Embed Size (px)
Citation preview

BRAINRESEARCH
BrainResearch732(1996)145-153
Research report
5-HTIAand benzodiazepine receptors in the basolateral amygdala modulateanxiety in the social interaction test, but not in the elevated plus-maze
Luis E. Gonzalez, Nick Andrews, Sandra E. File *Psychopharmacology Research Unit, UMDS Diuision of Pharmacology, Guy’s Hospital, London SE1 9RT, UK
Accepted23April 1996
Abstract
In order to investigate the role of the 5-HTIAreceptors of the amygdala in modulating anxiety, rats were implanted with bilateralcannulae aimed at the basolateral nucleus of the amygdala complex and infused with either artificial cerebrospinal fluid (aCSF) or theselective 5-HTIA receptor agonist 8-OH-DPAT (50–200 ng) and tested in two animal models of anxiety. In the elevated plus-maze test,no significant effects were detected in this dose range. In contrast, 8-OH-DPAT caused an overall reduction in levels of socialinvestigation, thus indicating anxiogenic actions in the social interaction test. At 50 ng, 8-OH-DPAT had a selective action on anxiety,while at 200 ng there was a concomitant reduction in locomotor activity and, in some animals, signs of the 5-HTIA syndrome. Evidencethat the rmxiogenic effect of 8-OH-DPAT(50 ng) was duetoactivatbnof 5-HTIA receptors came from the finding that ( – )-tertatolol, a5-HTIA receptor antagonist, reversed this effect at a dose (1.5 ~g) which was silent when given alone. The benzodiazepine receptoragonist, midazolam (1 and 2 ~g) was bilaterally administered into the basolateral nucleus of the amygdala and evoked clear-cut anxiolyticeffects in the social interaction test. These data indicate that the agonist activation of post-synaptic 5-HTIA receptors in the basolateralnucleus of the amygdala may produce anxiogenic effects, while agonist activation of BDZ receptors in the same areas evokes anxiolyticeffects. Our results from the social interaction test are similar to those previously reported from tests of anxiety using punished paradigms,but contrast with those found in the elevated plus-maze. Thus, it is concluded that either the two tests have different sensitivities tomidazolam and 8-OH-DPAT or more intriguingly, the tests are evoking fundamentally different states of anxiety, with that evoked by theplus-maze being mediated via brain areas or receptors different from those studied here.
Keywords: 5-HTIA; Benzodiazepine;Amygdala;8-OH-DpAT;Midwol~; plus-maze;Socialinteraction
1. Introduction
The amygdala has been critically implicated in themodulation of fear and anxiety [8]. For example,elec-trolytic or N-methyl-D-aspartate lesions of central or lateralnuclei of the arnygdala have been reported to block orreduce anxiety expression [22,27], while electrical stimula-tion of neurons within the arnygdaloid complex elicitclear-cut signs of fear in many animals including humans[26,7]. Furthermore, electrophysiological studies haveshown that both unconditioned and conditioned aversivestimuli can evoke neural activity in the amygdala [3,15,32]and mRNA levels of c-jiis proto-oncogene were increasedin the amygdala during conditioned and unconditioned fearsituations [5].
Serotonin (5-hydroxytryptamine; 5-HT) release has beenshown to be increased in the amygdaloid complex in the
*Correspondingauthor.Fax: +44 (171)955-4627.
anxiogenic situation of diazepam withdrawal [9], and in-creases in extracellular 5-HT have been detected in thebasolateral nucleus of the amygdala as measured by micro-dialysis techniques during stressful situations [20]. Admin-istration of 5-HT directly into the amygdala has beenshown to evoke anxiogenic effects as measured in thesocial interaction test [16] and in a punished paradigm [17].The amygdala is a main projection site from the dorsalraph6 nucleus [4,21], and it is now widely held thatneuronal inhibition by different manipulations of the sero-tonergic neurones from the dorsal raph6 nucleus leads toconsistent anxiolytic effects across several animal models(for review see [14]). All these findings indicate thatpost-synaptic serotonergic receptors in the amygdala maymodulate anxiety.
Although there is a relatively high density of 5-HTIAreceptors within the amygdaloid complex [6,33], the roleof these receptors in modulating anxiety has been ratherscarcely investigated. Hodges et al. [17] reported that
0006-8993/96/$15.00 Copyright@ 1996ElsevierScienceB.V. All rights reserved.PII S0006-8993(96)005 17-3

146 L.E. Gonzalez et al, /Brain Research 732 (1996) 145-153
infusions of the 5-HTIA receptor agonist 8-OH-DPAT (125or 500 ng/O.5 pl) into the lateral and basolateral nuclei ofthe amygdala decreased punished responding, thus indicat-ing anxiogenic actions. Conversely, Takao et al. [30],examined the effects of two 5-HT1~ receptor agonists,buspirone and d-AP159 (N-cyclohexyl-1,2,3,4-tetrahydro-benzo(b)thieno-(2,3c)pyridine)-3-carboarnide HC1) wheninjected into the central nucleus of the amygdala; d-AP159but not buspirone (a partial agonist) had anticonflict activ-ity. This suggests that the activation of 5-HTIA receptors inbasolateral and central nuclei may have opposite conse-quences for anxiety. However, there is not enough evi-dence to support the specificity of such effects. Thus, theactions of 5-HTIA ligands have not been replicated inthose or other animal tests of anxiety and it has not yetbeen shown whether the effects of direct infusions of5-HT1~ receptor agonists are blocked by 5-HTIA receptorantagonists.
In the following experiments we investigated the effectsof 8-OH-DPAT when injected into the basolateral nucleusof the amygdala in the social interaction and the elevatedplus-maze, two ethnologicallyderived tests of anxiety [11].The dose range of 8-OH-DPAT was selected on the basisof those doses found to be active in previous studies [1];the highest dose (200 ng) was determined by its volubilityin aCSF. The low light test condition of the social interac-tion test was used since this is the most sensitive toanxiogenic effects [10].
We used (–)-tertatolol as a selective 5-HTIA receptorantagonist [18], since it is soluble in artificial cerebrospinalfluid (aCSF) at pH 7.4 and, at the time of study, WAY100635 was not available. The dose of (–)-tertatolol (1.5Kg) was selected based on the findings of a previous study[2].
Drugs acting at the GABA–benzodiazepine receptorcomplex have consistent anxiolytic effects in conflictparadigms when administered into the arnygdaloid com-plex [17,28,31], and such effects have also been found inthe social interaction test [16]. However, there is no clearagreement regarding the relative importance of the centraland basolateral nuclei of the amygdala. For example, whileShibata et al. [29], using the Geller-Seifter conflictparadigm, found anticonflict effects of benzodiazepineswhen injected into the central but not the basolateralnucleus, Hodges et al. [17] found significant anticonflicteffects of benzodiazepines injected into the basolateralareas using the same paradigm. The lack of specific histo-logical procedures to identify nuclei boundaries may ac-count for these discrepancies. We therefore examinedwhether bilateral administration of midazolam to the baso-lateral nucleus of the amygdala was able to increase socialinvestigation, indicating an anxiolytic effect, and haveexamined the injection sites using an acetylcholinesterasestaining procedure which delineates the basolateral nucleusfrom the central nucleus of the arnygdala. Although Hig-gins et al. [16] used high light, unfamiliar test conditions,
the high light familiar condition was selected for this studyto determine whether high light was a sufficient aversivefactor in making the test sensitive to benzodiazepines.
2. Materials and methods
2.1. Animals
Male hooded Lister rats weighing 200–300 g (HarlanUK) were housed singly following surgery and allowed torecover for 7 days prior to behavioral testing. Food andwater were freely available and the room in which theywere housed was lit with dim light and maintained at 22”C.Lights were on from 07.00–19.00 h. In order to accustomthe animals to handling and to keep the cannulae patent,each day following surgery the rats were gently wrappedin a cloth and the stylets replaced. All the experimentalgroups had 12 animals per group before surgery. The finalgroup sizes reflect the loss of animals post-operatively dueto displacement of the cannula mounting, blockade of thecannula and the results of the histological analysis.
2.2. Apparatus
The plus-maze was made of wood and consisted of twoopposite open arms 50 X 10 cm, and two opposite armsenclosed by 40 cm high walls. The arm were connected bya central 10 X 10 cm square, and thus the maze formed a‘plus’ shape. The maze was elevated 50 cm from the floorand lit by dim light. A closed-circuit TV camera wasmounted vertically over the maze and the behaviour wasscored from a monitor in an adjacent room. All scoreswere entered directly into a IBM computer. Changes in thepercentage of time spent on the open arms indicate changesin anxiety, and number of closed arm entries is a reliablemeasure of locomotor activity [11].
The social interaction test arena was a wooden box60 X 60 cm, with 35 cm high walls and was lit by lowlight (30 Iux). The high light (350 IUX)was used only inExpt. 4. A camera was mounted vertically above the arenaand the rats were observed on a monitor in an adjacentroom. Test sessions were also video recorded for subse-quent analysis. Infrared photocells were mounted in thewalls, 4.5 and 12 cm from the floor and the interruption ofthese beams provided automated measures of locomotoractivity and rearing, respectively. Since there were reduc-tions in these automated measures following intra-amyg-daloid injections of 8-OH-DPAT that nearly reached sig-nificance, the motor activity of the operated animal alonewas also assessed retrospectively from the video film. Thesocial interaction test arena was therefore divided into ninesquares of 20 cm2 each by placing a grid over the videoscreen. The number of line crossings and rears made bythe injected animal were scored by an observer blind to thedrug treatment.

L.E. Gonzalez et al. /Brain Research 732 (1996) 145-153 147
2.3. Drugs and chemicals
(+)-8-OH-DPAT hydrobromide (Research Biochemi-cal, UK) and (– )tertatolol hydrochloride (1.R.I.S., France)were dissolved in artificial cerebrospinal fluid (aCSF) ofthe following composition (mM): NaCl 126.6, NaHCO~27.4, KC1 2.4, KHZPO1 0.5, CaClz 0.89, MgClz 0.8,Naz HPO1 0.48 and glucose 7.1; pH 7.4. All drug concen-trations reflect base weight. Midazolam maleate (RocheProducts) was dissolved in saline.
2.4. Surgeq
Stereotaxic coordinates were verified histologically ac-cording to Paxinos and Watson by cannulating three sub-jects from the batch of rats to be operated. Rats wereanaesthetised by inhalation of 3% halothane (May andBaker, UK) in oxygen and positioned in a stereotaxicframe (Kopf Instruments). The skull was exposed and theincisor bar adjusted for each rat such that bregma andlambda were at the same height. Three indentations weremade in the skull to accommodate screws which, togetherwith the application of dental cement, held the canmdae inplace. For bilateral cannulation of the basolateral nuclei ofamygdala, 12 mm long steel guide carmulae (23 gauge,Cooper’s Needle Works, UK) were positioned at 3.3 mmposterior to bregma, lateral +4.5 mm and vertical –5.7mm (this coordinate was taken from the dura), thus sitingthem 2 mm above the target area. Cannulae were keptpatent using 12 mm-long stainless steel stylets (30 gauge,Cooper’s Needle Works).
2.5. Behauioural testing
2.5.1. Elevated plus-maze
On the test day, rats were gently wrapped in a cloth andbilaterally injected using needles constructed from 30-gauge steel tubing which extended 2 mm below the tip ofthe indwelling carmulae. Injections were 0.5 I.L1,except for200 ng 8-OH-DPAT (0.6 I.J,l)and were made over a periodof 30 s; the needles were left in position a further 30 s toallow drug diffusion. Three min later each rat was placedin the central square of the plus-maze and its behaviourobserved for 5 rnin by an observer blind to the drugtreatment. The number of entries onto open and closedarms and the times spent in open and closed arms and inthe central square were scored. Testing took place between08.00 and 13.00 h in an order randomised for drug treat-ment in a room lit by dim light (< 50 IUX)from a lightsource directly above the plus-maze. The arena was thor-oughly wiped with damp tissue after each trial.
2.5.2. The social interaction testRats were allocated to unoperated partners on the basis
of body weight, such that members of a pair did not differby more than 5 g. They were given two 5-rein familiarisa-
tion sessions with the test arena; the two sessions wereseparated by 24 h.
On the test day, 24 h after the last familiarisation, theoperated rats were injected following the same proceduredescribed for the elevated plus-maze test. Three tin laterthe pair was placed in the social interaction arena. Thebehaviour was observed for 4.5 tin by an observer blindto the drug treatment who scored the time spent in activesocial investigation (sniffing, following and grooming thepartner) which was entered into a computer. Testing wasperformed between 08.00 and 12.00 h in an order ran-domised for drug treatment and the arena thoroughly wipedafter each trial. Testing took place over two days andvehicle control animals were tested on each day; differentsets of rats being used in each experiment.
Experiment 1. Rats were randomly allocated to thefollowing groups (the numbers in parentheses indicatethose with verified cannulae placements) and tested in theelevated plus-maze: vehicle (n = 8); 8-OH-DPAT 50 ng(n= 8), 100 ng (n= 9) or 200 ng (n= 8) and (-)-tertatolol 1.5 Kg (n= 9).
Experiment 2. Rats were randomly allocated to thefollowing groups and tested in the social interaction test:vehicle (n = 8); 8-OH-DPAT 50 ng (n = 8) and 200 ng(n= 8).
Experiment 3. Rats were randomly allocated to thefollowing groups and tested in the social interaction test:vehicle (n = 11); 8-OH-DPAT 50 ng (n = 10); (–)-tertatolol 1.5 p,g (n= 10); 8-OH-DPAT 50 ng + (–)-tertatolol 1.5 p,g (n= 10).
Experiment 4. Rats were randomly allocated to thefollowing groups and tested in the elevated plus-maze:vehicle (n = 9); midazolam 1 pg (n = 9); midazolam 2Kg (n= 10).
Experiment 5. Rats were randomly allocated to thefollowing groups and tested in the social interaction test:vehicle (n = 8); midazolam 1 pg (n = 8);midazolam 2 pg(n= 8).
2.6. Histology
At the end of behavioral testing all animals weresacrificed, the brains removed and the injection site veri-fied histologically (according to the atlas of Paxinos andWatson [23]) by a person blind to drug treatment. Frozenbrains were sectioned on a British Optical Cryocut at 10pm. Sections were obtained parallel to the stereotaxicplanes by adjusting the angle of cutting. Sections weretaken directly from the cryotome knife on coated slides(poly-L-lysine 10%) with the help of an antiroll device.
2.6.1. Acetylcholinesterase (AChE) histochemistryIn order to visualise the boundaries of the basolateral
nuclei, the method for demonstration of AChE was usedfollowing the procedures described by Paxinos and Watson[23]. Slides with fresh, unfixed tissue were incubated for

148 L.E. Gonzalez et al. /Brain Research 732 (1996) 145–153
15 h in the following solution: 100 ml of stock solution to for 10 min in 1% sodium sulphide (1.0 g in 100 ml ofwhich had been added 116 mg of SLacetylthiocholine HZO) at pH 7.5. They were then rinsed with water andiodide and 3.0 mg ethopropazine (May and Baker). Subse- immersed in 4% paraformaldehyde in phosphate buffer forquently, the slides were rinsed with water and developed 8 h and then allowed to dry. Finally they were cover-slipped
A
B
cdfalnucleus
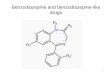
Fig. 1. A: diagrammatic representation of coronal sections from 2.8 to 3.8 mm posterior to bregma (Paxinos and Watson [23]) showing the target area(shaded)of the basolaterdamygdaloidnuclei.Theadjacentcentral~ygdaloid nucleiwe alsodrawn(unshaded).placementsf~ling outsidethe t~get meaare shownby filled circles marking the sites of the tips of the injection needles. B: coronrd section (approximately 3.3 mm Posterior to bregma) shined foracetylcholinesterase (graticule 1 mm), showing a typical bilateral carmulation of the basolateral amygdaloid nuclei (shown as dark areas at the bottom).

L.E. Gonzalez et al. /Brain Research 732 (1996) 145-153 149
with DPX (Raymond A. Lamb, London). The stock solu-tion was 50 mM sodium acetate buffer at pH 5 which wasmade by adding 6.8 g of sodium acetate, 1.0 g of coppersulfate crystals and 1.2 g of glycine to 1.0 1 of HZO andlowering the pH to 5.0 with HC1.
Fig. 1A, depicting coronal slices through the amygdalacomplex shows the target sites as shaded and the positionsof the tips of the injection needles for the animals excluded(not in the target area) from statistical analysis. A photo-graph illustrating a correct cannulation is also shown (Fig.IB) with the lower dark images corresponding to basolat-eral amygdaloid nuclei which were obtained by AChEhistochemistry.
2.7. Statistics
The social interaction scores were subjected to analysisof variance (ANOVA) with Duncan’s post-hoc tests fordifferences between individual groups. It is the signifi-cance of these that are shown in the figures. Antagonismbetween 8-OH-DPAT and tertatolol was assessed by two-way analysis of variance; antagonism being shown by asignificant 8-OH-DPAT X tertatolol interaction. To assessthe influence of the locomotor activity on anxiety mea-sures, analysis of covariance (ANCOVA) was performedwhen concomitant changes in locomotor activity and socialinvestigation were observed.
3. Results
3.1. Experiment 1. Effects of 8-OH-DPAT on behauiour inthe elevated plus-maze following intra-amygdaloid injec-tions
None of the doses tested had any significant effect onany of the measures recorded (see Table 1). However, thedrug at the dose of 200 ng produced reductions in the timespent in the central square that nearly reached significance(F(1,14) = 3.2, P < 0.07) and flat body postures in 3/8animals immediately after the injections. Since all the threedoses were without effect on measures of anxiety, we
L
o 60c10m
- 50
zo
L 40.Iih
v 50 2008-OH-OPAT (rig)
Fig. 2. Mean (+ S.E.M.) time spent in social investigation (seconds) byrats bilaterally injected into the basolateral nuclei of the amygdala withaCSF (V) or 8-OH-DPAT (50 and 200 rig). Testing was carried out underlow light, familiar test conditions for 4.5 min. “ P <0.05, compared withvehicle control.
reduced the number of drug groups to be tested in thesocial interaction test, selecting the two extremes (50 and200 rig).
3.2.Experiment 2. Effects of 8-OH-DPAT on social inter-action following intra-amygdaloid injections
Bilateral microinjections of 8-OH-DPAT (50 and 200ng) into the basolateral nuclei of amygdala significantlydecreased the time spent in social investigation (F(2,21) =5.0, P < 0.01). Post-hoc analysis showed that both doseswere significantly different from vehicle treated animals(see Fig. 2). Using the automatic system (infrared beambreaks), no significant reduction of locomotor activity wasobserved (F(2,21) = 2.5, P = 0.1) and although a signifi-cant reduction in the number of rears (F(2,21) = 4.2,P < 0.05)was observed, post-hoc tests failed to reachsignificance when comparing each group with vehicle.Since this automatic system assesses the locomotor activity
Table 1Mean (*s.E.M.) of% time on open arms, % number of entries to open -s, number of closed arm entries and time @ spent in the central sw~e by ratsin the plus-maze after intra-amygdaloid injections
Experiment Treatment % Time open arms % No. entries to open arms No. closed arm entries Time in cerrtre(s)
Exut. 1 Vehicle (aCSF) 25.5 + 4.0 49.3 * 6.8 8.9 + 1.9 67.9 + 8.9.DPAT 50 ng 33.8 + 4.9 44.5 k 2.4 10.8 ~ 0.8 73.2 + 5.8DPAT 100 ng 27.1 * 5.7 41.6 k 3.7 9.7 * 0.9 60.7 + 8.5
DPAT 200 ng 21.1 * 3.1 42.3 + 6.6 7.0 * 1.0 47.4 + 7,3
Tertatolol 1.5 pg 31.9 t 5.0 45.1 t 5.1 9.2 + 1.4 69.7 + 7.4
Expt. 4 Vehicle (satine) 36.2 * 4.8 42.5 k 4.5 8.7 * 1.1 72.8 ~ 4.8
Midazolam 1 u~ 36.0 + 4.7 43.2 h 4.7 9.9 * 1.5 66.9 + 8.3,-Midazolam 2 ~g 35.8 * 5.1 46.1 ~ 4.8 9.1 + 1.6 61.6 + 8.3
—

Tab
le2
Mea
n(~
S.E
.M.)
loco
mot
orac
tivi
tyan
dre
ars
auto
mat
ical
lyre
cord
edfo
rpa
irs
ofra
tsby
irrf
rear
edbe
ambr
eaks
orin
divi
dual
lysc
ored
for
the
inje
cted
rat
alon
e,un
der
low
ligh
tfa
mil
i~(L
F),
orhi
ghli
ght
fam
ilia
r(H
F)co
ndit
ions
ofth
eso
cial
inte
ract
ion
test
Exp
erim
ent
Tre
atm
ent
Loc
omot
orac
tivi
tybe
ambr
eaks
(pai
r)L
ocom
otor
acti
vity
(inj
ecte
dan
imal
)R
ears
beam
brea
ks(p
air)
Rea
rs(i
njec
ted
anim
al)
Exp
t.2
Veh
icle
(aC
SF)
304.
9*11
.520
6+
19.4
27.6
*5.
427
.1t
3.3
Con
diti
ons
8-O
H-D
PA
T50
ng32
0.8+
1820
2.6~
12.1
36.0
+4
35.5
+-4
.7L
F8-
OH
-DP
AT
200
ng26
6*2
2.6
128
+32
.7*
18.1
~3.
413
.4+
4.0
‘
Exp
t.3
Veb
icle
(aC
SF)
282.
7+17
.519
2.2+
13.3
35.3
*6.
526
.1+
3.3
Con
diti
ons
8-O
H-D
PA
T50
ng31
5.2*
18.5
220
*10
.444
.7+
4.8
31.6
+3.
1L
FT
erta
tolo
l1.
5pg
328.
5+
15.8
234
+16
.737
.6L
4.9
20.9
+2.
0T
er1.
5pg
+D
PA
T50
ng31
6.9
&20
.722
9.4*
11.8
44.1
*4.
032
.1*4
.6
Exp
t.5
Veh
icle
(sal
ine)
265.
3i
11.6
131.
1+9.
836
.1+
13.3
26.6
&6.
lC
ondi
tion
sM
idaz
olam
1pg
256.
5+
14.2
130.
2+8.
734
.8*
7.8
24.7
+7.
2H
FM
idaz
obm
r2p,
g27
0*7
.213
3+
6.9
35.1
+6.
123
.7&
4.3
Veh
icle
(aC
SFor
sali
ne),
8-O
H-D
PAT
,(–
)-te
rtat
olol
,8-
OH
-DP
AT
+(–
)-te
rtat
olol
orm
idaz
olam
(1an
d2
pg)
was
adm
inis
tere
dbi
late
rall
yin
toth
eba
sola
tera
lnu
cleu
sof
the
amyg
dala
via
chro
nica
lly
indw
elli
ngca
nnul
ae3
tin
prio
rto
test
ing.
Tes
ting
was
for
4.5
tin
and
scor
edby
anob
serv
erbl
ind
todr
ugtr
eatm
ent.
‘P
<0.
05,
com
pare
dw
ith
vehi
cle
cont
rol.

L.E. Gonzalez et al. /Brain Research 732 (1996) 145-153 151
. low light.: 90cou 80aJ
$’ 70zoI 60 –
z
50
40
30
20
10
0iv DPAT T T+
DPAT
high light
12vMDZ (ug)
Fig. 3. Mean (~S.E.M.) time spent in social interaction (seconds) by ratsbilaterally injected into the basolateral nuclei of amygdala either in lowlight, familiar conditions with aCSF (V), 8-OH-DPAT 50 ng (DPAT),(-)-tertatolol (T) and (–)-tertatolol (1.5 pg)+8-OH-DPAT (50 ng)(T+ DPAT) or in high light familiar conditions with saline (V) ormidazolam 1 and 2 Kg (MDZ). The scores are expressed as the cumula-tive time (seconds) during the 4.5 min of testing. ‘ F’<0.05 comparedwith vehicle control; * * P <0.01 compared with vehicle control.
and rears of a rat pair (both the injected and the unoperatedpartner), we scored only the locomotor activity and rearsof the injected animal by counting the number of squarescrossed and rears made by this rat alone. This showed asignificant reduction in locomotor activity (F(3,29) = 3.1,P < 0.05)and rears (F(3,29) = 5.3, P < 0.01), whichreached significance only for the dose of 200 ng (see Table2).
Since both social investigation and locomotor activityscores were decreased analysis of covariance was applied,to assess the independence of these effects. The effect of8-OH-DPAT (200 ng) on time spent in social investigation(F(1,14) = 6.0, P < 0.05)was no longer significant fol-lowing ANCOVA (F’(l ,14) = 2.0 P < 0.2),indicatingsome non-specific response reduction at this higher dose.
3.3.Experiment 3. Antagonism of the anxiogenic effect of8-OH-DPAT by (– )-tertatolol
The dose of 50 ng of 8-OH-DPAT was selected for thestudy of antagonism according to the results of Expt. 2.Once again, 8-OH-DPAT at a dose of 50 ng reduced thelevel of social investigation (P < 0.05). This anxiogeniceffect of 8-OH-DPAT (50 ng) was antagonised by (–)-tertatolol (1.5 p,g) (8-OH-DpAT x tertatolol interaction,F’(1,37)= 4.3, P < 0.05);see Fig. 3. There were no signif-icant changes in motor activity (see Table 2).
3.4.Experiment 4. Effects of intra-amygdala injections ofmidazolam (MDZ) on the behaviour in the elevated plus-maze
No effects were observed on any of the behaviorsrecorded in the elevated plus-maze after intra-amygdaloidinjections of midazolam (1 and 2 Lg); see Table 1.
3.5.Experiment 5. Effects of intra-amygdala injections ofmidazolam (MDZ) on the behauiour in the social interac-tion test
Bilateral administration of midazolam (1 and 2 pg) tothe basolateral nuclei of the amygdala increased time spentin social investigation (F(2,21) = 4.4, P < 0.05);see Fig.3. As shown in Table 2, this drug had no effects on eitherlocomotor activity or rears.
4.Discussion
Bilateral injections of the selective 5-HTIA receptoragonist, 8-OH-DPAT to the basolateral nucleus of theamygdala decreased levels of social investigation underlow light, familiar test conditions while having no effectson the level of open arm activity of rats on the plus-maze.The consistent anxiogenic effects of the drug in the socialinteraction test, obtained with the lowest dose used (50 ng)were blocked by ( – )-tertatolol, a 5-HTl~ receptor antago-nist [18], thereby indicating that they were caused byactivation of 5-HTIA receptors. In contrast to the anxio-genic effect of 5-HTIA receptor activation, the administra-tion of the BDZ receptor agonist midazolam (1 and 2 pg)evoked artxiolytic effects. The results obtained using 8-OH-DPAT and midazolam parallel and extend the findingsof Hodges et al. [17] who investigated the role of theamygdala in a modified Geller-Seifter paradigm. Thus,data from the social interaction and punished proceduressupport the suggestion that post-synaptic 5-HTIA receptorsand BDZ receptors may mediate, within the same area,anxiogenic and anxiolytic effects respectively. The anxi-olytic effects of midazolam are also a confirmation of theanxiolytic effects reported by Higgins et al. [16] withanother BZD receptor agonist (flurazepam, 200 ng) in thesocial interaction and a punished drinking test followingintra-amygdaloid injections.
The response of rats to intra-amygdaloid injections ofmidazolam again, like 8-OH-DPAT, highlight differencesbetween the two tests of anxiety with rnidazolam adminis-tration evoking a significant anxiolytic effect in the socialinteraction test while having no effect in the elevatedplus-maze. This would appear to suggest that the benzodi-azepine and 5-HTIA receptors of the basolateral amygdalamay not play a role in the anxiety elicited by the elevatedplus-maze. The lack of effect of midazolam in the plus-maze, however, contradicts the anxiolytic effects found by

152 L.E. Gonzalez et al. /Brain Research 732 (1996) 145-153
Green and Vale [13] and Pesold and Treit [25], but proce-dural differences may account for the discrepancies. Forexample, the former authors used 10-min instead of 5-reintrials and it has been found that trial length can change theresponse to benzodiazepines in the elevated plus-maze[12]. A second factor was that while their basal level ofcontrol (in terms of 70 time on the open arms) was about20%, ours was 36.5%. It is possible that a lower basallevel, i.e. higher anxiety level, favours the detection ofanxiolytic action, as more aversive test conditions correlatewith lower basal levels thereby facilitating the detection ofanxiolytic action, an effect clearly demonstrated in thesocial interaction test [10,1l]). A third, though possiblymore speculative, factor to be accounted for is that of adifferent method of cannulation. Green and Vale [13] usedan angle of 6° for their vertical coordinate. As can be seenin Paxinos and Watson [23], the cannula would passthrough the lateral ventricles. It is our experience that ifdiffusion occurs from the needle tip it is in an upwarddirection, which in this case would be towards the lateralventricles. It is therefore possible that the drug could reacheven remote areas such as the dorsal raph6 nucleus, asshown by Jolas et al. [19]. Regarding the study of Pesoldand Treit [25], these authors injected into the basolateralamygdala a dose of 10 ~g/O.5 pl (5-times bigger thanours) and their Yoopen arm time basal levels (about 15%)were even lower than those of Green and Vale. These twofactors may contribute to an increased chance of detectinganxiolytic activity. Even so, the anxiolytic effects reportedby Pesold and Treit are difficult to reconcile with the lackof effect in the elevated plus-maze following lesion of theamygdala found previously in that laboratory [24].
Changes in locomotor scores were detected only afteradministration of the highest dose of 8-OH-DPAT (200rig). No significant reduction in the number of closed armentries and time spent in the central square (see Table 1)was evident in the group of animals with flat body posture(n= 3). This group exhibited such a postural change im-mediately after the injection but they overcame the motorimpairment over the trial time, therefore the overall scoresof those parameters showed a normal distribution for thedrug group (200 rig). However, in the social interactiontest we detected a significant reduction of locomotor activ-ity and rears with the dose of 200 ng. Among this group,some animals also presented postural changes (flat bodyposture, n = 3) during the first minutes following thecentral injection; however these changes were not apparentin all of them at the end of the trial. As in the case of theelevated plus-maze parameters of motor activity, the loco-motor activity and rears of the 8-OH-DPAT group (200ng) had a pattern compatible with a normal distribution.
In conclusion, we have shown that intra-amygdaloidinjection of the 5-HTIA receptor agonist, 8-OH-DPAT, andthe benzodiazepine receptor agonist, midazolam, has oppo-site effects on anxiety levels in the social interaction test.At equivalent doses, in our animals, under the same surgi-
cal and environmental conditions, the drugs had no effecton behavioral performance in the elevated plus-maze.These data indicate that the tests may possess differentialsensitivity to the compounds tested here. However, moreintriguing is the proposal that there is a fundamentaldifference in the anxiety evoked by the two tests, with thatevoked by the plus-maze being mediated via brain areas orreceptors different from those studied here.
Acknowledgements
L.E.G. is supported by a grant from CONICIT-ULA,Venezuela. The authors are grateful for the expert techni-cal assistance of Mr P. Mabbutt, the use of the cryostat ofDr. M. Rattray (Div. of Biochemistry, UMDS) and thephotography by S. Smith (Div. of Anatomy and CellBiology, UMDS).
References
[1] Andrews, N., Hogg, S., Gonzalez, L.E. and File S.E., 5-HTIAreceptors in the median raphe nucleus and dorsal hippocampusmediate anxiolytic and auxiogenic behaviors respectively, Eur. 1Pharnracol., 264 (1994) 259-264.
[2] Andrews, N., Gonzalez, L.E. and File, S.E., 5-HTIA receptor media-tion of the anxiogenic effect of hippocampal injection of 8-OH-DPAT, Psychopharmacology, 9 (1995) A115.
[3] Applegate, C.D., Frysinger, R.C., Kapp, B.S. and Gallagher, M.,Multiple unit activity recorded from amygdala central during Pavlo-vian heart rate conditioning in rabbit, Brain Res., 238 (1982)457–462.
[4] Azrnitia, E.C. and Segal, M., An autoradiographic analysis of thedifferential ascending projections of the dorsal and median raphenuclei in the rat, J. Cornp. Neurol., 179 (1978) 641–668.
[5] Campeau, S., Hayward, M.D., Hope, B.T., Rosen, J.B., Nestler, E.J.and Davis, M., Induction of c-~os proto-oncogene in rat amygdaladuring unconditioned and conditioned fear, Brain Res., 565 (1991)349-352.
[6] Chalmers, D.T. and Watson, S.J., Comparative anatomical distribu-tion of 5-HTIA receptor and mRNA and 5-HTL~binding in rat brain—a combined in situ hybridisation on vitro receptor autoradiographicstudy, Brain Res., 561 (1991) 51–60.
[7] Chapman, W.P., Schroeder, H.R., Guyer, G., Brazier, M.A.B., Fager,C., Poppen, J.L., Solomon, H.C. and Yakolev, P.I., Physiologicalevidence concerning the importance of the amygdaloid nuclea regionin the integration of circulating function and emotion in man,Science, 129 (1954) 949-950.
[8] Davis, M., The role of the amygdala in conditioned fear. In J.P.Aggleton (Ed.), The Amygdala: Neurobiological Aspects of Emotion,Memory and Mental Dysfunction, Wiley-Liss, New York, 1992, pp.255-305.
[9] Femandes, C., Andrews, N. and File, S.E., Diazepam withdrawalincreases [3H15-HT release from rat amygdaloid slices, Pharmacol.Biochem. Behaw., 49 (1994) 359-362.
[10] File, S.E., The use of social interaction as a method for detectinganxiolytic activity of chlordiazepoxide-like drugs, 1 Neurosci.Methods, 2 (1980) 219-238.
[11] File, S.E., Behavioral detection of anxiolytic action. In J.M. Elliott,D.J. Heal and C.A. Marsden (Eds.), Experimental Approaches toAnxiety and Depression, John Wiley and Sons, Chichester, UK,1992, pp. 25-44.

LE. Gonzalez et al. /Brain Research 732 (1996) 145-153 153
[12] File, S.E., The interplay of learning and anxiety in the elevatedplus-maze, Behau. Brairs.Res., 58 (1993) 199–202.
[13] Green, S. and Vale, A.L., Role of urnygdaloid nuclei in the anxi-olytic effects of benzodiazepines in rats, Behau. Pharnracol., (1992)261-264.
[14] Griebel, G., 5-Hydroxytryptamine-interacting drugs in animal mod-els of anxiety disorders: more than 30 years research, Pharmacol,Ther., 65 (1995) 319-395.
[15] Henke, P.G., Unit-activity in the central amygdaloid nucleus of ratsin response to immobilization-stress, Brain. Res. Rev., 10 (1983)833-837.
[16] Higgins, G., Jones, B., Oakley, N, and Tyers, M., Evidence that the~ygdala is involved in the disinhibitory effects of 5-HT3 receptorantagonists, Psychopharmacology, 104 (1991) 545–551.
[17] Hodges, H., Green, S. and Glenn, B., Evidence that the amygdala isinvolved in benzodiazepine and serotonergic effects on punishedresponding but not on discrimination, Psychopharmacology, 92(1987) 491-504.
[18] Jolas, T., Haj-Dahmane, S., Lanfumey, L., Fattaccini, C.M., Kidd,E.J., Adrien, J., Gozlan, B., Guardiola-Lemaitre, B. and Harnon, M.,(–)-Tertatolol is a potent antagonist at pre- and postsynaptic sero-tonin 5-HTiA receptors in the rat brain, Arch. Pharmacol., 347(1993) 453-463.
[19] Jolas, T., Schreiber, R., Laporte, A.M., Chastanet, M., De Vry, J.,Glaser, T., Adrien, J. and Hamon, M., Are postsynaptic 5-HT1~receptors involved in the anxiolytic effects of 5-HTIA receptoragonists and in their inhibitory effects on the firing of serotonergicneurons in the rat?, J. Pharmacol. Exp. Ther., 272 (1995) 920-929.
[20] Kawahara, H.,Yoshida, M., Yokoo, H., Nishi, M. and Tanaka, M.,Psychological stress increases serotonin release in the arnygdala andprefrontal cortex assess by in vivo nricrodialysis, Nezsrosci. Left.,162 (1993) 81-84.
[21] Ma, Q.T., Yin, G.F., Ai, M.K. and Han, J.S., Serotonergic projec-tions from the nucleus raphe dorsalis to the amygdala in the rat,Nearosci. Lett., 134 (1991) 21-24.
[22] Melia, K.R., Sananesm, C.B. and Davis, M., Lesions of the central
nucleus of the amygdala block the excitatory effects of septalablation on the acoustic startle reflex, Physiol. Behau., 51 (1992)175-180.
[23] Paxinos, G. and Watson, C.H., The Rat Brain in Stereotaxic Coordi-nates, Academic Press, London, 1986.
[24] Pesold, C, and Treit, D., The septum and the amygdala differentiallymediate the anxiolytic effects of benzodiazepines, Brain Res., 638(1994) 295-301.
[25] Pesold, C. and Treit, D,, The central and basolateral amygdaladifferentially mediate the anxiolytic effects of benzodiazepines,Brain Res., 671 (1995) 213-221.
[26] Rosen, J.B. and Davis, M,, Enhancement of acoustic startle byelectrical stimulation of the amygdala, Behau. Neurosci., 102 (1988)195-202.
[27] Sananes, C.B. and Davis, M., NMDA lesions of the lateral andbasolateral nuclei of the amygdala block fear-potentiated startle andshock sensitization of startle, Behau. Neurosci., 106 (1992) 72-80.
[28] Scheel-Kriiger, J. and Petersen, E.N., Anticonflict effect of thebenzodiazepines mediated by GABAergic mechanism in the amyg-dala, Eur. 1 PharmacoZ., 83 (1982) 115-116.
[29] Shibata, K., Kataoka, Y., Gomita, Y. and Ueki, S., Localisation ofthe site of the anticonflict action of benzodiazepines in the amyg-daloid nucleus of rats, Brain Res., 234 (1982) 442-446.
[30] Takao, K., Nagatani, T., Kasahrrra, K. and Hashimoto, S., Role ofthe central serotonergic system in the anticonflict effect of d-AP159,Pharmacol. Biochem. Behau,, 43 (1992) 503-508.
[31] Thomas, S.R. and Iversen, S.D., Correlation of 3H-diazepam bindingdensity with anxiolytic locus in the amygdafoid complex of the rat,Brain Res., 342 (1985) 85-90.
[32] Umemoto, M. and Olds, M.E., Effects of chlordiazepoxide, di-azeparn and chlorpromazine on conditioned emotional behaviour andconditioned neuronal activity in Iimbic hypothalamic and genicuIateregions, Neuropsychopharmacology, 14 (1975) 413–425.
[33] Wright, D.E., Seroogy, K.B., Lundgren, K.H., Davis, B.M. andJennes, L., Comparative localization of serotonin ,a,lCand* receptorsubtype mRNAs in rat brain, J Comp. Neurol., 351 (1995) 357–373.