Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Apr. 1972, p. 583-594Copyright © 1972 American Society for Microbiology
Vol. 5, No. 4Printed in U.S.A.
Antigen-Mediated Fusion of Specifically SensitizedRabbit Alveolar Macrophages
B. GALINDOCenter of Microbiology anid Cell Biology, Instituto Venezolano de Investigacioines Cienitificas, Caracas, Ventezuela
Received for publication 2 December 1971
Rabbits sensitized intravenously with heat-killed Mycobacterium tuberculosis(strain H37Ra) suspended in mineral oil developed a strong pulmonary granulo-matous response which reached its peak about 3 to 4 weeks after injection. Alveolarcells (4 X 106 cells/ml of tissue culture medium 199) procured 6 weeks after sensi-tization showed extensive development of multinucleated giant cells after 12 hr ofincubation in tissue culture flasks containing heat-killed H37Ra (5 ;g/ml). Giantcells measured 80 ,um to 2.5 mm in length and contained between 30 and 700 nuclei.In contrast, no giant cells were observed when similar samples of the same cell pop-ulations were incubated in flasks containing: (i) no mycobacteria; (ii) heat-killedEscherichia coli; (iii) heat-killed Bacillus subtilis; (iv) latex particles; (v) ovalbu-min; or (vi) phytohemagglutinin. The addition of immune (anti-H37Ra) sera
potentiated the phenomenon of giant cell formation. In addition, supernatant fluidsobtained from sensitive alveolar cells incubated with H37Ra were capable of in-ducing giant cell formation when incubated with nonsensitized alveolar cells. Theresults suggest that fusion of alveolar macrophages is mediated by an immunologi-cal mechanism.
Giant cell formation is a characteristic cellularevent that occurs in granulomatous inflammation.It is particularly pronounced in lesions resultingfrom tuberculous infection.The discovery of multinucleated giant cells in
animal tissues dates from more than a centuryago, when J. Muller (quoted in reference 10) ob-served them in tumors. Multinucleated giant cellswere first observed in tuberculous tissue byRokitansky (24) and subsequently described inLanghans' (13, 14) classical papers.
Since the discovery of giant cells, an extensiveliterature has accumulated on this subject. Giantcells have been found in normal (26) as well as inpathological mammalian tissues (1, 5, 18, 26).
Giant cells in cultures of animal tissue werefirst described by Lambert (15) and subsequentlyby numerous investigators (3, 6, 8, 10, 11, 16, 17,22, 23, 25).The formation of multinucleated giant cells may
be the result of two possible mechanisms. First,they may result from repeated amitotic (16) ormitotic (25) nuclear division without cellulardivision of the cytoplasm. Second, giant cells maybe produced by fusion of mononucleated cells (8,12, 15, 20, 25).
It has been found that several viruses can causefusion of mononucleated cells to form multinu-
cleated giant cells in tissue culture. Among theseare: measles virus (6), mumps virus (11), infantilecroup virus (3), a virus of the parainfluenza group(17), parainfluenza HVJ virus (22, 23), and astrain of Sendai virus (10).
I am not aware of any evidence that suggeststhat giant cell formation is mediated by cellularimmune mechanisms. The data to be presentedprovide strong evidence that fusion of alveolarmacrophages can be induced by specific immuno-logic mechanisms which, in turn, depend onfactors associated with cellular immunity.
MATERIALS AND METHODSAnimals. A total of 141 outbred rabbits of either
sex weighing from 2 to 3 kg were used in the presentexperiments. These animals were maintained on astandard laboratory diet.
Sensitization. Ninety-one rabbits were sensitizedintravenously with a single injection of 100 Ag of heat-killed, lyophilized Mycobacterium tuberculosis (strainH37Ra) suspended in 0.1 ml of mineral oil. Fortyanimals maintained under the same conditions wereused as normal controls.
Experimental procedure. Animals were sacrificed atdifferent intervals (from 18 to 97 days) after sensitiza-tion. The alveolar cells were procured by flushing thelungs of rabbits by a technique previously described(21). The viability of the cell populations was esti-mated by the trypan blue exclusion technique (19).
583

INFECT. IMMUNITY
The alveolar cells were washed twice in tissue cul-ture medium (TCM) 199 without serum and recoveredby centrifugation at 400 X g for 15 min. The cellswere then counted and suspended in TCM 199 con-taining no serum at a concentration of 4 X 106 cellsper ml. All media used in the present experimentscontained 100 jlg of streptomycin per ml and 100 unitsof penicillin per ml. Samples (5 ml) of the alveolar cellsuspension containing 5 lAg of heat-killed H37Ra cellsper ml were incubated in a CO2 incubator (95% air,5%6 C02) at 37 C in 30-ml capacity plastic tissue cul-ture flasks (Falcon Plastics, Oxnard, Calif.). Samplescontaining either 1 X 106 or 2 X 106 cells per ml werealso assayed.
Flasks containing equal concentrations of alveolarcells were incubated under the same experimental con-ditions as follows: (i) no mycobacteria; (ii) 10 Mg ofprotein purified derivative (PPD) kindly supplied byMyron Fisher, Park Davis and Co.; (iii) 5 ,ug ofheat-killed Escherichia coli per ml; (iv) phytohemag-glutinin, 10 Mg/ml (Difco Laboratories, Detroit,Mich.); (v) 5 ,ug of heat-killed M. bovis BCG per ml;(vi) 5 Mug of heat-killed Nocardia brasiliensis per ml;or (vii) 5 Ag of heat-killed Bacillus subtilis per ml.
Additional controls included alveolar cells procuredfrom nonsensitized rabbits. All experiments were per-formed at least in triplicate. Results were recorded at2, 4, 6, 12, 18, 24, and 48 hr after incubation.
Continuous microscopy observations were per-formed under a model M Nikon inverted microscopeat 37 C with 95% air and 5% CO2.
Anti-H37Ra serum. Immune sera were obtainedfrom rabbits sensitized intravenously with H37Ra 4 to6 weeks after injection.
Migration inhibition test. The migration inhibitiontest used was similar to that described previously (7).
RESULTSNature of pulmonary granulomatous response
induced by intravenous injection of 100 mg ofkilled H37Ra in 0.1 ml of mineral oil. Rabbitssensitized intravenously with 100 ,ug of killed M.tuberculosis H37Ra developed a strong pulmonarygranulomatous response within 2 weeks aftersensitization. The intensity of the granulomatousresponse was estimated by both the lung weightto body weight ratio (LW/BW) and the volumeof packed alveolar cells obtained through trachealwashings of the lungs with 50 ml of Hanks'balanced salt solution. Figure 1 is a scatter dia-gram showing the variation of the LW/BW ratiowith the time interval. The peak of the granu-lomatous reaction as measured by the LW/BWratio was attained about 25 to 36 days aftersensitization (LW/BW = 0.65 i 0.07 X 10-2).The lungs of these animals showed extensiveconsolidation, and the cell yields always exceeded0.6 ml of packed cells. Many animals yielded 1 to1.5 ml of alveolar cells. After 36 days, the granu-lomatous reaction began to disappear. Between40 to 55 days after injection the lungs were slightlyconsolidated (LW/BW = 0.50 4 0.03 X 10-2)and usually yielded 0.4 to 0.5 ml of alveolar cells.After 55 days the granulomatous reaction was nolonger evident (LW/BW = 0.42 + 0.04 X 10-2),and the lungs yielded less than 0.4 ml of packedcells.The composition of the cell populations varied
with the time interval after sensitization. During
10 20 30 40 50 60
Doys ofter sensitizotion70 80 90 100
FIG. 1. Variation of the lung weight to body weight ratio (LW/BW with the time interval after sensitization.Note that the peak of the reaction is about 30 days.
001r
N-.3
0.005 F
584 GALINDO
0
0 ,
I .. iI ..: .
00 11

IMMUNE MACROPHAGE FUSION
the peak of the reaction, the cell populations werecomposed of 65% macrophages, epithelioid cells,and giant cells; 25% lymphocytes; and 10% poly-morphonuclear leukocytes. Giant cells were rela-tively numerous, comprising up to 1% of themacrophage population. They were round inshape, measured about 60 to 100 ,jm in diameterand contained 10 to 50 nuclei. Between 40 to 55days after sensitization the cell populations werecomposed of 85% macrophages; 10% lympho-cytes; and 5% polymorphonuclear leukocytes.Giant cells were rarely observed.
Antigen-induced giant cell formation of granu-lomatous alveolar cells in vitro. Seventy two of 86intravenously sensitized animals sacrificed 20 to60 days after injection yielded alveolar cells whichdeveloped a strong microscopic clumping reactionafter 12 to 18 hr of incubation with 5 jig ofH37Ra per ml of TCM 199 (Fig. 2 and 3). Theclumps were composed of 80 or more cells andwere distributed throughout the bottom of theculture flask; they were particularly concentratedat the periphery. Although clumps were micro-
scopically prominent after 12 hr of incubation,small clusters of cells were clearly distinguish-able after 3 to 4 hr of incubation. The clumpswere composed mainly of macrophages, but a fewlymphocytes and polymorphonuclear leukocyteswere also present.The most striking feature associated with the
clumping reaction was the extensive developmentof multinucleated giant cells, which in many casescrowded the bottom of the culture flask. In themajority of cases, when nonclumped cells wereremoved, either by treatment with trypsin or byshaking of the culture flask, almost all clumpsdescribed above contained one or more multi-nucleated giant cells (Fig. 4).Giant cells were oval or round, slightly flat-
tened, and characteristically showed numerouscytoplasmic pseudopods. Some of the pseudopodswere long filaments of 1 to 2 jim in diameter, andsome others were short cytoplasmic elongationsseveral micrometers in diameter. Both types ofpseudopods were frequently observed as cyto-plasmic bridges between two giant cells or be-
FIG. 2. Alveolar cells from a rabbit 40 days after intravenous sensitization. (A) Cells are seen after 18 hr ofincubation without H37Ra, forming an homogeneous layer on the bottom of the culture flask. X 1.56. (B) A sampleof the same cell population after 18 hr of incubation with 5 ,ug of heat-killed H37Ra per ml. Note the presence ofnumerous clumps distributed throughout the bottom of the culture flask. X 1.56.
585VOL. 5, 1972

GALINDO
0..
I') i
3
,i 4'X
-) OJ
00
0
J
-0
..
1) Z~ 0 1 0~
40 4)~~~~ 06
g Q# ei
o0 -,40*I
..,O 0 40%Ui )0d.30vV
'30 ..
I-) I w~~~~0 *
0QOO
o c 0
o
0~~~~
0
:t
0
FIG. 3. A higher magniification of Fig. 2. X 75.
tween a giant cell and the suriounding macro-
phages. This kind of arrangement gave a typicalmorphological pattern which is shown in Fig. 5
and 6. Giant cells exhibited 15 to 200 round nucleiconcentrated at the center of the cell and con-taining two to three nucleoli.When the reaction was particularly strong,
multinucleated giant cells were readily visiblewithout any shaking or trypsin treatment. Inthese cases the bottom of the culture flask was
layered by a cell syncytium (Fig. 7). Giant cellsappeared like large flakes of up to 2 mm in di-ameter interconnected to each other by cyto-plasmic bridges. Nuclei were numerous, rangingfrom 100 to 700 per cell, and were distributed inclusters throughout the cytoplasm. Nuclei wereround and contained two to six nucleoli (Fig. 8).The number and size of giant cells decreased ascell concentrations were reduced to 1 X 106 cellsper ml. However, the pattern of distribution ofthe nuclei within the giant cells was not affected.Giant cells adhered strongly to the bottom of
the culture flask. It was very difficult to detachthem by incubation for several hours with eithertrypsin (0.5 mg/ml) or ethylenediaminetetraace-tate (0.02%7,). Mechanical removal of the cellswith a rubber policeman disintegrated most of thegiant cells present in the syncytia, preventing anyaccurate giant cell count. For these reasons theintensity of the phenomenon of giant cell forma-
tion was estimated as follows: +, few giant cell,located at the periphery of the culture flask; + +snumerous round giant cells visible in almost allmicroscopic fields; + + +, numerous round giantcells filling practically the entire surface of theiculture flask; and +++ +, the presence of a
syncytium. The intensity of the phenomenon ofgiant cell formation in the majority of the caseswas classified as a + + + reaction.Animals sacrificed between 18 to 35 days after
sensitization yielded alveolar cells which fre-quently showed giant cell formation in vitro after12 to 16 hr of incubation in media containing nomycobacteria. The number of these cells observedin vitro far exceeded the number of giant cellspresent in the alveolar cell population immedi-ately after collection from the lung. The morpho-logical pattern of arrangement of the giant cellson the bottom of the culture flask was similar tothat induced by H37Ra, but the intensity of thephenomenon was attenuated. The intensityreached a +++ reaction in only 2 out of 18cases. The addition of H37Ra effected a significantincrement in the intensity of the phenomenon ofgiant cell formation. In these cases a + + + +reaction was the rule. During this interval aftersensitization, rabbits showed a strong granu-lomatous response (LW/BW > 0.6 X 10-2), andthe cell yields usually exceeded 0.6 ml of packedcells.
586 INFECT. IMMUNITY
)~0~b0~ ~f:0o *0
6
..w

IMMUNE MACROPHAGE FUSION
FIG. 4. Alveolarcells procuredfrom a rabbit 42 days after intravenous sensitization. (A). Cells were photographed12 hr after incubation without mycobacteria. Note the regularity in size and in distributionz of the cell population.Phase-contrast; X400. (B) A companiont sample of the cell population shown after 12 hr of incutbation with 5 Igof heat-killed H37Ra per ml. Giant cells are clearly visible. The thickness of the cells precludes ani adequate viewof the nuclei. Several attached alveolar cells are seeni ont top of the giant cells. The reactioni was classified as a+++ reaction. Phase-contrast; X400.
VOL. 5, 1972 587

INFECT. IMMUNITY
,,il.S
FiG. 5. An intercellular bridge connecting the cytoplasm of a macrophage with a giant cell. Note microvilliwhich are characteristic of these cells. Formol fixation and hematoxylin-eosin stain; X 780.
FiG. 6. Giant cells forming a syncytium. Multiple connections between giant cells are evident. Nuclei are par-
tially visible. Phase-contrast; X400.
588 GALINDO
.
I
.fex .r By_
.t ...a_._.}} "5+:*_r"
.4F "XX
a
'S r :;,...w.,:a. .*,..,
*a.*.D_/.,.-
v i',- i.,. '
__;ee
.. '., .'...
", i',7 Ue:W .:' i.
At ': :1:l, ; t;
* sz s4;thJ w*U.,2 oi /: .......
.*, dr t. -.S 2
v"rpi.

IMMUNE MACROPHAGE FUSION
FIG. 7. A microscopic field showing numerous giant cells. Nuclei are clearly visible accumulated at the centerof the giant cells. Plhase-contrast; X 200.
FIG. 8. Portions of two giant cells showing intercytoplasmic bridges. Nuclei are round and contain three orfour nucleoli. Phase-contrast; X 780.
VOL. 5, 1972 589

INFECT. IMMUNITY
Continuous microscopy observations of granu-lomatous cells incubated with the mycobacteriaprovided direct evidence that the phenomenon ofgiant cell formation in vitro was due to fusion ofalveolar macrophages. After 4 to 5 hr of incuba-tion, alveolar macrophages began to approacheach other and formed cell clusters composed of
about 5 to 10 cells. After 6 to 8 hr of incubation,cell clumps looked compact and increased in di-ameter by attachment of new macrophages. Dur-ing the following 8 hr, the demarcation betweenmacrophages located on top of the clumps disap-peared progressively, and portions of the under-laying giant cells began to be visible. The process
FIG. 9. Incorporationi of a macrophage i/ito the cytoplasm of a giant cell. The macrophage emits a pseudopodthat comes in contact with the gianit cell cytoplasm (a) anid is progressively incorporated (b-h) in a 6-hr intierval.X200.
590 GALINDO
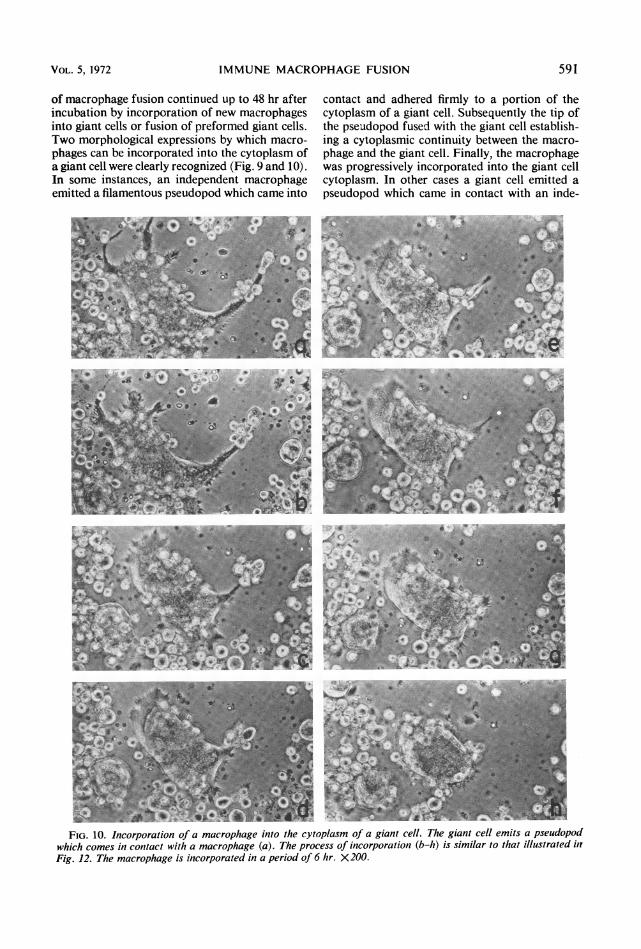
IMMUNE MACROPHAGE FUSION
of macrophage fusion continued up to 48 hr afterincubation by incorporation of new macrophagesinto giant cells or fusion of preformed giant cells.Two morphological expressions by which macro-phages can be incorporated into the cytoplasm ofa giant cell were clearly recognized (Fig. 9 and 10).In some instances, an independent macrophageemitted a filamentous pseudopod which came into
contact and adhered firmly to a portion of thecytoplasm of a giant cell. Subsequently the tip ofthe pseudopod fused with the giant cell establish-ing a cytoplasmic continuity between the macro-phage and the giant cell. Finally, the macrophagewas progressively incorporated into the giant cellcytoplasm. In other cases a giant cell emitted apseudopod which came in contact with an inde-
Y.T
>~Ik,'
,, 4.
,. /.6*,,. .
A
f -.*! W j ;;'i 1 11E
FIG. 10. Incorporation of a macrophage into the cytoplasm of a giant cell. The giant cell emits a pseudopodwhich comes in contact with a macrophage (a). The process of incorporation (b-h) is similar to that illustrated inFig. 12. The macrophage is incorporated in a period of 6 hr. X200.
VOL. 5, 1972 591
ll,,A.0,4-.. 49 4 '...w i - V17";;Id;: -,&'w Ar
* ...'..4

INFECT. IMMUNITY
pendent macrophage, and the process of fusioncontinued in a way similar to that describedabove.
Ziehl-Neelsen stain demonstrated that numer-ous intact acid-fast bacilli were present within thecytoplasm of independent macrophages. Medium-size giant cells contained relatively few bacilli,several of which looked disintegrated. Very fewbacilli were observed within the cytoplasm of largegiant cells present in the syncytia. Electron mi-croscopy revealed that most of the mycobacteriapresent in giant cells were disintegrated withinphagolysosomes.
Alveolar cells procured from intravenously sen-sitized animals were regularly inhibited frommigration in the presence of H37Ra. In a total of10 experiments, it was noted that in addition tothe inhibition of migration, a considerable num-ber of giant cells were present among the inhibitedcells. Giant cells were round, oval, or elongated,and some of them were 2.5 mm in length. De-velopment of giant cells in the migration cham-bers paralleled giant cell formation in cultureflasks. Alveolar cells procured from animals be-tween 18 to 35 days after sensitization frequentlydeveloped a limited number of giant cells in theabsence of H37Ra. The addition of H37Ra pro-duced a significant increment in the number ofgiant cells. Animals sacrificed during this intervalafter sensitization showed a considerable granu-lomatous response (LW/BW ratio = 0.6 x 10-2),and their lungs yielded cells which exhibited thephenomenon of autoinhibition previously de-scribed (7). At longer intervals (40 to 60 days aftersensitization), giant cells in the migration cham-bers were formed only in the presence of H37Ra.
Intravenously sensitized rabbits showing noskin reactivity to PPD usually yielded alveolarcells, which formed giant cells and were inhibitedfrom migration in the presence of H37Ra.
Specificity of the induction of giant cell forma-tion. Specificity was assayed by using sensitizedalveolar cell populations showing no spontaneousgiant cell formation. Consequently, any giant cellformation would be attributed to the effect of thetested agent. The following paragraphs sum-marize the results of 20 experiments.
In each experiment the alveolar cell populationswere divided into identical fractions. One fractionwas tested with 5 ,ug of H37Ra per ml. In all ex-periments the sensitized alveolar cell populationsexhibited extensive development of multinucle-ated giant cells in the presence of the mycobac-teria.No giant cell formation was observed when cell
populations were tested with one of the followingagents: (i) particulate nonrelated antigen such asB. subtilis or E. coli at a concentration of 5 ,ug/ml,
(ii) phagocytosis-stimulating agents which do notrequire opsonization, such as latex particles (1 to5%G), (iii) nonspecific agglutinating agents, suchas phytohemagglutinin agents, (10 ,g/ml), (iv)soluble nonrelated antigen at different concentra-tions, such as ovalbumin (5 to 40 ,ug/ml) andferritin (5 to 15 ,ug/ml).
Results with PPD were variable. In the majorityof the cases PPD induced only a limited formationof giant cells far less prominent than that inducedby H37Ra. In other instances giant cells were notobserved in media containing PPD, while samplesof the same cell population exhibited a conspicu-ous giant cell formation in the presence of H37Ra.The phenomenon of giant cell formation in
vitro could be easily induced when H37Ra sensi-tized alveolar cells were incubated with eitherBCG or N. brasiliensis at a concentration of 5,ug per ml.Enhancement of giant cell formation in the pres-
ence of anti-H37Ra serum. Alveolar cells fromintravenously sensitized rabbits developed bothspontaneous and H37Ra-induced giant cell for-mation in media containing either no serum, 10to 20%7/ fetal bovine serum, or 10 to 20%C normalhomologous serum. The intensity and the mor-phological appearance of the phenomenon weresimilar in all these media.
In five experiments, the addition of 20%C ofanti-H37Ra autologous or homologous serummarkedly increased the intensity of both spon-taneous and H37Ra-induced giant cell formation.Media containing both H37Ra (5 ,ug/ml) and
20% homologous anti-H37Ra serum were capableof inducing a limited giant cell formation innormal alveolar cells. No giant cell formationwas observed when normal alveolar cells wereincubated with media containing either H37Raor anti-H37Ra serum alone.
Antigen-antibody complexes formed with fer-ritin-antiferritin serum (at excess antibody, equiv-alence, or excess antigen) were not able to induceany giant cell formation in either normal orH37Ra-sensitized alveolar cells.Macrophage fusion factor(s) produced by granu-
lomatous cells. Normal and H37Ra-sensitized al-veolar cells at a concentration of 4 x 10f cells perml were incubated separately in 199 media con-taining 5 Mg of H37Ra per ml. Fractions of thesame cell populations were incubated in the samemedia but without H37Ra. After 16 to 18 hr ofincubation the cells were removed by centrifuga-tion, and the supernatant fluids were filteredthrough membrane filters (0.45 ,um, Millipore).Normal alveolar cells at a concentration of 2 x106 cells per ml were incubated in plastic flaskscontaining the respective undiluted supernatantfluids without the inclusion of additional H37Ra.
592 GALINDO

IMMUNE MACROPHAGE FUSION
Five of seven experiments clearly showed thatsupernatant fluids obtained from sensitized cellsincubated with H37Ra induced giant cell forma-tion of normal alveolar cells. The intensity of thephenomenon was classified as a + + to + + +reaction.
In migration chambers, samples of the samesupernatant fluids effected both inhibition of mi-gration of normal cells and extensive giant cellformation.
Supernatant fluids obtained from granuloma-tous cells incubated without H37Ra, but showingspontaneous giant cell formation, were capableof inducing giant cell formation when incubatedwith nonsensitized alveolar cells.
DISCUSSIONIn a previous communication (7), it was pro-
posed that the granulomatous lung reaction in-duced by mycobacteria may be a useful model inanalyzing the mechanisms responsible for severalexpressions of cellular immunity, especially al-lergic granuloma formation. In this regard, themain advantage of the model proposed is that thelung cell populations which can be readily pro-cured are actively participating in a cellular im-mune response.The present studies using this model have re-
vealed that sensitized alveolar cells develop ex-tensive multinucleated giant cell formation whenincubated in vitro with the sensitizing agent(H37Ra). The phenomenon of giant cell forma-tion in vitro was highly reproducible since it wasobserved in almost 90% of the cases.
Since numerous giant cells containing 400 ormore nuclei were seen as early as 12 to 16 hr afterincubation, giant cell formation observed in vitromust be due to fusion of alveolar macrophages.Endomitosis could not account for this number ofnuclei within a 12-hr period of incubation. Fur-thermore, no signs of either amitotic or mitoticactivity were observed.Continuous microscopy observation provided
direct evidence that the phenomenon of giant cellformation is due to fusion of alveolar macro-phages (Fig. 9 and 10). It was found that incor-poration of alveolar macrophages into a giantcell is a continuous process which takes place inapproximately 8 hr. Several macrophages can besimultaneously incorporated into a giant cell.
Nonspecific induction of giant cell formationin vitro with different cell lines has been exten-sively reported (3, 6, 8, 10, 11, 15, 16, 17, 22, 23,25). In the present study, specificity was tested inseveral ways. Either particulate (B. subtilis, E.coli) or soluble (ovalbumin, ferritin) nonrelatedantigens failed to induce giant cell formation ofH37Ra-sensitized alveolar cells. Nonspecific stim-
ulation of phagocytosis, such as that induced bylatex particles, was also ineffective in inducingfusion of alveolar cells. Finally, simple cell agglu-tination, as that caused by phytohemagglutinin,was not enough to induce giant cell formation ofeither normal or sensitized alveolar cells. Accord-ing to these results, the phenomenon of giant cellformation of granulomatous alveolar cells invitro appears to be unique in that it was specifi-cally induced by the sensitizing antigen. There-fore, it seems logical to conclude that this phe-nomenon is mediated by an immunologicalmechanism. The fact that N. brasiliensis wasable to induce giant cell formation of H37Ra-sensitized alveolar cells is not surprising becauseit is well known that Mycobacterium and No-cardia share common antigens.
It is of interest that PPD was far less effectivethan whole cells (H37Ra) in inducing giant cellformation in vitro. This result may imply thatcomponents other than PPD are responsible forthe elicitation of this phenomenon.Anti-H37Ra homologous serum consistently
induced a significant increment in the intensityof giant cell formation in media containing themycobacteria. Either normal homologous serumor unrelated immune serum, such as antiferritinserum did not effect any change in the intensityof the giant cell response. Although an easyexplanation is not readily available, it might bethat antigen-antibody complexes act synergis-tically with the basic fusion mechanism. In thisregard, limited giant ceU formation was observedwhen normal alveolar cells were incubated inmedia containing both H37Ra and anti-H37Raserum.
Antigen-antibody complexes prepared withferritin either in excess antigen, equivalence, orexcess antibody did not cause giant cell forma-tion of normal cells. Nor did they produce anyincrement in the intensity of H37Ra-inducedgiant cell formation.Granulomatous cells procured from animals
sacrificed between 20 to 35 days after sensitiza-tion frequently developed giant cells in mediacontaining no mycobacteria. The intensity of thisspontaneous giant cell formation tended toparallel the intensity of the granulomatousresponse. It was regularly observed when theLW/BW ratio was higher than 0.6 X 10-2. Anexplanation for this phenomenon is not readilyavailable. However, it is possible that alveolarcells contain antigenic fragments remainingfrom the sensitizing injection of H37Ra that maybe responsible for inducing giant cell formationin vitro.
Spontaneous giant cell formation may bearsome relation to the phenomenon of autoinhibi-
VOL. 5, 1972 593

INFECT. IMMUNITY
tion of migration previously described (7). Inthis regard, granulomatous cells exhibiting auto-inhibition invariably developed giant cells withinthe inhibited area of migration.The present experiments clearly indicate that
granulomatous cells obtained from H37Ra-sen-sitized rabbits produced a soluble material(s)when incubated with H37Ra, which is capableof inducing extensive giant cell formation ofnonsensitized alveolar cells. This substance(s)was also produced in vitro in the absence ofH37Ra when granulomatous alveolar cellsshowed spontaneous giant cell formation.
Studies are being conducted in this laboratoryto determine the nature of the macrophage fusionfactor(s) as well as the cell type responsible forits production. The results of this investigationwill be the subject of a future communication.Several experiments already performed in thisconnection indicate that a purified population oflymphocytes, obtained from lymph nodes drain-ing an H37Ra-induced granuloma, produced thefusion factor(s) only in media containing H37Ra.Nonsensitized lymphocytes fail to produce thefusion factor(s) when incubated under the same
conditions. It has also been found that the fusionfactor is inactivated after trypsin treatment butis not altered when incubated at 56 C for1 hr.
Because the chemical nature of the factorresponsible for alveolar cell fusion has not beendetermined, it is not possible to ascertain itsrelationship to other substances produced as a
conisequence of specific lymphocyte stimulation,such as migration inhibitory factor (2, 4), lym-photoxin (9), etc.
Disintegration of the mycobacteria appearedto be more effectively accomplished within thecytoplasm of multinucleated giant cells thanwithin individual macrophages. It is, then, con-
ceivable that the phenomenon of giant cell for-mation, so typical of granuloma formation, may
represent a mechanism of cell cooperation for a
greater antimicrobial capacity.
ACKNOWLEDG MENT
We thank Rafael Pingarr6n for his photographic assistanceand Rober-to Ca,stillo fo;- his technic:tl help.
LIr'SRATURE CITED
1. Alleni, J. M., E. M. Brieger, and R. J. W. Rees. 1965. Electron
micros:opy of the host-cell p.irasite relation in murinielepr-osy. J. Paithol. Bacteriol. 89:30f1-306.
2. Bloom, B. R.. aind B. Bennett. 1966. Mzc-ih:nism of a reaction
int vitro aissociated with del.yed-type hypersetisitivity.Scieisce 153:80 82.
3. Chanock, R. M. 1956. Associatio.s of a new type of cvto-pathogenic iniyxovirus with infantile croup. J. Exp. Med.101:555-575.
4. David, J. R. 1966.Delalyed hypersensitivity int vitro: itsmiiediationby cell-free substanceces for-medlylilphoid cell-antigen interaction. P-oc. Nat. Acad. Sci. U.S.A. 56:72- 77.
5. Dum ont, A., and M. Sheldon. 1965. Chainges in the finestructur-e of m-iacirophages in experiinmentally prodLtCedtuberculous granLtlOml1aS in hamnsters. Lah. Invest. 14:2034-2055.
6. Enders, J. F., aniid T. C. Peebles. 1954. Propag:ttion in tissuecultureof'cytopathogenic agents frotn patietits with mileasles.
Proc. Soc. Exp. Biol. Med. 86:277-286.7. Galindo, B.. anid Q. N. Myr-vik. 1970. Migratory response of
gr-anulomaitouls alveolar cells fromii BCG-sensitized rabbits.J.lImmuncl. 105 :227-237.
8.Go:noglio, P. M., G. Ottino, and D. Cantino. 1971. Experi-iniental study oni development and behaviour- of the multi-
nucleated gianit cells" itn vitro J. RetiCLuloenidothel. Soc.9:397 -408.
9. Granger, G. A., and T. W. Williaiiis. 1968. Lytnphiocytecytotoxicity intiitro: activation and release of a cytotoxicfactor.Na.turileLLondoni2 218:1253-1254.
10. Harris, H., J. F.WW tkins, C. E. Ford, and G.1 Schoefl.1'966. Artificial heterokaryons of' animal cells fromi differ-
ent species. J. Cell Sci. 1:1-30.1.1 Henle, G., F. Deinhardt, and A. Giratdi. 1954. Cytolytic
effect3 of' miumps virus in tissue cultur-es of epithelial cells,p. 299. Iit Cellular biology Ofmf AoVirulS infectiont s( Ciba
Fotindation Symiu-posiuut). Chi ttrchll, London.
l2. Lange, 0. 1875. Ueber die EntstehuLng derblutkrperhaltingenZellen und die Metmrrorphosen des Blutes im Lymiiphsacckdes Frosclhes. Virchows Ar-chh Pathol. An:tt. Physiol. 65:27-35.
13. Langhans, 1868. Ueher Riesenezelleni miiit wanidstiindigenKernen in Tuberkeln und die fibrbse Forti des Tuberkels.
Virchows Arch. Pathol. Anat. Physiol. 42:382-404.14. Latinghatns, T. 1870. Beobachtungen jiber Resorptiota der
Extr-atvast-te Lind Pigtneitbildung in denselben. Vir-chowsAr-ch. Paithol. Anat. Physiol. 49:66 116.
15. La3mbert, R. A. 1912. Thepr'oduction of' foreign body giantcells in*itro.J. Exp. Med. 15:510-515.
16. Lewis,W. H. 1927. The f'crimnation of giaint cells in tissue cul-tures and their- sitinilar-ity to those in tuberculous lesions.Atner. Rev. Tubere.15:616-6'8.
17. Mairston, R. Q. 1958. Cytopathogenic effects of hetinadsorp-tion virus type 1. Proc. Soc. Exp. Biol. Med. 98:853-856.
18. Matthews, J. L., J. H. Martini. and G.J. Raice. 1967. Gianiit-cell centrioles. Science 155:1423- 1424.
19. McLintians, W. R., E. V. Davis, F. L. Glover, and G. W.Ratke. 1957. The subinerged culture of miammailian cells:
the Spinner culture. J. Imtiiuiiol. 79:428-433.20. Metchnikoff E. 1884. Untersuchungen Uiber die intraicellul'dire
Verdaiuung bei wirbellosen Thieren .Arb. Zool. Itist.Utiiv. Wein 5:141 173.
21. Myrvik, Q. N., E. S. Leake, and B. Farliss. 1961. Studies on
pulmonary alveolarl macrophages fr-omii the nor-miail riabbitat techniqlue to proc-ure themii itn a high state of' puI-ity. J.Inmmunol. 86:128 133.
22. Okadai, Y. 1958. The fusiont of Ehlrlich's tUmilOr CAuISed byH.V.J. virus itt iitro. Biken J. 1:103-110.
23. Okaida, Y. 1962. Ats:tlysis cf giant polynuclear cell for-matiotic.used by H.V.J. vi,as front Ehilichs ascites tuIlloIr cells.Exp. Cell Res. 26.98-107.
24. Rokitansky, C. 1855. Lehrbuch der patliologischen Anazi-to:nie, 3rd ed., vol. 1, p. 295. Braumifller, Vienntt.
25. Sutton, J. S., antd L. Weiss. 1966. Transfor-tiiation of' ilmono-cytes in tissue cultuire into mnacirph.tges, epithelioid cells,
aind multinucleated giant cells. An electron n-ricrcscopestudy. J. Cell Biol. 23:303-33l.
26. Virchow, R. 1858. Reizung unid Reizbarkeit. Virchowvs Ar-ch.Pat"lol. Atiat. Physiol. 14:1-31.
594 GALINDO




























