Embed Size (px)
Citation preview

Botanical journal of the Linnean Society (1991), 105: 137-177. With 5 figures
A synopsis of the neotropical species of Dalechampia (Euphorbiaceae)
GRADY L. WEBSTER, F.L.S.
Department of Botany, University of California, Davis, California 95616, U.S .A .
AND
W. SCOTT ARMBRUSTER
Department of Biology and Wildlge and Institute of Arctic Biology, University of Alaska, Fairbanks, Alaska 99701, U .S .A .
Receiued June 1989, accepted f o r publication November 1989
WEBSTER, G. L. & ARMBRUSTER, W. S., 1991. A synopsis of the neotropical species of Dalechampia (Euphorbiaceae). The characters used in inferring phylogenetic relationship in Dalechampia are discussed, and a consistent terminology is devised for the parts of the distinctive pseudanthial inflorescences. The infrageneric classification is radically modified, with the 92 provisionally accepted binomials for the neotropical taxa referred to six sections: Rhopalostylis, Dioscoreifliae, C7emophyllum, Coriaceae, Ti l i fol iae and Dalechampia. As here interpreted, section Dalechampia includes 53 species in five subsections; one new subsection, Convolvuloides, is described.
ADDITIONAL KEY WORDS:-Phylogenetic relationship - terminology - infrageneric classification.
CONTENTS
Introduction . . . . Morphological characters .
Growth form . . . Leaves . . . . Indumentum . . . Inflorescence . . . Flowers . . . . Fruits and seeds . .
Ecology . . . . . Systematic treatment . .
Dalechampia L. . . Key to the sections . Section I . Rhopalostylis . Section 2. Dioscoreifoliae Section 3. Cremophyllum Section 4. Coriaceae . Section 5 . Tiliifoliae . Section 6. Dalechampia .
0024-4074/91/020137 + 41 $03.00/0
. . .
. . .
. . .
. . .
. . .
. . .
. . .
. . .
. . .
. . .
. . .
. .
. . . .
. . . .
. . . .
. . . .
. I . .
. . . .
. . . .
. . . .
. . . .
. . . .
. . . .
. . . . 137
. . . . . . . . 138
. . . . . . . . 139
. . . . . . . . 139
. . . . . . . . 141
. . . . . . . . 141
. . . . . . . . 142
. . . . . . . . 146
. . . . . . . . 147
. . . . . . . . 147
. . . . . . . . 148
. . . . . . . . 148
. . . . . . . . 149
. . . . . . . . 150
. . . . . . . . 151
. . . . . . . . 156
. . . . . . . . 157
. . . . . . . . 157
. . . . . . . . 158
0 1991 The Linnean Society of London

138 G. L. WEBSTER AND W. S. ARMBRUSTER
Species of uncertain status . . . . . . . . . . . . . . . I 7 I Addendum . . . . . . . . . . . . . . . . . . . 171 Excluded taxa. . . . . . . . . . . . . . . . . . . 171 Acknowledgements . . . . . . . . . . . . . . . . . 17 1 Indextotaxa. . . . . . . . . . . . . . . . . . . 173
INTRODUCTION
Dalechampia, a genus of over 100 tropical species, is unique within the Euphorbiaceae because of its bilabiate pseudanthia which are fundamentally different from the pseudanthia of other genera such as Peru and Pedilanthus. According to Mueller (1866) and Pax & Hoffmann (1919), Dalechampia is the only genus of the tribe Dalechampieae; however, i t is closely related to taxa in tribe Plukenetieae (Bentham, 1880).
The pseudanthium of Dalechampia is distinguished in most species by two large, often coloured, involucral bracts inserted below a 3-flowered pistillate cyme and a several-flowered staminate pleiochasium (Webster & Webster, 1972). The most unusual feature of the inflorescence, however, is the secretory gland located within the staminate subflorescence. In the great majority of species, the inflorescences are visited by female bees that collect resin for nest-building from the gland of the staminate subflorescence, but in a few species the gland produces aromatic compounds rather than resin, and their inflorescences are visited by male euglossine bees (Armbruster & Webster, 1979). The great variety in inflorescence characters such as size and colour of bracts and glands, number of stamens, and stylar column length reflects a considerable diversity in apoidean pollinators, especially in the Megachilidae and Euglossini (Armbruster & Webster, 1982; Armbruster & Herzig, 1984).
There is also notable variation in vegetative characters within the neotropical species of Dalechampia. Although the majority are twining vines, erect or decumbent subshrubs are widespread in Brazil and adjacent South America. Leaf shape is highly diverse, ranging from simple and unlobed to 3- or 5-lobed or palmately parted. Leaf polymorphism is marked in some species, where unlobed and lobed or parted leaves may be found on the same plant.
Despite the great diversity of the neotropical species of Dalechampia, progress in understanding phylogenetic relationships between the species has been very slow. Baillon (1858) divided the genus into three sections, two of these unnamed and the third, section Cremophyllum, based on the shrubby species now known as D . spathulata (Scheidw.) Baillon. Mueller (1866, 1874) created a new section, Champadelia, for Baillon’s section A, which included only the Brazilian D . houlletiana Baillon; he referred all of the other species (including Cremophyllum) to section Eudalechampia. Within section Eudalechampia, Mueller recognized a number of informal (unnamed) groups, largely defined on characters of habit and leaf shape. Pax (1890) at first accepted the arrangement of Mueller, but later Pax & Hoffmann (1919) resurrected section Cremophyllum and described ten new sections. These new sections were largely based on Mueller’s informal groups, with habit, leaf shape and pistillate sepals furnishing most of the diagnostic characters.
It would be surprising if the classification of Pax & Hoffmann, based on a very limited number of characters, proved to reflect accurately phylogenetic relationships. As pointed out by Webster & Webster (1972), ontogenetic and

NEOTROPICAL SPECIES OF DALECHAMPIA 139
geographic variation in leaf shape of many species raises serious doubts about the validity of such characters as ‘dimorphic foliage’ for defining species groups. A particularly serious problem has been the neglect of pseudanthial characters by most workers on the genus. This is understandable in view of the fact that most studies have been done on herbarium specimens. The study of Michaelis (1924), although based mostly on herbarium material, partially overcame these handicaps and provided the first significant advance beyond Baillon with improved insights into inflorescence structure and its relation to evolutionary trends.
As a result of over a decade of field, greenhouse, and herbarium observations on neotropical Dalechampia, we have concluded that considerable redefinition of the supraspecific taxa is necessary in order to better reflect natural relationships. Since Dalechampia, because of its production of resins as a floral reward, occupies a very unusual ecological niche among tropical plants (Armbruster, 1984a), we anticipate that it will receive increasing attention in ecological and evolutionary research. The present study is intended to provide the basis for future systematic, ecological and coevolutionary studies by ( 1 ) reviewing the morphological characters that are useful in determining phylogenetic relationship, and (2) providing a detailed synopsis of our revised classification, including keys and descriptions for the sections and subsections, and an enumeration of the species accepted as distinct.
MORPHOLOGICAL CHARACTERS
The only detailed accounts of the morphology of any species of Dalechampia are those of D. scandens by Webster & Webster (1972) and of D. spathulata by Froebe et al. (1983), although rather comprehensive taxonomic descriptions have been given for the species of Panama (Webster, 1968) and the Congo (Ltonard, 1962). Here we wish to provide a brief discussion of the salient characters that appear useful in defining species groups within the genus.
Growth form
The majority of the neotropical and all of the paleotropical species of Dalechampia are twining vines (Fig. 1 ) . Apparently the twining habit is primitive, as it also predominates in the related genera Plukenetia and Tragia. However, a number of neotropical species are not twining, and some of them (e.g. in subsection Caperonioides) are decumbent to erect subshrubs. The species of section Cremophyllum, including the popular ornamental species D . spalhulata, are erect single-stemmed shrubs (“Schopkaiimchen”) of the tropical rainforest understory.
These differences in habit appear to reflect adaptation to niches in various tropical vegetation types. Most climbing species occur in rain forest or moist forest, mainly along forest margins or in light gaps. Shrubby or suffruticose species, with the exception of section Cremophyllum, are in contrast at home in open habitats such as the cerrados or campos rupestres of Brazil. It is notable that all of the species of the most primitive section, Rhopalosblis, are confined to lowland tropical rainforest areas.

140 C. L. WEBSTER AND W. S. ARMBRUSTER
A
Figure 1 . Habit of Dalechampia species. A. D. micrantha. B. D. dioscoreijolia. C. D . spathulata. D. D. femandesii. E. D. stipulacea. F. D. scandens. C. D. caperonioides. H. D. cissijolia. These, and other figures, drawn from living plants in greenhouses at University of California, Davis.

NEOTROPICAL SPECIES OF DALECHAMPZA 141
Leaves
Leaf morphology in Dalechampia displays a protean diversity that in the neotropics is perhaps rivalled only by the genus Passgora. In the primitive section Rhopalost_ylis, the leaves are unlobed and entire with palmate venation. The species of section Cremophyllum are distinctive in having simple elongated pinnately veined leaves. In the remaining groups, leaf shape is highly variable at the individual, population, and species level. In sections Dioscoreifoliae and Dalechampia, there are three different kinds of leaves: unlobed, palmately lobed and palmately parted (compound); however, the three types do not normally coexist within a single species.
As noted by Webster & Webster (1972), interpretation of leaf shape patterns in Dalechampia is complicated by ontogenetic variation. In some species, such as D. scandens, the lower (proximal) leaves of a branch tend to have fewer lobes than those produced later on. In other species, such as D. tiliifolia Lam., the first leaves on a branch may be completely unlobed, followed by leaves with transitional lobing, and with distinctly trilobed leaves at the distal nodes. In a number of species of section Dalechampia subsection Triphyllae, the earlier unlobed leaves are succeeded by trifoliolate leaves, but transitional trilobed leaves are normally lacking.
The evolutionary history of leaf shape in Dalechampia appears to be complex. The transition between lobed and unlobed leaves is apparently so easy to accomplish that reversals or independent origins have probably been common. Within section Dalechampia, for example, i t seems probable that some species with unlobed leaves (e.g. D. convolvuloides) have evolved from ancestors with lobed leaves, whereas other taxa with unlobed leaves (e.g. D. boliviana, D. tenuiramea) have been derived from ancestors with trifoliolate leaves.
Stipules are an important systematic character in defining or recognizing many species of Dalechampia. In many species they are reflexed and persistent a t maturity, whereas in others (e.g. D. micrantha, D. aristolochiifolia, D. tiliifolia) they are deciduous. The stipules in section Cremophyllum are distinctive in becoming indurate and ribbed but not reflexed in age.
Indumentum
Most species of Dalechampia have two very different kinds of trichomes: simple unicellular hairs and specialized stinging hairs. The latter were carefully studied by Knoll (1905), who showed that these remarkable trichomes represent highly modified epidermal cells in which a greatly elongated arm of a druse crystal functions as a sort of hypodermic needle to inject irritant compounds. Although the stinging hairs in Dalechampia are usually smaller and less irritant than those in related genera of tribe Plukenetieae such as Acidoton and Tragia, foliage of some species such as D. jicifolia and D. pernambucensis can be distinctly irritating on contact. These stinging hairs are so distinctive in structure that Knoll appears to have been correct in concluding that they demonstrate a close connection between Dalechampia and Tragia (and related genera).
The pistillate sepals of most species of Dalechampia become armed in fruit with highly irritating trichomes. These are not crystalliferous, however, and these unicellular hairs produce merely mechanical irritation.

I42 G. L. WEBSTER AND W. S. ARMBRUSTER
\ upper invol. bract
adstarnipate brtl.
pistil flower
abstaminate br
C \lower invol. bract D
Figure 2. Morphology of the Da[e~hampiapseudan~hium. A, B. Pseudanthium of D. scandens: lateral and frontal views. C, D. Cross sectional diagrams of the pseudanthium of D. scandens ( C ) and D. aristolochiijolia (D).
A relatively uncommon trichome is that of glandular hairs, which are known mainly from the widespread South American species D. stipulacea. These glandular hairs give a viscid character to both foliage and bracts of this species.
InJo rescence
The inflorescence in Dalechampia is a bilaterally symmetrical bisexual pseudanthium that has a fundamentally similar ground-plan in the great

NEOTROPICAL SPECIES OF D A L E C H A M P Z A 143
Figure 3. Involucral bracts of Dalechampia species (upper bracts). A. D. micrantha. B. D. shankii. C . D . dioscorefolia. D. D. schottii. E . D. scandens. F . D. magnistipulata. G. D . convoluuloides.
majority of species (Fig. 2). The two, usually showy involucral bracts are attached at alternate nodes so closely juxtaposed that on casual inspection they appear to be opposite, but careful examination shows that the bract below the pistillate cymule is slightly lower than the other. Each bract has a pair of stipules clearly homologous to those of the foliage leaves, but the involucral stipules are often distinctively shaped.
The involucral bracts of the Dalechampia pseudanthium show a great range of diversity in size, shape and colour (Fig. 3). In section Rhopalostylis, the involucral bracts are usually less than 1 cm long and the lamina generally resembles the associated stipules. Within the other sections, the involucral bracts are relatively showy and larger, from 1 cm to well over 5 cm in length. Species in the two largest sections (Dioscoreifoliae and Dalechampia) show considerable diversity in colour, from greenish to white, pink, purplish or yellow. Bract shape varies independently of colour, from ovate or oblong and unlobed to deeply 3- or 5-lobed; bract margins may be entire, dentate or lacerate.
In all species of Dalechampia there is a single pistillate cyme inserted above the lower involucral bract and below the staminate flowers. Nearly always, the cyme has three flowers, but in D. Jiczfolia and rarely in a few other species, the number may be reduced to one. The pistillate flowers are subtended by a pistillate involucel of a single lower (proximal) bract and a pair of upper (distal) bractlets; the upper bractlets are very commonly fused into a single compound bractlet, so that the pistillate flowers (e.g. in D. scandens) appear to be enclosed by a 2-lipped involucel.
The staminate subflorescence in Dalechampia is much more variable in structure than the pistillate, and its interpretation has been more controversial. In the familiar species of section Dalechampia such as D. scandens it consists of a

144 G . L. WEBSTER AND W. S. ARMBRUSTER
F
Figure 4. Resiniferous bractlets of Dalechampia species. A. D . shankii B. D . aristolochiifolia. C. D . spathulata. D. D. schottii. E. D. fernandesii. F . D . scandens.
pedunculate 2-lipped staminate involucel with a row of nine or ten staminate flowers facing the pistillate cymule and a sticky resiniferous gland facing the upper involucral bract. In species considered to be less highly evolved, however, such as those in section Dioscoreifoliae, the involucel is composed of two pairs of discrete bracts that appear decussately arranged, although the studies of Froebe et al. (1983) on D. spathulata suggest that the bracts are alternate but approximated by contraction of internodes.
In some of the neotropical species of Dalechampia, the staminate involucel is basically tetramerous, as can be seen in sections Rhopalostylis and Dioscoreifoliae, where the four bracts are more or less separate and decussate. Froebe et al. (1983) interpret the gland of D. spathulata as mainly derived from bractlets of the single upper (proximal) branch. In other neotropical taxa, the staminate pleiochasium has five branches. In sections other than Rhopalostylis, the staminate flowers are suppressed on two of the arms, and the bractlets are resiniferous. The ‘gland’ of the inflorescence of most neotropical species therefore represents the tightly packed modified staminate bractlets of two arms of the staminate pleiochasium. However, there is considerable variation in the number and position of the modified staminate bractlets making up the resiniferous gland; within section Dioscoreifoliae, a progression can be seen from species, such as D. shankii in which the resiniferous bractlets do not form a distinct gland, to D. dioscoreifolia and D. aristolochiifolia in which there is a well-defined resin gland similar in appearance to those in section Dalechampia (Figs 2, 4) . In a few species in section Dalechampia, such as D. brownsbergensis, the resin gland has been secondarily lost.
The resiniferous gland in D. scandens is composed of laminar structures with secretory epithelium (illustrated by Webster & Webster, 1972); other species vary greatly in the number, size and location of the laminae. In section Dioscoreifoliae the resin is produced by lacerate bractlets that evidently belong to the fertile arms as well as one sterile arm of the staminate subflorescence. One can trace among various species a morphological series from this relatively slightly modified arrangement to the extreme laminar structures of section

NEOTROPICAL SPECIES OF DALECHAMPIA I45
Dalechampia (Fig. 4). The glandular structure in section Cremophyllum is distinctive because the laminae end in long finger-like projections, presumably reflecting the production of aromatic compounds rather than resin.
Although Bailion ( 1858) correctly interpreted the resiniferous bractlets as modified bracts of branches of the staminate inflorescence, Urban (1888) claimed that the parts of the resin gland represent modified staminate flowers, but later workers such as Michaelis (1924), Venkata Rao (1971) and Webster & Webster (1972) have upheld the original view of Baillon. There remains, however, some uncertainty about the basic pattern of the staminate subflorescence as a whole. Michaelis (1924) claimed that the primitive condition is found in the Madagascar species D . tamzfolia, in which the staminate pleiochasium is radially symmetrical, with five or six bracts subtending monochasial branches surrounding the single central (and terminal) staminate flower. In the South American species of sections Rhopalosblis, Dioscoreifoliae and Cremophyllum, there are usually only four pleiochasial branches, while 5 branches occur in section Delechampia. At present, i t is difficult to decide whether the primitive state is four branches or five.
Venkata Rao (1971) has attempted to trace the evolution of inflorescence types in the Euphorbiaceae, and suggested that the pseudanthium in Dalechampia can be derived from the inflorescence of Acalypha. There is indeed a general correspondence between the thyrse (‘pseudoraceme’) of Acalypha, with its bracteate proximal pistillate flowers and distal staminate cymules, and the pseudanthium of Dalechampia. In Acalypha the inflorescence ends in a pistillate flower, whereas in Dalechampia the terminal flower is staminate. The analysis by Froebe et al. (1983) describes the inflorescence of Dalechampia spathulata as a ‘complex sciadioid’ with two basal prophylls (involucral bracts), a pistillate cymule, four staminate subflorescences and a terminal staminate flower. Except for the sex of the terminal flower, there is a general correspondence with the Acalypha inflorescence, as suggested by Venkata Rao, although the two genera are so different in many respects that the inflorescence type in Acalypha can scarcely be regarded as directly ancestral to that in Dalechampia.
The terminology applied to the inflorescence of Dalechampia has remained rather imprecise because of the unfamiliarity of most investigators with fresh or preserved material of a wide range of species. In our diagram (Fig. 2) we have indicated the terms used €or parts in a typical inflorescence such as is found in section Dalechampia. Normally, the central (first opening) staminate flower of the staminate involucel represents the apex of the pseudanthial axis, as indicated by Froebe et al. (1983). However, because of the contraction of the inflorescence axes and superimposition of bilateral symmetry, use of the terms “proximal” and
distal” is not very helpful in descriptive terminology. It seems more convenient, and less misleading, to indicate the relative position of parts in the inflorescence by using the qualifying terms abstaminate and adstaminate for parts in the pistillate subflorescence, and adpistillate and abpistillate for parts in the staminate subflorescence. The pistillate flowers, typically in a contracted 3-flowered cyme, are then said to be subtended by an abstaminate bract subtending the central flower, and two adstaminate bractlets (sometimes fused into one) subtending the two lateral flowers. The lip of the staminate involucel adjacent to the upper involucral bract, and its constituent bracts, are said to be abpistillate, and the lip or bracts away from the involucral bract are then adpistillate.
“
146 G . L. WEBSI’ER AND W. S. ARMBRUSTER
Flowers
The staminate flowers of Dalechampia are similar to those found in various genera of Plukenetieae, although they differ from those of Tragia, for example, in lacking a floral disk. The sepals are always valvate, and recurve in a characteristic way at anthesis. Because the pedicel is articulate, flowers may fall off the staminate involucel onto the pistillate cymule below, thus increasing the chances of self-pollination; usually, however, the entire staminate subflorescence also disarticulates. The androecium consists of a variable number of stamens, typically between 10 and 50, with the filaments united into a column. The pollen grains of Dalechampia are very characteristic, as shown by Punt (1962) and Webster & Webster (1972); they are usually large (50 pm in length or more) and ellipsoidal, with long colpi and a very coarse ektexinal reticulum. There is very little difference in pollen morphology between species of different sections, although the grains in section Cremophyllum differ in being more nearly spherical in shape.
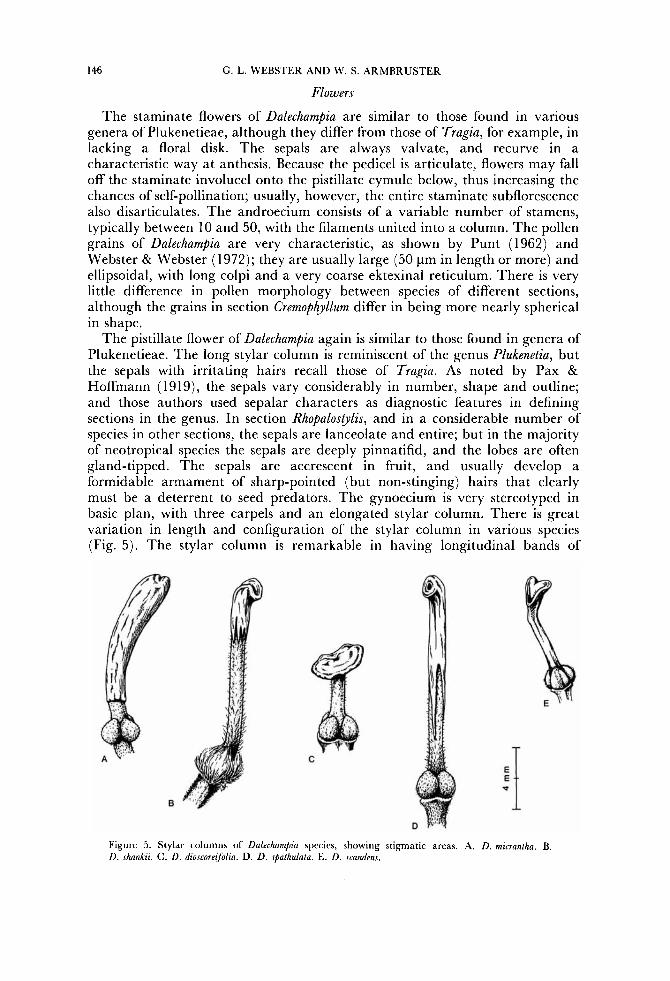
The pistillate flower of Dalechampia again is similar to those found in genera of Plukenetieae. The long stylar column is reminiscent of the genus Plukenetia, but the sepals with irritating hairs recall those of Tragia. As noted by Pax & Hoffmann (1919), the sepals vary considerably in number, shape and outline; and those authors used sepalar characters as diagnostic features in defining sections in the genus. In section Rhopalostylis, and in a considerable number of species in other sections, the sepals are lanceolate and entire; but in the majority of neotropical species the sepals are deeply pinnatifid, and the lobes are often gland-tipped. The sepals are accrescent in fruit, and usually develop a formidable armament of sharp-pointed (but non-stinging) hairs that clearly must be a deterrent to seed predators. The gynoecium is very stereotyped in basic plan, with three carpels and an elongated stylar column. There is great variation in length and configuration of the stylar column in various species (Fig. 5 ) . The stylar column is remarkable in having longitudinal bands of
Figure 5. Stylar columns of Dalechampia species, showing stigmatic areas. A. D. micrantha. B. D. shankii. C. D. dioscoreifolia. D. D. spathulata. E. D . scandens.

NEOTROPICAL SPECIES OF DALECHAMPIA 147
stigmatic surface running halfway or more towards the base. This has led to an unfortunate confusion in the descriptive terminology, since the ‘stigma’ of Mueller, Pax and Hoffmann, and other writers refers to the apex of the stylar column but not to the actual stigmatic surface. To be accurate, therefore, the apparent ‘stigma’ should be referred to as the apex of the stylar column.
Fruits and seeds
The fruits of Dalechampia are always 3-celled capsules and offer little in the way of distinctions except for size. The ecarunculate seeds are subglobose and usually smooth, although a few species have a distinctly roughened seed coat. The pedicels of the pistillate flowers elongate considerably during maturation of the fruit, and fruiting pedicel length is a character of some systematic utility. Dispersal of the seeds occurs as a result of explosive dehiscence of the capsule; Armbruster ( 1982) has discussed the ecological parameters involved in seed dispersal in D. spathulata.
ECOLOGY
The purpose of the present work is to present a systematic review of the neotropical taxa. However, since the peculiar morphology of the Dalechampia inflorescence reflects a long history of evolution by the plant in response to its apoidean pollinators, a brief mention of the functioning of the inflorescence in pollination seems appropriate.
Although visits of bees to Dalechampia were observed in Brazil in the 19th century, little was understood about the pollination until the observations of Armbruster and Webster in Mexico in 1976 (Armbruster & Webster, 1979) and Sazima, Sazima & Carvalho-Okano in Brazil in 1979 (Sazima et al., 1985). Additional observations have been reported from Panama (Armbruster & Herzig, 1984), Ecuador (Armbruster & Webster, 1982), Amazonian Brazil (Armbruster & Webster, 1981) and Costa Rica (Armbruster, 1983, 1984b). It is now apparent that the majority of neotropical species are pollinated by female bees that visit the inflorescences to gather resin from the gland in the staminate subflorescence. As shown by Armbruster (1984a, 1985, 1988b), species with smaller inflorescences are primarily pollinated by megachilid bees, and those with larger inflorescences by euglossines. In addition to the standard measurements of style length and gland size, there are other dimensions of particular interest to pollination ecologists, especially the distance from the gland or stamens to the tips of the stylar columns (Armbruster, 1985, 1988b).
Pollination by fragrance-collecting male euglossine bees (Apidae: Euglossini) is the second most common system of pollination in Dalecharnpia; perhaps a dozen species are pollinated in this way. It is striking that male euglossine pollination is scattered widely throughout the genus, occurring in sections Rhopalostylis (Armbruster et al., 1990), Cremopfyllum (Armbruster & Webster, 1979; Armbruster et al., 1989), and Dalechampia (Webster & Armbruster, 1982; Armbruster et al., 1990). This indicates that male euglossine pollination has originated at least three times within the genus.
One of the striking characters related to pollination is the colour of the involucral bracts. These may lack any distinctive colour in some species (e.g.

148 G. L. WEBSTER AND W. S. ARMBRUSTER
they are deep green in some taxa of subsection Triphyllae), but they are often white or pink, and distinctly yellow bracts occur in some species of section Dalechampia. The reasons for these colour differences are still obscure, although it seems possible that they may function as species recognition signals in regions of sympatry. In most neotropical species, the bracts play a dual role, since they inclose the developing fruit, presumably to deter predation; during fruit maturation, the bright colours are generally lost, and the fruiting bracts turn green.
SYSTEMATIC TREATMENT
Because of our limited knowledge of the neotropical species of Dalechampia, a detailed monograph cannot yet be prepared. However, i t is clear from knowledge at hand that the supraspecific taxa recognized by Pax & Hoffmann (1919) must be drastically modified. The following synoptic treatment is based on field work, greenhouse, laboratory and herbarium studies of the neotropical species. We have examined living plants of about one half of the species. Type specimens of the majority of species have been examined in European herbaria by the senior author, and a large number of lectotypifications have been made as a result. Unless specified to the contrary, lectotypifications in the following synopsis are made here for the first time. Typification of a considerable number of species has been difficult because Mueller and later Pax & Hoffmann cited specimens in Berlin that have since been destroyed. Where possible, lectotypes have been selected from duplicate material in other herbaria, but in a number of instances it has appeared more prudent to postpone selection of lectotypes or neotypes until the taxa are better understood.
We have adopted a conservative policy in enumerating species in this synopsis, i.e. we have given earlier authors the benefit of the doubt and accepted proposed species unless i t is quite clear that they must be reduced to synonymy. This has resulted in the acceptance of a number of names of rather questionable status, especially in section Dalechampia subsections Humiles and Triphyllae. Allem & Waechter (1977) have reduced to synonymy a large number of binomials in those two subsections, and their suggestions need careful appraisal by future workers.
Dalechampia
Dalechampia Linnaeus, Species Plantarum, 1054 ( 1753) ; Genera Plantarum, edition 5: 473 (1754). Subshrubs, shrubs or vines, usually twining; indumentum of simple and
stimulose hairs (sometimes nearly glabrous). Leaves alternate, stipulate, usually petiolate; blades entire or dentate, usually palmately veined, unlobed to lobed or partite. Monoecious; inflorescences pseudanthial, usually bilaterally symmetrical, subtended by a pair of subopposite often showy involucral bracts, axillary or terminating short shoots of 1 to few nodes. Involucral bracts with blade usually much larger than stipules, palmately veined, unlobed to deeply palmatifid, margins entire or dentate. Pistillate flowers 3 in a contracted dichasium inserted above lower involucral bract, subtended by a lower (proximal) abstaminate bract and 1 or 2 sometimes fused upper (distal)

NEOTROPICAL SPECIES OF DALECHAMFZA 149
adstaminate bractlets; staminate pleiochasium terminal but apparently inserted between pistillate cymule and upper involucral bract, mostly with 8-1 2 flowers; involucel of staminate subflorescence of distinct or united bracts; bractlets subtending staminate flowers all or partly resiniferous (or producing aromatic compounds). Staminate flowers pedicellate, pedicel usually articulate above the base; sepals mostly 4 or 5, valvate, recurving at anthesis, entire; stamens 8-100 or more, mostly 20-50, filaments connate, anthers bilocular, longitudinally dehiscen t; pollen grains ellipsoidal, brevi-3-colporate, ektexine with prominent raised reticulum; pistillode absent. Pistillate flowers subsessile or short-pedicellate a t anthesis; sepals 5- 12, entire or pinnatifid, lobes glandular or eglandular; ovary 3-celled, glabrous or pubescent; styles united into an elongated column, sometimes lobed or discoidal at apex; stigmatic surface usually extending down upper one-third to three-quarters of stylar column; ovules anatropous. Fruit capsular, subtended by an accrescent calyx usually armed with stiff hairs; columella persistent; seeds ecarunculate, globose or subglobose, smooth or rugose; endosperm copious; cotyledons flat, broader and longer than the radicle.
TYPE: Dalechampia scandens L. The neotropical species are here assigned to six sections. Apparently all of the
20 paleotropical species can be accommodated in the sections recognized for the neotropics, but the placement of these paleotropical species must await detailed study.
Key to the sections
1. Distal involucral bract small (less than 8 mm long); staminate involucel radially symmetrical, of 4 distinct bracts, these whorled rather than decussate; staminate flowers 7-1 3; stamens 5-15; staminate bractlets not aggregated into a distinct resiniferous gland; vegetative parts generally lacking stinging hairs; leaves unlobed, obtuse to cordate or hastate at base, with arcuate secondary veins usually confined to distal half of midrib.
1. Rhopalostylis Distal involucral bract larger (usually more than 10 mm long); staminate involucel bilaterally or radially symmetrical, bracts distinct or fused; staminate flowers 4-16; stamens usually 12 or more; staminate bractlets usually aggregated into a distinct gland producing resin or aromatic compounds.
2. Staminate involucel of 4 free decussate bracts; bractlets in staminate pleiochasium with deeply laciniate to papillate margins.
3. Vines (except one species a branched shrub); bractlets of staminate pleiochasium deeply laciniate, producing resin; pollen grains prolate; seeds smooth to rugulose.
1.
2. Dioscoreifoliae 3. Monopodial subshrubs (rarely sparsely branched);
bractlets of staminate pleiochasium papillate, producing aromatic compounds rather than resin; pollen grains subglobose; seeds tuberculate.
3. Cremophyllum

150
2. G . L. WEBSTER AND W. S. ARMBRUSTER
Staminate pleiochasium with a two-lipped involucel of 4 or 5 more or less fused bracts; resiniferous bractlets with entire to shallowly laciniate margins.
4. Staminate involucel of 4 or 5 incompletely connate bracts, with 3 adpistillate bracts subtending the staminate flowers opposite the 1 or 2 abpistillate bracts subtending the resiniferous gland; resiniferous bractlets wth pectinate to shallowly laciniate margins.
5. Leaves unlobed, rigid; involucral bracts not over 1 cm long; staminate involucel of 5 bracts; staminate flowers 13-15; anthers glandular. 4. Coriaceae
5. Leaves lobed or unlobed, chartaceous; involucral bracts over I cm long; staminate involucel of 4 bracts; staminate flowers c. 10; anthers not glandular.
4. Staminate involucel bilabiate, the bracts completely connate; resiniferous bractlets of staminate pleiochasium with entire or crenulate margins. 6. Dalechampia
Section 1. Rhopalostylis Pax & Hoffmann in A. Engler (Ed.), Das
TYPE: Dalechampia micrantha Poeppig. SYNONYM: Megalostylis Spencer Moore, Journal of Botany, 54: 208 (1916). TYPE:
Megalostylis poeppigii Spencer Moore ( = Dalechampia micrantha Poeppig) , Twining vines, vegetative parts nearly or quite lacking stinging hairs; leaves
simple, unlobed (at most hastate at base); distal involucral bracts less than 0.5 cm long, usually stipuliform, entire, greenish or whitish; staminate bracts 4, discrete; staminate bractlets 1 to several per arm, sparsely and obscurely resiniferous; staminate flowers 7-13; sepals mostly 3 or 4; stamens 5-15; pistillate flowers with 5 or 6 entire sepals; stylar column usually clavate, not abruptly dilated at apex; cocci of capsule with massive walls; seeds smooth.
1. Dalechampia attenuistylus Armbruster, Brittonia, 41: 44, fig. 1 (1989). TYPE: Venezuela, Bolivar, Santa Elena de Uairkn, Armbruster, Keller @ Matsuki
DISTRIBUTION: rain forests, southern Venezuela.
2. Dalechampia fragrans Armbruster, sp. nov. TYPE: Suriname, Brownsberg, Nature Park, c. km 13 Plateau Weg, 8 xi 1985,
Armbruster @ Herzig 85-101 (MO, holotype; ALA, NY, UVS, isotypes). Frutex volubilis; lamina folii ovata, acuminata, basi truncata ad cordatam;
stipulate deltatae, reflexae, c. 3 mm longae; petiolus 2-6 cm longus; flores staminati 12-13; stamina 4-6; sepala pistillata 3-4 mm longa, integra; stylus clavatus, 7-10 mm longus, 2.5-4.5 mm latus. DISTRIBUTION: rain forests, Suriname and French Guiana. NOTE: related to D. micrantha Poeppig, but differing in its slightly to strongly
5. Tiliifoliae
Pjanzenreich, ZV, 147. XII (Hejit 68) : 7 ( 19 19).
87-153 (MO, holotype; ALA, MY, NY, VEN, isotypes).
cordate leaf bases and its stouter stylar column.

NEOTROPI CAL SPECIES OF DALECHAMPIA 151
3. Dalechampia hastata Webster, sp. nov. TYPE: Brazil, Amazonas, Manaus, terra firme, km 9 da BR-17, 16 iii 1955,
Chagas INPA 1886 (MG, holotype; INPA, isotype). Frutex volubilis; lamina folii lanceolata, 5-8 x 1-2 cm, acuminata, ad basin
hastatum (lobis 5-20 mm longis); stipulae lineari-lanceolatae, reflexae; petiolus 0.5-1.5 cm longus; flores staminati 9; stamina 9-12; sepala pistillata c. 4 mm longa, integra; stylus 5 mm longus, clavatus, 1.5-2.0 mm latus; capsula c. 9 mm diametro; semina subglobosa, 3.6-3.8 mm longa, laevia.
Climbing vine; leaves lanceolate, acuminate, hastate at base (lobes 5-20 mm long), 5-8 x 1-2 cm; stipules linear-lanceolate, reflexed; petioles 5-15 mm long; pseudanthium with 9 staminate flowers; stamens 9-12 per flower; pistillate sepals c. 4 mm long, entire; style 5 mm long, clavate, 1.2-2.0 mm thick; capsule G. 9 mm in diameter; seeds subglobose, 3.6-3.8 mm long, smooth. DISTRIBUTION: rain forests, vicinity of Manaus, Amazonas, Brazil. NOTE: related to D. attenuistylus Armbruster, but differing in its hastate leaves
with shorter petioles and its differently shaped stylar column.
4. Dalechampia liesneri Huft, Annals o f the Missouri Botanical Garden, 76: 1078
TYPE: Venezuela, Amazonas, Atures, 0.5-2 km E of Rio Coro-Coro, Liesner @
DISTRIBUTION: rain forests, southern Venezuela.
(1989).
Holst 21244 (MO, holotype; U, VEN, isotypes).
5. Dalechampia micrantha Poeppig, Nova Genera ac Species Plantarum, 3: 19, pl.
TYPE: Brazil, Amazonas, Ega (Teft.), Poeppig 2807 (W, holotype; K, isotype) . SYNONYM: Megalostylis poeppigii Spencer Moore, Journal of Botany, 54: 208 ( 19 16).
TYPE: Brazil, Poeppig (BM, holotype, probably part of the type collection of D. micrantha) .
222 (1841).
DISTRIBUTION: Amazonian forests, Peru, Venezuela and Brazil (Amazonas).
6. Dalechampia olympiana Kuhlmann & Rodrigues, Amazonia, Botanica, 5: 1
TYPE: Brazil, Amazonas, Manaus, Rodrigues 225 (INPA, holotype) . DISTRIBUTION: endemic to Amazonas. Brazil.
( 1954).
7 . Dalechampia parvibracteata Lanjouw, Recueit des travaux botanigues
TYPE: Guyana, Upper Demarara River, Jenman 4088 (K, holotype). DISTRIBUTION: lowland rain forests, eastern Venezuela, Guiana, Suriname and
Nlerlandais, 31: 463 ( 1934).
northern Brazil (Amapi, ParA).
Section 2. Dioscoreifoliae Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Dalechampia dioscorezfolia Poeppig. SYNONYM: Dalechampia section Syluaticae Pax & Hoffman, in A. Engler (Ed.), Das
Pjanzenreich, IV. 147. 12 (Hef t 68): 3 (19 19). LECTOTYPE: Dalechampia sylvestris Spencer Moore (chosen here).
PJEanzenreich, IV. 147. X I I (He f t 68) : 45 ( 19 19).

152 G. L. WEBSTER AND W. S. ARMBRUSTER
Twining or clambering vines (rarely erect subshrubs); vegetative parts with stinging hairs; leaves simple, lobed or palmately compound; floriferous shoot terminal, or axillary with 1-3 nodes below peduncle; involucral bracts unlobed or 3-5-lobed, denticulate to dentate or lacerate, whitish or greenish to pink or purplish, sessile or unguiculate; staminate involucel of 4 free bracts, these mostly decussate, rigid and concave; staminate bractlets lanceolate to obovate, distally blunt or lacerate, slightly to copiously resiniferous; staminate flowers 6-10 (-1 3); staminate sepals 3-5, stamens 7-70; pistillate involucel of 1 lower (proximal) usually eglandular abstaminate bract and 1 or 2 small (or obsolete) upper (distal) adstaminate bractlets; pistillate sepals 5-1 2, entire or pinnatifid, not glandular; stylar column sometimes clavate or apically dilated; seeds often angled or rugulose.
The species of this section with compound leaves were referred by Pax & Hoffmann to a separate section Sylvaticae, but they agree in reproductive characters with the species with entire leaves, and it does not seem appropriate to recognize two different sections for these relatively primitive species with discrete staminate bracts and lacerate resiniferous bractlets.
With 25 neotropical species, section Dioscoreifoliae is the largest section after section Dalechampia, and there is striking diversity in leaf and inflorescence morphology. Eventually it may be possible to define formal subsections, but at present it seems best to list the species under five informal groups.
Group 1. Leaves palmately divided; stylar column cylindrical, usually without a markedly dilated tip; pistillate sepals 5 or 6, entire, bifid, or pinnatifid; seeds round and smooth to slightly roughened.
8. Dalechampia alata Mueller Argoviensis, Linnaea, 34: 220 ( 1865). TYPE: Brazil, Rio de Janeiro, Tocaia, Schott 4198 (W, lectotype; erroneously cited
DISTRIBUTION: coastal rain forests, eastern Brazil (Bahia to Rio de Janeiro). by Mueller as Pohl 4198).
9. Dalechampia juruana Ule, Verhandlungen des Botanischen Vereins fur die Provinz
T Y P E : Brazil, Amazonas, lower Rio Juruii, Ule 5256 (holotype at B destroyed;
DISTRIBUTION: rain forests, Amazonian Colombia, Venezuela, Peru, and Brazil
Brandenburg, 50: 83 ( 1909).
photograph, MICH).
(Amazonas).
10. Dalechampia luetxelburgii Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Brazil, Bahia, Sgo Bento das Lages, von Lutselburg 132; Paraiba, Serra do
DISTRIBUTION: forests of north-eastern Brazil (CearA to Bahia) .
Pflansenreich, IV. 147. XVII (Hgt 85): 188 (1924).
Araripe, von Lutzelburg 12682 (not seen; syntypes at B destroyed).
11. Dalechampia papillistigma Armbruster, Brittonia, 41: 49, fig. 3 (1989). TYPE: Venezuela, Bolivar, Santa Elena de UairCn, Armbruster, Keller & Matsuki
DISTRIBUTION: Amazonian rain forests, south-eastern Venezuela. 87-155 (MO, holotype; ALA, MY, NY, VEN, isotypes).

NEOTROPICAL SPECIES OF DALECHAMPIA 153
12. Dalechampia peckoltiana Mueller Argoviensis, in Martius, Flora
TYPE: Brazil, Rio de Janeiro, Canta Gallo, Peckolt 93 (M, holotype). Perhaps
DISTRIBUTION: apparently restricted to the state of Rio de Janeiro, Brazil.
Brasiliensis, 11(2): 647 (1874).
merely an aberrant form of D. pentaphylla.
13. Dalechampia pentaphylla Lamarck, Enylopkdie Mdhodique, Botanique, 2: 258 (17861.
TYPE: Brazil, probably near Rio de Janeiro, 1790, Vandelli (Herb. Jussieu, P,
DISTRIBUTION: tropical and subtropical evergreen and semi-deciduous forests, holotype) .
Minas Geraes and Bahia to Parana and SHo Paulo.
14. Dalechampia psilogyne Mueller Argoviensis, in Martius, Flora Brasiliensis, 11(2): 648 (1874).
TYPE’: Brazil, Goias,’ Gardner 3447 (G, holotype; BM, K, isotypes). DISTRIBUTION: known only from cerrado, Goias, Brazil.
15. Dalechampia subintegra (Mueller Argoviensis) Mueller Argoviensis, in Martius, Flora Brasiliensis, 11(2): 650 (1874).
BASIONYM: Dalechampia triphylla variety subintegra Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1240 ( 1866).
TYPE: Bahia, Blanchet (G, holotype) . DISTRIBUTION: forests, Bahia and perhaps Minas Geraes, Brazil.
16. Dalechampia sylvestris Spencer Moore, Transactions of the Linnean Society
TYPE: Brazil, Mato Grosso, between Santa Cruz and TapirapuH, Spencer Moore
DISTRIBUTION: seasonal forests, Bolivia and Paraguay to western Brazil (Mato
of London, Botany, series 4, 2: 467 (1895).
376 (BM, holotype).
Grosso, Mato Grosso do Sul).
17. Dalechampia violacea Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Brazil, Mato Grosso do Sul, Corumba, Malme 2753 (B, holotype,
DISTRIBUTION: seasonal forests, Mato Grosso do Sul, Brazil.
18. Dalechampia websteri Armbruster, Systematic Botany, 9: 272 (1984). TYPE: Costa Rica, Heredia, Finca La Selva, Armbruster €9 Herzig 79-207 (DAV,
DISTRIBUTION: rain forests, Atlantic lowlands and slopes, western Panama to
Pjanzenreich, ZV, 147. XVZZ. (Hejit 85): 189 (1924).
destroyed; S, lecotype).
holotype; ALA, CR, MO, NY, isotypes).
Costa Rica and Nicaragua.
Group 2. Leaves unlobed; stylar column slender, clavate, apex not dilated; pistillate sepals entire or bifid.
19. Dalechampia arciana Baillon, Adansonia, 5 3 14 ( 1865). TYPE: Brazil, Bahia, Jacobina, Pouqo d’Areia, Blanchet 3884 (P, lectotype; G, UC,
isotypes) .

154 G. L. WEBSTER AND W. S. ARMBRUSTER
SYNONYM: Dalechampia schizoloba Mueller Argoviensis, Linnaea, 34: 22 1 (1865).
DISTRIBUTION: endemic to seasonal thickets and woodlands, Bahia, Brazil. TYPE: Brazil, Bahia, Blanchet 132 (G, lectotype).
20. Dalechampia megacarpa Armbruster, Brittonia, 41: 47, fig. 2 (1989). TYPE: Venezuela, Bolivar, San Ignacio, Armbruster, Herag €9 Otero 85-115 (MO,
holotype; ALA, MY, NY, VEN, isotypes). DISTRIBUTION: rain forests, southern Venezuela.
Group 3. Leaves lobed or unlobed; stylar column clavate, apex not greatly expanded; pistillate sepals 5 or 6, pinnatifid; seeds generally round, smooth to roughened.
21. Dalechampia canescens H.B.K., Nova Genera et Species Plantarum, 2: 98 (181 7). 21a. Dalechampia canescens subspecies canescens
TYPE: Colombia, Mariquita, Bonpland (P, holotype) . SYNONYM: Dalechampia smilacina H.B.K., op. cit. 99. TYPE: Venezuela, “in ripa
fluminis Orinoci, Carichanam inter et cataractam Aturensium”, Bonpland (P, holotype) . Kunth, in the original description, questioned the accuracy of Bonpland’s locality designation, and indeed D. canescens is not known to occur in the Orinoco valley area.
DISTRIBUTION: rain forests, Colombia and Venezuela.
21b. Dalechampia canescens subspecies friedrichsthalii (Mueller Argoviensis) Webster & Huft, Annals o f the Missouri Botanical Garden, 75: 1109 (1988).
BASIONYM: Dalechampia friedrichsthalii Mueller Argoviensis, Flora, 55: 45 ( 1872). TYPE: Nicaragua (incorrectly cited as “Guatemala”), Rio San Juan, Friedrichsthal
DISTRIBUTION: rain forests, Nicaragua to northwestern Colombia (Chocb) . 683 (G, holotype).
22. Dalechampia osana Armbruster, Systematic Botany, 13: 303 (1988). TYPE: Costa Rica, Puntarenas, Osa Peninsula, Burger €9 Liesner 7278 (F, holotype;
DISTRIBUTION: rain forests, Costa Rica. CR, isotype).
23. Dalechampia shankii (Molina) Huft, Annals o f the Missouri Botanical Garden,
TYPE: Costa Rica, Limbn, Rio Reventazbn, Shank tY Molina 4427 (F-1563051,
BASIONYM: Tragia shankii Molina, Ceiba, 11: 68 (1965). DISTRIBUTION: rain forests, Atlantic lowlands, Nicaragua to Panam6.
71: 341 (1984).
holotype).
24. Dalechampia uleana Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Brazil, Amazonas, Rio Acre, Nova York, Ule 9541 (B, holotype, destroyed;
DISTRIBUTION: rain forests, Amazonian Peru, Bolivia and Brazil (Amazonas).
Pjanrenreich, IF’. 147. XII. (HeJt 68) : 3 1 ( 19 19).
G, lectotype; MG, US, isotypes).

NEOTROPICAL SPECIES OF DALECH'4MPIA 155
Group 4. Leaves lobed or unlobed; stylar column apically dilated; pistillate sepals mostly 8-12, pinnatifid.
25. Dalechampia albibracteosa Rusby, Memoirs of the New York Botanical
TYPE: Bolivia, El Beni, Rurrenabaque, Cardenas 1793 (NY, holotype; MICH,
DISTRIBUTION: rain forests, Bolivia.
Garden, 7: 287 (1927).
isotype) .
26. Dalechampia aristolochiifolia H.B. K., Nova Genera et Species Plantarum, 2:
TYPE: Peru, Bracamoros, San Felipe, Humboldt DISTRIBUTION: seasonal forests, Peru.
99 (1817). Bonpland (P, holotype).
27. Dalechampia cujabensis Mueller Argoviensis, Linnaea, 34: 222 (1865). TYPE: Brazil, Mato Grosso, CuiabA, Manso in Herb. Martius 845 (G, lectotype; W,
DISTRIBUTION: cerrado and cerradgo, Mato Grosso, Brazil. isotype) .
28. Dalechampia dioscoreifolia Poeppig, in Poeppig & Endlicher, Nova
TYPE: Peru, Maynas, Poeppig 2163 (B, holotype, destroyed; W, lectotype). SYNONYM: Dalechampia dioscorezfolia variety pubescens Mueller Argoviensis, in De
Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1248 ( 1866). TYPE: Peru, Tarapoto, Spruce 4476 (K, lectotype). Field observations by the junior author indicate that this entity may be recognizable as a distinct taxon.
DISTRIBUTION: rain forests, Nicaragua south to Bolivia and western Brazil (Amazonas).
Genera ac Species Plantarum, 3: 20 ( 1841).
29. Dalechampia hutchisoniana Webster, Annals o f the Missouri Botanical
TYPE: Peru, Celendin, Cajamarca, Hutchison G' Wright 5446 (MO, holotype;
DISTRIBUTION: Andean xeric scrub, Peru.
Garden, 54: 193 (1967, as D. hutchinsoniana).
DAV, UC, isotypes).
30. Dalechampia kavsteniana Pax & Hoffmann, in Engler (Ed.), Das Pjanzenreich, ZV. 147. XZZ ( H ~ t 68) : 30 ( 19 19).
TYPE: Colombia, Bogota, l r iana 3557 (P, lectotype). DISTRIBUTION: forests, northern Colombia.
Group 5. Staminate bracts thin, not clearly decussate; involucral bracts less than 1 cm long; staminate flowers 4; seeds angled, rugulose.
31. Dalechampia schottii Greenman, Publications o f the Field Columbian Museum,
TYPE: Mexico, YucatAn, Mkrida, Schott 534, 956 (F, syntypes). SYNONYM: Dalechampia schottii variety trijoliolata Greenman, loc. cit. TYPE: Mexico,
Yucathn, Shichankanab, Gaumer 1430, 1512 (F, syntypes).
Botanical Series, 2: 255 (1907).

156 G . L. WEBSTER AND W. S. ARMBRUSTER
DISTRIBUTION: lowland seasonal forests, Mexico (Campeche, Quintana Roo, Yucatan).
NOTE: Dalechampia schottii is a very isolated species, and it is not certain that it can be accommodated within section Dioscorezfoliae. I t is provisionally located here pending further study.
Section 3. Cremophyllum (Scheidweiler) Baillon, Etude du Groupe des Euphorbiacbes: 487 ( 1858).
BASIONYM: Cremophyllum Scheidweiler, Bulletin de l'dcadimie Royale des Sciences et Belles-lettres de Bruxelles, 9(1) : 23 ( 1842).
TYPE: Cremophyllum spathulatum Scheidweiler = Dalechampia spathulata (Scheidweiler) Baillon. Erect shrubs with unbranched or sparsely branching stems; indumentum
sparse, with some stinging hairs on young parts; leaves unlobed, elongated, entire or dentate, pinnately veined; stipules persistent, not recurved; pseudanthia strictly axillary; involucral bracts unlobed, usually pink or yellow at anthesis, cordate at base, persisting in fruit; staminate involucel bilaterally symmetrical, of 4 bracts; staminate bractlets laminar with terminal cylindrical projections producing aromatic compounds; staminate flowers 6- 12; stamens 10-30; pistillate flowers with 5 or 6 unlobed sepals, margins irregularly glandular- dentate; stylar column slender, scarcely clavate, not dilated at tip; seeds tuberculate.
32. Dalechampia brevipedunculata Ule, Verhandlungen des Botanischen Vereins
TYPE: Brazil, Amazonas, Rio Juruh, Marary, Ule 5257 (B, holotype, destroyed;
DISTRIBUTION: rain forests, Amazonian Brazil.
fur die Provinz Brandenburg, 50: 82 ( 1909).
G, lectotype).
33. Dalechampia magnoliaj'olia Mueller Argoviensis, Linnaea, 34: 2 19 ( 1865). TYPE: Brazil, Para, Martius (B, holotype, destroyed; M, lectotype). SYNONYM: Dalechampia roezliana variety amazonica Ule, Verhandlungen des Botanischen
Vereins fur die Provinz Brandenburg, 50: 82 ( 1909). Dalechampia spathulata var. amazonica (Ule) Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, IV. 147 XI1 (He) 68) : 1 1 ( 19 19). Dalechampia amazonica (Ule) Macbride, Candollea, 8: 26 (1940). TYPE: Brazil, Amazonas, Jurua Miry, Ule 5865 (G, lectotype).
DISTRIBUTION: Amazonian rain forests, Peru (Loreto, Madre de Dios, San Martin), Venezuela (Amazonas, Bolivar), and Brazil (Amazonas, Amaph, Para, Mato Grosso).
34. Dalechampia spathulata (Scheidweiler) Baillon, Etude GinLrale du Groupe des Euphorbiacbes: 487 ( 1858).
BASIONYM: Cremophyllum spathulatum Scheidweiler, Bulletin de l'dcadhnie Royale des Sciences et Belles-lettres de Bruxelles, 9(1) : 23 ( 1842).
TYPE: Mexico, without locality, Scheidweiler (described from living plants in cultivation, specimens apparently not preserved; neotype here selected as the specimen cited by Baillon: Herb. Houllet, P).
SYNONYM: Dalechampia roezliana Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1233 ( 1866). TYPE (variety rosea

NEOTROPICAL SPECIES OF DALECHAMPIA 157
Mueller Argoviensis) : Mexico, Veracruz, Sontecomapan, Roezl (G, holotype) . Dalechampia roezliana variety viridis Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2): 1233 ( 1866). TYPE: cultivated, pan Houtte (G, holotype). Dalechampia roezliana variety alba Nicholson, Dictionary of Gardening, 1: 439 (1885). TYPE: cultivated, Nicholson (specimen perhaps not preserved).
DISTRIBUTION: Atlantic rain forests, Mexico (Veracruz) and Belize south to Costa Rica.
Section 4. Coriaceae Pax & Hoffmann, in A. Engler (Ed.), Das PJanzenreich, ZV. 147. XZZ (Heft 68): 55 (1919).
TYPE: Dalechampia coriacea Klotzsch ex Mueller Argoviensis Twining vines with thick woody stems; leaves unlobed, coriaceous, pinnately
veined (or somewhat triplinerved); involucral bracts unlobed, entire, greenish, unguiculate; staminate involucel 2-lipped, of 5 slightly to distinctly connate bracts, with embedded glands; resiniferous bractlets laminar, apically pectinate; staminate flowers 12-15; stamens mostly 25-100; pistillate flowers with 10-12 pinnately lobed sepals; stylar column apically dilated.
This monotypic section was recognized by Pax & Hoffmann almost entirely on the basis of the unlobed coriaceous leaves. Except for the pectinate resiniferous bractlets, the plant resembles species of section Dalechampia (subsection Convolvuloides) in the same area of northeastern Brazil.
35. Dalechampia coriacea Klotzsch ex Mueller Argoviensis, Linnaea, 35: 223
TYPE: Brazil, without locality (but evidently Bahia), Sellow (B, holotype,
DISTRIBUTION: seasonal woodlands, northern Bahia, Brazil.
(1865).
destroyed; G, lectotype; P, W, isolectotypes).
Section 5. Tiliifoliae Webster & Armbruster, Systematic Botany, 13: 310 (1988). TYPE: Dalechampia tiliifolia Lamarck.
Twining vines; leaves unlobed to palmately lobed; floriferous shoot usually with two nodes below the peduncle; involucral bracts unlobed to deeply 3-lobed, whitish or greenish; staminate involucel 2-lipped, with 3 adaxial and 1 abaxial bracts more or less connate at base; staminate bractlets more or less laminar, distally toothed or lacerate, distinctly resiniferous; staminate flowers 8- 12; sepals 4-6; stamens 15-35; pistillate flowers with 9-12 pinnatifid sepals; stylar column apically dilated; seeds smooth or reticulate.
This small mainly South American section is transitional betwen section Dioscoreifliae and section Dalechampia, but appears closer to the latter.
36. Dalechampia aflnis Mueller Argoviensis, Linnaea, 34: 223 ( 1865). TYPE: French Guyana, La Mana, 1856, Sagot (G, lectotype). DISTRIBUTION: rain forests, southern Venezuela and the Guyanas to Brazil
(AmapS, Amazonas, Par& Mato Grosso, MaranhBo, Cear6) .
37. Dalechampia fernandesii Webster, Brittonia, 41: 1, fig. 1 (1989). TYPE: Brazil, Cear6, Chapada da Ibiapaba, Webster, Fernandes -3 Matos 25598
DISTRIBUTION: montane thickets, western CearS, Brazil. (EAC, holotype; DAV, NY, R, isotypes).

158 G. L. WEBSTER AND W. S. ARMBRUSTER
38. Dalechampia ilheotica Wawra, Oesterreichische Botanische Zeitschrift, 13: 222
TYPE: Brazil, Bahia, Ilheus, Wawra €9 Maly 365 (W, holotype). SYNONYM: Dalechampia bahiensis Mueller Argoviensis, in De Candolle, Prodromus
Systematis Universalis Regni Vegetabilis, 15(2): 1247 ( 1866). TYPE: Brazil, Bahia, Blanchet 128 (G, lectotype) . Although Mueller himself reduced his D. bahiensis to synonymy under D. ilheotica, the Bahia populations form a systematically difficult complex, and the status of D. bahiensis still requires further investigation.
(1863).
DISTRIBUTION: coastal rain forests and restingas, Pernambuco to Bahia, Brazil.
39. Dalechampia tiliifolia Lamarck, Encyclopddie MLthodique, Botanique, 2: 257
TYPE: without definite locality (possibly Peru), Herb. Jussieu (P, holotype). SYNONYMS: Dalechampia colorata Linnaeus filius, Supplementum Plantarum: 42 1
(1 781). TYPE: Nova Granada, Mutis (US, isotype) . Dalechampia peruviana Lamarck, loc. cit. TYPE: Peru, Joseph Jussieu (P, holotype) . Dalechampia t i l i i f l ia variety peruviana (Lamarck) 0. Kuntze, Revisio Generum Plantarum, I : 596 (1891). Dalechampia heterophylla Vahl, Eclogue Americanae, 3 44 ( 1807). Dalechampia ~ ~ l i ~ o l ~ a variety heterophylla (Vahl) 0. Kuntze, Revisio Generum Plan~arum, I : 596 (1891). TYPE: Cayenne, van Rohr (C, holotype, not seen). Dalechampia pruriens Grisebach, Flora of the British West Indian Islands: 5 1 (1 859). TYPE: Trinidad, Crueger (GOET, holotype) .
DISTRIBUTION: rain forests and seasonal forests, southern Mexico to Bolivia and Brazil.
( 1 786).
Section 6. Dalechampia TYPE: Dalechampia scandens Linnaeus
Subshrubs or vines; vegetative parts usually with stinging hairs; leaves simple to lobed or palmately parted; floriferous shoot usually with a single node below the inflorescence; involucral bracts unlobed to 3- or 5-lobed, usually dentate, white, yellow, or pink, usually persistent in fruit; staminate involucel distinctly 2-lipped, bracts definitely connate; staminate bractlets laminar, entire or obscurely crenate at apex, resiniferous (rarely abortive); staminate flowers 7- 15; stamen 15-100; pistillate flowers wth 5-12 entire to pinnatifid sepals; stylar column sometimes clavate or apically dilated; seeds mostly globose and smooth.
About half of the species of Dalechampia, including most of those in the Old World, belong to this section. A number of the species groups that were treated by Pax & Hoffmann (1919) as distinct sections are here recognized as subsections of section Dalechampia.
Kty to the subsections
A. Twining vines; inflorescences axillary; involucral bracts white, yellow, or green.
B. Leaves on flowering branches lobed or unlobed but never compound.

NEOTROPICAL SPECIES OF DALECHAMPIA 159
C. Leaves on flowering branches lobed (at least in part); involucral bracts white or greenish. 6a. Dalechampia
C. Leaves on flowering branches all unlobed; bracts green or yellowish. 6b. Convolvuloides
B. Leaves on flowering branches 3-5-foliolate (at least in part); bracts green. 6c. Triphyllae
Erect or decumbent subshrubs; inflorescences mostly terminal; involucral bracts white, pink, or yellow.
Leaves lobed or unlobed; staminate flowers 8-1 1.
Leaves palmately parted; staminate flowers 7.
A.
D.
D. 6d. Humiles
6e. Brevipedes
Subsection 6a. Dalechampia TYPE: Dalechampia scandens Linnaeus. SYNONYM: Dalechampia section Scandentes Pax & Hoffmann, in A. Engler (Ed.),
Das Pjanzenreich, IV. 147. XII ( H g t 68) : 27 ( 19 19). Twining vines; indumentum with copious stinging hairs; leaves mainly
3-lobed (occasionally unlobed as well), palmately veined; involucral bracts whitish to yellowish-green, 3-lobed; staminate involucel with 7- 15 flowers; stamens (5-) 10-100 (-200); pistillate sepals pinnatifid; stylar column clavate or distinctly dilated at apex; seeds mostly smooth.
40. Dalechampia armbrusteri Webster, Brittonia, 41: 3 ( 1989). TYPE: Brazil, Bahia, grounds of CEPLAC, Webster €8 Armbruster 250UU (CEPEC,
DISTRIBUTION: coastal rain forests, Bahia and Espirito Santo, Brazil. holotype; DAV, GH, MO, NY, SP, UEC, isotypes).
41. Dalechampia brasiliensis Lamarck, Encyclopidie Mithodique, Bolanique, 2: 258 ( 1786). Dalechampia scandens variety brasiliensis (Lamarck) Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2): 1244 (1866).
TYPE: Brazil, Rio de Janeiro, Dombey (P, lectotype; NY, isotype). SYNONYMS: Dalechampia papposa Endlicher, Atakta Botanica: 22, pls. 20, 2 1 ( 1834).
TYPE: Brazil, Bahia, 1840, Blanchet (W, holotype). Dalechampia brasiliensis variety viridis Mueller Argoviensis, in Martius, Flora Brasiliensis, lZ(2): 658 (1874). TYPE: Brazil, Bahia, Sulzman 504 (G, lectotype).
DISTRIBUTION: evergreen and seasonal forests and capoeiras, Pernambuco to Minas Geraes and Rio de Janeiro, Brazil.
42. Dalechampia brownsbergensis Webster & Armbruster, Systematic Botany,
TYPE: Suriname, Brokopondo, Brownsberg, Webster & Armbruster 24124 (DAV,
DISTRIBUTION: rain forests, eastern Venezuela (Bolivar, Delta Amacuro) to
7: 484 (1982).
holotype; ALA, BBS, GH, MO, NY, isotypes).
Suriname and perhaps Paraiba, Brazil.

160 G. L. WEBSTER AND W. S. ARMBRUSTER
43. Dalechampia jicifolia Lamarck, Encylope‘die Me‘thodique, Botanique, 2: 258
TYPE: Brazil, Dombey (P, holotype). SYNONYM: Dalechampia tiliifolia variety f i c i f l i a (Lamarck) 0. Kuntze, Revisio
Generum PLantarum, 1: 596 (1891). DISTRIBUTION: rain forests, Bahia to Rio de Janeiro, Brazil.
( 1786).
44. Dalechampia grandilla Baillon, Adansonia, 5: 3 12 ( 1865). TYPE: Brazil, Rio de Janeiro, Uba, St. Hilaire A’ 507 (P, holotype). DISTRIBUTION: rain forests, Rio de Janeiro, Brazil.
45. Dalechampia herxogiana Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Bolivia, Santa Cruz, between Rio Pirai and Rio Cuchi, Herzog 1455 (G,
DISTRIBUTION: seasonal forests, Bolivia and western Brazil (Mato Grosso do Sul).
46. Dalechampia magnistipulata Webster & Armbruster, Brittonia, 31: 352 (1979).
TYPE: Mexco, Veracruz, “Los Tuxtlas”, Webster & Armbruster 20986 (DAV, holotype; ALA, GH, MEXU, MO, NY, isotypes).
DISTRIBUTION: rain forests, southern Mexico (Veracruz) .
Pjansenreich, IV. 147. XII (HeJt 68): 36 (1919).
lectotype). Perhaps a form of D. scandens.
47. Dalechampia pernambucensis Baillon, Adansonia, 5 3 1 1 ( 1865). TYPE: Brazil, Pernambuco, Gardner 1130 (G, holotype; BM, K, isotypes). SYNONYM: Dalechampia scandens variety pernambucensis (Baillon) Pax & Hoffmann,
DISTRIBUTION: seasonal woodlands and caatinga, CearA to Pernambuco, in A. Engler (Ed.), Das Pjanzenreich, IV. 147. XII (Hef t 68): 34 (1919).
northeastern Brazil.
48. Dalechampia riparia Smith & Downs, SeLLowia, 11: 153 (1959). TYPE: Brazil, Santa Catarina, Ibirama, Reitz & Klein 3834 (US 2249811,
holotype). Reduced to a synonym of D. stipulacea by Allem & Waechter (1977); however, although doubtless close to that species, the Santa Catarina plant is distinctive, and needs to be further investigated.
DISTRIBUTION: subtropical forests, Santa Catarina, Brazil.
49. Dalechampia scandens Linnaeus, Species Plantarum: 1054 ( 1753). TYPE: West Indies, illustrated by Plumier, Plantarum Americanarum fasciculus, 5: pl.
101 (original plate at P).
49a. Dalechampia scandens variety scandens SYNONYMS: Dalechampia latifoLia Lamarck, Encyclope‘die Me‘thodique, Botanique, 2: 257
( 1 786). Dalechampia scandens forma LatifoLia (Lamarck) Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1244 (1866). TYPE: Antilles, Surian (Herb. Jussieu, P, holotype). It seems possible that the Surian collection is the base of Plumier’s plate, and thus D. Latzfolia is an exact synonym of D. scandens.
Dalechampia villosa Lamarck, Encyclope‘die Me‘thodique, Botanique, 2: 257 ( 1786).

NEOTROPICAL SPECIES OF DALECHAMPZA 161
TYPE: cult. Hort. Trianon, 1764, Commerson (Herb. Jussieu 16295, P, holotype) .
Dalechampia mollis Vahl, Eclogae Americanae, 3: 44 ( 1807). Dalechampia scandens variety molis (Vahl) Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15 (2 ) : 1244 (1866). TYPE: Colombia, Santa Marta, uon Rohr (C, holotype, not seen).
Dalechampia jimbriata H.B.K., Noua Genera et Species Plantarum, 2: 100 ( 18 1 7). Dalechampia scandens variety jimbriata (H.B.K.) Mueller Argoviensis, in De Candolle, Prodromus Qstematis Universalis Regni Vegetabilis, 15 (2 ) : 1244 ( 1866). TYPE: Mexico, Campeche, Humboldt €3 Bonpland (P, holotype).
Dalechampia mollis H.B.K., Nova Genera et Species Plantarum, 2: 101 (1817); non D. mollis Vahl, 1807. TYPE: Colombia, Mariquita, Humboldt €5' Bonpland (P, holotype) .
Dalechampia ruboides H.B.K., Noua Genera et Species Plantarum, 2: 102 ( 18 17). Dalechampia rubformis Sprengel, Systema Vegetabilium, 3: 86 ( 1826). TYPE: Venezuela, Orinoco, Angostura, Humboldt G3 Bonpland (P, holotype) .
Dalechampia sidaefolia H.B.K., Noua Genera et Species Plantarum, 2: 200 (1817). TYPE: Venezuela, Orinoco, Pararuma Island and Angostura, Humboldt G3 Bonpland (P, syntypes) .
Dalechampia breuipes Briquet, Annuaire du Conservatoire et Jardin Botaniques Genive, 4: 609 (1900). TYPE: Colombia, Guanaguana, Funck 64 (G, holotype).
Dalechampia passflora Chodat and Hassler, Bulletin de I'Herbier Boissier, se'rie 2, 5: 609 (1905). TYPE: Paraguay, Chaco-y Island, near Concepcih, Hassler 7231 (G, holotype; S, isotype).
DISTRIBUTION: various plant communities, usually secondary, northern Mexico (Sinaloa, Tamaulipas) south through Central America and the West Indies to subtropical South America (Argentina, Paraguay, Uruguay, and southern Brazil).
49b. Dalechampia scandens variety falkax Mueller Argoviensis, in Martius, Flora Brasiliensis, l l (2 j : 658 (1874).
TYPE: Brazil, Para, Santarem, Spruce 199 (G, holotype). DISTRIBUTION: rain forests. Amazonian Brazil.
49c. Dalechampia scandens variety heterodonta Mueller Argoviensis,
TYPE: western Brazil, Tamberlik (G, holotype; W, isotype). Perhaps not distinct
DSTRIBUTION: Mato Grosso, Brazil.
in Martius, Flora Brasiliensis, 11 (2) : 658 ( 1874).
from variety fallax.
49d. Dalechampia scandens variety hibiscoides (H.B.K.) Mueller Argoviensis, in De Candolle, Prodromus Syslematis Universalis Regni Vegetabilis, 15(2j: 1245 (1866). Dalechampia hibiscoides H.B.K. Nova Genera et Species Plantarum, 2: 102 ( 18 1 7).
TYPE: Ecuador, near Guayaquil, Humboldt @ Bonpland (P, holotype). DISTRIBUTION: rain forests and seasonal forests, Pacific slopes and lowlands,
Ecuador. As interpreted here, Dalechampia scandens is a polytypic species confined to the
neotropics; paleotropical varieties recognized by Mueller Argoviensis ( 1866) and

162 G . L. WEBSTER AND W. S. ARMBRUSTER
Pax & Hoffmann (1919) appear to belong to different species. The four varieties enumerated here represent only a provisional sketch of the geographic variation within D. scandens.
50. Dalechampia stipulacea Mueller Argoviensis, Linnaea, 34: 22 1 ( 1865). TYPE: Peru, Pavon (G, holotype). SYNONYMS: Dalechampia stipulacea variety minor (Mueller Argoviensis) Mueller
Argoviensis, in Martius, Flora Brasiliensis, 11 (2) : 656 ( 1874). Dalechampia stipulacea forma minor Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1243 (1 866). TYPE: Brazil, Riedel (B, syntype, destroyed); Bahia, Blanchet 3909 ex p . (G, lectotype).
Dalechampia stipulacea variety membranacea Mueller Argoviensis, in Martius, Flora Brasiliensis, 22(2): 656 (1874). TYPE: Brazil, Minas Gerais, Lagoa Santa, Warming (C, holotype, not seen).
Dalechampia stipulacea variety piauhyensis Mueller Argoviensis, in Martius, Flora Brasiliensis, l l (2 ) : 667 (1874). TYPE: Brazil, Piauhy, Martius (G, holotype).
Dalechampia stipulacea variety bogotensis Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, ZV. 247. XZI (Hdt 68) : 28 ( 19 19). TYPE: Colombia, Bogoth, Triana 3555 (B, holotype, destroyed; G, lectotype).
PROBABLE SYNONYM: Dalechampia scandens variety Pallida Mueller Argoviensis, in Martius, Flora Brasiliensis, l l ( 2 ) : 659 (1874). TYPE: Brazil, Minas Geraes, Lagoa Santa, Warming (C, holotype) . Although Mueller referred this specimen to D. scandens, the characteristic glandular-fringed involucral bracts and stipules suggest a closer affinity to D. stipulacea.
DISTRIBUTION: seasonal tropical and subtropical forests, Colombia and Venezuela south to Argentina, Paraguay, and southern Brazil (Parani).
51. Dalechampia variij'olia Mueller Argoviensis, Linnaea, 34: 22 1 ( 1865). TYPE: Brazil, Minas Geraes, Widgren 3 (UPS, holotype, not seen; G, isotype
DISTRIBUTION: Minas Geraes, Brazil. [fragment]).
52. Dalechampia viridissima Webster, Brittonia, 41: 6, fig. 3 (1989). TYPE: Brazil, Bahia, CEPLAC grounds, Webster LY Armbruster 25165 (CEPEC,
DISTRIBUTION: Atlantic rain forests, Bahia and Espirito Santo, Brazil. NOTE: Dalechampia uiridissima is anomalous within subsection Dalechampia in its
reduced pubescence and deep green bracts, characters that suggest possible affinity with subsection Triphyllae. However, on the basis of leaf morphology it is referable to subsection Dalechampia.
holotype; DAV, GH, MO, NY, R. SP, UEC, isotypes).
Subsection 6b. Convolvuloides Webster & Armbruster, subsection nov. TYPE: Dalechampia convolvuloides Lamarck.
Frutices volubiles; lamina foliorum indivisa, acuminata, palmatinervia; bracteae involucratae virides vel lutescentes, indivisae vel tricuspidatae; involucellus staminalis floribus 7-1 0; stamina 12-40; sepala pistillata pinnatifida; columna stylaris vix dilatata; semina laevia.

NEOTROPICAL SPECIES OF DALECHAMPIA 163
Twining vines; leaves unlobed, acuminate, more or less cordate at base, palmatinerved, entire or dentate; involucral bracts green to yellowish, unlobed to tricuspidate; staminate involucel with 7-10 flowers; stamens 12-40; pistillate sepals pinnatifid; stylar column scarcely dilated at apex; seeds smooth.
53. Dalechampia arenalensis Armbruster, Systematic Botany, 9: 275 ( 1984). TYPE: Costa Rica, Alajueia, Volcan Arenal, Armbruster €9 Herzig 79-215 (DAV,
DISTRIBUTION: seasonal forests, northern Costa Rica. holotype; ALA, CR, MO, NY, isotypes).
54. Dalechampia boliviana Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Bolivia, Bang 2091 (NY, hololectotype; K, isotype). DISTRIBUTION: seasonal forests and thickets, Bolivia, Paraguay, and adjacent
Pjanzenreich, IV. 147. X I I (Hef t 68) : 50 (1919).
Brazil (Mato Grosso do Sul).
55. Dalechampia convolvuloides Lamarck, Encyclope'die Me'thodique, Botanique,
TYPE: Brazil, Dombey (P, holotype) . SYNONYM: DaLechampia m o n 5 ~ ~ l l a Vellozo, Flora Flum~nensis, 10: pl. 6 1 ( 183 1 );
Archivos do Museu NacionaL do Rio de Janeiro, 5: 409 (1881). TYPE: Brazil, Rio de Janeiro, Santa Cruz, Rio Taguay, Vellozo (type apparently lost; lectotype pl. 61).
2: 256 (1786).
DISTRIBUTION: coastal forests and restingas, Bahia to Rio de Janeiro, Brazil.
56. Dalechampia denticulata Wright ex Grisebach, Nachrichten der Akademie der
TYPE: Cuba, probably Pinar del Rio, Wright 2011 (GOET, holotype; GH, S,
DISTRIBUTION: western Cuba.
Wissenschaften in Gottingen, 1865: 18 1 ( 1865).
isotypes).
57. Dalechampia hispida Poeppig, Nova Genera ac Species Plantarum, 3: 19
TYPE: Peru, Mission Tocache, 1830, Poeppig 2016 (W, holotype). DISTRIBUTION: Amazonian rainforests, Peru.
(1841).
58. Dalechampia leandrii Baillon, Adansonia, 5: 3 15 ( 1865). TYPE: Brazil, Rio de Janeiro, Leandro di Sacramento 110 (P, lectotype). SYNONYM: Dalechampia vulptina Mueiler Argoviensis, Linnaea, 34: 222 (1865). TYPE:
DISTRIBUTION: rain forests along Serra do Mar, Rio de Janeiro, Brazil. Rio de Janeiro, Gaudichaud 1133 (G, lectotype).
59. Dalechampia schenckiana Pax & Hoffmann, in A. Engler (Ed.) , Das
TYPE: Brazil, Pernambuco, Garahuns, Schenck 4228 (B, holotype, destroyed;
DISTRIBUTION: agreste thickets, Pernambuco, Brazil.
Pjanzenreich, I V . 147. X I I (Hef t 68) : 49 (1919).
neotype here designated as Webster et al. 25G48, R; DAV, isotype).

164 G. L. WEBS'TER AND W. S. ARMBRUSTER
60. Dalechampia tenuiramea Mueller Argoviensis, Linnaea, 34: 222 ( 1865). TYPE: Brazil, Amazonas, Borba, Riedel22 (B, holotype, destroyed; G, lectotype). SYNONYMS: Dalechampia cynanchoides Spencer Moore, Transactions o f the Linnean
Society of London, Botany, series 4, 2: 468 ( 1895). Dalechampia tenuiramea variety cynanchoides (Spencer Moore) Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, I V . 147. X I I (Hejit 68) : 51 (1919). TYPE: Bolivia, Santa Cruz, Spencer Moore 483 (BM, lectotype).
DISTRIBUTION: seasonal and evergreen forests, Bolivia and western Brazil (Amazonas, Acre, Mato Grosso) to southern Venezuela (Amazonas).
Subsection 6c. Humiles (Pax & Hoffmann) Webster & Armbruster, stat.
BASIONYM: Dalechampia section Humiles Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Dalechampia humilis Mueller Argoviensis. SYNONYMS: Dalechampia section Guarantiticae Pax & Hoffmann, in A. Engler (Ed.),
Das Pjanzenreich, IV . 147. X I I (Hejl 68) : 40 (1919). TYPE: Dalechampia guaranitica Chodat & Hassler.
Dalechampia section Caperonioides Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, I V . 147. X I I ( H g t 68) : 1 1 ( 19 19). TYPE: Dalechampia caperonioides Baillon.
Dalechampia section Leucophyllae Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, I V . 147. X I I (Hejit 68) : 9 ( 19 19). TYPE: Dalechampia leucophylla Mueller Argoviensis. Erect or decumbent subshrubs (rarely clambering vines); leaves simple to
palmately lobed; floriferous shoots terminal or if axillary then with 1 or 2 nodes below the peduncle; involucral bracts white, yellow, or pink, unlobed to 3-lobed; staminate involucel with 8-1 1 flowers; stamens 10-25; pistillate sepals entire or pinnatifid; styles sometimes apically dilated; seeds smooth.
As circumscribed here, subsection Humiles includes most of the non-climbing species within section Dalechampia. Pax & Hoffmann separated sections Caperonioides and Leucophyllae because of their entire pistillate sepals, but the three species with this character are here treated as merely a group of subsection Humiles; their status requires further evaluation. However, there is no justification for recognition of their section Guaraniticae, which was defined by leaf polymorphism; it does not merit retention at any level.
nov.
Pjanzenreich, I V . 147. X I I ( H ~ t 68): 53 (1919).
Group 1. Pistillate sepals 5 or 6, entire (sections Caperonioides and Leucophyllae of Pax & Hoffmann).
61. Dalechampia caperonioides Baillon, Adansonia, 5: 3 16 ( 1865). TYPE: Brazil, Minas Gerais, Patrocinio, St. Hilaire C' 495 (P, lectotype). SYNONYMS: Dalechampia caperonioides variety lanceolata Mueller Argoviensis in De
Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1236 (1866). TYPE: Brazil, Minas Geraes, Rio Paranaiba, Pohl 626 (G, lectotype).
Dalechampia caperonioides variety obouata Mueller Argoviensis, in De Candolle, ~ r o d r o m ~ s Systemalis Universalis Regni Vegetabilis, 15(2) : 1235 ( 1866). TYPE:
Brazil, Goibs, Serra de Cristaes, Pohl 795 (G, lectotype). Dalechampia caperonioides variety rhomboidalis Mueller Argoviensis, in Martius,

165 NEO'I'ROPICAL SPECIES OF DALECHAMPIA
Flora Brasiliensis, 11 (2): 642 ( 1874). TYPE: Brazil, Minas Geras, Rio Paranaiba, Pohl 626 (G, lectotype).
Dalechampia caperonioides variety stenophylla Mueller Argoviensis, in Martius, Flora Brasiliensis, 11(2): 643 (1874). TYPE: Brazil, GoiAs, Chapadgo de S2o Marcos, Riedel2826 (G, holotype)
DISTRIBUTION: cerrados, central Brazil (Goias, Distrito Federal, Minas Geraes) . NOTE: despite the variability of this common cerrado species, it does not appear
practicable at this time to recognize the varieties accepted by Mueller or by Pax & Hoffmann.
62. Dalechampia francisceana Baillon, Adalzsonia, 5: 3 16 ( 1865). TYPE: Brazil, Minas Geraes', S5o Francisco, Weddell 2879 (P, holotype). DISTRIBUTION: Minas Geraes, Brazil.
63. Dalechampia leucophylla Mueller Argoviensis, Linnaea, 34: 2 19 ( 1865). TYPE: Brazil, Goib, Serra de Sgo Felix, Pohl 1937 (B, holotype, destroyed; G,
DISTRIBUTION: cerrados, Goias, Brazil. lectotype).
Group 2. Pistillate sepals usually more than 6, pinnatifid.
64. Dalechampia adscendens (Mueller Argoviensis) Mueller Argoviensis, in
BASIONYM: Dalechampia humilis variety adscendens Mueller Argoviensis, Linnaea, 34:
TYPE: Bolivia, Chiquitos, Santa Cruz, d'Orbigny 922 (G, hololectotype; P,
DISTRIBUTION: eastern Bolivia and western Brazil (Mato Grosso do Sul).
Martius, Flora Brasiliensis 12(2) : 640 ( 1874).
223 (1865).
isotype) .
65. Dalechampia glechomifolia Baillon, Adansonia, 5: 3 14 ( 1865). TYPE: Brazil, S2o Paulo, Gaudichaud 104, St. Hilaire c 1567 (P, syntypes). SYNONYM: Dalechampia microphylla Mueller Argoviensis, Linnaea, 34: 224 ( 1865).
DISTRIBUTION: campos, Argentina and southern Brazil (S2o Paulo to Rio Grande TYPE: Brazil, Sellow (B, holotype, destroyed).
do Sul).
66. Dalechampia guaranitica Chodat & Hassler, Bulletin de l'Herbier Boissier,
TYPE: Paraguay, Apepu (Tapiraguay), Hassler 4334 (G, holotype). DISTRIBUTION: Paraguay. Very close to D. weddelliana, differing only in its more or
series 5, 2: 608 ( 1905).
less deeply lobed leaves.
67. Dalechampia humilis Mueller Argoviensis, Linnaea, 34: 223 ( 1865). TYPE (variety procumbens Mueller Argoviensis) : Brazil, Goihs, Rio das Velhas,
DISTRIBUTION: cerrados, central Brazil (Goias, Distrito Federal, Minas Geraes) . Riedel (G, holotype) .
68. Dalechampia linearis Baillon, Adunsonia, 5: 3 15 ( 1865). TYPE: Brazil, Goias, Rio PilGes, St. Hilaire C' 801 (P, hololectotype).

166 G. L. WEBSTER AND W. S. ARMBRUSTER
SYNONYMS: Dalechampia breuipes variety monophylla Mueller Argoviensis, Linnaea, 34: 224 (1865). TYPE: Brazil, Piauhy, Gardner 2955 (G, lectotype).
Dalechampia linearis variety hilariana Baillon, Adansonia, 5: 3 16 (1865). TYPE: Brazil, Goias (?), St. Hiluire 676 (P, holotype).
Dalechampia linearis variety subintegra Mueller Argoviensis, in Martius, Flora Brasiliensis, 11(2): 644 (1874). TYPE: Brazil, Goias, Pohl3936 (G, lectotype).
Dalechampia goyazensis Mueller Argoviensis, in Martius, Flora Brasiliensis, 11(2) : 643 ( 1874). Dalechampia linearis variety goyazensis (Mueller Argoviensis) Pax & Hoffman, in A. Engler (Ed.), Das Pjanzenreich, ZV. 147. XIZ (Hef t 68 ) : 42 ( 19 19). TYPE: Brazil, GoiAs, Burchell 7491, Gardner 3980 (G, syntypes) .
DISTRIBUTION: cerrados, Goi as, Distrito Federal and western Minas Geraes, Brazil.
69. Dalechampia occidentalis Mueller Argoviensis, in Martius, Flora Brasiliensis, l l ( 2 ) : 641 (1874).
TYPE: western Brazil, Tamberlik (W, holotype; G, isotype). DISTRIBUTION: Mato Grosso, Brazil.
70. Dalechampia parvula Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Argentina, Misiones, San Pedro, Niederlein 15046 (B, holotype, destroyed). Pjanzenreich, I V . 147. XZZ (Hejit 68) : 55 (1919).
Perhaps a synonym of D. glechomifolia, as suggested by Allem & Waechter (1977).
DISTRIBUTION: northern Argentina.
71. Dalechampia rubrivenia Pax & Hoffmann, in A. Engler (Ed.), Das PfEanzenreich, IV. 147. XZI (Heft 68 ) : 43 (1919).
TYPE: Paraguay, Caaguazh, Hassler 9266 ex p . (B, holotype, destroyed). A dubious species, the flowers of which are unknown.
DISTRIBUTION: Paraguay.
72. Dalechampia serrula Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Paraguay, Sierra de Maracayu, Ipehu, Hassler 5296 (B, holotype,
DISTRIBUTION: Paraguay.
Pjanzenreich, IV. 147. XZZ (Heft 68 ) : 44.
destroyed; G, lectotype).
73. Dalechampia ulmifolia Chodat & Hassler, Bulletin de I’Herbier Boissier, se‘rie 2, 5: 608 (1905). TYPE: Paraguay, Valenzuela, Hassler 6949 (G, holotype; S, isotype) .
SYNONYM: Dalechampia grueningiana Pax, Reportorium Specierum Novarum Regni Vegetabilis, 8: 16 1 ( 19 10). Dalechampia ulmifolia variety grueningiana (Pax) Pax, in A. Engler (Ed.), Das Pjanzenreich, ZV. 147. XIZ (Hef t 68 ) : 54 ( 19 19). TYPE: Brazil, Rio Grande do Sul, Estancia Lourenqo Gomes, Neu Wurttemberg, Bornmuller 181 (B, holotype, destroyed).
DISTRIBUTION: campos, northern Argentina, Paraguay and southern Brazil (Rio Grande do Sul).

NEOTROPICAL SPECIES OF DALECHAMPIA 167
74. Dalechampia weddelliana Baillon, Adansonia, 5: 3 15 ( 1865). TYPE: Brazil, between GoiAs and CuiabA, Weddell 2974 (P, holotype). SYNONYMS: Dalechampia amambayensis Pax & Hoffmann, in A. Engler (Ed.), Das
Pjanzenreich, IV. 147. X I I (Heft 68): 43 (1919). TYPE: Paraguay, Sierra de Amambay, Punta Para, Hassler 9763 (B, holotype, destroyed; W, lectotype; K, isolectotype). Dalechampia rnorifolia Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, IV. 147. X I I (Heft 68): 42 (1919). TYPE: Paraguay, Apepu (Tapiraguay), Hassler 4335 (B, holotype, destroyed; G, lectotype). Dalechampia trichophila Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, IV. 147. X I I ( H q t 68): 44 (1919). TYPE: Paraguay, Rio Capibary, Hassler 4476 (G, lectotype).
DISTRIBUTION: cerrados and campos, Brazil (GoiBs, Mato Grosso, Mato Grosso do Sul).
Group 3. Stems clambering; pistillate sepals more than 6, pinnatifid; involucral bracts white or pink.
75. Dalechampia schippii Standley, Fieldiana, Botany, 11: 133 ( 1932). TYPE: Belize, Sarawee, Schipp S-181 (F, holotype). DISTRIBUTION: pinelands and thickets, Belize and adjacent Guatemala.
This common species of Belize appears to have much in common with the South American species of subsections Convoluuloides and Hurniles, but does not seem to be closely related to any of them; it is placed here tentatively.
Subsection 6d. Brevipedes (Pax & Hoffmann) Webster & Armbruster, stat. nov.
BASIONYM: Dalechampia section Brevipedes Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, IV. 147. X I I (Hefi 68): 26 (1919).
TYPE: Dalechampia brevipes Mueller Argoviensis. Erect subshrubs; leaves palmately parted, nearly sessile; inflorescences
terminal; bracts yellowish, 3- or 5-lobed; staminate involucel with 7- 10 flowers; stamens 10-30; pistillate sepals subentire to pinnatifid; seeds smooth.
76. Dalechampia brevipes Mueller Argoviensis, Linnaea, 34: 224 ( 1865). TYPE (variety pentaphylla Mueller Argoviensis) : Brazil, S2o Paulo, Rio Pardo,
DISTRIBUTION: cerrados, western Brazil (Mato Grosso, Mato Grosso do Sul, SBO Riedel582 (B, holotype, destroyed; NY, lectotype).
Paulo) .
77. Dalechampia riedeliana Mueller Argoviensis, in De Candolle, Prodromus
SYNONYM: Dalecharnpia breu$es variety triphylla Mueller Argoviensis, Linnaea, 34:
TYPE: Brazil, Mato Grosso, Serra do Diamantina, Riedel 1120 (B, holotype,
DISTRIBUTION: cerrados, Mato Grosso, Brazil.
Systematis Universalis Regni Vegetabilis, 15(2) : 1236 ( 1866).
225 (1865).
destroyed; NY, lectotype).

168 G. L. WEBSTER AND W. S. ARMBRUSTER
Subsection 6e. Triphyllae (Pax & Hoffmann) Webster & Armbruster, stat. nov.
BASIONYM: Dalechampia section Triphyllae Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, ZV. 147. XZZ (Hgt 68) : 13 ( 191 9).
TYPE: Dalechampia triphylla Lamarck. Twining vines, indumentum often sparse; leaves trifoliolate (rarely
5-foliolate), sometimes also with simple leaves; floriferous shoots axillary ; bracts greenish, unlobed to 3-lobed; staminate involucel with 7-1 1 flowers; stamens 10- 100; pistillate sepals 6- 12, entire to deeply pinnatifid; seeds smooth.
Most of the species of this mainly South American group were reduced to synonyms of the type species, D. triphylla, by Allem & Waechter (1977). However, although it is clear that Pax & Hoffmann (1919) were too liberal in recognizing species, there is no doubt that D. triphylla is distinct from all of the other taxa by virtue of its very shallowly toothed pistillate sepals. Much more field and experimental work needs to be done to elucidate this difficult complex.
78. Dalechampia anomala Pax & Hoffmann, in A. Engler (Ed.), Das
TYPE: Paraguay, between Rio Apa and Rio Aquidaban, Fiebrig 4673 (G,
DISTRIBUTION: seasonal woodlands, Paraguay and adjacent Brazil (Parana) .
Pjanzenreich, ZV. 147. XI I (Hej l 68 ) : 24, fig. 6 (1919).
lectotype; K, isotype).
79. Dalechampia bangii Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, ZV. 147. XZI (Hgt 68) : 24 ( 19 19).
TYPE: Bolivia, Bang 2181 (B, holotype, destroyed; NY, lectotype, K, US, isolectotypes).
SYNONYM: Dalechampia boliviana Gandoger, Bulletin de la SociCte' Botanique de France, 66: 286 (1920, non Dalechampia boliviana Pax & Hoffmann, 1919). TYPE: also Bang 2181.
DISTRIBUTION: seasonal forests, Bolivia, Paraguay, and adjacent Brazil (Mato Grosso do Sul).
80. Dalechampia burchellii Mueller Argoviensis, in Martius, Flora Brasiliensis,
TYPE: Brazil, Goias, Burchell 6661 (G, holotype; K, isotype). DISTRIBUTION: seasonal woodlands (cerradgo) , GoiAs, Brazil.
11(2) : 649 (1874).
81. Dalechampia cissifolia Poeppig, Nova Genera ac Species Plantarum, 3: 20 (1841). 81a. Dalechampia cissifolia subspecies cissifolia:
SYNONYM: Dalechampia triphylla variety cissifolia (Poeppig) Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1239 (1866). TYPE: Peru, Maynas, Yurimaguas, Poeppig 2085 (W, holotype; G, isotype).
81b. Dalechampia cissifolia subspecies panamensis (Pax &r Hoffmann) Webster, Annals o f the Missouri Botanical Garden, 54: 193 (1967).
BASIONYM: Dalechampia panamensis Pax & Hoffmann, in A. Engler (Ed.), Das Pjanzenreich, IF'. 147. XZZ (He$ 68) : 19 ( 19 19).
DISTRIBUTION: evergreen forests. Amazonian Peru, Venezuela, and Brazil.

NEOTROPICAL SPECIES OF DALECHAMPZA 169
TYPE: Panama, Cerro Gordo, Pittier 2311 (US67656 1, hololectotype; designated
DISTRIBUTION: evergreen forests, Nicaragua to Panama. by Webster, 1967).
82. Dalechampia claussenialta Baillon, Adansonia, 5: 3 10( 1865). TYPE: Brazil, Rio de Janeiro, Novo Friburgo, Claussen 115 (P, holotype; G,
isotype) . SYNONYMS: Dalechampia triphylla variety clausseniana (Baillon) Mueller Argoviensis,
in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1240 (1866).
Dalechampia triphylla variety villosa (forma pohliana) Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1240 ( 1866). TYPE: Brazil, Villa Boa, Pohl (G, holotype; W, isotype).
Dalechampia triphylla variety villosa forma riedeliana Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1240 (1866). TYPE: Brazil, Mandioca, Riedel (G, holotype).
DISTRIBUTION: rain forests or seasonal forests, Goias and SBo Paulo to Espirito Santo and Rio de Janeiro, Brazil.
83. Dalechampia hassleriana Chodat, Bulletin de /'Herbier Boissier, serie 2, 5:
TYPE: Paraguay, Rio Corrientes, Hassler 5852 (G, holotype) . DISTRIBUTION: seasonal woodlands, Paraguay and adjacent Brazil (Parana).
609 (1905).
84. Dalechampia heteromorpha Pax & Hoffmann, in A. Engler (Ed.), Das PJlanzenreich, IV. 147. XII (Ht$ 68) : 26 ( 19 19).
TYPE: Costa Rica, Rio Grande, San R a m h , Brenes 14414 (B, holotype, destroyed; K. lectotype).
SYNONYMS: Dalechampia triphylla variety mexicana Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 1239 ( 1866). TYPE: Mexico, Veracruz, Liebmann (C, syntypes).
Dalechampia scandens variety trisecta Donneli Smith, Botanical Gazette, 13: 199 (1888). TYPE: Guatemala, Escuintla, San Juan Mixtan, Donnell Smith 2070 (F, holotype; US, isotype).
Dalechampia guatemalensis Gandoger, Bulletin de la Sociite' Botanique de France, 66: 286 (1920). TYPE: Guatemala, Alta Verapaz, Tuerckheim (holotype not seen; US, isotype).
Dalechampia molliuscula Blake, Contributions to the United States National Herbarium, 24: 12 (1922). TYPE: Guatemala, Izabal, Quebradas, Blake 7547 (US 98963 1 , holotype).
DISTRIBUTION: rain forests. southern Mexico (Veracruz) to Costa Rica.
85. Dalechampia laevigata Standley, Publications o f the Field Columbian Museum, Botanical Series, 4: 3 12 ( 1929),
TYPE: Honduras, Lancetilla Valley, Standley 53531 (F 583076, holotype). These plants of the Lancetilla Valley may prove to represent only a constantly unlobed leaf form of D. heteromorpha.
DISTRIBUTION: Atlantic rain forests, Honduras.

170 G. L. WEBSTER AND W. S. ARMBRUSTER
86. Dalechampia meridionalis Mueller Argoviensis, in Martius, Flora
TYPE: Brazil, Sgo Paulo, Burchell4299 (G, lectotype). SYNONYM: Dalechampia pallida Klotzsch ex Pax & Hoffmann, in A. Engler (Ed.),
Das PJancenreich, ZV. 147. XIZ (Hdt 68): 22 (1919): TYPE: Brazil, Sellow 636exp . (B, holotype, destroyed; K, lectotype).
DISTRIBUTION: seasonal tropical and subtropical forests, Siio Paulo and Minas Geraes, Brazil, south to Uruguay.
Brasiliensis, 11 (2) : 648 ( 1874).
87. Dalechampia micromeria Baillon, Adansonia, 5: 310 (1865). TYPE: Brazil, Rio de Janeiro, Gaudichaud 1130 (G, holotype; P, isotype). SYNONYMS: Dalechampia sellowiana variety serrata Mueller Argoviensis, Linnaea, 34:
220 (1865). TYPE: Brazil, S2o Paulo, Sellow 4177 (B, holotype, destroyed; P, lectotype).
Dalechampia sellowiana var. subintegra Mueller Angoviensis, Linnaea, 34: 220 (1865). TYPE: Brazil, Rio de Janeiro, Casaretto 1178 (G, lectotype).
Dalechampia micromeria variety angustifolia Pax & Hoffmann, in A. Engler (Ed.), Das PJanzenreich. ZV. 147. XZZ (Hdt 68): 23 (1919). TYPE: Paraguay, Cordillera de Altos, Hassler 6067 (G, lectotype).
Dalechampia crenulata Pax & Hoffmann, in A. Engler (Ed.), Bas Pjancenreich, ZV. 147. XVII (Hdt 85): 189 (1924). TYPE: Brazil, Siio Paulo, Serra Caracol, Moskn 1626 (not seen).
DISTRIBUTION: thickets and restingas, southern Brazil (Rio de Janeiro to Rio Grande do Sul).
88. Dalechampia olfersiana Mueller Argoviensis, Linnaea, 34: 280 ( 1865). TYPE: Brazil, Sellow 636 ex p . (B, holotype, destroyed; G, lectotype [fragment]). DISTRIBUTION: seasonal woodlands, Minas Geraes, Brazil.
89. Dalechampia regnellii Mueller Argoviensis, in Martius, Flora Brasiliensis,
TYPE: Brazil, Minas Gerais, Caldas, Regnell 1053 (G, holotype; S, isotype) . DISTRIBUTION: Minas Geraes, Brazil.
l l ( 2 ) : 647 (1874).
90. Dalechampia stenosepala Mueller Argoviensis, in De Candolle, Prodromus
TYPE: Brazil, Sellow (B, holotype, destroyed; G, lectotype [fragment]). SYNONYM: Dalechampia patagonica Pax & Hoffmann, in A. Engler (Ed.), Das
Pjanrenreich, ZV. 147. XVZZ (Hejit 85): 190 (1922). TYPE: Brazil, Paranh, Santa Cruz, Ponta Grossa, Duskn 2497 (B, holotype, destroyed). Lourteig (1954) pointed out that Pax & Hoffmann incorrectly located their proposed new species in Argentina and that it is not distinguishable from D. stenosepala.
DISTRIBUTION: seasonal topical and subtropical forests, Argentina, Paraguay, and southern Brazil (Rio de Janeiro to Rio Grande do Sul).
Systematis Universalis Regni Vegetabilis, 15(2) : 1241 ( 1866).
91. Dalechampia triphylla Lamarck, Encyclopkdie Mkthodique, Botanique, 2: 258
TYPE: Brazil, Dombey (P, holotype [Herb. Jussieu 162921; NY, isotype). SYNONYM: Dalechampia crenulata Gandoger, Bulletin de la Socikttt Botanique de France,
(1 786).

NEOTROPICAL SPECIES OF DALECHAMPIA 171
66: 286 (1920) [non D. crenulata Pax & Hoffm., 19241. TYPE: Brazil, S2o Paulo, Campinas, Nouaes 269 (not seen).
Dalechampia triphylla is here construed in a narrower sense than that of Mueller (1866) or Allern & Waechter (1977), to include only populations in southern Brazil with broad (not narrowly pinnatisect) fruiting pistillate sepals. DISTRIBUTION: seasonal tropical and subtropical forests, south-eastern Brazil (S2o
Paulo, Rio de Janeiro).
92. Dalechampia weberbaueri Pax & Hoffrnann, in A. Engler (Ed.), Das
TYPE: Peru, Jatn, Cajamarca, Weberbauer 6192 (B, holotype, destroyed; G,
DISTRIBUTION: montane thickets, Peru.
Fjansenre~ch, IV. 147. X I I (Hejit 68): 21 (1919).
lectotype).
SPECIES OF UNCERTAIN STATUS
93. Dalechampia martiana Klotzsch ex Pax & Hoffmann, in A. Engler (Ed.),
TYPE: Brazil, Sgo Paulo, Martius (B, holotype, destroyed). Probably belongs in Das Pjansenreich, IV. 147. X I I (Hejit 68): 29 (1919).
subsection Dalechampia.
94. Dalechampia ternata Parodi, Anales Cient@as Argentina, 11: 52 (1881). TYPE: not seen. NOTE: referred with doubt to D. micromeria by Pax & Hoffmann (1919).
ADDENDUM
95. Dalechampia allemii Webster, Annals o f the Missouri Botanical Garden, in press.
TYPE: Brazil, Bahia, Mun. Andarai, 50 km NW of Andarai, Allem, Webster, €9 Werneck 2980 (CEN, holotype; DAV, isotype). The species is referable to subsection Triphyllae.
EXCLUDED ‘I‘AXA
Section Champadelia Mueller Argoviensis, in De Candolle, Prodromus Systematis Universalis Regni Vegetabilis, 15(2) : 664 ( 1866).
TYPE: Dalechampia houlletiana Baillon, Etude Glnnkale du Groupe des Euphorbiacles: 486, pl. 3, figs 31, 32 (1858).
TYPE: Brazil, Pernambuco, Houllet (P, holotype) . This mysterious plant proves not to belong to the Euphorbiaceae. The
fragmentary specimen has distinctive tack-shaped glands that are totally foreign to Dalechampia. I t probably represents a species of Cissus (Vitaceae).
ACKNOWLEDGEMENTS
We are indebted to the curators of the herbaria that have loaned or given access to critical specimens examined during this study: F, G, GH, K, M, MO, NY, P, RJ, S, UC, W. The assistance of Dr Alicia Lourteig at the Museum d’Histoire Naturelle, Paris, was especially valuable. Helpful comments on the manuscript were provided by Dr Adrian Juncosa and other reviewers. The illustrations were drawn by Ms Leslie Randall.

172 G . L. WEBSTER AND W. S. ARMBRUSTER
REFERENCES
ALLEM, A. C., 1977. Notas taxonbmicas sobre as tribos Phyllantheae, Dalechampieae e Manihoteae
ALLEM, A. C. & WAECHTER, J. L., 1977. Notas sistemiticas y nuevos sinbnimos en Euphorbiaceae de
ARMBRUSTER, W. S., 1982. Seed predation and dispersal in Dalechampia (Euphorbiaceaej: divergent
ARMBRUSTER, W. S., 1983. Dalechampia scandens. In D. H . Janzen (Ed.), Costa Rican Natural History:
ARMBRUSTER, W. S., 1984a. The role of resin in angiosperm pollination: ecological and chemical
ARMBRUSTER, W. S., 1984b. Two new species of Dalechampia from Mesoamerica. Systematic Botany, 9:
ARMBRUSTER, W. S., 1985. Patterns of character divergence and the evolution of reproductive ecotypes of Dalechampia scandens (Euphorbiaceaej. Evolution, 39: 733-752.
ARMBRUSTER, W. S., 1988a. A new species, section, and synopsis of Dalechampia (Euphorbiaceae) from Costa Rica. Systematic Botany, 13: 303-312.
ARMBRUSTER, W. S., 1988b. Multilevel comparative analysis of morphology, function, and evolution of Dalechampia blossoms. Ecology, 69: 1746-1 761.
ARMBRUSTER, W. S., 1989. Three new species of Dalechampia (Euphorbiaceae) from Venezuelan Guayana. Brittonia, 41: 44-52.
ARMBRUSTER, W. S. & HERZIG, A. L., 1984. Partitioning and sharing of pollinators by four sympatric species of Dalechampia (Euphorbiaceae) in Panama. Annals of the Missouri Botanical Garden, 71: 1-16.
ARMBRUSTER, W. S. & WEBSTER, G . L., 1979. Pollination of two species ofDalechampia (Euphorbiaceae) in Mexico by euglossine bees. Biotropica, 11: 278-283.
ARMBRUSTER, W. S. & WEBSTER, G. L., 1981. Sistemas de polinizaGio de duas especies simpitricas de Dalechampia (Euphorbiaceaej no Amazonas, Brasil. Acta Amazonica, 11(1): 13-17.
ARMBRUSTER, W.. S. & WEBSTER, G. L., 1982. Divergent pollination systems in sympatric species of South American Dalechampia (Euphorbiaceae). American Midland Naturalist, 108: 325-337.
ARMBRUSTER, W. S., HERZIG, A. L. & CLAUSEN, T. P., 1990. Incomplete partitioning of male euglossine pollinators by two sympatric species of Dalechampia (Euphorbiaceae) in Suriname. Journol of Tropical Ecology, in press.
ARMBRUSTER, W. S., KELLER, S., MATSUKI, M. & CLAUSEN, T . P., 1989. Pollination of Dalechampia magnoliiflia (Euphorbiaceae) by male euglossine bees (Apidae: Euglossini). American Journal of Botany, 76:
(Euphorbiaceaej no Rio Grande do Sul. Iheringia, Botbnica, 22: 3-15.
America del Sur-11. Revista Brasileira de Biologia, 37: 91-101.
patterns and ecological consequences. American Journal of Botany, 69: 1429-1440.
230-233. Chicago: University of Chicago Press.
considerations. American Journal of Botany, 71: 1149-1 160.
272-278.
1279-1285. BAILLON, H., 1858. Etude Gkndrale du Groupe des Euphorbiacies. Pans: Victor Masson. CROIZAT, L., 1940. O n the phylogeny of the Euphorbiaceae and some of their presumed allies. Revista
CROIZAT, L., 1942. Peculiarities of the inflorescence in the Euphorbiaceae. Batanical Gazette, 103: 771-779. FROEBE, H. A,, MAGIN, N., JOHLINGER, H. & NETZ, M., 1983. A re-evaluation of the inflorescence of
Dalechampia spathulala (Scheidw. j Baillon (Euphorbiaceae). Botanische Jahrbucher f u r Systematik, 104: 261-287. HUFT, M. J., 1984. A new combination in Dalechampia (Euphorbiaceae). Annals of the Missouri Botanical Garden,
71: 341. KNOLL, F., 1905. Die Brennhaare der Euphorbiaceen-Gattungen Dalechampia und Tragia. Sitzungsberichte der
Kaiserlichen natunvissenschaftliche Klasse, 114: 29-48. MICHAELIS, P., 1924. Blutenmorphologische Untersuchungen an den Euphorbiaceen. Botanische
Abhandlungen, 3: 1-150. LEONARD, J., 1962. Euphorbiaceae, Dalechampieae. Flore du Congo et du Rwanda-Burundi, Spermatophytes, 8 ( 1 ) :
LOURTEIG, A,, 1954. Euphorbiaceae Argentinae, Addendum 11. Arkiv f i r Botanik, I I . 3 : 71-87. MUELLER, J., 1866. Euphorbiaceae, in A. P. De Candolle (Ed.), Prodromus $stematis Universalis Regni
MUELLER, J., 1874. Euphorbiaceae, Dalechampieae, in von Martius, Flora Brasiliensis, 11 (2 ) : 633-664. PAX, F., 1890. Euphorbiaceae. In A. Engler & K. Prantl (Eds), Die Naturlichen Pjanrenfamilien, Erste Aujage,
PAX, F. & HOFFMANN, K., 1919. Euphorbiaceae-Dalechampieae. In A. Engler (Ed.), Das Pjanzenreich, I?'.
PUNT, W., 1962. Pollen morphology of the Euphorbiaceae with special reference to taxonomy. Wenlza, 7:
SAZIMA, M., SAZIMA, I. & CARVALHO-OKANO, R. M., 1985. Biologia floral de Dalechampia stipulacea (Euphorbiaceaej e sua polinizaqio por Euglossa melanotricha (Apidae). Revista Brasileira de Biologia, 45: 85-93.
URBAN, I., 1888. Uber den Blutenstand von Dalechampia. Jahrbuch des Koniglichen Botanischen Gartens und des Botanischen Museums zu Berlin, 4: 252-259.
VENKATA RAO, C., 1971. Anatomy of the inflorescence ofsome Euphorbiaceae. With a discussion on the phylogeny and evolution of the inflorescence, including the cyathium. Botaniska Notisel-, 124: 39-64.
Universitdria, Santiago, 25: 205-220.
194-202.
Vegetabilis, 15(2) : 189- 126 1.
III . 3 ( 5 ) : 1-119.
147. XI1 (Heft 68): 1-59.
1-1 16.

NEOTROPICAL SPECIES OF DALECHAMPIA 173
WEBSTER, G. L., 1968. Dalechampia. In Flora of Panama, Part VI. Annals of the Missouri Botanical Garden, 54: 308-3 16.
WEBSTER, G. L., 1989. Three new species of Dalechampia (Euphorbiaceae) from Brazil. Brzttonia, 41: 1-9. WEBSTER, G. L. & ARMBRUSTER, W. S., 1979. A new euglossine-pollinated species of Dalechampia from
WEBSTER, G. L. & ARMBRUSTER, W. S., 1982. An unusual new species of Dalechampia from Surinam.
WEBSTER, G. L. & HUFT, M. J., 1988. Revised synopsis of Panamanian Euphorbiaceae. Annals of the
WEBSTER, G. L. & WEBSTER, B. D., 1972. The morphology and relationships of Dalechampia scandens
Veracruz, Mexico. Brittonia, 31: 352-357.
Systematic Botany, 7: 484-488.
Missouri Botanical Ga~den, 75: 1087- 1 144.
(Euphorbiaceae). American Journal of Botany, 59: 573-586.
INDEX T O 'I'AXA
Numbers of species and subspecific taxa indicate the species numbers assigned in the text; numbers in bold face indicate numbers assigned to supraspecific taxa. Taxa accepted in the synopsis at the rank indicated are given in roman, synonyms in italics, new taxa in bold face.
Cremophyllum Scheidw. 3 Cremophyllum spathulatum Scheidw. 34. Dalechampia -section Dalechampia section Dalechampia section Dalechampia section Dalechampia section Dalechampia section Dalechampia section Dalechampia section Dalechampia section Dalechampia section Dalechampia section Dalechampia section Dalechamoia section
Brevipedes Pax & Hoffm. 6d Caperonioides 6c Champadelia [excluded taxa] Coriaceae Pax & Hoffm. 4 Cremophyllum (Scheidw.) Baillon 3 Dalechampia 6a Guaraniticae Pax & Hoffm. 6c Humiles Pax & Hoffm. 6c Leucophyllae Pax & Hoffm. 6c Rhopalostylis Pax & Hoffm. 1 Scandentes 6a Tiliifoliae Webster & Armbruster 5 Triphyllae Pax & Hoffm. 6e
Dalechampia subsection Brevipedes (Pax & Hoffm.) Webster &
Dalechampia subsection Convolvuloides Webster & Armbruster Dalechampia subsection Dalechampia 6a Dalechampia subsection Humiles (Pax & Hoffm.) Webster &
Dalechampia subsection Triphyllae (Pax & Hoffm.) Webster &
Dalechampia adscendens (Muell. Arg.) Muell. Arg. Dalechampia affinis Muell. Arg. 36 Dalechampia alata Muell. Arg. 8 DaIechampia albibracteosa Rusby 25 Dalechampia allemii 95 Dalechampia amambayensis Pax & Hoffm. Dalechampia amaronica (Ule) Macbr. 33 Dalechampia anomala Pax & Hoffm. 78 Dalechampia arenalensis Armbruster 53 Dalechampia arciana Baillon 19
Armbruster 6d 6b
Armbruster 6c
Armbruster 6e 64
74

174 G . L. WEBSTER AND W. S. ARMBRUSTER
Dalechampia aristolochiifolia H. B. K. Dalechampia attenuistylus Armbruster 1 Dalechampia bahiensis Muell. Arg. 38 Dalechampia bangii Pax & Hoffm. Dalechampia boliviana Gandoger 79 Dalechampia boliviana Pax & Hoffm. Dalechampia brasiliensis Lam. 4 1 Dalechampia brasiliensis var. viridis Muell. Arg. Dalechampia brevipedunculata Ule 32 Dalechampia brevipes Briq. 49a Dalechampia brevipes Muell. Arg. 76 Dalechampia brevipes var. monophylla Muell. Arg. 68 Dalechampia brevipes var. pentaphylla Muell. Arg. 76 Dalechampia brevipes var. triphylla Muell. Arg. Dalechampia brownsbergensis Webster & Armbruster Dalechampia burchellii Muell. Arg. 80 Dalechampia canescens H. B. K. Dalechampia canescens subsp. canescens Dalechampia canescens subsp. friedrichtsthalii (Muell. Arg.) Webster &
Dalechampia caperonioides Baillon 6 1 Dalechampia caperonioides var. lanceolata Muell. Arg. Dalechampia caperonioides var. obovata Muell. Arg. 6 1 Dalechampia caperonioides var. rhomboidalis Muell. Arg. Dalechampia caperonioides var. stenophytla Muell. Arg. Dalechampia cissifolia Poepp. 8 1 Dalechampia cissifolia subsp. cissifolia Dalechampia cissifolia subsp. panamensis (Pax & Hoffm.) Webster Dalechampia clausseniana Baillon 82 Dalechampia colorata L. f, 39 Dalechampia convolvuloides Lam. 55 Dalechampia coriacea K1. ex Muell. Arg. Dalechampia crenulata Gandoger 9 1 Dalechampia crenulata Pax & Hoffm. Dalechampia cujabensis Muell. Arg. 27 Dalechampia cyanchoides Sp. Moore 60 Dalechampia denticulata Wright ex Griseb. Dalechampia dioscoreifolia Poepp. 28 Dalechampia dioscoreifolia var. pubescens Muell. Arg. Dalechampia ficifolia Lam. 43 Dalechampiajimbriata H. B. K. 49a Dalechampia fragrans Armbruster 2 Dalechampia francisceana Baillon 62 Dalechampia friedrichsthalii Muell. Arg. Dalechampia glechomifolia Baillon 65 Dalechampia goyazensis Muell. Arg. 68 Dalechampia granadilla Baillon 44 Dalechampia grueningiana Pax 73 Dalechampia guaranitica Chod. & Hassl.
26
79
54
4 1
77 42
21 2 1 a
Huft 2 1 b
6 1
6 1 6 1
8 1 a 81 b
35
87
56
28
2 1 b
66

NEOTROPICAL SPECIES OF DALECHAMPZA
Dalechampia guatemalensis Gandoger 84 Dalechampia hassleriana Chodat 83 Dalechampia hastata Webster 3 Dalechampia herzogiana Pax & Hoffm. Dalechampia heteromorpha Pax & Hoffm. Dalechampia heterophylla Vahl 39 Dalechampia hibisczformis Spreng. 49d Dalechampia hibiscoides H. B. K . 49d Dalechampia hispida Poepp. 57 Dalechampia houlletiana Baillon [excluded taxa] Dalechampia humilis Muell. Arg. 67 Dalechampia humilis var. procumbens Muell. Arg. Dalechampia humilis var. adscendens Muell. Arg. Dalechampia hutchisoniana Webster 29 Dalechampia ilheotica Wawra 38 Dalechampia juruana Ule 9 Dalechampia karsteniana Pax & Hoffm. Dalechampia laevigata Standley 85 Dalechampia latzfolia Lam. 49a Dalechampia leandrii Baillon 58 Dalechampia leucophylla Muell. Arg. 63 Dalechampia liesneri Huft 4 Dalechampia linearis Baillon 68 Dalechampia linearis var. goyazensis (Muell. Arg.) Pax & Hoffm. Dalechampia linearis var. hilariana Baillon Dalechampia linearis var. subintegra Muell. Arg. Dalechampia luetzelburgii Pax & Hoffm. Dalechampia magnoliifolia Muell. Arg. 33 Dalechampia martiana K1. ex Pax & Hoffm. Dalechampia megacarpa Armbruster 20 Dalechampia meridionalis Muell. Arg. 86 Dalechampia micrantha Poepp. 5 Dalechampia micromeria Baillon 8 7 Dalechampia micromeria var. angustifolia Pax & Hoffm. Dalechampia microphylla Muell. Arg. 65 Dalechampia mollis Vahl 49a Dalechampia monophylla Vell. 55 Dalechampia molliuscula Blake 84 Dalechampia morzfolia Pax & Hoffm. 74 Dalechampia occidentalis Muell. Arg. 69 Dalechampia olfersiana Muell. Arg. 88 Dalechampia olympiana Kuhlm. & Rodr. Dalechampia osana Armbruster 22 Dalechampia Pallida K1. ex Pax & Hoffm. Dalechampia panamensis Pax & Hoffm. 8 1 b Dalechampia papillistigma Armbruster 1 1 Dalechampia papposa Endl. 4 1 Dalechampia parvibracteata Lanj. 7 Dalechampia parvula Pax & Hoffm.
45 84
67 64
30
68 68
10 68
93
88
6
86
70
I75

176 G . L. WEBSTER AND W. S. ARMBRUSTER
Dalechampia passijlora Chod. & Hassl. 49a Dalechampia patagonica Pax & Hoffm. 90 Dalechampia peckoltiana Muell. Arg. 12 Dalechampia pentaphylla Lam. 13 Dalechampia pernambucensis Baillon 47 Dalechampia peruviana Lam. 39 Dalechampia pruriens Griseb. 39 Dalechampia psilogyne Muell. Arg. 14 Dalechampia regnellii Muell. Arg. 89 Dalechampia riedeliana Muell. Arg. 77 Dalechampia riparia Smith & Downs Dalechampia roezliana Muell. Arg. 34 Dalechampia roezliana var. alba Nicholson Dalechampia roezliana var. amazonica Ule 33 Dalechampia roezliana var. viridis Muell. Arg. Dalechampia rubifarmis Spreng. 49a Dalechampia ruboides H. B. K. 49a Dalechampia rubrivenia Pax & Hoffm. Dalechampia scandens L. 49 Dalechampia scandens var. brasiliensis (Lam.) Muell. Arg. Dalechampia scandens var. fallax Muell. Arg. 49b Dalechampia scandens var.jmbriata (H. B. K.) Muell. Arg. Dalechampia scandens var. heterodonta Muell. Arg. Dalechampia scandens var. hibiscoides (H. B. K.) Muell. Arg. Dalechampia scandens f. latifolia (Lam.) Muell. Arg. Dalechampia scandens var. mollis (Vahl) Muell. Arg. Dalechampia scandens var. pallida Muell. Arg. Dalechampia scandens var. pernambucensis (Baillon) Pax & Hoffm. Dalechampia scandens var. trisecta Donn. Sm. Dalechampia schenckiana Pax & Hoffm. 59 Dalechampia schizoloba Muell. Arg. 19 Dalechampia schippii Standley 75 Dalechampia schottii Greenm. 3 1 Dalechampia schottii var. trifoliata Greenm. Dalechampia sellowiana Muell. Arg. 87 Dalechampia sellowiana var. serrata Muell. Arg. Dalechampia sellowiana var. subintegra Muell. Arg. Dalechampia serrula Pax & Hoffm. Dalechampia sidif l ia H. B. K. Dalechampia smilacina H. B. K. 21a Dalechampia spathulata (Scheidw.) Baillon 34 Dalechampia spathulata var. alba (Nicholson) Pax & Hoffm. Dalechampia spathulata var. amazonica (Ule) Pax & Hoffm. 33 Dalechampia spathulata var. rosea (Muell. Arg.) Pax & Hoffm. Dalechampia spathulata var. viridis (Muell. Arg.) Pax & Hoffm. Dalechampia stenosepala Muell. Arg. 90 Dalechampia stipulacea Muell. Arg. 50 Dalechampia stipulacea var. bogotensis Pax & Hoffm. Dalechampia stipulacea var. major Muell. Arg.
48
34
34
71
41
49a 49c
49a 49a
49d
50 47
84
3 1
87 87
72 49a
34
34 34
50 50

177 NEOTROPICAL SPECIES OF DALECHAMPIA
Dalechampia stipulacea var. minor Muell. Arg. 50 Dalechampia stipulacea var. membranacea Muell. Arg. Dalechampia stipulacea var. piauhyensis Muell. Arg. Dalechampia subintegra (Muell. Arg.) Muell. Arg. Dalechampia sylvestris Sp. Moore 16 Dalechampia tenuiramea Muell. Arg. 60 Dalechampia tenuiramea var. cynanchoides (Sp. Moore) Pax & Hoffm. Dalechampia ternata Parodi 94 Dalechampia tiliifolia Lam. 39 Dalechampia tiliifolia var. jcifolia (Lam.) 0. Ktze. Dalechampia tiliifolia var. heterophylla 0. Ktze. 39 Dalechampia tiliifolia var. peruviana (Lam.) 0. Ktze. Dalechampia trichophila Pax & Hoffm. Dalechampia triphylla Lam 9 1 Dalechampia triphylla var. cissgolia (Poepp.) Muell. Arg. Dalechampia triphylla var. clausseniana (Baillon) Muell. Arg. Dalechampia triphylla var. mexicana Muell. Arg. Dalechampia triphylla var. subintegra Muell. Arg. Dalechampia triphylla var. uillosa Muell. Arg. f. pohliana Muell. Arg. Dalechampia triphylla var. uillosa f. riedeliana Muell. Arg. Dalechampia uleana Pax & Hoffm. 24 Dalechampia ulmifolia Chod. & Hassler Dalechampia ulmifolia var. grueningiana (Pax) Pax Dalechampia uahliana Steud. 49a Dalechampia variifolia Muell. Arg. Dalechampia uillosa Lam. 49a Dalechampia violacea Pax & Hoffm. Dalechampia viridissima Webster 52 Dalechampia uulpina Muell. Arg. 58 Dalechampia weberbaueri Pax & Hoffm. Dalechampia websteri Armbruster 18 Dalechampia weddelliana Baillon 74 Megalostylis Sp. Moore 1 Megalostylis poeppigii Sp. Moore 5
50 50 15
60
43
39 74
8 1 a 82
84 15
82 82
73 73
5 1
17
92















![Neotropical cervidology_12[1]](https://img.dokumen.tips/doc/110x75/547f4ab5b37959a22b8b56e0/neotropical-cervidology121.jpg)













