Embed Size (px)
Citation preview

Transport od endoplazmatskog retikulado Golgijevog aparata i kroz njega


Stanica tankog crijeva koja luči mukus – sluz
Golgi proizvodi glikoproteine i proteoglikane
polarizirana stanica: lučenje apikalno
Krstić 1984
Golgijev aparat je naročito izražen kod sekretornih stanica

FoLu (gray fox lung) cells transduced with Organelle Lights Golgi-GFP fluorescent protein (O36215). Imaging was performed on fixed cells using fluorescence and DIC microscopy. Image courtesy of Michael Davidson,
Molecular Expressions Inc., Tallahassee, FL.

Rothman i sur.
Rekonstitucija vezikularnogtransporta
-virusom inficirana mutantna stanicane može dodavati N acetilglukozaminu Golgiju-virusni proteini se mogu prepoznati
u epruveti pomiješani GA normalne neinficirane stanice i GA mutirane inficiranestanice
-glikozilacija virusnih proteina:dokaz prijenosa enzima koji dodaju Nacetil-glukozamin glikoproteinima vezikulamaizmeđu mutiranog i normalnog Golgija

Struktura Golgija: cisterne (6-60)-u blizini jezgre i centrosoma, lokalizirani mikrotubulima
cis strana (ER-ulaz)cis Golgi mreža CGNcis cisternamedijalna cisternatrans cisternatrans Golgi mreža TGNtrans strana (sekrecija – izlaz)
Go
lgijev stog

Funkcije Golgijevog aparata
sinteza složenih glikoproteinaO glikozilacija proteinasfingomijelin i glikolipidiglikozaminoglikani ekstracelularnog matriksasloženi polisaharidi biljke (hemiceluloza, pektin)
razvrstavanje proteina i vezikularni transport-oligosaharidne grupe i proteinski uzorci usmjeravajusortiranje

Proteini na putu od ER do Golgija i dalje do površine stanice prolaze niz modifikacija.
Mora postojati ravnoteža puteva transporta prema naprijed inazad (vraćanje rezidentnih proteina u prethodni odjeljak)
sortiranje, selekcija proteina za pakiranje i transport

Glikozilacija u Golgijevom aparatu
ER: N-vezani oligosaharidi reguliraju pravilno savijanje
GA: dodatne modifikacije:
kompleksni oligosaharidioriginalni N oligosaharidi iz ER se skraćuju i dodaju dodatnišećeri (N-acetilglukozamini, galaktoza, sijalinska kiselina, fukoza)
oligosaharidi bogati manozom:nalikuju onima iz ER (AGA, manoza)
tip glikoproteina ovisi o dostupnosti enzima GAmodifikacije ovise o položaju oligosaharida: ako je dostupan enzimimanastat će složeni glikoprotein, ako je uz proteinsku površinu nastat će visokomanozni
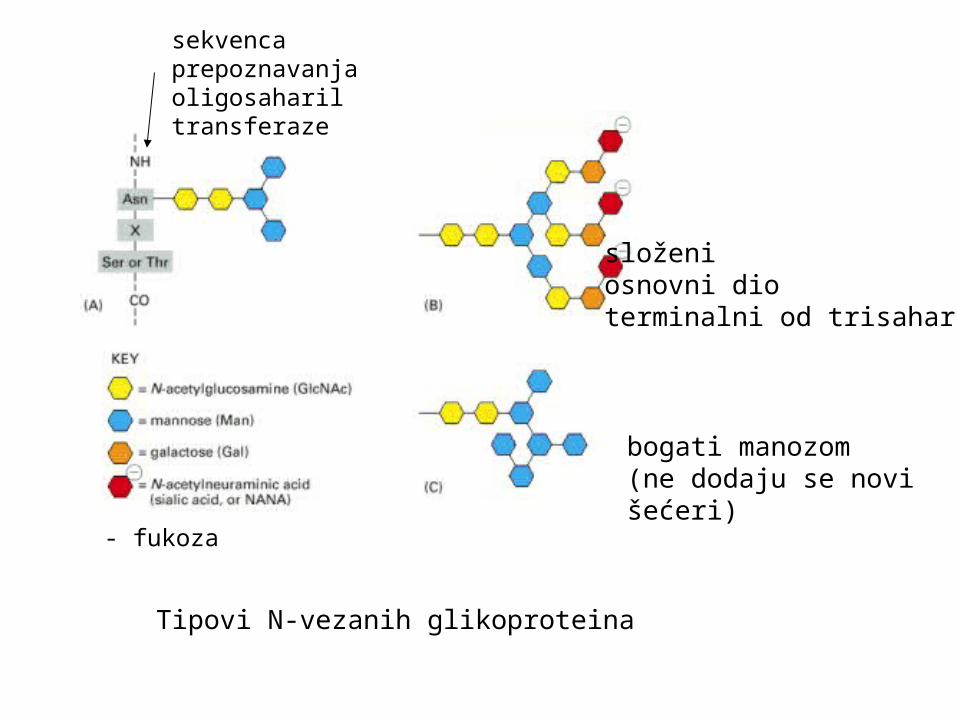
Tipovi N-vezanih glikoproteina
složeniosnovni dioterminalni od trisaharida
bogati manozom(ne dodaju se novišećeri)
- fukoza
sekvenca prepoznavanja oligosaharil transferaze
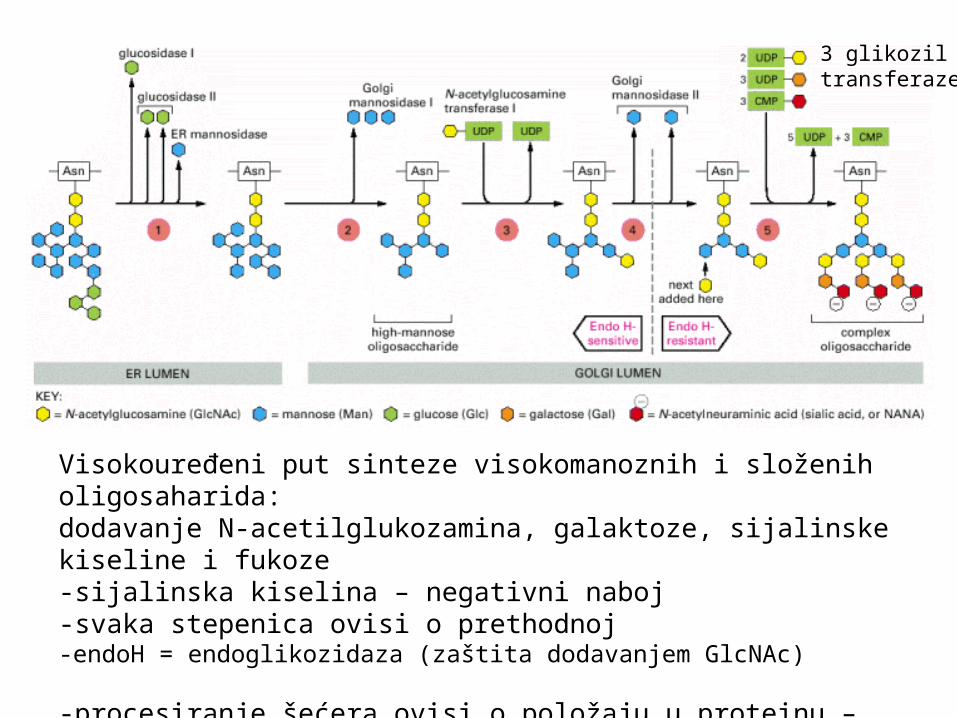
Visokouređeni put sinteze visokomanoznih i složenih oligosaharida:dodavanje N-acetilglukozamina, galaktoze, sijalinske kiseline i fukoze-sijalinska kiselina – negativni naboj-svaka stepenica ovisi o prethodnoj-endoH = endoglikozidaza (zaštita dodavanjem GlcNAc)
-procesiranje šećera ovisi o položaju u proteinu – dostupnosti enzimima -ako je uz sam protein i nedostupan – ostaje visokomanozni
3 glikozil transferaze

O-vezana glikozilacija na Ser ili Thr (OH)glikoziltransferaze dodaju šećerne nukleotideobično N-acetilgalaktozamin + do desetak šećera
nastanak proteoglikana: modifikacija proteoglikanskih proteina u proteoglikanepolimerizacija glikozaminoglikanskih lanaca – povezani preko ksiloze na Sermnogi se sekretiraju kao dio ekstracelularnog matriksa,drugi ostaju vezani za membranumukus kao zaštita epitela
sulfatacija glikozaminoglikana – negativni nabojaminokiseline tirozina
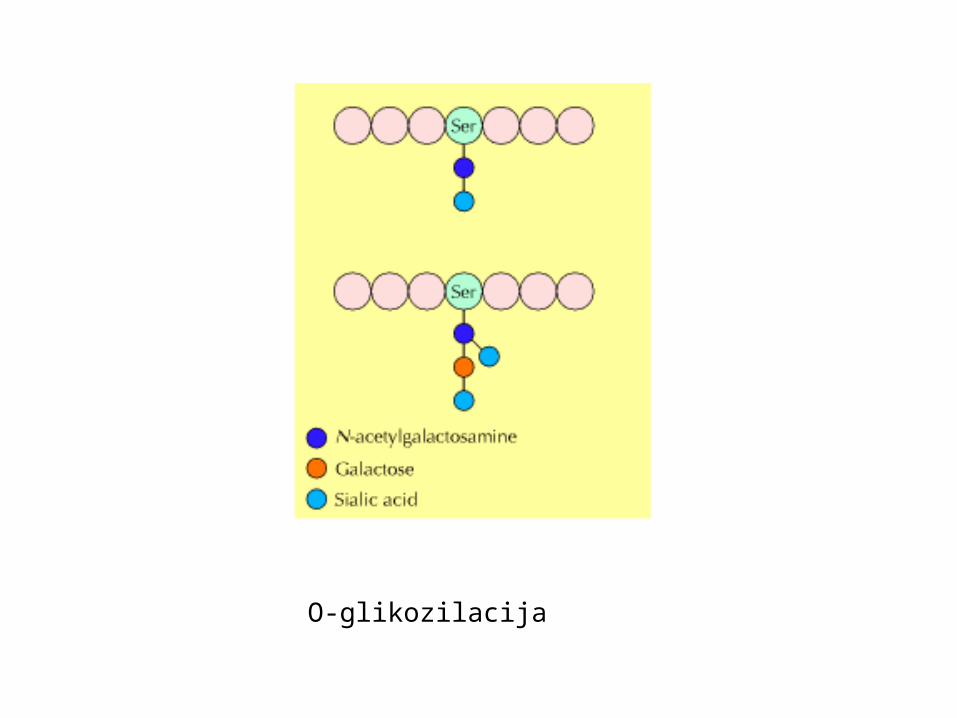
O-glikozilacija

Uloga glikozilacije proteina
za razliku od sinteze DNA, RNA, proteinaglikozilacija zahtijeva za svaku reakciju drugi enzim –složen proces koji upućuje na važnost glikozilacije
N-vezana glikozilacija: eukarioti (+ kvasac)
“signalizacija”proteinski folding u ERlektini (proteini koji vežu ugljikohidrate) usmjeravaju ER-Gatransport i sortiranje u trans Golgi mreži
zaštita od proteaza
selektini – vrsta lektina koja sudjeluje u međustaničnoj adheziji
prijenos signala – Notch receptor ovisi o O-glikozilaciji

Uloge glikolipida
ovisi o lokalizaciji:
-apikalna površina epitelne stanice:
zaštita od nepogodnih uvjeta: niskog pH i enzima
-nabijeni glikolipidi: prijenos iona i signala
-prepoznavanje: lektini
adhezija stanica
-ulaz za bakterijske toksine: gangliozid Gm1 za toksin kolere – proljev, porast cAMP, ulaz Na i vode u stanicu

Gangliozidi: oligosaharidi s jednom ili više sijaličnih ostataka - negativan naboj40 gangliozida, najobilniji u živčanim stanicama-prijenos živčanog impulsa
Galaktocerebrozid-neutralan
-složeni glikolipidi nastaju u Golgijevom aparatuiz ceramida
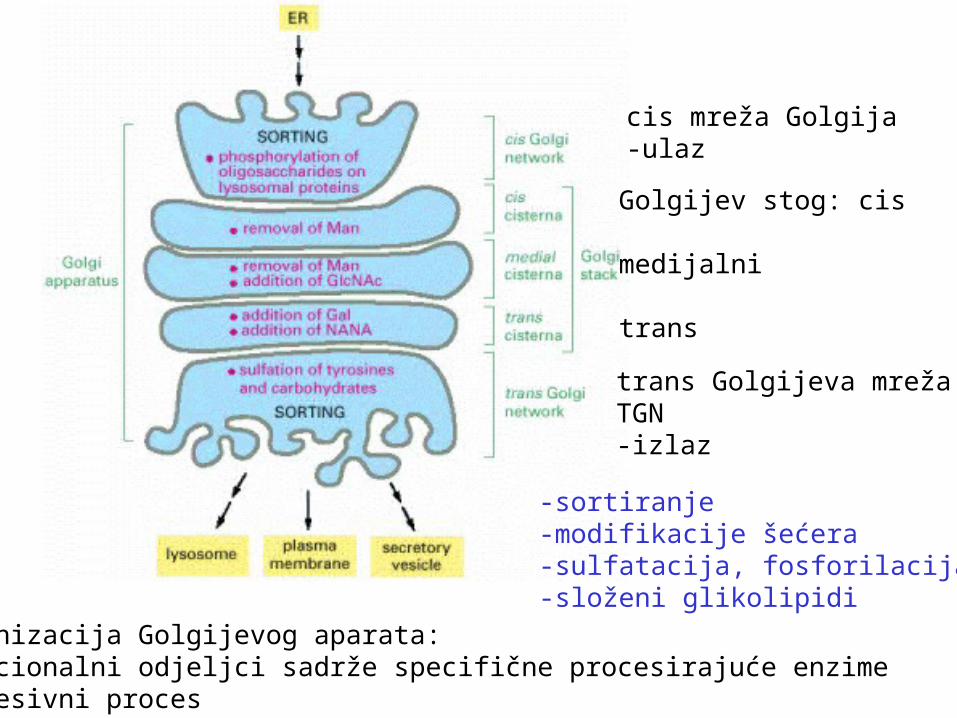
Organizacija Golgijevog aparata: funkcionalni odjeljci sadrže specifične procesirajuće enzimesukcesivni proces
cis mreža Golgija-ulaz
Golgijev stog: cis
medijalni
trans
trans Golgijeva mrežaTGN-izlaz
-sortiranje-modifikacije šećera-sulfatacija, fosforilacija-složeni glikolipidi

Vezikularni transport između odjeljaka-definicija karaktera odjeljka: kompozicija membrane – specifični markeripo principu donora i receptora, ako su na više odjeljaka, specifična kombinacija daje jedinstvenu molekularnu adresu-Lumen vezikula i organela topološki ekvivalentan-stalna komunikacija odjeljaka-donori i ciljni odjeljci izmjenjuju transportne vezikulevezikule “pupaju” s jedne membrane i stapaju se s drugomnoseći “teret” -principi signala i receptora

Biosintetski sekretorni put:
proteini sintetizirani na ER i usmjereni prema Golgijupakiraju se u male COPII obložene transportne vezikule
ERGIC: ER – Golgi intermediate compartment: struktura membranaizmeđu endoplazmatske mrežice i Golgija
tv. pupaju na mjestima ER “exit” sites: mjestima izlaza ERproteini se koncentriraju: dio ima transportne “izlazne” signale kojeprepoznaju receptori i pakiraju ih u pupajuće vezikule
rezidentni proteini polako “cure” iz ER – retrogradni transportiako nemaju signala, u maloj se količini “spakiraju” u vezikule
ERGIC53 – lektin koji usmjerava iz ER u GA

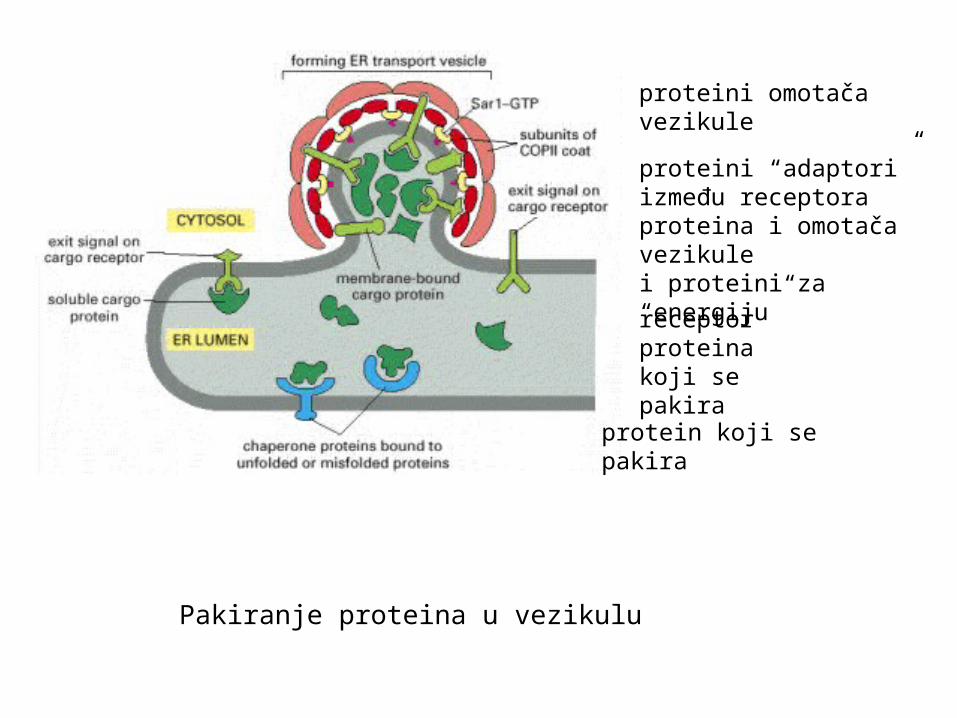
Pakiranje proteina u vezikulu
protein koji se pakira
receptor proteina koji se pakira
proteini “adaptori”između receptora proteina i omotača vezikule
proteini omotača vezikule

-Vezikule čine cjevaste nakupine – tubularne klastere-Mikrotubuli pomažu u transportu klastera i vezikula prema trans GA-Iz vezikularnih tubularnih klastera se odvajaju vezikuleCOPII za transport naprijed iz ERCOPI za povratni transport rezidentnih proteina ERTransport iz ER u cis Golgijevu mrežu
mikrotubuli: transport vezikula i klastera
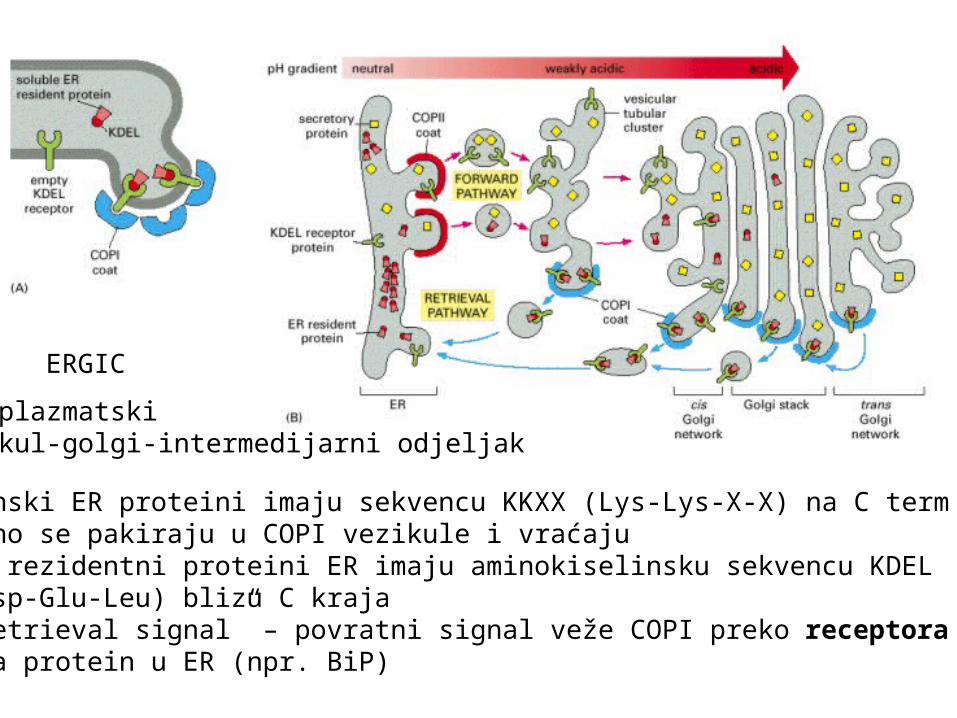
Membranski ER proteini imaju sekvencu KKXX (Lys-Lys-X-X) na C term i direktno se pakiraju u COPI vezikule i vraćajuTopivi rezidentni proteini ER imaju aminokiselinsku sekvencu KDEL (Lys-Asp-Glu-Leu) blizu C kraja-ER “retrieval signal” – povratni signal veže COPI preko receptora KDELi vraća protein u ER (npr. BiP)
ERGIC
endoplazmatski retikul-golgi-intermedijarni odjeljak

Transmembranski proteini ER: direktno vežu COPI Topivi rezidentni proteini ER: vežu receptor KDEL sekvencea on veže COPI, formira se vezikula i vraća nazad u ERsam receptor KDEL se mora vratiti u GA
afinitet vezanja za protein reguliran je promjenom pH:u blago kiselom GA receptor veže KDEL sekvencuu neutralnom ER otpuštakiselost se regulira H+ pumpama na membrani GA
neki proteini nemaju “signale” i slučajno i sporo odlaze i vraćaju se u GA iz ER
-mutiran KDEL rezidentnih proteina ne dovodi do masovne sekrecijepretpostavlja se da su proteini ipak zadržani u ER stvaranjem agregata kojisu preveliki za transport (kin recognition u GA)

Duljina transmembranskog dijela proteina također određujesmjer transporta:kolesterolne membrane su “deblje”, a čine vanjsku membranuproteini usmjereni prema njoj imaju dulje transmembranskedijeloverezidentni proteini ER koji se ne ugrađuju u vanjske membraneimaju kraći transmembranski dio

Proteini ulaze u cis Golgi i i pomiču se naprijed u GA ili se vraćaju u ER ako su rezidentni proteini
proteini koji izlaze iz trans Golgija sortiraju se ovisnokamo su usmjereni: u lizosome, sekretorne vezikule,staničnu površinu, ili se vraćaju u prethodni odjeljak

Organizacija Golgijevog aparata: funkcionalni odjeljci sadrže specifične procesirajuće enzimesukcesivni proces
cis mreža Golgija-ulaz
Golgijev stog: cis
medijalni
trans
trans Golgijeva mrežaTGN-izlaz
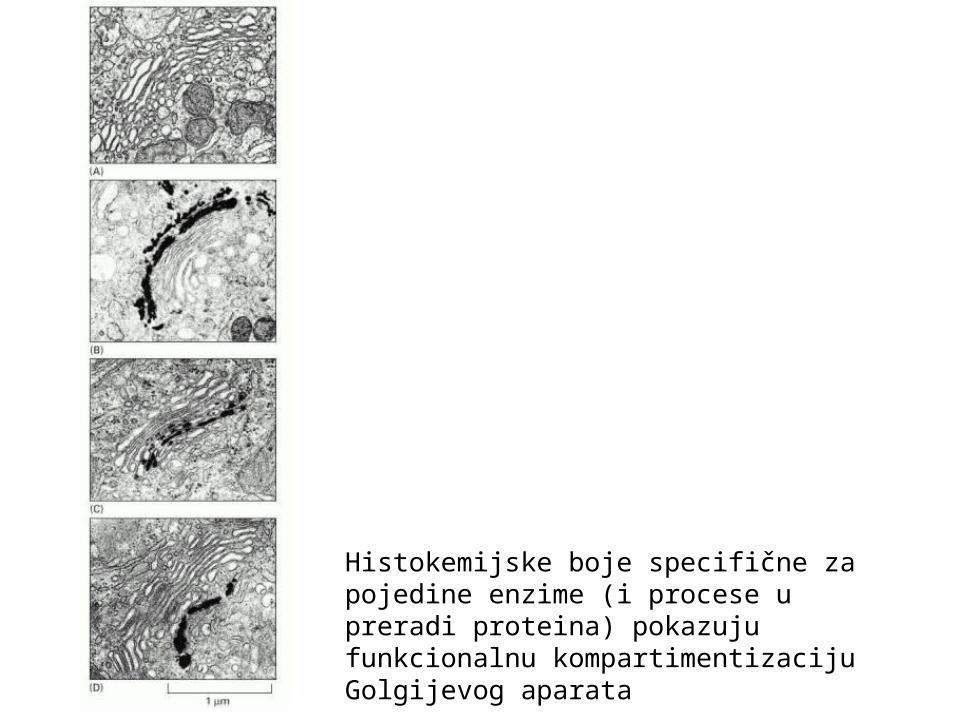
Histokemijske boje specifične za pojedine enzime (i procese u preradi proteina) pokazuju funkcionalnu kompartimentizaciju Golgijevog aparata
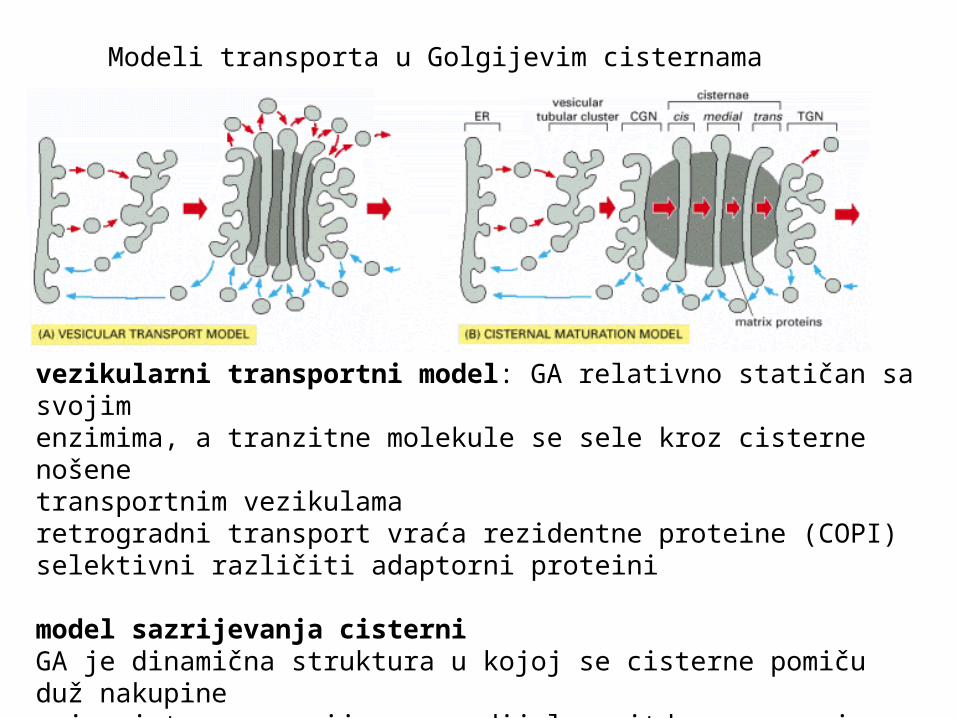
Modeli transporta u Golgijevim cisternama
vezikularni transportni model: GA relativno statičan sa svojimenzimima, a tranzitne molekule se sele kroz cisterne nošene transportnim vezikulamaretrogradni transport vraća rezidentne proteine (COPI)selektivni različiti adaptorni proteini
model sazrijevanja cisterni GA je dinamična struktura u kojoj se cisterne pomiču duž nakupine-cis cisterna sazrijeva u medijalnu, itd, a nove cis kontinuirano nastajuretrogradna struja enzima-moguća kombinacija modela

Lumenal targeted GFP, used as a marker of soluble cargo, visualises rapid ERGIC to Golgi traffic by a tubulo-vesicular network (Blum R, Stephens D J, Schulz I: J.Cell Sci,113, 3151-3159, 2000.)
-mikrotubuli: lokalizacija Golgija, transport vezikula-matriks Golgija: regulacija fosforilacijom u mitozi – raspad GA

Vezikularni transport

Dvosmjerni promet u stanici: endocitički (uzimanje tvari)i biosintetički-sekretorni koji završava egzocitozom
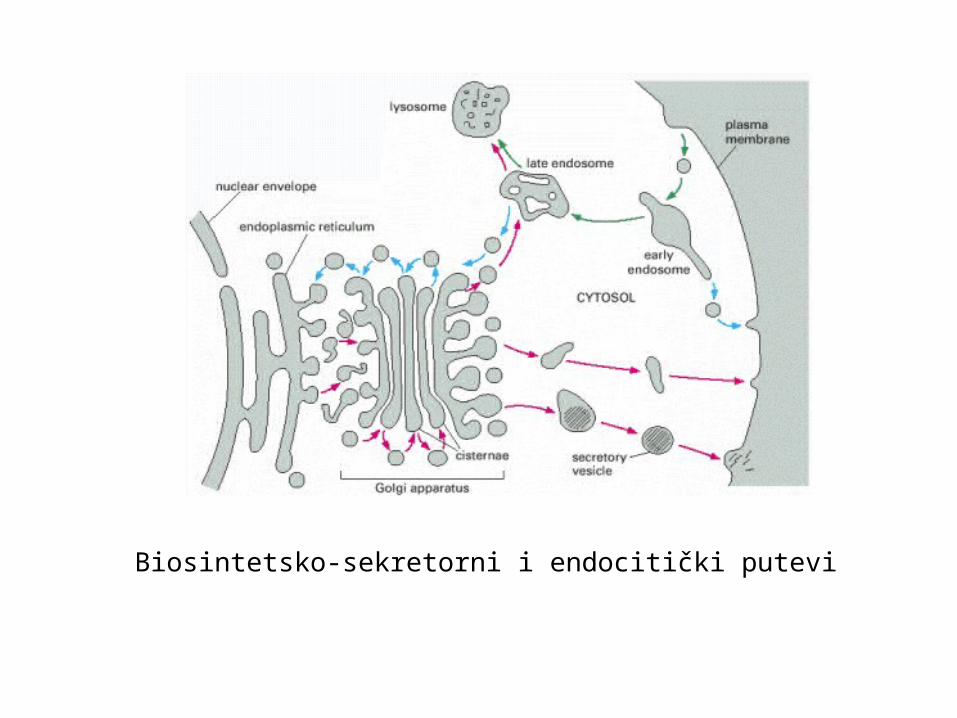
Biosintetsko-sekretorni i endocitički putevi
Pakiranje proteina u vezikulu
protein koji se pakira
receptor proteina koji se pakira
proteini “adaptori”između receptora proteina i omotača vezikulei proteini za “energiju”
proteini omotača vezikule

Transportne vezikule nastaju iz posebnih dijelova membrane koji sadrže specifične proteine koji sudjeluju u transportu – “omotače”
omogućuju koncentriranje određenih molekula koje se prenosestvaraju zaobljenu strukturu nalik košarici s citosolne strane i oblikuju vezikuluprije fuzije s ciljnom membranom uklanja se “omotač”
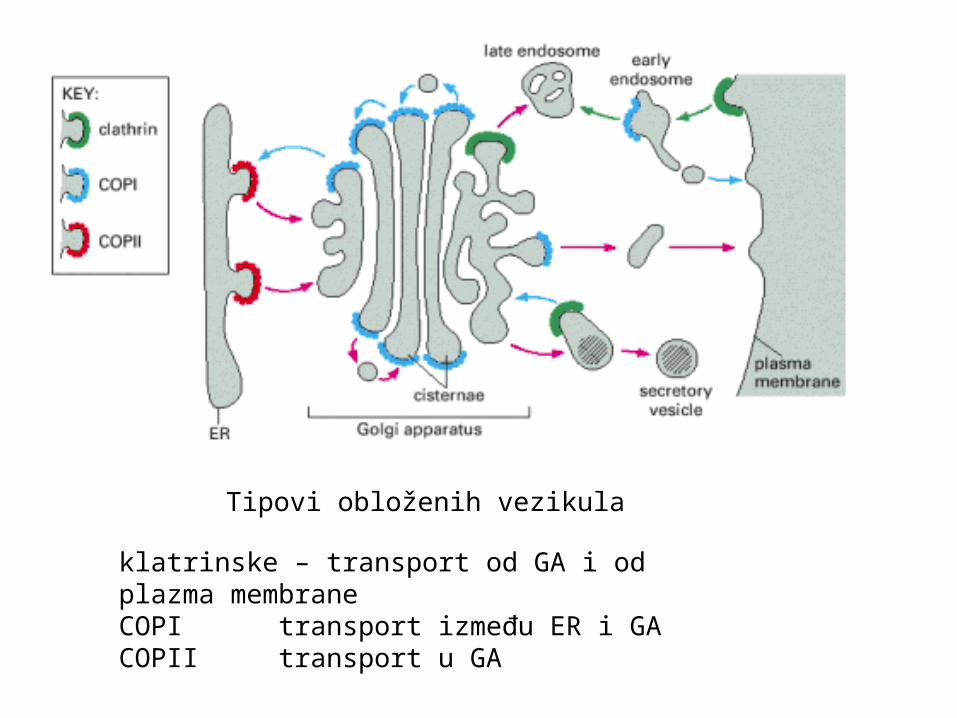
Tipovi obloženih vezikula
klatrinske – transport od GA i od plazma membraneCOPI transport između ER i GACOPII transport u GA

Tri tipa vezikula

Triskelion klatrina (podjedinica vezikule) – čine strukturu nalik košarici
-tri teška i tri laka lanca-struktura penta i heksamera

Klatrinske vezikule i jamice

Sastav klatrinske vezikule
klatrinski triskelion - spontano udruživanje heksagona i pentagonau mrežicu i vezikulu
adaptin: veže klatrin za membranu veže transmembranske proteine i transmembranske receptore (cargo receptors) koji reagiraju sa specifičnim proteinima koji se pakiraju i prenose
dinamin: kako raste klatrinski “pup”, dinamin čini prsten oko vratapupaGTPaza omogućuje otkidanje vezikule od membrane
-“odmatanje” vezikule od klatrina u citoplazmi

Nastanak i odmatanje klatrinske vezikule-rigidnost membrane – vezikule se odcjepljuju na zaobljenimdijelovima GA
auksilin – aktivacija ATP-aze koja omogući odmatanje

Dinamin odcijepa vezikulu
vezikule pričvršćene za membranukad je dinamin mutiran

COPII vezikula se formira pomoću G proteina Sar-1-regrutacija proteina omotača
ARF : COPI i klatrinski omotač na Golgijevoj membraniSar1: COPII omotač na ER membraninepoznat: klatrinski omotač na plazma membrani

Specifičnost transporta vezikula: svaka vezikula ima svoj cilj“kodiran” površinskim markerskim molekulama ovisnim o porijeklu itipu tereta, a ciljne membrane imaju komplementarne receptore iprepoznaju markere
Prepoznavanje je kontrolirano putem
SNARE – kataliziranje fuzije vezikula
Rab –GTPaza – početno pristajanje i privlačenje vezikule na ciljnu membranu

SNARE komplementarni parovi (v i t SNARE: vezikularni i ciljni - target)omogućavaju fuziju približavanjem dviju membranaspecifičnost interakcijealfa heliksi se omotavaju jedan oko drugog- mete toksina tetanusa i botulizma- homotipska i heterotipska fuzija: istovrsne i različite membrane/odjeljci

Proteini SNARE upravljaju vezikularnim transportom
-dodatne kontrolne molekule za trenutnu fuziju
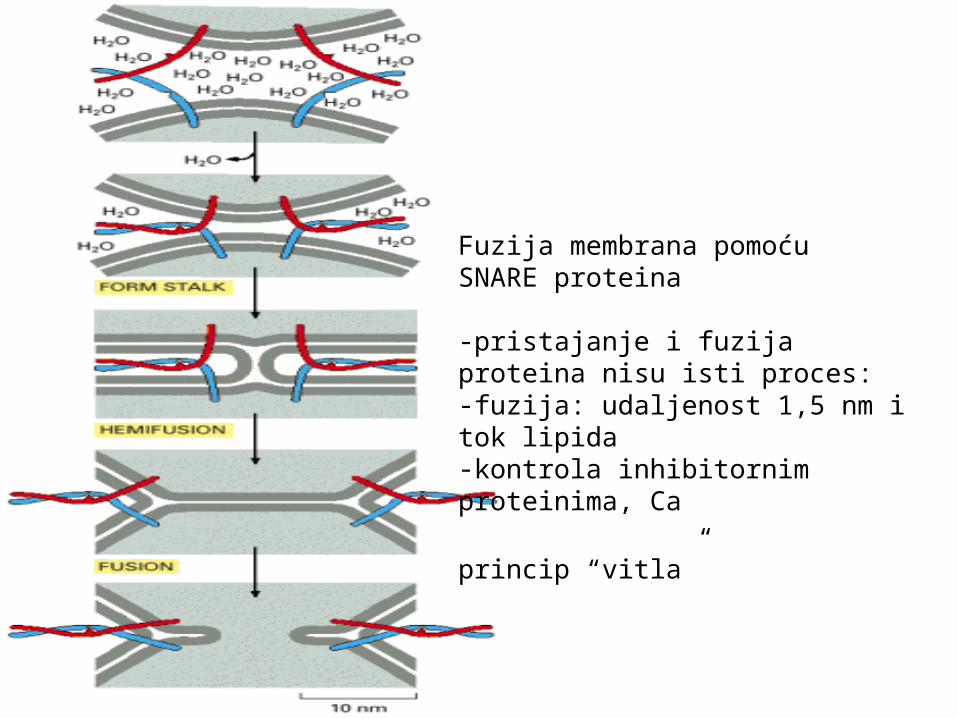
Fuzija membrana pomoćuSNARE proteina
-pristajanje i fuzija proteina nisu isti proces:-fuzija: udaljenost 1,5 nm i tok lipida-kontrola inhibitornim proteinima, Ca
princip “vitla”

Protein NSF koristi ATP za “reaktiviranje” SNARE koji su fuzionirani, da bise odvojili. Tako se dodatno kontrolira kada i gdje će se vezikule spojiti.

Rab proteini: GTPazevežu GTP i mijenjaju konformaciju, prikvače se za ciljnu membranuvezani su za transportnu vezikulu kad se odvoji od donorske membrane i vežu odgovarajuće Rab efektorske proteine na ciljnoj membraniRab i njegov receptor olakšavaju sparivanje v i t SNARE
dodatni proteini drže ga u inaktivnom stanju
specifični su Rab na pojedinim membranama (“adrese”)
koncentriraju i privlače vezikule na njihova ciljna mjesta i okidaju oslobađanje kontrolnih SNARE proteinanakon pristajanja hidroliziraju GTP i vraćaju se inaktivni u citosol
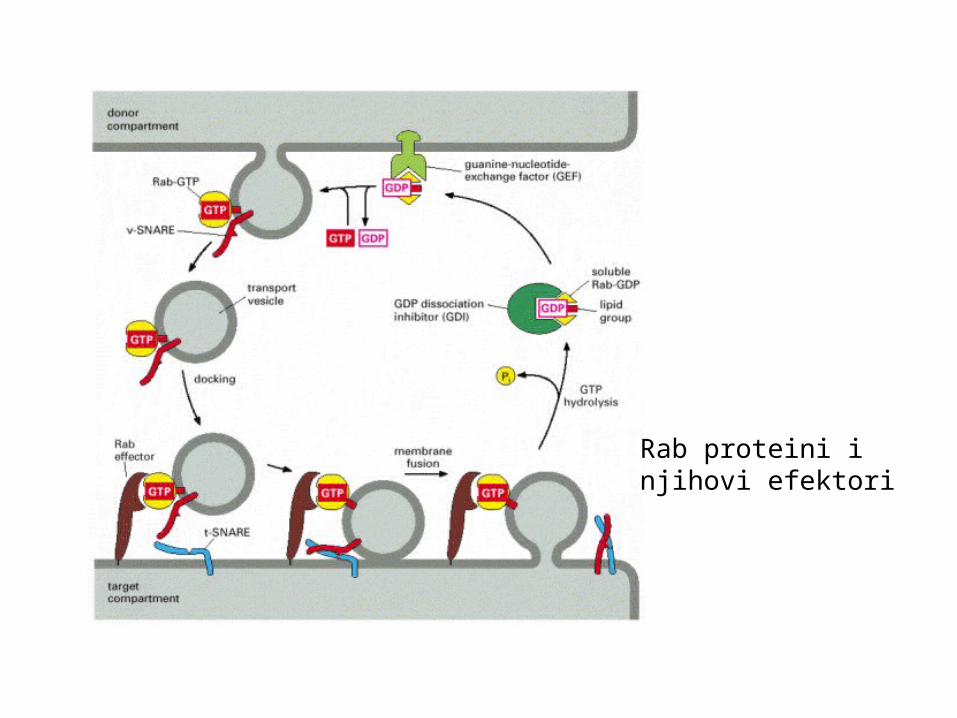
Rab proteini injihovi efektori

Rab proteini koji vežu GTP i mjesta njihova djelovanja
Korak u prijenosu Uključeni Rab proteini
egzocitozaod prijelaznog ER do GA Rab1, Rab1b, Rab2od GA natrag u ER Rab6, Rab6bunutar GA Rab1, Rab6, Rab6bod trans G mreže do stanične membrane Rab11a, Rab11b
endocitozaod stanične m. do ranih endosoma Rab5a, Rab5b, Rab5cod ranih endosoma do st. membrane Rab4, Rab15, Rab18od ranih do kasnih endosoma Rab7
posebne ulogeegzocitoza sekretornih zrnaca Rab8bod kasnih endosoma do trans G mreže Rab9, Rab11a, Rab11bod trans G mreže do bazolateralne membrane Rab8aod trans G mreže do apikalne membrane Rab21
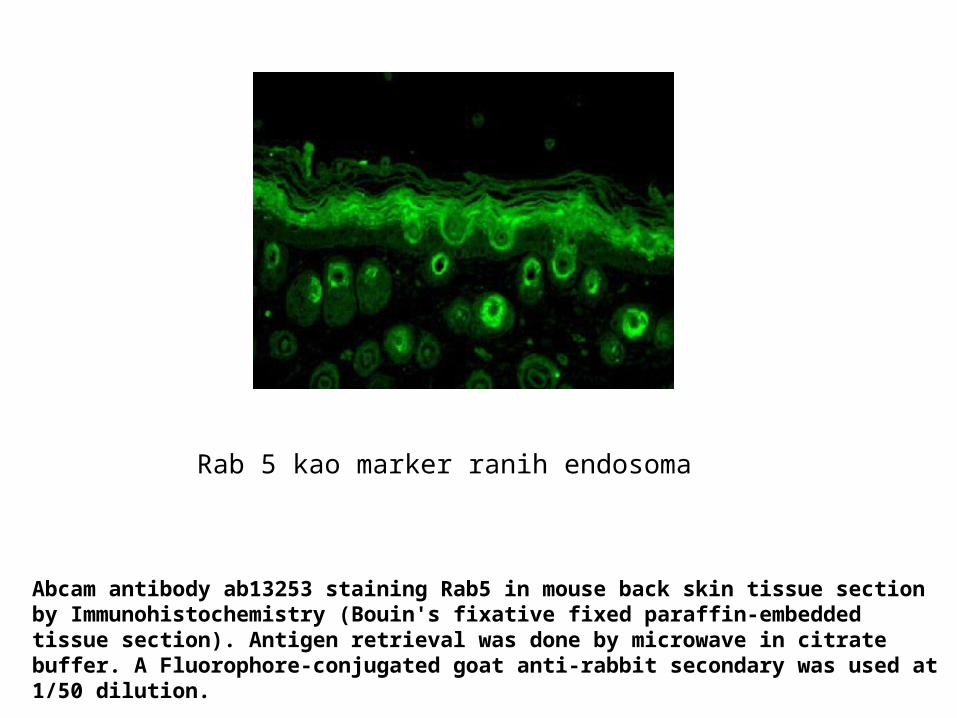
Rab 5 kao marker ranih endosoma
Abcam antibody ab13253 staining Rab5 in mouse back skin tissue section by Immunohistochemistry (Bouin's fixative fixed paraffin-embedded tissue section). Antigen retrieval was done by microwave in citrate buffer. A Fluorophore-conjugated goat anti-rabbit secondary was used at 1/50 dilution.

Fuzija stanica:
oplodnja
mioblasti
infekcija virusa

Ulaz virusa HIV-a u stanicu fuzijom

Transport od trans Golgija u lizosome
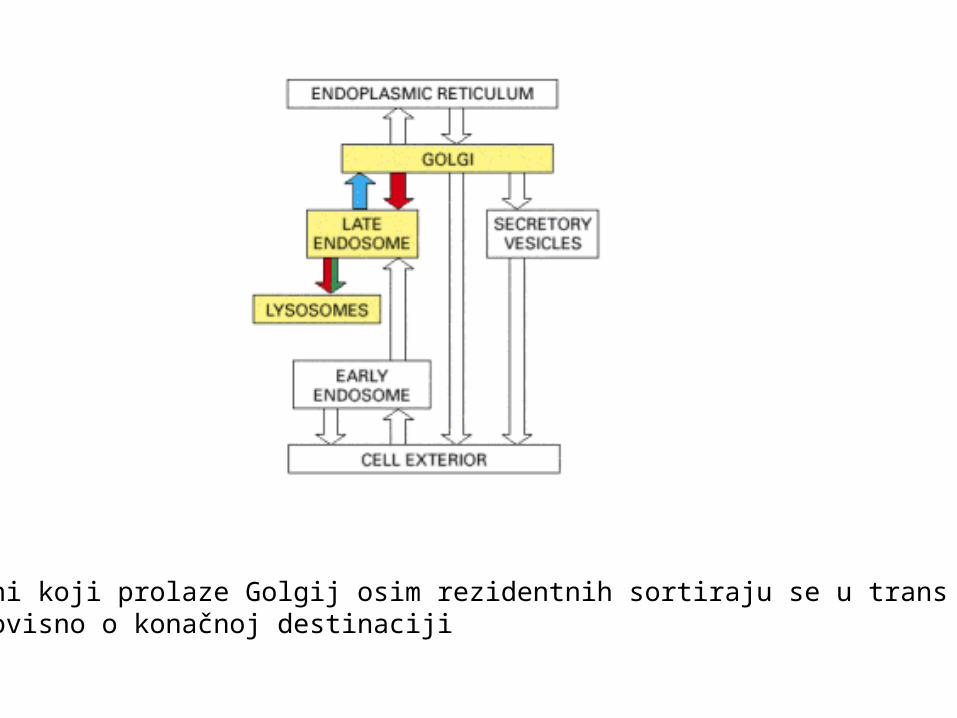
Proteini koji prolaze Golgij osim rezidentnih sortiraju se u trans Golgijevojmreži ovisno o konačnoj destinaciji

Vakuole
specifične za biljke i gljive
jedna ili više velikih tekućinom ispunjenih mjehurića
srodni lizosomima po hidrolazama, ali druge funkcije (različite vakuole)
biljne stanice:
pohrana hranjivih tvari i otpadnih tvari
odjeljak za degradaciju
metoda povećanja veličine stanice
kontrola turgorskog tlaka (osmotski pritisak)
-kontrolirano cijepanje i sinteza polimera održava turgorski tlak
homeostatsko sredstvo kod varijacije uvjeta okoline
transport H+ u vakuolu održava kiselost citosola
specifični transporteri

Specifični vakuolni sastojci kod biljaka
vakuolna bojila: antocijani i antoksantini
glikozidi (cijanovodik, digitalis)
alkaloidi (kofein, kodein, kinin, strihnin, kokain, nikotin, kolhicin)
treslovine (tanini) - zaštita od mikroorganizama
ulja i masti – pohrana rezervne hrane, alge, sjemenke
terpenski derivati i eterična ulja (smole, balzami, terpentin,
kaučuk)

Vakuole biljaka-30-90% volumena stanice

http://www.biologyjunction.com/images/3rhoeo.jpg
Obojene vakuole biljke Rheo discolor
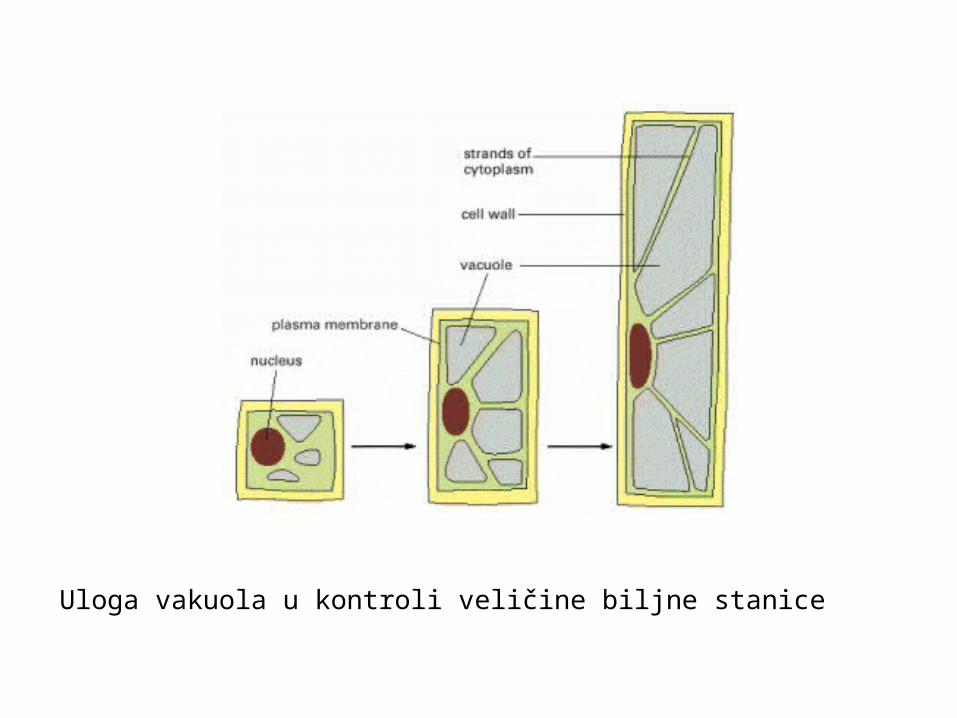
Uloga vakuola u kontroli veličine biljne stanice

Lizosomi
membranom odjeljene vezikule ispunjene hidrolitičkim enzimima40-tak vrsta enzima: nukleaze, proteaze, glikozidaze, lipaze, fosfolipaze,fosfataze, sulfatazekisele hidrolaze: potreban pH 5.0 (neaktivni u citosolu)specifična membrana koja omogućava transport molekula u citosolH+ pumpa koristi ATP hidrolizu za zakiseljavanjelizosomski membranski proteini su glikozilirani zbog zaštite od lize
sličnosti: kisele hidrolazeheterogenost: razne funkcije: razgradnja intra i ekstracelularnog debrijarazgradnja fagocitiranih mikroorganizamaproizvodnja hranjivih tvari
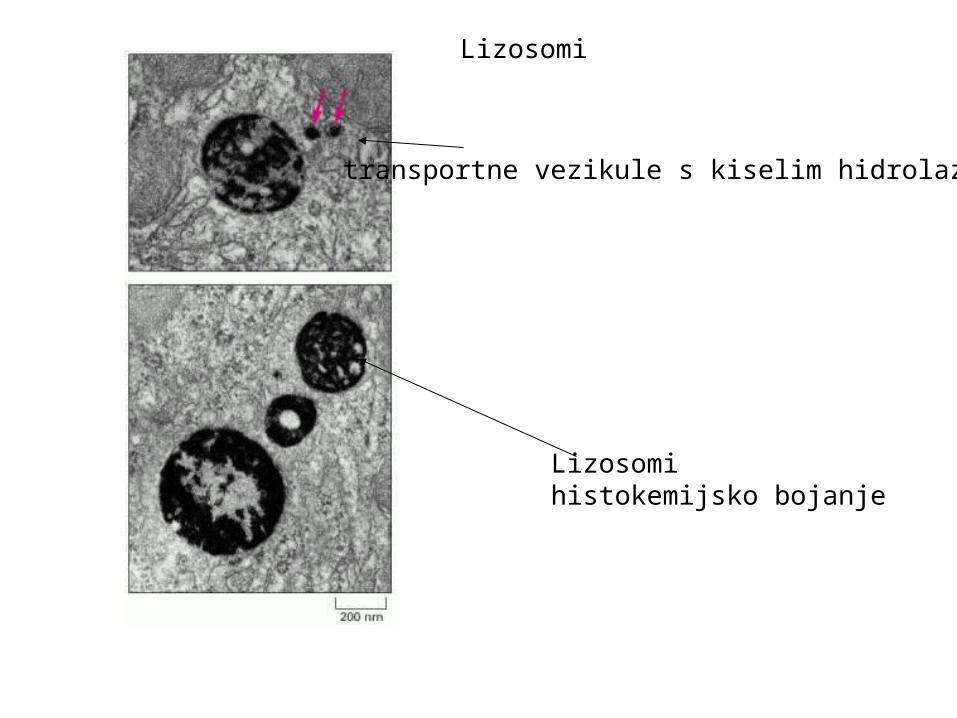
Lizosomihistokemijsko bojanje
transportne vezikule s kiselim hidrolazama
Lizosomi

Lizosomi: kisele hidrolaze

Tri puta degradacije u lizosomima1. makromolekule uzete iz okoline endocitozom (rani→kasni endosom + transportne vezikule s enzimima→lizosom:promjena pH; transport molekula u citoplazmu)2. autofagija- uklanjanje nepotrebnih ili oštećenih organelaorganel se okruži membranom – autofagosom3. fagocitoza velikih čestica i mikroorganizama – fagosomifuzija s lizosomom i digestija

Put prijenosa lizosomskih hidrolaza i membranskih proteina ulizosome
Sintetiziraju se u zrnatoj ER i prenose kroz GA do trans Golgi mreževezikule ugrađuju lizosomske proteine i isključuju ostale
lizosomske hidrolaze imaju marker: manoza-6 fosfat (M6P)
cis Golgij: N vezan oligosaharid se fosforilira na manozitrans Golgij mreža: M6P receptorski protein veže hidrolazereceptor s hidrolazama i adaptin pakiraju se u klatrinske vezikule vezikule pupaju i donose sadržaj kasnim endosomima
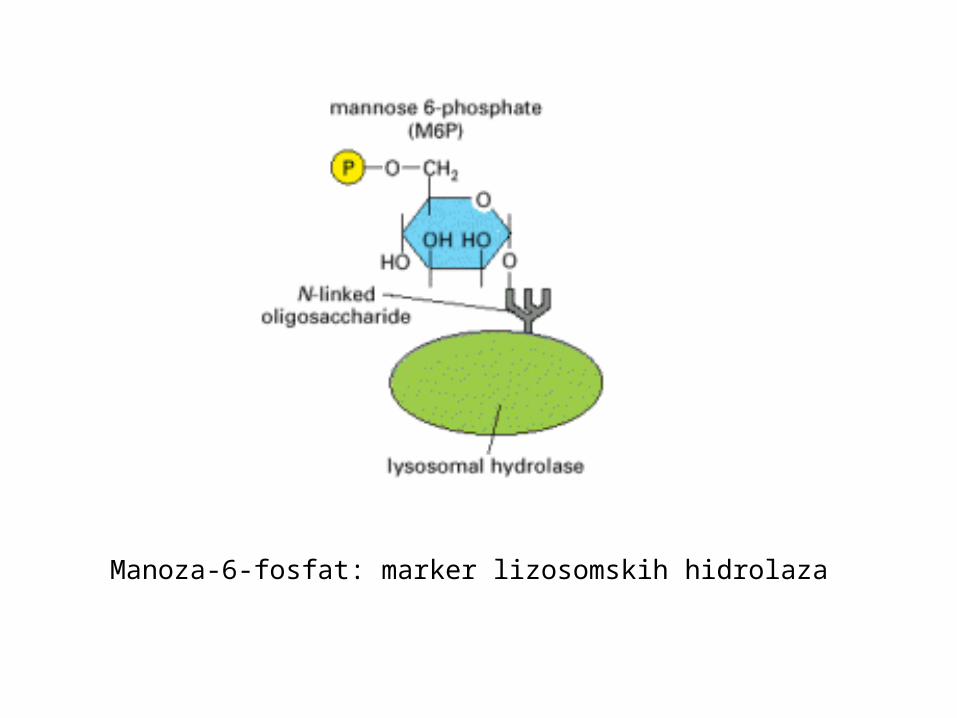
Manoza-6-fosfat: marker lizosomskih hidrolaza

Transport novosintetiziranih lizosomskih hidrolaza u lizosom
kasni endosom
pH 6

M6P receptorski protein veže oligosaharid kod pH 6,5 -6,7 u trans Golgiju
u kasnim endosomima je pH 6 i tada disocira receptor od hidrolazetijekom endosomskog sazrijevanja opada pH i tada se hidrolaze aktivirajuM6P receptori se izuzimaju u transportne vezikule koje pupaju iz kasnih endosomavraćaju se na membranu TGN i ponovo koriste
neke hidrolaze “zalutaju” na staničnu površinu, ali nisu aktivne zbog pHtransportiraju se i receptori M6P koji ih hvataju i vraćaju endocitozom posredovanom receptorima
M6P se dodaje samo na N-vezane oligosaharide koji su vezani za specifične proteine koji imaju određeni signalni uzorak

Prepoznavanje lizosomskih hidrolaza:manoza se fosforilira samo kod prepoznatih proteinasignalni uzorak – skupine susjednih aminokiselina na površini proteina
-cijepanje N acetilglukozaminskog ostatka, ostaje M6P-puno šećernih ostataka – visok afinitet signala
N acetilglukozaminfosfotransferaza
vezan GlcNAc-Pna manozu-ubikvitarne hidrolaze

Metaboličke bolesti:
I bolest (inclusion cell disease)recesivnanedostaju gotovo svi hidrolitički enzimi u fibroblastima nerazgrađene tvari se akumuliraju u lizosomimaenzimi se sekretiraju umjesto da se prenesu u lizosomeGlcNAc-fosfotransferaza u cis GN nedostaje pa se ne fosforiliraju i ne pakiraju u lizosome idu drugim transportnim putemu drugim tipovima stanica - drugi tipovi transporta, lizosomi normalnimembranski proteini lizosoma se transportiraju putem neovisnim o M6P
lizosomi:50 različitih enzima za razgradnju proteina, DNA RNA, polisaharida i lipida30 različitih nasljednih bolesti: lizosomske bolesti odlaganja

nedostatak pojedinih lizosomskih hidrolaza – nakupljanje supstrata
Hurlerova bolest (enzimi za cijepanje glikozaminoglikana)
Gaucherova bolest: nedostatak glukocerebrozidaze-liječenje: egzogeno uzimanje enzima;

Osnovni tipovi lizosomskih bolesti
mukopolisaharidozeglikoproteinozesfingolipidoze: Fabrijeva bolest Farberova Gaucherova Tay Sachsova metakromatska leukodistrofija Niemann-Pickova tip A i BDruge lipidozeBolest pohrane glikogena: Pompeova bolestVišestruka deficijencija enzimaPoremećaj transporta lizosomaDrugi poremećaji lizosomskih proteina

s
cerebrozid gangliozid
sulfat
sfingozinmasna kiselina
sfingomijelin
Sandhofovabolest
Fabrijeva
laktozilceramidoza
Gauscherova
Nieman-Pick
Farberova lipogranulomatoza
Krabbe
Metakromatska leukodistrofija
Tay-SachsGM2 gangliozidoza
GM1 gangliozidoza

Drugi putevi lizosoma:
lizosomska sekrecija (defekacija) – izbacivanje neprobavljivog debrisasamo u slučaju stresa
neke stanice: specifična fuzija lizosoma s plazma membranommelanociti imaju melanosome koji oslobađaju melanin ekstracelularnoegzocitozomuzimaju ga keratinociti za pigmentaciju kože
nepoznati svi putevi transporta u lizosome

-autofagosomi: neselektivna degradacija dugoživućih citoplazmatskih proteina, organela-makroautofagija (mitohondriji, peroksisomi, mikroorganizmi)-mikroautofagija-posredovana šaperonima (prepoznavanje motiva, unos u autofagosomputem receptora)
-gladovanje:selektivna razgradnja normalnih proteina koji se mogu nadomjestitiprepoznavanje sekvence Lys-Phe-Glu-Arg-Gln“žrtvovanje” za aminokiseline i energiju

Autofagija tijekom gladovanja: specifični receptori na mitohondriju-signali srodni signalima apoptoze (proteinima porodice Bcl-2)-uloga u tumorima-signalni putevi preživljenja: mTOR, PI3 kinaze
Hailey, D.W.,, Rambold, A.S., Satpute-Krishnan, P.,Mitra, K., Sougrat, R., Kim, P.KK., and Lippincott-Schwartz, Cell 140, 2010)..

Stanice eksprimiraju LC3 (homolog ATG8)fuzioniran s CFP. LC3 je u citosolu(gornja slika) i gladovanjem seregurutira u autofagosom i tu ostajedo fuzije s lizosomom.(Lippincott-Schwartz lab.)
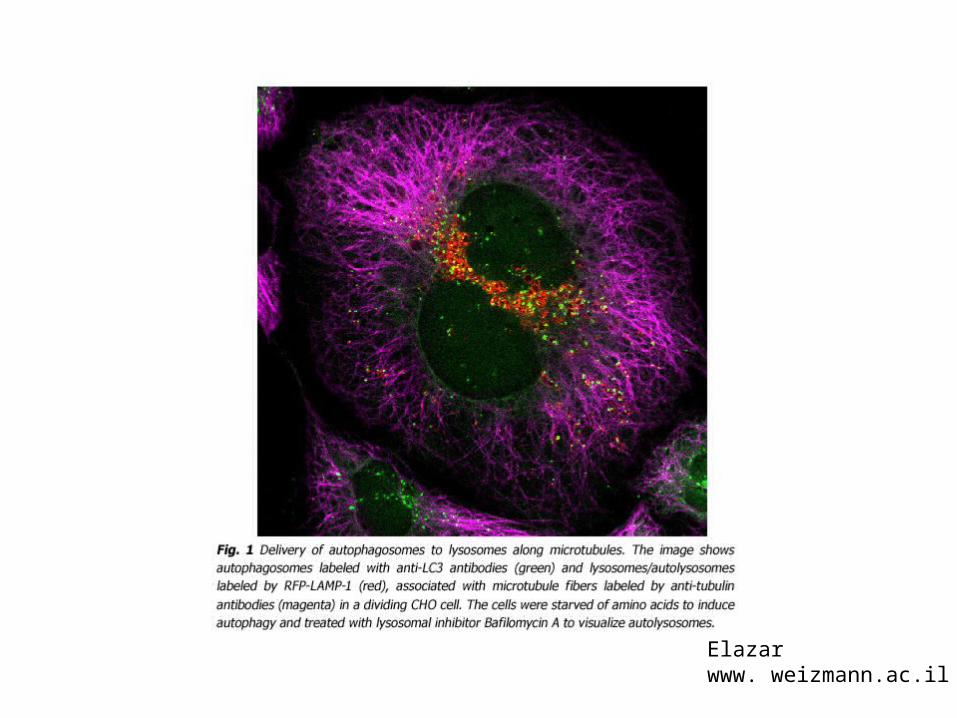
Elazarwww. weizmann.ac.il

Transport u stanicu s plazma membraneendocitoza


Stanica uzima hranjive tvari ili dijelove stanica ili čak stanice endocitozomMaterijal se uklapa u mali dio plazma membrane koji seinvaginira i zatim odvaja u obliku endocitičke vezikule
Dva tipa endocitoze:
fagocitoza (“stanično jedenje”): velike čestice: mikroorganizmi,mrtve stanice, putem velikih vezikula fagosoma (>250 nm)
pinocitoza (“stanično pijenje”): unošenje tekućine i otopina malim pinocitoznim vezikulama (100 nm) često kontinuirani proces
- endocitoza posredovana receptorima

Fagocitozaprotozoa: način prehrane“profesionalni” fagociti: makrofagi, neutrofili, dendritične staniceuklanjaju ostarjele stanice, apoptotske, mikroorganizme
veličina fagosoma ovisi o veličine unešenog materijalapinocitoza: male i uniformne vezikulerezidualno tijelo: neprobavljive tvari

pinocitoza: konstitutivan proces
fagocitoza: kontrolirani proces – receptorsko vezanje “okida” proces
-antitijela oblože mikroorganizam i Fc kraj antitijela je izložen prepoznaje ga Fc receptor na makrofagu i okida fagocitozustanica produži pseudopodije i okruži česticu, fuzijom membrananastaje fagosom-receptori za komponente komplementa okidaju fagocitozu-receptori za oligosaharide mikroorganizama-fosfatidilserin apoptotskih stanica
-stanice fagocitiraju i anorganske tvari (staklo, lateks, azbest)
-ne fagocitiraju žive animalne stanice – antagonistički signali

Makrofag fagocitira dva eritrocita

Pinocitozastanice “gutaju” dijelove svoje plazma membrane i zatim ih vraćajuna površinumakrofagi pojedu 100% svoje membrane u pola sata 25% tekućine svaki sat
ravnoteža endo i egzocitoze da se održi površina i obujam
endocitoza započinje formiranjem klatrinske jamice na plazmatskoj membranibrzo se odvaja od membrane kao klatrinska vezikulavezikula se oslobađa omotača
velik broj jamica i vezikula nastaje endocitozom kako se ekstracelularna tekućina zarobi u invaginaciji koja stvori vezikulu,bilo koja tvar otopljena u ekstracelularnoj tekućini se internalizira:endocitoza tekuće faze

Nastanak klatrinske vezikule na plazmatskoj membraniPerry and Gilbert 1979
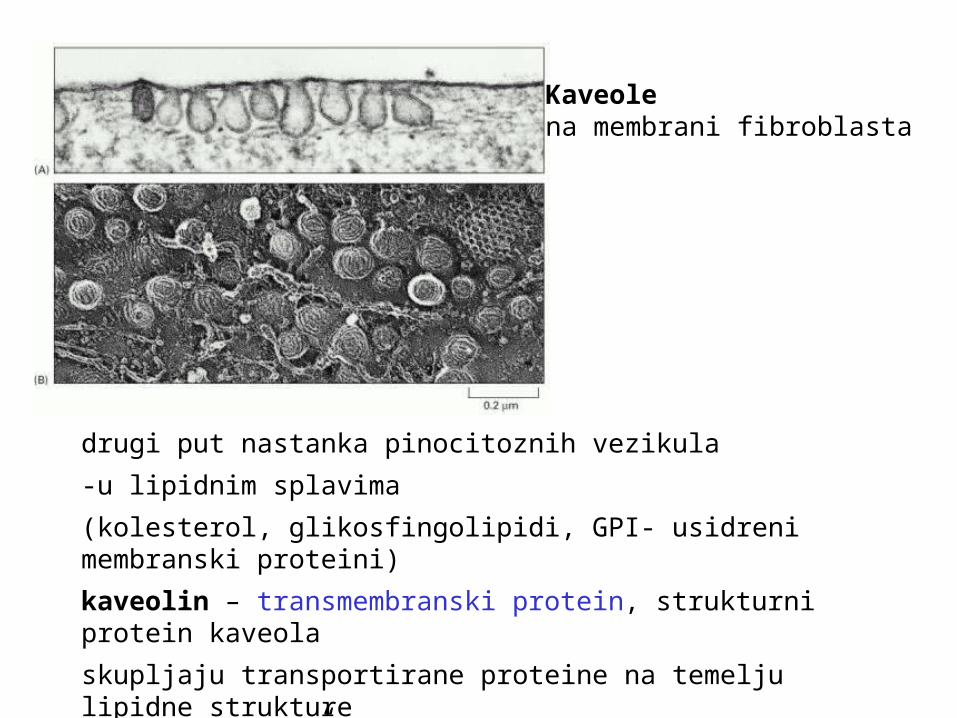
Kaveole na membrani fibroblasta
drugi put nastanka pinocitoznih vezikula
-u lipidnim splavima
(kolesterol, glikosfingolipidi, GPI- usidreni membranski proteini)
kaveolin – transmembranski protein, strukturni protein kaveola
skupljaju transportirane proteine na temelju lipidne strukture
prenose “teret” do endosoma ili na suprotnu plazmatsku membranu stanice (transcitoza)

Receptorom posredovana endocitoza:
mogućnost unosa i koncentriranja specifičnih makromolekulamakromolekula veže receptor, akumulira se u obloženoj jamicii ulazi kao klatrinska vezikula u stanicu
u istoj jamici razni receptori

Čestica LDL (low density lipoprotein)kolesterol se unosi posredstvom receptorau krvi se prenosi u obliku LDL
3 x 106 Da1500 molekula estera kolesterolamembrana: 800 fosfolipida500 neesterificiranih kolesterolnih molekulaprotein: 500 kDa
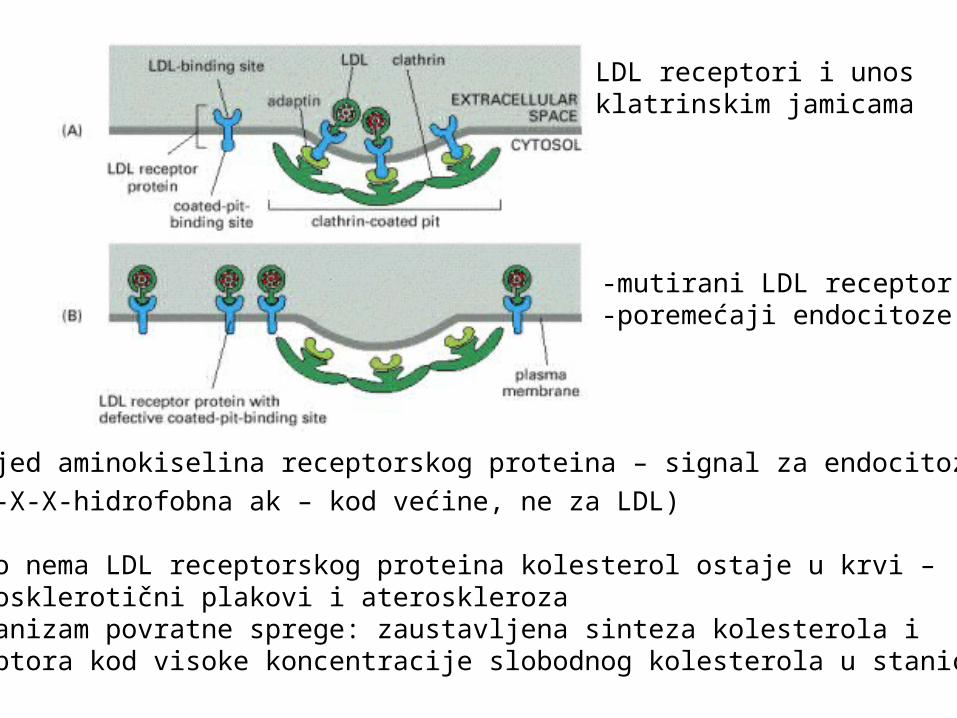
-slijed aminokiselina receptorskog proteina – signal za endocitozu
(Tyr-X-X-hidrofobna ak – kod većine, ne za LDL) -ako nema LDL receptorskog proteina kolesterol ostaje u krvi –aterosklerotični plakovi i ateroskleroza-mehanizam povratne sprege: zaustavljena sinteza kolesterola i receptora kod visoke koncentracije slobodnog kolesterola u stanici
-mutirani LDL receptor-poremećaji endocitoze
LDL receptori i unos klatrinskim jamicama

rani endosomi su glavna sortirajuća stanica u endocitičkom putu
rani endosom
→ kasni endosom zakiseljavanjem (vakulolarna H+ATPaza) i djelovanjem hidrolaza razgradnja u lizosomu
→neki receptori se vraćaju na istu ili drugu membranu
→hidrolaze se oslobađaju “biljega” i završavaju u lizosomu
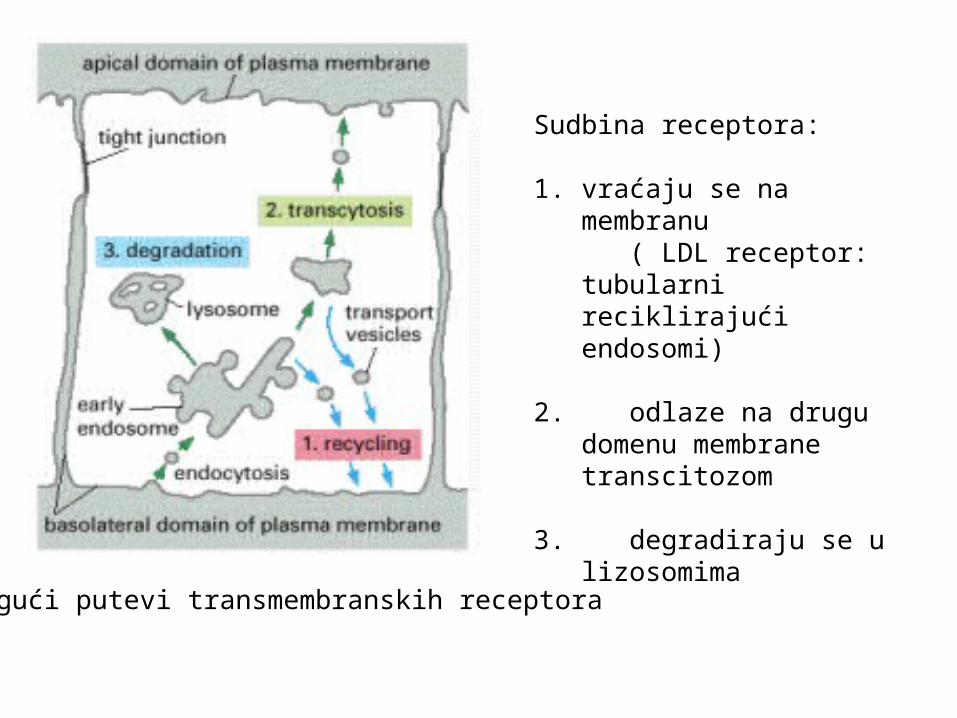
Mogući putevi transmembranskih receptora
Sudbina receptora:
1. vraćaju se na membranu ( LDL receptor: tubularni
reciklirajući endosomi)
2. odlaze na drugu domenu membrane transcitozom
3. degradiraju se u lizosomima
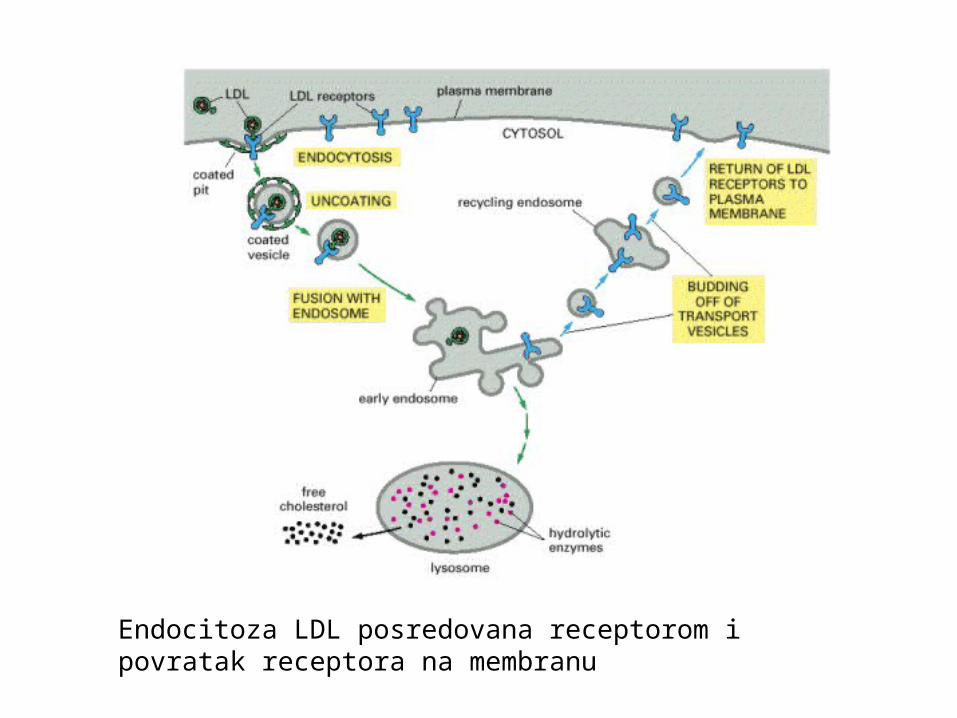
Endocitoza LDL posredovana receptorom ipovratak receptora na membranu

Recikliranje transferinskogreceptora (crveno) i opioidnireceptori u kasnim endosomima(zeleno) žuto: rani endosomi s oba receptora
transferinski receptor veže transferin, a transferin željezorecikliranje liganda – apotransferina, oslobađanje željeza-apotransferin se odvaja od receptoratek u neutralnom pH ekstracelularne tekućine

Transport multivezikularnogtjelešca mikrotubulimaspajaju se s transportnim vezikulamas kiselim hidrolazama i završavajukao lizosomi zakiseljavanjem
Multivezikularno tijelo – povezani migrirajući endosomi

Multivezikularno tjelešce biljke
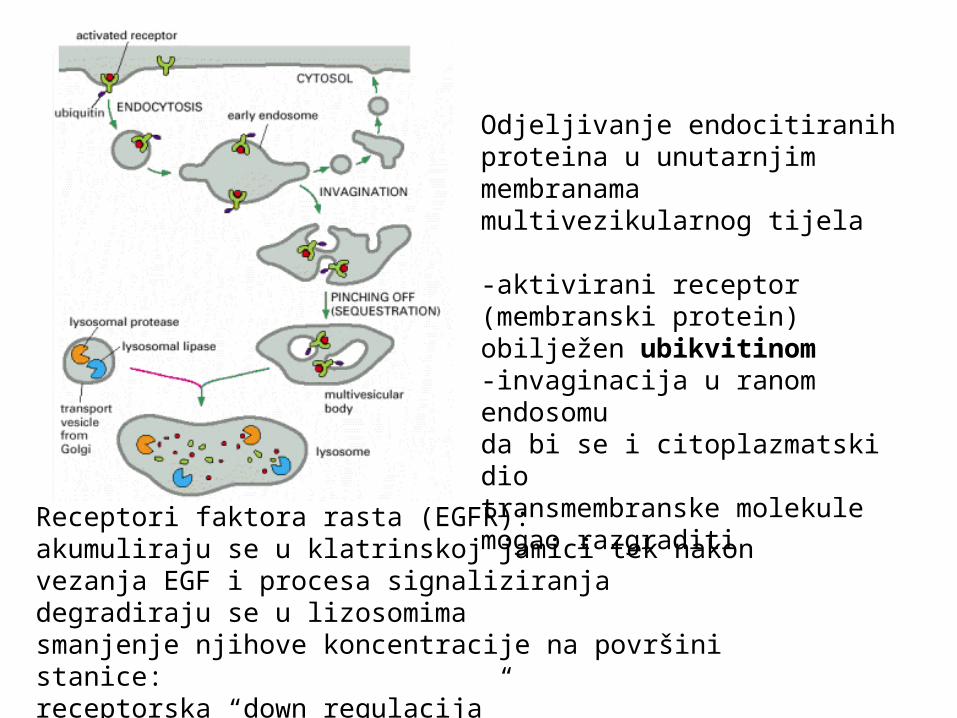
Receptori faktora rasta (EGFR): akumuliraju se u klatrinskoj jamici tek nakon vezanja EGF i procesa signaliziranjadegradiraju se u lizosomimasmanjenje njihove koncentracije na površini stanice: receptorska “down regulacija”
Odjeljivanje endocitiranih proteina u unutarnjim membranama multivezikularnog tijela
-aktivirani receptor (membranski protein) obilježen ubikvitinom-invaginacija u ranom endosomuda bi se i citoplazmatski dio transmembranske molekule mogao razgraditi

ESCRT: endosomal sorting complex required for transport:-sustav konzerviran od bakterija do viših eukariota-katalizira izrezivanje vrata membrane u biogenezi multivezikularnih tijela (MVB), citokinezu i pupanje virusa s omotačem-ubikvitinacija: signal za usmjeravanje na put ESCRT i lizosomsku degradaciju-funkcija nalik dinaminu, ali obrnute orijentacije (iznutra prema van)
Osnovne funkcije kompleksa ESCRT Modeli nastanka kompleksa
Hanson, Curr. Op. Cell Biol. 2009
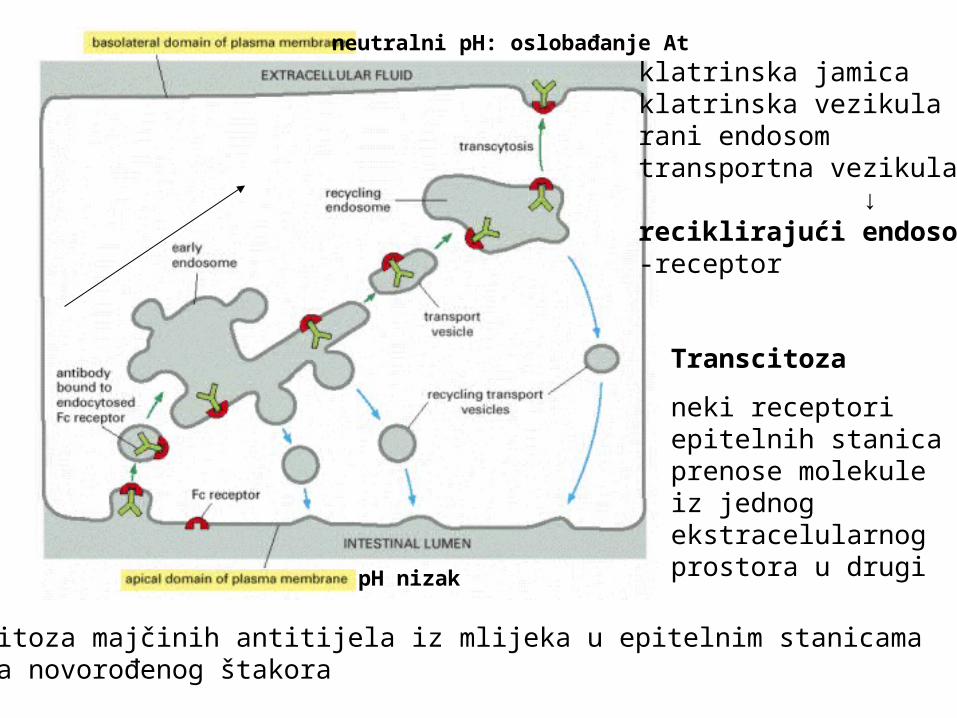
Transcitoza majčinih antitijela iz mlijeka u epitelnim stanicama crijeva novorođenog štakora
klatrinska jamicaklatrinska vezikularani endosomtransportna vezikula ↓reciklirajući endosom-receptor
Transcitoza
neki receptori epitelnih stanica prenose molekule iz jednog ekstracelularnog prostora u drugi
pH nizak
neutralni pH: oslobađanje At

Vezikule u epitelnoj stanici-bazolateralna i apikalna domena membrane komuniciraju različitim ranimendosomskim odjeljcima. Molekule bez signala za transcitozu ili recikliranjeimaju zajednički kasni endosomski odjeljak i razgradnju u lizosomima.
polarna stanica:
apikalna membrana
bazolateralnamembrana
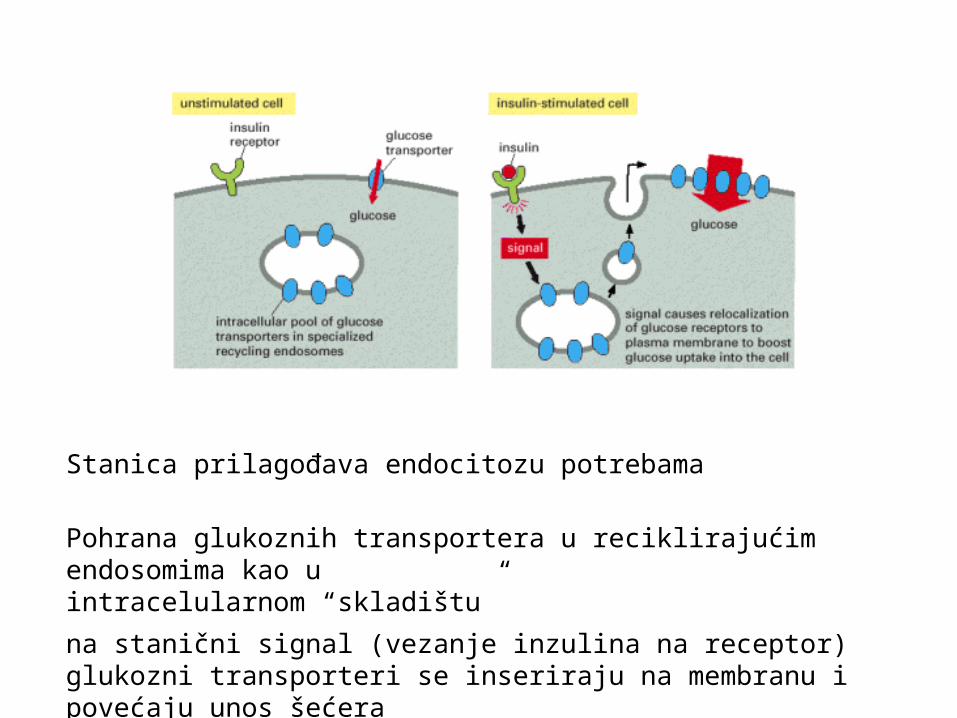
Stanica prilagođava endocitozu potrebama
Pohrana glukoznih transportera u reciklirajućim endosomima kao u intracelularnom “skladištu”
na stanični signal (vezanje inzulina na receptor) glukozni transporteri se inseriraju na membranu i povećaju unos šećera

Transport od trans Golgijeve mreže do stanične okolineegzocitoza

-membrana vezikula donosi komponente plazma membrane, a sadržajvezikule topive proteine koji se izluče u vanstanični prostorEgzocitoza: fuzija vezikula s plazma membranom
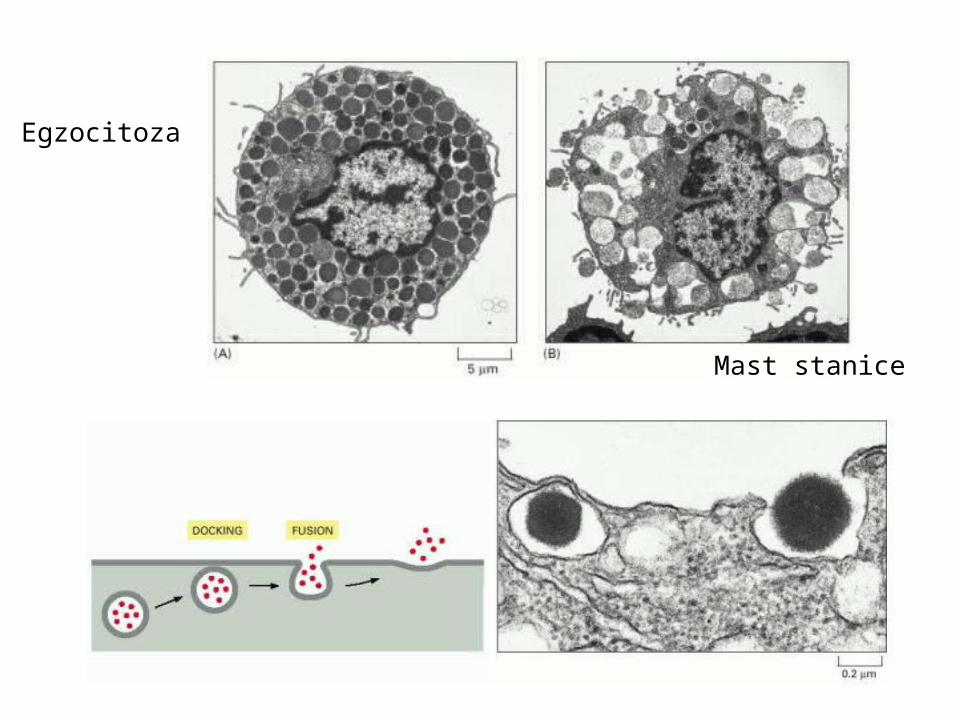
Egzocitoza
Mast stanice

Transportne vezikule koje stižu na plazmatsku membranu napuštaju trans Golgijevu mrežu.Nose membranske proteine i lipide potrebne za izgradnju membranei topive proteine koji se sekretiraju u ekstracelularni prostor.-sekrecija proteoglikana i glikoproteina ekstracelularnog matriksa
Konstitutivni sekretorni put
Regulirani sekretorni put: sekretorne vezikule se pohranjuju za kasnije oslobađanje, nakon signala (hormoni, neurotransmiteri, digestivni enzimi)

Tipovi sekretornih puteva

Tri klase proteina moraju se separirati prije napuštanja TGN:oni namijenjeni lizosomima (preko kasnih endosoma – biljezi) sekretornim vezikulama (biljezi)oni koji se direktno odnose do stanične površine
kod lizosomskih proteina postoji signal – manoza-6-fosfatni biljeganalogni signali postoje kod sekretornih proteina i vezikula
ako nema biljega kod nepolarne stanice proteini idu direktno navanjsku staničnu membranu – “obavezni” put (konstitutivni)
kod polariziranih stanica postoje složeni sustavi usmjeravanja
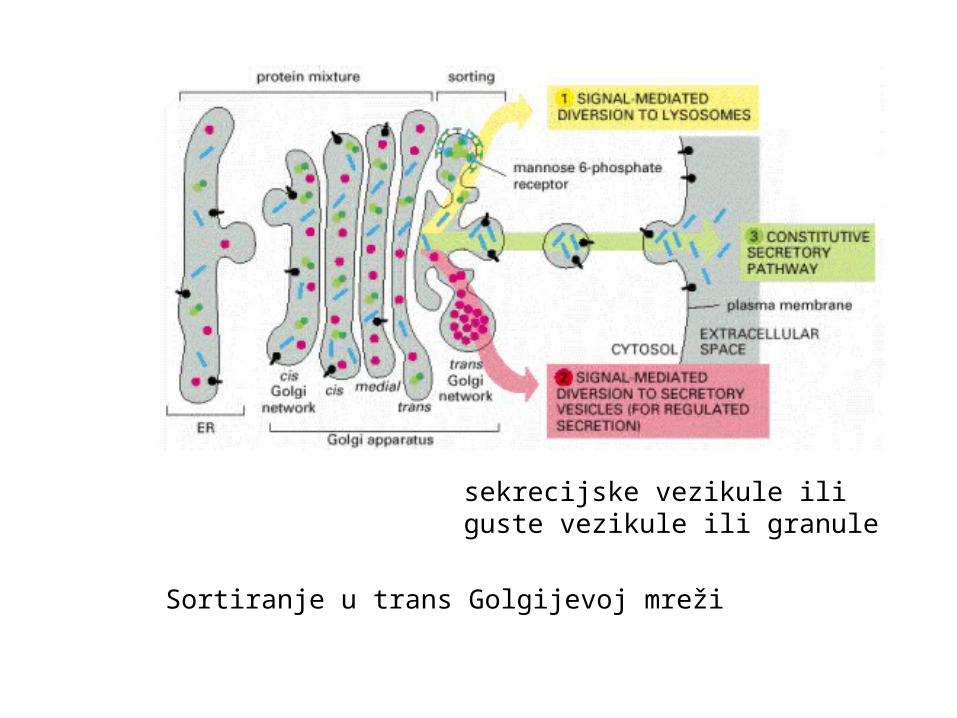
Sortiranje u trans Golgijevoj mreži
sekrecijske vezikule iliguste vezikule ili granule

Nastanak sekrecijskih vezikula: kako sazrijevaju bivaju koncentriranije i kiselije sadržaj lumena i višak membrane “odnose” klatrinske vezikule-opći signali sortiranja postoje u svim stanicama
sekrecija beta stanica gušterače-vezikule s proinzulinom
zrele sekretorne vezikule višenemaju klatrin - crne točke (At)

Egzocitoza sekretornih vezikula:inzulin stanica gušterače-naglo oslobađanje velike količine tvari
-kondenzacija

Proteini se mogu proteolitički procesirati tijekom nastanka sekretorne vezikule
Mnogi hormoni i neuropeptidi i probavni enzimi sintetizirajuse kao pre-pro-peptidi: signal za ERpoliproteini koji se cijepaju na više proteinapoliproteini za različite proteine ovisno o stanici
cijepanje može početi u trans Golgiju, u sekretornim vezikulama i dovršiti seekstracelularno
-regulacija djelovanja - odgađanje aktivacije u stanici-premaleni za kotranslacijski transport

Procesirajući putevi u nastanku prohormona proopiomelanokortina: jedan prekursor proizvodi razne peptidne hormone
encefalini: premaleni (5 amk) za kotranslacijski transport i pakiranje

Neuropeptidi imaju reguliranu sekreciju
putuju duž aksona pomoću mikrotubula (usmjereno)
čekaju signal:neurotransmiter → akcijski potencijal → otvaranje kalcijevih kanala reguliranih naponom → ulaz kalcijavezanje Ca za specifične senzore okida fuziju plazma membrane(sinaptičke vezikule) i oslobađanje u ekstracelularni prostorsudjeluju proteini SNARE koji su vjerojatno već inicirani, tako dainfluks kalcija izazove fuziju
upotreba samo nekoliko vezikula istovremenotrenutni odgovorlokalna reakcija

Živčana stanica i egzocitoza sinaptičkih vezikula-da bi se omogućila trenutna reakcija, sinaptičke se vezikule nose do membrane -nova se vezikula stvara endocitozom i donosi do endosoma-pupa iz endosoma i “puni” se neurotransmiterom pomoću proteina nosača-egzocitoza i sekrecija kao odgovor na akcijski potencijal-recikliranje samih sinaptičkih membrana

Egzocitoza kao lokalizirani odgovor (kontakt sa zrncem)mast stanice otpuštaju histamin

Komponente sekretorne vezikule se odmah uklanjajus plazma membrane, endocitozom, da se ne bi povećalapovršina stanice
rast stanice: egzocitoza ↑ i porast membrane
nepoznati mehanizmi koordinacije egzocitoze i endocitoze

Polarizirane stanice mogu imati različite proteine sekretirane narazličitim polovima
Proteini usmjereni bazolateralno imaju sortirajući signal u citoplazmatskom repu.
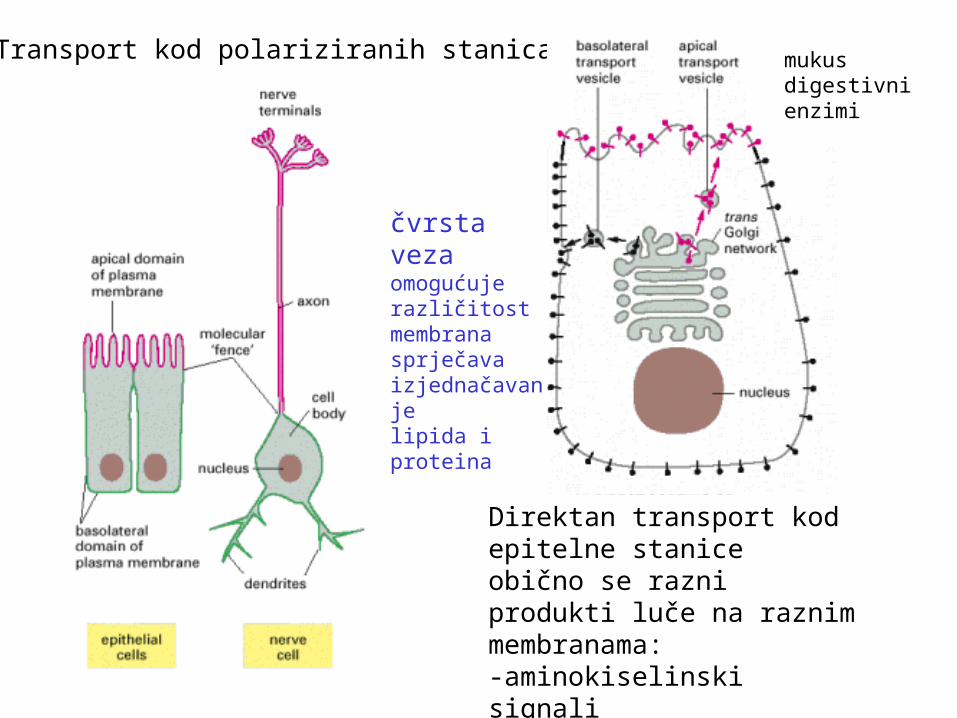
Transport kod polariziranih stanica
Direktan transport kod epitelne staniceobično se razni produkti luče na raznim membranama:-aminokiselinski signali-indirektan put - transcitoza
čvrstavezaomogućuje različitost membranasprječavaizjednačavanjelipida i proteina
mukusdigestivni enzimi

-polarni (apikalni) transport komponenata lipidnih splavi -apikalno su glikosfingolipidi koji štite od pH i digestivnih enzima proteini usidreni glikozilfosfatidilinozitolom GPI-proteini s GPI udružuju se s glikosfingolipidima u lipidnim splavima u trans GN-lektini (vežu ugljikohidrate) stabiliziraju lipidne splavi-sa specifičnim teretom, splavi pupaju iz trans Golgija u transportne vezikuleusmjerene prema apikalnoj membrani

Lipid Rafts enriched in SM and Cholesterol(screen capture from: http://multimedia.mcb.harvard.edu/anim_innerlife.html

Prijenos signala putem transsinaptičkog prijenosa transmembranskog proteina i liganda (Wnt) vezikulama
Korkut i sur. Cell 139, 393-404,2009.
Hipoteza prijenosa vezikule

DNARNA
proteini
proteinimodifikacija proteina
lipidi
modifikacija lipidaproteiniproteoglikani
peroksisomi
hER: sinteza proteinaN glikozilacijaglikolipidna sidra proteina
gER: fosfolipidi steroli, ceramidi detoksifikacija spremište Ca
Golgi:složeni glikoproteinisloženi glikolipidiproteoglikanisloženi polisaharidi





























