Embed Size (px)
Citation preview

Preclinical Development
Assessing the Activity of Cediranib, a VEGFR-2/3 TyrosineKinase Inhibitor, against VEGFR-1 and Members of theStructurally Related PDGFR Family
Sandra R. Brave1, Kirsty Ratcliffe1, Zena Wilson1, Neil H. James1, Sue Ashton1, Anna Wainwright1,Jane Kendrew1, Philippa Dudley1, Nicola Broadbent1, Graham Sproat1, Sian Taylor1, Claire Barnes1,Jeffrey C. Silva2, Charles L. Farnsworth2, Laurent Hennequin3, Donald J. Ogilvie1,Juliane M. J€urgensmeier1, Masabumi Shibuya4, Stephen R. Wedge1, and Simon T. Barry1
AbstractCediranib is a potent inhibitor of the VEGF receptor (VEGFR)-2 and VEGFR-3 tyrosine kinases. This study
assessed the activity of cediranib against the VEGFR-1 tyrosine kinase and the platelet-derived growth factor
receptor (PDGFR)-associated kinases c-Kit, PDGFR-a, and PDGFR-b. Cediranib inhibited VEGF-A–stimu-
lated VEGFR-1 activation in AG1-G1-Flt1 cells (IC50 ¼ 1.2 nmol/L). VEGF-A induced greatest phosphoryla-
tion of VEGFR-1 at tyrosine residues Y1048 and Y1053; this was reversed by cediranib. Potency against
VEGFR-1 was comparable with that previously observed versus VEGFR-2 and VEGFR-3. Cediranib also
showed significant activity against wild-type c-Kit in cellular phosphorylation assays (IC50¼ 1–3 nmol/L) and
in a stem cell factor–induced proliferation assay (IC50 ¼ 13 nmol/L). Furthermore, phosphorylation of wild-
type c-Kit in NCI-H526 tumor xenografts was reduced markedly following oral administration of cediranib
(�1.5 mg/kg/d) to tumor-bearing nude mice. The activity of cediranib against PDGFR-b and PDGFR-a was
studied in tumor cell lines, vascular smooth muscle cells (VSMC), and a fibroblast line using PDGF-AA and
PDGF-BB ligands. Both receptor phosphorylation (IC50 ¼ 12–32 nmol/L) and PDGF-BB–stimulated cellular
proliferation (IC50 ¼ 32 nmol/L in human VSMCs; 64 nmol/L in osteosarcoma cells) were inhibited. In vivo,
ligand-induced PDGFR-b phosphorylation in murine lung tissue was inhibited by 55% following treatment
with cediranib at 6 mg/kg but not at 3 mg/kg or less. In contrast, in C6 rat glial tumor xenografts in mice,
ligand-induced phosphorylation of both PDGFR-a and PDGFR-bwas reduced by 46% to 61%with 0.75mg/kg
cediranib. Additional selectivity was showed versus Flt-3, CSF-1R, EGFR, FGFR1, and FGFR4. Collectively,
these data indicate that cediranib is a potent pan-VEGFR kinase inhibitorwith similar activity against c-Kit but
is significantly less potent than PDGFR-a and PDGFR-b. Mol Cancer Ther; 10(5); 861–73. �2011 AACR.
Introduction
Inhibition of VEGF-A–mediated signaling providestherapeutic benefit in oncology as monotherapy in the
treatment of renal cell cancer (1, 2) and, when combinedwith certain cytotoxic therapies, in other disease settings(3). Approaches used to inhibit VEGF receptor (VEGFR)signaling include small molecule inhibitors that bind intothe ATP-binding pocket within the kinase domain ofVEGFR-2 to prevent ATP catalysis and propagation ofreceptor signaling. This receptor predominantly trans-duces the angiogenic and permeability activity of VEGF-A. Two other VEGF receptors are also described withdifferential binding of VEGF family ligands. Each smallmolecule differs in its potency against the VEGF recep-tors, selectivity versus other kinases, physicochemicalproperties, and pharmacokinetic profile (4, 5). SomeVEGFR tyrosine kinase inhibitors have additional activityagainst 1 ormoremembers of the platelet-derived growthfactor receptor (PDGFR) family of kinases (class III),which comprises PDGFR-a, PDGFR-b, c-Kit, colony-sti-mulating factor receptor-1, and Flt-3; these receptors havesome structural homology with the VEGFR family mem-bers in that each harbors a kinase-insert sequence inits intracellular domain (6). However, VEGFR kinase
Authors' Affiliations: 1Oncology iMED, AstraZeneca, Alderley Park, Mac-clesfield, Cheshire, United Kingdom; 2Cell Signaling Technology, Danvers,Machigan; 3AstraZeneca Pharma, Centre de Recherches, Z.I. La Pom-pelle, Chemin de Vrilly, Reims, France; and 4Department of MolecularOncology, Tokyo Medical and Dental University, Tokyo, Japan
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
Current address for D.J. Ogilvie: Paterson Institute for Cancer Research,Wilmslow Rd., Manchester M20 4BX, United Kingdom.
Corresponding Author: Simon Barry, Oncology iMED, AstraZeneca,Alderley Park, Macclesfield, Cheshire SK10 4TG, United Kingdom. Phone:44-1625-513350; Fax: 44-1625-513350.E-mail: [email protected] or Steve Wedge, OncologyiMED, AstraZeneca, Alderley Park, Macclesfield, Cheshire, SK10 4TG,United Kingdom. Phone: 44-1625-523236; Fax: 44-1625-513350.E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-10-0976
�2011 American Association for Cancer Research.
MolecularCancer
Therapeutics
www.aacrjournals.org 861
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

inhibitors have also been described with activity againsttyrosine kinases outside the PDGFR class and, in somecases, against serine/threonine kinases. Inhibiting multi-ple kinases simultaneously may provide additional ther-apeutic opportunities in defined disease settings but mayimpact adversely on tolerability, particularly if chronicadministration or usage in combination with concurrentcytotoxic treatment is required.
Cediranib has been previously shown to be a potentinhibitor of VEGFR-2 and VEGFR-3 signaling in cellularassays and to inhibit the growth of both angiogenic bloodvessels and lymphangiogenic vessels in vivo (7–9). Inrecombinant kinase assays, cediranib also inhibitsVEGFR-1 kinase activity (IC50 ¼ 5 nmol/L) within asimilar concentration range to VEGFR-2 and VEGFR-3(IC50 values of <0.1 and �3 nmol/L, respectively; ref. 7).However, a more quantitative assessment of inhibitoractivity against endogenous VEGFR-1 kinase has proventechnically challenging in endothelial cells because of thelow intrinsic activity of this receptor. Cediranib showsselectivity against other kinases (7). Cediranib has similarpotency against c-Kit when compared with VEGFR-2 inphosphorylation assays but less potency against PDGFR-a and PDGFR-b, particularly in a PDGF-AA/PDGFR-a–driven tumor cell proliferation assay. However, theactivity of cediranib against a c-Kit–driven phenotypicendpoint, PDGFR-a and -b signaling in normal cell types(which may also influence therapeutic response), or acomparative inhibition of these targets in vivo was notpreviously examined.
This study primarily aimed to further describe thepharmacology of cediranib by (i) determining activityagainst VEGFR-1 in cells, (ii) using a wider complementof cell lines and assays to examine activity against c-Kitand PDGFR-a/b signaling in vitro, and (iii) to examinepharmacodynamic inhibition of c-Kit, PDGFR-a, andPDGFR-b in vivo over a dose range in which cediranibhas previously shown activity in tumor models. The dataconfirm that cediranib is primarily a pan-VEGFR inhibi-tor that can inhibit wild-type c-Kit. The data also suggestthat cediranib may have some partial pharmacodynamicactivity against PDGFR-a and PDGFR-b receptor activa-tion in tumors, although this inhibition may be of limitedfunctional relevance but does inhibit FGFR1 and FGFR4.
Materials and Methods
ReagentsCediranib [(4-fluoro-2-methyl-1H-indol-5-yl)oxy]-6-me-
thoxy-7-(3-pyrrolidin-1-ylpropoxy)quinazoline(AZD2171;ref. Fig. 1) was synthesized according to the processesdescribed inWO 00/47212, in particular those describedin example 240 of WO/47212. The free base of cediranibwas used in these preclinical studies, with a molecularweight of 450.51. For all in vitro assays, cediranib wasprepared initially as a 10 mmol/L stock solution indimethyl sulphoxide (DMSO) and diluted in the rele-vant assay media, such that the final concentration of
DMSO did not exceed 0.01%, with the exception ofstudies examining direct effects on tumor cells in which1% DMSO was required to examine higher concentra-tions of cediranib. All in vivo studies were conducted byonce-daily oral gavage. For studies in mice, cediranibwas suspended in 1% (w/v) aqueous polysorbate 80[polyoxyethylene (20) sorbitan monooleate in deionizedwater] and dosed at 0.1 mL/10 g of body weight.
Cell cultureNCI-H526 [a human small cell lung cancer (SCLC)
line], U118MG (a human glioblastoma line), MG63 (ahuman osteosarcoma line), and C6 (a rat glial line) cellswere purchased from the American Type Culture Collec-tion); no further authentication was done on these lines.The M07e [a human acute myelogenous leukemia (AML)line] cells were purchased from the German Collection ofMicroorganisms and Cell Cultures (Deutsche Sammlungvon Mikroorganismen und Zellkulturen GmbH (GermanCollection of Microorganisms and Cell Cultures); nofurther authentication was done. NIH 3T3 (mouse fibro-blast) cells were obtained fromA.Wong, Jefferson CancerInstitute; no further authentication was done. Humanaortic and coronary vascular smoothmuscle cells (VSMC)were purchased from PromoCell GmbH. All cell lineswere routinely passaged less than 10 times with theexception of the primary vascular cells, which werepassaged no more than 4 times. NCI-H526, U118MG,MG63, C6, M07e, NIH 3T3, human aortic, and coronaryVSMCs were maintained in culture as per providers’recommendation. M07e cells were maintained in culturein the presence of interleukin-3 (5 ng/mL) and granulo-cyte macrophage colony stimulating factor (5 ng/mL).
Inhibition of growth factor–stimulated receptorphosphorylation in vitro
The ability of cediranib to inhibit receptor phosphor-ylation in cells was determined using Western blotting.Cells were serum starved overnight in the presence(M07e and NCI-H526) or absence (aortic and coronaryVSMC) of 0.1% bovine serum albumin or in the presenceof 1% charcoal-stripped serum (MG63, U118MG, C6, andNIH 3T3 cells). Cells were then incubated with cediranibfor 60 to 120 minutes and stimulated with the relevantligand: stem cell factor (SCF; 50 ng/mL) and PDGF-AA or
O
N
N
O
O
F
N
N
Figure 1. The structure of cediranib.
Brave et al.
Mol Cancer Ther; 10(5) May 2011 Molecular Cancer Therapeutics862
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

PDGF-BB (50 ng/mL) for 5 to 10 minutes. SCF wasobtained from R&D Systems and PDGF-AA andPDGF-BB from Sigma-Aldrich. Cell lysates of NCI-H526, M07e, and aortic and coronary VSMCs were pre-pared in lysis buffer I (50 mmol/L Tris-HCl, pH 7.6; 137mmol/L sodium chloride; 10% glycerol; 0.1% Igepal;0.1% SDS; 50 mmol/L sodium fluoride; 1 mmol/Lsodium orthovanadate and cocktail protease inhibitortablets (Roche Diagnostics Ltd.). Cell lysates of MG63,U118MG, C6, and NIH 3T3 cells were prepared in lysisbuffer 2 (10% glycerol, 2% SDS, 50 mmol/L Tris-HCl, 200mmol/L 2-mercaptoethanol).The protein concentration in the lysates was deter-
mined using a bicinchoninic acid assay kit (Pierce) andWestern blottingwas done onwhole cell lysates (50–75 mgof protein loaded per lane), using standard SDS-PAGEmethods with detection by enhanced chemilumines-cence. Total and phosphorylated proteins weremeasuredusing antibodies to c-Kit [Cell Signaling Technology(CST) #3391], and phosphorylated c-Kit [(pc-Kit)SC5535; Santa Cruz Biotechnology]; PDGFR-a (AF-307;R&D Systems), PDGFR-a (CST #3164), and phosphory-lated PDGFR-a [(pPDGFR-a) SC12910; Santa CruzBiotechnology]; PDGFR-b (AF-385; R&D Systems),PDGFR-b (1469-1; Epitomics), phosphorylated PDGFR-b [(pPDGFR-b) CST #3161]; mitogen-activated proteinkinase [(MAPK) CST #9102) and phosphorylated MAPK[(pMAPK) CST #9101]. Phosphorylation was quantitatedusing the ChemiGenius Imaging System for Chemilumi-nescence (Syngene) with the exception of the humancoronary VSMCs, which were quantified by ELISA (goatanti-PDGFR Alpha R&D Systems AF-307-NA Capture,DuoSet detection; R&D Systems).AG1-G1-Flt-1 cells were established with the permis-
sion of the Ethics Committee for Scientific Research at theInstitute of Medical Science, University of Tokyo, Tokyo,Japan. Briefly, a human adult benign angioma wasexcised surgically and plated with Ham’s F-12 nutrientmixture medium (Invitrogen) supplemented with 10%FBS (JRH Biosciences) and 40 mg/mL kanamycin (Wako).A pEF1a-SV40 large T antigen plasmid was introducedinto the cells, using DMSO and polybrene (SIGMA-Aldrich). An SV40 large T-positive clone AG1-G1 cellwas isolated and then pBCMGS-Neo-Flt-1 carrying thefull length of Flt-1 cDNA (10), or the empty vectorpBCMGS-Neo plasmid, was transfected into AG1-G1 cellby the Effectene Transfection Reagent (Qiagen). Cloneselection and culture were done with Ham’s F-12 med-ium containing 10% FBS, 40 mg/mL kanamycin, and 400mg/mL geneticin G418 (Wako). G418 was decreased to200 mg/mL in regular culture. To examine inhibition ofVEGFR-1 phosphorylation, cells were placed in serum-free media overnight and then incubated with cediranibfor 90 minutes and stimulated with VEGF 50 ng/mL(R&D Systems) for the last 5 minutes of incubation. Celllysates were prepared in lysis buffer 1 and phosphory-lated VEGFR-1 (pVEGFR-1) was evaluated using MesoScale methodology [Meso Scale Discovery (MSD)]. The
pVEGFR-1 was analyzed by MSD ELISA. Total VEGFR-1antibody (MAB321; R&D Systems Inc.) was spotted ontohigh-binding MSD plates and incubated for 2 hours atroom temperature, after which time plates were blockedand then washed. Cell lysates were added and incubatedovernight at 4�C. Plates were washed and then the sul-photagged-pY20 detection antibody was added and leftto incubate for 1 hour at room temperature. Followingwashing, read buffer was added and plates were readimmediately on the SECTOR Imager 6000 (MSD). Tovisualize pVEGFR-1 by Western blotting, VEGFR-1 wasimmunoprecipitated with an antibody to total VEGFR-1(SC316; Santa Cruz Biotechnology) and immunoblottedwith the anti-phosphotyrosine antibody PY20 (SC508;Santa Cruz Biotechnology). Levels of total VEGFR-1 wereconfirmed by immunoblotting with SC316.
Sample preparation and mass spectrometry foridentification of phosphotyrosine modification ofVEGFR-1
Analysis of VEGFR-1–phosphorylated epitope changeswas done using Cell Signaling Technology’s proprietaryPhosphoScan methodology. AG1-G1-Flt-1 cells wereplaced in serum-free media overnight and stimulatedwith VEGF (50 ng/mL) or placenta growth factor (PlGF;100 ng/mL; R&D Systems) for 5 minutes [the former alsowith and without cediranib (100 nmol/L; 85 minutespreincubation and 5 minutes coincubation with ligand)].Protein extracts fromAG1-G1-Flt1 cells were prepared bysuspending cells in Urea Lysis Buffer (20 mmol/LHEPES, pH 8.0, 9.0 mol/L urea, 1 mmol/L sodiumorthovanadate, 2.5 mmol/L sodium pyrophosphate, 1mmol/L b-glycerol phosphate). Lysates generated fromapproximately 2 � 108 cells were prepared for eachsample condition (control, VEGF-treated, PlGF-treated,and VEGF þ cediranib-treated). The resulting proteinextracts (40 mg total protein each) were then reducedwith dithiothreitol (4.5 mmol/L), carboxamidomethy-lated using iodoacetamide (10.0 mmol/L), and subse-quently digested with trypsin (1:100 weight, trypsin tototal protein). Peptides were separated from nonpeptidematerial by solid-phase extraction with Sep-Pak C18cartridges. Lyophilized peptides were redissolved, andphosphorylated peptides were isolated using a slurry ofimmobilized phosphorylated tyrosine antibody (CST#9411) conjugated to protein G agarose beads (Roche).Peptides were eluted from antibody resin into a totalvolume of 100 mL in 0.15% trifluoroacetic acid. Elutedpeptides were concentrated with PerfectPure C18 tipsimmediately before liquid chromatography/mass spec-trometry (LC/MS) analysis. Peptides were loadeddirectly onto a 10 cm � 75 mm PicoFrit capillary columnpacked with Magic C18 AQ reversed-phase resin. Thecolumn was developed with a 45-minute linear gradientof acetonitrile in 0.125% formic acid delivered at280 nL/min. Tandem mass spectra were collected witha linear trap quadrupole (LTQ)-orbitrap hybrid massspectrometer, using a top-ten method, a dynamic
Assessment of Cediranib Selectivity
www.aacrjournals.org Mol Cancer Ther; 10(5) May 2011 863
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

exclusion repeat count of 1, and a repeat duration of 30seconds. MS spectra were collected in the orbitrap com-ponent of the mass spectrometer, and MS/MS spectrawere collected in the LTQ. MS/MS spectra were evalu-ated using TurboSequest in the Proteomics Browserpackage (v. 27, rev. 13). The ratios of the integrated peakheight intensities for phosphopeptide quantificationwere obtained using the XCalibur software 2.0.7 (Ther-moFinnigan). A reduction in peak intensity in VEGF-treated cells compared with control, PlGF-treated cellscompared with control, or VEGF plus cediranib-treatedcells compared with control was expressed as a negativefold change value. All integrated peak intensity calcula-tions were manually reviewed to ensure proper integra-tion of consistently shaped, coeluting peaks. Changes inphosphorylated peptide levels were measured by takingthe ratio of raw intensities between control and treatedcells, with the untreated sample as the reference (denomi-nator) in each case. Raw intensity ratios were normalizedusing a median adjustment technique whereby the log2ratios comprising each binary comparison (inhibitor trea-ted versus control) was independently and globallyadjusted such that the normalized median log2 ratio iszero. The normalized log2 ratios were then converted totheir corresponding normalized fold changes.
Inhibition of growth factor–mediated cellularproliferation
NCI-H526 cells were used to determine the effect ofcediranib on SCF-stimulated proliferation. Cells wereseeded at a density of 1� 105 per mL in 96-well microtiterplates in phenol red–free low-serum containing media(0.2% FBS) overnight. The following day cells were pre-treated with cediranib (0.1–100 nmol/L) for 30 minutesbefore stimulation with 50 ng/mL SCF and then incuba-tion for 72 hours at 37�C. Cell proliferation was deter-mined using an XTT endpoint (Roche Diagnostics Ltd.).All assays were done in triplicate, and the mean � SEMwas calculated from 6 independent experiments. Humanaortic VSMCs were used to determine the effect of cedir-anib on PDGF-BB–stimulated proliferation. Cells wereseeded at 10,000 cells per well in black-walled 96-wellplates in smooth muscle cell growth medium 2 (Promo-cell GmbH) and incubated overnight at 37�C. The follow-ing day, the medium was replaced with Dulbecco’sModified Eagle’s Medium (DMEM; Gibco) containing0.1% FBS, PDGF-BB (50 ng/mL), and cediranib. After24-hour incubation, a bromodeoxyuridine (BrdU) reagent(Amersham) was added and cells were incubated for afurther 24 hours at 37�C. Cells were fixed in formalin for15minutes, and proliferationwas assessed by staining forBrdU by using the Cell Proliferation Fluorescence Kit(Amersham). Cells were imaged on the ArrayScan (Cel-lomics). All assays were done in triplicate, and the mean� SEM was calculated from 3 independent experiments.MG63 cells were used to determine the effect of cediranibon PDGF-AA- and PDGF-BB–stimulated proliferation.Cells were seeded at 1,500 cells per well in 96-well plates
in phenol red DMEM containing 1% charcoal-strippedserum for 24 hours at 37�C. The following day, themedium was replaced with DMEM (Sigma-Aldrich) con-taining PDGF-AA or PDGF-BB (50 ng/mL) and cediranibfor a further 72 hours. Cell proliferation was determinedas described earlier. All assays were done in triplicate,and the mean � SEM was calculated from 3 independentexperiments.
Inhibition of receptor phosphorylation in vivoThe activity of cediranib was evaluated in an NCI-
H526 human SCLC tumor xenograft model. Tumorswere implanted subcutaneously in the hind flank offemale nude (nu/nu genotype) mice of at least 8 weeksof age. When tumors reached a volume of 0.36 � 0.02cm3, mice were randomized (8–10 per group) and dosedwith cediranib (0.75–6 mg/kg/d) or vehicle adminis-tered once daily by oral gavage. Tumor volumes wereassessed by bilateral Vernier caliper measurement atleast twice weekly and calculated using the formula(length � width) � H (length � width) � (p/6), wherelength was taken to be the longest diameter across thetumor and width the corresponding perpendicular. Toremove any size dependency before statistical evalua-tion (the variation in mean tumor volume data increasesproportionally with volume and is therefore dispropor-tionate between groups), data were log-transformedbefore statistical evaluation by using a 1-tailed 2-samplet test. NCI-H526 xenograft tumors were evaluatedfor c-Kit receptor phosphorylation ex vivo by usingimmunoprecipitation, following acute (2 days; 8 ani-mals per group) or chronic (17 days; 3 animals pergroup) treatment with cediranib. Tumors were homo-genized in lysis buffer I, and following a protein assay,5 mg of protein from each sample was immunopreci-pated overnight at 4�C with anti-c-Kit–conjugated agar-ose beads. The immune complexes were washed, andproteins were eluted by boiling in SDS sample buffer.Standard SDS-PAGE methods were done to enabledetection of total and pc-Kit, using antibodies as pre-viously described. Protein phosphorylation was quan-titated using the ChemiGenius as described earlier.
The activity of cediranib was also evaluated in a C6 ratglial tumor xenograftmodel inmice. Cellswere cultured in199 media (Life Technologies) supplemented with 10%fetal calf serum and 2 mmol/L glutamine and maintainedin 7.5% CO2. For all tumor studies, C6 glial cells wereresuspended in sterile PBS and inoculated subcutaneously(1� 105 in 100 mL) in the hind flank of male athymic mice.Tumor volumes were assessed as described earlier.Approximately 21dayspostimplantationwhen theC6glialtumor volume was between 0.5 and 0.8 cm3, mice wererandomized into groups of 10 before treatment. Micereceived a single-bolus oral dose of either cediranib orvehicle control. Animalswere sacrificed 4 hours post–doseadministration. Five minutes before sacrifice, VEGF-A (20mg per mouse; AstraZeneca) and PDGF-BB (75 mg permouse; Biosource) were coadministered intravenously
Brave et al.
Mol Cancer Ther; 10(5) May 2011 Molecular Cancer Therapeutics864
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

(0.3 mL/mouse in PBS) to all animals. Terminal bloodsamples were taken from the vena cava into lithiumheparin tubes to collect plasma samples for pharmacoki-netic analysis. Tumors and lungswere excised and halved,and each tissue was half weighed and snap frozen imme-diately in liquid nitrogen. Tissue samples were stored at�80�C until processed for Western blot analysis for totaland phosphorylated VEGFR-2 (pVEGFR-2) and PDGFR-b.Lungswerehomogenized in lysisbuffer I, and tumorswerehomogenized in lysis buffer III [60 mmol/L Tris, 150mmol/L sodium chloride, 1 mmol/L ethylenediaminete-traacetic acid, 1% Igepal, 0.25% sodium deoxycholate, 40%glycerol, 8% SDS, 200 mmol/L Tris-HCl, phosphataseinhibitor cocktails 1 and 2 (Sigma-Aldrich)].
Results
Cediranib is a potent inhibitor of VEGFR-1To determine the potency of cediranib against VEGFR-
1 in cells, a cell line stably transfected with full-lengthVEGFR-1 (AG1-AG-Flt-1) was used. Cediranib inhibitedVEGF-A–driven VEGFR-1 phosphorylation with an IC50
value of 1.2 nmol/L (Fig. 2A). This is comparable with thecellular potency versus VEGFR-2 (0.5 nmol/L; ref. 7) andVEGFR-3 (<1 nmol/L; ref. 8) and consistent with theprimary pharmacology of the compound being that ofa potent pan-inhibitor of VEGFR-1, VEGFR-2, andVEGFR-3 tyrosine kinase activity.To identify the tyrosine phosphorylation sites on
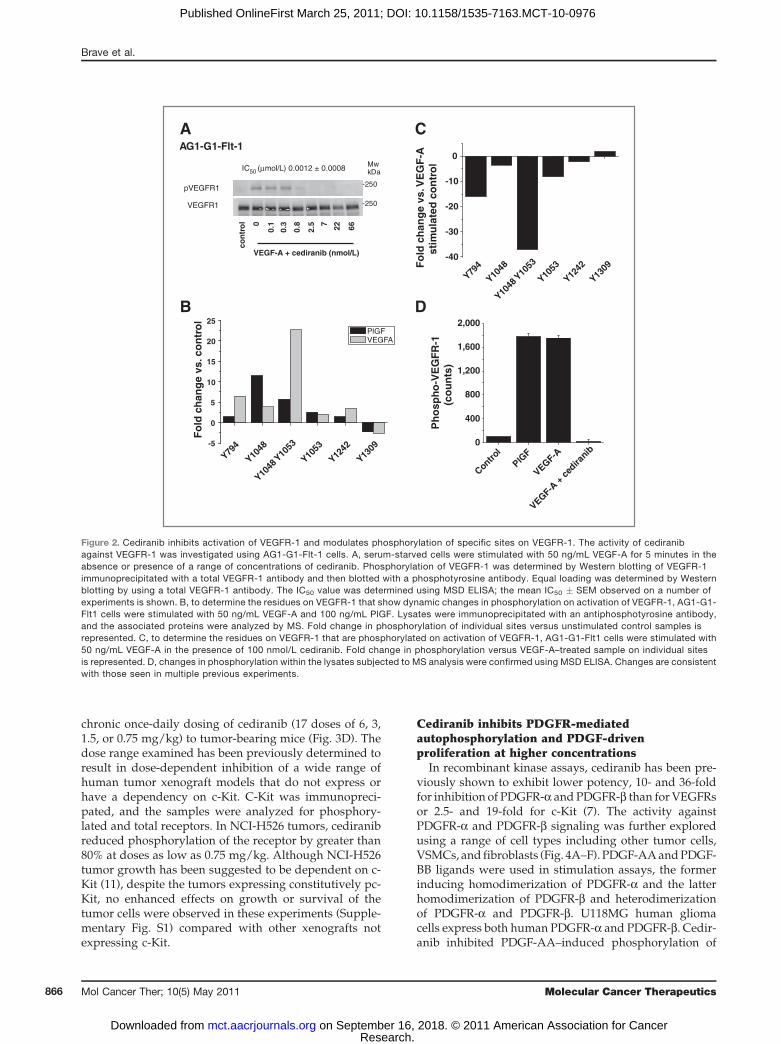
VEGFR-1 modulated by ligand-induced autophosphor-ylation and inhibition by cediranib, Phosphoscan wasdone on VEGFR-1 isolated from the AG1-G1 cells treatedwith VEGF-A and with VEGF-A in the presence of 100nmol/L cediranib. Phosphorylated receptor wasenriched via a total phospho-tyrosine immunoprecipita-tion. The residues phosphorylated on VEGFR-1 in eachtreated lysate were examined by specifically identifyingphosphorylated peptides corresponding to VEGFR-1(Fig. 2A and B; Supplementary Table S1). On stimulationwith VEGF-A or PlGF, significant induction of phosphor-ylation of peptides incorporating tyrosine residuesY1053, Y1048/Y1053, and Y1048 was observed. Modestinduction of phosphorylation was also detected at resi-dues 794 and 1242, but the magnitude of change waslower (Fig. 2A). The pattern of ligand-induced phosphor-ylation by both VEGF-A and PlGF was similar, althoughthe magnitude of induction was higher with VEGF-Athan with PlGF. Serine-phosphorylated peptides werealso detected, although the significance of these modifi-cations is unclear (Supplementary Table S1). This showsthat under these conditions, the phosphorylation status ofVEGFR-1 is dynamically regulated on a restricted num-ber of residues on engaging VEGF-A or PlGF, with Y1048and Y1053 showing the greatest fold changes.To determine which residues were dynamically regu-
lated by cediranib, we compared protein extracts fromcells stimulated with VEGF-A with those from cellsstimulated with VEGF in the presence of 100 nmol/L
cediranib (Fig. 2C). There was a marked reduction in therelative abundance of peptides corresponding to Y794,Y1053, Y1053/Y1048, and Y1048 in cediranib-treatedsamples, with a 37-fold reduction in the presence ofthe peptide corresponding to pY1053/48 in the cedira-nib-treated samples (Fig. 2C). The total tyrosine phos-phorylation status of VEGFR-1 in the lysates used for thisspecific analysis was also assessed by ELISA. VEGFR-1from each lysate was captured, and the level of tyrosinephosphorylation was detected using an antiphosphotyr-osine antibody (Fig. 2D). Both VEGF-A and PlGF inducedsignificant phosphorylation of VEGFR-1 in the lysates.Cediranib inhibited the VEGF-A–induced phosphoryla-tion of VEGFR-1.
Cediranib inhibits c-Kit phosphorylation andSCF-induced proliferation
Cediranib inhibits c-Kit with a similar potency to thatwith which it inhibits the tyrosine kinase activity ofVEGFRs (7). The activity of cediranib against c-Kit wastested in 2 cell lines,M07e andNCI-H526. SCF-stimulatedc-Kit phosphorylation was inhibited with IC50 values of 3and 1 nmol/L, respectively (Fig. 3A and B). MAPK as adownstream signaling marker was also inhibited with anIC50 value similar to that for inhibition of receptor phos-phorylation (Fig. 3A and B). The relationship betweeninhibition of acute ligand-induced phosphorylation andSCF-stimulated c-Kit-dependent proliferation was deter-mined using NCI-H526 cells. Cediranib inhibited SCF-stimulated proliferation of NCI-H526 cells after 72 hourswith an IC50 value of 0.013 nmol/L (Fig. 3C), and fullinhibition being achieved at concentrations between 20and 50 nmol/L. From these experiments, it seems thatabout 10-fold higher concentrations were required toinhibit functional consequences of c-Kit signaling (i.e.,c-Kit–dependent cellular proliferation) than for the inhi-bition of receptor phosphorylation.
Mutations in c-Kit are associated with certain tumorssuch as gastrointestinal stromal tumors (GIST) and AMLin which they drive tumor growth. The activity of cedir-anib against a range of common c-Kit mutations was alsodetermined using a panel of cell lines that eitherexpressed mutated c-Kit endogenously or were transi-ently transfected with mutated receptors. The c-Kit muta-tions assessed were V560G, V559D, W557R, Del 557–558,V654A, T670I, D816V, D816Y, and N822K. To assess thepotential of the compound to inhibit phosphorylation ofthese receptors, cells were incubated in the presence andabsence of 20 nmol/L of cediranib. Cediranib inhibitedphosphorylation of c-Kit mutants V560G, V559D,W557R,and Del 557–558, V654A, and N822K markedly, but it didnot inhibit constitutive phosphorylation of c-Kit mutantsT670I, D816V, and D816Y (Supplementary Table S2).
Inhibition of c-Kit phosphorylation by cediranibin vivo
Inhibition of c-Kit phosphorylation was examined invivo in establishedNCI-H526 tumor xenografts, following
Assessment of Cediranib Selectivity
www.aacrjournals.org Mol Cancer Ther; 10(5) May 2011 865
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976
chronic once-daily dosing of cediranib (17 doses of 6, 3,1.5, or 0.75 mg/kg) to tumor-bearing mice (Fig. 3D). Thedose range examined has been previously determined toresult in dose-dependent inhibition of a wide range ofhuman tumor xenograft models that do not express orhave a dependency on c-Kit. C-Kit was immunopreci-pated, and the samples were analyzed for phosphory-lated and total receptors. In NCI-H526 tumors, cediranibreduced phosphorylation of the receptor by greater than80% at doses as low as 0.75 mg/kg. Although NCI-H526tumor growth has been suggested to be dependent on c-Kit (11), despite the tumors expressing constitutively pc-Kit, no enhanced effects on growth or survival of thetumor cells were observed in these experiments (Supple-mentary Fig. S1) compared with other xenografts notexpressing c-Kit.
Cediranib inhibits PDGFR-mediatedautophosphorylation and PDGF-drivenproliferation at higher concentrations
In recombinant kinase assays, cediranib has been pre-viously shown to exhibit lower potency, 10- and 36-foldfor inhibition of PDGFR-a andPDGFR-b than forVEGFRsor 2.5- and 19-fold for c-Kit (7). The activity againstPDGFR-a and PDGFR-b signaling was further exploredusing a range of cell types including other tumor cells,VSMCs, and fibroblasts (Fig. 4A–F). PDGF-AAandPDGF-BB ligands were used in stimulation assays, the formerinducing homodimerization of PDGFR-a and the latterhomodimerization of PDGFR-b and heterodimerizationof PDGFR-a and PDGFR-b. U118MG human gliomacells express both human PDGFR-a and PDGFR-b. Cedir-anib inhibited PDGF-AA–induced phosphorylation of
AG1-G1-Flt-1 A C
B D
VEGFR1
pVEGFR1
con
tro
l 0
0.1
0.3
0.8
2.5 7 22 66
MwkDa
-250
-250
VEGF-A + cediranib (nmol/L)
IC50 (μmol/L) 0.0012 ± 0.0008
Y794
Y1048
Y1048
Y1053
Y1053
Y1242
Y1309
-5
0
5
10
15
20
25PlGFVEGFA
Fo
ld c
han
ge
vs. c
on
tro
l
Y794
Y1048
Y1048
Y1053
Y1053
Y1242
Y1309
-40
-30
-20
-10
0
Fo
ld c
han
ge
vs. V
EG
F-A
stim
ula
ted
co
ntr
ol
Contro
lPlG
F
VEGF-A
VEGF-A +
ced
irani
b0
400
800
1,200
1,600
2,000
Ph
osp
ho
-VE
GF
R-1
(co
un
ts)
Figure 2. Cediranib inhibits activation of VEGFR-1 and modulates phosphorylation of specific sites on VEGFR-1. The activity of cediranibagainst VEGFR-1 was investigated using AG1-G1-Flt-1 cells. A, serum-starved cells were stimulated with 50 ng/mL VEGF-A for 5 minutes in theabsence or presence of a range of concentrations of cediranib. Phosphorylation of VEGFR-1 was determined by Western blotting of VEGFR-1immunoprecipitated with a total VEGFR-1 antibody and then blotted with a phosphotyrosine antibody. Equal loading was determined by Westernblotting by using a total VEGFR-1 antibody. The IC50 value was determined using MSD ELISA; the mean IC50 � SEM observed on a number ofexperiments is shown. B, to determine the residues on VEGFR-1 that show dynamic changes in phosphorylation on activation of VEGFR-1, AG1-G1-Flt1 cells were stimulated with 50 ng/mL VEGF-A and 100 ng/mL PlGF. Lysates were immunoprecipitated with an antiphosphotyrosine antibody,and the associated proteins were analyzed by MS. Fold change in phosphorylation of individual sites versus unstimulated control samples isrepresented. C, to determine the residues on VEGFR-1 that are phosphorylated on activation of VEGFR-1, AG1-G1-Flt1 cells were stimulated with50 ng/mL VEGF-A in the presence of 100 nmol/L cediranib. Fold change in phosphorylation versus VEGF-A–treated sample on individual sitesis represented. D, changes in phosphorylation within the lysates subjected to MS analysis were confirmed using MSD ELISA. Changes are consistentwith those seen in multiple previous experiments.
Brave et al.
Mol Cancer Ther; 10(5) May 2011 Molecular Cancer Therapeutics866
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

PDGFR-a and PDGF-BB–induced phosphorylation ofPDGFR-b, with mean IC50 values of 20 and 32 nmol/L,respectively. In C6 rat glioma cells, a similar IC50 value of24 nmol/Lwas observed versus PDGF-AA stimulation ofPDGFR-a (Fig. 4C). In NIH 3T3 cells (mouse fibroblastline), cediranib was slightly more potent, inhibitingPDGF-BB–mediated phosphorylation of PDGFR-b withan IC50 value of 12 nmol/L (Fig. 4D). Comparable activitywas found in smoothmuscle cell types. In culturedhumancoronary VSMCs, the primary PDGFR is PDGFR-a (datanot shown). Cediranib inhibited PDGF-AA–stimulatedreceptor phosphorylation with an IC50 value of 15nmol/L (Fig. 4E). In contrast, in human aortic VSMCs,theprimaryPDGFR isPDGFR-b (datanot shown). In thesecells, cediranib inhibits PDGF-BB–induced phosphoryla-tion of PDGFR-bwith an IC50 value of 23 nmol/L (Fig. 4F).To determine how effectively cediranib inhibits the func-
tional consequences of PDGFR activation, its potencywasassessed in both PDGF-AA- and PDGF-BB–driven pro-liferation assays. In humanaorticVSMCs, cediranib inhib-ited PDGF-BB–stimulated proliferation after 48 hourswith an IC50 value of 36 nmol/L (Fig. 4G), similar tothe potency versus PDGFR-bphosphorylation in the samecells. In MG63 cells, cediranib inhibited PDGF-BB–stimu-lated proliferation with an IC50 value of 63.5 nmol/L(Fig. 4H), similar to the previously reported IC50 valueof 40 nmol/L versus PDGF-AA-induced proliferation inthe same cell line (7).
Cediranib gives differential inhibition of PDGFRsignaling in C6 tumors and murine lung tissue inligand-induced acute pharmacodynamic assays
We have previously shown time- and dose-dependentinhibition of VEGFR-2 in murine lung tissue by using a
NCI-H526 A B
C D
-160
-160
-35
-35
pc-Kit
pMAPK
MAPK
c-Kit
MWkDa
SCF + [cediranib] (nmol/L)
Co
ntr
ol 0
0.00
1
0.01 0.1 1 10 100
IC50 (μmol/L) 0.001 ± 0.0003
M07e
IC50 (μmol/L) 0.003 ± 0.0020
-160
-160
-35
-35
pc-Kit
pMAPK
MAPK
c-Kit
MWkDa
SCF + [cediranib] (nmol/L)
Co
ntr
ol 0
0.00
1
0.01 0.1 1 10 100
0 0.1 1 5 10 20 50 100
0.00
0.05
0.10
0.15
0.20
0.25
Res
pons
e (O
D)
[Cediranib] (nmol/L)
NCI-H526 proliferation
IC50 (μmol/L) 0.013 ± 0.005
Control 6 3 1.5 0.75 0
50,000
100,000
150,000
200,000
250,000
300,000
pc-K
it si
gnal
Dose of cediranib (mg/kg)
*** ** ****
Control 6 3 1.5 0.75
Cediranib (mg/kg)MWkDa
pc-Kit(pY719)
c-Kit
160-
160-
NCI-H526 tumor PD
Figure 3. Cediranib inhibits SCF-driven c-Kit phosphorylation and proliferation in tumor cells. To assess phospho c-Kit inhibition NCI-H526 (A) and M07e (B)cells, serum-starved cells were incubated in the presence or absence of cediranib, as indicated, for 90 to 120 minutes and then stimulated with 50 ng/mLSCF for 5 to 10 minutes. Representative Western blots showing the inhibition of phosphorylated c-Kit and phosphorylated MAPK in NCI-H526 cells(A) and M07e cells (B) are shown. All data are representative of at least 3 similar experiments. The mean IC50� SEM observed over a number of experiments isshown. C, to determine the ability of cediranib to inhibit SCF-stimulated proliferation, NCI-H526 cells were stimulated with 50 ng/mL cediranib in thepresence and absence of cediranib for 72 hours and proliferation was assessed by BrdU incorporation. The mean IC50 � SEM observed over a numberof experiments is shown. D, to show that cediranib inhibits phosphorylation of c-Kit in chronically dosed NCI-H526 tumor xenografts, mice bearingNCI-H526 tumors were treated once-daily orally with 6, 3, 1.5, and 0.75 mg/kg/d cediranib and the levels of pc-Kit were determined by ex vivo analysis.c-Kit was immunoprecipitated from tumor lysates by using a total c-Kit antibody. c-Kit and phospho c-Kit were detected by Western blotting andchemiluminescence; a representative Western blot is shown. Phospho c-Kit was quantified, and the mean levels in each group� SEM are shown. **, P < 0.01;***, P < 0.001. PD, pharmacodynamics.
Assessment of Cediranib Selectivity
www.aacrjournals.org Mol Cancer Ther; 10(5) May 2011 867
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

ligand-induced pharmacodynamic assay (12). Thisapproach was taken because the interanimal variabilityin pVEGFR-2 levels was high, making accurate assess-ment of inhibitor dose responses extremely difficult. The
addition of exogenous ligand to stimulate receptor phos-phorylation overcame this issue. Here we used a similarapproach to assess the inhibition of PDGFR activationrelative to VEGFR-2 to gain greater insight into the effects
MWkDa
PDGF-AA + cediranib (nmol/L)
Co
ntr
ol 0
0.15
0.46 1.
44.
1 12 37 100
300
-250
-250PDGFR-α
pPDGFR-α
IC50 (μmol/L) 0.020 ± 0.008
U118MGA B
C D
E F
G H
MWkDa
PDGF-AA + cediranib (nmol/L)
Co
ntr
ol 0
0.15
0.46 1.
44.
1 12 37 100
300
-250
-250PDGFR-α
pPDGFR-α
PDGFR-α
pPDGFR-α
IC50 (μmol/L) 0.024 ± 0.004
C6MWkDa
PDGF-BB + cediranib (nmol/L)
Co
ntr
ol 0
0.15
0.46 1.
44.
1 12 37 100
300
-250
-250PDGFR-β
pPDGFR-β
PDGFR-β
pPDGFR-β
IC50 (μmol/L) 0.012 ± 0.0027
NIH 3T3
MWkDa
PDGF-AA + cediranib (nmol/L)
Co
ntr
ol 0
0.3
0.8
2.5 7 22 66 200
-250
-250
Human coronary VSMC
IC50 (μmol/L) 0.015 ± 0.003MWkDa
PDGF-BB + cediranib (nmol/L)
Co
ntr
ol 0
-250
-250
IC50 (μmol/L) 0.023 ± 0.007
0.3
0.8
2.5 7 22 66 200
Human aortic VSMC
0 0.3
0.8
2.5 7 22 66 200
0
5
10
15
20
Res
pons
e (B
rDu
units
)
[Cediranib] (nmol/L)
Human aortic VSMC (PDGF-BB)
IC50 (μmol/L) 0.032 ± 0.004 IC50 (μmol/L) 0.0635 ± 0.0009
MG63 (PDGF-BB)
0 1.5 4.613.7 41.3 123
3701,111
3,3330
2
4
6
8
10
12
Res
pons
e (O
D x
100)
[Cediranib] (nmol/L)
MWkDa
PDGF-BB + cediranib (nmol/L)
Co
ntr
ol 0
0.15
0.46 1.
44.
1 12 37 100
300
-250
-250PDGFR-β
pPDGFR-β
IC50 (μmol/L) 0.032 ± 0.010
U118MG
Figure 4. Cediranib inhibitsboth PDGF ligand–stimulatedphosphorylation of PDGFR-aand PDGFR-b and ligand-dependent proliferation.To determine the activity ofcediranib against PDGFRs, arange of cell lines U118MG (A andB), C6 (C), NIH 3T3 (D), humancoronary VSMCs (E), and humanaortic VSMCs (F) were stimulatedwith PDGF-AA (50 ng/mL) orPDGF-BB (50 ng/mL) for 5 to 10minutes and then lysed. Inhibitionof pPDGFR-b in human aorticVSMCs (D) and inhibition ofpPDGFR-a in human coronaryVMSCs and C6 cells (E).Representative Western blotsshowing pPDGFR-a andpPDGFR-b, as well as levels oftotal receptors, are shown asindicated. The mean IC50 � SEMobserved over a number ofexperiments is shown. Todetermine the effect of cediranibon PDGF-stimulated proliferation,human aortic VSMCs (G) andMG63 cells (H) were stimulatedwith 50 ng/mL PDGF-BB for 72hours in the presence or absenceof cediranib. The mean IC50 �SEM observed over a number ofexperiments is shown.
Brave et al.
Mol Cancer Ther; 10(5) May 2011 Molecular Cancer Therapeutics868
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

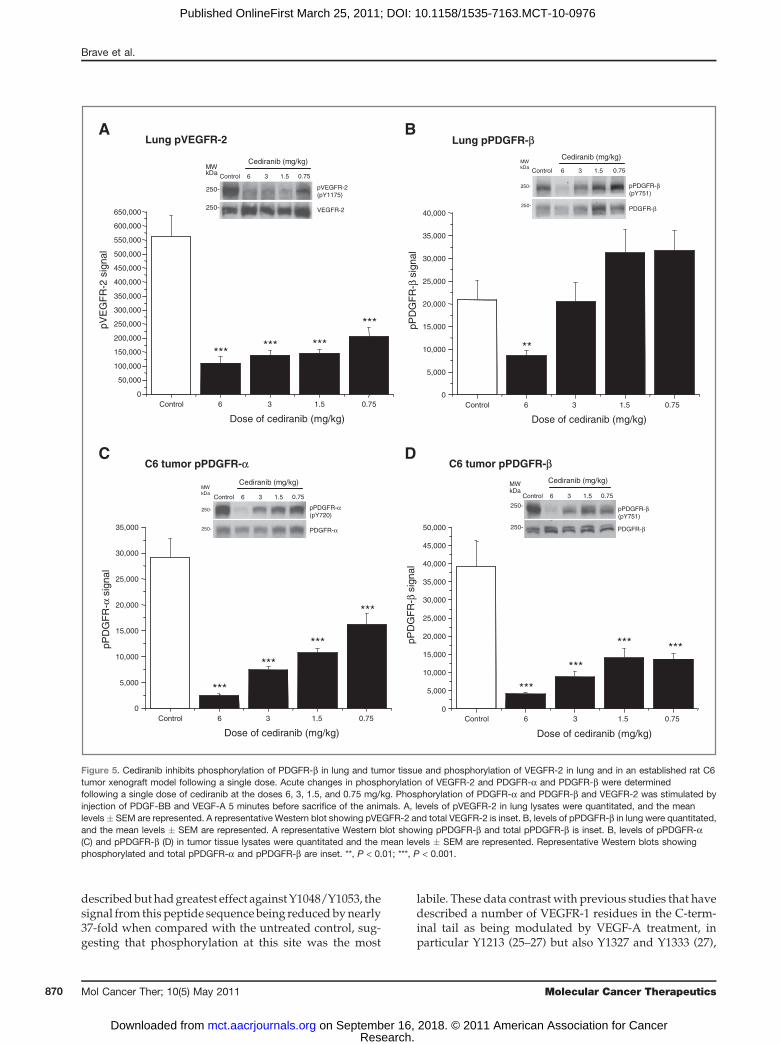
of cediranib on these receptors in vivo. The relativepotency of cediranib versus VEGFR-2 and PDGFR-bwas compared directly in vivo in the same animal. Tonormalize levels of pVEGFR-2 and stimulate PDGFR-aand PDGFR-b phosphorylation, animals were injectedwith both VEGF-A and PDGF-BB immediately beforesacrifice. Lungs from animals bearing C6 tumors receiv-ing cediranib 6, 3, 1.5, or 0.75 mg/kg for 4 hours wereassessed for levels of pVEGFR-2 (Fig. 5A) and pPDGFR-b(Fig. 5B) 4 hours after dosing. This time point was chosen,as we established that the maximal exposure of cediraniboccurs between 2 and 3 hours in mice (data not shown).Consistent with previous data (12), VEGFR-2 phosphor-ylation in lung was significantly reduced over a range ofdoses from 6mg/kg down to 0.75 mg/kg (Fig. 5A). Incontrast, phosphorylation of PDGFR-b was only signifi-cantly inhibited in lung at 6 mg/kg, with no significantinhibition achieved at lower doses (Fig. 5B).To further explore the relative activity against activity
PDGFR-a and PDGFR-b expressed in tumor cells, weexamined C6 tumor xenografts. C6 cells express bothPDGFR-a and PDGFR-b. Although PDGFR-a is consti-tutively phosphorylated in C6 cells in vivo, the injectionof PDGF-BB results in the additional phosphorylation ofPDGFR-b, enabling inhibition of both receptors to bestudied in the same tumor. In separate studies (acute andchronic), we have assessed the activity of cediranibagainst PDGFR-a in C6 tumor xenografts (at 6 hours)and established that cediranib gives up to 60% inhibitionof PDGFR-a phosphorylation. Again because of varia-bility in phosphorylation between tumors, these datawere variable (data not shown). Adopting an acuteligand stimulation approach allowed us to further exam-ine inhibition of both PDGFRs in tumor and lung withinthe individual animals. In contrast to the modulation ofreceptor phosphorylation in lung tissue, phosphoryla-tion of both PDGFR-a (Fig. 5C) and PDGFR-b (Fig. 5D) inC6 tumors, in the same animals, was partially inhibitedby all doses of cediranib examined.
Selectivity against other tyrosine kinase receptorsSomeVEGFR tyrosine kinase inhibitors also have activ-
ity against other kinases. Selectivity of cediranib againstmutFlt-3 and CSF-1R members of the PDGFR family hasbeen shown previously (Wedge and colleagues 2005). Inaddition to determining the relative activity againstVEGFR-1, c-Kit, and PDGFRs, we also tested its activityagainst wild-type Flt-3 and fibroblast growth factorreceptor (FGFR) 1 and FGFR4 (SupplementaryTable S3). The activity against Flt-3 was determined usingOCI-AML-5 cells stimulated with Flt-3 ligand (Flt-3L).The activity against FGFR1 and FGFR4 was determinedby transiently overexpressing the receptor in Cos-1 cells,which resulted in constitutive phosphorylation of thereceptor. Cediranib was inactive against wild-type Flt-3(IC50 value >1 mmol/L) and had marginal activity versusFGFR-1 and -4 (IC50 values of 0.35 and 2.17 mmol/L,respectively). These data indicate that cediranib is sig-
nificantly less active against these receptors than againstthe VEGF receptors, or c-Kit.
Discussion
We have previously shown that cediranib is a potentinhibitor of VEGFR-2 and VEGFR-3 and that it reducesgrowth of a wide range of tumor models by targetingtumor vasculature (7, 8, 12). Cediranib has selectivity forVEGFR-2 against a wide range of kinases, including thePDGFR family members CSF-1R and Flt-3 in cellularphosphorylation assays (420- and >20,000-fold selectivity,respectively, vs. VEGFR-2; ref. 7). Here we explore thepharmacology of cediranib in more depth, examining itsactivity against VEGFR-1 in cells and the PDGFR familymembers c-Kit, PDGFR-a, and PDGFR-b in vitro and invivo within a dose range of 0.75 to 6 mg/kg which hasbeen routinely examined within preclinical tumor xeno-graft experiments.
Establishing the potency of small molecule inhibitorsagainst VEGFR-1 signaling in ligand-induced endothelialcell assays has proven challenging due to the low intrinsickinase activity associated with this receptor. Despite thislow kinase activity, there is some evidence to implicateVEGFR-1 signaling in pathologic angiogenesis (13–15) aswell as in the recruitment of macrophages and myeloidprescursor cell recruitment to tumors (16–18), a processwhich has been linked with resistance to VEGF signalinginhibitors (19–21). The role of the VEGFR-1 kinasedomain in the recruitment of bone marrow–derived cellsinto tumors has been confirmed using VEGFR-1 TK�/�
transgenic mice (22). Consequently, concurrent inhibitionof VEGFR-1 and -2 signaling may afford added thera-peutic benefit. To examine inhibition of VEGFR-1, weused a cell line derived from a human benign angiomainto which the full-length receptor was overexpressed bystable transfection. This cell line does not express VEGFR-2 and therefore enables a more accurate assessment ofactivity against VEGFR-1 by avoiding any confoundersthat could result from VEGFR heterodimerization. Theinhibition of VEGF-induced VEGFR-1 phosphorylationby cediranib, as determined by Western blotting, wasevident at a potency that is comparable with that deter-mined against VEGFR-2 and VEGFR-3 activation in cel-lular assays, thereby confirming that cediranib is a pan-VEGFRkinase inhibitor.Although it is clear that VEGFR-1does induce specific signaling (10, 23, 24), there is limitedinformation on the residues involved. We therefore alsoused an MS-based method to examine the residues acti-vated onVEGFR-1 by VEGF or PlGF in AG1-G1-Flt-1 cellsand inhibition of the VEGF-induced response by cedir-anib. VEGF-A and PlGF were found to induce a broadlysimilar pattern of change in VEGFR-1 increasing thephosphorylation of tyrosine residues Y794, Y1048,Y1053, and Y1242. The greatest fold change evident wasat Y1048 and Y1053, which are within the tyrosine kinasedomain of the receptor. Cediranib treatment abolished allVEGF-stimulated phosphorylation on the 4 residues
Assessment of Cediranib Selectivity
www.aacrjournals.org Mol Cancer Ther; 10(5) May 2011 869
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976
describedbuthadgreatest effect againstY1048/Y1053, thesignal from this peptide sequence being reducedbynearly37-fold when compared with the untreated control, sug-gesting that phosphorylation at this site was the most
labile. These data contrast with previous studies that havedescribed a number of VEGFR-1 residues in the C-term-inal tail as being modulated by VEGF-A treatment, inparticular Y1213 (25–27) but also Y1327 and Y1333 (27),
Lung pVEGFR-2A B
C D
Control 6 3 1.5 0.750
50,000
100,000
150,000
200,000
250,000
300,000
350,000
400,000
450,000
500,000
550,000
600,000
650,000
pVE
GF
R-2
sig
nal
Dose of cediranib (mg/kg)
*** *** ***
***
pVEGFR-2 (pY1175)
VEGFR-2
250-
250-
MWkDa Control 6 3 1.5 0.75
Control 6 3 1.5 0.75 Control 6 3 1.5 0.75
Cediranib (mg/kg)
C6 tumor pPDGFR-α
Control 6 3 1.5 0.750
5,000
10,000
15,000
20,000
25,000
30,000
35,000
pPD
GF
R-α
sig
nal
Dose of cediranib (mg/kg)
***
***
***
***
pPDGFR-α(pY720)
PDGFR-α
250-
250-
MWkDa
Cediranib (mg/kg)
Lung pPDGFR-β
Control 6 3 1.5 0.750
5,000
10,000
15,000
20,000
25,000
30,000
35,000
40,000
pPD
GF
R-β
sig
nal
Dose of cediranib (mg/kg)
**
PDGFR-β
250-
250-
MWkDa
Control 6 3 1.5 0.75
Cediranib (mg/kg)
pPDGFR-β(pY751)
C6 tumor pPDGFR-β
Control 6 3 1.5 0.750
5,000
10,000
15,000
20,000
25,000
30,000
35,000
40,000
45,000
50,000
pPD
GF
R-β
sig
nal
Dose of cediranib (mg/kg)
***
***
*** ***
PDGFR-β
250-
250-
MWkDa
Cediranib (mg/kg)
pPDGFR-β(pY751)
Figure 5. Cediranib inhibits phosphorylation of PDGFR-b in lung and tumor tissue and phosphorylation of VEGFR-2 in lung and in an established rat C6tumor xenograft model following a single dose. Acute changes in phosphorylation of VEGFR-2 and PDGFR-a and PDGFR-b were determinedfollowing a single dose of cediranib at the doses 6, 3, 1.5, and 0.75 mg/kg. Phosphorylation of PDGFR-a and PDGFR-b and VEGFR-2 was stimulated byinjection of PDGF-BB and VEGF-A 5 minutes before sacrifice of the animals. A, levels of pVEGFR-2 in lung lysates were quantitated, and the meanlevels � SEM are represented. A representative Western blot showing pVEGFR-2 and total VEGFR-2 is inset. B, levels of pPDGFR-b in lung were quantitated,and the mean levels � SEM are represented. A representative Western blot showing pPDGFR-b and total pPDGFR-b is inset. B, levels of pPDGFR-a(C) and pPDGFR-b (D) in tumor tissue lysates were quantitated and the mean levels � SEM are represented. Representative Western blots showingphosphorylated and total pPDGFR-a and pPDGFR-b are inset. **, P < 0.01; ***, P < 0.001.
Brave et al.
Mol Cancer Ther; 10(5) May 2011 Molecular Cancer Therapeutics870
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

and Y1309 in response to PlGF stimulation (24). Althoughpeptides indicating phosphorylation at Y1213 weredetected in our study, thesewere notmodulated by ligandactivation. Thesedifferencesmaybe attributable to the cellline examined or technical approach used. We wereunable to develop a pharmacodynamic assay to measureinhibition of VEGFR-1 activity in vivo, due to both the lowlevel of receptor phosphorylation and the inability toidentify selective phosphorylation-specific antibodies toVEGFR-1. However, given the similar potency of cedir-anib against each VEGF receptor in cells, it would bereasonable to assume that it has the ability to inhibitVEGFR-1–driven signaling responses in vivo.In addition to having activity against the VEGF recep-
tors, cediranib also inhibits the kinase activity of c-Kit.Inhibition of wild-type c-Kit signaling in M07e and NCI-H526 cells prevented downstream MAPK phosphoryla-tion. Cediranib inhibited the SCF-induced proliferation ofNCI-H526 cells and reduced an associated increase inAKT phosphorylation (data not shown). However, adecrease in potency (of �10-fold) was observed for inhi-bition of SCF-induced proliferation, suggesting that up to90% of SCF signaling through c-Kit and MAPK needs tobe suppressed to deliver a cytostatic effect in NCI-H526cells. In vivo, inhibition (�85%) of the constitutive phos-phorylation of c-Kit in established NCI-H526 xenografttumors was observed after 17 days of chronically dosingcediranib at 0.75 to 6 mg/kg. This suggests that cediranibmay elicit a pharmacodynamic affect great enough toinfluence the phenotypic consequences of c-Kit signalingin vivo, although an enhanced antitumor effect was notobserved in xenografts derived from this particulartumor line.Mutation or aberrant activation of c-Kit and its ligand
SCF is associated with the progression of numeroussolid and hematologic malignancies, including GIST(28), SCLC (29), and AML (30). Approximately 95% ofGIST cases are positive for c-Kit, with 60% to 70%positive for the c-Kit exon 11 mutations (V560G,V559D, W557R, and Del 557–558) against which the c-Kit/PDGFR/Abl kinase inhibitor imatinib shows activ-ity (31). More recently, secondary mutations of c-Kithave been identified that confer acquired resistance toimatininb (32, 33). Cediranib was found to inhibit phos-phorylation of all of the imatinib sensitive c-Kit mutantforms found in GIST, as well as inhibited 2 of thesecondary point mutations which confer acquired resis-tance to imatinib (V654A and N822K; refs. 34–37). How-ever, cediranib was not active against the T670Igatekeeper mutation in c-Kit (38) or the D816V/D816Yc-Kit mutations (35–37).Previous data generated in cellular phosphorylation
assays showed that cediranib was 10- to 16-fold lessactive against PDGFR-a and PDGFR-b than againstVEGFR, and this margin increased to 100-fold when acomparative assessment of ligand-induced proliferationwas done. The in vitro data generated with cediranib inthis study against receptor phosphorylation in multiple
cell types is consistent with our previous data, with IC50
values in the range of 15 to 32 nmol/L being slightlyhigher than existing data in MG63 cells (cediranib IC50
values of 5 and 8 nmol/L against PDGFR-a and PDGFR-b, respectively). A previously observed drop-off inpotency of approximately 7-fold between phosphoryla-tion and proliferation end points for PDGF-AA/PDGFR-a signaling inMG63 cells was also apparent in the presentstudies when these cells were stimulated with PDGF-BB.The activity of cediranib against PDGF-BB–inducedPDGFR-b phosphorylation and cellular proliferationwas more comparable in the primary VSMCs, suggestingthat PDGFR-mediated signaling responses may be cell-type dependent. In vivo, cediranib inhibited PDGFR-bsignaling in C6 rat tumor xenografts across the dose rangeexamined, in contrast to an effect being evident only at adose of 6 mg/kg in normal lung tissue. This apparentdiscrepancy is unlikely due to species-specificity differ-ences, given the high degree of receptor homologybetween mouse and rat. Although a distribution effectcannot be ruled out, the tissue concentrations of cediranibin C6 tumors did not exceed those in normal lung tissue(data not shown), nor is the inhibition of PDGFR phos-phorylation in C6 tumors due to a bystander effect that issecondary to the antivascular effects of cediranib, becausecompound treatment does not affect the phosphorylationof other receptor tyrosine kinases, such as EGFR in Lovohuman colorectal tumor xenografts, at doses that signifi-cantly inhibit tumor growth (data not shown). An alter-native explanation for the divergent effect observed inlung and C6 tumors could relate to the differential regula-tion or function of PDGFR-b in these tissue compartments,the receptor driving significant cellular proliferation in C6tumors but not in normal lung tissue.
Although cediranib inhibited PDGFR-a and PDGFR-bphosphorylation in C6 tumors, thismodel did not seem tohave increased sensitivity to the antitumor effects of thecompound. A dose of 3mg/kg cediranib, which inhibitedPDGFR-a and PDGFR-b phosphorylation by 73% and76%, respectively, 4 hours after an acute dose, inhibitedtumor growth by 52% (� 4% SEM) after 10 to 14 days ofcontinuous once-daily dosing (data not shown), an effectnot dissimilar to that observed in non–PDGFR-depen-dent tumor models as a consequence of inhibiting VEGFsignaling (5). This finding reinforces the fact that verysignificant inhibition of PDGFR signaling may berequired to prevent phenotypic signaling responses.The activity of cediranib against PDGFR-a andPDGFR-b would therefore not be expected to contributesignificantly to an effect on tumor growth or survival,unless a tumor has a particularly high dependency onsignaling from these receptors.
This work highlights the significant challenge to accu-rately describe the relative activity of anATP-competitiveinhibitor potent against more than 1 kinase. This requiresconsideration of activity at the recombinant kinase level,within multiple cellular phosphorylation and prolifera-tion assays, and then in vivo potency against the pertinent
Assessment of Cediranib Selectivity
www.aacrjournals.org Mol Cancer Ther; 10(5) May 2011 871
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

kinase in target tissues. These data are all required to fullyinterpret observations made in preclinical models. The invivo pharmacodynamic data that show that across thedose range cediranib is primarily a VEGF signaling inhi-bitor with activity against c-Kit. That a significant drop-off in potency is observed between ligand-induced recep-tor phosphorylation and cellular proliferation for c-Kit,PDGFR-a, and PDGFR-b, but not for VEGFR-2, inendothelial cell assays (7), combined with the relativeorder of potency against these targets within a number ofin vitro assays, suggests that cediranib is primarily aVEGFR inhibitor.
Disclosure of Potential Conflict of Interest
S.R. Brave, K. Ratcliffe, Z. Wilson, N.H. James, S. Ashton, A. Wain-wright, J. Kendrew, P. Dudley, N. Broadbent, G. Sproat, S. Taylor, C.Barnes, L. Hennequin, D.J. Ogilvie, J.M. J€urgensmeier, S.R. Wedge, andS.T. Barry are current or former employees and shareholders of
AstraZeneca. J.C. Silva and C.L. Farnsworth are employees of CellSignaling Technology.
Acknowledgments
The authors thank the staff of CDMG, Inc., for xenograft studies, SusanLovick for statistical analysis, Jonathon Orme for FGFR-1 and FGFR-4assays and analyses, Jonathan Dry for mutation analysis, and JamesSherwood and John Smith for c-Kit sequencing and mutation analyses.
Grant Support
The Phosphoscan analysis of VEGFR-1 was done by Cell Signaling Tech-nology and funded by AstraZeneca.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received October 27, 2010; revised February 15, 2011; acceptedFebruary 23, 2011; published OnlineFirst March 25, 2011.
References1. Escudier B, Pluzanska A, Koralewski P, Ravaud A, Bracarda S,
Szczylik C, et al. Bevacizumab plus interferon alfa-2a for treatmentof metastatic renal cell carcinoma: a randomised, double-blind phaseIII trial. Lancet 2007;370:2103–11.
2. Motzer RJ, Hutson TE, Tomczak P, Michaelson MD, Bukowski RM,Oudard S, et al. Overall survival and updated results for sunitinibcompared with interferon alfa in patients with metastatic renal cellcarcinoma. J Clin Oncol 2009;27:3584–90.
3. Hurwitz H, Fehrenbacher L, Novotny W, Cartwright T, Hainsworth J,Heim W, et al. Bevacizumab plus irinotecan, fluorouracil and leucov-orin for metastatic colorectal cancer. N Engl J Med 2004;350:2335–42.
4. Ivy SP, Wick JY, Kaufman BM. An overview of small-molecule inhi-bitors of VEGFR signaling. Nat Rev Clin Oncol 2009;6:569–79.
5. Wedge SR, J€urgensmeier JM. VEGF receptor tyrosine kinase inhibi-tors for the treatment of cancer. In: Marme D, Fusenig N, editors.Tumour Angiogenesis—Basic Mechanisms and Cancer Therapy.Heidelberg, Germany: Springer-Verlag; 2008. p. 395–424. ISBN978-3-540-33176-6.
6. McTigue MA, Wickersham JA, Pinko C, Showalter RE, Parast CV,Tempczyk-Russell A, et al. Crystal structure of the kinase domain ofhuman vascular endothelial growth factor receptor 2: a key enzyme inangiogenesis. Structure 1999;7:319–30.
7. Wedge SR, Kendrew J, Hennequin LF, Valentine PJ, Barry ST, BraveSR, et al. AZD2171: a highly potent, orally bioavailable, vascularendothelial growth factor receptor-2 tyrosine kinase inhibitor for thetreatment of cancer. Cancer Res 2005;65:4389–400.
8. Heckman CA, Holopainen T, Wirzenius M, Keskitalo S, Jeltsch M, Yl€a-Herttuala S, et al. The tyrosine kinase inhibitor cediranib blocks ligand-induced vascular endothelial growth factor receptor-3 activity andlymphangiogenesis. Cancer Res 2008;68:4754–62.
9. Padera TP, Kuo AH, Hoshida T, Liao S, Lobo J, Kozak KR, et al.Differential response of primary tumor versus lymphatic metastasis toVEGFR-2 and VEGFR-3 kinase inhibitors cediranib and vandetanib.Mol Cancer Ther 2008;7:2272–79.
10. Seetharam L, Gotoh N, Maru Y, Neufeld G, Yamaguchi S, Shibuya M.A unique signal transduction from FLT tyrosine kinase, a receptor forvascular endothelial growth factor VEGF. Oncogene 1995;10:135–47.
11. Abrams TJ, Lee LB, Murray LJ, Pryer NK, Cherrington JM. SU11248inhibits KIT and platelet-derived growth factor receptor beta in pre-clinical models of human small cell lung cancer. Mol Cancer Ther2003;2:471–78.
12. Smith NR, James NH, Oakley I, Wainwright A, Copley C, Kendrew J,et al. Acute pharmacodynamic and antivascular effects of the vascular
endothelial growth factor signaling inhibitor AZD2171 in Calu-6human lung tumor xenografts. Mol Cancer Ther 2007;6:2198–208.
13. Carmeliet P, Moons L, Luttun A, Vincenti V, Compernolle V, De Mol M,et al. Synergism between vascular endothelial growth factor andplacental growth factor contributes to angiogenesis and plasmaextravasation in pathological conditions. Nat Med 2001;7:575–83.
14. Fischer C, Jonckx B, Mazzone M, Zacchigna S, Loges S, Pattarini L,et al. Anti-PlGF inhibits growth of VEGF(R)-inhibitor-resistant tumorswithout affecting healthy vessels. Cell 2007;131:463–75.
15. Wu Y, Zhong Z, Huber J, Bassi R, Finnerty B, Corcoran E, et al. Anti-vascular endothelial growth factor receptor-1 antagonist antibody asa therapeutic agent for cancer. Clin Cancer Res 2006;12:6573–84.
16. Luttun A, Tjwa M, Moons L, Wu Y, Angelillo-Scherrer A, Liao F, et al.Revascularization of ischemic tissues by PlGF treatment, and inhibi-tion of tumor angiogenesis, arthritis and atherosclerosis by anti-Flt1.Nat Med 2002;8:831–40.
17. Kaplan RN, Riba RD, Zacharoulis S, Bramley AH, Vincent L, Costa C,et al. VEGFR1-positive haematopoietic bone marrow progenitorsinitiate the pre-metastatic niche. Nature 2005;438:820–7.
18. Murakami M, Zheng Y, Hirashima M, Suda T, Morita Y, Ooehara J,et al. VEGFR1 tyrosine kinase signaling promotes lymphangiogenesisas well as angiogenesis indirectly via macrophage recruitment. Arter-ioscler Thromb Vasc Biol 2008;28:658–64.
19. Shojaei F, Wu X, Malik AK, Zhong C, Baldwin ME, Schanz S, et al.Tumor refractoriness to anti-VEGF treatment is mediated byCD11bþGr1 þmyeloid cells. Nat Biotechnol 2007:25;911–20.
20. Shojaei F, Wu X, Zhong C, Yu L, Liang XH, Yao J, et al. Bv8 regulatesmyeloid-cell-dependent tumour angiogenesis. Nature 2007;450:825–31.
21. Shojaei F, Wu X, Qu X, Kowanetz M, Yu L, Tan M, et al. G-CSF-initiated myeloid cell mobilization and angiogenesis mediate tumorrefractoriness to anti-VEGF therapy in mouse models. Proc Natl AcadSci U S A 2009;106:6742–7.
22. Muramatsu M, Yamamoto S, Osawa T, Shibuya M. Vascular endothe-lial growth factor receptor-1 signaling promotes mobilization ofmacrophage lineage cells from bone marrow and stimulates solidtumor growth. Cancer Res 2010;70:8211–21.
23. Waltenberger J, Claesson-Welsh L, Siegbahn A, Shibuya M, HeldinCH. Different signal transduction properties of KDR and Flt1, tworeceptors for vascular endothelial growth factor. J Biol Chem1994;269:26988–95.
24. Autiero M, Waltenberger J, Communi D, Kranz A, Moons L, Lam-brechts D, et al. Role of PlGF in the intra- and intermolecular cross talkbetween the VEGF receptors Flt1 and Flk1. Nat Med 2003;9:936–43.
Brave et al.
Mol Cancer Ther; 10(5) May 2011 Molecular Cancer Therapeutics872
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

25. Igarashi K, Isohara T, Kato T, Shigeta K, Yamano T, Uno I. Tyrosine1213 of Flt-1 is a major binding site of Nck and SHP-2. BiochemBiophys Res Commun 1998;246:95–9
26. Yu Y, Hulmes JD, Herley MT, Whitney RG, Crabb JW, Sato JD. Directidentification of a major autophosphorylation site on vascularendothelial growth factor receptor Flt-1 that mediates phosphatidy-linositol 3’-kinase binding. Biochem J 2001;358:465–72.
27. Ito N, Wernstedt C, Engstr€om U, Claesson-Welsh L. Identification ofvascular endothelial growth factor receptor-1 tyrosine phosphoryla-tion sites and binding of SH2 domain-containing molecules. J BiolChem 1998;273:23410–8.
28. Rubin BP, Singer S, Tsao C, Duensing A, Lux ML, Ruiz R, et al. KITactivation is a ubiquitous feature of gastrointestinal stromal tumors.Cancer Res 2001;61:8118–21.
29. Camps C, Sirera R, Bremnes RM, Garde J, Safont MJ, Blasco A, et al.Analysis of c-Kit expression in small cell lung cancer: prevalence andprognostic implications. Lung Cancer 2006;52:343–7.
30. Advani AS. C-Kit as a target in the treatment of acute myelogenousleukemia. Curr Hematol Rep 2005;4:51–8.
31. Heinrich MC, Corless CL, Demetri GD, Blanke CD, von Mehren M,Joensuu H, et al. Kinase mutations and imatinib response in patientswith metastatic gastrointestinal stromal tumor. J Clin Oncol 2003;21:4342–9.
32. Wardelmann E, Merkelbach-Bruse S, Pauls K, Thomas N, SchildhausHU, Heinicke T, et al. Polyclonal evolution of multiple secondary KIT
mutations in gastrointestinal stromal tumors under treatment withimatinib mesylate. Clin Cancer Res 2006;12:1743–9.
33. Desai J, Shankar S, Heinrich MC, Fletcher JA, Fletcher CD, Manola J,et al. Clonal evolution of resistance to imatinib in patients withmetastatic gastrointestinal stromal tumors. Clin Cancer Res 2007;13:5398–405.
34. Frost MJ, Ferrao PT, Hughes TP, Ashman LK. JuxtamembranemutantV560GKit is more sensitive to Imatinib (STI571) compared with wild-type c-Kit whereas the kinase domain mutant D816VKit is resistant.Mol Cancer Ther 2002;1:1115–24.
35. Antonescu CR, Besmer P, Guo T, Arkun K, Hom G, Koryotowski B,et al. Acquired resistance to imatinib in gastrointestinal stromal tumoroccurs through secondary gene mutation. Clin Cancer Res 2005;11:4182–90.
36. Shinomura Y, Kinoshita K, Tsutsui S, Hirota S. Pathophysiology,diagnosis, and treatment of gastrointestinal stromal tumors. J Gas-troenterol 2005;40:775–80.
37. Heinrich MC, Corless CL, Blanke CD, Demetri GD, Joensuu H,Roberts PJ, et al. Molecular correlates of imatinib resistance ingastrointestinal stromal tumors. J Clin Oncol 2006;24:4764–74.
38. Tamborini E, Bonadiman L, Negri T, Greco A, Staurengo S, Bidoli P,et al. Detection of over-expressed and phosphorylated wild-type kitreceptor in surgical specimens of small cell lung cancer. Clin CancerRes 2004;10:8214–9.
Assessment of Cediranib Selectivity
www.aacrjournals.org Mol Cancer Ther; 10(5) May 2011 873
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976

2011;10:861-873. Published OnlineFirst March 25, 2011.Mol Cancer Ther Sandra R. Brave, Kirsty Ratcliffe, Zena Wilson, et al. Structurally Related PDGFR FamilyKinase Inhibitor, against VEGFR-1 and Members of the Assessing the Activity of Cediranib, a VEGFR-2/3 Tyrosine
Updated version
10.1158/1535-7163.MCT-10-0976doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2011/03/24/1535-7163.MCT-10-0976.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/10/5/861.full#ref-list-1
This article cites 37 articles, 20 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/10/5/861.full#related-urls
This article has been cited by 5 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://mct.aacrjournals.org/content/10/5/861To request permission to re-use all or part of this article, use this link
Research. on September 16, 2018. © 2011 American Association for Cancermct.aacrjournals.org Downloaded from
Published OnlineFirst March 25, 2011; DOI: 10.1158/1535-7163.MCT-10-0976





























