Embed Size (px)
Citation preview

AbstractTelomeres consist of repetitive GC-rich DNA and associated proteins. These repetitive sequences are known to be natural impediments to the DNA replication fork machinery. A novel telomere associated complex, CTC1-STN1-TEN1 (CST), was recently identified in mammals and plants. It shares significant homology with the budding yeast CST (Cdc13-Stn1-Ten1) complex. Moreover, CST shows remarkable similarity to the single-stranded binding protein RPA. In yeast, CST regulates telomere and G-overhang length through interactions with telomerase and DNA polymerase (pol) α. Interestingly, the recently discovered mammalian CST also interacts with DNA pol α. Our initial studies showed that depletion of human CST leads to telomere loss and increased G-overhang length as well as more general genomic instability, i.e increased H2AX foci and anaphase bridges. We hypothesize that CST plays a dual role in replication of the telomeric tract and C-strand fill-in synthesis after telomerase extension of the telomere. Consistent with our hypothesis, current studies show that depletion of CST results in defects associated with replication of the telomeric DNA and with C-strand fill-in synthesis. Intriguingly, knockdown of CST shows delayed replication fork reinitiation after replication fork stalling, suggesting that CST may also play a role in general DNA replication at other sites of difficult-to-replicate DNA. Our findings suggest that CST functions as a novel replication/repair factor that acts in situations necessitating a specialized mechanism for pol α recruitment. (NIH-GM041803; NIH-T32CA117846)
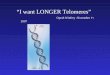
Telomere Structure & Replication
A, Mammalian telomeres consist of repetitive sequence (TTAGGG) and are bound by a six subunit complex, named shelterin. Telomeres are proposed to form a t-loop structure, which prevents DNA damage signaling. B, Telomere replication consist of three major stages. First, replication of the telomeric DNA. Second, the ends are processed to form a 3' overhang and the telomere is extended by telomerase. Third, the C-strand is filled-in, creating a short G-strand overhang for t-loop formation.
POT1
TPP1
TRF2TRF15’
RAP1 TIN2A B
Stage I
5’
5’3’
3’
+
G-strand
C-strand
Stage II
Stage III3’5’

• Shelterin consists of six subunits, of which all but one are essential in mice.
• Each subunit performs a specific task in telomere maintenance.
• In budding yeast, no shelterin-like proteins have been identified.
• Instead, a complex consisting of Cdc13, Stn1 and Ten1 (CST) protects and maintains the telomere.
• Until recently, a CST-like complex had not been identified in higher eukaryotes.
• Studies from our group and others identified a CST-like complex (CTC1-STN1-TEN1) in plants and humans.
• The mammalian CST complex appears to function in telomere maintenance.
Shelterin vs. CST Complex
Ten1 Stn1
S. cerevisiae Est1 TelomeraseCdc13
Polα/primase
??
Pot1
Tpp1
Trf2
Trf1 Tin2
Rap1
Vertebrates
Telomerase
Polα/primase
CST
??

DNA polymerase (pol ) accessory factor:
-CTC1 and STN1 were originally identified as pol accessory factors (AAF)132 and AAF44, respectively1,2
-Increases pol binding and polymerase/primase activities2
RPA-like characteristics:
-Contains multiple OB-folds and binds ssDNA3,4
-Heterotrimeric
-Yeast structure of Stn1-Ten1 shows structural homology to RPA2-RPA35
-NMR structure of mouse STN1 resembles that of RPA26
Mammalian CST complex
siRNA knockdown of CST subunits:
At the telomere:
- Loss of telomeric signal, by telomere FISH (STN1)3
- Increase in G-overhang length (Both CTC1 and STN1)3,4
Genome instability (CTC1):
-Increase in anaphase bridges3
-Increased H2AX staining3
1. Casteel et al., JBC, 20092. Goulin et al., JBC, 19903. Surovtseva et al., Mol Cell, 20094. Miyake et al., Mol Cell, 20095. Sun et al., Genes Dev, 20096. PDBID: 1WJ5
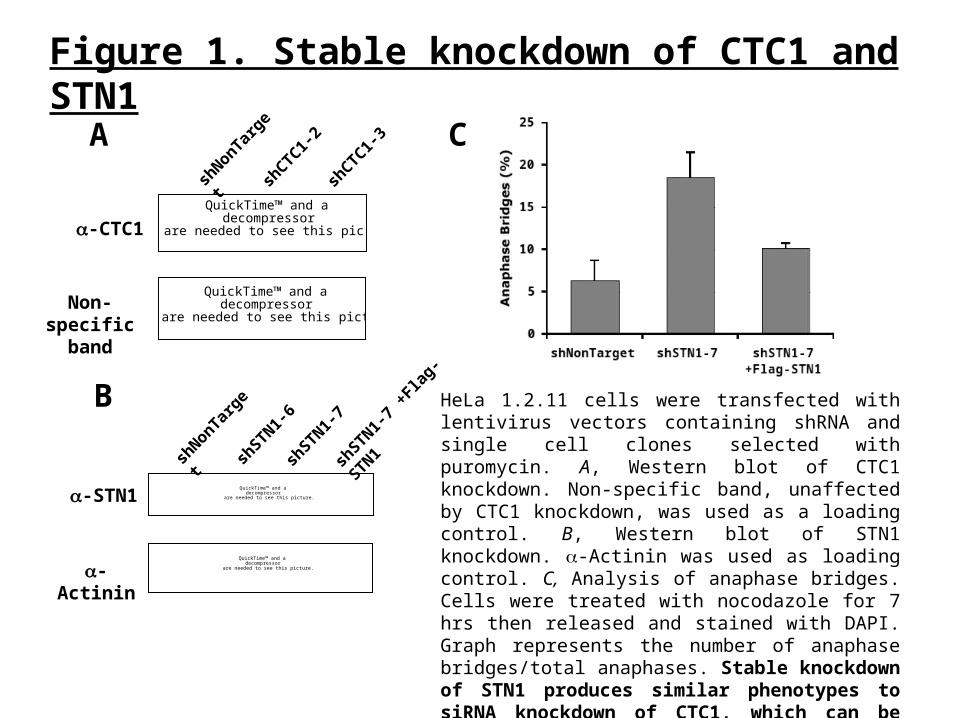
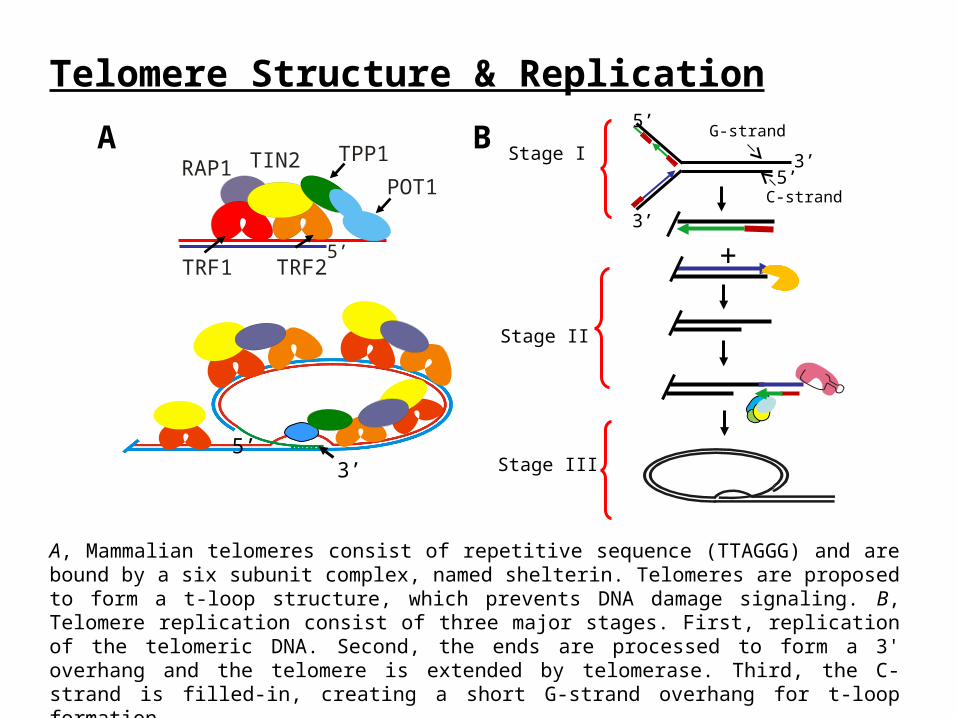
Figure 1. Stable knockdown of CTC1 and STN1
HeLa 1.2.11 cells were transfected with lentivirus vectors containing shRNA and single cell clones selected with puromycin. A, Western blot of CTC1 knockdown. Non-specific band, unaffected by CTC1 knockdown, was used as a loading control. B, Western blot of STN1 knockdown. -Actinin was used as loading control. C, Analysis of anaphase bridges. Cells were treated with nocodazole for 7 hrs then released and stained with DAPI. Graph represents the number of anaphase bridges/total anaphases. Stable knockdown of STN1 produces similar phenotypes to siRNA knockdown of CTC1, which can be rescued by expression of shRNA resistant Flag-STN1.
QuickTime™ and a decompressor
are needed to see this picture.
QuickTime™ and a decompressor
are needed to see this picture.
shCTC1-
2sh
CTC1-3
-CTC1
Non-specific band
A
shNonTar
get
Csh
NonTarget
shSTN1-
7
shSTN1-
6
-STN1
-Actinin
B
QuickTime™ and a decompressor
are needed to see this picture.
shSTN1-
7 +F
lag-S
TN1
QuickTime™ and a decompressor
are needed to see this picture.
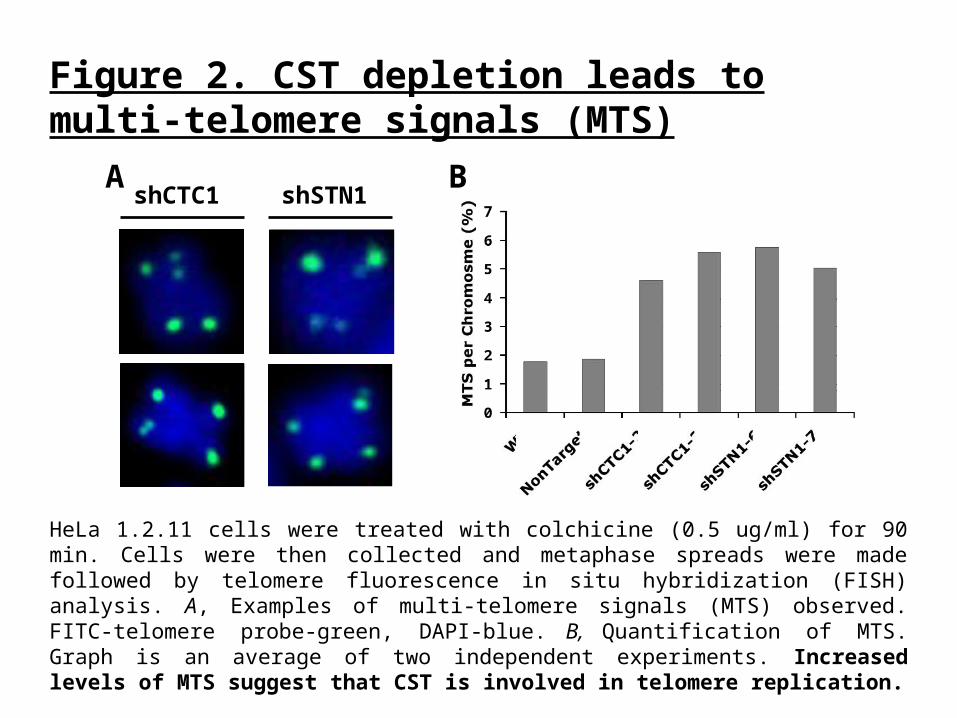
Figure 2. CST depletion leads to multi-telomere signals (MTS)
HeLa 1.2.11 cells were treated with colchicine (0.5 ug/ml) for 90 min. Cells were then collected and metaphase spreads were made followed by telomere fluorescence in situ hybridization (FISH) analysis. A, Examples of multi-telomere signals (MTS) observed. FITC-telomere probe-green, DAPI-blue. B, Quantification of MTS. Graph is an average of two independent experiments. Increased levels of MTS suggest that CST is involved in telomere replication.
shCTC1 shSTN1A B

What are MTS?• MTS are proposed to arise from replication defects at the telomere due to the
repetitive G-rich sequences.
• Other occurrences of MTS:
-Previously seen in cells from TRF1 knockout mice (TRF1 was shown to function in telomere replication)1,2
-Also seen with FEN1 knockdown in primary cell lines (BJ fibroblast)3
• MTS are similar to common fragile sites:
-Arise when cells are treated with aphidicolin, which inhibits DNA pol /
-ATR-dependent
• Common fragile sites
-Cytogenetically characterized as gaps or breaks in the chromosome
-Induced with replication-stress
-Proposed to occur at sites of DNA secondary structure formation4
5’3’
Fork stalling
5’3’
Accumulationof ssDNA
Single strandgaps
Discontinuities inchromatin structure
Possible cause of MTS/fragile telomeres
1. Sfeir et al., Cell, 2009 2. Martinez et al., Genes Dev, 2009 3. Saharia et al., JBC, 2010 4. Burrow et al., NAR, 2010
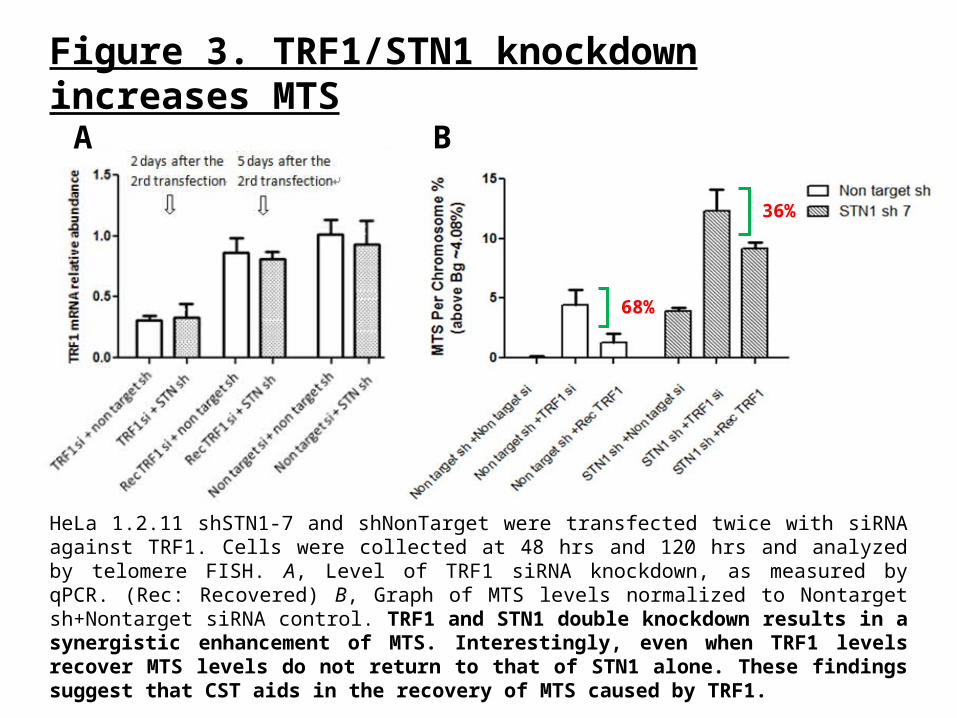
Figure 3. TRF1/STN1 knockdown increases MTS
HeLa 1.2.11 shSTN1-7 and shNonTarget were transfected twice with siRNA against TRF1. Cells were collected at 48 hrs and 120 hrs and analyzed by telomere FISH. A, Level of TRF1 siRNA knockdown, as measured by qPCR. (Rec: Recovered) B, Graph of MTS levels normalized to Nontarget sh+Nontarget siRNA control. TRF1 and STN1 double knockdown results in a synergistic enhancement of MTS. Interestingly, even when TRF1 levels recover MTS levels do not return to that of STN1 alone. These findings suggest that CST aids in the recovery of MTS caused by TRF1.
A B
68%
36%
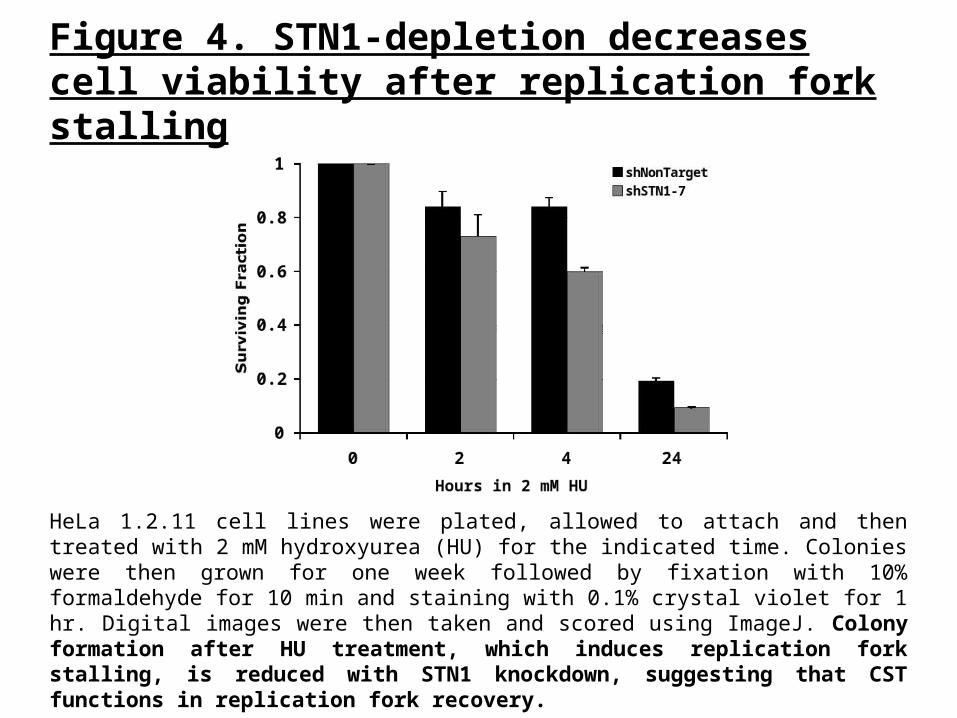
Figure 4. STN1-depletion decreases cell viability after replication fork stalling
HeLa 1.2.11 cell lines were plated, allowed to attach and then treated with 2 mM hydroxyurea (HU) for the indicated time. Colonies were then grown for one week followed by fixation with 10% formaldehyde for 10 min and staining with 0.1% crystal violet for 1 hr. Digital images were then taken and scored using ImageJ. Colony formation after HU treatment, which induces replication fork stalling, is reduced with STN1 knockdown, suggesting that CST functions in replication fork recovery.

HeLa 1.2.11 cells were treated for 2 hrs with either 2 mM HU or 7.5 ug/ml APH and released into media containing 50 M EdU for 30 minutes. Coverslips were then processed using the Click-iT EdU Alexa-Fluor 488 Imaging Kit (Invitrogen). Blue-DAPI, Green-EdU. Recovery after either HU or APH treatment results in decreased replication reinitiation, as measured by the incorporation of the nucleoside analog EdU after fork stalling.
Figure 5. STN1-depletion delays DNA replication reinitiation after fork stalling
BshNonTarget +HU
shSTN1-7 +HU
Merge EdU
A
shSTN1-7 + Flag -STN1 + HU

Conclusions
• The heterotrimeric CST complex has been shown to bind ssDNA, interact with pol and partially localize to telomeres
• Depletion of either CTC1 or STN1 leads to an increase in MTS, which suggests dysfunctional telomere replication
• Knockdown of TRF1 in the STN1-depleted cell line increases MTS relative to either TRF1 or STN1 knockdown alone, suggesting that CST promotes the recovery of TRF1 induced MTS
• Induction of replication fork stalling in STN1-depleted cells leads to reduced cell survival and delayed fork reinitiation, indicting that CST is involved in DNA replication reinitiation after fork stalling
• Our studies suggest that the CST complex functions as a novel DNA replication/repair factor, which promotes DNA replication through the recruitment of DNA pol .
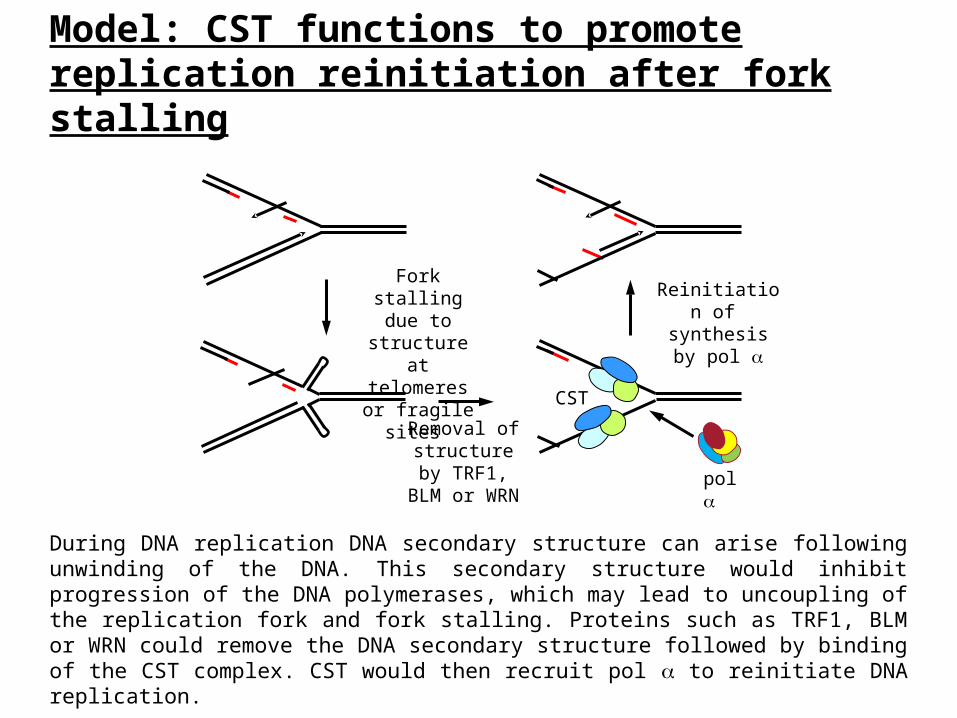
Model: CST functions to promote replication reinitiation after fork stalling
During DNA replication DNA secondary structure can arise following unwinding of the DNA. This secondary structure would inhibit progression of the DNA polymerases, which may lead to uncoupling of the replication fork and fork stalling. Proteins such as TRF1, BLM or WRN could remove the DNA secondary structure followed by binding of the CST complex. CST would then recruit pol to reinitiate DNA replication.
Removal of structure by TRF1, BLM
or WRN
Fork stalling due to structure at telomeres or
fragile sites
Reinitiation of synthesis by
pol
pol
CST












![Mrc1 protects uncapped budding yeast telomeres from ... · proteins, Cdc13, Stn1 and Ten1 [6–11]. If telomeres become uncapped, they activate a DNA dam-age response pathway leading](https://img.dokumen.tips/doc/110x75/5f1e3fac2ea8c846df63c904/mrc1-protects-uncapped-budding-yeast-telomeres-from-proteins-cdc13-stn1-and.jpg)