Embed Size (px)
Citation preview

Aspects of the life cycle of Amblyomma parvum
(Acari: Ixodidae) under natural conditions
Santiago Nava *, Atilio J. Mangold, Alberto A. Guglielmone
Instituto Nacional de Tecnologıa Agropecuaria, Estacion Experimental Agropecuaria Rafaela,
CC 22, CP 2300 Rafaela, Santa Fe, Argentina
Received 6 March 2008; received in revised form 9 May 2008; accepted 22 May 2008
Abstract
Monthly samplings of the rodent Galea musteloides (Caviidae) were carried out from January 2005 to December 2006 in
Quilino, Cordoba Province, Argentina, to determine the seasonal distribution of immature stages of Amblyomma parvum. In the
same locality, cattle and goats were also monthly examined to asses the seasonality of the females of this species. Engorged ticks
collected on their natural hosts were maintained in the laboratory at 25 8C and 83–86% relative humidity to study biological
parameters, and groups of larvae and nymphs were also maintained at three different photoperiods [12 h light (L)–12 h dark (D), 14
L–10 D, and 10 L–14 D]. Larvae were found from late summer to early winter with the peak in autumn, nymphs were collected from
early winter to early spring with the peak in middle winter, and females were detected on cattle and goats during spring and summer,
with the peak in early and middle summer. The pre-moult-period of engorged immature ticks exposed at different photoperiods in
the laboratory show no indication of morphogenetic diapause, being the maximum period 16.4 days for larvae and 21.8 days for
nymphs. The difference in the mean engorgement weight of nymphs moulting to females (6.2 mg � 1.8) and to males
(2.1 mg � 0.1) was statistically significant (P < 0.01), but the difference in the pre-moult period of the nymphs moulting to
females (20.1 days � 2.0) and to males (18 days � 2.0) was not significant (P > 0.01). The average weight of females was
206.1 mg � 23.6, the mean number of eggs laid was 1500.4 � 298.2, the average of pre-oviposition period was 6.5 days � 0.8, and
the minimum incubation period of eggs was 33.1 days � 1.2. There was not significant correlation between the weight of females
and the pre-oviposition period and between weight of immature stages and pre-moult period, but there was significant positive
correlation between weight of females and number of eggs laid. A. parvum has a life cycle under natural conditions with one
generation per year characterized by distinct seasonal peaks of activity for each parasitic stage, where adults are abundant in the
hottest months and the immature stages prevail in the dry season. Apparently, this life cycle is not regulated via diapause. Additional
studies on the life cycle of A. parvum in nature should be performed in other countries to confirm the results obtained in Argentina.
# 2008 Elsevier B.V. All rights reserved.
Keywords: Amblyomma parvum; Seasonality; Life cycle; Argentina
www.elsevier.com/locate/vetpar
Available online at www.sciencedirect.com
Veterinary Parasitology 156 (2008) 270–276
1. Introduction
Amblyomma parvum is a Neotropical tick species
found from southern Mexico to Argentina (Guglielmone
* Corresponding author. Tel.: +54 3492440121;
fax: +54 3492440114.
E-mail address: [email protected] (S. Nava).
0304-4017/$ – see front matter # 2008 Elsevier B.V. All rights reserved.
doi:10.1016/j.vetpar.2008.05.029
et al., 2003; Nava et al., 2008). Although the distribution
of this tick species is wide, there is little knowledge of
ecological aspects such as seasonal distribution of
immature stages and biological information of A. parvum
feeding in nature.
Adult of A. parvum feed on several large and
medium-sized mammals species, including domestic
animals (cattle, dog, cat, donkey, goat, horse and sheep)

S. Nava et al. / Veterinary Parasitology 156 (2008) 270–276 271
and man (Nava et al., 2008). Particularly in Argentina,
the findings of adults of A. parvum on domestic animals
such as cattle and goats are frequent (Guglielmone and
Nava, 2006). In contrast, the records of hosts for larvae
and nymphs of A. parvum are less numerous. There are
occasional records of subadults in Argentina on wild
and domestic artiodactyls, on the white-eared opossum
Didelphis albiventris, on the Caviidae (Dolichotinae)
rodent Dolichotis salinicola, and on domestic and wild
carnivores (Ivancovich and Luciani, 1992). Serra Freire
et al. (1996) found immature stages on the Cervidae
Mazama guazoubira in Brazil and Jones et al. (1972) on
the Lagomorpha Sylvilagus brasiliensis in Venezuela.
Dunn (1923) and Floch and Fauran (1958) mention
larvae and nymphs of A. parvum parasitizing wild
rodents in Panama and French Guiana. Early reports of
host-association for the subadults stages should be
considered carefully, because some records were made
before the description of larvae and nymphs by
Guglielmone et al. (1990a) and often do not specify
the characters used to identify the specimens. Recently,
in a monthly survey of birds and rodents conducted in
north-western of Cordoba Province, Nava et al. (2006a)
stated that the Caviidae rodent Galea musteloides is the
principal host for A. parvum subadults.
Guglielmone et al. (1981, 1990b) described infesta-
tions of adults on cattle in Argentina from late spring to
early autumn, and Szabo et al. (2007) found similar
results in Brazil, with host-seeking adults of A. parvum
in summer and autumn. Also in Argentina, Guglielmone
et al. (1991) studied the life cycle of A. parvum in the
laboratory using a colony started with the offspring of
two engorged females obtained from cattle naturally
parasitized. The results of this study showed that the
mean duration of the life cycle was 99.6 days, and the
incubation period of the eggs was the longest part of the
cycle.
Fig. 1. Monthly rainfall and temperature at the stud
A. parvum may have public health and veterinary
importance. Adults frequently infest domestic animals
and man and have been reported to harbour a novel
Rickettsia sp. strain belonging to the spotted fever group
(Pacheco et al., 2007). Knowledge of the ecology of
A. parvum is necessary to design appropriate control
strategies and to make epidemiological inferences. In
this work we describe the seasonal distribution of all
stages of this tick and provide knowledge of its biology
under field and laboratory conditions.
2. Materials and methods
The field study was carried out in Quilino (308120S,
648320W; altitude 450 m), Cordoba Province Argentina.
The area corresponds to the Western Chaqueno District
of the Chaco Phytogeogrpahic Province according to
the definition given by Cabrera (1994). This district is
characterized by no permanent watercourses, and the
vegetation consists of forests of Aspidosperma queb-
racho blanco, Prosopis nigra, Prosopis pugionata,
Cercidium australe, Zizyphus mistol and Stetsonia
coryne, bushes belonging to the species Larrea
divaricada, Mymozyganthus carinatus, Senna aphylla,
Mimosa detinens, Celtis pallida and Geoffroea deco-
ticans and a herbaceous stratum formed mainly by
perennial grasses. The study area has a climate with a
mean temperature of 24 8C in summer and 12 8C in
winter, and rainfall concentrated on summer (500–
800 mm annually). Fig. 1 includes the monthly rain and
temperature of the study site along the sampling period.
The seasonal distribution of immature stages of A.
parvum was determined by examining G. musteloides
(Caviidae: Caviinae), which were captured in 50
Tomahawk live-trap-type (32 cm in length, 10 cm in
height and 10 cm in width) placed in the field twice a
month. Traps were set at 5 m intervals baited with
y area from January 2005 to December 2006.

S. Nava et al. / Veterinary Parasitology 156 (2008) 270–276272
carrot, and the captured rodents were processed and
examined for ticks following the methodology detailed
in Nava et al. (2006a).
To assess the seasonality of adults ticks, counts were
performed monthly during 2 years on one side of 5 cows
and 5 goats taken at random in the same properties
where rodents were caught. For the statistical analysis
the number of collected ticks on cows and goats was
duplicated. Cattle and goats were not treated with
acaricides through the study period.
The ticks removed from rodents were preserved in
96% ethanol and identified according to Guglielmone
et al. (1990a). The engorged larvae and nymphs
collected on the examined hosts were maintained at
25 8C � 1 and 80–83% relative humidity until moult-
ing, to obtain nymphs and adults to confirm identifica-
tion. Precise determination of immature ticks can be
done with PCR procedures. Therefore, sequences of a
ca. 460 bp fragment of the mitochondrial 16S rDNA
gene were obtained from representative larvae, nymphs
and adults ticks to be compared to each other and with
those of A. parvum from Argentina deposited in
GenBank. DNA extraction and polymerase chain
reaction (PCR) amplification was set-up as described
Table 1
Percentage of prevalence (P) median (M), first and third quartiles (1Q–3Q) of
musteloides between January 2005 and December 2006 in Quilino, Cordob
Month n Larvae
P
January 2005 4 0
February 2005 9 0
March 2005 2 50
April 2005 2 50
May 2005 6 100
June 2005 9 100
July 2005 10 88.8
August 2005 16 0
September 2005 7 0
October 2005 12 0
November 2005 3 0
December 2005 3 0
January 2006 2 0
February 2006 2 0
March 2006 3 33.3
April 2006 7 28.5
May 2006 10 81.8
June 2006 14 86.6
July 2006 8 12.5
August 2006 16 0
September 2006 6 0
October 2006 11 0
November 2006 7 0
December 2006 3 0
The monthly number of hosts examined for ticks (n) is also indicated.
by Mangold et al. (1998a,b). The sequences were edited
using BioEdit Sequence Alignment Editor (Hall, 1999)
and aligned with Clustal W (Thompson et al., 1994). To
the analysis of the sequences the program Mega version
4.0 (Tamura et al., 2007) was used. The adults ticks
were determined following Guglielmone et al. (1990a)
and Guglielmone and Vinabal (1994). The prevalence
(P), median (M), and first and third quartiles (1Q–3Q)
were obtained for each type of infested host, and the site
of attachment of each tick stage was recorded.
Larvae and nymphs were weighed and maintained in
the laboratory at 25 8C and 83–86% relative humidity,
at three daily photoperiods, namely, 12 h light (L)–12 h
dark (D), 14 L–10 D, and 10 L–14 D. Moulting success
and pre-moult period were recorded, and analysis of
variance (ANOVA) was used to test differences among
photoperiodic regimens. A t-test was used to compare
the weight and pre-moult periods of nymphs moulting to
males or females. Due to the fact that most of the
females were observed on cattle and goats in spring and
summer, where the day length is between 13 h and 14 h,
all engorged females were exposed in the laboratory at
14 L–10 D, 25 8C, and 83–86% relative humidity. The
weight, pre-oviposition period, number of eggs laid and
larvae and nymphs of Amblyomma parvum monthly collected on Galea
a Province, Argentina
Nymphs
M (1Q–3Q) P M (1Q–3Q)
0 0 0
0 0 0
1.5 (0–3) 0 0
4.5 (0–9) 0 0
77 (40–92) 0 0
20 (7.5–32.5) 80 3 (0.5–6)
2 (1–4) 100 19.5 (14–25)
0 93.8 21 (9.5–28)
0 100 12 (1–18)
0 0 0
0 0 0
0 0 0
0 0 0
0 0 0
0 (0–5) 0 0
0 (0–10) 0 0
18.5 (10–29) 0 0
2 (1–5) 21.5 0
0 (0–2) 62.5 4 (0–7.5)
0 87.5 3 (1.5–7)
0 50 0.5 (0–2)
0 9.1 0
0 0 0
0 0 0

S. Nava et al. / Veterinary Parasitology 156 (2008) 270–276 273
Table 2
Percentage of prevalence (P) median (M), first and third quartiles (1Q–
3Q) of females of Amblyomma parvum monthly collected on 5 cattle
and 5 goats between January 2005 and December 2006 in Quilino,
Cordoba Province, Argentina
Month Females on cattle Females on goats
P (%) M (1Q–3Q) P (%) M (1Q–3Q)
January 2005 100 12 (10–17) 100 10 (5–20)
February 2005 100 10 (8–19) 100 10 (10–15)
March 2005 100 8 (6–9) 100 6 (4–11)
April 2005 40 0 (0–4) 100 2 (2–5)
May 2005 0 0 0 0
June 2005 0 0 0 0
July 2005 0 0 0 0
August 2005 0 0 0 0
September 2005 0 0 0 0
October 2005 20 0 (0–1) 20 0 (0–1)
November 2005 60 2 (2–4) 100 6 (4–8)
December 2005 100 4 (2–6) 100 6 (4–9)
January 2006 100 8 (8–14) 100 14 (11–20)
February 2006 100 8 (6–14) 100 10 (8–12)
March 2006 100 4 (3–8) 100 8 (4–10)
April 2006 60 2 (0–3) 60 2 (0–4)
May 2006 0 0 0 0
June 2006 0 0 0 0
July 2006 0 0 0 0
August 2006 0 0 0 0
September 2006 0 0 0 0
October 2006 40 0 (0–3) 80 2 (2–2)
November 2006 100 4 (2–7) 100 2 (2–3)
December 2006 100 2 (2–2) 100 6 (4–9)
The monthly number of hosts examined for ticks (n) is also indicated.
minimum incubation periods of the eggs were recorded.
The linear correlation coefficient (r) was used to detect
relationships between the weight of female ticks and the
number of eggs laid and pre-oviposition period, and
between weight of immature and pre-moult period.
Additionally, the deterministic coefficient (r2) for the
significant correlations (P < 0.01) was calculated.
Fig. 2. Seasonal distribution of larvae and nymphs of Amblyomma parvum on
Quilino, Cordoba Province, Argentina, from January 2005 to December 20
3. Results
A total of 829 female (all from cattle and goats), 819
nymphs and 1039 larvae (all from G. musteloides) of A.
parvum were collected. All larvae and 97.5% of nymphs
were attached on the ears of G. musteloides, and more
than 95% of females collected were attached on the
head of cattle and goats, predominantly on ears and
periocular region, where inflammation occurs at the site
of tick attachment.
Divergence in the 16S rDNA sequences among
larvae, nymphs and females of ticks determined as A.
parvum was low, with a maximum difference of 1.1%
and a minimum difference of 0% (GenBank accession
numbers EU306136–EU306138 and EU543570–
EU543572), and the genetic divergence between A.
parvum specimens from Quilino and A. parvum
specimens from other Argentinean localities (GenBank
accession numbers EU306139–EU306156) was also
low, with a maximum difference of 1.1% and a
minimum difference of 0%. These findings support
standard morphological identifications.
Prevalence and M � 1Q–3Q of immature stages
and females of A. parvum collected per month are
shown in Tables 1 and 2, respectively. Analysis of the
monthly distribution of A. parvum demonstrated
distinct peaks of infestation for each of the three
stages (Fig. 2). Larvae were collected from March to
July with a peak of infestation in May in the two study
years; nymphs were found from June to September in
2005 and from June to October in 2006, with a peak of
infestation in August; females were found on cattle
and goats from October to April. Adult infestations
were concentrated principally in spring and summer in
the 2 years of samplings, with a peak in January and
February.
Galea musteloides and females ticks on cattle and goats (combined) in
06.
S. Nava et al. / Veterinary Parasitology 156 (2008) 270–276274
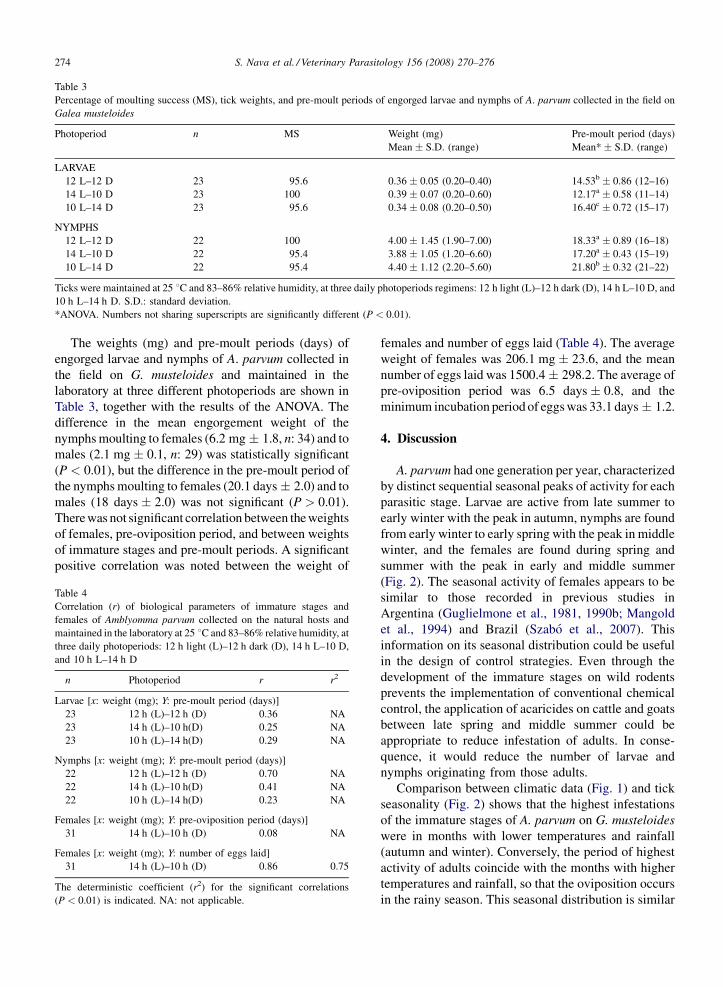
Table 3
Percentage of moulting success (MS), tick weights, and pre-moult periods of engorged larvae and nymphs of A. parvum collected in the field on
Galea musteloides
Photoperiod n MS Weight (mg) Pre-moult period (days)
Mean � S.D. (range) Mean* � S.D. (range)
LARVAE
12 L–12 D 23 95.6 0.36 � 0.05 (0.20–0.40) 14.53b � 0.86 (12–16)
14 L–10 D 23 100 0.39 � 0.07 (0.20–0.60) 12.17a � 0.58 (11–14)
10 L–14 D 23 95.6 0.34 � 0.08 (0.20–0.50) 16.40c � 0.72 (15–17)
NYMPHS
12 L–12 D 22 100 4.00 � 1.45 (1.90–7.00) 18.33a � 0.89 (16–18)
14 L–10 D 22 95.4 3.88 � 1.05 (1.20–6.60) 17.20a � 0.43 (15–19)
10 L–14 D 22 95.4 4.40 � 1.12 (2.20–5.60) 21.80b � 0.32 (21–22)
Ticks were maintained at 25 8C and 83–86% relative humidity, at three daily photoperiods regimens: 12 h light (L)–12 h dark (D), 14 h L–10 D, and
10 h L–14 h D. S.D.: standard deviation.
*ANOVA. Numbers not sharing superscripts are significantly different (P < 0.01).
The weights (mg) and pre-moult periods (days) of
engorged larvae and nymphs of A. parvum collected in
the field on G. musteloides and maintained in the
laboratory at three different photoperiods are shown in
Table 3, together with the results of the ANOVA. The
difference in the mean engorgement weight of the
nymphs moulting to females (6.2 mg � 1.8, n: 34) and to
males (2.1 mg � 0.1, n: 29) was statistically significant
(P < 0.01), but the difference in the pre-moult period of
the nymphs moulting to females (20.1 days � 2.0) and to
males (18 days � 2.0) was not significant (P > 0.01).
There was not significant correlation between the weights
of females, pre-oviposition period, and between weights
of immature stages and pre-moult periods. A significant
positive correlation was noted between the weight of
Table 4
Correlation (r) of biological parameters of immature stages and
females of Amblyomma parvum collected on the natural hosts and
maintained in the laboratory at 25 8C and 83–86% relative humidity, at
three daily photoperiods: 12 h light (L)–12 h dark (D), 14 h L–10 D,
and 10 h L–14 h D
n Photoperiod r r2
Larvae [x: weight (mg); Y: pre-moult period (days)]
23 12 h (L)–12 h (D) 0.36 NA
23 14 h (L)–10 h(D) 0.25 NA
23 10 h (L)–14 h(D) 0.29 NA
Nymphs [x: weight (mg); Y: pre-moult period (days)]
22 12 h (L)–12 h (D) 0.70 NA
22 14 h (L)–10 h(D) 0.41 NA
22 10 h (L)–14 h(D) 0.23 NA
Females [x: weight (mg); Y: pre-oviposition period (days)]
31 14 h (L)–10 h (D) 0.08 NA
Females [x: weight (mg); Y: number of eggs laid]
31 14 h (L)–10 h (D) 0.86 0.75
The deterministic coefficient (r2) for the significant correlations
(P < 0.01) is indicated. NA: not applicable.
females and number of eggs laid (Table 4). The average
weight of females was 206.1 mg � 23.6, and the mean
number of eggs laid was 1500.4� 298.2. The average of
pre-oviposition period was 6.5 days � 0.8, and the
minimum incubation period of eggs was 33.1 days � 1.2.
4. Discussion
A. parvum had one generation per year, characterized
by distinct sequential seasonal peaks of activity for each
parasitic stage. Larvae are active from late summer to
early winter with the peak in autumn, nymphs are found
from early winter to early spring with the peak in middle
winter, and the females are found during spring and
summer with the peak in early and middle summer
(Fig. 2). The seasonal activity of females appears to be
similar to those recorded in previous studies in
Argentina (Guglielmone et al., 1981, 1990b; Mangold
et al., 1994) and Brazil (Szabo et al., 2007). This
information on its seasonal distribution could be useful
in the design of control strategies. Even through the
development of the immature stages on wild rodents
prevents the implementation of conventional chemical
control, the application of acaricides on cattle and goats
between late spring and middle summer could be
appropriate to reduce infestation of adults. In conse-
quence, it would reduce the number of larvae and
nymphs originating from those adults.
Comparison between climatic data (Fig. 1) and tick
seasonality (Fig. 2) shows that the highest infestations
of the immature stages of A. parvum on G. musteloides
were in months with lower temperatures and rainfall
(autumn and winter). Conversely, the period of highest
activity of adults coincide with the months with higher
temperatures and rainfall, so that the oviposition occurs
in the rainy season. This seasonal distribution is similar

S. Nava et al. / Veterinary Parasitology 156 (2008) 270–276 275
to the pattern reported for another Neotropical species,
Amblyomma cajennense (Mangold et al., 1987;
Guglielmone et al., 1990b; Labruna et al., 2002;
Oliveira et al., 2003).
The pre-moult and pre-oviposition periods and the
minimum incubation period of eggs were similar to the
results obtained in the laboratory by Guglielmone et al.
(1991) where ticks were fed on rabbits. In both studies,
the mean incubation period of eggs was the longest
development phase. The engorged ticks exposed at
different photoperiods in the laboratory show no
indication of morphogenetic diapause as defined by
Belozerov (1982). The pre-moult period of larvae and
nymphs showed statistically significant differences
among groups exposed to different light/dark regimens
(Table 3), but these results are probably of little
biological significance, because the maximum differ-
ences observed was 4.2 days for larvae and 4.6 days for
nymphs. As expected for ticks, nymphs moulting to
females were heavier than those moulting to males, and
there was a significant correlation between weight of
females and number of eggs laid.
In Argentina, the life cycle of A. parvum in nature
includes use of wild and domestic animals as hosts of
preimagos and adults, respectively, and the involvement
of G. musteloides as host of larvae and nymphs in the
life cycle of A. parvum also occur in the natural cycle of
Amblyomma tigrinum (Aguirre et al., 2005; Nava et al.,
2006b), a tick species whose distribution is sympatric
with A. parvum in several regions of South America.
Thus, the role of Caviidae rodents as hosts of immature
stages for other Amblyomma species from South
America should be evaluated.
Acknowledgements
We acknowledge to O. Warnke for help in the field
work, and INTA, Fundacion ArgenINTA, Asociacion
Cooperadora INTA Rafaela and CONICET for their
financial assistance.
References
Aguirre, D.H., Mangold, A.J., Cafrune, M.M., Guglielmone, A.A.,
2005. Amblyomma tigrinum (Acari: Ixodidae): new data on hosts
and biology of immature stages and on DNA composition. Vet.
Parasitol. 134, 267–272.
Belozerov, V.N., 1982. Diapause and biological rhythms in ticks. In:
Obenchain, F.D., Galun, R. (Eds.), Physiology of Ticks. Pergamon
Press, Oxford, UK, pp. 469–500.
Cabrera, A.L., 1994. Enciclopedia Argentina de Agricultura y Jardi-
nerıa. Fascıculo 1. Regiones fitogeograficas argentinas. Tomo II.
Editorial ACME, Buenos Aires, Argentina, 85 pp.
Dunn, L.H., 1923. The ticks of Panama, their hosts, and the disease
they transmit. Am. J. Trop. Med. Hyg. 3, 41–104.
Floch, H., Fauran, P., 1958. Ixodides de la Guyane et des Antilles
Francaises. Arch. Inst. Pasteur Guyane 446, 1–94.
Guglielmone, A.A., Vinabal, A.E., 1994. Claves morfologicas dico-
tomicas e informacion ecologica para la identificacion de garra-
patas del genero Amblyomma Koch, 1844 de la Argentina. Rev.
Inv. Agropec. 25, 39–67.
Guglielmone, A.A., Nava, S., 2006. Las garrapatas argentinas del
genero Amblyomma (Acari: Ixodidae): distribucion y hospeda-
dores. Rev. Inv. Agropec. 35, 135–155.
Guglielmone, A.A., Hadani, A., Mangolg, A.J., De Haan, L., Bermu-
dez, A., 1981. Garrapatas (Ixodoidea-Ixodidae) del Ganado
bovino en la provincia de Salta: especies y carga en 5 zonas
ecologicas. Rev. Med. Vet. 62, 194–205.
Guglielmone, A.A., Mangold, A.J., Keirans, J.E., 1990a. Redescrip-
tion of the male and female of Amblyomma parvum Aragao, 1908,
and description of the nymph and larva, and description of all
stages of Amblyomma pseudoparvum sp.n. (Acari: Ixodida: Ixo-
didae). Acarologia 31, 144–159.
Guglielmone, A.A., Mangold, A.J., Aguirre, D.H., Gaido, A.B.,
1990b. Ecological aspects of four species of ticks found on cattle,
in Salta, Northwest Argentina. Vet. Parasitol. 35, 93–101.
Guglielmone, A.A., Mangold, A.J., Garcia, M.D., 1991. The life cycle
of Amblyomma parvum Aragao, 1908 (Acari: Ixodidae) under
laboratory conditions. Exp. Appl. Acarol. 13, 129–136.
Guglielmone, A.A., Estrada Pena, A., Keirans, J.E., Robbins, R.G.,
2003. Ticks (Acari: Ixodida) of the Neotropical Zoogeographic
Region. International Consortium on Ticks and Tick-Borne Dis-
eases (ICTTD-2) Atalanta, Houten, The Netherlands, 173 pp.
Hall, T.A., 1999. BioEdit: a user friendly biological sequence align-
ment editor and analysis program for Windows 95/98/NT. Nucl.
Acids Symp. Ser. 41, 95–98.
Ivancovich, J.C., Luciani, C.A., 1992. Las garrapatas de Argentina.
Monografıa de la Asociacion Argentina de Parasitologıa Veter-
inaria, 95 pp.
Jones, E.K., Clifford, C.M., Keirans, J.E., Kohls, G.M., 1972. The
ticks of Venezuela (Acarina: Ixodoidea) with a key to the species
of Amblyomma in the Western Hemisphere. Brigham Young Univ.
Biol. Ser. Sci. Bull. Biol. Ser. 17, 1–40.
Labruna, M.B., Kasai, N., Ferreira, F., Faccini, J.L.H., Gennari, S.M.,
2002. Seasonal dynamics of ticks (Acari: Ixodidae) on horses in
the state of Sao Paulo, Brazil. Vet. Parasitol. 105, 65–77.
Mangold, A.J., Bermudez, A.C., Kuhne, G.I., Guglielmone, A.A.,
1987. Garrapatas (Ixodoidea-Ixodidae et Argasidae) del ganado
bovino en Santiago del Estero. II. Especies y carga en el noroeste y
sudoeste de la provincia. Rev. Med. Vet. 68, 142–146.
Mangold, A.J., Aguirre, D.H., Gaido, A.B., Guglielmone, A.A.,
1994. Seasonal variation of ticks (Ixodidade) in Bos taurus x
Bos indicus cattle under rotational grazing in forested and
deforested habitats in northwestern Argentina. Vet. Parasitol.
54, 289–395.
Mangold, A.J., Bargues, M.D., Mas-Coma, S., 1998a. 18S rRNA gene
sequence and phylogenetic relationships of European hard-tick
species (Acari: Ixodidae). Parasitol. Res. 84, 31–37.
Mangold, A.J., Bargues, M.D., Mas-Coma, S., 1998b. Mitochondrial
16S rRNA sequences and phylogenetic relationships of Rhipice-
phalus and other tick genera among Metastriata (Acari: Ixodidae).
Parasitol. Res. 84, 478–484.
Nava, S., Mangold, A.J., Guglielmone, A.A., 2006a. The natural hosts
for larvae and nymphs of Amblyomma neumanni and Amblyomma
parvum (Acari: Ixodidae). Exp. Appl. Acarol. 40, 123–131.

S. Nava et al. / Veterinary Parasitology 156 (2008) 270–276276
Nava, S., Mangold, A.J., Guglielmone, A.A., 2006b. The natural hosts
of larvae and nymphs of Amblyomma tigrinum Koch, 1844 (Acari:
Ixodidae). Vet. Parasitol. 140, 124–132.
Nava, S., Szabo, M.P.J., Mangold, A.J., Guglielmone, A.A., 2008.
Distribution, hosts. 16rDNA sequences and phylogenetic position
of the Neotropical tick Amblyomma parvum (Acari: Ixodidae).
Ann. Trop. Med. Parasitol. 102, 409–425.
Oliveira, P.R., Borges, L.M.F., Leite, R.C., Freitas, C.M.V., 2003.
Seasonal dynamics of the Cayenne tick, Amblyomma cajennense
on horses in Brazil. Med. Vet. Entomol. 17, 412–416.
Pacheco, R.C., Moraes-Filho, J., Nava, S., Brandao, P.E., Richtzen-
hain, L.J., Labruna, M.B., 2007. Detection of a novel spotted fever
group rickettsia in Amblyomma parvum ticks (Acari: Ixodidae)
from Argentina. Exp. Appl. Acarol. 43, 63–71.
Serra Freire, N.M., Amorim, M., Gazeta, G.S., Guerim, L., Desiderio,
M.H.G., 1996. Ixodofauna de cervıdeos no Brasil. Rev. Bras.
Cienc. Vet. 3, 51–54.
Szabo, M.P.J., Olegario, M.M.M., Santos, A.L.Q., 2007. Tick fauna
from two locations in the Brazilian savannah. Exp. Appl. Acarol.
43, 73–84.
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: molecular
evolutionary genetics analysis (MEGA) software version 4.0. Mol.
Biol. Evol. 24, 1596–1599.
Thompson, J.D., Higgins, D., Gibson, T.J., 1994. CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalities and weight matrix choice. Nucleic Acids Res. 22,
4673–4680.





























