Embed Size (px)
Citation preview

Immunity, Vol. 18, 121–130, January, 2003, Copyright 2003 by Cell Press
Virus Subversion of the MHC Class IPeptide-Loading Complex
occupied with peptide, fully assembled class I mole-cules (H-chain, �2m, and peptide) are released from theloading complex, exit the ER, and transit to the cell
Lonnie Lybarger,1,4 Xiaoli Wang,1,4
Michael R. Harris,2 Herbert W. Virgin IV,3
and Ted H. Hansen1,*surface (Cresswell, 2000; Williams et al., 2002a).1Department of Genetics
The importance of the class I antigen presentation2 Department of Pediatricspathway in controlling intracellular pathogens is high-3 Department of Pathology & Immunologylighted by the diverse mechanisms that many virusesWashington University School of Medicinehave evolved to specifically disrupt this pathwaySt. Louis, Missouri 63110(Ploegh, 1998; Tortorella et al., 2000). Recently, a newfamily of virus-encoded molecules has been discoveredthat interferes with class I expression and, in someSummarycases, the expression of other immune modulators.These proteins all share a conserved zinc binding do-Many viral proteins modulate class I expression, yet,main characteristic of the PHD/LAP family (Ashland etin general, their mechanisms of specific class I recog-al., 1995; Saha et al., 1995) at their N termini. Genesnition are poorly understood. The mK3 protein ofencoding such proteins have been described in herpes-�2-Herpesvirus 68 targets the degradation of nascentviruses (Albrecht et al., 1992; Nicholas et al., 1997; vanclass I molecules via the ubiquitin/proteasome path-Santen, 1991; Virgin et al., 1997) and poxviruses (Guerinway. Here, we identify cellular components of the MHCet al., 2002). The best-characterized examples are theclass I assembly machinery, TAP and tapasin, that arekK3 and kK5 molecules of Kaposi’s sarcoma associatedrequired for mK3 function. mK3 failed to regulate classherpesvirus (KSHV) and the mK3 protein of �-Herpesvi-I in TAP- or tapasin-deficient cells, and mK3 interactedruses 68. These proteins possess a characteristic mem-with TAP/tapasin, even in the absence of class I. Ex-brane topology (type III) with the N- and C termini bothpression of mK3 resulted in the ubiquitination of TAP/located in the cytosol and only a short connecting pep-tapasin-associated class I, and mutants of class I inca-tide within the ER or Golgi lumen (Boname and Steven-pable of TAP/tapasin interaction were unaffected byson, 2001; Sanchez et al., 2002). All of these moleculesmK3. Thus, mK3 subverts TAP/tapasin to specificallydownregulate surface class I, but their sites of actiontarget class I molecules for destruction.appear to be quite distinct. kK5 and kK3 enhance endo-cytosis of class I molecules from the cell surface (Cos-Introductioncoy and Ganem, 2000; Ishido et al., 2000; Haque et al.,2001) and direct them to lysosomes for degradationCD8� T cells play a critical role in the control of virus(Hewitt et al., 2002; Lorenzo et al., 2002; Means et al.,infection, through the recognition of viral peptides pre-2002; Coscoy and Ganem, 2000). Furthermore, kK3 hassented by MHC class I molecules on infected cells. Therecently been shown to associate with class I in a post-class I antigen presentation pathway initiates in the ERER compartment (Hewitt et al., 2002). Though structur-and involves the concerted activity of several accessoryally related, mK3 induces the turnover of ER-residentmolecules (Cresswell, 2000; Williams et al., 2002a). Newlyclass I molecules (Stevenson et al., 2000). mK3-medi-synthesized class I heavy chains (H-chains) first associateated degradation is quite rapid and involves physicalwith calnexin (Degan and Williams, 1991). Next, in theassociation of mK3 with class I (Stevenson et al., 2000;presence of �2-microglobulin (�2m), H-chain/�2m hetero-Boname and Stevenson, 2001; Yu et al., 2002). However,
dimers enter the peptide-loading complex, a collectionit remains unclear whether mK3 binds directly to class
of molecules that facilitate the acquisition of high-affinityI or through an intermediate host protein(s).
peptides by class I (Ortmann et al., 1994; Suh et al., The common feature of class I targeting by mk3 homo-1994). This complex contains H-chain, �2m, TAP, ta- logs is their ubiquitin ligase activity imparted by thepasin, calreticulin, and Erp57 (Cresswell, 2000; Williams conserved N-terminal PHD/LAP domain. Ubiquitinatedet al., 2002a). TAP, a heterodimer of TAP-1 and TAP-2 H-chains have been detected in the presence of all threesubunits, forms a peptide pump that transports class I of these proteins, and the PHD/LAP domains are re-binding peptides from the cytosol to the ER lumen. Ta- quired for this process (Boname and Stevenson, 2001;pasin is a class I-specific chaperone that significantly Hewitt et al., 2002; Coscoy et al., 2001). Direct ubiquitinimproves the quality of peptides bound by class I and ligase activity has been demonstrated for the kK5 pro-is required for the formation of the loading complex tein (Coscoy et al., 2001), and it is therefore likely that(Sadasivan et al., 1996; Williams et al., 2002b). Calreti- all of the members of this family have similar activity.culin and Erp57, though serving as chaperones for many Class I molecules are ubiquitinated on their cytosolicglycoproteins, are also bona fide members of the loading tails (Boname and Stevenson, 2001; Hewitt et al. 2002;complex and increase the efficiency of peptide loading Coscoy et al., 2001), and this modification triggers theonto the H-chain (Gao et al., 2002; Dick et al., 2002). downstream events in mK3-, kK3-, or kK5-mediatedOnce the peptide binding groove of the H-chain is stably degradation. Although a motif has been identified in
the tail of class I that is required for mK3-mediateddegradation (Boname and Stevenson, 2001), this se-*Correspondence: [email protected]
4 These authors contributed equally to this work. quence alone is unlikely to account for the remarkable
Immunity122
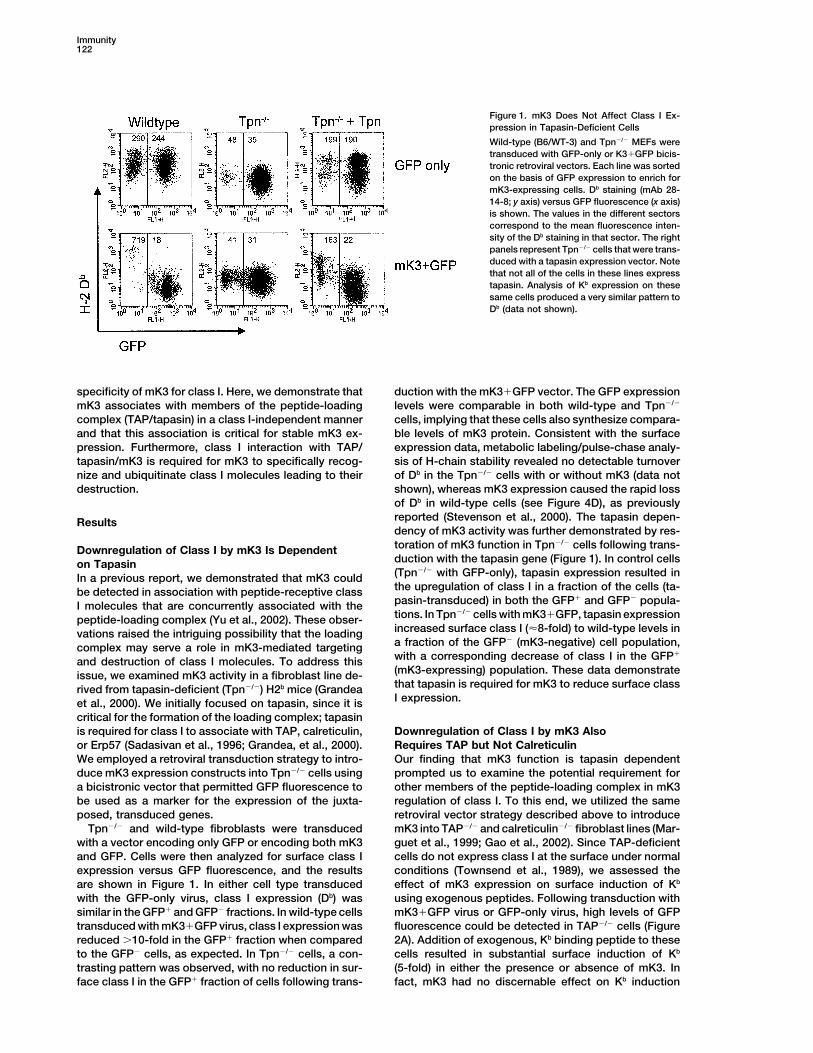
Figure 1. mK3 Does Not Affect Class I Ex-pression in Tapasin-Deficient Cells
Wild-type (B6/WT-3) and Tpn�/� MEFs weretransduced with GFP-only or K3�GFP bicis-tronic retroviral vectors. Each line was sortedon the basis of GFP expression to enrich formK3-expressing cells. Db staining (mAb 28-14-8; y axis) versus GFP fluorescence (x axis)is shown. The values in the different sectorscorrespond to the mean fluorescence inten-sity of the Db staining in that sector. The rightpanels represent Tpn�/� cells that were trans-duced with a tapasin expression vector. Notethat not all of the cells in these lines expresstapasin. Analysis of Kb expression on thesesame cells produced a very similar pattern toDb (data not shown).
specificity of mK3 for class I. Here, we demonstrate that duction with the mK3�GFP vector. The GFP expressionlevels were comparable in both wild-type and Tpn�/�mK3 associates with members of the peptide-loading
complex (TAP/tapasin) in a class I-independent manner cells, implying that these cells also synthesize compara-ble levels of mK3 protein. Consistent with the surfaceand that this association is critical for stable mK3 ex-
pression. Furthermore, class I interaction with TAP/ expression data, metabolic labeling/pulse-chase analy-sis of H-chain stability revealed no detectable turnovertapasin/mK3 is required for mK3 to specifically recog-
nize and ubiquitinate class I molecules leading to their of Db in the Tpn�/� cells with or without mK3 (data notshown), whereas mK3 expression caused the rapid lossdestruction.of Db in wild-type cells (see Figure 4D), as previouslyreported (Stevenson et al., 2000). The tapasin depen-Resultsdency of mK3 activity was further demonstrated by res-toration of mK3 function in Tpn�/� cells following trans-Downregulation of Class I by mK3 Is Dependentduction with the tapasin gene (Figure 1). In control cellson Tapasin(Tpn�/� with GFP-only), tapasin expression resulted inIn a previous report, we demonstrated that mK3 couldthe upregulation of class I in a fraction of the cells (ta-be detected in association with peptide-receptive classpasin-transduced) in both the GFP� and GFP� popula-I molecules that are concurrently associated with thetions. In Tpn�/� cells with mK3�GFP, tapasin expressionpeptide-loading complex (Yu et al., 2002). These obser-increased surface class I (�8-fold) to wild-type levels invations raised the intriguing possibility that the loadinga fraction of the GFP� (mK3-negative) cell population,complex may serve a role in mK3-mediated targetingwith a corresponding decrease of class I in the GFP�
and destruction of class I molecules. To address this(mK3-expressing) population. These data demonstrateissue, we examined mK3 activity in a fibroblast line de-that tapasin is required for mK3 to reduce surface classrived from tapasin-deficient (Tpn�/�) H2b mice (GrandeaI expression.et al., 2000). We initially focused on tapasin, since it is
critical for the formation of the loading complex; tapasinis required for class I to associate with TAP, calreticulin, Downregulation of Class I by mK3 Also
Requires TAP but Not Calreticulinor Erp57 (Sadasivan et al., 1996; Grandea, et al., 2000).We employed a retroviral transduction strategy to intro- Our finding that mK3 function is tapasin dependent
prompted us to examine the potential requirement forduce mK3 expression constructs into Tpn�/� cells usinga bicistronic vector that permitted GFP fluorescence to other members of the peptide-loading complex in mK3
regulation of class I. To this end, we utilized the samebe used as a marker for the expression of the juxta-posed, transduced genes. retroviral vector strategy described above to introduce
mK3 into TAP�/� and calreticulin�/� fibroblast lines (Mar-Tpn�/� and wild-type fibroblasts were transducedwith a vector encoding only GFP or encoding both mK3 guet et al., 1999; Gao et al., 2002). Since TAP-deficient
cells do not express class I at the surface under normaland GFP. Cells were then analyzed for surface class Iexpression versus GFP fluorescence, and the results conditions (Townsend et al., 1989), we assessed the
effect of mK3 expression on surface induction of Kbare shown in Figure 1. In either cell type transducedwith the GFP-only virus, class I expression (Db) was using exogenous peptides. Following transduction with
mK3�GFP virus or GFP-only virus, high levels of GFPsimilar in the GFP� and GFP� fractions. In wild-type cellstransduced with mK3�GFP virus, class I expression was fluorescence could be detected in TAP�/� cells (Figure
2A). Addition of exogenous, Kb binding peptide to thesereduced �10-fold in the GFP� fraction when comparedto the GFP� cells, as expected. In Tpn�/� cells, a con- cells resulted in substantial surface induction of Kb
(5-fold) in either the presence or absence of mK3. Intrasting pattern was observed, with no reduction in sur-face class I in the GFP� fraction of cells following trans- fact, mK3 had no discernable effect on Kb induction
TAP/Tapasin Requirement for mK3 Function123
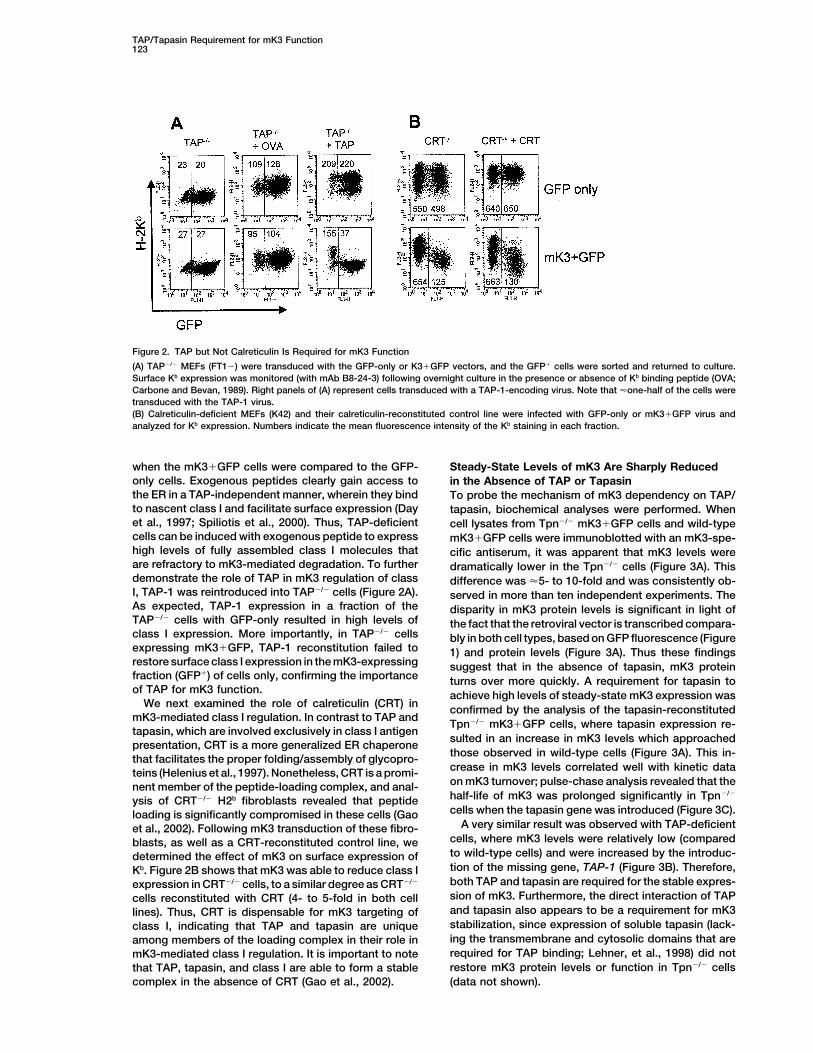
Figure 2. TAP but Not Calreticulin Is Required for mK3 Function
(A) TAP�/� MEFs (FT1�) were transduced with the GFP-only or K3�GFP vectors, and the GFP� cells were sorted and returned to culture.Surface Kb expression was monitored (with mAb B8-24-3) following overnight culture in the presence or absence of Kb binding peptide (OVA;Carbone and Bevan, 1989). Right panels of (A) represent cells transduced with a TAP-1-encoding virus. Note that �one-half of the cells weretransduced with the TAP-1 virus.(B) Calreticulin-deficient MEFs (K42) and their calreticulin-reconstituted control line were infected with GFP-only or mK3�GFP virus andanalyzed for Kb expression. Numbers indicate the mean fluorescence intensity of the Kb staining in each fraction.
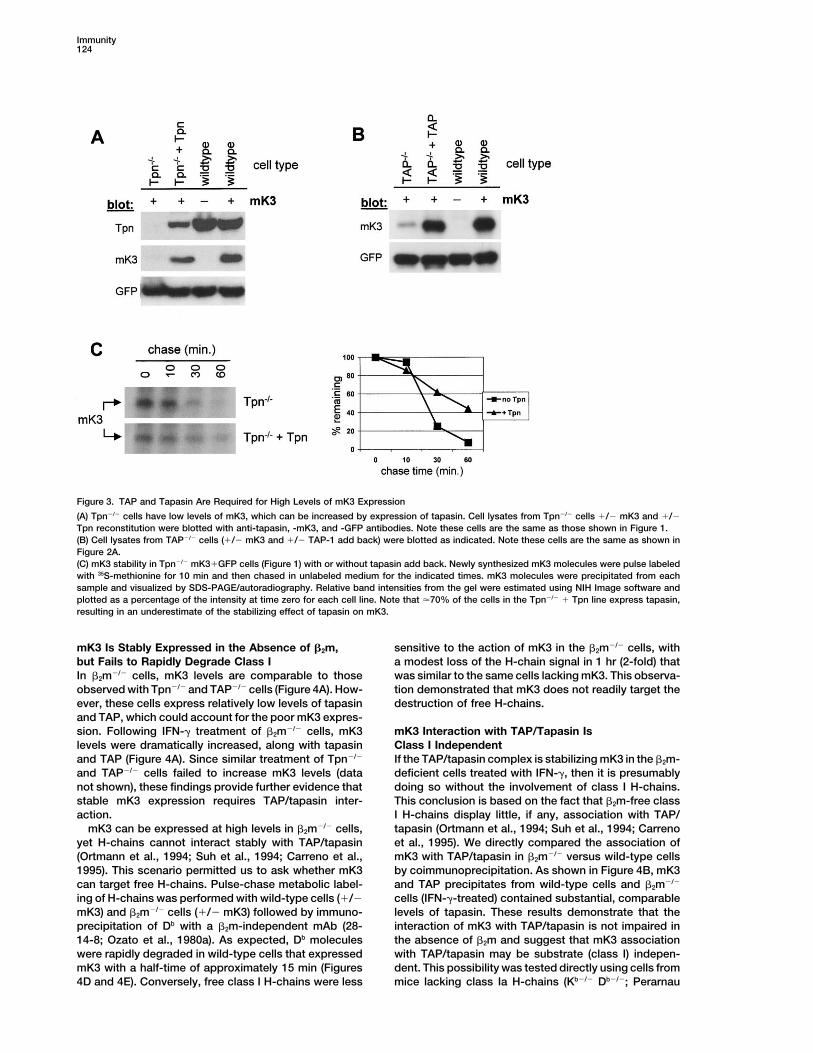
when the mK3�GFP cells were compared to the GFP- Steady-State Levels of mK3 Are Sharply Reducedonly cells. Exogenous peptides clearly gain access to in the Absence of TAP or Tapasinthe ER in a TAP-independent manner, wherein they bind To probe the mechanism of mK3 dependency on TAP/to nascent class I and facilitate surface expression (Day tapasin, biochemical analyses were performed. Whenet al., 1997; Spiliotis et al., 2000). Thus, TAP-deficient cell lysates from Tpn�/� mK3�GFP cells and wild-typecells can be induced with exogenous peptide to express mK3�GFP cells were immunoblotted with an mK3-spe-high levels of fully assembled class I molecules that cific antiserum, it was apparent that mK3 levels wereare refractory to mK3-mediated degradation. To further dramatically lower in the Tpn�/� cells (Figure 3A). Thisdemonstrate the role of TAP in mK3 regulation of class difference was �5- to 10-fold and was consistently ob-I, TAP-1 was reintroduced into TAP�/� cells (Figure 2A). served in more than ten independent experiments. TheAs expected, TAP-1 expression in a fraction of the disparity in mK3 protein levels is significant in light ofTAP�/� cells with GFP-only resulted in high levels of the fact that the retroviral vector is transcribed compara-class I expression. More importantly, in TAP�/� cells bly in both cell types, based on GFP fluorescence (Figureexpressing mK3�GFP, TAP-1 reconstitution failed to 1) and protein levels (Figure 3A). Thus these findingsrestore surface class I expression in the mK3-expressing suggest that in the absence of tapasin, mK3 proteinfraction (GFP�) of cells only, confirming the importance turns over more quickly. A requirement for tapasin toof TAP for mK3 function. achieve high levels of steady-state mK3 expression was
We next examined the role of calreticulin (CRT) in confirmed by the analysis of the tapasin-reconstitutedmK3-mediated class I regulation. In contrast to TAP and
Tpn�/� mK3�GFP cells, where tapasin expression re-tapasin, which are involved exclusively in class I antigen
sulted in an increase in mK3 levels which approachedpresentation, CRT is a more generalized ER chaperone
those observed in wild-type cells (Figure 3A). This in-that facilitates the proper folding/assembly of glycopro-crease in mK3 levels correlated well with kinetic datateins (Helenius et al., 1997). Nonetheless, CRT is a promi-on mK3 turnover; pulse-chase analysis revealed that thenent member of the peptide-loading complex, and anal-half-life of mK3 was prolonged significantly in Tpn�/�
ysis of CRT�/� H2b fibroblasts revealed that peptidecells when the tapasin gene was introduced (Figure 3C).loading is significantly compromised in these cells (Gao
A very similar result was observed with TAP-deficientet al., 2002). Following mK3 transduction of these fibro-cells, where mK3 levels were relatively low (comparedblasts, as well as a CRT-reconstituted control line, weto wild-type cells) and were increased by the introduc-determined the effect of mK3 on surface expression oftion of the missing gene, TAP-1 (Figure 3B). Therefore,Kb. Figure 2B shows that mK3 was able to reduce class Iboth TAP and tapasin are required for the stable expres-expression in CRT�/� cells, to a similar degree as CRT�/�
sion of mK3. Furthermore, the direct interaction of TAPcells reconstituted with CRT (4- to 5-fold in both celland tapasin also appears to be a requirement for mK3lines). Thus, CRT is dispensable for mK3 targeting ofstabilization, since expression of soluble tapasin (lack-class I, indicating that TAP and tapasin are uniqueing the transmembrane and cytosolic domains that areamong members of the loading complex in their role inrequired for TAP binding; Lehner, et al., 1998) did notmK3-mediated class I regulation. It is important to noterestore mK3 protein levels or function in Tpn�/� cellsthat TAP, tapasin, and class I are able to form a stable
complex in the absence of CRT (Gao et al., 2002). (data not shown).
Immunity124
Figure 3. TAP and Tapasin Are Required for High Levels of mK3 Expression
(A) Tpn�/� cells have low levels of mK3, which can be increased by expression of tapasin. Cell lysates from Tpn�/� cells �/� mK3 and �/�Tpn reconstitution were blotted with anti-tapasin, -mK3, and -GFP antibodies. Note these cells are the same as those shown in Figure 1.(B) Cell lysates from TAP�/� cells (�/� mK3 and �/� TAP-1 add back) were blotted as indicated. Note these cells are the same as shown inFigure 2A.(C) mK3 stability in Tpn�/� mK3�GFP cells (Figure 1) with or without tapasin add back. Newly synthesized mK3 molecules were pulse labeledwith 35S-methionine for 10 min and then chased in unlabeled medium for the indicated times. mK3 molecules were precipitated from eachsample and visualized by SDS-PAGE/autoradiography. Relative band intensities from the gel were estimated using NIH Image software andplotted as a percentage of the intensity at time zero for each cell line. Note that �70% of the cells in the Tpn�/� � Tpn line express tapasin,resulting in an underestimate of the stabilizing effect of tapasin on mK3.
mK3 Is Stably Expressed in the Absence of �2m, sensitive to the action of mK3 in the �2m�/� cells, witha modest loss of the H-chain signal in 1 hr (2-fold) thatbut Fails to Rapidly Degrade Class I
In �2m�/� cells, mK3 levels are comparable to those was similar to the same cells lacking mK3. This observa-tion demonstrated that mK3 does not readily target theobserved with Tpn�/� and TAP�/� cells (Figure 4A). How-
ever, these cells express relatively low levels of tapasin destruction of free H-chains.and TAP, which could account for the poor mK3 expres-sion. Following IFN-� treatment of �2m�/� cells, mK3 mK3 Interaction with TAP/Tapasin Is
Class I Independentlevels were dramatically increased, along with tapasinand TAP (Figure 4A). Since similar treatment of Tpn�/� If the TAP/tapasin complex is stabilizing mK3 in the �2m-
deficient cells treated with IFN-�, then it is presumablyand TAP�/� cells failed to increase mK3 levels (datanot shown), these findings provide further evidence that doing so without the involvement of class I H-chains.
This conclusion is based on the fact that �2m-free classstable mK3 expression requires TAP/tapasin inter-action. I H-chains display little, if any, association with TAP/
tapasin (Ortmann et al., 1994; Suh et al., 1994; CarrenomK3 can be expressed at high levels in �2m�/� cells,yet H-chains cannot interact stably with TAP/tapasin et al., 1995). We directly compared the association of
mK3 with TAP/tapasin in �2m�/� versus wild-type cells(Ortmann et al., 1994; Suh et al., 1994; Carreno et al.,1995). This scenario permitted us to ask whether mK3 by coimmunoprecipitation. As shown in Figure 4B, mK3
and TAP precipitates from wild-type cells and �2m�/�can target free H-chains. Pulse-chase metabolic label-ing of H-chains was performed with wild-type cells (�/� cells (IFN-�-treated) contained substantial, comparable
levels of tapasin. These results demonstrate that themK3) and �2m�/� cells (�/� mK3) followed by immuno-precipitation of Db with a �2m-independent mAb (28- interaction of mK3 with TAP/tapasin is not impaired in
the absence of �2m and suggest that mK3 association14-8; Ozato et al., 1980a). As expected, Db moleculeswere rapidly degraded in wild-type cells that expressed with TAP/tapasin may be substrate (class I) indepen-
dent. This possibility was tested directly using cells frommK3 with a half-time of approximately 15 min (Figures4D and 4E). Conversely, free class I H-chains were less mice lacking class Ia H-chains (Kb�/� Db�/�; Perarnau
TAP/Tapasin Requirement for mK3 Function125
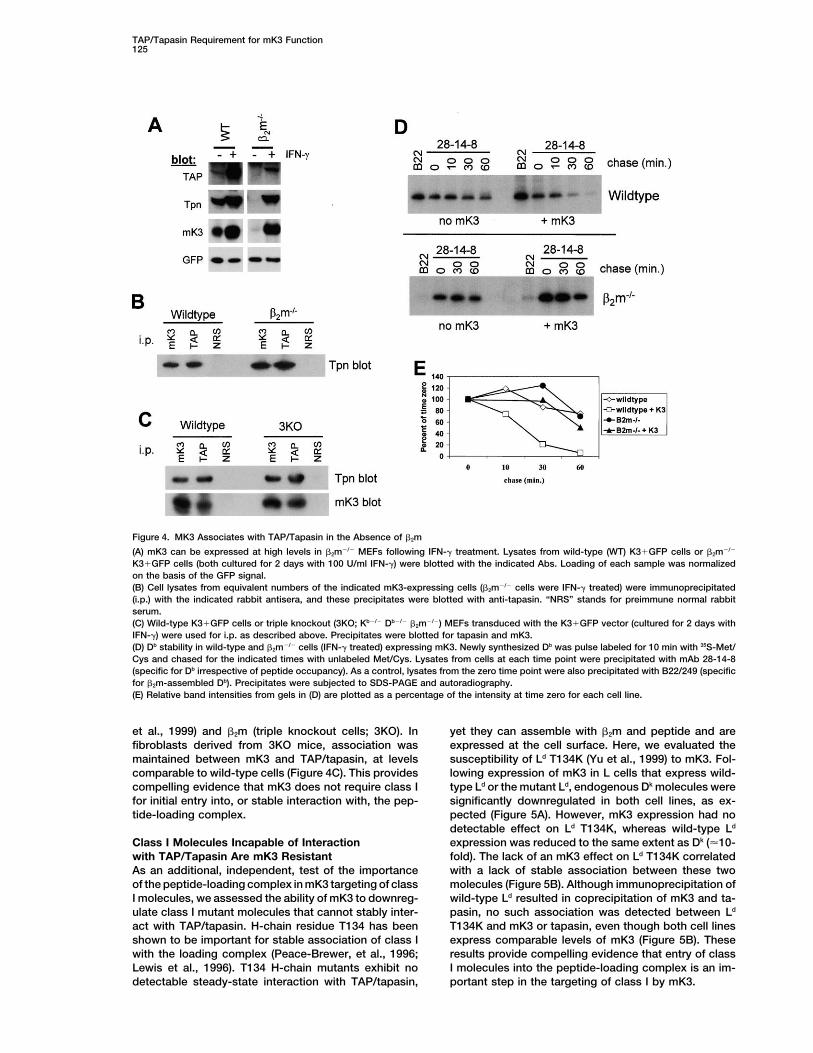
Figure 4. MK3 Associates with TAP/Tapasin in the Absence of �2m
(A) mK3 can be expressed at high levels in �2m�/� MEFs following IFN-� treatment. Lysates from wild-type (WT) K3�GFP cells or �2m�/�
K3�GFP cells (both cultured for 2 days with 100 U/ml IFN-�) were blotted with the indicated Abs. Loading of each sample was normalizedon the basis of the GFP signal.(B) Cell lysates from equivalent numbers of the indicated mK3-expressing cells (�2m�/� cells were IFN-� treated) were immunoprecipitated(i.p.) with the indicated rabbit antisera, and these precipitates were blotted with anti-tapasin. “NRS” stands for preimmune normal rabbitserum.(C) Wild-type K3�GFP cells or triple knockout (3KO; Kb�/� Db�/� �2m�/�) MEFs transduced with the K3�GFP vector (cultured for 2 days withIFN-�) were used for i.p. as described above. Precipitates were blotted for tapasin and mK3.(D) Db stability in wild-type and �2m�/� cells (IFN-� treated) expressing mK3. Newly synthesized Db was pulse labeled for 10 min with 35S-Met/Cys and chased for the indicated times with unlabeled Met/Cys. Lysates from cells at each time point were precipitated with mAb 28-14-8(specific for Db irrespective of peptide occupancy). As a control, lysates from the zero time point were also precipitated with B22/249 (specificfor �2m-assembled Db). Precipitates were subjected to SDS-PAGE and autoradiography.(E) Relative band intensities from gels in (D) are plotted as a percentage of the intensity at time zero for each cell line.
et al., 1999) and �2m (triple knockout cells; 3KO). In yet they can assemble with �2m and peptide and areexpressed at the cell surface. Here, we evaluated thefibroblasts derived from 3KO mice, association was
maintained between mK3 and TAP/tapasin, at levels susceptibility of Ld T134K (Yu et al., 1999) to mK3. Fol-lowing expression of mK3 in L cells that express wild-comparable to wild-type cells (Figure 4C). This provides
compelling evidence that mK3 does not require class I type Ld or the mutant Ld, endogenous Dk molecules weresignificantly downregulated in both cell lines, as ex-for initial entry into, or stable interaction with, the pep-
tide-loading complex. pected (Figure 5A). However, mK3 expression had nodetectable effect on Ld T134K, whereas wild-type Ld
expression was reduced to the same extent as Dk (�10-Class I Molecules Incapable of Interactionwith TAP/Tapasin Are mK3 Resistant fold). The lack of an mK3 effect on Ld T134K correlated
with a lack of stable association between these twoAs an additional, independent, test of the importanceof the peptide-loading complex in mK3 targeting of class molecules (Figure 5B). Although immunoprecipitation of
wild-type Ld resulted in coprecipitation of mK3 and ta-I molecules, we assessed the ability of mK3 to downreg-ulate class I mutant molecules that cannot stably inter- pasin, no such association was detected between Ld
T134K and mK3 or tapasin, even though both cell linesact with TAP/tapasin. H-chain residue T134 has beenshown to be important for stable association of class I express comparable levels of mK3 (Figure 5B). These
results provide compelling evidence that entry of classwith the loading complex (Peace-Brewer, et al., 1996;Lewis et al., 1996). T134 H-chain mutants exhibit no I molecules into the peptide-loading complex is an im-
portant step in the targeting of class I by mK3.detectable steady-state interaction with TAP/tapasin,

Immunity126
Figure 5. Ld T134K Mutants Do Not BindTAP/Tapasin and Are Resistant to mK3
L-Ld cells �/� mK3 were compared to L-Ld
T134K cells (Yu et al., 1999) �/� mK3.(A) Surface class I expression on the indi-cated cells, staining for Ld (mAb 30-5-7; leftpanels) or the endogenous class I molecule,Dk (mAb 15-5-5; right panels). The heavy linesindicate staining of the parental cells lines,and the dashed lines indicate staining of themK3-expressing cells. L cells stained withsecondary Ab alone were included as a con-trol (shaded peaks).(B) Lysates from L-Ld cells (�/� mK3) or L-Ld
T134K cells (�/� mK3) were precipitated withanti-mK3 serum or mAb 64-3-7 (specific foropen forms of Ld). Precipitates were then blot-ted with the Ab listed along the bottom of thefigure.
MK3 Induces the Rapid Loss of TAP-Associated pressing comparable amounts of mK3 (see Figure 5B)that resulted in ubiquitination of wild-type Ld molecules.Class I
Given that mK3 interaction with TAP/tapasin was re- This demonstrates that class I molecules require TAP/tapasin interaction to be ubiquitin-conjugated by mK3.quired for mK3 function, we next examined the turnover
of loading complex-associated class I molecules in thepresence of mK3 by pulse-chase labeling of newly syn- Discussionthesized class I molecules followed by anti-TAP immu-noprecipitation. Figures 6A and 6B reveal that in the Here, we describe a novel example of a virus that ex-
ploits host proteins interacting uniquely with class I aspresence of mK3, class I H-chains were relatively heavilyassociated with TAP after a short label, and these mole- a means to specifically target class I destruction. We
show that TAP and tapasin, two ER proteins that facili-cules were rapidly lost with time, similar to the total poolof Db (precipitated with mAb 28-14-8). This increased tate full assembly of class I heterotrimers, are required
for the targeting of class I by mK3. In support of therate of association between newly synthesized class IH-chains and TAP was highly reproducible. Conversely, critical role of TAP and tapasin, mK3 fails to regulate
class I in cell lines lacking either of these molecules.in the absence of mK3, fewer H-chains were associatedwith TAP at time zero, and the levels of TAP-associated Consistent with this requirement, class I H-chains in
�2m-deficient cells fail to associate with TAP/tapasinH-chains increased slightly over the time course of theexperiment, consistent with previous findings (Suh et (Ortmann et al. 1994; Suh et al., 1994; Carreno et al.,
1995) and are not efficiently degraded by mK3. Further-al., 1996). Thus, in the presence of mK3, class I mole-cules enter the loading complex quickly but are subse- more, we demonstrate that interaction with TAP/tapasin
has two separable outcomes on mK3: (1) stabilizationquently lost over time, indicating that the TAP/tapasin-associated pool of class I is targeted for mK3-mediated and (2) facilitation of class I recognition. Separation of
these two features of TAP/tapasin involvement in mK3degradation.To determine whether class I molecules are ubiquitin- function was provided by studies of �2m-deficient cells.
When �2m-deficient cells were treated with IFN-�, mK3conjugated while in the loading complex, TAP moleculeswere precipitated, followed by reprecipitation of class I protein was stabilized (presumably resulting from its
interaction with TAP/tapasin), but class I molecules were(Ld) molecules. The TAP-associated Ld molecules werethen immunoblotted with an anti-ubiquitin antibody. As relatively poor targets for degradation (due to a failure
to bind TAP/tapasin).shown in Figure 6C, TAP-associated class I moleculeswere clearly ubiquitinated in the presence of mK3 only, In regard to the role of TAP/tapasin in mK3 stabiliza-
tion, our preliminary data indicate that in the absencefurther demonstrating that this pool of H-chains is atarget of mK3. We next tested whether a class I molecule of TAP or tapasin, mK3 molecules are turned-over two
to three times more rapidly (Figure 3C and data notincapable of association with TAP/tapasin (Ld T134K)could be ubiquitinated. As shown in Figure 6D, Ld T134K shown). Interestingly, mutations of the PHD domain of
mK3 that abrogate its ubiquitin-conjugation activity (Bo-molecules were not ubiquitin-conjugated in cells ex-
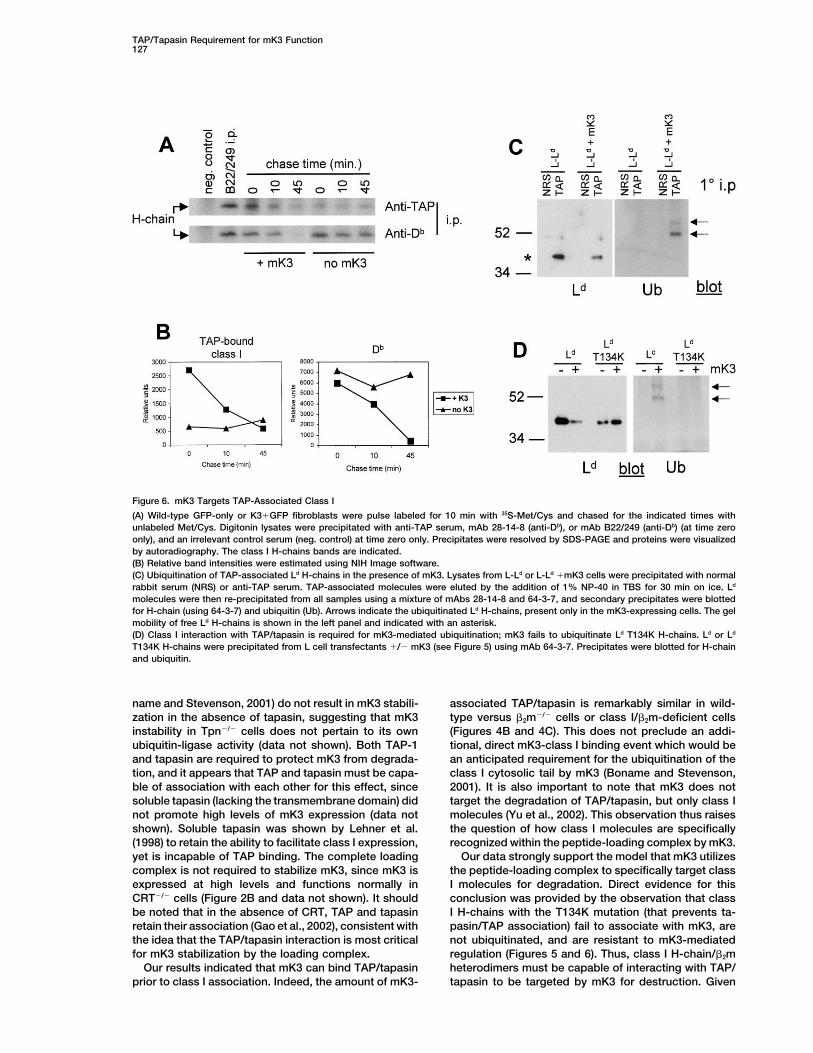
TAP/Tapasin Requirement for mK3 Function127
Figure 6. mK3 Targets TAP-Associated Class I
(A) Wild-type GFP-only or K3�GFP fibroblasts were pulse labeled for 10 min with 35S-Met/Cys and chased for the indicated times withunlabeled Met/Cys. Digitonin lysates were precipitated with anti-TAP serum, mAb 28-14-8 (anti-Db), or mAb B22/249 (anti-Db) (at time zeroonly), and an irrelevant control serum (neg. control) at time zero only. Precipitates were resolved by SDS-PAGE and proteins were visualizedby autoradiography. The class I H-chains bands are indicated.(B) Relative band intensities were estimated using NIH Image software.(C) Ubiquitination of TAP-associated Ld H-chains in the presence of mK3. Lysates from L-Ld or L-Ld �mK3 cells were precipitated with normalrabbit serum (NRS) or anti-TAP serum. TAP-associated molecules were eluted by the addition of 1% NP-40 in TBS for 30 min on ice. Ld
molecules were then re-precipitated from all samples using a mixture of mAbs 28-14-8 and 64-3-7, and secondary precipitates were blottedfor H-chain (using 64-3-7) and ubiquitin (Ub). Arrows indicate the ubiquitinated Ld H-chains, present only in the mK3-expressing cells. The gelmobility of free Ld H-chains is shown in the left panel and indicated with an asterisk.(D) Class I interaction with TAP/tapasin is required for mK3-mediated ubiquitination; mK3 fails to ubiquitinate Ld T134K H-chains. Ld or Ld
T134K H-chains were precipitated from L cell transfectants �/� mK3 (see Figure 5) using mAb 64-3-7. Precipitates were blotted for H-chainand ubiquitin.
name and Stevenson, 2001) do not result in mK3 stabili- associated TAP/tapasin is remarkably similar in wild-type versus �2m�/� cells or class I/�2m-deficient cellszation in the absence of tapasin, suggesting that mK3
instability in Tpn�/� cells does not pertain to its own (Figures 4B and 4C). This does not preclude an addi-tional, direct mK3-class I binding event which would beubiquitin-ligase activity (data not shown). Both TAP-1
and tapasin are required to protect mK3 from degrada- an anticipated requirement for the ubiquitination of theclass I cytosolic tail by mK3 (Boname and Stevenson,tion, and it appears that TAP and tapasin must be capa-
ble of association with each other for this effect, since 2001). It is also important to note that mK3 does nottarget the degradation of TAP/tapasin, but only class Isoluble tapasin (lacking the transmembrane domain) did
not promote high levels of mK3 expression (data not molecules (Yu et al., 2002). This observation thus raisesthe question of how class I molecules are specificallyshown). Soluble tapasin was shown by Lehner et al.
(1998) to retain the ability to facilitate class I expression, recognized within the peptide-loading complex by mK3.Our data strongly support the model that mK3 utilizesyet is incapable of TAP binding. The complete loading
complex is not required to stabilize mK3, since mK3 is the peptide-loading complex to specifically target classI molecules for degradation. Direct evidence for thisexpressed at high levels and functions normally in
CRT�/� cells (Figure 2B and data not shown). It should conclusion was provided by the observation that classI H-chains with the T134K mutation (that prevents ta-be noted that in the absence of CRT, TAP and tapasin
retain their association (Gao et al., 2002), consistent with pasin/TAP association) fail to associate with mK3, arenot ubiquitinated, and are resistant to mK3-mediatedthe idea that the TAP/tapasin interaction is most critical
for mK3 stabilization by the loading complex. regulation (Figures 5 and 6). Thus, class I H-chain/�2mheterodimers must be capable of interacting with TAP/Our results indicated that mK3 can bind TAP/tapasin
prior to class I association. Indeed, the amount of mK3- tapasin to be targeted by mK3 for destruction. Given

Immunity128
the variable dependency of different class I molecules unpublished data). Thus, mK3 does not impair H-chain/on the loading complex for surface expression (reviewed �2m association, per se, but merely alters the balanceby Williams et al., 2002a), it is intriguing to speculate of the interaction favoring the incorporation of unlabeledthat class I molecules with minimal dependency on the �2m. Collectively, the kinetic data presented here andloading complex may have evolved to counteract viral in our previous report (Yu et al., 2002) are consistentimmune evasion strategies similar to this one. with an obligatory interaction of mK3 with TAP/tapasin
It is interesting to note that the first six residues for the destruction of H-chain/�2m complexes.(KRRRNT) of the Db tail are required for mK3 association In summary, our combined data demonstrate that(Boname and Stevenson, 2001). A comparison of the mK3 exploits the peptide-loading complex for two noveltails of class I molecules susceptible to mK3-mediated interactions. First, mK3 associates with the loadingdownregulation does not yield a strong consensus, ex- complex, in a class I-independent manner, as a meanscept for the presence of multiple basic residues (data to increase its steady-state concentration, and second,not shown). In addition, the transmembrane domains of mK3 uses TAP/tapasin to impart target specificity byclass I, which have been implicated in KSHV kK3/kK5 bringing mK3 and nascent class I molecules into proxim-binding to class I (Ishido et al., 2000; Sanchez et al., ity with one another. These findings reveal a heretofore2002; Lorenzo et al., 2002; Hewitt et al., 2002), do not unappreciated, yet essential contextual framework forreveal an obvious consensus sequence. As noted by mK3 that must be considered for all subsequent studiesBoname and Stevenson (2001), sequences similar to of its recognition and function. Furthermore, we revealKRRRNT are present in other cellular surface proteins, a novel example of a virus-encoded class I inhibitoryyet mK3 displays remarkable specificity for class I even molecule that employs host cofactors to facilitate targetthough it is oriented on the opposite side of the ER recognition. Although unique to date, it seems likely thatmembrane from the class I ectodomain. Our data resolve target recognition by many other virus inhibitors of classthis enigma by demonstrating that mK3 requires an in- I will also require specific host cofactor molecules.teraction with the TAP/tapasin complex to specificallytarget class I molecules for destruction. Experimental Procedures
Through our analysis of TAP/class I association kinet-ics, we made the interesting observation that mK3 ex- Cell Lines
B6/WT-3 (Pretell et al., 1979) is an SV-40 transformed C57BL/6pression expedites the kinetics of class I interaction with(H2b) murine embryo fibroblast (MEF) line and was a gift of Stephenthe loading complex. Specifically, our data show thatJennings (Louisiana State University Health Sciences Center). Ta-maximal association of Db with TAP/tapasin is achievedpasin-deficient (Tpn�/�; Grandea et al., 2000), TAP-1-deficient (FT1�;
within 10 min in the presence of mK3 (Figure 6A). By Marguet et al., 1999), and calreticulin-deficient (K42, Gao et al.,comparison, in the absence of mK3, Db does not reach 2002) fibroblasts were gifts from Luc Van Kaer (Vanderbilt Universitymaximal association with TAP/tapasin until about 45 School of Medicine), Michael Edidin (Johns Hopkins University),
and Tim Elliott (University of South Hampton School of Medicine),min (Suh et al., 1996). A likely explanation for this kineticrespectively. �2m-deficient cells (B6.B2M�) were derived in this labdifference is that the rapid mK3-mediated degradationby serial passage of fibroblasts from day 16 C57BL/6 �2m�/� em-of class I H-chains results in significantly more availablebryos (Jackson Laboratories, Bar Harbor, ME). Triple knockout fibro-TAP/tapasin docking sites. The rapid association ofblasts (Kb�/� Db�/� �2m�/�; 3KO) were derived from day 16 embryos
class I with TAP/tapasin in mK3-expressing cells also from 3KO mice. These mice were derived by breeding Kb/Db doublyhelps to explain other enigmas regarding mK3 activity. deficient mice (H2b) (Perarnau, et al., 1999), a gift of F. LemmonierFor example, nascent class I molecules are turned over (Institut Pasteur) with �2m�/� (H2b) mice. F1 mice from this cross were
interbred to obtain 3KO founders. Mouse L cells (H2k) expressing Ld,rapidly in mK3-expressing cells (e.g., Ld and Db haveLd T134K, and Ld � mK3 have been described (Yu et al., 1999, 2002).half-lives of 15 min; Stevenson et al., 2000; Yu et al.,293T cells (Du Bridge et al., 1987) were used for ecotropic retrovirus2002). However, in these same cells, a relatively highproduction, and ProPakA.6 cells (ATCC, Rockville, MD) were usedlevel of class I molecules is associated with TAP/tapasinfor production of amphotropic retrovirus for transduction of L cells.
(Yu et al., 2002). The fact that class I associates much All cells were maintained in RPMI 1640 (Invitrogen, Carlsbad, CA)more rapidly with TAP/tapasin in mK3 positive cells ex- supplemented with 10% fetal calf serum (HyClone, Logan, UT), 2 mMplains how TAP/tapasin can be required for mK3 activity L-glutamine, 0.1 mM nonessential amino acids, 1.25 mM HEPES, 1
mM sodium pyruvate, and 100 U/ml penicillin/streptomycin (all fromand, at the same time, mK3-mediated class I degrada-the Tissue Culture Support Center at Washington University Schooltion can also be rapid.of Medicine). Where indicated, cells were cultured for 2–3 days inThis model for mK3 action also helps resolve anotherthe presence of 100 units/ml interferon-� (Biosource, Sunny-puzzle, in this case regarding the role of �2m. We show vale, CA).
here that (1) TAP/tapasin are required for mK3-mediatedclass I degradation (Figures 1 and 2) and that (2) �2m is
DNA Constructs and Retroviral Transductionsrequired for mK3 targeting of class I (Figure 4D), presum- For these studies, a biscistronic retroviral vector was used thatably by providing access of the H-chains to the peptide- encodes the gene of interest from the upstream cistron and GFPloading complex. Yet, in an earlier study, we showed from the downstream cistron (pMIG; Van Parijs et al., 1999). Thethat when mK3-expressing cells were labeled with a parental vector is referred to here as GFP-only. A vector containing
the mK3 gene as the upstream cistron was generated and is referredshort pulse of 35S-Met, Ld molecules exhibited impairedto as K3�GFP. The TAP-1-encoding retroviral vector (pMSCV-association with labeled �2m (Yu et al., 2002). A unifyingTAP-1b) was a gift of Robert Schreiber (Washington University Medi-explanation for these combined findings recently be-cal School). Retrovirus-containing supernatants were generated us-
came apparent. In mK3-expressing cells, the ratio of �2m ing either (1) the Vpack vector system (Stratagene, La Jolla, CA) forto H-chain is dramatically increased (due to selective transient production in 293T cells to generate ecotropic virus forH-chain loss), resulting in the preferential association infection of H2b cells or (2) ProPakA.6 cells for transient production
of virus for infection of H2k cells (L cells). Packaging cells wereof newly synthesized H-chains with unlabeled �2m (our

TAP/Tapasin Requirement for mK3 Function129
transfected using FuGene 6 (Roche Diagnostics, Indianaoplis, IN). viding critical reagents. We are grateful to Steven Truscott, NancyMyers, Lisa Kimpler, and Rose Connors for invaluable technicalVirus-containing supernatants were collected 48–72 hr post-
transfection, and 1–2 ml was added to �106 target cells per infec- assistance.This work was supported by NIH grants AI19687 (T.H.H. andtion. Hexadimethrine bromide (Sigma, St. Louis, MO) was also added
to a final concentration of 8 �g/ml. In most cases, the GFP� cells M.R.H.), AI42793 (T.H.H.), AI45019 (H.W.V.), CA74730 (H.W.V.), andT32AI07163 (L.L. and X.W.).from the transduced lines were sorted. These lines stably maintained
a high proportion (�90%) of GFP� cells for �12 weeks in culture.Received: July 17, 2002Revised: November 5, 2002Antibodies
A rabbit antiserum to C-terminal sequences of K3 (residues 167–187)Referenceswas generated by immunization with KLH-coupled peptide. Anti
mouse TAP-1 serum and anti mouse tapasin mAb 5D3 have beenAlbrecht, J.C., Nicholas, C.J., Biller, D., Cameron, K.R., Biesinger,described (Carreno et al., 1995; Harris et al., 2001). The anti GFPB., Newman, C., Wittmann, S., Craxton, M.A., Coleman, H., Fleck-mAb was obtained from Covance (Princeton, NJ). MAb B8-24-3enstein, B., et al. (1992). Primary structure of the herpesvirus saimirirecognizes folded Kb (ATCC, Rockville, MD), mAb 28-14-8 recog-genome. J. Virol. 66, 5047–5058.nizes the conformed �3 domain of Db irrespective of peptide occu-
pancy (Ozato et al., 1980a), and B22-249 recognizes fully assembled Ashland, R., Gibson, T.J., and Stewart, A.F. (1995). The PHD finger:Db (Hammerling et al., 1979). MAb 64-3-7 is specific for open forms implications for chromatin-mediated transcriptional regulation.(unassembled) of Ld (Smith et al., 1993), mAb 30-5-7 is specific for Trends Biochem. Sci. 20, 56–59.folded forms of Ld (Smith et al., 1993), and mAb 15-5-5 recognizes Boname, J.M., and Stevenson, P.G. (2001). MHC class I ubiquitina-folded Dk (Ozato et al., 1980b). Anti-ubiquitin blots were performed tion by a viral PHD/LAP finger protein. Immunity 15, 627–636.using mAb PD41 (Santa Cruz Biotech, Santa Cruz, CA).
Carbone, F.R., and Bevan, M.J. (1989). Induction of ovalbumin-spe-cific cytotoxic T cell in vivo. J. Exp. Med. 169, 603–612.
Flow CytometryCarreno, B.M., Solheim, J.C., Harris, M., Stroynowski, I., Connolly,All flow cytometric analyses were performed using a FACSCaliburJ.M., and Hansen, T.H. (1995). TAP associates with a unique class(Becton Dickinson, San Jose, CA). Data were analyzed using Cell-I conformation, whereas calnexin associates with multiple class IQuest software (Becton Dickinson). Staining was performed as de-forms in mouse and man. J. Immunol. 155, 4726–4733.scribed (Yu et al., 2002). PE-conjugated goat anti-mouse IgG (BDCoscoy, L., and Ganem, D. (2000). Kaposi’s sarcoma-associatedPharmingen, San Diego, CA) was used to visualize class I staining.herpesvirus encodes two proteins that block cell surface display ofGFP fluorescence was collected in the FITC channel, and electronicMHC class I chains by enhancing their endocytosis. Proc. Natl.compensation was applied to segregate the two signals.Acad. Sci. USA 97, 8051–8056.
Coscoy, L., Sanchez, D.J., and Ganem, D. (2001). A novel class ofImmunoprecipitations and ImmunoblotHerpesvirus-encoded membrane-bound E3 ubiquitin ligases regu-For coimmunoprecipitations, cells were lysed in Tris-buffered saline,lates endocytosis of proteins involved in immune recognition. J. CellpH 7.4 (TBS) � 1.0% digitonin (Wako, Richmond, VA) that containedBiol. 155, 1265–1273.a saturating concentration of precipitating antibody. After lysis for
30 min on ice, postnuclear lysates were incubated with Protein Cresswell, P. (2000). Intracellular surveillance: controlling the as-A-sepharose CL-4B (Amersham Pharmacia Biotech, Uppsala, Swe- sembly of MHC class I-peptide complexes. Traffic 1, 301–305.den) for 1 hr. Beads were washed four times in TBS � 0.1% digitonin, Day, P.M., Yewdell, J.W., Porgador, A., Germain, R.N., and Bennink,and proteins were eluted by boiling in LDS sample buffer (Invitrogen). J.R. (1997). Direct delivery of exogenous MHC class I molecule-For immunoblot of cell lysates, cells were lysed in TBS � 1.0% binding oligopeptides to the endoplasmic reticulum of viable cells.NP-40 (Sigma). Postnuclear lysates were mixed with LDS sample Proc. Natl. Acad. Sci. USA 94, 8064–8069.buffer, and 2-mercaptoethanal was added to a final concentration
Degen, E., and Williams, D.B. (1991). Participation of novel 88-kDof 1%. Immunoblot of all samples was performed following SDS-protein in the biogenesis of murine class I histocompatibility mole-PAGE separation of precipitated proteins essentially as describedcules. J. Cell Biol. 112, 1099–1115.(Yu et al., 2002). Specific proteins were visualized by chemilumines-Dick, T., Banjia, N., Peaper, D.R., and Cresswell, P. (2002). Disulfidecence using the ECL system (Amersham, Boston, MA).bond isomerization and the assembly of MHC class I-peptide com-plexes. Immunity 16, 87–98.Metabolic Labeling/Pulse-ChaseDu Bridge, R.B., Tang, P., Hsia, H.C., Leong, P.M., Miller, J.H., andAfter �45 min preincubation in Met/Cys-free medium (DMEM withCalos, M.P. (1987). Analysis of mutation in human cells by using an5% dialyzed FCS), cells (at 1 � 107 cells/ml) were pulse labeledEpstein-Barr virus shuttle system. Mol. Cell. Biol. 7, 379–387.with Express 35S-Met/Cys labeling mix (Perkin Elmer Life Sciences,
Boston, MA) at 175 �Ci/ml for 10 min. Chase was initiated by the Gao, B., Adhikari, R., Howarth, M., Nakamura, K., Gold, M.C., Hill,addition of an excess of unlabeled Met/Cys (5 mM each). Cells were A.B., Knee, R., Michalak, M., and Elliott, T. (2002). Assembly andthen washed, an aliquot removed for the zero time point, and the antigen-presenting function of MHC class I molecules in cells lack-remaining cells placed back in culture at 37 degrees for the indicated ing the ER chaperone calreticulin. Immunity 16, 99–109.times. For immunoprecipitations, labeled cells were lysed in 1% Grandea, A.G., III, Golovina, T.N., Hamilton, S.E., Sriram, V., Spies,NP-40 or 1% digitonin (for coimmunoprecipitations) dissolved in T., Brutkiewicz, R.R., Harty, J.T., Eisenlohr, L.C., and Van Kaer, L.TBS with 20 mM IAA and 5 mM PMSF (both from Sigma). Postnuclear (2000). Impaired assembly yet normal trafficking of MHC class Ilysates were precleared over protein A-sepharose CL-4B with nor- molecules in tapasin mutant mice. Immunity 13, 213–222.mal rabbit serum for �60 min on ice, then protein A-sepharose
Guerin, J.-L., Gelfi, J., Boullier, S., Delverdier, M., Ballanger, F.-A.,alone for �60 min on ice. Lysates were then transferred to proteinBertagnoli, S., Drexler, I., Sutter, G., and Messud-Petit, F. (2002).A-sepharose pellets containing the appropriate pre-bound mAbs.Myxoma Virus leukemia-associated protein is responsible for majorAfter binding for 45 min on ice, protein A pellets were washed fourhistocompatibility class I and Fas-CD95 down-regulation and de-times with 0.1% NP-40 or 0.1% digitonin in TBS, and bound proteinsfines scrapins, a new group of surface cellular receptor abductorwere eluted by boiling in 1X LDS sample buffer. Samples wereproteins. J. Virol. 76, 2912–2923.subjected to SDS-PAGE, and gels were treated with Amplify (Amer-Hammerling, G.J., Hammerling, U., and Lemke, H. (1979). Isolationsham), dried, and exposed to BioMax-MR film.of twelve monoclonal antibodies against Ia and H-2 antigens. Immu-nogenetics 8, 433–445.AcknowledgmentsHaque, M., Ueda, K., Nakano, K., Hirata, Y., Parravicini, C., Cor-bellino, M., and Yamanishi, K. (2001). Major histocompatibility com-We would like to thank Drs. Glen Grandea, Luc van Kaer, Michael
Edidin, Tim Elliott, Stephen Jennings, and Robert Schreiber for pro- plex class I molecules are down-regulated at the cell surface by the

Immunity130
K5 protein encoded by Kaposi’s sarcoma-associated Herpesvirus/ Saha, V., Chaplin, T., Gregorini, A., Ayton, P., and Young, D.B. (1995).The leukemia-associated protein (LAP) domain, a cysteine-rich mo-human Herpesvirus-8. J. Gen. Virol. 82, 1175–1180.tif, is present in a wide range of proteins, including MLL, AF10, andHarris, M.R., Lybarger, L., Yu, Y.Y.L., Myers, N.B., and Hansen, T.H.MLLT6 proteins. Proc. Natl. Acad. Sci. USA 92, 9737–9741.(2001). Association of ERp57 with mouse MHC class I molecules isSanchez, D.J., Coscoy, L., and Ganem, D. (2002). Functional organi-tapasin dependent and mimics that of calreticulin and not calnexin.zation of MIR2, a novel viral regulator of selective endocytosis. J.J. Immunol. 166, 6686–6692.Biol. Chem. 277, 6124–6130.Helenius, A., Trombetta, E.S., Herbert, D.N., and Simon, J.F. (1997).Spiliotis, E.T., Manley, H., Osorio, M., Zuniga, M.C., and Edidin, M.Calnexin and calreticulin and the folding of glycoproteins. Trends(2000). Selective export of MHC class I molecules from the ER afterCell Biol. 7, 193–200.their dissociation from TAP. Immunity 13, 841–851.Hewitt, E.C., Duncan, L., Mufti, D., Baker, J., Stevenson, P.G., andSmith, J.D., Myers, N.B., Gorka, J., and Hansen, T.H. (1993). ModelLehner, P.J. (2002). Ubiquitylation of MHC class I by the K3 viralfor the in vivo assembly of nascent Ld class I molecules and for theprotein signals internalization and TSG101-dependent degradation.expression of unfolded Ld molecules at the cell surface. J. Exp. Med.EMBO J. 10, 2418–2429.178, 2035–2045.Ishido, S., Wang, C., Lee, B.-S., Cohen, G.B., and Jung, J.U. (2000).Stevenson, P.G., Efstathiou, S., Doherty, P.C., and Lehner, P.J.Downregulation of major histocompatibility complex class I mole-(2000). Inhibition of MHC class I-restricted antigen presentation bycules by Kaposi’s sarcoma-associated herpesvirus K3 and K5 pro-�2-herpesviruses. Proc. Natl. Acad. Sci. USA 97, 8455–8460.teins. J. Virol. 74, 5300–5309.Suh, W.-K., Cohen-Doyle, F., Fruh, K., Wang, K., Peterson, P.A., andLehner, P.J., Surman, M.J., and Cresswell, P. (1998). Soluble tapasinWilliams, D.B. (1994). Interaction of MHC class I molecules withrestores MHC class I expression and function in the tapasin-nega-the transporter associated with antigen processing. Science 264,tive cell line 220. Immunity 8, 221–231.1322–1326.Lewis, J.W., Neisig, A., Neefjes, J., and Elliott, T. (1996). Point muta-Suh, W.-K., Mitchell, E.K., Yang, Y., Peterson, P.A., Waneck, G.L.,tions in the �2 domain of HLA-A2.1 define a functionally relevantand Williams, D.B. (1996). MHC class I molecules form ternary com-interaction with TAP. Curr. Biol. 6, 873–883.plexes with calnexin and TAP and undergo peptide-regulated inter-Lorenzo, M.E., Jung, J.U., and Ploegh, H.L. (2002). Kaposi’s sarcomaaction with TAP via their extracellular domains. J. Exp. Med. 184,associated Herpesvirus K3 utilizes the ubiquitin-proteasome system337–348.in routing class I major histocompatibility complexes to late endo-Tortorella, D., Gewurz, B.E., Furman, M.H., Schust, D.J., and Ploegh,cytic compartments. J. Virol. 76, 5522–5531.H.L. (2000). Viral subversion of the immune system. Annu. Rev. Im-
Marguet, D., Spiliotis, E.T., Pentcheva, T., Liebowitz, M., Schneck,munol. 18, 861–926.
J., and Edidin, M. (1999). Lateral diffusion of GFP-tagged H2Ld mole-Townsend, A., Ohen, C., Bastin, J., Lunggren, H.-G., Foster, L., andcules and GFP-TAP1 reports on the assembly and retention of theseKarre, K. (1989). Association of class I major histocompatility heavymolecules in the endoplasmic reticulin. Immunity 11, 231–240.and light chian induced with viral peptides. Nature 340, 443–448.
Means, R.E., Ishido, S., Alvarez, X., and Jung, J.U. (2002). MultipleVan Parijs, L., Refaeli, Y., Lord, J.D., Nelson, B.H., Abbas, A.K., andendocytic trafficking pathways of MHC class I molecules inducedBaltimore, D. (1999). Uncoupling IL-2 signals that regulate T cellby a Herpesvirus protein. EMBO J. 21, 1638–1649.proliferation, survival, and Fas-mediated activation-induced cell
Nicholas, J., Ruvolo, V., Zong, J., Ciufo, D., Guo, H.G., Reitz, M.S., death. Immunity 11, 281–288.and Hayward, G.S. (1997). A single 13-kilobase divergent locus in
van Santen, V.L. (1991). Characterization of the bovine herpesvirusthe Kaposi’s sarcoma associated herpesvirus (human herpesvirus4 major immediate-early transcript. J. Virol. 65, 5211–5224.8) genome contains nine open reading frames that are homologousVirgin, H.W., IV, Latreille, P., Wamsley, P., Hallsworth, K., Weck,to or related to cellular proteins. J. Virol. 71, 1963–1974.K.E., Dal Canto, A.J., and Speck, S.H. (1997). Complete sequenceOrtmann, B., Androlewicz, M., and Cresswell, P. (1994). MHC classand genomic analysis of murine gammaherpesvirus 68. J. Virol. 71,I/�2-microglobulin complexes associate with the TAP transporter5894–5904.before peptide binding. Nature 368, 864–867.Williams, A.P., Au Peh, C., and Elliott, T. (2002a). The cell biologyOzato, K., Hansen, T.H., and Sachs, D.H. (1980a). Monoclonal anti-of MHC class I antigen presentation. Tissue Antigens 59, 3–17.bodies to MHC antigens II: antibodies to the H-2Ld antigen, theWilliams, A.P., Peh, A.P., Purcell, A.W., McCluskey, J., and Elliott,product of a third polymorphic locus for the mouse major histocom-T. (2002b). Optimization of the MHC class I peptide cargo is depen-patibility complex. J. Immunol. 125, 2473–2477.dent on tapasin. Immunity 16, 509–520.Ozato, K., Mayer, N., and Sachs, D. (1980b). Hybridoma cell linesYu, Y.L.Y., Turnquist, H.R., Myers, N.B., Balendiran, G.K., Hansen,secreting monoclonal antibodies to mouse H-2 and Ia antigens. J.T.H., and Solheim, J.C. (1999). An extensive region of an MHC classImmunol. 124, 533–540.I �2 domain is loop influences interaction with the assembly com-
Peace-Brewer, A.L., Tussey, L.G., Matsui, M., Li, G., Quinn, D.G.,plex. J. Immunol. 163, 4427–4433.
and Frelinger, J.A. (1996). A point mutation in HLA-A*0201 resultsYu, L.L.Y., Harris, M.R., Lybarger, L., Kimpler, L.A., Myers, N.B.,in failure to bind the TAP complex and to present virus-derivedVirgin, H.W., IV, and Hansen, T.H. (2002). Physical association of thepeptides to CTL. Immunity 4, 505–514.K3 protein of gamma-2 Herpesvirus 68 with major histocompatibility
Perarnau, B., Saron, M.F., San Martin, B.R., Bervas, N., Ong, H.,complex class I molecules with impaired peptide and �2-microglob-
Soloski, M.J., Smith, A.G., Ure, J.M., Gairin, J.E., and Lemonnier,ulin assembly. J. Virol. 76, 2796–2803.
F.A. (1999). Single H2Kb, H2Db and double H2KbDb knockout mice:peripheral CD8� T cell repertoire and anti-lymphocytic choriomen-ingitis virus cytolytic responses. Eur. J. Immunol. 29, 1243–1252.
Ploegh, H.L. (1998). Viral strategies of immune evasion. Science 280,248–253.
Pretell, J., Greenfield, R.S., and Tevethia, S.S. (1979). Biology ofsimian virus 40 (SV40) transplantation antigen (TrAg). V. In vitrodemonstration of SV40 TrAg in SV40 infected nonpermissive mousecells by the lymphocyte mediated cytotoxicity assay. Virology 97,32–41.
Sadasivan, B., Lehner, P.J., Ortmann, B., Spies, T., and Cresswell,P. (1996). Roles for calreticulin and a novel glycoprotein, tapasin,in the interaction of MHC class I molecules with TAP. Immunity 5,103–114.







![[Monolingual Sounds, [No] Translation as Subversion and the Hope for Polyphony](https://img.dokumen.tips/doc/110x75/635977a1debc1859f604cad2/monolingual-sounds-no-translation-as-subversion-and-the-hope-for-polyphony.jpg)





















