK14,IL1RNIL-15 Signaling 2.1 0.0263 LCK,MAPK14Rapoport-Luebering Glycolytic Shunt 2.06 0.2 BPGMPEDF Signaling 2.02 0.0238 MAPK14,CFLARUrea Cycle 1.98 0.167 ARG1Arginine Degradation VI (Arginase 2 Pathway) 1.98 0.167 ARG1Glycogen Biosynthesis II (from UDP-D-Glucose) 1.98 0.167 GYG1PPAR Signaling 1.96 0.0222 IL1R2,IL1RNIL-1 Signaling 1.95 0.022 GNAI3,MAPK14Cholecystokinin/Gastrin-mediated Signaling 1.87 0.0198 MAPK14,IL1RNAirway Pathology in Chronic Obstructive Pulmonary Disease 1.86 0.125 MMP9Citrulline Biosynthesis 1.86 0.125 ARG1Corticotropin Releasing Hormone Signaling 1.82 0.0187 GNAI3,MAPK14Role of Macrophages, Fibroblasts and Endothelial Cells in Rheumatoid Arthritis 1.81 0.01 IL1R2,MAPK14,IL1RNPaxillin Signaling 1.8 0.0182 MAPK14,ITGAMHIF1α Signaling 1.78 0.0177 MAPK14,MMP9Role of Tissue Factor in Cancer 1.73 0.0168 LCK,MAPK14FXR/RXR Activation 1.69 0.016 IL1RN,SERPINA1CCR3 Signaling in Eosinophils 1.69 0.0159 GNAI3,MAPK14
Supplementary Table 4: Canonical pathways of differentially expressed genes between cases and controls in combined microarrayand RT- qPCR experiments
Page 30 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
GNRH Signaling 1.68 0.0157 GNAI3,MAPK14Role of IL-17A in Psoriasis 1.65 0.0769 S100A9Fatty Acid Activation 1.65 0.0769 ACSL1Superpathway of Citrulline Metabolism 1.61 0.0714 ARG1IL-12 Signaling and Production in Macrophages 1.58 0.0139 MAPK14,SERPINA1Parkinson's Signaling 1.56 0.0625 MAPK14Relaxin Signaling 1.55 0.0134 GNAI3,MMP9γ-linolenate Biosynthesis II (Animals) 1.53 0.0588 ACSL1Mitochondrial L-carnitine Shuttle Pathway 1.53 0.0588 ACSL1Oxidative Ethanol Degradation III 1.53 0.0588 ACSL1Hepatic Cholestasis 1.51 0.0127 IL1R2,IL1RNPPARα/RXRα Activation 1.47 0.012 IL1R2,MAPK14Tec Kinase Signaling 1.46 0.0119 GNAI3,LCKEthanol Degradation IV 1.44 0.0476 ACSL1Differential Regulation of Cytokine Production in Intestinal Epithelial Cells by IL-17A and IL-17F1.4 0.0435 LCN2Role of NFAT in Regulation of the Immune Response 1.4 0.0111 GNAI3,LCKHepatic Fibrosis / Hepatic Stellate Cell Activation 1.4 0.011 IL1R2,MMP9Dendritic Cell Maturation 1.39 0.011 MAPK14,IL1RNEndothelin-1 Signaling 1.39 0.0109 GNAI3,MAPK14IL-22 Signaling 1.38 0.0417 MAPK14Glycolysis I 1.38 0.0417 BPGMRegulation of the Epithelial-Mesenchymal Transition Pathway 1.37 0.0107 MMP9,PSEN1IL-17A Signaling in Gastric Cells 1.37 0.04 MAPK14Role of JAK family kinases in IL-6-type Cytokine Signaling 1.37 0.04 MAPK14Gluconeogenesis I 1.37 0.04 BPGMRole of NFAT in Cardiac Hypertrophy 1.36 0.0106 GNAI3,MAPK14Production of Nitric Oxide and Reactive Oxygen Species in Macrophages 1.35 0.0104 MAPK14,SERPINA1Clathrin-mediated Endocytosis Signaling 1.33 0.0102 NUMB,SERPINA1Thrombin Signaling 1.32 0.01 GNAI3,MAPK14Fatty Acid β-oxidation I 1.29 0.0333 ACSL14-1BB Signaling in T Lymphocytes 1.28 0.0323 MAPK14G Protein Signaling Mediated by Tubby 1.28 0.0323 LCKEthanol Degradation II 1.26 0.0312 ACSL1Systemic Lupus Erythematosus Signaling 1.26 0.00922 LCK,IL1RNCoagulation System 1.22 0.0286 SERPINA1Cardiac Hypertrophy Signaling 1.2 0.00862 GNAI3,MAPK14April Mediated Signaling 1.19 0.0263 MAPK14Inhibition of Matrix Metalloproteases 1.19 0.0263 MMP9Role of PKR in Interferon Induction and Antiviral Response 1.17 0.025 MAPK14B Cell Activating Factor Signaling 1.17 0.025 MAPK14Neuroprotective Role of THOP1 in Alzheimer's Disease 1.17 0.025 MMP9Stearate Biosynthesis I (Animals) 1.17 0.025 ACSL1Role of Hypercytokinemia/hyperchemokinemia in the Pathogenesis of Influenza 1.16 0.0244 IL1RNUVC-Induced MAPK Signaling 1.15 0.0238 MAPK14iNOS Signaling 1.14 0.0233 MAPK14Primary Immunodeficiency Signaling 1.13 0.0227 LCKGraft-versus-Host Disease Signaling 1.13 0.0227 IL1RN
Page 31 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Role of Cytokines in Mediating Communication between Immune Cells 1.06 0.0192 IL1RNRetinoic acid Mediated Apoptosis Signaling 1.01 0.0172 CFLARMSP-RON Signaling Pathway 1.01 0.0169 ITGAMCalcium-induced T Lymphocyte Apoptosis 0.999 0.0167 LCKIL-2 Signaling 0.973 0.0156 LCKUVB-Induced MAPK Signaling 0.96 0.0152 MAPK14Role of IL-17A in Arthritis 0.954 0.0149 MAPK14EGF Signaling 0.948 0.0147 MAPK14Melatonin Signaling 0.942 0.0145 GNAI3Glioma Invasiveness Signaling 0.936 0.0143 MMP9Role of MAPK Signaling in the Pathogenesis of Influenza 0.936 0.0143 MAPK14ErbB4 Signaling 0.936 0.0143 PSEN1GPCR-Mediated Integration of Enteroendocrine Signaling Exemplified by an L Cell 0.93 0.0141 GNAI3Role of PI3K/AKT Signaling in the Pathogenesis of Influenza 0.919 0.0137 GNAI3Ephrin B Signaling 0.919 0.0137 GNAI3STAT3 Pathway 0.919 0.0137 MAPK14BMP signaling pathway 0.913 0.0135 MAPK14CD40 Signaling 0.897 0.013 MAPK14IL-17A Signaling in Airway Cells 0.897 0.013 MAPK14ATM Signaling 0.887 0.0127 MAPK14FLT3 Signaling in Hematopoietic Progenitor Cells 0.872 0.0122 MAPK14Communication between Innate and Adaptive Immune Cells 0.872 0.0122 IL1RNAltered T Cell and B Cell Signaling in Rheumatoid Arthritis 0.867 0.012 IL1RNGPCR-Mediated Nutrient Sensing in Enteroendocrine Cells 0.862 0.0119 GNAI3α-Adrenergic Signaling 0.857 0.0118 GNAI3IL-17 Signaling 0.857 0.0118 MAPK14Neuregulin Signaling 0.852 0.0116 PSEN1LPS-stimulated MAPK Signaling 0.852 0.0116 MAPK14NF-κB Activation by Viruses 0.852 0.0116 LCKBladder Cancer Signaling 0.852 0.0116 MMP9TGF-β Signaling 0.848 0.0115 MAPK14G Beta Gamma Signaling 0.843 0.0114 GNAI3Factors Promoting Cardiogenesis in Vertebrates 0.838 0.0112 MAPK14FGF Signaling 0.834 0.0111 MAPK14Death Receptor Signaling 0.83 0.011 CFLARReelin Signaling in Neurons 0.825 0.0109 LCKErbB Signaling 0.804 0.0103 MAPK14CTLA4 Signaling in Cytotoxic T Lymphocytes 0.8 0.0102 LCKCDK5 Signaling 0.8 0.0102 MAPK14RANK Signaling in Osteoclasts 0.796 0.0101 MAPK14Antioxidant Action of Vitamin C 0.796 0.0101 MAPK14UVA-Induced MAPK Signaling 0.788 0.0099 MAPK14Virus Entry via Endocytic Pathways 0.784 0.0098 CD55SAPK/JNK Signaling 0.78 0.00971 LCKMouse Embryonic Stem Cell Pluripotency 0.772 0.00952 MAPK14Type I Diabetes Mellitus Signaling 0.769 0.00943 MAPK14T Cell Receptor Signaling 0.761 0.00926 LCK
Page 32 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Androgen Signaling 0.754 0.00909 GNAI3p53 Signaling 0.751 0.00901 MAPK14Axonal Guidance Signaling 0.741 0.00452 GNAI3,MMP9iCOS-iCOSL Signaling in T Helper Cells 0.73 0.00855 LCKPancreatic Adenocarcinoma Signaling 0.726 0.00847 MMP9Fc Epsilon RI Signaling 0.723 0.0084 MAPK14Renin-Angiotensin Signaling 0.723 0.0084 MAPK14Natural Killer Cell Signaling 0.72 0.00833 LCKfMLP Signaling in Neutrophils 0.72 0.00833 GNAI3Gαi Signaling 0.72 0.00833 GNAI3Sphingosine-1-phosphate Signaling 0.713 0.0082 GNAI3Type II Diabetes Mellitus Signaling 0.704 0.008 ACSL1CD28 Signaling in T Helper Cells 0.701 0.00794 LCKPKCθ Signaling in T Lymphocytes 0.698 0.00787 LCKCdc42 Signaling 0.692 0.00775 MAPK14p70S6K Signaling 0.689 0.00769 GNAI3Th1 Pathway 0.689 0.00769 PSEN1Role of Pattern Recognition Receptors in Recognition of Bacteria and Viruses 0.686 0.00763 C3AR1HMGB1 Signaling 0.686 0.00763 MAPK14P2Y Purigenic Receptor Signaling Pathway 0.686 0.00763 GNAI3Synaptic Long Term Depression 0.66 0.00714 GNAI3Ovarian Cancer Signaling 0.652 0.00699 MMP9Th2 Pathway 0.644 0.00685 PSEN1Regulation of eIF4 and p70S6K Signaling 0.624 0.00649 MAPK14Dopamine-DARPP32 Feedback in cAMP Signaling 0.614 0.00633 GNAI3CXCR4 Signaling 0.6 0.0061 GNAI3Mitochondrial Dysfunction 0.598 0.00606 PSEN1Gap Junction Signaling 0.595 0.00602 GNAI3Germ Cell-Sertoli Cell Junction Signaling 0.589 0.00592 MAPK14RhoGDI Signaling 0.582 0.00581 GNAI3Ephrin Receptor Signaling 0.582 0.00581 GNAI3Sertoli Cell-Sertoli Cell Junction Signaling 0.58 0.00578 MAPK14Th1 and Th2 Activation Pathway 0.565 0.00556 PSEN1CREB Signaling in Neurons 0.561 0.00549 GNAI3B Cell Receptor Signaling 0.561 0.00549 MAPK14RAR Activation 0.551 0.00535 MAPK14AMPK Signaling 0.549 0.00532 MAPK14NRF2-mediated Oxidative Stress Response 0.545 0.00526 MAPK14ILK Signaling 0.541 0.00521 MMP93-phosphoinositide Biosynthesis 0.539 0.00518 LCKBreast Cancer Regulation by Stathmin1 0.523 0.00495 GNAI3Integrin Signaling 0.505 0.00472 ITGAMcAMP-mediated signaling 0.492 0.00455 GNAI3Superpathway of Inositol Phosphate Compounds 0.481 0.00441 LCKPhospholipase C Signaling 0.47 0.00427 LCKColorectal Cancer Metastasis Signaling 0.458 0.00413 MMP9Signaling by Rho Family GTPases 0.453 0.00407 GNAI3
Page 33 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from

Confidential: For Review Only
Protein Ubiquitination Pathway 0.441 0.00394 BAG1Xenobiotic Metabolism Signaling 0.422 0.00372 MAPK14G-Protein Coupled Receptor Signaling 0.42 0.0037 GNAI3Protein Kinase A Signaling 0.313 0.00266 GNAI3
Page 34 of 34
https://mc.manuscriptcentral.com/bmjpo
BMJ Paediatrics Open
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
on June 3, 2020 by guest. Protected by copyright.
http://bmjpaedsopen.bm
j.com/
bmjpo: first published as 10.1136/bm
jpo-2017-000092 on 31 August 2017. D
ownloaded from




![University of Groningen Pneumococcal conjugate vaccines ... › ws › files › 67431176 › Fortanier_et... · [Intervention Review] Pneumococcal conjugate vaccines for preventing](https://img.dokumen.tips/doc/110x75/5f03b7c17e708231d40a6d8d/university-of-groningen-pneumococcal-conjugate-vaccines-a-ws-a-files-a.jpg)
























