Embed Size (px)
Citation preview

NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999 907
ARTICLES
The ability to screen the blood supply for hepatitis B virus (HBV)and to prevent infection by protective vaccination has substan-tially reduced HBV transmission and the development of the car-rier state1. However, neither of these approaches is helpful to themore than 300 million HBV carriers worldwide who are at highrisk for the development of chronic hepatitis, cirrhosis and hepa-tocellular carcinoma. Interferon and/or lamivudine are beingused to treat carriers with chronic liver disease (CLD), althougheach of these drugs has its limitations2–4.
Part of the problem in understanding the pathogenesis ofchronic HBV infection and in testing new therapeutic ap-proaches is the lack of a suitable preclinical model. HBV has anarrow host range5. Naturally occurring HBV-like viruses inground squirrels, woodchucks and ducks6,7 have been useful forantiviral studies8,9, but much less so in understanding the patho-genesis of CLD. This is because none of these wild animal modelshave inbred strains necessary to facilitate the independent ma-nipulation of both host and virus to establish cause and effect.Transgenic mice expressing HBV genes10,11 or supporting virusreplication12,13 have been made, although all are tolerant to thetransgene, so that adoptive transfer of primed cells has resultedin acute or fulminant disease and not chronic hepatitis14,15.Intrahepatic injection of HBV DNA into nude mice resulted inCLD (ref. 16), demonstrating that chronic hepatitis is immune-mediated. Adoptive transfer of cytotoxic T lymphocytes specificfor hepatitis B surface antigen (HBsAg) into thymectomized, irra-diated and bone marrow-reconsitituted HBV transgenic mice re-sulted in the development of hepatocellular carcinoma17,confirming that the pathogenesis of hepatocellular carcinoma isimmune-mediated.
Here we created an HBV transgenic severe combined immun-odeficiency (SCID) mouse that consistently supports HBV geneexpression and replication. After adoptive transfer of syngeneic,unprimed splenocytes, these mice reproducibly cleared virusmarkers from liver and serum, and developed chronic hepatitis.
The development of hepatitis in a nontolerant host is analogousto that in chronic human infections, and provides an easily ma-nipulated laboratory-based model to elucidate the pathogenesisof CLD and to evaluate new approaches aimed at both the virusand disease.
HBV transgenic SCID mice replicate virusWe used the C.B-17 SCID mouse as the host strain to make trans-genic mice with the plasmid pTKHH2 (ref. 18). We testedfounder mice for the transgene by analysis of DNA obtainedfrom tail tissue. When mouse genomic DNA was digested withHindIII (which does not cleave HBV DNA), Southern blot analy-sis showed two bands (data not shown), indicating two sites ofintegration. Digestion of whole-cell DNA with EcoR1, whichcleaves HBV DNA once, resulted in a band with a size of 3 kb, in-dicating that the integrated transgene consisted of full-lengthHBV DNA (data not shown). To minimize the amount of DNAneeded to detect the transgene, we extracted whole-cell DNA,then amplified it by PCR using HBV core region primers, and an-alyzed the products by agarose gel electrophoresis and ethidiumbromide staining.
We then assessed HBV DNA in serum samples obtained fromtransgene-positive mice, by CsCl density equilibrium centrifuga-tion. After centrifugation, we assayed each gradient fraction forHBV DNA by PCR, followed by Southern blot analysis. A band ofthe expected size and specificity at a density of 1.28 was pro-duced, which is characteristic of virus particles obtained fromhuman serum (Fig. 1a). Parallel analyses of serum samples fromnontransgenic littermates yielded no PCR products (data notshown). We immunoprecipitated aliquots from HBV-positivegradient fractions with antibody against HBs, then extracted pel-lets and amplified DNA by PCR; HBV DNA was also produced(Fig. 1b). Immunoprecipitation with an irrelevant antibody re-sulted in no PCR products (data not shown). These PCR productswere also resistant to pretreatment of serum samples with DNase
Hepatitis B virus transgenic mouse model of chronic liver disease
JONATHAN LARKIN1, MARCY CLAYTON1, BILL SUN1, CLAIRE E. PERCHONOCK1,JUDITH L. MORGAN2 LINDA D. SIRACUSA2, FRANK H. MICHAELS2 & MARK A. FEITELSON1,2
1Department of Pathology, Anatomy and Cell Biology, Thomas Jefferson University, Room 222 Alumni Hall, 1020 Locust Street, Philadelphia, Pennsylvania 19107, USA
2Department of Microbiology and Immunology, Kimmel Cancer Center, Thomas Jefferson University, Philadelphia,Pennsylvania 19107, USA
J.L.M. present address: Fox Chase Cancer Center, 7701 Burholme Ave, Philadelphia, Pennsylvania 19111, USAF.H.M. present address: OraGen, 3600 Inverness Drive, Chevy Chase, Maryland 20815
Correspondence should be addressed to M.A.F.; email: [email protected]
A model for hepatitis B virus-associated chronic liver disease has been made using cloned he-patitis B virus DNA as a transgene in a severe combined immunodeficient host. These mice con-sistently support virus gene expression and replication. After adoptive transfer of unprimed,syngeneic splenocytes, these mice cleared virus from liver and serum, and developed chronicliver disease. This model will permit identification of the host and virus contributions to chronicliver disease in the absence of tolerance.
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m

Fig. 1 Evidence of viral replication in serum andliver of HBV transgenic SCID mice. a, Southern blotanalysis of PCR products derived from the serumsample of a transgenic mouse after density equilib-rium centrifugation. Far left, HBV-positive humanserum sample, which shows the expected core re-gion PCR product at 0.5 kb. b, Immunoprecipitationof virus-containing density equilibrium gradientfractions with antibody against HBs before PCR am-plification and Southern blot analysis of serum sam-ples from three transgenic mice (1–3) and a humancarrier (4). c, Southern blot analysis of whole-cell DNA isolated from theliver of a nontransgenic mouse (lane 1) and from two transgenic littermates(lanes 2 and 3). The replicative forms are ≤ 3 kb. d, Northern blot analysis of
908 NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999
ARTICLES
(data not shown). These results indicate that the transgenic SCIDmice have detectable virus in blood.
The presence of virus in blood is indicative of replication inthe liver. Thus, we extracted whole-cell DNA from the livers oftransgenic and nontransgenic mice and assessed it for replicativeforms by Southern blot analysis (Fig. 1c). Transgenic miceshowed growing replicative forms of HBV DNA in liver (Fig. 1c,lanes 2 and 3) similar to those in infected human and chim-panzee livers5,19,20. Nontransgenic littermates had no evidence ofviral replication (Fig. 1c, lane 1). As pregenomic RNA is also asso-ciated with replication, we did northern blot analysis usingwhole-cell RNA prepared from the livers of transgenic and non-transgenic mice. Pregenomic RNA at the expected size of 3.6 kbwas present in the livers of transgenic mice (Fig. 1d, lanes 2 and3) but not in a nontransgenic littermate (Fig. 1d, lane 1). Theblots also showed a major RNA at 2.0 kb, which probably en-codes the major HBsAg polypeptide21. These results provide fur-ther evidence of viral replication in these mice.
Consistent virus production in transgenic miceHBV in the blood of transgenic mice has been detected for sevengenerations, indicating stable transmission of a functional trans-gene (data not shown). For individual mice, quantitative PCRanalysis of serial serum samples showed that each mouse pro-duced a consistent baseline level of virus that did not changewith age (Fig. 2a). As the amount of virus was different in individ-ual mice, we determined the range of virus production and num-ber of mice producing different levels of virus (Fig. 2b). Aboutone-third of the mice had steady-state levels of HBV at about 1 × 106 virus genome equivalents per ml blood. Most mice in thecolony produced 1 × 105–1 × 107 virus genome equivalents per mlblood. To confirm these PCR results, we quantified virus DNA in200 µl of serum from randomly selected mice, by slot–blot hy-
bridization (Fig. 2c and d). The results are in complete agreementwith those of quantitative PCR. The reason for the different levelsof virus in individual mice is not known, but they did not corre-late with age or gender, nor did the mean levels of virus changewith subsequent generations of mice (data not shown). Thus, thetransgenic mouse colony consists of mice that produce consistentlevels of virus that are different for each mouse.
HBV gene expression in serum and liverThe immunoprecipitation of HBV DNA from the sera of trans-genic mice with antibody against HBs (Fig. 1b) indicates thatthese sera should also be positive for HBsAg. Serum samples fromrandomly chosen transgenic mice assessed by SDS–PAGE andwestern blot analysis with antibody against HBs (ref. 22) pro-duced a pair of bands at 25 and 29 kDa (Fig. 3a, lanes 2–5). Theseco-migrated with bands representing the major protein (25 kDa)and glycoprotein (gp29) of HBsAg from human sera (Fig. 3a, lane1). These polypeptides were not detected in the sera of nontrans-genic littermates (Fig. 3a, lane 6). Thus, HBsAg is present in theserum of HBV transgenic SCID mice.
To assess HBV gene expression in the liver, we killed 3-month-old mice with average levels of virus in blood (4 × 105–1 × 107
virus genome equivalents/ml) and stained their livers for HBsAg,hepatitis B core antigen (HbcAg) and HBxAg by immunofluores-cence. We detected cytoplasmic HBsAg in 30–40% of hepatocytesfrom most transgenic mice (Fig. 3b) but not in the livers of non-transgenic littermates (data not shown). HBcAg was strongly pos-itive in scattered hepatocellular nuclei from transgenic mice (Fig.3c and d) but not from nontransgenic littermates (data notshown). We also found cytoplasmic HBcAg in up to 30% of hepa-tocytes (Fig. 3d), which is consistent with virus replication23.Cytoplasmic HBxAg was detectable in more than 75% of hepato-cytes in most transgenic mice (Fig. 3e) but was undetectable inthe livers of nontransgenic littermates (Fig. 3f). HBxAg was notdetectable in transgenic livers stained with preimmune serum orstained with antibody against HBx preincubated with HBxAg be-
virus RNAs from a nontransgenic SCID mouse (lane 1), and from two differ-ent HBV transgenic SCID mice (lanes 2 and 3) using full-length HBV DNA asprobe. Left margin, molecular size markers.
Fig. 2 Concentration of HBV DNA in the sera of transgenic mice. a, Bloodobtained from five different transgenic mice (horizontal axis, age inmonths) and was assessed for the levels of virus DNA by semiquantitativePCR. Each line represents the levels of virus DNA in an individual mouse. b,Sera from each of 129 mice (about 3 months of age) were assayed for HBVDNA by semiquantitative PCR. c, Virus DNA from serum was measured byslot–blot analysis of 200 µl of serum from each of eight transgenic mice ran-domly chosen from b (slots 1–8) and from two nontransgenic littermates(slots 9 and 10). d, Hybridization with increasing dilutions of pTKHH2 gen-erated a standard curve (line); results from individual mice (�) in c werecompared with this curve.
b c d
Gradient fractions
Den
sity
of C
SCI
(g/m
l)
a
a b
c dNumber of mice
Gel scan (relative area)
Age of mice
Viri
ons/
ml ×
105
Log
of H
BVD
NA
/ml
Log
of H
BVD
NA
/ml
1 4 7 10
0.3 2.1 8 12
1 8 15 22 29 36
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m

Fig. 3 Detection of viral antigens in the serum and liver oftransgenic mice. a, Western blot analysis of HBsAg in sera. Lane1, HBsAg from a human carrier; lanes 2–5, four transgenic mice3 months old; lane 6, 3-month-old nontransgenic littermate.b–f, Immunoflorescent staining for HBV antigens in transgenicSCID mouse livers. Liver sections from transgenic mice werestained with antibody against HBs (b) or HBc (c and d). Liversections from a transgenic mouse (e) or a nontransgenic litter-mate (f) were stained with antibody against HBx. Original mag-nifications: ×200 (b and c), ×400 (d) and ×100 (e and f). Theliver tissues in b–e were derived from mice with 2.3 × 106–5.8 ×106 virus genome equivalents/ml of blood. The results are rep-resentative of mice with about 4 × 105–1 × 107 virus genomeequivalents/ml in their blood.
NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999 909
ARTICLES
fore staining (data not shown). The staining patterns of intrahep-atic HBsAg, HBcAg and HBxAg were the same in tissues evaluatedat 9 months of age (data not shown; analogous to Fig. 3). Thesedata show that HBV antigens are consistently expressed in pat-terns similar to those in the livers of human carriers23–25.
Extrahepatic HBV gene expression and replicationGiven that there are extracellular sites of HBV infection26, we didnorthern blot analysis and immunohistochemical staining ofkidneys, hearts, brains, pancreata, skeletal muscles, lungs,spleens and peripheral blood lymphocytes from transgenic mice.Reproducibly detectable virus transcripts and proteins werefound only in the kidney (data not shown). Only trace amountsof HBV transcripts (in heart, pancreas and brain), and variable,low levels of antigen (HBsAg in heart and pancreas) were foundin other organs (data not shown). Thus, HBV expression occursin several extrahepatic sites, as reported in human infections26
and in other transgenic models of hepatitis B (refs. 10,11,27).
Chronic liver disease in adoptively transferred miceWe sought to determine whether immunological reconstitutionresulted in CLD. Transgenic mice and nontransgenic littermateswere subjected to partial hepatectomies, allowed to recover, and
then adoptively transferred with 1 × 107 unprimed, syngeneicsplenocytes. We injected additional transgenic mice with anequal volume of saline. We then determined alanine amino-transferase (ALT) values in serial serum samples, and 35 weekslater, killed the mice and removed the livers for further analysis.Among transgenic mice, half developed increased ALT within 6weeks of adoptive transfer (Fig. 4a). By week 10, ALT values wereincreased in all adoptively transferred transgenic mice, and thispersisted, with fluctuations, for at least another 6 months (Fig.4a). None of the transgenic mice injected with saline or adop-tively transferred nontransgenic mice developed ALT levels morethan 300% above baseline at any time. Thus, a single adoptivetransfer of 1 × 107 unprimed, syngeneic splenocytes into trans-genic mice resulted in the development and persistence of in-creased ALT in serum, which is consistent with underlying CLD.
Adoptive transfers into transgenic mice were also accompa-nied by the clearance of HBsAg and viral DNA from serumwithin a few weeks. Levels of viral DNA often decreased to0.001% of baseline levels within 2 weeks of adoptive transfer,and the clearance of HBsAg was often accompanied by serocon-version to antibody against HBs (Fig. 4b and c). However, HBVDNA often reappeared in one or more serum samples collected atlater time points in mice with persistently increased ALT (Fig. 4b
and c). Antibody responses against the core, Xand/or polymerase gene products were also ob-served in some mice. Nontransgenic littermatesadoptively transferred with splenocytes hadnormal ALT values and no virus markersthroughout the observation period (Fig. 4d).Among the ten transgenic mice adoptivelytransferred, all cleared HBsAg and serocon-verted to antibody against HBs within the first
Fig. 4 ALT and HBV markers in the blood of adoptivelytransferred mice. a, Average ALT values over time in tentransgenic mice (�–�) and ten nontransgenic littermates(�–�) adoptively transferred with 1 × 107 normal, syn-geneic splenocytes, or in transgenic mice injected withPBS (�–�). Each curve shows the average ALT enzyme ac-tivity in units/liter. b–d, ALT (�–�) and viral DNA in serum(�– –�) and serological profiles (chart under graphs) oftwo transgenic mice (b and c) and a nontransgenic litter-mate (d) (time, horizontal axis). anti-pol, antibody againstviral polymerase.
a b c
d e f
a
b c dWeeks after adoptive transfer
ALT
act
ivity
[uni
ts/I
]
ALT
act
ivity
[uni
ts/I
(�–
�)]
Log of H
BVD
NA
( �--�
)
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m
910 NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999
ARTICLES
2–4 weeks after transfer. In nine of these, HBV DNA became un-detectable by PCR within the first 6 weeks after adoptive trans-fer, but viral DNA reappeared later in eight of these. Half of themice developed antibodies against HBc, 40% developed antibod-ies against the viral polymerase and 30% developed antibodiesagainst HBxAg. Thus, adoptive transfer of normal splenocytesinto transgenic mice results in increased ALT, in the clearance ofvirus markers and in the development of virus antibodies.
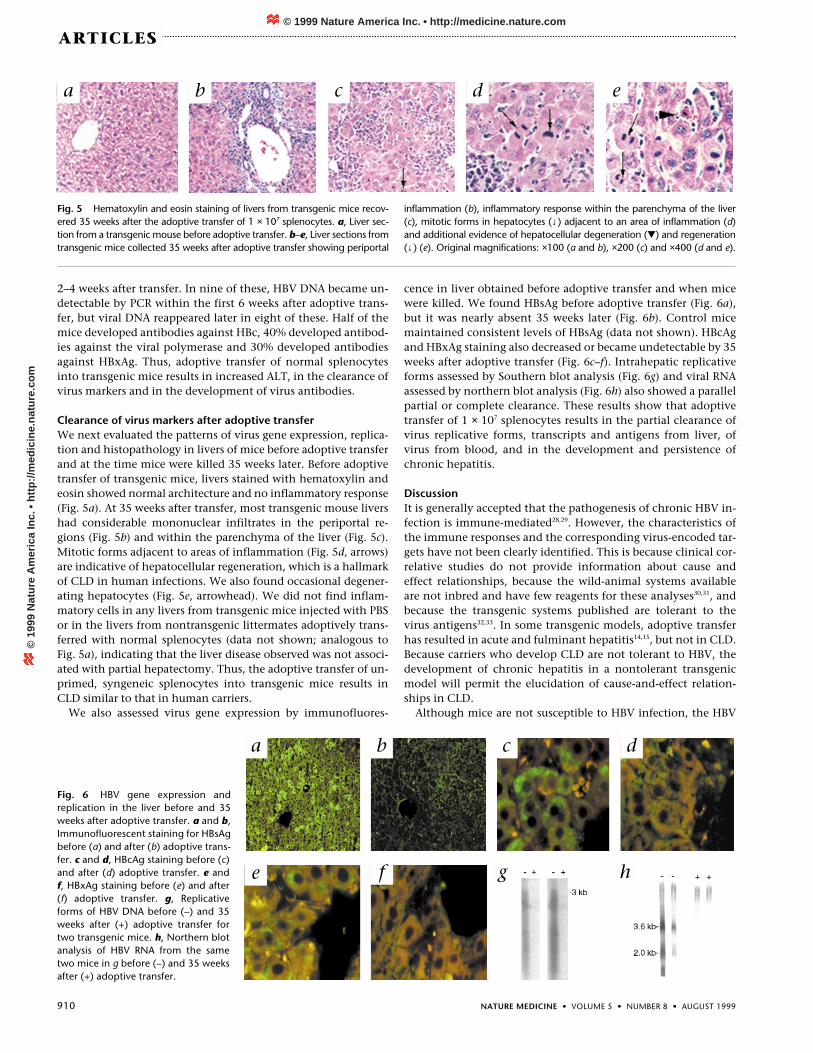
Clearance of virus markers after adoptive transferWe next evaluated the patterns of virus gene expression, replica-tion and histopathology in livers of mice before adoptive transferand at the time mice were killed 35 weeks later. Before adoptivetransfer of transgenic mice, livers stained with hematoxylin andeosin showed normal architecture and no inflammatory response(Fig. 5a). At 35 weeks after transfer, most transgenic mouse livershad considerable mononuclear infiltrates in the periportal re-gions (Fig. 5b) and within the parenchyma of the liver (Fig. 5c).Mitotic forms adjacent to areas of inflammation (Fig. 5d, arrows)are indicative of hepatocellular regeneration, which is a hallmarkof CLD in human infections. We also found occasional degener-ating hepatocytes (Fig. 5e, arrowhead). We did not find inflam-matory cells in any livers from transgenic mice injected with PBSor in the livers from nontransgenic littermates adoptively trans-ferred with normal splenocytes (data not shown; analogous toFig. 5a), indicating that the liver disease observed was not associ-ated with partial hepatectomy. Thus, the adoptive transfer of un-primed, syngeneic splenocytes into transgenic mice results inCLD similar to that in human carriers.
We also assessed virus gene expression by immunofluores-
cence in liver obtained before adoptive transfer and when micewere killed. We found HBsAg before adoptive transfer (Fig. 6a),but it was nearly absent 35 weeks later (Fig. 6b). Control micemaintained consistent levels of HBsAg (data not shown). HBcAgand HBxAg staining also decreased or became undetectable by 35weeks after adoptive transfer (Fig. 6c–f). Intrahepatic replicativeforms assessed by Southern blot analysis (Fig. 6g) and viral RNAassessed by northern blot analysis (Fig. 6h) also showed a parallelpartial or complete clearance. These results show that adoptivetransfer of 1 × 107 splenocytes results in the partial clearance ofvirus replicative forms, transcripts and antigens from liver, ofvirus from blood, and in the development and persistence ofchronic hepatitis.
DiscussionIt is generally accepted that the pathogenesis of chronic HBV in-fection is immune-mediated28,29. However, the characteristics ofthe immune responses and the corresponding virus-encoded tar-gets have not been clearly identified. This is because clinical cor-relative studies do not provide information about cause andeffect relationships, because the wild-animal systems availableare not inbred and have few reagents for these analyses30,31, andbecause the transgenic systems published are tolerant to thevirus antigens32,33. In some transgenic models, adoptive transferhas resulted in acute and fulminant hepatitis14,15, but not in CLD.Because carriers who develop CLD are not tolerant to HBV, thedevelopment of chronic hepatitis in a nontolerant transgenicmodel will permit the elucidation of cause-and-effect relation-ships in CLD.
Although mice are not susceptible to HBV infection, the HBV
Fig. 5 Hematoxylin and eosin staining of livers from transgenic mice recov-ered 35 weeks after the adoptive transfer of 1 × 107 splenocytes. a, Liver sec-tion from a transgenic mouse before adoptive transfer. b–e, Liver sections fromtransgenic mice collected 35 weeks after adoptive transfer showing periportal
inflammation (b), inflammatory response within the parenchyma of the liver(c), mitotic forms in hepatocytes (↓) adjacent to an area of inflammation (d)and additional evidence of hepatocellular degeneration () and regeneration(↓) (e). Original magnifications: ×100 (a and b), ×200 (c) and ×400 (d and e).
Fig. 6 HBV gene expression andreplication in the liver before and 35weeks after adoptive transfer. a and b,Immunofluorescent staining for HBsAgbefore (a) and after (b) adoptive trans-fer. c and d, HBcAg staining before (c)and after (d) adoptive transfer. e andf, HBxAg staining before (e) and after(f) adoptive transfer. g, Replicativeforms of HBV DNA before (–) and 35weeks after (+) adoptive transfer fortwo transgenic mice. h, Northern blotanalysis of HBV RNA from the sametwo mice in g before (–) and 35 weeksafter (+) adoptive transfer.
a b c d e
a b c d
e f g h
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m

NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999 911
ARTICLES
transgenic SCID mouse model has many characteristics similar tothose in human infections. These include the development ofCLD and partial clearance of virus markers from the blood andliver of transgenic mice after adoptive transfer, which seems to re-flect in vivo recognition and priming of immunocompetent cellsto virus antigens (Figs. 4–6), as in human infections29,34. The per-sistence and fluctuations of ALT in transgenic mice over nearly 9months of observation (Fig. 4) also resembles the recurrent boutsof CLD characteristic of patients who eventually develop cirrho-sis35,36. So far, we have not found cirrhosis and/or hepatocellularcarcinoma in adoptively transferred transgenic mice, even after14 months (of their approximately 18-month life span). The per-sistence of increased ALT long after the clearance of virus fromthe blood of the transgenic mice (Fig. 4) is analogous to the per-sistence of CLD in patients who have seroconverted from hepati-tis B envelope antigen (HBeAg) to antibody against HBe (refs.29,34,36). Hallmarks of CLD also include degenerating hepato-cytes (Fig. 5e) and mitotic figures among hepatocytes (Fig. 5d ande), indicating hepatocellular regeneration. These features willcontribute to the identification of the lymphocyte subsets andthe corresponding virus antigens that serve as targets for im-munological recognition and attack during the course of CLD.
Both HBV carriers and transgenic mice with CLD expressHBcAg in hepatocellular nuclei and cytoplasm, and expressHBxAg in a mostly cytoplasmic location in hepatocytes23–25 (Fig.3). Both clear HBV DNA and virus antigens from serum and liverduring the course of CLD (Figs. 4 and 6)(refs. 28,29,35). This in-dicates that in vivo priming of both cellular and humoral im-mune responses have taken place. Future work will characterizethe host–virus relationship that results from adoptive transfersof lymphocyte subsets or with virus antigen primed splenocytes.In this way, cause-and-effect relationships relevant to CLD willbe elucidated; this will form the basis from which antiviral andimmunomodulatory approaches directed against CLD will beevaluated. In addition, the model established here can be used asan example for the construction of other systems aimed at un-derstanding the immune-mediated pathogenesis of infectiousand autoimmune diseases in other organ systems.
MethodsConstruction of transgenic mice. The plasmid pTKHH2, which contains ahead-to-tail EcoR1 dimer of HBV DNA and is infectious in chimpanzees18,was used to make transgenic mice. pTKHH2 was partially digested withEcoR1 and the HBV dimer was resolved by agarose gel electrophoresis. TheHBV DNA dimer (6.4 kb) was then transferred to a DEAE membrane, eluted,extracted with phenol/chloroform and then with chloroform, and finallyprecipitated with ethanol. The extraction and precipitation were then re-peated. The pTKHH2 insert was then microinjected into two strains of miceat concentrations of 1 ng/µl and 5 ng/µl. All microinjected embryos wereimplanted into pseudopregnant Swiss Webster female mice (TaconicFarms, Germantown, New York). B6C3F1 and C.B-17 SCID female micewere superovulated as described37 and mated to males of the same strains.Twenty B6C3F1 females released 674 eggs, 394 of which were fertilizedand microinjected; 264 fertilized eggs survived microinjection. After beingcultured overnight, 238 embryos were implanted, and 13 of 64 livebornoffspring carried the transgene. In contrast, C.B-17 SCID females were poorsuperovulators, with 118 C.B-17 SCID females releasing 1875 eggs, 690 ofwhich were fertilized. C.B-17 SCID fertilized eggs were fragile; of the 637microinjected, only 256 survived. Half of the embryos were culturedovernight before implantation and half were implanted directly after mi-croinjection. Of the 219 embryos implanted, only 1 of 15 liveborn offspringcarried the transgene. This single C.B-17 SCID transgenic founder was theprogenitor of all mice analyzed for this research.
Whole-cell DNA extracted from tail tissues of founder mice (F0) wasscreened for the transgene by Southern blot analysis of undigested and
EcoR1-digested samples or by PCR amplification. Mice with the HBV trans-gene were tested for virus in blood. Mice with the virus were mated toC3H/SCID mice (C3H SMN.C SCID; The Jackson Laboratory, Bar Harbor,Maine). Littermates with the virus were mated to each other to establish atransgenic colony.
Detection of the transgene. DNA was extracted from mouse tail tissue38
and 1 µg was analyzed for the HBV transgene by PCR using HBV core regionprimers, as described39. Samples (10 µl) of amplified products were ana-lyzed by 1.5% agarose gel electrophoresis in TAE buffer with 0.5 µg ethid-ium bromide/ml.
Detection of HBV DNA in serum. Virion DNA in blood of mice was de-tected by semiquantitative PCR. About 100 µl of blood drawn from theretro-orbital plexus was allowed to clot, and serum was collected after cen-trifugation at 6,610g for 10 min at 4 °C. Then, 25 µl of serum from eachmouse was digested with DNase 1 at 37 °C for 30 min. The DNase was in-activated and the samples were digested with proteinase K. Each samplewas passed through a Qiamp column (Qiagen, Chatsworth, California) andeluted in 200 µl of nuclease-free water. Semiquantitative PCR used 6-µlaliquots of isolated DNA template containing 10-fold dilutions of DNA am-plified with the same primers but smaller than the HBV amplicon (‘spike’) incompetitive PCR reactions, as described39. PCR products were separated byagarose gel electrophoresis as described above. Semiquantitation was ac-complished by densitometry scans of a photographic negative image froman ethidium bromide-stained gel in which lanes had equal amounts of am-plified samples and ‘spike’.
Slot–blot analysis of HBV DNA from serum. DNA purified from 200 µl ofserum as described above was applied to a nylon membrane (Schleicher& Schuell, Keene, New Hampshire) through a slot–blot apparatus(BioRad, Hercules, California). Samples were hybridized in stringent con-ditions38 to a full-length, HBV DNA probe radiolabeled with α-32P-ATP byrandom priming.
Immunoflorescence. Liver tissues were fixed in 10% buffered formalin(Fisher) and embedded in paraffin, and sections 5 µm in thickness weremounted on poly-L-lysine coated slides. Slides were deparaffinized and in-cubated with (normal pig) blocking serum diluted 1:100 in PBS for 20 minat room temperature. Slides were washed twice for 5 min in PBS and incu-bated with primary antibody for 1 h at room temperature. The primary an-tibodies, against HBs or HBc, were used according to manufacturer’sinstructions (Dako, Carpinteria, California). The antibody against HBx wasused according to published procedures40. Slides were rinsed as describedabove and incubated with FITC-conjugated secondary antibody against IgGfor 30 min at room temperature. After being washed and mounted, slideswere visualized using an epifluorescent microscope (Bx60 UniversalFlorescent Microscope; Olympus, Melville, New York) equipped with FITCfilters and a camera (DKC 5000 Sony 3 CCD; Sony, New York, New York).
Partial hepatectomy and adoptive transfers. Blood was obtained fromtransgenic mice and nontransgenic littermates before they were subjectedto a 30–40% partial hepatectomy41,42. Hemoclips (Weck, Research Triangle,North Carolina) were used for ligation of the remaining liver tissue.Recovery time was 1 week, to allow for complete regeneration of liver mass.Ten transgenic mice were adoptively transferred with 1 × 107 normalsplenocytes from syngenic donors (C3H/CB17 F1) by injection of the tailvein. Another ten transgenic mice were injected with an equal volume ofsaline. Groups of nontransgenic littermates adoptively transferred with 1 ×107 normal splenocytes were analyzed in parallel. Serial serum samples werecollected weekly and assayed for HBV by semiquantitative PCR. ALT levelswere determined in serum samples by measuring enzyme activity with acommercial kit (ALT/AST 50; Sigma). Mice were killed 35 weeks after adop-tive transfer, and their livers were removed for further analysis.
AcknowledgmentsThe authors thank C. Calkins and L. Eisenlohr for discussions, and J. Song forassisting in screening mice for virus. This work was supported by NIH grantsCA48656 and CA66971, and by a grant from the Alcoholic Beverage Medical
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m

912 NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999
ARTICLES
Research Foundation to M.A.F. This work was also supported by a pilot projectfrom AA07186 to E. Rubin. The Kimmel Cancer Center Transgenic andKnockout Mouse Facility is supported, in part, by NIH grant CA56036.
RECEIVED 16 APRIL; ACCEPTED 7 JUNE 1999
1. Blumberg, B.S. The current state of the prevention of HBV infection, the carrierstate and hepatocellular carcinoma. Res. Virol. 148, 91–94 (1997).
2. Hoofnagle, J.H., & Lau, D. New therapies for chronic hepatitis B. J. Viral Hepat. 4,41–50 (1997).
3. Saracco, G., & Rizzetto, M. A practical guide to the use of interferons in the man-agement of hepatitis virus infections. Drugs 53, 74–85 (1997).
4. Honkoop, P., Niesters, H.G., de Man, R.A., Osterhaus, A.D., & Schalm, S.W.Lamivudine resistance in immunocompetent chronic hepatitis B. Incidence andpatterns. J. Hepatol. 26, 1393–1395 (1997).
5. Pugh, J.C., & Bassendine, M.F. Molecular biology of hepadnavirus replication.British Med. Bull. 46, 329–353 (1990).
6. Robinson, W.S., Marion, P., Feitelson, M.A., & Siddidjui, A. in Viral Hepatitis,1981 International Symposium. (eds. Szmuness, W., Alter, H.J., & Maynard, J.E.)57–68 (Franklin Institute Press, Philadelphia, 1982).
7. Summers, J. Three recently described animal virus models for hepatitis B virus.Hepatology 1, 179–183 (1981).
8. Mason, W.S. et al. Characterization of the antiviral effects of 2’ car-bodeoxyguanosine in ducks chronically infected with duck hepatitis B virus.Hepatology 19, 398–411 (1994).
9. Korba, B.A. et al. Liver-targeted antiviral nucleosides: enhanced antiviral activityof phosphatidyl-dideoxyguanosine versus dideoxyguanosine in woodchuck he-patitis virus infection in vivo. Hepatology 23, 958–963 (1996).
10. Babinet, C., Farza, H., Morello, D., Hadchouel, M., & Pourcel, C. Specific expres-sion of hepatitis B surface antigen (HBsAg) in transgenic mice. Science 230,1160–1163 (1985).
11. Chisari, F.V. et al. A transgenic mouse model of the chronic hepatitis B surfaceantigen carrier state. Science 230, 1157–60 (1985).
12. Farza, H. et al. Replication and gene expression of hepatitis B virus in a transgenicmouse that contains the complete viral genome. J. Virol. 62, 4144–4152 (1988).
13. Araki, K. et al. Expression and replication of hepatitis B virus genome in trans-genic mice. Proc. Natl. Acad. Sci. USA 86, 207–211 (1989).
14. Moriyama, T. et al. Immunobiology and pathogenesis of hepatocellular injury inhepatitis B virus transgenic mice. Science 248, 361–364 (1990).
15. Ando, K. et al. Mechanisms of class I restricted immunopathology. A transgenicmouse model of fulminant hepatitis. J. Exp. Med. 178, 1541–1554 (1993).
16. Feitelson, M.A., DeTolla, L,J. & Zhou, X.D. A chronic carrierlike state is estab-lished in nude mice injected with cloned hepatitis B virus DNA. J. Virol. 62,1408–1415 (1988).
17. Nakamoto, Y., Guidotti, L.G., Kuhlen, C.V., Fowler, P., & Chisari, F.V. Immunepathogenesis of hepatocellular carcinoma. J. Exp. Med. 188, 341–350 (1998).
18. Will, H. et al. Cloned HBV DNA causes hepatitis in chimpanzees. Nature 299,740–742 (1982).
19. Blum, H.E., Haase A.T., Harris J,D., Walker, D. & Vyas, G.N. Asymmetric replica-tion of hepatitis B virus DNA in human liver: demonstration of cytoplasmicminus-strand DNA by blot analyses and in situ hybridization. Virology 139, 87–96(1984).
20. Ruiz-Opazo, N., Chakraborty, P.R. & Shafritz, D.A. Characterization of viralgenomes in the liver and serum of chimpanzee long-term hepatitis B virus carri-
ers: a possible role for supercoiled HBV DNA in persistent HBV infection. J. Cell.Biochem. 19, 281–292 (1982).
21. Tiollais, P., Pourcel, C. & Dejean A. The hepatitis B virus. Nature 317, 489–495(1985).
22. Lerner, R.A. et al. Chemically synthesized peptides predicted from the nucleotidesequence of the hepatitis B virus genome elicit antibodies reactive with the nativeenvelope protein of Dane particles. Proc. Natl. Acad. Sci. USA 78, 3403–3407(1981).
23. Ray, M.B. Hepatitis B Virus Antigens in Tissues (University Park Press, Baltimore,Maryland, 1979).
24. Wang, W., London, W.T., Lega, L. & Feitelson, M.A. Hepatitis B x antigen in liverfrom carrier patients with chronic hepatitis and cirrhosis. Hepatology 14, 29–37(1991).
25. Wang, W., London, W.T. & Feitelson, M.A. Hepatitis B x antigen in hepatitis Bvirus carrier patients with liver cancer. Cancer Res. 51, 4971–4977 (1991).
26. Omata, M. Significance of extrahepatic replication of hepatitis B virus. Hepatology12, 364–366 (1990).
27. Guidotti, L.G., Matzke, B., Schaller, H. & Chisari, F.V. High level hepatitis B virusreplication in transgenic mice. J. Virol. 69, 6158–6169 (1995).
28. Dienstag, J.L. in Viral Hepatitis and Liver Disease (eds. Vyas, G.N., Dienstag, J.L., &Hoofnagle, J.H.) 135–166 (Grune & Stratton, New York, 1984).
29. Feitelson, M.A. Hepatitis B virus gene products as immunological targets inchronic infection. Mol. Biol. Med. 6, 367–393 (1989).
30. Shanmuganathan, S., Waters, J.A., Karayiannis, P., Thursz, M. & Thomas, H.C.Mapping of the cellular immune responses to woodchuck hepatitis core antigenepitopes in chronically infected woodchucks. J. Med. Virol. 52, 128–135 (1997).
31. Lohrengel, B., Lu, M. & Roggendorf, M. Molecular cloning of the woodchuck cy-tokines: TNF-alpha, IFN-gamma, and IL-6. Immunogenetics 47, 332–335 (1998).
32. Milich, D.R. et al. Autoantibody production in hepatitis B e antigen transgenicmice elicited with a self T-cell peptide and inhibited with nonself peptides. Proc.Natl. Acad. Sci. USA 88, 4348–4352 (1991).
33. Wirth, S., Guidotti, L.G., Ando, K., Schlicht, H.J. & Chisari, F.V. Breaking toleranceleads to autoantibody production but not autoimmune liver disease in hepatitis Bvirus envelope transgenic mice. J. Immunol. 154, 2504–2515 (1995).
34. Thomas, H.C. The hepatitis B virus and the host response. J. Hepatol. 11(Suppl 1),S83–S89 (1990).
35. Hoofnagle, J.H., & Alter, H.J. in Viral Hepatitis and Liver Disease (eds. Vyas, G.N.,Dienstag, J.L., & Hoofnagle, J.H.) 97–114 (Grune & Stratton, New York, 1984).
36. Hoofnagle, J.H., & Schafer, D.F. Serologic markers of hepatitis B virus infection.Semin. Liver Dis. 6, 1–10 (1986).
37. Hogan, B., Beddington, R., Costantini, F., & Lacy, E. in Manipulating the MouseEmbryo: A Laboratory Manual 2nd edn. (Cold Spring Harbor Press, Cold SpringHarbor, New York, 1994).
38. Sambrook, J., Fritsch E.F., & Maniatis, T. in Molecular Cloning: A LaboratoryManual 2nd edn. (Cold Spring Harbor Press, Cold Spring Harbor, New York,1989).
39. Feitelson, M.A. et al. Pathogenesis of post-transfusion viral hepatitis in childrenwith β-thalassemia. Hepatology 19, 558–568 (1994).
40. Feitelson, M.A. & Clayton, M.M. The X antigen/antibody markers in hepad-navirus infections. II. Antibodies to the X gene product(s). Gastroenterology 99,500–507 (1990).
41. Schaeffer, D.O., Hosgood, G., Oakes, M.G., St. Amant, L.G. & Koon, C.E. An al-ternative technique for partial hepatectomy in mice. Lab. Animal Sci. 44, 189–190(1994).
42. Waynforth, H.B. in Experimental and Surgical Techniques in the Rat 2nd edn.(Academic, London, 1992).
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m





























