Embed Size (px)
Citation preview

The Prostate 68:129 ^138 (2008)
Anti-PSMAImmunotoxin asNovel Treatment forProstateCancer?Highand SpecificAntitumor
ActivityonHumanProstateXenograftTumors in SCIDMice
Philipp Wolf,1 Karen Alt,1,2 Patrick Buhler,1,2 Arndt Katzenwadel,1
Ulrich Wetterauer,1 Marlene Tacke,3 and Ursula Elsasser-Beile1*1DepartmentofUrology, Experimental Urology,Universityof Freiburg, Freiburg,Germany
2Facultyof Biology,Universityof Freiburg, Freiburg,Germany3Institute ofMolecularMedicineandCell Research,Universityof Freiburg, Freiburg,Germany
BACKGROUND. Expression of the prostate specific membrane antigen (PSMA) is highlyrestricted to prostate epithelial cells. Therefore, toxin-based immunotherapy against thisantigenmay represent an alternative therapeutic option for prostate cancer. For these purposes,the effects of the recombinant anti-PSMA immunotoxin A5-PE40 on prostate tumor growthwere investigated in vitro and in vivo.METHODS. The in vitro binding and cytotoxicity of A5-PE40 were tested on the PSMA-expressing prostate cancer cell line C4-2 and on the PSMA-negative cell line DU145 by flowcytometry andWST assays. The binding of the immunotoxin to SCIDmouse xenografts and tovariousmouse organswas examined byWestern blot analysis. In vivo, the antitumor activity ofthe immunotoxin was tested by injecting A5-PE40 in mice bearing C4-2 or DU145 xenografts.RESULTS. In vitro, a specific binding of A5-PE40 to C4-2 cells could be shown witha concentration-dependent cytotoxicity (IC50 value¼ 220 pM). In the next step, a specificbinding of the immunotoxin to C4-2 xenografts could be demonstrated. In contrast, no bindingonmouse organs expressing high homologousmouse PSMAwas found. The treatment of micewith C4-2 tumors caused a significant inhibition of tumor growth in vivo, whereas DU145xenografts remained totally unaffected.CONCLUSIONS. A5-PE40 represents a recombinant anti-PSMA immunotoxin with potentantitumor activity in mice bearing human prostate cancer xenograft tumors. Therefore, A5-PE40 could be a promising candidate for therapeutic applications in patients with prostatecancer. Prostate 68: 129–138, 2008. # 2007 Wiley-Liss, Inc.
KEY WORDS: PSMA; prostate cancer; recombinant immunotoxin; immunotherapy;Pseudomonas Exotoxin A
INTRODUCTION
There is a distinct medical need for new treatmentstrategies in patients with prostate cancer withadvanced disease after primary therapies. Over thepast years, novel strategies have been under inves-tigation for the management of prostate cancer includ-ing targeted therapy with prodrugs, immunotherapy,gene therapy, and angiogenesis inhibition [1–3].
P. Wolf and K. Alt equally contributed o this work.
Grant sponsor: Deutsche Krebshilfe, Bonn, Germany.
*Correspondence to: Ursula Elsasser-Beile, MD, PhD, Department ofUrology, Experimental Research Group, University of Freiburg,Breisacher Str. 117, D-79106 Freiburg, Germany.E-mail: [email protected] 16 July 2007; Accepted 21 September 2007DOI 10.1002/pros.20684Published online 28 November 2007 in Wiley InterScience(www.interscience.wiley.com).
� 2007Wiley-Liss, Inc.

Recent advances in expression profiling have iden-tified a number of candidate markers for prostatecancer [2,4]. One of which was the prostate specificmembrane antigen (PSMA), which has been proven tobe an excellent target because it is (a) primarilyexpressed in the prostate, (b) abundantly expressed asprotein at all stages, (c) upregulated in androgen-insensitive or metastatic disease, (d) presented at thecell surface but not released into the circulation, and (e)internalized after antibody binding by receptor-medi-ated endocytosis [5]. Moreover, PSMA shows aconsistent and restricted expression in the neovascula-ture of most other solid tumors but not in normalvasculature [6–8]. In this respect, PSMAmay also serveas a target for specific antiangiogenetic drugs.
The PSMA protein is expressed as a homodimerwith a compact three-dimensional structure on the cellsurface of prostate epithelial cells and exhibits anextensive extracellular domain of 707 aa [9]. Antibodiesor antibody fragments targeting this specific extrac-ellular structure can bind to and can be internalized inPSMA-expressing cells, which is a precondition for thedelivery of cytotoxic agents that act intracellularly[10,11].
The binding domains of recombinant immunotoxinsare mostly single-chain antibody fragments (scFv),representing the smallest functional component of anantibody. A scFv consists of two variable domains, oneof the heavy (VH) and one of the light chain (VL) ofthe parental antibody connected by a flexible peptidelinker. Themajor advantage of an scFv is its smaller sizecompared to the full antibody. This results in a bettertumor penetration, an improved tumor distribution,and a faster blood clearance [12,13].
The truncated formofPseudomonasExotoxinA, PE40(aa 252–613) is a qualified toxin domain for construct-ing immunotoxins [14]. It lacks the natural bindingdomain Ia, and only consists of domains II, Ib and III.Domain II serves as a translocation domain andincludes an amino acid motiv (aa 274–280), which canbe cleaved intracellularly by the enzyme furin. Thisproteolytic activation is essential for the enzymaticactivity of themolecule. The domains Ib and III, exhibita mono-(ADP-Ribosyl-)transferase activity, mimic aribosome moiety and thereby achieve universal recog-nition of eukaryotic elongation factor-2 (eEF-2) [15].This leads to a ribosylation of eEF-2 resulting in aninhibition of protein synthesis and finally in apoptosis.
Recently we could show that the recombinantimmunotoxin A5-PE40, consisting of the anti-PSMA-scFv A5 as binding domain and PE40 as toxin domain,specifically binds and effectively kills PSMA-positiveLNCaP prostate cancer cells in vitro [11]. In the presentstudy we describe the specific and potent antitumoractivity of A5-PE40 in vivo in SCID-mice bearing
xenograft tumors from a highly aggressive and andro-gen-independent growing PSMA-expressing prostatecancer cell-line.
MATERIALSANDMETHODS
Cell Lines
The PSMA-positive, androgen-independent pros-tate cancer LNCaP subline C4-2 and the PSMA-negative prostate cancer cell line DU145 were obtainedfromATCC (Manassas, VA). Cells were propagated byserial passage in RPMI 1640 medium, supplementedwith penicillin (100 U/ml), streptomycin (100 mg/l),and 10% FCS (Biochrom, Berlin, Germany) at 378C in ahumidified atmosphere of 5% CO2.
Anti-PSMAscFvA5 and RecombinantImmunotoxinA5-PE40
The generation of the anti-PSMA scFv A5 and theconstruction of the recombinant anti-PSMA-immuno-toxinA5-PE40 havebeendescribedpreviously [11,16]. Inbrief, scFv A5 was obtained from the monoclonal anti-PSMA antibody 3/A12 by phage display technology.The immunotoxin A5-PE40 was constructed by fusing agene encoding PE40 (bp 252–613) from PseudomonasExotoxin A to a C-terminal position of the scFv A5including a human c-myc-tag for detection and a His6-tag for purification. The immunotoxin was expressed inE. coli, purifiedbyaffinity chromatography, anddialyzedagainst PBS. About 100 mg of the purified protein weregained routinely from 1 L bacterial culture.
RNAIsolation and RT-PCRAnalysis
From freshly removed tissues (tumors, kidneys,brains, livers) 20–25 mg were given into 700 ml guani-dine-thiocyanate lysisbuffer (Qiagen,Hilden,Germany).Samples were quick-frozen in nitrogen and homogen-ized after thawing. Total RNA was isolated withsilicagel-based membranes (Rneasy, Qiagen) accordingto the manufacturer’s protocol. Synthesis of cDNA wasperformed at 428C for 60 min in a final volume of 100 mlcontaining 50 ml of denatured RNA, 20 ml 5� buffer(Promega, Heidelberg, Germany), 5 ml of a dNTP-Mix at10 mM (Qiagen), 1.5 ml RNAsin (40 units/ml, Promega),2.5 ml of 150 pM random hexamer primers, and 2.5 ml ofAMV reverse transcriptase (10 units/ml, Promega).
For PCR the cDNA samples were diluted 1:10 and2 ml were combined with the PCR mixture, containing17 ml water, 2.5 ml 10� buffer (Qiagen), 2.5 ml of 2 mMdNTP, 1 ml of 25 pM sense- and antisense-primer and0.1 ml (1 U) Taq polymerase (Qiagen). The reactionmixture was amplified with a thermal cycler for30 cycles. PCR products were separated on 1% agarosegels and detected by ethidiumbromide staining.
The Prostate DOI 10.1002/pros
130 Wolf et al.

The followingoligonucleotide 50 and 30 primerswereused: for human PSMA 50-ATGTGGAATCTCCTT-CAC-30 and 50-CTCTGCAATTCCACGCCTAT-30 (508C);formurine PSMA 50-TTTAACCGGAACCTTCCTCA-30
and 50-GGGTGGCTGTTCAGATAATG-30 (548C); forhuman GAPDH 50-TGGGGAAGGTGAAGGTCGG-30
and 50-TTTTCTAGACGGCAGGTCAG-30 (628C); formurine GAPDH 50-TTTGCAGTGGCAAAGTGGA-GATTG-30 and 50-ATTGGGGGTAGGAACACGGA-30
(548C).
Western Blotting
For the generation of tissue lysates 20–25 mg oftissues were quick-frozen in nitrogen. After thawing700 ml of the lysis buffer containing 50 mM Tris–HCl,pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% NP-40 and acocktail of protease inhibitors (Roche Diagnostics,Mannheim, Germany) were added and samples werehomogenized. After centrifugation the protein contentof the supernatant was determined by Micro BCAProtein Reagent Kit (Pierce Biotechnology, Rockford,IL). Then 50 mg protein per lane were separated bySDS–page and electrotransferred onto nitrocellulosemembranes (Invitrogen, Karlsruhe, Germany). Blotswere blocked with 5% non-fat dry milk in 0.5% PBSTween-20 for 0.5 hr at RT and incubated for 1 hr at RTwith immunotoxin A5-PE40 (10 mg/ml) or with themAb 3/A12 (20 mg/ml). For development a HRPconjugated mouse anti-human-c-myc-mAb (RocheDiagnostics) or a rabbit-anti-mouse-Ig-HRP (Dako,Hamburg, Germany) were added.
A monoclonal mouse anti-(mouse/hu) b-actin anti-body (Biozol Diagnostica GmbH, Eching, Germany)was used as standard for protein loading. Blots weredeveloped with 3,30-diaminobenzidine as substrate.
FlowCytometry
The binding of the immunotoxin A5-PE40 and of thescFv A5 was tested by flow cytometry as describedpreviously [16]. In brief, C4-2 or DU145 cells wereincubated with scFv or immunotoxin at differentconcentrations between 0.5 and 500 nM. Staining wasdone with a mouse anti-human c-myc mAb (RocheDiagnostics) and goat anti-mouse Ig-RPE (BectonDickinson, Mountain View, CA). Cells were resus-pended in PBS containing propidium iodide andrelative fluorescence of stained cells was measuredusing a FACScan1 flow cytometer and the CellQuest1
Software (Becton Dickinson). Mean fluorescence inten-sity values (MFI) were determined after subtraction ofthe background staining with mouse anti-humanc-myc mAb and RPE-labeled goat antimouse Ig alone.
InVitroCytotoxicity
The in vitro cytotoxicity of A5-PE40 was measuredby using the Cell Proliferation Reagent WST-1 (RocheDiagnostics). C4-2 or DU145 cells were seeded at1.5� 104 cells/well in a 96-well plate and grown for24 hr until a subconfluent cell layer was formed. Thendilutions of the immunotoxin A5-PE40 in aliquots of50 ml/well were added. In a second experiment theimmunotoxin was added together with 25% of SCIDmouse serum.After incubation for 48 hr at 378Cand 5%CO2 the cultures were pulsed with 15 ml/well WST-1reaction solution and incubated for another 90 min.After this the spectrophotometrical absorbances of thesamples were measured at 450 nm (ref. 690 nm). TheIC50-value was obtained from dose-dependent cellviability curves.
InVivoTestingof A5-PE40
Male SCID mice (5–6 weeks old, 20–25 g) wereobtained from Charles River Laboratories (Sulzfeld,Germany) and kept under standardized sterile envi-ronmental conditions. All experiments were per-formed according to the German animal protectionlaw with permission from the local authorities.
To test the tolerability of the immunotoxin, varioussingle doses between 1 mg (14 nM) and 20mg (280 nM) ofA5-PE40 diluted in 500 ml PBS (pH 7.4) were injectedinto the tail vein.
For tumor xenografting, 2� 106 C4-2 or DU145 cellsin 100 ml PBSwere admixedwith 100 ml of reconstitutedbasement membrane (Matrigel, Collaborative Biomed-ical Products, Chicago, IL) at 48C and administeredsubcutaneously into the right flank of each animal.Growing tumors were palpated, diameters weremeasured by a caliper rule and recorded three timesa week. Tumor volumes were calculated according tothe formula V¼ 4/3 p r3.
At the end of the experiment orwhen the tumors hadreached more than 15 mm diameter animals wereasphyxiated by CO2.
Statistical comparison of tumor volumes within thedifferent treatment or control groups was determinedwith the Student’s t-test. P-values of P� 0.05 wereconsidered as significant, P� 0.01 as highly significantP� 0.001 as extremely significant.
RESULTS
Bindingof the ImmunotoxinA5-PE40toC4-2 Target Cells
The concentrations for 50% saturation of PSMA siteson C4-2 cells were determined by flow cytometry as31 nM for A5-PE40 and 33 nM for scFv A5 (Fig. 1a). At
The Prostate DOI 10.1002/pros
ImmunotoxinAgainst Prostate Cancer 131
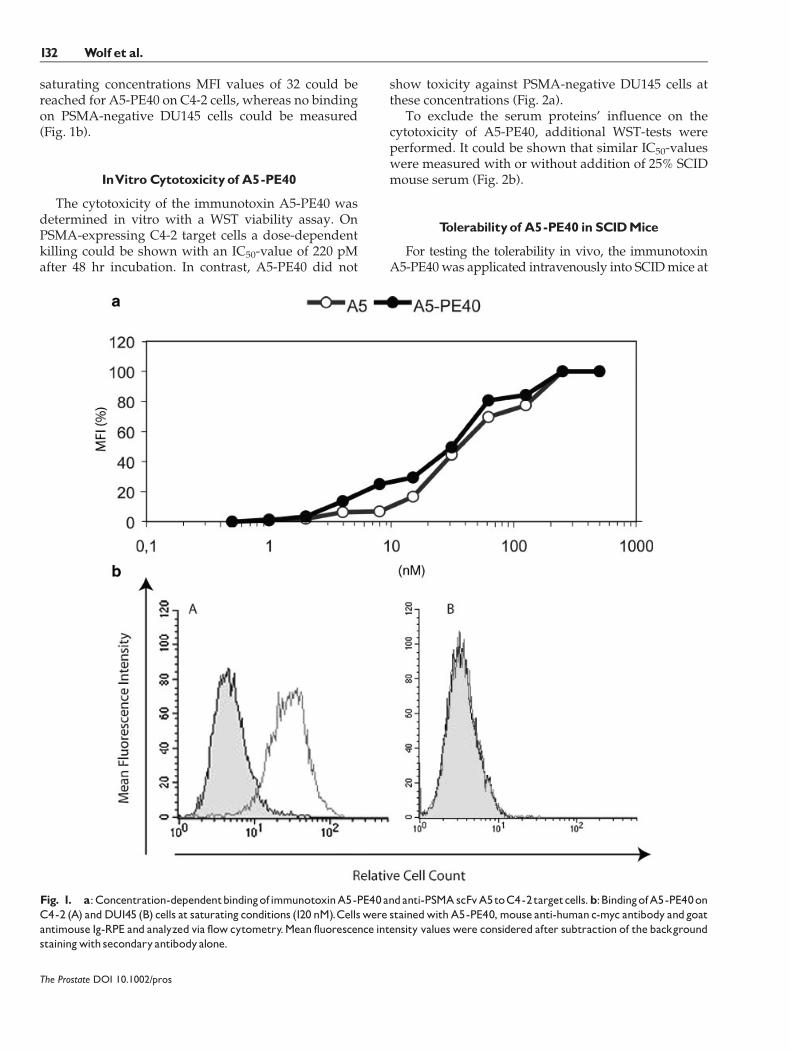
saturating concentrations MFI values of 32 could bereached for A5-PE40 on C4-2 cells, whereas no bindingon PSMA-negative DU145 cells could be measured(Fig. 1b).
InVitroCytotoxicityof A5-PE40
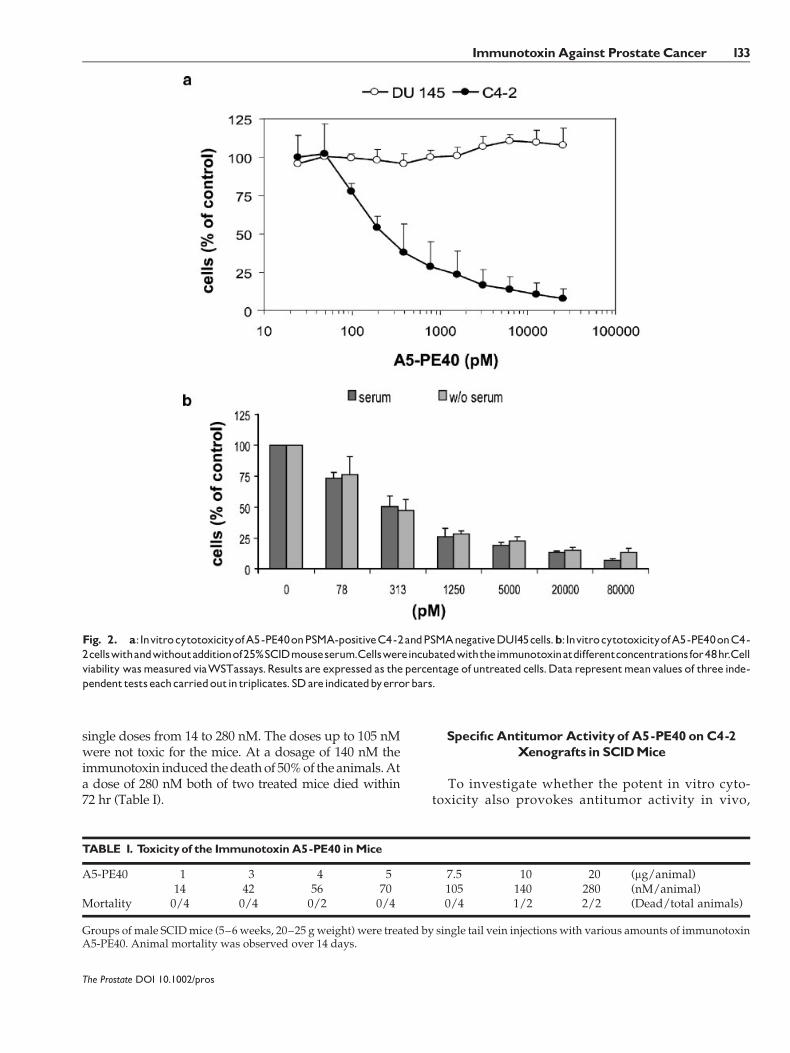
The cytotoxicity of the immunotoxin A5-PE40 wasdetermined in vitro with a WST viability assay. OnPSMA-expressing C4-2 target cells a dose-dependentkilling could be shown with an IC50-value of 220 pMafter 48 hr incubation. In contrast, A5-PE40 did not
show toxicity against PSMA-negative DU145 cells atthese concentrations (Fig. 2a).
To exclude the serum proteins’ influence on thecytotoxicity of A5-PE40, additional WST-tests wereperformed. It could be shown that similar IC50-valueswere measured with or without addition of 25% SCIDmouse serum (Fig. 2b).
Tolerabilityof A5-PE40 in SCIDMice
For testing the tolerability in vivo, the immunotoxinA5-PE40was applicated intravenously into SCIDmice at
The Prostate DOI 10.1002/pros
Fig. 1. a:Concentration-dependentbindingofimmunotoxinA5-PE40andanti-PSMAscFvA5toC4-2targetcells.b:BindingofA5-PE40onC4-2 (A) andDU145 (B) cells at saturating conditions (120 nM).Cellswere stainedwithA5-PE40,mouse anti-humanc-myc antibody andgoatantimouse Ig-RPE and analyzed via flowcytometry.Mean fluorescence intensity valueswere considered after subtraction of thebackgroundstainingwith secondaryantibodyalone.
132 Wolf et al.
single doses from 14 to 280 nM. The doses up to 105 nMwere not toxic for the mice. At a dosage of 140 nM theimmunotoxin induced thedeathof 50%of theanimals.Ata dose of 280 nM both of two treated mice died within72 hr (Table I).
Specif|c AntitumorActivityof A5-PE40 onC4-2Xenografts in SCIDMice
To investigate whether the potent in vitro cyto-toxicity also provokes antitumor activity in vivo,
The Prostate DOI 10.1002/pros
Fig. 2. a:InvitrocytotoxicityofA5-PE40onPSMA-positiveC4-2andPSMAnegativeDU145cells.b:InvitrocytotoxicityofA5-PE40onC4-2cellswithandwithoutadditionof25%SCIDmouseserum.Cellswereincubatedwiththeimmunotoxinatdifferentconcentrationsfor48hr.Cellviability wasmeasured viaWSTassays.Results are expressed as thepercentage of untreated cells.Data representmeanvalues of three inde-pendenttestseachcarriedoutin triplicates. SDareindicatedbyerrorbars.
TABLE I. Toxicityof the ImmunotoxinA5-PE40 inMice
A5-PE40 1 3 4 5 7.5 10 20 (mg/animal)14 42 56 70 105 140 280 (nM/animal)
Mortality 0/4 0/4 0/2 0/4 0/4 1/2 2/2 (Dead/total animals)
Groups of male SCIDmice (5–6weeks, 20–25 g weight) were treated by single tail vein injections with various amounts of immunotoxinA5-PE40. Animal mortality was observed over 14 days.
ImmunotoxinAgainst Prostate Cancer 133

A5-PE40 was tested in a SCID mouse xenograftmodel. In this model about 90% of the micexenotransplanted with C4-2 cells developed solidtumors within 9 days, whereas mice inoculatedwith DU145 cells already generated tumors within6 days.
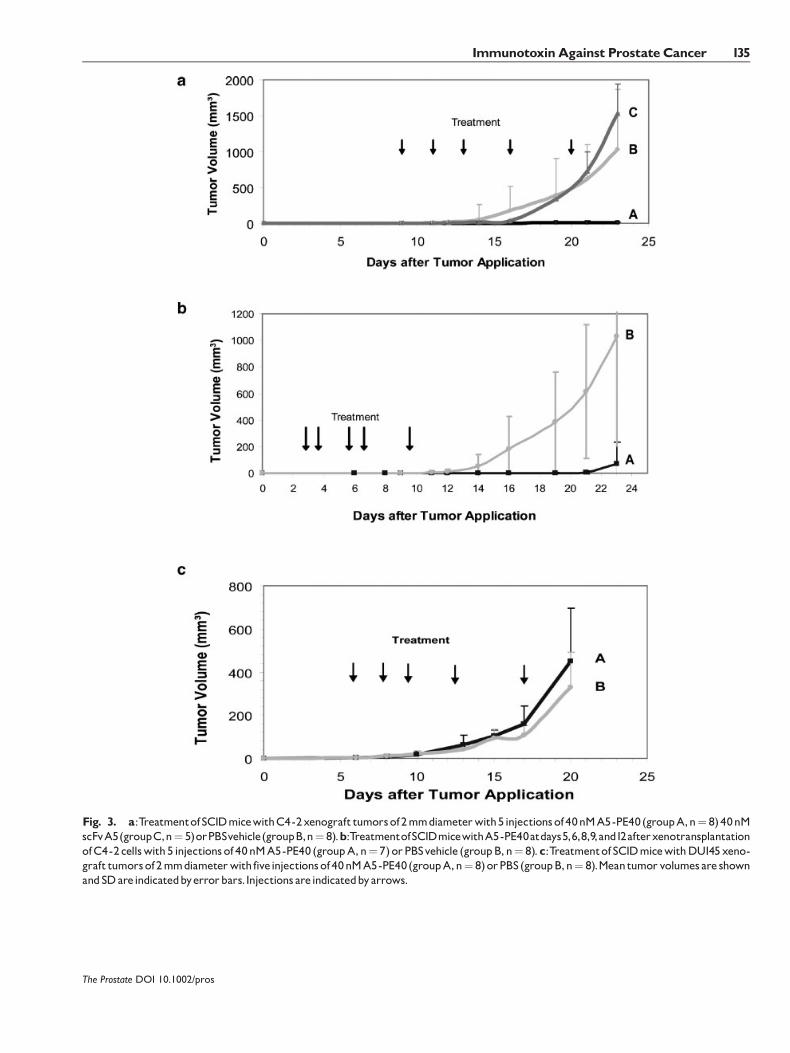
In a first series of experiments, 21 mice bearingpalpable C4-2 xenograft tumors with a diameter ofabout 2 mm diameter were randomized into threegroups:GroupA (n¼ 8)was treatedwith five injectionsof 40 nM immunotoxin A5-PE40 within 12 days,whereas the animals of the control group B (n¼ 8)received PBS as vehicle. The animals of group C (n¼ 5)were treated with five injections of 40 nM scFv A5within 12 days. The observation of tumor growth wasdiscontinued on day 22.
As shown in Figure 3a, all mice treated with A5-PE40 (group A) developed very slow growing tumorsuntil the end of the experiment. The mean tumorvolume in this group increased from 2.37 (�1.12) mm3
to 5.58 (�7.5) mm3. In contrast, the mice treated withscFv A5 (group C) showed a rapid tumor growth withmean tumor volumes increasing from 1.77 (�0.12) mm3
to 1528 (�411) mm3. Also in the PBS control group(group B) the mean tumor volumes increased from2.87 (�2.57) to 1032 (�831) mm3 during the experi-ment. In this group six animals showed a very fasttumor growth, whereas two animals had slowergrowing tumors. The differences in tumor sizebetween groups A and C were statistically significant(P� 0.001) and also between group A and the PBScontrol group B (P� 0.01).
In a second series of experiments mice wereinoculated with C4-2 cells and treated before tumorswere palpable. Sevenmicewere treatedwith 40 nMA5-PE40 at day 5, 6, 8, 9, and 12 after xenotransplantation ofC4-2 cells (group A) and compared to eight controlanimals that were treated with PBS vehicle (group B).As shown in Figure 3b, there was a significant lowertumor growth in the A5-PE40 treated group comparedto the PBS group. In theA5-PE40 treatment group threeanimals had no tumor, three animals had very slowgrowing tumors, and one animal had a fast growingtumor, in the PBS group two animals had slow tomediate tumor growth, and six animals had fastgrowing tumors. At the end of the experiment (day23) the mean tumor volume in the treatment groupwas 67.9 (�168) mm3 compared to 1,032 (�830) mm3
in the control group. The tumor volume differencesin the two groups were statistically significant(P� 0.001).
In a third series 16 mice bearing PSMA-negativeDU145 xenograftswere randomizedwhen the palpabletumors had reached about 2 mm diameter. Eightanimals were then treated with five injections of
40 nM A5-PE40 (group A) and eight animals with PBSvehicle (group B) within 12 days. As shown inFigure 3c, mice treated with A5-PE40 (n¼ 8) had amean final tumor volume of 449 (�246) mm3, whereasmice of the PBS control group (n¼ 8) showed a meantumor volume of 329 (�163) mm3 at the end of theexperiment. The differences of tumor volumes in bothgroups were not statistically significant, which provesthat A5-PE40 lacks an antitumorous effect against thePSMA-negative DU145 tumors.
Bindingof A5-PE40 toTumorXenograftsButNottoMouseOrgans
For demonstrating the reactivity of A5-PE40 onhuman PSMA but yet not on the highly homologousmouse PSMA [17], a discrimination of the expression ofthese proteins in the xenograft tumors and mouseorgans was necessary. Therefore, RT-PCRs for humanand mouse PSMA as well as GAPDH were performedwith lysates of the C4-2 and DU145 xenografts, mousekidneys, brains, and livers. As expected, C4-2 andDU145 tumors expressed human GAPDH and onlya minimal amount of mouse GAPDH, which can betraced back to a fewmurine cells in the tumors. The C4-2 tumors but not DU145 tumors strongly expressedhuman PSMA. Mouse PSMA could be stronglydetected in the kidney, weakly in the brain and wasundetectable in the liver, which served as control organ(Fig. 4).
On the basis of these expression experiments it couldbe demonstrated byWestern blot that A5-PE40 and theparental monoclonal antibody 3/A12 only bound tolysates of humanPSMAexpressingC4-2 xenografts butnot to mouse PSMA expressing kidney and brainlysates (Fig. 5).
DISCUSSION
In the present studywe tested the antitumor activityofA5-PE40 in a SCID-mouse C4-2 xenograftmodel. C4-2 is a subline of the human prostate cancer cell lineLNCaP [18]. C4-2 cells express PSMA, grow androgen-independent and have a high proclivity for bonemetastases [19]. Therefore, these cells represent avaluable model for hormone-refractory advancedhuman prostate cancer.
As a precondition for the in vivo experiments, astrong binding of theA5-PE40 to C4-2 target cells couldbe shown, comparable to that of scFvA5. This indicatesthat the toxin domain does not sterically inhibit thebinding domain. The binding of the A5-PE40 resultedin a specific in vitro cytotoxicity on C4-2 cells whereasnon-PSMAexpressingDU145 cellswerenot affected bythe immunotoxin.
The Prostate DOI 10.1002/pros
134 Wolf et al.
The Prostate DOI 10.1002/pros
Fig. 3. a:TreatmentofSCIDmicewithC4-2xenograft tumorsof2mmdiameterwith5injectionsof40nMA5-PE40(groupA,n¼ 8)40nMscFvA5(groupC,n¼ 5)orPBSvehicle(groupB,n¼ 8).b:TreatmentofSCIDmicewithA5-PE40atdays5,6,8,9,and12afterxenotransplantationofC4-2cellswith5 injectionsof40nMA5-PE40 (groupA,n¼ 7)orPBSvehicle (groupB,n¼ 8).c:TreatmentofSCIDmicewithDU145xeno-graft tumorsof2mmdiameterwithfiveinjectionsof40nMA5-PE40(groupA,n¼ 8)orPBS(groupB,n¼ 8).Meantumorvolumesare shownandSDareindicatedbyerrorbars.Injections areindicatedbyarrows.
ImmunotoxinAgainst Prostate Cancer 135

Anticancer activities of oncotoxins are often limited,mainly due to their inactivation in the serum, due totheir binding to serum components and enzymaticdegradation [20,21]. Testing the immunotoxinA5-PE40for 48 hr in thepresence or absence of 25%mouse serumin WST assays showed no difference in cytotoxicityagainst C4-2 target cells. This suggests that thestructure and enzymatic activity of the immunotoxinis not influenced by serum components.
A further important aspect to be considered was apossible cross reactivity of the anti-PSMA immuno-toxin with murine PSMA, which is highly homologousto human PSMA [17]. Although mouse PSMA expres-sion could be detected in mouse kidney and brain,according to former publications [17,22] the immuno-toxin A5-PE40 did not show binding to these organs inWestern blot analyses, which proves that it does notbind to mouse PSMA.
Concerning tolerability and unspecific toxicity it canbe stated that the immunotoxin was well tolerated inSCIDmicewith a dosage up to 105 nM.At higher doses
A5-PE40 proved to be toxic, which is in accordance toother published PE-based immunotoxins [23,24].
Toxic side effects of immunotoxins in animals andhumans generally consist of two types: (a) targetedkilling of normal cells that have the same or ahomologous target antigen or (b) non-specific bindingto cells of other organs. From themissing binding ofA5-PE40 to several mouse organs, a targeted killing ofnormal cells in the SCID mouse seems improbable. Forunspecific toxicity, the liver is particularly vulnerableand a liver toxicity of PE-based immunotoxins hasalready been shown in literature [25–27]. Therefore ahepatotoxicity may also be assumed in our study. Areduction of animal toxicity caused by PE-basedimmunotoxins could be achieved by reducing theisoelectric point (pI) of the Fv, [25] by inhibition of theTNF-a production by Kupffer cells [26] or by chemicalmodification with polyethylene glycol [28].
Another problem of A5-PE40 is the possible immu-nogenicity of the non-human fusion protein. In order toreduce this immunogenicity, we are going to humanizethe mouse framework of the A5 scFv.
In the present study, A5-PE40 was applicated at twodifferent times of tumor growth: In a first series whenxenografts were grown up to 2–3 mm diameter(corresponding to about 14 days after tumor induction)and in a second series already at day 6 after tumorinduction. In both therapy schemes a significantinhibition of tumor growth was observed. However,the result was somewhat worse in the second group,due to one rather fast growing tumor.
Generally, an early treatment of relatively smalltumors with immunotoxins has been reported to givethe best results. Thus in a mouse renal cell carcinomamodel the administration of an anti-erbB-2-ETAimmunotoxin over a 10-day period starting 1 day aftertumor cell transplantation completely suppressed theformation of metastases, whereas treatment of animals11 days after tumor cell transplantation only led to areduction in their number and size [29]. The immuno-toxin dosage in our study is comparable to otherpublished studies with PE-based immunotoxins. DiPaolo et al. [30] could inhibit tumor growth of Ep-CAMexpressing xenograft-tumors of 160 mm3 size with ananti-Ep-CAM-ETA immunotoxin at a dosage of9� 5 mg or 3� 10 mg per animal every second day.Beginning at day 5 after tumor induction Kuan andPastan could demonstrate a significant dose dependentantitumor activity of an anti-Lewis-ETA immunotoxin.Complete remissions of Lewis-positive xenografttumors were obtained with a dosage of only3� 12 mg/kg [31]. A remission of small MCF7 andOHS-M1 tumors after application of 0.075 mg/kg or0.15 mg/kg recombinant immunotoxin 8H9(Fv)-PE38could be shown by Onda et al. [23].
The Prostate DOI 10.1002/pros
Fig. 4. Gene expressionofhumanandmurine PSMAinC4-2 andDU145xenograftsandmouseorgans.a:HumanGAPDH,(b)murineGAPDH,(c)humanPSMA,and (d)murinePSMA.
Fig. 5. Western blot with the immunotoxin A5-PE40 and theparentalmonoclonal antibody 3/A12 on lysates of C4-2 and DU145xenografts andmouse organs.HumanPSMAwas detected inC4-2tumorsas100kDaprotein.b-Actin(42kDa)wasdetectedasloadingcontrolinall samples.
136 Wolf et al.

With respect to the PSMA target antigen no otherrecombinant immunotoxin has been published. There-fore, only a comparison with chemically conjugatedimmunotoxins is possible. Huang et al. could preventprogression of LNCaP xenograft tumors of 150 mm3
size on nudemice with 2� 50 mg Immunotoxin E6-dgA[32]. Also Russell et al. could obtain a 48% tumorreduction of LNCaP-LN3 xenografts with a chemicallyconjugated anti-PSMA immunotoxin. The only partialtumor inhibition was suggested to be due to impuritiesof the preparation and the rather big tumors treated[33]. Better results were obtained by Ma et al. [34] whoapplied high doses of a chemically conjugated anti-PSMA-Auristatin immunotoxin to nude mice bearingC4-2 tumors.
CONCLUSION
We have demonstrated that A5-PE40 provides apotent and specific antitumor efficacy on C4-2 xeno-grafts, even if the subcutaneous localization and thebad vascularization of the fast growing human xeno-graft tumors may not represent an optimal model. Ourresults indicate that the immunotoxin A5-PE40 may beeffective in eliminating or preventing tumors of lowburden, for example, micrometastases or tumors aftervolume reduction by brachytherapy or hormonaltreatment. Additionally, from the C4-2 model it maybe concluded that the immunotoxin could be effectiveonhormone refractory cancer, forwhichno treatment isavailable so far. Therefore, this approachmerits furtherdevelopment as a molecularly targeted therapy ofprostate cancer.
ACKNOWLEDGMENTS
We thank D. Gierschner, I. Rother-Jehle, and I.Schaber for technical assistance. This work wassupported in part by the Deutsche Krebshilfe, Bonn,Germany.
REFERENCES
1. Isaacs JT. New strategies for the medical treatment of prostatecancer. BJU Int 2005;96 (Suppl 2):35–40.
2. SonpavdeG,HutsonTE.Newapproaches in hormone refractoryprostate cancer. Am J Clin Oncol 2006;29(2):196–201.
3. Stewart AB, Lwaleed BA, Douglas DA, Birch BR. Current drugtherapy for prostate cancer: An overview. Curr Med ChemAnticancer Agents 2005;5(6):603–612.
4. Ross JS, Gray KE, Webb IJ, Gray GS, Rolfe M, Schenkein DP,Nanus DM, Millowsky MI, Bander NH. Antibody-basedtherapeutics: Focus on prostate cancer. Cancer Metastasis Rev2005;24(4):521–537.
5. GhoshA,HestonWD. Tumor target prostate specificmembraneantigen (PSMA) and its regulation in prostate cancer. J CellBiochem 2004;91(3):528–539.
6. LiuH,Moy P, Kim S, Xia Y, RajasekaranA,Navarro V, KnudsenB, Bander NH. Monoclonal antibodies to the extracellulardomain of prostate-specific membrane antigen also reactwith tumor vascular endothelium. Cancer Res 1997;57:3629–3634.
7. Chang SS, Reuter VE, Heston WD, Bander NH, Grauer LS,Gaudin PB. Five different anti-prostate-specific membraneantigen (PSMA) antibodies confirm PSMA expression intumor-associated neovasculature. Cancer Res 1999;59(13):3192–3198.
8. Milowsky MI, Nanus DM, Kostakoglu L, Sheehan CE, Vallab-hajosula S, Goldsmith SJ, Ross JS, Bander NH. Vascular targetedtherapy with anti-prostate-specific membrane antigen mono-clonal antibody J591 in advanced solid tumors. J Clin Oncol2007;25(5):540–547.
9. Mesters JR, Barinka C, Li W, Tsukamoto T, Majer P, Slusher BS,Konvalinka J, Hilgenfeld R. Structure of glutamate carboxypep-tidase II, a drug target in neuronal damage and prostate cancer.EMBO J 2006;25(6):1375–1384.
10. Schulke N, Varlamova OA, Donovan GP, Ma D, Gardner JP,Morrissey DM, Arrigale RR, Zhan C, Chodera AJ, Surowitz KG,MaddonPJ,HestonWD,OlsonWC.Thehomodimer of prostate-specific membrane antigen is a functional target for cancertherapy. Proc Natl Acad Sci USA 2003;100(22):12590–12595.
11. Elsasser-Beile U, Wolf P, Gierschner D, Buhler P, Schultze-SeemannW,WetterauerU.Anewgenerationofmonoclonal andrecombinant antibodies against cell-adherent prostate specificmembrane antigen for diagnostic and therapeutic targeting ofprostate cancer. Prostate 2006;66(13):1359–1370.
12. Kang N, Hamilton S, Odili J, Wilson G, Kupsch J. In vivotargeting of malignant melanoma by 125Iodine- and 99mTech-netium-labeled single-chain Fv fragments against high molec-ular weight melanoma-associated antigen. Clin Cancer Res2000;6(12):4921–4931.
13. Wu AM, Senter PD. Arming antibodies: Prospects and chal-lenges for immunoconjugates. Nat Biotechnol 2005;23(9):1137–1146.
14. Kreitman RJ. Chimeric fusion proteins—Pseudomonas exo-toxin-based. Curr Opin Investig Drugs 2001;2(9):1282–1293.
15. Jorgensen R, Merrill AR, Yates SP, Marquez VE, Schwan AL,Boesen T, Andersen GR. Exotoxin A-eEF2 complex structureindicates ADP ribosylation by ribosome mimicry. Nature 2005;436(7053):979–984.
16. Wolf P, Gierschner D, Buhler P, Wetterauer U, Elsasser-Beile U.A recombinant PSMA-specific single-chain immunotoxin haspotent and selective toxicity against prostate cancer cells. CancerImmunol Immunother 2006;55(11):1367–1373.
17. Bacich DJ, Pinto JT, TongWP, HestonWD. Cloning, expression,genomic localization, and enzymatic activities of the mousehomologof prostate-specificmembrane antigen/NAALADase/folate hydrolase. Mamm Genome 2001;12(2):117–123.
18. Thalmann GN, Anezinis PE, Chang SM, Zhau HE, Kim EE,Hopwood VL, Pathak S, von Eschenbach AC, Chung LW.Androgen-independent cancer progression and bonemetastasisin the LNCaP model of human prostate cancer. Cancer Res1994;54(10):2577–2581.
19. ThalmannGN, Sikes RA,WuTT,DegeorgesA, Chang SM,OzenM, Pathak S, Chung LW. LNCaP progression model of humanprostate cancer: Androgen-independence and osseous meta-stasis. Prostate 2000;44(2):91–103, Jul 101; 144(102).
20. Candiani C, Franceschi A, Chignola R, Pasti M, Anselmi C,Benoni G, Tridente G, Colombatti M. Blocking effect of humanserum but not of cerebrospinal fluid on ricin A chain
The Prostate DOI 10.1002/pros
ImmunotoxinAgainst Prostate Cancer 137

immunotoxin potentiation by monensin or carrier protein-monensin conjugates. Cancer Res 1992;52(3):623–630.
21. Franceschi A, Dosio F, Anselmi C, Chignola R, Candiani C, PastiM, Tridente G, Colombatti M. Mechanisms involved in serum-dependent inactivation of the immunotoxin enhancers mon-ensin and carrier-protein-monensin. Eur J Biochem 1994;219(1–2):469–479.
22. Schmittgen TD, Zakrajsek BA, Hill RE, Liu Q, Reeves JJ, AxfordPD, Singer MJ, Reed MW. Expression pattern of mouse homologof prostate-specific membrane antigen (FOLH1) in the transgenicadenocarcinoma of the mouse prostate model. Prostate 2003;55(4):308–316.
23. Onda M, Wang QC, Guo HF, Cheung NK, Pastan I. In vitro andin vivo cytotoxic activities of recombinant immunotoxin8H9(Fv)-PE38 against breast cancer, osteosarcoma, and neuro-blastoma. Cancer Res 2004;64(4):1419–1424.
24. Bang S, Nagata S, OndaM, KreitmanRJ, Pastan I. HA22 (R490A)is a recombinant immunotoxinwith increasedantitumor activitywithout an increase in animal toxicity. Clin Cancer Res 2005;11(4):1545–1550.
25. OndaM,KreitmanRJ, VasmatzisG, LeeB, Pastan I. Reduction ofthe nonspecific animal toxicity of anti-Tac(Fv)-PE38 by muta-tions in the framework regions of the Fv which lower theisoelectric point. J Immunol 1999;163(11):6072–6077.
26. Onda M, Willingham M, Wang QC, Kreitman RJ, Tsutsumi Y,Nagata S, Pastan I. Inhibition of TNF-alphaproducedbyKupffercells protects against the nonspecific liver toxicity of immuno-toxin anti-Tac(Fv)-PE38, LMB-2. J Immunol 2000;165(12):7150–7156.
27. Kennedy PE, Bera TK, Wang QC, Gallo M, Wagner W, LewisMG, Berger EA, Pastan I. Anti-HIV-1 immunotoxin 3B3(Fv)-PE38:Enhancedpotencyagainst clinical isolates inhumanPBMCsand macrophages, and negligible hepatotoxicity in macaques.J Leukoc Biol 2006;80(5):1175–1182.
28. Tsutsumi Y,OndaM,Nagata S, Lee B, KreitmanRJ, Pastan I. Site-specific chemical modification with polyethylene glycol ofrecombinant immunotoxin anti-Tac(Fv)-PE38 (LMB-2) improvesantitumor activity and reduces animal toxicity and immunoge-nicity. Proc Natl Acad Sci USA 2000;97(15):8548–8553.
29. Maurer-Gebhard M, Schmidt M, Azemar M, Altenschmidt U,Stocklin E, Wels W, Groner B. Systemic treatment with arecombinant erbB-2 receptor-specific tumor toxin efficientlyreduces pulmonarymetastases in mice injected with geneticallymodified carcinoma cells. Cancer Res 1998;58(12):2661–2666.
30. Di Paolo C, Willuda J, Kubetzko S, Lauffer I, Tschudi D, WaibelR, Pluckthun A, Stahel RA, Zangemeister-Wittke U. A recombi-nant immunotoxin derived from a humanized epithelial celladhesion molecule-specific single-chain antibody fragment haspotent and selective antitumor activity. Clin Cancer Res 2003;9(7):2837–2848.
31. Kuan CT, Pastan I. Improved antitumor activity of a recombi-nant anti-Lewis(y) immunotoxin not requiring proteolyticactivation. Proc Natl Acad Sci USA 1996;93(3):974–978.
32. Huang X, Bennett M, Thorpe PE. Anti-tumor effects and lack ofside effects in mice of an immunotoxin directed against humanand mouse prostate-specific membrane antigen. Prostate 2004;61(1):1–11.
33. Russell PJ, Hewish D, Carter T, Sterling-Levis K, OwK, HattarkiM, Doughty L, Guthrie R, Shapira D, Molloy PL, WerkmeisterJA, Kortt AA. Cytotoxic properties of immunoconjugatescontaining melittin-like peptide 101 against prostate cancer: Invitro and in vivo studies. Cancer Immunol Immunother 2004;53(5):411–421.
34. Ma D, Hopf CE, Malewicz AD, Donovan GP, Senter PD,GoeckelerWF,Maddon PJ, OlsonWC. Potent antitumor activityof anauristatin-conjugated, fullyhumanmonoclonal antibody toprostate-specificmembrane antigen. Clin Cancer Res 2006;12(8):2591–2596.
The Prostate DOI 10.1002/pros
138 Wolf et al.





























