Embed Size (px)
Citation preview

IMUNOLOGIAAnticorpos e Antígenos

INTRODUÇÃO1/2 Os anticorpos (Ac) são proteínas circulantes produzidas
em resposta à exposição a estruturas estranhas conhecidas como antígenos (Ag).
São diversificados e específicos na sua capacidade de reconhecer formas estranhas, e constituem os principais mediadores da imunidade humoral contra todas as classes de micróbios.
Os anticorpos, as moléculas do complexo de histocompatibilidade (MHC) e os receptores de antígenos das células T são as três classes de moléculas usadas no reconhecimento de antígenos da imunidade adquirida.
Desses três, os anticorpos ligam a maior variedade de estruturas antigênicas, mostram a maior habilidade para discriminar entre diversos antígenos, e apresentam a maior força de ligação com os antígenos.

INTRODUÇÃO 2/2 Ac conectados à membrana na superfície dos linfócitos B funcionam
como receptores para Ag. Ac secretados que residem na circulação, tecidos e locais mucosos
conectam Ag, neutralizam toxinas e evitam a entrada e disseminação de patógenos.
O reconhecimento do Ag por anticorpos conectados à membrana em células B naïves específicas ativa esses linfócitos e inicia uma resposta humoral.
A eliminação do Ag geralmente requer a interação do Ac com componentes como as proteínas do complemento e células fagocíticas e eosinófilos.
As funções efetoras mediadas por Ac incluem a neutralização dos m.o. ou de produtos tóxicos; ativação do sistema complemento; opsonização dos patógenos para aumentar a fagocitose; citotoxicidade celular dependente de anticorpo (ADCC) – marcador para células NKs – e hipersensibilidade imediata – ativação dos mastócitos.

CARACTERÍSTICA DA LIGAÇÃO DOS AG PELAS MOLÉCULAS DE RECONHECIMENTO DE AG DO SISTEMA IMUNE

DISTRIBUIÇÃO NATURAL E PRODUÇÃO DE AC Os Ac estão distribuídos pelos líquidos biológicos do corpo e são
encontrados nas superfícies de um número limitado de tipos celulares.
As células B inicialmente expressam uma forma integrante da membrana da molécula de Ac na superfície celular, onde ela atua como receptor a Ag.
Após a exposição a Ag, a maior parte da resposta inicial dos Ac ocorre nos tecidos linfóides, mas plasmócitos produtores de Ac de longa duração podem persistir na medula óssea.
Formas secretadas de Ac estão presentes no plasma, nas secreções mucosas e líquido intersticial dos tecidos. Essas formas geralmente se ligam a superfície de células efetoras como fagócitos mononucleares, NK e mastócitos.
Quando o plasma ou o sangue formam um coágulo, Ac permanecem no soro. O soro que contém uma quantidade detectável de moléculas de Ac que conectam um Ag é chamado de anti-soro.
O estudo dos Ac e suas reações com os Ag é chamado de Sorologia. A concentração de moléculas de Ac no soro específicas para determinado Ag é geralmente estimada pela determinação de quantas diluições seriais podem ser feitas até que não se observe mais nenhuma ligação; diz-se que o soro com alta concentração de moléculas de Ac específicos para um Ag tem um “título alto”.

ESTRUTURA MOLECULAR DOS AC
• Método para imortalizar células secretoras de Ac individuais de um animal imunizado produzindo um hidridoma, e cada qual secretou Ac Monoclonais individuais de especificidade predeterminada.
• A disponibilidade de populações homogêneas de Ac e de plasmócitos permitiu a determinação da sequência completa de aa e a clonagem de moléculas individuais de Ac. A pronta disponibilidade de Ig monoclonais culminou na determinação tridimensional, pela cristalografia por raios X, da estrutura de vários Ac e Ac ligados a Ag.

CARACTERÍSTICAS GERAIS DA ESTRUTURA DOS AC 1/4
As proteínas do soro são tradicionalmente separadas por sua solubilidade em albumina e globulina, e podem, ainda, ser subdivididas pelo processo de eletroforese.
A maioria dos Ac é encontrada no terceiro grupo de globulinas (de migração mais rápida), chamado de gamaglobulinas.
Outro nome comum para Ac é Imunoglobulina (Ig), que se refere à imunidade conferida pela fração de gamaglobulina.
Todas as moléculas de Ac possuem as mesmas características estruturais básicas, mas apresentam uma grande variedade nas regiões que conectam os Ag. Essa variabilidade das regiões de ligação de Ag é responsável pela capacidade dos diversos Ac de conectar um número enorme de Ag estruturalmente distintos.

CARACTERÍSTICAS GERAIS DA ESTRUTURA DOS AC 2/4
Uma molécula de Ac possui uma estrutura básica simétrica composta de duas cadeias leves idênticas e duas cadeias pesadas idênticas.
Tanto as cadeias leves quanto as pesadas contêm uma série de unidades homólogas repetidas, que se dobram independentemente em forma globular chamada de domínio da Ig.
As duas cadeias estão ligadas entre si por uma ponte de dissulfeto.
Todas as moléculas que contêm esse padrão pertencem à chamada superfamília das Igs, e acredita-se que todas as sequências genéticas que codificam os domínios de Ig dessas moléculas evoluíram de um único gene.

CARACTERÍSTICAS GERAIS DA ESTRUTURA DOS AC 3/4
Tanto as cadeias pesadas quanto as leves possuem uma região aminoterminal (N-terminal) variável (V) que participa no reconhecimento dos Ag e de regiões constantes (C) carboxiterminais (C-terminais)
As regiões C das cadeias pesadas possuem as funções efetoras. Nas cadeias pesadas, a região V é composta de um domínio Ig e a região C
é composta de 3 ou 4 domínios Ig. Cada cadeia leve é composta de um domínio Ig na região V e de um
domínio Ig na região C. As regiões variáveis (V) são assim determinadas porque contêm regiões em
que a sequência de aa é variável e distingue os Ac feitos por um clone de uma célula B dos Ac feitos por outros clones.
A região V de uma cadeia pesada (Vh) é justaposta com a região V de uma cadeia leve (Vl) para formar o local de conexão com o Ag.
Os domínios da região C são separados dos locais de conexão com o Ag. Os domínios C da cadeia pesada interagem com outra moléculas efetoras e células do Sistema Imune, participando, assim, como mediadora da maioria das funções biológicas dos Ac.
A região C-terminal das cadeias pesadas ancora os Ac ligados à membrana nas membranas plasmáticas dos linfócitos B

CARACTERÍSTICAS GERAIS DA ESTRUTURA DOS AC 4/4
As moléculas de IgG são clivadas pelas enzimas papaína e pepsina nos locais indicados pelas setas.
A digestão com papaína permite a separação de duas regiões: Fab (fragment antigen binding) que retêm a capacidade de se ligar ao Ag; Fc (fragment crystallizable) que tem uma propensão de se auto-associar e cristalizar em trama, se liga a moléculas ou células efetoras.
Quando se usa a pepsina, a proteólise gera um único fragmento F(ab)2, conector de Ag com dobradiça e os laços dissulfetos intercadeias intactos.
A organização básica da molécula de IgG é comum a todas as moléculas Ig de todos os isótipos e de todas as espécies.
CARACTERÍSTICAS ESTRUTURAIS DAS REGIÕES VARIÁVEIS E SEU RELACIONAMENTO COM A LIGAÇÃO DE AG
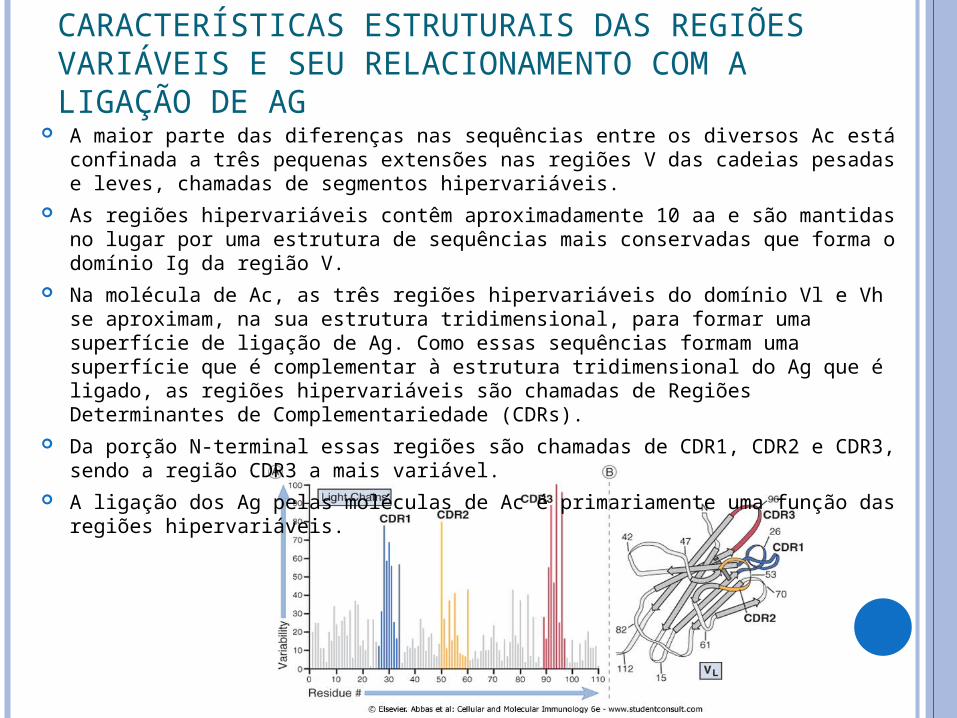
A maior parte das diferenças nas sequências entre os diversos Ac está confinada a três pequenas extensões nas regiões V das cadeias pesadas e leves, chamadas de segmentos hipervariáveis.
As regiões hipervariáveis contêm aproximadamente 10 aa e são mantidas no lugar por uma estrutura de sequências mais conservadas que forma o domínio Ig da região V.
Na molécula de Ac, as três regiões hipervariáveis do domínio Vl e Vh se aproximam, na sua estrutura tridimensional, para formar uma superfície de ligação de Ag. Como essas sequências formam uma superfície que é complementar à estrutura tridimensional do Ag que é ligado, as regiões hipervariáveis são chamadas de Regiões Determinantes de Complementariedade (CDRs).
Da porção N-terminal essas regiões são chamadas de CDR1, CDR2 e CDR3, sendo a região CDR3 a mais variável.
A ligação dos Ag pelas moléculas de Ac é primariamente uma função das regiões hipervariáveis.

CARACTERÍSTICAS ESTRUTURAIS DAS REGIÕES CONSTANTES E SEU RELACIONAMENTO COM AS FUNÇÕES EFETORAS 1/3
As moléculas de Ac podem ser divididas em classes e subclasses distintas com base nas diferenças na estrutura das regiões C das cadeias pesadas.
As classes das moléculas de Ac são chamadas de isótipos e são denominadas IgA, IgD, IgE, IgG e IgM.
As regiões C das cadeias pesadas de todas as moléculas de um isótipo apresentam essencialmente a mesma sequência de aa. Essa sequência é diferente em Ac de outros isótipos. As cadeias pesadas são designadas pela letra do alfabeto grego correspondente ao isótipo do Ac.
Os Ac podem agir como Ag quando introduzidos em outros hospedeiros, estimulando a produção de antianticorpos específicos para uma classe de Ig.
Os diversos isótipos desempenham funções efetoras diferentes. Isso porque a maior parte das funções efetoras é mediada pela ligação das regiões Ch a receptores Fc em várias células e proteínas plasmáticas.

CARACTERÍSTICAS ESTRUTURAIS DAS REGIÕES CONSTANTES E SEU RELACIONAMENTO COM AS FUNÇÕES EFETORAS 2/3

CARACTERÍSTICAS ESTRUTURAIS DAS REGIÕES CONSTANTES E SEU RELACIONAMENTO COM AS FUNÇÕES EFETORAS 3/3
Flexibilidade das moléculas de Ac: os dois locais de ligação de Ag de um monômero de Ig podem se ligar simultaneamente a dois determinantes separados por distâncias variáveis. Essa flexibilidade se deve principalmente á região da dobradiça, localizada entre Ch1 e Ch2 o que permite o movimento independente dos locais de conexão de Ag em relação ao restante da molécula.
Existem duas classes de isótipos de cadeias leves, chamados de k e l, uma molécula de Ac tem duas cadeias leves k ou duas l, mas nunca uma de cada.

FORMAS LIGADAS À MEMBRANAS E SECRETADAS DAS CADEIA PESADAS DE IGS
As formas ligadas à membrana das cadeias pesadas das Igs, mas não as formas secretadas, contêm regiões transmembrana compostas de aa hidrofóbicos e domínos citoplasmáticos que diferem significativamente entre os diferentes isótipos. As formas secretadas dos Ac terminam em cauda hidrofílica.

SÍNTESE, AGREGAÇÃO E EXPRESSÃO DAS MOLÉCULAS DE IG
As cadeias pesadas e leves são sintetizadas no RER. A proteína é translocada para dentro do RER, e as cadeias pesadas de Ig são N-glicosiladas. A dobra apropriada das cadeias pesadas e a sua união às cadeias leves são reguladas por chaperones. Essas proteínas, que incluem as calnexina e BiP (binding protein), se ligam aos polipeptídeos de Ig recém-sintetizados para assegurar que eles sejam retidos ou levados para degradação, a não ser que sejam dobrados de maneira apropriada e unidos para formar moléculas de Ig. Após associação co-valente das cadeias pesadas e leves por pontes de dissulfeto, as moléculas se dirigem as cisternas do Golgi, onde os carboidratos são modificados e os Ac são, então, transportados para membrana plasmática em vesículas ancorando-se ou sendo secretados.
O amadurecimento das células B de progenitores na medula óssea é acompanhada de mudanças específicas na expressão dos genes das Igs, resultando na produção de moléculas de Ig de diversas formas.

CONEXÃO DOS ANTÍGENOS PELOS ANTICORPOS: CARACTERÍSTICAS DOS ANTÍGENOS BIOLÓGICOS 1/2
Um Ag é qualquer substância que pode ser especificamente conectada por um Ac ou receptor da célula T.
Os Ac podem reconhecer praticamente qualquer tipo de molécula biológica, incluindo metabólitos intermediários, açucares, lipídeos, autacóides e hormônios, assim como macromoléculas como carboidratos complexos, fosfolipídeos, ácidos nucléicos e proteínas.
Em contraste, as células T reconhecem principalmente os peptídeos.
As moléculas que estimulam as respostas imunológicas são chamadas de imunógenos. Somente macromoléculas são capazes de estimular os linfócitos B para iniciar respostas imunes humorais, porque a ativação das células B exige a junção (ligação cruzada) de múltiplos receptores a Ag, ou exige Ag protéicos para evocar ajuda das células T.
As macromoléculas são geralmente muito maiores do que a região de ligação de Ag a Ac. Consequentemente, qualquer Ac se liga apenas a uma parte da macromolécula, chamada de determinante ou epitopo. A presença de vários determinantes idênticos é chamada de polivalência ou multivalência. Ag polivalentes podem induzir aglomeração do recptor da célula B e iniciar o processo de ativação dessa célula.

CONEXÃO DOS ANTÍGENOS PELOS ANTICORPOS: CARACTERÍSTICAS DOS ANTÍGENOS BIOLÓGICOS 2/2
A configuração espacial dos diversos epitopos em uma única molécula de proteína pode influenciar na ligação dos Ac de várias formas, causando efeitos alostéricos.
Os determinantes antigênicos podem depender da dobra da proteína, assim como sua estrutura co-valente. Alguns determinantes são acessíveis nas proteínas nativas e se perdem quando elas são desnaturadas, enquanto outros só são expostos quando a proteína se desdobra. Novos determinantes (neodeterminantes) surgem de modificações co-valentes como a quebra de ligações peptídicas.

CONEXÃO DOS ANTÍGENOS PELOS ANTICORPOS: BASES ESTRUTURAIS E QUÍMICAS DA LIGAÇÃO COM OS ANTÍGENOS 1/2
Os locais de ligação de Ag da maioria dos Ac são superfícies planas que podem acomodar epitopos conformativos de macromoléculas, permitindo que os Ac se liguem a essas macromoléculas.
O reconhecimento do Ag pelo Ac envolve uma ligação não co-valente reversível (forças eletrostáticas, pontes de H, forças de van der Waals e interações hidrofóbicas). A importância relativa de cada uma delas depende da estrutura dos locais de ligação;
A força de ligação entre um único local de ligação de Ac e epitopo é chamada de afinidade. A afinidade é normalmente representada por uma constante de dissociação (Kd), que indica a facilidade com a qual se pode separar um complexo Ag-Ac. Uma Kd menor indica uma afinidade mais alta, porque uma concentração menor de Ag e Ac é necessária para formação de complexos.

CONEXÃO DOS ANTÍGENOS PELOS ANTICORPOS: BASES ESTRUTURAIS E QUÍMICAS DA LIGAÇÃO COM OS ANTÍGENOS 2/2
Apesar de a afinidade de qualquer local de conexão de Ag ser a mesma para cada epitopo de um Ag polivalente, a força de ligação do Ac com o Ag deve levar em consideração a ligação de todos os locais a todos os epitopos disponíveis. Essa força de ligação geral é chamada de alvitez e é muito maior do que a afinidade do local de ligação de modo isolado.
O tamanho dos complexos (imunes) Ag-Ac depende das concentrações relativas de Ag e Ac. Grandes complexos são formados em concentrações de Ag-Ac multivalentes denomindas zona de equivalência; os complexos são menores quando há excesso relativo de Ag ou Ac.

RELAÇÃO ENTRE ESTRUTURA E FUNÇÃO DAS MOLÉCULAS DE ANTICORPOS: CARACTERÍSTICAS RELACIONADAS COM O RECONHECIMENTO DE AG 1/2
Todas as características do reconhecimento de Ag refletem as propriedades das regiões V dos Ac.
Especificidade: apresentam especificidade impressionante, distinguindo pequenas diferenças na estrutura química de um Ag. É necessário que haja esse alto grau de especificidade para que os Ac gerados em resposta aos Ag não reajam com moléculas do hospedeiro estruturalmente semelhantes ou com os Ag de outros m.o. (reação cruzada)
Diversidade: a capacidade de Ac em qualquer indivíduo de se ligar a um grande número de diferentes Ag é reflexo da diversidade do Ac, e a coleção total de Ac com especificidades diferentes representa o repertório de Ac. Os mecanismos genéticos que geram esse repertório ocorrem exclusivamente nos linfócitos, são baseados na recombinação aleatória de um número limitado de sequências de DNA, herdadas da linhagem germinativa em genes funcionais que codificam as regiões V.
RELAÇÃO ENTRE ESTRUTURA E FUNÇÃO DAS MOLÉCULAS DE ANTICORPOS: CARACTERÍSTICAS RELACIONADAS COM O RECONHECIMENTO DE AG 2/2
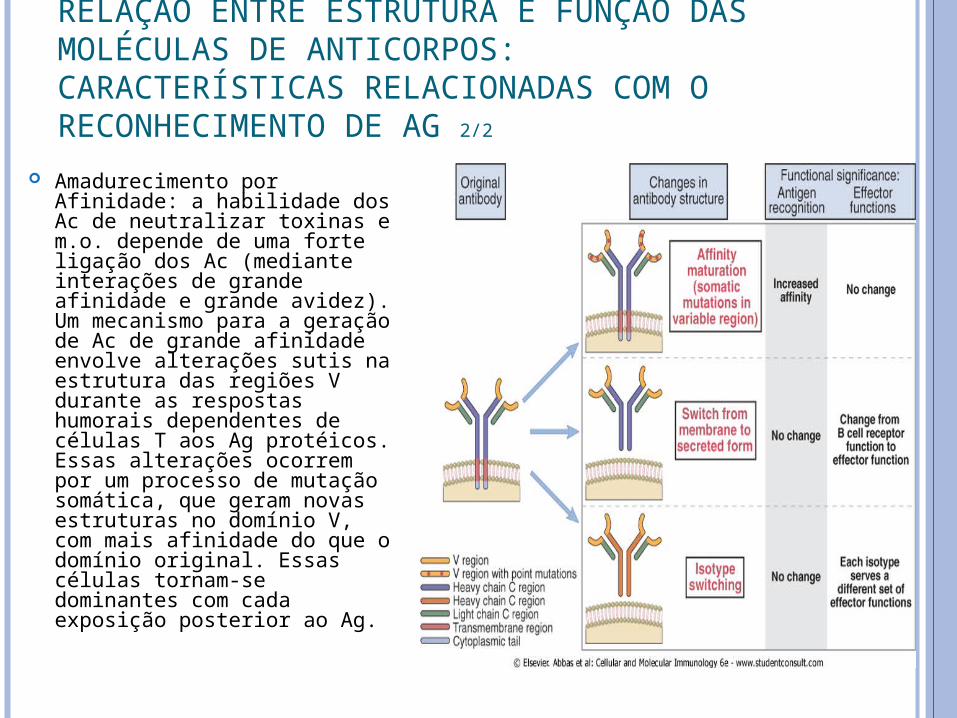
Amadurecimento por Afinidade: a habilidade dos Ac de neutralizar toxinas e m.o. depende de uma forte ligação dos Ac (mediante interações de grande afinidade e grande avidez). Um mecanismo para a geração de Ac de grande afinidade envolve alterações sutis na estrutura das regiões V durante as respostas humorais dependentes de células T aos Ag protéicos. Essas alterações ocorrem por um processo de mutação somática, que geram novas estruturas no domínio V, com mais afinidade do que o domínio original. Essas células tornam-se dominantes com cada exposição posterior ao Ag.

RELAÇÃO ENTRE ESTRUTURA E FUNÇÃO DAS MOLÉCULAS DE ANTICORPOS: CARACTERÍSTICAS RELACIONADAS COM AS FUNÇÕES EFETORAS
Várias funções efetoras das Igs são mediadas pelas porções Fc das moléculas, e os isótipos de Ac que diferem nessas regiões desempenham funções distintas.
As funções efetoras dos Ac só são iniciadas pelos Ac que estão ligados a Ag, e não pela molécula livre. É necessária a presença de pelo menos duas porções Fc adjacentes para conectar e iniciar os vários sistemas efetores, tais como as proteína do complemento e os FcRs dos fagócitos.
Alterações nos isótipos dos Ac durante a resposta humoral influenciam em como e quando as respostas irão reagir para erradicar o Ag.
O processo de recombinação de troca em que ocorre a mudança no tipo da região Ch e, consequentemente, do isótipo do Ac produzido pela célula B não altera as regiões V e a especificidade.
A região C das cadeias pesadas determina a distribuição tecidual das moléculas de Ac.